SELEKSI PADI HIBRIDA TURUNAN GALUR MANDUL JANTAN TIPE
SITOPLASMA
WILD ABORTIVE
, KALINGA, DAN GAMBIACA
UNTUK TOLERANSI TERHADAP CEKAMAN KEKERINGAN
YUNI WIDYASTUTI
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Dengan ini saya menyatakan bahwa tesis berjudul “Seleksi Padi Hibrida Turunan Galur Mandul Jantan Tipe Sitoplasma Wild Abortive, Kalinga, dan Gambiaca untuk Toleransi terhadap Kekeringan” adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
RINGKASAN
YUNI WIDYASTUTI. Seleksi Padi Hibrida Turunan Galur Mandul Jantan Tipe Sitoplasma Wild Abortive, Kalinga, dan Gambiaca untuk Toleransi terhadap Cekaman Kekeringan. Dibimbing oleh BAMBANG SAPTA PURWOKO dan MUHAMAD YUNUS.
Kekeringan merupakan salah satu cekaman abiotik yang dapat menyebabkan penurunan hasil gabah padi. Bagi pemulia tanaman, perakitan varietas padi toleran cekaman kekeringan sekaligus memiliki potensi hasil tinggi menjadi sebuah tantangan. Teknologi padi hibrida merupakan alternatif untuk mencapai potensi hasil tinggi pada kondisi optimum, namun demikian penelitian pemanfaatan padi hibrida pada kondisi non optimum terutama kekeringan belum banyak dilakukan. Sebagian besar penelitian padi hibrida di Indonesia menggunakan galur mandul jantan (GMJ) tipe wild abortive (WA) sebagai tetua betina. Pemanfaatan GMJ tipe sitoplasma Gambiaca dan Kalinga diharapkan dapat meningkatkan keragaman padi hibrida di Indonesia dan memperluas preferensi petani. Penelitian ini bertujuan untuk: (1) menyeleksi galur-galur tetua dan F1 hibrida padi hibrida terhadap simulasi cekaman kekeringan pada fase
perkecambahan, (2) mengindentifikasi daya gabung umum (DGU) dan khusus (DGK) untuk karakter toleransi terhadap simulasi cekaman kekeringan pada fase perkecambahan, (3) mengindentifikasi daya gabung umum dan khusus untuk karakter hasil dan komponen hasil, (4) memverifikasi gen pengendali pemulih kesuburan GMJ tipe WA, Kalinga, dan Gambiaca, dan (5) mendapatkan F1 padi
hibrida potensi hasil lebih tinggi dibanding varietas Limboto dan Ciherang, memiliki sifat agronomi baik serta toleran terhadap simulasi cekaman kekeringan.
Serangkaian penelitian dilakukan untuk mengkombinasikan perakitan padi hibrida dari tiga sumber sitoplasma berbeda sekaligus memiliki toleransi terhadap simulasi cekaman kekeringan melalui: (1) Identifikasi toleransi kekeringan tetua dan F1 padi hibrida pada fase perkecambahan menggunakan Polietilen Glikol
(PEG) 6000 konsentrasi 25%, (2) Analisis daya gabung dan pengaruh genetik beberapa karakter toleransi kekeringan pada fase perkecambahan, (3) Analisis daya gabung dan heterosis karakter hasil dan komponen hasil hibrida turunan galur mandul jantan tipe Wild Abortive (WA), Kalinga, dan Gambiaca, dan (4) Verifikasi gen pemulih kesuburan yang mengendalikan pemulihan kesuburan tiga tipe GMJ melalui marka SSR.
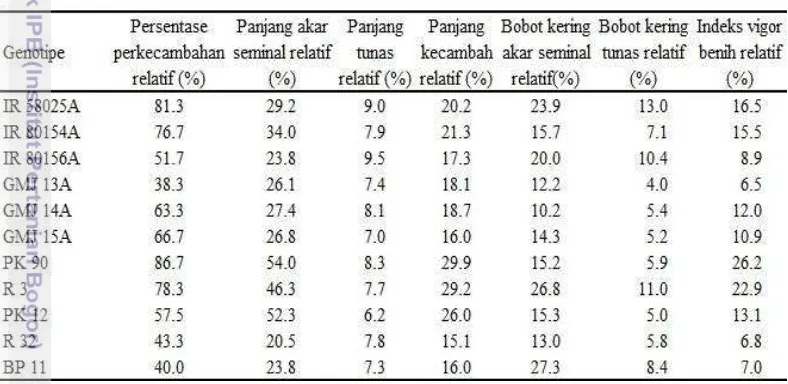
panjang akar seminal dan panjang kecambah. Di antara galur pemulih kesuburan, PK 90 merupakan penggabung baik untuk persentase perkecambahan dan indeks vigor benih. Kombinasi hibrida IR 80154A/PK 90, GMJ 13A/R 3, GMJ 14A/R 3, dan GMJ 15A/PK 90 memiliki DGK nyata dan positif untuk semua karakter perkecambahan yang diamati.
Analisis daya gabung dan heterosis karakter hasil dan komponen hasil hibrida turunan galur mandul jantan tipe Wild Abortive, Kalinga, dan Gambiaca berhasil mengidentifikasi GMJ IR 58025A sebagai penggabung umum terbaik untuk karakter jumlah gabah isi per malai, GMJ 14A untuk umur 50% berbunga dan persentase pengisian gabah, serta GMJ 15A untuk karakter jumlah gabah hampa per malai. Di antara tester, galur PK 12 penggabung umum terbaik untuk karakter umur 50% berbunga, jumlah gabah hampa per malai, dan persentase gabah isi per malai, sedangkan galur BP 11 untuk karakter jumlah gabah isi per malai. Hibrida baru dengan nilai standar heterosis > 20% dibanding Maro, Hipa 8, Ciherang, dan Limboto pada setiap latar belakang sistem GMJ adalah GMJ 13A/BP 11, GMJ 14A/PK 90, dan GMJ 15A/PK 90 masing-masing untuk WA, Kalinga, dan Gambiaca.
Pada kegiatan terakhir yaitu verifikasi gen pemulih kesuburan yang mengendalikan pemulihan kesuburan tiga tipe GMJ melalui marka SSR telah berhasil didapatkan informasi adanya kemiripan tingkat fertilitas serbuk sari dan spikelet dari hibrida-hibrida turunan Gambiaca dan Kalinga dengan hibrida tipe WA. Hal ini mengindikasikan bahwa terdapat kesamaan proses biologi yang mempengaruhi ketiga sistem sitoplasma ini. Pola segregasi pada populasi F2 untuk
karakter fertilitas serbuk sari pada GMJ tipe WA dan Gambiaca mengindikasikan keterlibatan dua gen pemulih kesuburan yang dominan, sedangkan tipe Kalinga diduga dikendalikan oleh gen dominan tunggal. Galur restorer PK 12 mampu memulihkan ketiga GMJ dengan ketiga sitoplasma lebih baik dibanding galur restorer PK 90 dan BP 11. Primer RM490 dan RM258 potensial digunakan sebagai penanda gen Rf3 dan Rf4 pada pemulihan kesuburan tipe WA.
Dari rangkaian penelitian ini dapat disimpulkan bahwa galur-galur tetua yang teridentifikasi toleran terhadap simulasi cekaman kekeringan berpotensi dapat digunakan untuk merakit F1 hibrida toleran cekaman kekeringan. Diperoleh
kombinasi hibrida hasil dan heterosis tinggi yang berasal dari GMJ tipe WA, Kalinga, dan Gambiaca. Galur-galur R yang selama ini digunakan untuk merakit hibrida dengan GMJ tipe WA berpotensi untuk dikembangkan sebagai tetua hibrida GMJ tipe Gambiaca dan Kalinga. Diperoleh dua F1 hibrida potensi hasil
tinggi, memiliki sifat agronomi baik serta toleran terhadap simulasi cekaman kekeringan pada fase perkecambahan yaitu GMJ 13A/R 3, dan GMJ 14A/R 3. Kata kunci: Galur mandul jantan, Wild Abortive, Gambiaca, Kalinga, padi hibrida,
SUMMARY
YUNI WIDYASTUTI. Selection of Hybrid Rice Derived from Wild Abortive, Gambiaca, and Kalinga Cytoplamic Male Sterile System for Breeding Program of Tolerant to Drought Stress. Supervised by BAMBANG SAPTA PURWOKO as chairman dan MUHAMAD YUNUS as member of the advisory committee.
Drought is an abiotic stress causing significant reduction in rice yield. This challenge breeder to breed new varieties with high yield potential and tolerance to drought. Hybrid rice is an alternative technology for achieving high yield potential under favorable conditions, but the performance of hybrids under drought stress has not yet been fully evaluated. In Indonesia, most of hybrid rice research used WA-cytoplasmic male sterile (CMS) source, which accounted for 90% commercial hybrids rice varieties. Utilization of CMS type Kalinga and Gambiaca is expected to increase diversity of hybrid rice and broader preference of farmers to cultivate it. The objectives of this research were (1) to select parental lines and F1 hybrids tolerant to drought at germination stage, (2) to identify general
combining ability (GCA) and specific combining ability (SCA) of germination traits in drought stress, (3) to identify GCA and SCA of yield components of parental lines and F1 hybrids, (4) to verify restoring ability genes of WA, Kalinga,
and Gambiaca CMS, and (5) to get new hybrid rice having higher yield potential than Limboto and Ciherang and tolerant to drought.
The studies were conducted to breed hybrid rice tolerant to drought from different CMS systems, through a series of experiments i.e. (1) Identification drought tolerant of parents and F1 rice hybrids at germination phase using 25%
concentration of Polyethylene Glycol (PEG) 6000, (2) The combining ability and genetic effects for some germination traits, (3) The combining ability analysis and heterosis for yield and yield components of rice hybrids derived from WA, Kalinga, and Gambiaca CMS systems, and (4) Verification of fertility restorer genes controlling the restoring fertility of three CMS systems throught SSR markers.
