Nilaparvata lugens STÅL (HEMIPTERA: DELPHACIDAE)
TERHADAP TUJUH VARIETAS TANAMAN PADI
RAHMINI
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI DISERTASI DAN SUMBER INFORMASI
Dengan ini saya menyatakan bahwa disertasi “Respon Biologi Wereng Batang Cokelat Nilaparvata lugens Stål (Hemiptera: Delphacidae) terhadap Tujuh Varietas Tanaman Padi” adalah hasil penelitian saya, dengan arahan dari komisi pembimbing selama mengikuti program S3 di Sekolah Pascasarjana, Institut Pertanian Bogor. Hasil penelitian ini belum pernah dipublikasi dalam bentuk apa pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka yang ada dalam disertasi ini.
Bogor, Juli 2012
RAHMINI. Biological Responses of Brown Planthopper Nilaparvata lugens Stål (Hemiptera: Delphacidae) on Seven Rice Varieties. Under supervision of PURNAMA HIDAYAT, ENDANG SRI RATNA, I WAYAN WINASA, and SYAFRIDA MANUWOTO
Brown planthopper is one of the major pest on rice in Indonesia. Rice resistance is one of the components to control brown planthopper outbreak. The problem arises when brown planthopper could develop into new biotype that could adapt on previous resistant varieties. In developing new resistant varieties, it is necessary to understand the responses of the brown planthopper to the various rice varieties which have different level of resistance. The purposes of the researches were: (1) to study the biological responses of brown planthopper on biophysical factors of rice varieties; and (2) to study the biological responses of brown planthopper on biochemical factors of rice varieties. These studies were conducted in Research Station of the Indonesian Center for Rice Research, Muara-Bogor, the Indonesian Center for Biology and Genetic Research, Bogor, and the Indonesian Institute of Science, Cibinong-Bogor during April 2010-May 2012. Several kinds of biological responses of brown planthopper to rice varieties were studied in terms adult settling preference, feeding activity trough honeydew test, and life performance through life table analysis. Biochemical factors such as sucrose and oxalic acid contents in some rice varieties were analyzed in the Indonesian Center for Biotechnology and Genetics. Microtechnique of rice tissues were prepared in the Indonesian Institute of Science, Cibinong -Bogor. Rice varieties such as TN1 (no resistance gene), IR26 (Bph1), IR42 (bph2), IR64 (Bph1+), IR74 (Bph3), PTB33 (bph2, Bph3) and Inpari13 (unknown resistance gene) were used in this study. Adults showed same non-preference reactions to resistant varieties, included Inpari13 for settling. Physical factors such as thickness of epidermis were different between TN1 and PTB, IR64, but there were no differences among PTB33, IR64, IR74, IR42, IR26 ad Inpari13. Number of trichomes were not different among varieties studied. There were no contribution of epidermis thickness and number of trichomes in rice resistance to brown planthopper. Feeding activity of adult female was highest on susceptible TN1. The difference in feeding activity between biotype 2 and 3 was on IR64. The highest mortality occurred during the immature stages, especially in the first and second instars. The intrinsic rate of increase (rm), net reproductive rate (Ro) of brown planthopper were higher and shorter doubling time (DT) on susceptible variety such as TN1. High oxalic acid and low sucrose in the resistant variety such as PTB33 could play role in rice resistance to brown planthopper.