The results of first experiment showed that the germination percentage, seed vigor index, seminal root length, and seedling length were the primary evaluation indicator for tolerance to drought at germination phase. The use of PEG 6000 concentration of 25% at the germination phase could identify maintainer lines (i.e. IR 58025B, IR 80154B, GMJ 14B, and GMJ 15B) and restorer lines (i.e. PK 90, R 3, dan PK 12) as genotypes tolerant to drought simulation. Twelve F1 hybrids
identified tolerant to drought were IR 58025A/R 32, IR 80154A/PK 90, IR 80154A/ R3, IR 80154A/PK 12, IR 80154A/BP 11, IR 80156A/BP 11, GMJ 13A/PK 90, GMJ 13A/R 3, GMJ 13A/BP 11, GMJ 14A/R 3, GMJ 14A/BP 11, and GMJ 15A/PK 90.
appeared that IR 58025A line was a good general combiner for number of filled grains per panicle, GMJ 14A line was a good general combiner for 50% days of maturity and and filled grain percentage per panicle, while GMJ 15A was good combiner for number of unfilled grains per panicle. Among the testers, PK 12 line was the best general combiner for 50% days maturity, number of unfilled grain per panicle, and filled grain percentage per panicle, while BP 11 was the best general combiner for number of filled grain per panicle. The highest specific combining ability for weight of grain per hill was achieved by GMJ 13A/PK 90 (WA-CMS), IR 80154A/R 32 (Gambiaca-CMS), and GMJ 14A/ R 3 (Kalinga-CMS). The highest value of standard heterosis better than Maro, Hipa 8, Ciherang, and Limboto were GMJ 13A/BP 11, GMJ 14A/PK 90, and GMJ 15A/BP 11 for CMS systems i.e. WA, Kalinga, and Gambiaca repectively.
The genetic relationship among three cytoplasmic male sterility (CMS) systems, consisting of wild abortive (WA), Gambiaca, and Kalinga was studied. The results showed that the F1 plants derived from crosses involving A and R
lines of the respective cytoplasm and their cross-combination showed similar pollen and spikelet fertility values, indicating that similar biological processes governed fertility restoration in these three CMS systems. The results from the inheritance of fertility restoration was studied in crosses involving three elite restorer lines of rice viz. PK 90, PK 12, and BP 11 and three male sterile line IR 58025A WA), IR 80154A Gambiaca), and IR 80156A (CMS-Kalinga). The results from the inheritance study showed that the pollen fertility restoration in WA and Gambiaca CMS systems was governed by two independent and dominant genes with classical duplicate gene action, while Kalinga system controlled by one single dominant gene. The restorer line PK 12 have restoring ability stronger than PK 90 and BP 11. The SSR primers RM490 and RM258 revealed potential to be markers of Rf3 and Rf4 genes in restoring fertility of WA system.
It can be concluded that the parental lines identified as tolerant to drought potential to be use as parents of hybrid rice tolerant to drought. The Kalinga and Gambiaca CMS system could restored the fertility by CMS-WA. These results have important implications for the hybrid rice breeding using different cytoplasmic backgrounds.
© Hak Cipta Milik IPB, Tahun 2016
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB.
Tesis
Sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Pemuliaan dan Bioteknologi Tanaman
SELEKSI PADI HIBRIDA TURUNAN GALUR MANDUL JANTAN
TIPE SITOPLASMA
WILD ABORTIVE
, KALINGA, DAN
GAMBIACA UNTUK TOLERANSI TERHADAP CEKAMAN
KEKERINGAN
YUNI WIDYASTUTI
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Nama NIM
Yuni Widyastuti
A 253130271
Disetujui oleh
Komisi Pembimbing
Prof Dr Ir S. MSc
Ketua
Ketua Program Studi
Pemuliaan dan Bioteknologi Tanaman
Dr Ir Wahyu E.K., MS
Tanggal Ujian Tesis :
18 Januari 2016
Diketahui oleh
PRAKATA
Alhamdulillaahirabbil’aalamiin. Puji dan syukur penulis panjatkan kepada Allah Subhanahu Wa Ta’ala atas limpahan karunia-Nya sehingga tesis ini dapat diselesaikan dengan baik.
Perubahan iklim global, salah satunya mengakibatkan banyak terjadi gagal panen karena cekaman kekeringan. Teknologi padi hibrida toleran cekaman kekeringan diharapkan mampu menjadi alternatif solusi penurunan hasil akibat kekeringan. Terlebih lagi, belum optimalnya penggunaan galur mandul jantan tipe Kalinga dan Gambiaca sebagai tetua pada perakitan padi hibrida di Indonesia, maka penulis memilih tema Seleksi Padi Hibrida Turunan Galur Mandul Jantan Tipe Sitoplasma Wild Abortive, Kalinga, dan Gambiaca untuk Toleransi terhadap Cekaman Kekeringan.
Terimakasih penulis sampaikan kepada Prof Dr Ir Bambang S. Purwoko, MSc dan Dr Ir Muhamad Yunus, MSi selaku komisi pembimbing yang dengan ikhlas memberikan masukan, arahan, bimbingan, dorongan dan motivasi dalam penulisan tesis ini. Penulis juga menyampaikan terimakasih kepada Badan Litbang Pertanian Kementerian Pertanian atas dukungan dana beasiswa dan dana penelitian melalui DIPA Balitbang Tahun 2015. Terimakasih juga penulis sampaikan kepada Kepala Balai Besar Penelitian Padi, ketua tim penelitian padi hibrida Dr Satoto, tim peneliti dan teknisi: Dr Indrastuti A. Rumanti, Nita Kartina SP, Bayu Pramono SP, R. Noviadi SSi, Sarmadi, Soewarto, Cecep, Firman, Dede, dan Kamsi, atas segenap bantuan dan dukungannya. Terimakasih kepada teman-teman seperjuangan di program studi Pemuliaan dan Bioteknologi Tanaman Angkatan 2013 atas persahabatan dan kekeluargaannya.
Kepada suami tercinta, Sudradjat, SE Ak CA, ananda terkasih Najwa Kamila, Ahza Syarif Al Auni, dan Attar Syauqi Azzaini, terimakasih atas cinta, doa, dorongan dan kasih sayang yang tidak terhingga. Keluarga besar Bani Muhdayat dan Sudjai terimakasih atas dukungan yang telah diberikan.
Tiada yang sempurna pada karya manusia, tetapi besar harapan penulis, karya ini dapat bermanfaat dalam pengembangan padi, khususnya padi hibrida di Indonesia.
Bogor, Maret 2016
Mekanisme Mandul Jantan dan Pemulihan Kesuburan 6
Toleransi Kekeringan 7
Pemanfaatan Marka Molekuler pada Padi Hibrida 9
3 IDENTIFIKASI TOLERANSI KEKERINGAN TETUA DAN F1 PADI HIBRIDA ADA FASE PERKECAMBAHAN
4 ANALISIS DAYA GABUNG DAN PENGARUH GENETIK BEBERAPA KARAKTER TOLERANSI KEKERINGAN PADA
6 VERIFIKASI GEN PEMULIH KESUBURAN YANG MENGENDALIKAN PEMULIHAN KESUBURAN TIGA TIPE GMJ MELALUI MARKAH SSR
66
Abstrak 66
Abstract 67
Pendahuluan 68
Bahan dan Metode 69
Hasil dan Pembahasan 73
Simpulan 79
7 PEMBAHASAN UMUM 80
Metode Seleksi Cepat pada Fase Perkecambahan sebagai Alternatif
Seleksi Genotipe Tolerang Cekaman Kekeringan 81
Peran Tetua dalam Perakitan Padi Hibrida Toleran Cekaman
Kekeringan dan Hasil Tinggi 82
Pemanfaatan GMJ Tipe Sitoplasma WA, Kalinga, dan Gambiaca
dalam Peningkatan Produksi Padi Hibrida 84
Kendali Genetik Pemulihan Kesuburan pada Tipe Sitoplasma WA,
Kalinga, dan Gambiaca 85
8 SIMPULAN DAN SARAN 87
Simpulan 87
Saran 87
DAFTAR PUSTAKA 88
RIWAYAT HIDUP 99
1 Materi genetik skrining tetua padi hibrida menggunakan PEG 6000
konsentrasi 25% pada fase perkecambahan 14
2 Materi genetik skrining F1 padi hibrida menggunakan PEG 6000
konsentrasi 25% pada fase perkecambahan 16
3 Analisis ragam pada perlakuan genotipe dan PEG 6000 pada variabel
perkecambahan 21 genotipe padi 17
4 Pengaruh simulasi cekaman kekeringan terhadap penurunan rata-rata enam variabel perkecambahan pada 21 genotipe padi 18 5 Koefisien korelasi variabel perkecambahan padi 20 6 Analisis komponen utama terhadap 8 variabel perkecambahan padi 21 7 Nilai koefisien toleransi cekaman kekeringan empat variabel utama
toleransi kekeringan pada fase perkecambahan padi 22 8 Pengaruh larutan PEG 6000 konsentrasi 25% terhadap persentase
perkecambahan dan panjang akar seminal padi 24
9 Pengaruh larutan PEG 6000 konsentrasi 25% terhadap panjang tunas
dan panjang kecambah padi 25
10 Pengaruh PEG 6000 konsentrasi 25% terhadap variabel bobot kering
akar seminal dan bobot kering tunas padi 27
11 Nilai koefisien toleransi cekaman kekeringan empat variabel utama toleransi kekeringan pada fase perkecambahan padi 28 12 Material genetik analisis daya gabung karakter toleransi fase
perkecambahan padi 33
13 Karakter perkecambahan galur tetua padi hibrida pada kondisi
cekaman kekeringan 35
14 Analisis ragam karakter perkecambahan padi pada kondisi cekaman
kekeringan 36
15 Nilai kuadrat tengah hasil analisis ragam karakter perkecambahan
genotipe padi 36
16 Proporsi kontribusi lini, tester dan interaksi lini tester terhadap karakter
perkecambahan padi 37
17 Nilai duga daya gabung umum karakter toleransi cekaman kekeringan fase perkecambahan padi menggunakan persilangan lini x tester 38 18 Nilai duga daya gabung khusus karakter toleransi cekaman kekeringan
fase perkecambahan padi menggunakan persilangan lini x tester 30 19 Materi genetik uji daya gabung karakter hasil dan komponen hasil padi
hibrida, tetua, dan varietas pembandingnya 46
20 Struktur analisis ragam untuk analisis lini x tester 47 21 Nilai kuadrat tengah hasil analisis ragam daya gabung karakter hasil
dan komponen hasil padi 49
22 Proporsi kontribusi lini, tester, interaksi lini x tester karakter hasil dan
komponen hasil padi 51
23 Nilai duga daya gabung umum tetua padi hibrida pada karakter hasil
dan komponen hasil 52
24 Nilai duga daya gabung khusus karakter hasil dan komponen hasil 30
padi hibrida 54
25 Nilai rata-rata karakter hasil dan komponen hasil tetua padi hibrida
yang digunakan 56
26 Nilai rata-rata karakter hasil dan komponen hasil 30 F1 hibrida padi 57
27 Nilai heterosis dibanding tetua terbaik (heterobeltiosis) karakter hasil
dan komponen hasil padi hibrida 60
28 Nilai heterosis dibanding rata-rata kedua tetua (mid-parent heterosis) karakter hasil dan komponen hasil padi hibrida 62 29 Hasil gabah 30 F1 hibrida padi dan standar heterosis terhadap 4 varietas
pembanding 64
30 Materi genetik padi hibrida untuk verifikasi gen pemulih kesuburan 70
31 Kode dan sekuen dari pasangan primer SSR 72
32 Fertilitas serbuk sari pada tanaman F1 hibrida padi turunan AxR kombinasi GMJ sistem WA, Kalinga, dan Gambiaca dengan 3 galur R 74 33 Fertilitas spikelet pada tanaman F1 hibrida padi turunan AxR
kombinasi GMJ sistem WA, Kalinga, dan Gambiaca dengan 3 galur R 74 34 Analisis Chi-Kuadrat karakter fertilitas serbuk sari pada tanaman
populasi F2 padi turunan GMJ tipe WA, Kalinga, dan Gambiaca 76
35 Primer polimorfis untuk gen Rf3 dan Rf4 77
36
Hasil analisis Chi-Kuadrat untuk fertilitas serbuk sari pada tanaman padi populasi F2 IR 58025A/PK 90 (WA) berdasarkan pola pita DNA
yang diperoleh
78
37
Hasil analisis Chi-Kuadrat untuk fertilitas serbuk sari pada tanaman padi populasi F2 IR 80154A/PK 12 (Gambiaca) berdasarkan pola pita
DNA yang diperoleh
78
37
Hasil analisis Chi-Kuadrat untuk fertilitas serbuk sari pada tanaman padi populasi F2 IR 80156A/BP 11 (Kalinga) berdasarkan pola pita
DNA yang diperoleh
79
1 Bagan alur kegiatan penelitian. 3
2 Metode tiga galur pada perakitan padi hibrida 5
3 Keragaan kecambah tetua padi hibrida pada kondisi cekaman
menggunakan PEG 6000 konsentrasi 25%. 21
4 Keragaan padi galur tetua maintainer 58
5 Keragaan padi galur tetua pemulih kesuburan 58
6 Keragaan F1 hibrida padi terseleksi 66
7 Klasifikasi sterilitas serbuk sari pada padi 71
8 Butir serbuk sari GMJ dengan tipe sitoplasma berbeda 71
1 PENDAHULUAN
Latar Belakang
Padi (Oryza sativa L.), merupakan salah satu tanaman pangan yang berhasil ditingkatkan produksinya melalui pemanfaatan fenomena heterosis atau vigor hibrida. China merupakan negara yang merintis penelitian padi hibrida pada tahun 1970 an dan berhasil menunjukkan bahwa padi hibrida mampu memberikan hasil 1-1.5 ton ha-1 atau 20-30% lebih tinggi dibanding varietas konvensional inbrida
(Lin & Yuan 1980; Ma & Yuan 2003). Pemanfaatan teknologi ini diharapkan dapat menurunkan masalah kelaparan dan malnutrisi di Asia, Afrika, dan Amerika Latin yang sebagian besar penduduknya bergantung pada beras (Toriyama et al. 2010). Sampai tahun 2013, Badan Penelitian dan Pengembangan Pertanian telah melepas 19 varietas unggul hibrida ke masyarakat (BB Padi 2014). Target perakitan padi hibrida sampai saat ini terfokus pada hibrida untuk lahan sawah irigasi dengan kondisi lingkungan optimum.
Perubahan pola iklim pada saat ini menyebabkan kekeringan tidak hanya mengancam lahan gogo atau tadah hujan (Balasubramanian et al. 2007) tetapi juga lahan sawah irigasi terutama irigasi semi teknis (Susanto et al. 2012). Hal tersebut ditambah dengan kerusakan infrastruktur pengairan karena kurangnya pemeliharaan menjadikan pertanaman padi sawah irigasi pun terancam kekeringan. Akibatnya produksi beras terancam turun karena meluasnya areal lahan dengan risiko gagal panen. Perakitan varietas unggul padi toleran cekaman kekeringan menjadi salah satu komponen teknologi yang penting untuk mengantisipasi lahan-lahan tercekam kekeringan.
Selama ini penggunaan padi hibrida masih terbatas di lahan irigasi. Pada kondisi optimum padi hibrida dilaporkan mampu memberikan hasil 20-30% lebih tinggi dibanding padi inbrida (Virmani et al. 1982). Virmani (2003) melaporkan terdapat superioritas padi hibrida pada lingkungan tercekam baik kekeringan atau pada lahan bermasalah lainnya. Pada kondisi tercekam kekeringan, padi hibrida toleran kekeringan tetap mampu mengekspresikan potensi heterosisnya lebih baik dibanding padi hibrida yang tidak toleran (Yaqoob et al. 2012). Atlin et al. (2008) berhasil mengidentifikasi dua hibrida yang memiliki potensi hasil tinggi saat ditanam di lahan sawah yang memiliki irigasi minimum. Xie (2010) melaporkan padi hibrida yang dikembangkan di lahan irigasi menunjukkan heterosis tinggi pada saat dievaluasi pada kondisi tercekam kekeringan. Luo (2010) berhasil merakit padi hibrida toleran kekeringan melalui tetua galur mandul jantan yang juga toleran terhadap kekeringan.
Perakitan padi hibrida di Indonesia sampai saat ini menggunakan sistem tiga galur terdiri atas galur mandul jantan (GMJ=A=CMS) sebagai tetua betina dan galur pemulih kesuburan (R=restorer) sebagai tetua jantan. Galur pelestari (B=maintainer) digunakan sebagai tetua jantan pada perbanyakan benih GMJ. Sistem cytoplasmic male sterility (CMS) pada padi dimanfaatkan untuk eksploitasi heterosis dan dapat menghasilkan hasil gabah tinggi apabila dikombinasikan dengan galur restorer (pemulih kesuburan) yang efektif (Nematzadeh & Kiani 2010).
pemulihan kesuburan, dan pelestari kemandulan. Sattari et al. (2008) melaporkan hasil studi genetik terhadap GMJ tipe WA (subspesies Indica), Gambiaca (subspesies Indica dari Afrika Barat), dan Dissi (subspesies Indica, DS 97A, dari Senegal), menunjukkan bahwa galur pelestari dari satu GMJ dapat melestarikan GMJ dengan tipe sitoplasma berbeda. Selain itu proses biologi yang mirip mempengaruhi pemulihan kesuburan ketiga tipe sitoplasma tersebut. Hal ini disebabkan ketiga GMJ tersebut berasal dari sistem sitoplasma yang sama yaitu sporofitik. Penelitian Khrisnalata dan Sharma (2012) terhadap GMJ tipe WA dan Kalinga menunjukkan bahwa galur yang dapat memelihara kemandulan dan memulihkan kesuburan pada GMJ tipe WA tidak selalu dapat berperan sama pada GMJ tipe Kalinga. Hal ini diduga karena adanya perbedaan gen pengendali pemulihan kesuburan pada GMJ tipe WA dan Kalinga. Menurut Li et al. (2007), GMJ tipe WA termasuk tipe sporofitik yang gugur serbuk sarinya pada fase uninukleat sedangkan GMJ tipe Kalinga termasuk tipe gametofitik yang gugur saat fase binukleat dan trinukleat. Namun demikian, Sahu et al. (2014) melaporkan dari lima galur restorer yang diuji, satu galur dapat memulihkan baik GMJ tipe WA maupun Kalinga.
Sebagian besar penelitian padi hibrida di Indonesia terarah pada CMS-WA yang ditunjukkan dengan 90% varietas padi hibrida komersial menggunakan tipe ini sebagai galur mandul jantan. Pada tahun 1970, sebesar 85% pertanaman jagung hibrida di AS hancur karena Helminthosporium maydis yang menyebabkan kerusakan parah terutama pada hibrida berbasis CMS-T (Sofi et al. 2007). Kasus serupa dapat mengancam pertanaman padi, akibat penanaman secara monokultur pada lahan luas, jangka waktu lama, dan latar belakang genetik yang sama. Untuk menghindari hal tersebut, maka penggunaan sistem CMS lainnya pada perakitan padi hibrida sangat diperlukan (Toriyama et al. 2010).
IRRI telah mengidentifikasi beberapa sumber sitoplasma lain, antara lain Gambiaca, Dissi, ARC, mutagen IR 62829B dan Kalinga (Virmani & Toledo 2000). Rumanti (2011) melalui disertasinya berhasil mengembangkan GMJ tipe WA, Gambiaca dan Kalinga yang bereaksi tahan terhadap patogen HDB patotipe III, berturut-turut yaitu BI 703A, BI 855A dan BI 669A. Namun kedua tipe GMJ yaitu Kalinga dan Gambiaca tersebut belum banyak dieksplorasi keunggulannya pada perakitan tanaman F1 hibrida di Indonesia.
Analisis molekuler untuk mengetahui gen-gen yang mengendalikan pemulihan kesuburan pada CMS dengan tipe sitoplasma berbeda telah banyak dilakukan. Melalui penggunaan marka RFLP, Zhang et al. (1997) berhasil memetakan satu gen Rf pada kromosom nomor 1 (Rf3). Yao et al. (1997) mengkonfirmasi keberadaan Rf3 pada kromosom nomor 1 dan memetakan gen Rf lain yaitu Rf4 pada kromosom nomor 10 menggunakan marka RAPD dan RFLP. Jing et al. (2001), memetakan gen Rf4 yang mempengaruhi pemulihan kesuburan pada kromosom nomor 10 melalui marka SSLP sedangkan Zhang et al. (2002) menggunakan marka RFLP untuk mengidentifikasi gen Rf4. Adapun Bazrkar et al. (2008) berhasil menandai empat gen Rf untuk CMS-WA menggunakan marka SSR pada kromosom nomor 1 (Rf3), 7 (Rf6), 10 (Rf4), dan 12 (Rf7) begitu pula Jie et al. (2008) yang memetakan gen pemulih kesuburan pada kromosom nomor 1 dan 10 melalui marka SSR.
Serangkaian kegiatan penelitian dilakukan untuk membentuk F1 hibrida padi
sitoplasmik WA, Kalinga, dan Gambiaca. Alur penelitian disajikan pada Gambar 1.
Gambar 1 Bagan alur kegiatan penelitian.