RAHMINI. Respon Biologi Wereng Batang Cokelat Nilaparvata lugens Stål (Hemiptera: Delphacidae) terhadap Tujuh Varietas Tanaman Padi. Dibimbing oleh PURNAMA HIDAYAT, ENDANG SRI RATNA, I WAYAN WINASA, dan SYAFRIDA MANUWOTO
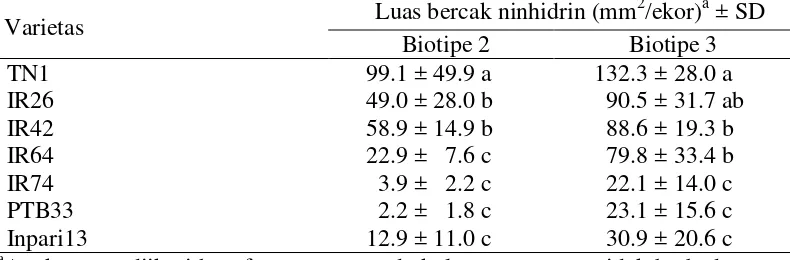
tahan PTB33. Respon tersebut diduga berkaitan erat dengan ukuran ovipositor WBC yang nyata lebih kecil pada WBC biotipe 2 dibandingkan biotipe 3. Sebaliknya, jumlah trikhoma pada permukaan tanaman dan ketebalan jaringan epidermis batang tidak berpengaruh terhadap perilaku hinggap dan peletakan telur. Respon kemampuan makan WBC ditunjukkan dengan luasan bercak embun madu yang nilainya 23.1 dan 2.2 mm2/ekor berturut-turut dihasilkan oleh WBC biotipe 3 dan 2 pada varietas standar tahan PTB33 dibandingkan 132.3 dan 99.1 mm2/ekor pada varietas standar rentan TN1. Kematian WBC sebagian besar terjadi pada individu muda, dan mortalitas rendah pada umur lebih tua. Terjadi penundaan peneluran pada varietas tahan IR74, yaitu puncak peneluran dicapai pada hari ke-25 sebanyak 14,4 butir telur/induk, sedangkan pada varietas inang biotipe 3 yaitu IR42 terjadi pada hari ke-21 sebanyak 16 butir/induk, Kemampuan tumbuh dan berkembang WBC yang lebih tinggi pada TN1 diduga berkaitan erat dengan tingginya kadar sukrosa sebagai sumber nutrisi pakan dan rendahnya kadar asam oksalat sebagai bahan penghambat makan yang terkandung di dalam jaringan tanaman. Semua respon tersebut di atas diekspresikan lebih lanjut pada neraca kehidupan WBC. Pada tanaman standar rentan TN1, WBC memiliki laju pertumbuhan intrinsik dan laju reproduksi bersih yang tinggi, serta waktu penggandaan populasi relatif lebih pendek daripada varietas tahan IR74.
©Hak cipta milik Institut Pertanian Bogor, tahun 2012
Hak cipta dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya utuk kepentigan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan yang wajar IPB
Nilaparvata lugens STÅL (HEMIPTERA: DELPHACIDAE)
TERHADAP TUJUH VARIETAS TANAMAN PADI
RAHMINI
Disertasi
sebagai salah satu syarat untuk memperoleh gelar Doktor pada
Program Studi Entomologi
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Stål (Hemiptera: Delphacidae) terhadap Tujuh Varietas Tanaman Padi
Nama : Rahmini
NIM : A361070041
Program Studi : Entomologi
Disetujui Komisi Pembimbing
Ketua
Dr.Ir. Purnama Hidayat, M.Sc. Dra. Endang Sri Ratna, Ph.D. Anggota
Anggota
Dr.Ir. I Wayan Winasa, M.Si.
Anggota
Prof.Dr.Ir. Syafrida Manuwoto, M.Sc.
Diketahui
Ketua Program Studi Entomologi
Dr.Ir. Pudjianto, M.S.
Dekan Sekolah Pascasarjana IPB
Dr.Ir. Dahrul Syah, M.Sc.Agr.
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah SWT karena dengan rahmat dan hidayah-Nya, maka disertasi yang berjudul “Respon Biologi Wereng Batang Cokelat Nilaparvata lugens Stål (Hemiptera: Delphacidae) terhadap Tujuh Varietas Tanaman Padi” dapat diselesaikan.
Penelitian dan penulisan disertasi dapat terlaksana atas bimbingan dan arahan komisi pembimbing yaitu Dr.Ir. Purnama Hidayat, M.Sc, sebagai Ketua, Dra. Endang Sri Ratna, Ph.D, Dr.Ir. I Wayan Winasa, M.Si, Prof.Dr.Ir. Syafrida Manuwoto, M.Sc, masing-masing sebagai Anggota Komisi Pembimbing, atas semua bimbingan dan arahan tersebut, penulis menyampaikan terima kasih.
Penulis juga menyampaikan terimakasih kepada Kepala Badan Litbang Pertanian, Ketua Komisi Pembinaan Tenaga Badan Litbang Pertanian, Kepala Puslitbangtan, Kepala Balai Besar Penelitian Tanaman Padi, Ketua Kelompok Peneliti Entomologi-Fitopatologi BB Padi yang telah menugaskan dan memberi kesempatan kepada penulis utuk melanjutkan pendidikan, serta Pemimpin dan staf Bendahara Badan Litbang Pertanian yang telah membantu mempermudah penyaluran dana pendidikan penulis. Penulis juga menyampaikan terimakasih kepada Kepala Kebun Percobaan BB Padi di Muara, Bogor atas fasilitas rumah kaca dan laboratorium yang digunakan selama penelitian, juga kepada KKP3T yang telah membiayai penelitian.
Kepada Ketua dan seluruh staf dosen Program Studi Entomologi-Fitopatologi IPB, penulis menyampaikan terimakasih atas ilmu yang telah diberikan, juga kepada Dr.Ir. Kikin Hamzah Mutaqin, M.Si atas bantuan dokumentasi hasil penelitian.