Perumusan Masalah
Pemanfaatan galur mandul jantan pada perakitan padi hibrida di Indonesia sampai saat ini masih banyak menggunakan tipe sitoplasma Wild Abortive. Penggunaan satu tipe sitoplasma dalam waktu yang lama dan terus menerus dapat mempersempit keragaman genetik F1 hibrida yang dihasilkan sehingga rawan
perakitan tanaman F1 hibrida. Sementara itu perakitan hibrida toleran cekaman
kekeringan dapat menjadi salah satu alternatif solusi untuk memberikan kelebihan hasil terutama untuk lahan sawah tadah hujan dan irigasi semi teknis.
Berdasarkan uraian di atas, timbul pertanyaan penelitian berkaitan dengan upaya untuk memanfaatkan galur mandul jantan dengan tipe sitoplasma WA, Kalinga, dan Gambiaca pada perakitan padi hibrida toleran kekeringan. Secara khusus dapat dirumuskan permasalahan sebagai berikut :
1. Adakah respon galur-galur tetua dan F1 padi hibrida dengan uji cepat terhadap
simulasi cekaman kekeringan pada fase perkecambahan.
2. Bagaimana daya gabung tetua padi hibrida dalam membentuk F1 hibrida
toleran simulasi cekaman perkecambahan dan berdaya hasil tinggi.
3. Adakah perbedaan gen pengendali pemulihan kesuburan GMJ tipe WA, Kalinga, dan Gambiaca.
4. Adakah F1 padi hibrida yang memiliki potensi hasil lebih tinggi dibanding
varietas inbrida Limboto dan Ciherang, memiliki sifat agronomi baik serta toleran terhadap cekaman kekeringan pada fase perkecambahan.
Tujuan Penelitian
1. Menyeleksi galur-galur tetua dan F1 hibrida padi hibrida dengan uji cepat
terhadap simulasi cekaman kekeringan pada fase perkecambahan.
2. Mengindentifikasi daya gabung umum dan khusus untuk karakter toleransi terhadap simulasi cekaman kekeringan pada fase perkecambahan dari tetua dan F1 hibrida.
3. Mengindentifikasi daya gabung umum dan khusus untuk karakter hasil dan komponen hasil dari tetua dan F1 hibrida.
4. Memverifikasi gen pengendali pemulih kesuburan GMJ tipe WA, Kalinga, dan Gambiaca.
5. Mendapatkan F1 padi hibrida dengan potensi hasil lebih tinggi dibanding
varietas inbrida Limboto dan Ciherang, memiliki sifat agronomi baik serta toleran terhadap cekaman kekeringan. untuk karakter-karakter toleransi terhadap cekaman kekeringan pada fase perkecambahan.
3. Terdapat daya gabung yang baik antara GMJ dan galur pemulih kesuburan untuk karakter hasil dan komponen hasil
4. Terdapat perbedaan secara genetik pengendali pemulih kesuburan pada GMJ tipe WA, Kalinga, dan Gambiaca.
5. Terdapat kombinasi F1 hibrida padi baru potensi hasil tinggi dan toleran
2. TINJAUAN PUSTAKA
Perakitan Padi Hibrida
Perakitan atau pemuliaan tanaman hibrida dilandasi oleh fenomena heterosis yaitu suatu kecenderungan bahwa F1 akan tampil lebih baik dibandingkan dengan
salah satu atau rata-rata kedua tetuanya. Penampilan gejala heterosis atau vigor hibrida dapat dinyatakan dengan berbagai indikator seperti hasil, tinggi tanaman, sistim perakaran, jumlah malai, jumlah biji, kandungan lemak, kandungan protein, besar tongkol, dan sebagainya (Virmani 2004). Pemanfaatan gejala heterosis dalam produksi tanaman pertanian secara komersial adalah pembentukan varietas hibrida.
Cytoplasmic male sterile (CMS) atau mandul jantan sitoplasma merupakan kondisi ketidakmampuan tanaman untuk memproduksi serbuk sari fungsional (Eckardt 2006). Sistem ini dimanfaatkan untuk memproduksi benih hibrida beberapa jenis tanaman seperti: jagung, padi, kapas, dan tanaman sayuran lain sehingga tanaman tersebut dapat meningkatkan produktifitas melalui fenomena heterosis pada turunan pertama.
Padi merupakan tanaman menyerbuk sendiri yang secara alami bunga jantan (serbuk sari) menyerbuki putik (bunga betina) pada bunga dan tanaman yang sama. Penyerbukan silang pada dua tanaman padi yang berbeda untuk menghasilkan benih hibrida dapat dimungkinkan jika bunga jantan pada tanaman betina bersifat mandul atau dibuat tidak berfungsi yang dapat dibuat dengan cara memasukkan gen CMS. Penggunaan gen CMS ini mengharuskan perakitan varietas padi hibrida menggunakan tiga galur, yaitu galur mandul jantan (GMJ) atau CMS (galur A), galur pelestari atau maintainer (galur B), dan galur pemulih kesuburan atau restorer (galur R) (Yuan et al. 1994; Virmani 1994).
Galur mandul jantan yang digunakan sebagai tetua betina, memiliki sitoplasma mandul dan gen inti pemulih kesuburan resesif (gen rf) yang tidak berfungsi sehingga galur ini akan tetap mandul jantan. Galur B mempunyai gen inti resesif namun sitoplasma normal sehingga tanaman bersifat fertil (dapat membentuk biji) dan dapat digunakan sebagai polinator untuk perbanyakan benih GMJ. Galur R memiliki gen inti untuk pemulihan kesuburan dominan (gen Rf) dan sitoplasma normal/mandul. Apabila galur GMJ disilangkan dengan galur R maka akan terbentuk F1 hibrida dengan konstitusi genetik gen sitoplasma
mewarisi induknya (steril) namun gen inti bersifat heterozigot (Rfrf) sehingga tanamannya bersifat fertil (Scnable & Wise 1998). Hubungan ketiga galur (A, B, dan R) dapat dilihat pada Gambar 2.
Mekanisme Mandul Jantan dan Pemulihan Kesuburan
Male sterile (mandul jantan) merupakan fenomena gagal berfungsinya kepala sari (anter) atau serbuk sari (polen) pada proses pembuahan. Beberapa tanaman memanfaatkan mekanisme ini untuk produksi benih hibrida secara komersial, antara lain pada jagung, tomat, sorgum, dan padi. Karakter ini diturunkan secara maternal dan berkaitan erat dengan ketidaknormalan open reading frame (ORF) pada genom mitokondria (Hanson & Bentolila 2004) sehingga menyebabkan gangguan fertilitas. Sterilitas yang terjadi ini dapat dipulihkan oleh gen Rf (restorer of fertility gene) dalam inti sel yang berfungsi untuk memulihkan kesuburan. Sistem CMS/Rf terjadi karena adanya interaksi antara genom inti dan mitokondria.
Menurut Yuan et al. (2003), berdasarkan perilaku genetik dari gen Ms galur mandul jantan padi diklasifikasikan menjadi tipe sporofitik dan gametofitik. Pada mandul jantan tipe sporofitik, sterilitas atau fertilitas serbuk sari ditentukan oleh genotipe sporofit. Apabila genotipe sporofitiknya S (rfrf), maka semua serbuk sari akan mengalami aborsi. Sebaliknya, jika genotipenya N (RfRf) atau S (RfRf), mikrospora. Pada GMJ tipe gametofitik, fertilitas GMJ gametofitik ditentukan oleh genotipe gametofit (serbuk sari) saja. Gen inti, Rf, dan rf pada gametofit, berturut-turut akan menentukan fertilitas dan sterilitas serbuk sari. Pengguguran serbuk sari biasanya terjadi pada fase akhir perkembangan mikrospora. Termasuk pada tipe ini adalah GMJ Kalinga. Malai dari galur tipe ini biasanya hanya sedikit tertutup oleh pelepah daun bendera atau bahkan tidak tertutup sama sekali (Satoto & Rumanti 2011).
Tetua dengan sitoplasma steril dan gen rfrf digunakan sebagai tetua betina sedangkan gen Rf berkontribusi sebagai tetua jantan.
Toleransi Kekeringan
Salah satu kendala produksi padi lahan sawah tadah hujan adalah ancaman cekaman kekeringan (Mackill et al. l996). Penggunaan varietas toleran cekaman kekeringan dapat mengurangi ketidastabilan hasil padi sawah tadah hujan (Curtois et al. 2008; Babu et al. l996). Selain itu varietas unggul padi yang adaptif pada kondisi cekaman kekeringan menjadi salah satu komponen teknologi yang penting untuk mengantisipasi dampak pemanasan global khususnya lahan-lahan rawan cekaman kekeringan.
Pengembangan galur-galur toleran kekeringan di Indonesia saat ini masih terfokus pada padi inbrida. Susanto et al. (2012) mengidentifikasi enam galur inbrida yang ditanam pada lahan sawah irigasi tercekam kekeringan dengan hasil lebih tinggi dibanding Inpari 10 dan Situ Bagendit. Varietas gogo hasil rakitan pemulia BB Padi yang toleran terhadap cekaman kekeringan untuk lahan tadah hujan antara lain: Limboto, Batutugi, Situ Bagendit, Inpago 4, Inpago5, dan Inpago 6. Varietas padi sawah inbrida yang toleran kekeringan antara lain: Dodokan, Silugonggo, dan Inpari 10 Laeya (BB Padi 2014).
Beberapa faktor yang menyebabkan penurunan produksi gabah padi akibat cekaman kekeringan antara lain: terhambatnya pertumbuhan daun dan anakan produktif (Kramer & Boyer 1995) serta menurunnya tingkat fotosintesis dan area daun akibat senesen yang terjadi lebih awal (Nooden 1988). Cekaman kekeringan dapat mengakibatkan proses sintesis karbohidrat turun secara perlahan dan metabolisme sink menjadi terhambat pada saat fase reproduksi sehingga diikuti dengan menurunnya fertilitas spikelet (Rahman et al. 2002). Penurunan potensial air di luar sel memproduksi larutan di dalam sel yang lebih rendah tekanan osmotiknya sehingga menyebabkan turgor turun. Fenotipe yang terlihat jelas pada tanaman adalah penggulungan daun (Ji et al. 2012). Menurut Clarke (1986), penggulungan daun merupakan salah satu respon tanaman pada saat air di sekitarnya mengalami defisit, hal ini adalah mekanisme alami untuk mengurangi transpirasi secara berlebihan. Pada kondisi cekaman kekeringan pertumbuhan akar pada tanaman yang mampu beradaptasi akan terlihat lebih panjang menembus ke dalam tanah sebagai salah satu respon kurangnya cakupan air (Ji et al. 2012).
(alokasi biomassa, modifikasi anatomi spesifik, mekanisme fisiologi yang lebih rumit) terhadap kekeringan (Pugnaire et al. 2004)
Fenomena heterosis pada padi hibrida salah satunya mencakup peningkatan volume perakaran yang menyebabkan kemampuan penyerapan air dari lapisan tanah yang lebih dalam sehingga tanaman bisa terhindar dari kekeringan. Menurut Fukai et al. (1999), hal tersebut merupakan salah satu mekanisme toleransi tanaman terhadap cekaman kekeringan. Fukai dan Cooper (1995) menyatakan bahwa padi hibrida toleran merupakan padi yang pada kondisi tercekam kekeringan tetap dapat memberikan hasil gabah lebih tinggi dibanding padi inbrida. Sejumlah penelitian menunjukkan bahwa pada kondisi tercekam kekeringan, padi hibrida mampu memberikan hasil lebih tinggi dibanding dengan padi inbrida (Villa et al. 2012; Yaqoob et al. 2012)
Beberapa metode seleksi untuk tanaman toleran cekaman kekeringan telah dilakukan. Breseghello et al. (2008) melakukan seleksi padi secara langsung pada kondisi lahan sawah tadah hujan yang tercekam kekeringan. Seleksi secara tidak langsung dilakukan dengan mengkondisikan laboratorium atau rumah kaca serupa dengan kondisi cekaman kekeringan. Di laboratorium, seleksi umumnya dilakukan dengan menggunakan PEG (polyethylene glycol) baik pada kultur in vitro (Lestari & Mariska 2006) atau fase perkecambahan (Cahyadi et al. 2013). Afa et al. (2012) dengan menggunakan PEG 6000 konsentrasi 25% (setara dengan -0.99 MPa atau -9.9 Bar), berhasil mendapatkan tiga galur hibrida dan satu varietas unggul hibrida yaitu Maro yang menunjukkan toleransi kekeringan setara dengan Limboto. Seleksi secara tidak langsung di rumah kaca, umumnya dilakukan pada fase vegetatif dengan menggunakan media pasir (Efendi 2009), media campuran perlit dan tanah (Praba et al. 2009), atau campuran parafin dan vaselin untuk menguji daya tembus akar (Suardi 2002).
Senyawa PEG telah dikenal secara luas dapat menginduksi terjadinya cekaman terhadap air pada tahap awal pertumbuhan sehingga menyebabkan terjadinya kondisi kekeringan (Badiane et al. 2004). Beberapa penelitian menggunakan PEG untuk skrining benih terhadap kekeringan dengan memperhitungkan indeks kekeringan (Bouslama & Schapaugh 1984). Menurut Ariyanti (2011) pemberian larutan PEG 6000 dengan konsentrasi yang semakin pekat akan menurunkan kecepatan berkecambah dan pertumbuhan kecambah. Penurunan pertumbuhan akar dan tunas disebabkan karena PEG mengikat air sehingga air menjadi tidak tersedia bagi kecambah (Verslues et al. 2006). Berkurangnya air yang masuk ke dalam kecambah menyebabkan kurang sempurnanya proses perkecambahan terutama pada radikula, hal ini dapat menjadi indikator pengujian cekaman kekeringan (Kamil, 1979).
Pemanfaatan Marka Molekuler pada Padi Hibrida
Padi merupakan salah satu tanaman yang berkembang pesat dalam hal penelitian secara molekuler. Hal tersebut didasarkan pada ukuran genom padi yang kecil dan relatif sedikit jumlah DNA berulang, bersifat diploid, dan kemudahan dalam pengkulturan jaringan. Sejak dekade 1990, penelitian molekuler padi semakin banyak dilakukan (Akhmadikhah et al. 2007). Teknik molekuler telah banyak diterapkan pada pemuliaan padi hibrida antara lain: studi pemulihan kesuburan pada GMJ dengan sumber sitoplasma berbeda, mengidentifikasi lokus yang bertanggungjawab terhadap fertilitas, dan marka molekuler yang terpaut dengan gen Rf, mengetahui keragaman genetik galur-galur tetua, dan memanfaatkan marker assisted selection untuk mempercepat proses perakitan padi hibrida (Jing et al. 2001; Xu 2002; Sattari et al. 2008; Ghara et al. 2012). Selain itu, Kumar et al. (2012) memanfaatkan teknik molekuler untuk mengidentifikasi dan monitoring kemurnian benih padi hibrida.
Penanda morfologi digunakan untuk mengindentifikasi galur mandul jantan, pelestari, atau pemulih kesuburan seperti bentuk gabah, sterilitas atau fertilitas serbuk sari, bentuk tanaman dan sebagainya (Shah et al. 2012). Kelemahan penanda morfologi, seperti rendahnya kontrol genetik dan besarnya pengaruh lingkungan terhadap ekspresi fenotipik menyebabkan ketidakstabilan hasil pada analisis keragaman. Oleh karena itu, marka molekuler dapat dimanfaatkan untuk perakitan tetua padi hibrida.
Penemuan gen-gen yang mengendalikan karakter penting pada tanaman dapat digunakan sebagai marker assisted selection (MAS), meningkatkan perakitan varietas unggul baru, dan menemukan alel-alel baru pada plasma nutfah. Salah satu kemajuan dalam teknik molekuler adalah pengembangan marka SSR atau mikrosatelit (McCouch et al. 1997). Marka ini polimorfik sehingga dapat digunakan untuk mendeteksi genotipe yang dekat kekerabatannya dan mudah dideteksi. Mikrosatelit atau SSR (simple sequens repeat) merupakan tandem arrays dari 2-5 pasangan basa nukleotida berulang. Penanda atau marka SSR merupakan penanda genetik yang teruji karena dapat mendeteksi keanekaragaman alel tingkat tinggi, bersifat ko-dominan, berbasis reaksi PCR (polymerase chain reaction), mudah diaplikasikan dan ekonomis (Jain et al. 2004). Kelebihan utama dari teknik ini yaitu ketelitian dalam pembacaan fragmen DNA (sampai 1 bp) dan dalam sekali pendeteksian dapat diproses marka yang berbeda ukuran fragmen DNA (Santoso et al. 2006).
Marka SSR telah digunakan untuk mempelajari keragaman genetik dan pemetaan gen pemulih kesuburan (Ahmadikhah et al. 2007; Jing et al. 2001; Bazrkar et al. 2008; Sheeba et al. 2006; Sattari et al. 2008; Shah et al. 2012). Selain itu, marka SSR juga telah digunakan untuk membantu pengelompokan tetua hibrida. Pengelompokan menggunakan marka SSR terbukti sama dengan pengelompokan berdasar informasi silsilah tetua (Xangsayasane et al. 2010; Xu et al. 2002). Tasliah et al. (2004), Zainal dan Bahagiawati (2005), dan Victoria et al. (2007) melaporkan marka SSR dan RAPD mampu mendeteksi jarak genetik pada padi selain menggunakan karakter morfologi.
pada studi pengelompokan untuk menentukan jarak genetik antara lain: panjang malai, bobot malai jumlah gabah/malai, persentase pengisian gabah, dan hasil gabah (Kulsum et al. 2011; Hasan et al. 2013). Proses identifikasi, evaluasi, dan karakterisasi genotipe padi baik secara morfologi maupun level molekuler untuk mengetahui dasar perbedaan genetik merupakan hal yang penting dalam perakitan varietas baru (Ogunbayo et al. 2005).
3 IDENTIFIKASI TOLERANSI KEKERINGAN TETUA DAN
F
1PADI HIBRIDA PADA FASE PERKECAMBAHAN
MENGGUNAKAN POLIETILEN GLIKOL (PEG) 6000
KONSENTRASI 25%
Abstrak
Perakitan padi hibrida toleran cekaman kekeringan memerlukan tetua yang memiliki toleransi terhadap kekeringan. Polietilen glikol (PEG) telah banyak digunakan sebagai larutan osmotik untuk mendeteksi toleransi kekeringan pada fase perkecambahan padi. Penelitian ini bertujuan untuk mengevaluasi beberapa variabel toleransi terhadap simulasi cekaman kekeringan selama fase perkecambahan dan menyeleksi galur-galur tetua serta F1 padi hibrida toleran
cekaman kekeringan melalui uji cepat PEG 6000 pada konsentrasi 25%. Percobaan dilakukan di rumah kaca BB Biogen Cimanggu Bogor, menggunakan rancangan kelompok lengkap teracak (RKLT) 2 faktor 3 ulangan dengan faktor pertama adalah genotipe padi sedangkan faktor kedua adalah PEG 6000 2 taraf , yaitu konsentrasi 0 dan 25%. Analisis komponen utama menunjukkan variabel utama untuk skrining toleransi kekeringan yaitu persen perkecambahan, indeks vigor benih, panjang akar seminal, dan panjang kecambah. Berdasarkan keempat variabel tersebut, teridentifikasi galur tetua padi hibrida yaitu galur B (IR 58025B, IR 80154B, GMJ 14B, dan GMJ 15B) dan galur R (PK 90, R 3, dan PK 12) sebagai genotipe toleran kekeringan. Dua belas hibrida yang diduga toleran simulasi cekaman kekeringan fase perkecambahan adalah IR 58025A/R 32, IR 80154A/PK 90, IR 80154A/ R3, IR 80154A/PK 12, IR 80154A/BP 11, IR 80156A/BP 11, GMJ 13A/PK 90, GMJ 13A/R 3, GMJ 13A/BP 11, GMJ 14A/R 3, GMJ 14A/BP 11, dan GMJ 15A/PK 90.
Abstract
Development of drought tolerant hybrid rice varieties requires parental lines which have tolerance to drought. Polyethylene glycol (PEG) has been widely used as an osmotic solution for detecting drought tolerant genotypes at germination stage of rice. The objectives of this experiment were to evaluate variables of drought tolerance at germination stage and to select parental lines and F1 hybrids tolerant to drought using osmotic solution PEG 6000 at
concentration of 25%. The experiment was conducted at the greenhouse of ICABIOGRAD Cimanggu Bogor. The experiment was arranged in factorial randomized block design with three replications. The first factor was genotype and the second factor (osmotic potential) had two levels (0 and 25% concentration levels of PEG 6000). Based on the principal component analysis, four primary indicators were proposed for drought tolerance of rice at germination stage i.e. germination percentage, seed vigor index, seminal root length, and seedling length. Based on coefficient of drought tolerance, the genotypes identified as tolerance to drought were IR 58025B, IR 80154B, GMJ 14B, and GMJ 15B (maintainer lines), PK 90, R 3, and PK 12 (restorer lines). Further, twelve F1 hybrids identified as tolerant to drought were IR 58025A/R 32,
IR 80154A/PK 90, IR 80154A/ R3, IR 80154A/PK 12, IR 80154A/BP 11, IR 80156A/BP 11, GMJ 13A/PK 90, GMJ 13A/R 3, GMJ 13A/BP 11, GMJ 14A/R 3, GMJ 14A/BP 11, and GMJ 15A/PK 90.