Kepada rekan-rekan Sekolah Pascasarjana Program Studi Entomologi-Fitopatologi, para staf dan rekan mahasiswa di Laboratorium Virologi, Laboratorium Fisiologi dan Toksikologi, serta Laboratorium Taksonomi, Program Studi Entomologi-Fitopatologi Fakultas Pertanian Institut Pertanian Bogor, penulis menyampaikan terimakasih atas saran, bantuan dan kebersamaannya.
Penulis menyampaikan penghargaan dan terimakasih yang tak terhingga kepada Mamah, kakak-kakak, para keponakan atas doa yang tak putus, dorongan dan dukungannya, serta suamiku Kus Adrianto dan ananda Dani atas doa, pengertian dan pengorbanannya selama penulis mengerjakan dan menyeselesaikan pendidikan.
Penulis berharap tulisan ini dapat bermanfaat bagi kita semua.
Bogor, Juli 2012 Rahmini
Penulis dilahirkan di Bandung pada tanggal 18 Nopember 1970 dari ayah Abidin Gandaatmaja (Alm.) dan ibu Jenab. Penulis merupakan putri ketiga dari tiga bersaudara.
Penguji pada Ujian Tertutup: Dr. Pudjianto, M.S. Dr. I Nyoman Widiarta
Halaman
Biologi Wereng Batang Cokelat ………...……… 7
Seleksi Tanaman Inang oleh Wereng Batang Cokelat ……...……. 7
Mekanisme Ketahanan Tanaman ……….……..……... 9
Ketahanan Tanaman Padi Terhadap Wereng Batang Cokelat Sebagai Dasar Pembentukan Perakitan Varietas Padi Tahan ……... 12 III RESPON BIOLOGI WERENG BATANG COKELAT TERHADAP KARAKTER BIOFISIK TANAMAN PADI ………. 17
V PEMBAHASAN UMUM ……….……….... 45
VI KESIMPULAN DAN SARAN ………. 49
DAFTAR PUSTAKA ………..………..……… 51
Halaman 3.1 Preferensi hinggap imago WBC pada berbagai varietas tanaman padi
dan pada waktu yang berbeda setelah dilepaskan ...
23
3.2 Preferensi peletakan telur WBC pada berbagai varietas tanaman padi
25
3.3 Luas ovipositor WBC biotipe 2 dan biotipe 3 berdasarkan morfometri ovipositor ………..…..……….
26
3.4 Jumlah trikhoma dan tebal epidermis pada pelepah daun berbagai varietas tanaman padi ………..
27
4.1 Kemampuan makan WBC pada berbagai varietas tanaman padi melalui uji embun madu ………..………
37
4.2 Nilai laju reproduksi bersih (Ro), periode rata-rata satu generasi (T),
laju pertumbuhan intrinsik (rm), dan waktu penggandaan populasi
(DT) WBC biotipe 3 yang diinfestasi pada berbagai varietas tanaman padi ………..…………