Key words: drought tolerant, germination, hybrid rice, PEG
Pendahuluan
Kekeringan merupakan salah satu cekaman abiotik yang dapat menyebabkan penurunan hasil dan kualitas gabah padi (Pandey & Bhandari 2008). Kehilangan hasil disebabkan oleh cekaman kekeringan diperkirakan mencapai 58% (Ouk et al. 2006). Oleh karena itu, menjadi tantangan bagi pemulia untuk merakit varietas padi toleran cekaman kekeringan sekaligus memiliki potensi hasil tinggi.
Teknologi padi hibrida merupakan alternatif untuk mencapai potensi hasil tinggi pada kondisi optimum, namun demikian penelitian pemanfaatan padi hibrida pada kondisi non optimum terutama kekeringan belum banyak dilakukan (Xie 2010). Di Indonesia, program perakitan padi hibrida menggunakan tiga galur yaitu galur mandul jantan (GMJ), galur maintainer (B), dan galur pemulih kesuburan (R). Walaupun hasil penelitian Bueno et al. (2010) menunjukkan ketidakkonsistenan toleransi cekaman kekeringan hibrida dengan tetuanya, namun identifikasi galur tetua toleran merupakan hal penting untuk perakitan padi hibrida toleran kekeringan. Sebagai contoh, Luo (2010) di China yang mengembangkan padi hibrida melalui perakitan GMJ toleran kekeringan.
Teknik penapisan yang cepat dan akurat untuk toleransi terhadap kekeringan merupakan tahapan pemuliaan yang penting (Boopathi et al. 2013). Salah satunya dengan mensimulasi kondisi cekaman kekeringan di laboratorium menggunakan media osmotik polietilen glikol (PEG) 6000. PEG 6000 merupakan zat kimia inert dan non toksik dengan bobot molekul tinggi. Pada konsentrasi tertentu, PEG 6000 dapat menginduksi kondisi kekurangan air sebagaimana yang terjadi pada tanah kering (Mirbahar et al. 2013). Penggunaan PEG 6000 untuk mengidentifikasi toleransi kekeringan telah banyak dilakukan pada tanaman pangan seperti padi, gandum, jagung, dan kedelai (Afa et al. 2013; Govindaraj et al. 2010; Hamayun et al. 2010). Pada saat kondisi kapasitas lapang, tanah mempunyai potensial osmotik -0.33 bar sedangkan pada kondisi titik kelembaban kritis mencapai potensial osmotik -15 bar. Berbagai penelitian melaporkan bahwa penggunaan PEG 6000 dengan konsentrasi 20-25% setara dengan -6.7 sampai -9.9 bar mampu membedakan genotipe padi yang toleran maupun peka cekaman kekeringan (Lestari & Mariska 2006; Meutia et al. 2010; Afa et al. 2012).
Fase perkecambahan dan pertumbuhan bibit merupakan fase potensial dan kritis terhadap cekaman kekeringan (Ahmad et al. 2009). Skrining pada fase ini diharapkan dapat mereduksi materi genetik untuk mempercepat proses perakitan. Sejumlah studi respon tanaman padi terhadap cekaman kekeringan pada fase perkecambahan telah banyak dilakukan (Jiang & Lafitte 2007; Ballo et al. 2012). Mereka mengidentifikasi beberapa variabel perkecambahan yang dapat digunakan sebagai indikator toleransi kekeringan seperti: persentase perkecambahan, panjang akar seminal, panjang tunas, rasio panjang akar seminal/tunas, dan indeks vigor.
Penelitian ini bertujuan untuk: (1) mengevaluasi beberapa variabel toleransi terhadap cekaman kekeringan pada fase perkecambahan padi; (2) menyeleksi galur tetua dan F1 padi hibrida toleran cekaman kekeringan melalui uji cepat
Bahan dan Metode
I. Seleksi Toleran Kekeringan Tetua Padi Hibrida Menggunakan
PEG 6000 Konsentrasi 25 % pada Fase Perkecambahan
Waktu dan Tempat
Percobaan simulasi toleransi kekeringan pada tetua padi hibrida dilakukan pada bulan April-Mei 2014 di rumah kaca, Balai Besar Penelitian dan Pengembangan Bioteknologi dan Sumber Daya Genetik Pertanian di Cimanggu, Bogor.
Bahan Penelitian
Materi yang digunakan adalah tetua padi hibrida meliputi 6 galur maintainer (B), 10 galur restorer (R), 3 varietas pembanding toleran cekaman kekeringan yaitu Gajah Mungkur, Salumpikit (Suardi 2000), dan Limboto serta 2 varietas pembanding peka cekaman kekeringan yaitu IR 20 dan IR 64 (Afa et al. 2012). Total genotipe yang akan diskrining adalah 21 (Tabel 1).
Tabel 1 Materi genetik skrining tetua padi hibrida menggunakan PEG 6000 konsentrasi 25% pada fase perkecambahan
No Genotipe Keterangan No Genotipe Keterangan
1. IR 58025B Galur B 12. PK 21 Galur R
Percobaan menggunakan rancangan kelompok lengkap teracak (RKLT) 2 faktor yaitu faktor pertama adalah 21 genotipe padi dan faktor kedua adalah konsentrasi PEG (BM 6000) yang dibedakan menjadi 2 taraf, yaitu 0 (tanpa perlakuan PEG) dan 25% yang setara -9.9 bar (Michel & Kaufmann 1973). Penentuan konsentrasi 25% didasarkan skrining padi hibrida yang dilakukan Afa et al. (2012). Percobaan diulang 3 kali sehingga terdapat 126 satuan percobaan. Unit percobaan ialah satu cawan petri yang berisi 20 kecambah.
sebagai kontrol dengan pemberian 10 mL tiap cawan petri sebagai media perkecambahan. Perlakuan diberikan selama 7 hari.
Pengamatan untuk setiap variabel percobaan dilakukan dengan cara sebagai berikut:
1. Persentase perkecambahan (PP), pengamatan dilakukan terhadap jumlah kecambah normal pada hari ke-7 (Scott et al. 1984) yaitu:
PP = Jumlah kecambah normal pada hari ke-7 x 100% Jumlah benih yang dikecambahkan
2. Panjang akar seminal (PAS), diukur mulai dari ujung akar hingga pangkal akar dengan satuan centimeter (cm) pada pengamatan hari ke-7
3. Panjang tunas (PT), diukur mulai dari ujung tunas hingga pangkal tunas dengan satuan centimeter (cm) pada pengamatan hari ke-7
4. Panjang kecambah (PK), diukur mulai ujung tunas sampai dengan pangkal akar dengan satuan centimeter (cm) pada pengamatan hari ke-7
5. Bobot kering akar seminal (BKAS), dihitung dengan menimbang bagian akar kecambah umur 7 hari yang telah dikeringkan dengan oven pada suhu konstan 60 0C selama 3 hari, satuan yang digunakan adalah gram (g)
6. Bobot kering tunas (BKT), dihitung dengan menimbang bagian tunas kecambah umur 7 hari yang telah dikeringkan dengan oven pada suhu konstan 60 0C selama 3 hari, satuan yang digunakan adalah gram (g)
7. Indeks vigor benih (IVB), dihitung menggunakan rumus Khodarahmpour (2011) yaitu:
IVB = (Panjang akar seminal + Panjang tunas) x Persentase perkecambahan
Analisis Data
Respon 21 genotipe padi terhadap simulasi cekaman kekeringan dihitung menggunakan nilai indeks penurunan rata-rata (Jiang & Lafitte 2007) sebagai berikut:
Penurunan rata-rata (%) = 1- Variabel x kondisi tercekam x 100% Variabel x kondisi kontrol
Data hasil pengamatan dianalisis dengan analisis ragam (uji F) sesuai rancangan percobaan yang digunakan. Analisis ragam dihitung menggunakan nilai tingkat penurunan (TP) variabel perkecambahan (Xie et al. 2013) yaitu:
TP = Nilai variabel x kondisi kontrol – Nilai variabel x kondisi tercekam x 100% Nilai variabel x kondisi kontrol
Apabila analisis ragam menunjukkan pengaruh nyata pada taraf 5% dilanjutkan dengan uji Dunnett untuk membandingkan semua genotipe dengan semua varietas pembanding toleran dan peka.
Perbedaan kemampuan toleransi genotipe pada kondisi tercekam dihitung dengan koefisien toleransi kekeringan (DC-drought resistance coefficient) untuk masing-masing variabel (Qi et al. 2010) yaitu:
Selanjutnya nilai DC digunakan pada analisis korelasi (Pearson) untuk mengetahui hubungan antar variabel perkecambahan. Analisis komponen utama (AKU) digunakan untuk mengidentifikasi variabel yang berkontribusi besar terhadap keragaman (Golmoghani et al. 2011). Berdasarkan nilai DC tiap variabel, AKU dilakukan untuk menghitung nilai vektor ciri dan tingkat kontribusi tiap komponen utama (Xie et al. 2013). Pengklasifikasian genotipe toleran dan peka simulasi cekaman kekeringan berdasarkan nilai rata-rata DC variabel perkecambahan yang teridentifikasi pada Komponen Utama 1.
Software yang digunakan untuk analisis ragam, korelasi, dan komponen utama adalah SAS versi 6.12, Minitab 14, dan Microsoft Excel 2003.
II. Seleksi Toleran Kekeringan pada F1 Padi Hibrida Menggunakan
PEG 6000 Konsentrasi 25% pada Fase Perkecambahan.
Waktu dan Tempat
Percobaan dilakukan pada Februari – Maret 2015 di rumah kaca Balai Besar Penelitian dan Pengembangan Bioteknologi dan Sumber Daya Genetik Pertanian, Cimanggu, Bogor Jawa Barat.
Bahan Penelitian
Materi yang digunakan adalah 30 F1 hibrida turunan dari tetua yang
teridentifikasi toleransinya terhadap simulasi cekaman kekeringan pada kegiatan I, varietas Salumpikit, Gajah Mungkur, Limboto, Maro, dan Hipa 8 sebagai varietas pembanding toleran, serta IR 20 dan IR 64 sebagai varietas pembanding peka cekaman kekeringan (Tabel 2). Pelaksanaan penelitian dan analisis data dilakukan seperti pada kegiatan I.
Tabel 2 Materi genetik skrining F1 padi hibrida menggunakan PEG 6000
konsentrasi 25% pada fase perkecambahan
No Genotipe No Genotipe No Genotipe 1 IR 58025A/PK 90 14 IR 80156A/R 32 27 GMJ 15A/R 3 2 IR 58025A/R 3 15 IR 80156A/BP 11 28 GMJ 15A/PK 12 3 IR 58025A/PK 12 16 GMJ 13A/PK 90 29 GMJ 15A/R 32 4 IR 58025A/R 32 17 GMJ 13A/R 3 30 GMJ 15A/BP 11 5 IR 58025A/BP 11 18 GMJ 13A/PK 12 31 Salumpikit 6 IR 80154A/PK 90 19 GMJ 13A/R 32 32 Gajah Mungkur 7 IR 80154A/R 3 20 GMJ 13A/BP 11 33 Limboto 8 IR 80154A/PK 12 21 GMJ 14A/PK 90 34 Maro 9 IR 80154A/R 32 22 GMJ 14A/R 3 35 Hipa 8 10 IR 80154A/BP 11 23 GMJ 14A/PK 12 36 IR 64 11 IR 80156A/PK 90 24 GMJ 14A/R 32 37 IR 20 12 IR 80156A/R 3 25 GMJ 14A/BP 11
Hasil dan Pembahasan
I. Seleksi Toleran Kekeringan Tetua Padi Hibrida Menggunakan
PEG 6000 Konsentrasi 25 % pada Fase Perkecambahan
Pengaruh PEG 6000 Konsentrasi 25% terhadap Perkecambahan
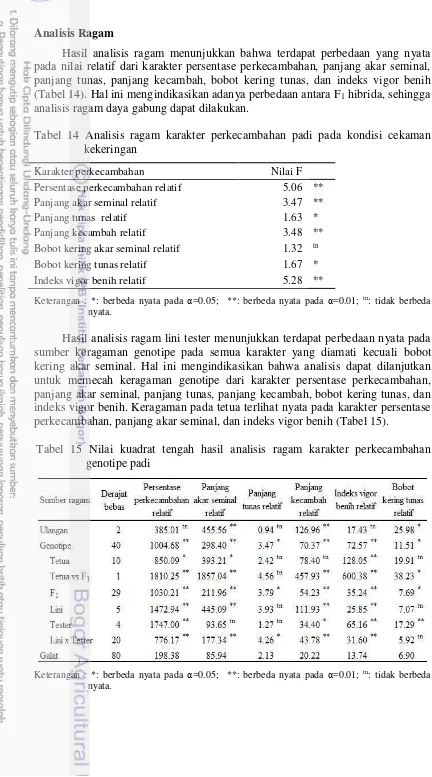
Hasil analisis ragam menunjukkan bahwa genotipe, dan perlakuan taraf PEG berpengaruh nyata terhadap variabel persentase perkecambahan, indeks vigor benih, panjang akar seminal, panjang tunas, panjang kecambah, bobot kering akar seminal, dan bobot kering tunas. Begitu pula interaksi genotipe x taraf PEG menunjukkan pengaruh nyata terhadap semua variabel kecuali bobot kering tunas (Tabel 3). Hal ini mengindikasikan bahwa antar genotipe memiliki respon berbeda terhadap perlakuan PEG 6000 konsentrasi 25% pada beberapa variabel perkecambahan.
Tabel 3 Analisis ragam pada perlakuan genotipe dan PEG 6000 pada variabel perkecambahan 21 genotipe padi
Variabel perkecambahan Genotipe Perlakuan G x P KK (%)
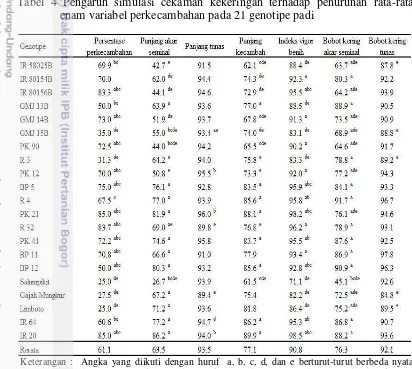
Perlakuan PEG 6000 menghasilkan penurunan persentase perkecambahan paling rendah pada varietas Salumpikit dan Limboto yang merupakan pembanding toleran kekeringan yaitu sebesar 25%. Galur GMJ 15B dan R 3 menunjukkan penurunan persentase perkecambahan yang tidak berbeda nyata dengan Salumpikit, Gajah Mungkur, dan Limboto. Sementara itu, sepuluh galur tetua padi hibrida lainnya menunjukkan respon penurunan persentase perkecambahan yang tidak berbeda nyata dengan varietas IR 20 yang merupakan pembanding peka kekeringan (Tabel 4). Menurut Jatoi et al. (2014), kenaikan tingkat osmotik media menyebabkan jumlah air yang diabsorbsi benih rendah sehingga mengakibatkan turunnya persentase perkecambahan.
Tabel 4 Pengaruh simulasi cekaman kekeringan terhadap penurunan rata-rata enam variabel perkecambahan pada 21 genotipe padi
Keterangan : Angka yang diikuti dengan huruf a, b, c, d, dan e berturut-turut berbeda nyata dengan Salumpikit, Gajah Mungkur, Limboto, IR 64, dan IR 20 berdasarkan uji Dunnett taraf 0.05.
lain, artinya pembanding toleran kekeringan ini tetap mampu menghasilkan akar panjang pada kondisi cekaman PEG. Galur GMJ 15B dan PK 90 merupakan genotipe yang memiliki respon setara dengan Salumpikit, sedangkan genotipe IR 58025B, IR 80154B, IR 80156B, GMJ 14B, R 3, dan PK 12 tidak berbeda nyata dengan Gajah Mungkur dan Limboto yang juga merupakan pembanding toleran kekeringan tetapi berbeda nyata dengan Salumpikit (Tabel 4).
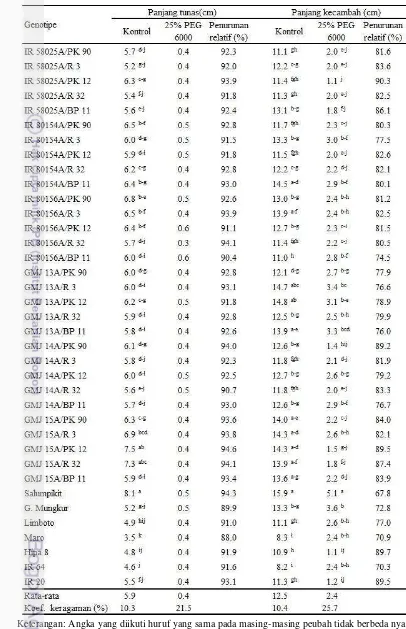
Persentase penurunan panjang tunas terrendah adalah R 32 yang tidak berbeda nyata dibandingkan dengan Gajah Mungkur yaitu sebesar 89.8%. Sebagian besar genotipe yang diuji menunjukkan persentase penurunan panjang tunas yang tinggi termasuk Salumpikit dan Limboto yang merupakan pembanding toleran kekeringan (Tabel 4). Menurut Zapico et al. (2008) pada kondisi kekurangan air pertumbuhan tunas lebih terhambat dibandingkan akar seminal hal ini disebabkan pasokan karbohidrat yang lebih banyak ditujukan ke bagian akar dibanding tunas pada fase perkecambahan. Persentase penurunan panjang kecambah yang tidak berbeda nyata dengan Salumpikit ditunjukkan oleh genotipe IR 58025B, GMJ 14B, dan PK 90 yaitu masing-masing sebesar 62.1, 67.8, dan 65.5% (Tabel 4).
Perlakuan PEG 6000 konsentrasi 25% menghasilkan persentase penurunan indeks vigor benih paling rendah pada varietas pembanding Salumpikit yaitu sebesar 71.1%, diikuti oleh Gajah Mungkur dan Limboto masing-masing sebesar 82.2% dan 86.4%. Galur IR 58025B, GMJ 13B, GMJ 15B, dan R 3 merupakan genotipe yang memiliki persentase penurunan vigor benih yang tidak berbeda nyata dengan ketiga varietas pembanding toleran tersebut. Sebagian besar genotipe turun drastis indeks vigornya akibat cekaman PEG. Rendahnya indeks vigor merupakan akibat pertumbuhan akar dan tunas yang terhambat.
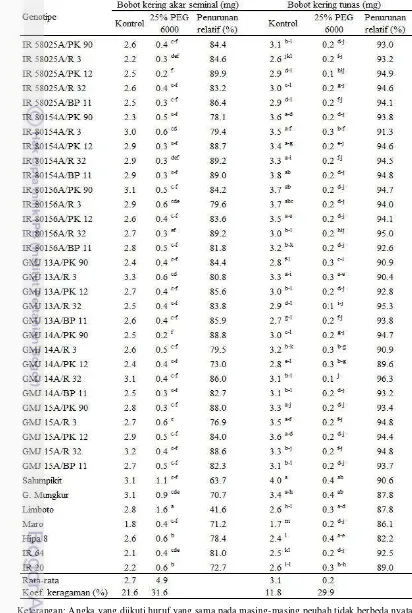
Akibat terjadinya penurunan panjang akar seminal mengakibatkan turunnya bobot kering akar. Salumpikit merupakan varietas pembanding paling rendah persentase penurunan bobot kering akar seminalnya yaitu 45.1%. Bobot kering akar seminal yang persentase penurunannya tidak berbeda nyata dengan Gajah Mungkur dan Limboto adalah IR 58025B, IR 80156B, GMJ 14B, GMJ 15B, PK 90, PK 12, dan PK 21 (Tabel 4). Bobot kering tunas relatif seragam, genotipe IR 58025B, GMJ 15B, dan R 3 menunjukkan persentase penurunan yang berbeda nyata dengan IR 20 sebagai varietas pembanding peka kekeringan (Tabel 4).
Secara umum pada Tabel 4 terlihat keragaman respon genotipe pada beberapa variabel perkecambahan. Beberapa peneliti melaporkan bahwa perbedaan toleransi tanaman terhadap cekaman kekeringan antara lain disebabkan oleh persentase perkecambahan, panjang akar seminal, panjang tunas, bobot kering tunas, bobot kering akar seminal, dan vigor benih (Ballo et al. 2012; Daksa et al. 2014; Widoretno 2011). Selain faktor ketersediaan air pada media, faktor genetik juga berpengaruh terhadap respon perkecambahan (Molphe-Balch et al. 1996).
Analisis Korelasi
menunjukkan beberapa karakter penting saling berhubungan antar variabel perkecambahan. Variabel yang memberikan kontribusi besar terhadap keragaman dan saling berkorelasi dapat digunakan sebagai indikator dalam mengelompokkan genotipe toleran kekeringan (Afa et al. 2013). Pada penelitian ini teridentifikasi persentase perkecambahan memiliki korelasi positif dan signifikan dengan indeks vigor benih, panjang akar seminal, panjang tunas, panjang kecambah, bobot akar seminal dan bobot kering tunas (Tabel 5).
Tabel 5 Koefisien korelasi variabel perkecambahan padi
Variabel X1 X2 X3 X4 X5 X6
X2 0.81 **
X3 0.43 * 0.84 **
X4 0.60 * 0.46 tn 0.06 tn
X5 0.55 * 0.91 ** 0.97 ** 0.29 tn
X6 0.79 ** 0.67 ** 0.38 tn 0.59 * 0.51 *
X7 0.58 * 0.45 tn 0.17 tn 0.79 ** 0.36 tn 0.74 ** Keterangan: Nilai relatif untuk variabel X1=persentase perkecambahan; X2=indeks vigor benih;
X3=panjang akar seminal; X4=panjang tunas; X5=panjang kecambah; X6= bobot
kering akar seminal; X7= bobot kering tunas * = nyata pada α= 0.05; ** = sangat
nyata pada α = 0.01.
Analisis Komponen Utama
Pemilihan variabel untuk menentukan toleransi suatu genotipe terhadap cekaman kekeringan sebaiknya dilakukan secara komprehensif, karena penentuan tingkat toleransi hanya berdasar satu variabel akan menghasilkan penilaian yang tidak valid (Zhang & Wang 2012). Analisis komponen utama dapat digunakan untuk menganalisis multi-variabel penentu toleransi kekeringan (Golmoghani et al. 2011). Metode ini dapat digunakan untuk memilih faktor yang penting sebagai acuan toleransi kekeringan sehingga dapat mencerminkan kondisi genotipe sesungguhnya (Reza et al. 2011; Bibi et al. 2012).
Tabel 6 Analisis komponen utama terhadap 8 variabel perkecambahan padi
Variabel Komponen 1 Komponen 2
Persentase perkecambahan 0.96 * 0.26
Indeks vigor benih 0.98 * 0.15
Panjang akar seminal 0.99 * 0.04
Panjang tunas 0.23 0.99 *
Panjang kecambah 0.96 * 0.26
Bobot kering akar seminal relatif 0.02 0.99 *
Bobot kering tunas relatif 0.41 0.90 *
Nilai eigen 5.45 2.41
Tingkat kontribusi (%) 68.09 30.14
Akumulasi tingkat kontribusi (%) 68.09 98.22
Keterangan: * Nilai maksimum tiap variabel dari dua komponen utama.
Keragaan beberapa galur tetua padi hibrida yang mengalami simulasi cekaman kekeringan pada fase perkecambahan dapat dilihat pada Gambar 3. Terlihat perbedaan yang jelas antara genotipe yang toleran cekaman PEG dengan genotipe yang peka adalah pada pertumbuhan akar seminalnya.
Gambar 3 Keragaan kecambah tetua padi hibrida pada kondisi cekaman menggunakan PEG 6000 konsentrasi 25%.
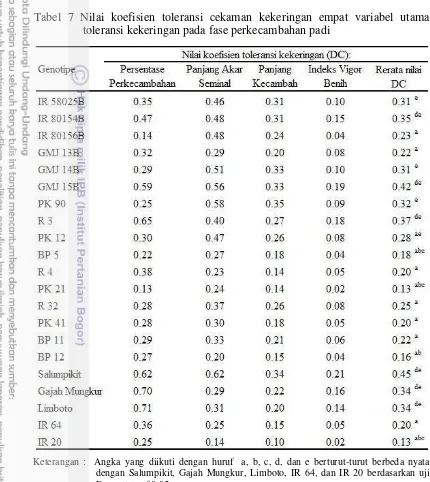
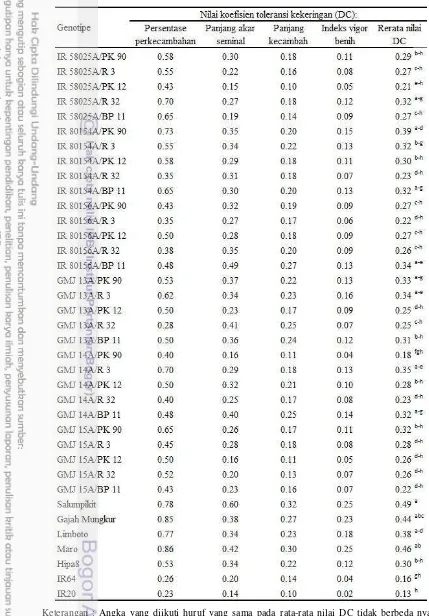
Berdasarkan nilai rata-rata koefisien DC dari empat indikator toleransi cekaman kekeringan (Tabel 7), GMJ yang menunjukkan rata-rata nilai DC tidak
berbeda nyata dengan Salumpikit, Gajah Mungkur, dan Limboto adalah IR 58025B, IR 80154B, GMJ 14B, dan GMJ 15B. Galur R yang memiliki
80154B, GMJ14B, GMJ 15B, PK 90, R 3, dan PK 12 diduga toleran cekaman PEG 6000 konsentrasi 25%, sedangkan genotipe dengan rata-rata nilai DC tidak berbeda nyata dengan IR 64 dan atau IR 20 diduga peka terhadap simulasi cekaman kekeringan fase perkecambahan yaitu IR 80156B, GMJ 13B (galur mandul jantan), BP 5, R 14, PK 21, R 32, PK 41, BP 11, dan BP 12 (Tabel 7). Tabel 7 Nilai koefisien toleransi cekaman kekeringan empat variabel utama
toleransi kekeringan pada fase perkecambahan padi
Keterangan : Angka yang diikuti dengan huruf a, b, c, d, dan e berturut-turut berbeda nyata dengan Salumpikit, Gajah Mungkur, Limboto, IR 64, dan IR 20 berdasarkan uji Dunnett taraf 0.05.
Selanjutnya galur mandul jantan baik yang toleran maupun peka cekaman PEG 6000 konsentrasi 25% tersebut disilangkan dengan lima galur R yang terdiri 3 genotipe toleran (PK 90, R 3, dan PK 12) dan 2 galur R peka simulasi cekaman kekeringan (R 32 dan BP 11) untuk mendapatkan benih F1 hibrida sebagai materi
II. Seleksi Toleran Kekeringan F1 Padi Hibrida Menggunakan PEG 6000 Konsentrasi 25 % pada Fase Perkecambahan
Hasil analisis ragam konsisten dengan kegiatan I, menunjukkan bahwa genotipe, taraf PEG, dan interaksi antara genotipe dan taraf PEG berpengaruh nyata terhadap variabel persentase perkecambahan, indeks vigor benih, panjang akar seminal, panjang tunas, panjang kecambah, bobot kering akar seminal, dan bobot kering tunas.
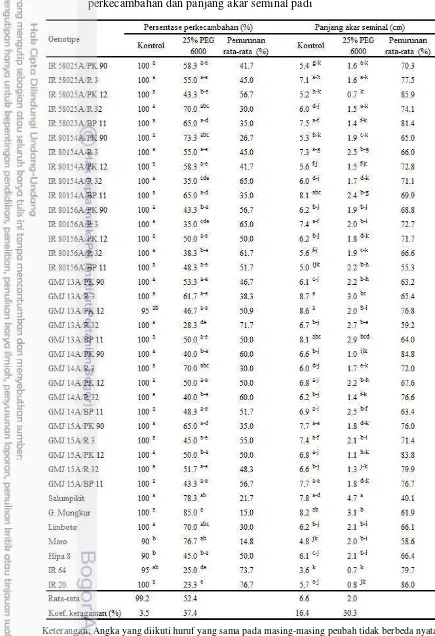
Perlakuan 25% PEG 6000 mempengaruhi persentase perkecambahan. Pada kondisi normal, seluruh genotipe yang diuji menunjukkan persentase perkecambahan yang normal dengan kisaran 90 – 100%. Rata-rata persentase perkecambahan varietas pembanding toleran pada kondisi tercekam kekeringan lebih tinggi dibanding varietas peka. Persentase perkecambahan tertinggi akibat perlakuan PEG 6000 diperoleh pada varietas pembanding Gajah Mungkur (85.0%), diikuti oleh Salumpikit, Maro, dan Limboto dengan kisaran antara 70.0 – 78.3%. Tiga hibrida IR 58025A/R 32, IR 80154A/PK 90, dan GMJ 14A/R 3 memiliki persentase perkecambahan yang tidak berbeda nyata dibandingkan Gajah Mungkur, Salumpikit, Maro, dan Limboto (varietas toleran kekeringan) yaitu berturut-turut masing-masing 70, 73.3, dan 70% dengan persentase penurunan masing-masing sebesar 30, 26.7, dan 30%. Varietas pembanding peka kekeringan yaitu IR 64 dan IR 20 masing-masing mengalami penurunan persentase perkecambahan sebesar 73.7 dan 76.7%, sedangkan Hipa 8 menunjukkan penurunan persentase perkecambahan sebesar 50% (Tabel 8)
Variabel panjang akar seminal pada kondisi kontrol tidak menunjukkan perbedaan secara signifikan, sebagian besar hibrida menampilkan akar yang lebih panjang dibanding varietas pembanding peka IR 64 dan IR 20. Akar seminal terpanjang akibat perlakuan PEG 6000 diperoleh pada genotipe GMJ 13A/R 3 yang tidak berbeda nyata dibandingkan dengan Gajah Mungkur, Limboto, Maro dan Hipa 8, tetapi berbeda nyata dengan Salumpikit. Selain itu teridentifikasi 11 hibrida lainnya berbeda nyata dengan varietas peka kekeringan IR 64 dan IR 20. Rata-rata panjang akar genotipe tersebut berkisar antara 2.0 – 3.0 cm, dengan persentase penurunan akar antara 55.3 – 76.8% sedangkan IR 64 dan IR 20 memiliki panjang akar hanya 0.7 dan 0.8 cm dengan persentase penurunan masing-masing 79.7 dan 86.0% (Tabel 8).
Tabel 8 Pengaruh larutan PEG 6000 konsentrasi 25% terhadap persentase perkecambahan dan panjang akar seminal padi
Tabel 9 Pengaruh larutan PEG 6000 konsentrasi 25% terhadap panjang tunas dan panjang kecambah padi