40
Halaman 1.1 Skema faktor-faktor yang mempengaruhi respon WBC terhadap
ketahanan varietas tanaman padi. .………
5
2.1 Perkembangan biotipe dan varietas tahan WBC di Indonesia …….... 14
3.1 Trikhoma pada permukaan pelepah daun varietas Inpari13 berdasarkan irisan membujur (perbesaran 200x) .………...………….
28
4.1 Skema pengujian embun madu ……..……….. 34
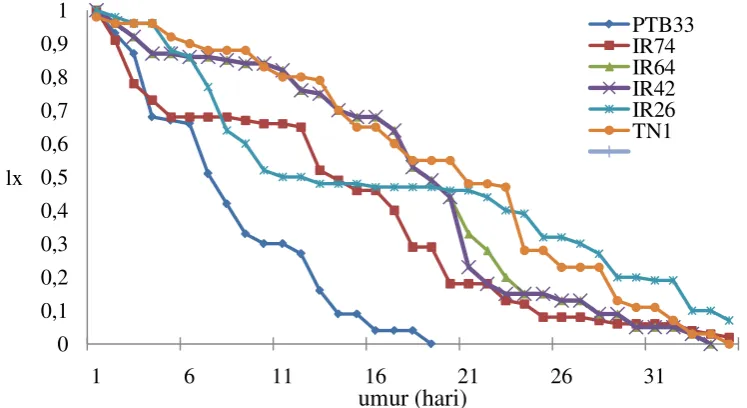
4.2 Kurva kelangsungan hidup (lx) WBC yang diinfestasikan pada berbagai varietas tanaman padi ………....
38
4.3 Peneluran harian WBC pada berbagai varietas tanaman padi (kurva mx) ……….…..
Halaman
Lampiran1. Aktivitas makan wereng cokelat biotipe3 berdasarkan uji ninhidrin pada berbagai varietas ………
55
Lampiran 2. Aktivitas makan wereng cokelat biotipe 2 berdasarkan uji ninhidrin pada berbagai varietas ………
552
I. PENDAHULUAN
Latar BelakangHama tanaman merupakan salah satu kendala yang dapat menurunkan
produktivitas tanaman. Salah satu hama penting pada tanaman padi adalah wereng
batang cokelat (Nilapavarta lugens Stål). Hama ini menyerang berbagai varietas tanaman padi khususnya varietas unggul tipe baru (VUTB), padi hibrida yang
diketahui rentan terhadap serangan organisme pengganggu tanaman, serta padi
varietas unggul baru (VUB) (Baehaki 2008). Adanya kebutuhan akan peningkatan
produksi beras nasional, maka tanaman padi berdaya hasil tinggi dikembangkan
seperti VUTB dan padi hibrida, meskipun padi tipe ini rentan terhadap wereng
batang cokelat (WBC). Saat ini beberapa varietas VUTB berstatus agak tahan
terhadap biotipe 3, sedangkan varietas hibrida berstatus rentan terhadap biotipe 3
(Suprihatno et al. 2010). Berkembangnya populasi WBC hingga terjadinya ledakan populasi, sering dikaitkan dengan penggunaan pestisida yang berlebihan,
introduksi varietas unggul yang rentan terhadap WBC, serta pemupukan yang
tinggi (Widiarta dan Suharto 2009).
Di Indonesia, serangan serangga ini dilaporkan pertama kali pada tahun
1931-1940 (Baehaki dan Widiarta 2009). Hingga saat ini WBC masih merupakan
masalah di pertanaman padi, seiring dengan berkembangnya biotipe WBC
(Munawar dan Baehaki 2008). WBC hidup dengan mengisap cairan floem
tanaman padi, yang mengakibatkan daun berubah warna menjadi kuning oranye
sebelum menjadi cokelat, mengering dan kemudian mati. Kondisi ini, disebut
“hopperburn” karena tanaman terlihat seperti terbakar. Perubahan yang terjadi pada tanaman akibat serangan WBC diantaranya adalah penurunan kandungan air
(Cagampang et al. 1974), penurunan kandungan protein; ditunjukkan dengan perubahan bagian tanaman yaitu daun yang menjadi cokelat, dan mengandung
73% protein lebih rendah dari daun sehat (Sogawa 1971). Penelitian di rumah
kaca menunjukkan bahwa WBC yang diinfestasikan pada tanaman padi
menurunkan penyerapan nutrien oleh akar, terutama fosfor (P) dan potasium (K)
(Wu et al. 2003). Salah satu penyebab hopperburn adalah berkurangnya laju translokasi fotosintat ke perakaran karena mengeringnya cairan floem dan
WBC secara terus-menerus dan eksploitasi penyerapan cairan makanan (Sogawa
1982). Selain menyerang langsung tanaman padi, WBC juga dapat menularkan
penyakit kerdil hampa dan kerdil rumput yang disebabkan oleh virus (Hibino et al. 1977).
WBC mempunyai keragaman genetik yang cukup luas serta relatif mudah
menyesuaikan diri dengan lingkungan. Hama ini cepat beradaptasi dan dapat
mematahkan ketahanan varietas yang semula bereaksi tahan. Penanaman varietas
yang sama secara terus-menerus, pemakaian insektisida yang kurang bijaksana,
dan sanitasi yang kurang baik akan mendorong munculnya biotipe baru WBC
(Harahap et al. 1987, Hanarida 1998, Soewito et al. 1995). Biotipe WBC didefinisikan sebagai suatu populasi serangga yang memiliki reaksi yang sama
terhadap suatu varietas tanaman (Bernays dan Chapman 1994).
Varietas tanaman padi tahan dikembangkan berdasarkan pemanfaatan
ketahanan alami beberapa varietas padi dan padi liar terhadap WBC. Berbagai
ketahanan tanaman padi terhadap WBC ditentukan oleh berbagai gen dalam
bentuk pertahanan biofisik dan biokimia. Ketahanan varietas padi terhadap hama
WBC ditentukan oleh beberapa faktor, yaitu faktor biofisik seperti ketebalan
jaringan tanaman, adanya trikhoma dan faktor biokimia seperti nutrisi atau
interaksi kedua faktor tersebut. Faktor-faktor tersebut mempengaruhi aktivitas
makan yang menentukan asupan nutrisi, yang pada gilirannya berakibat pada
pertumbuhan dan perkembangan wereng, kemampuan bertahan hidup dan
produksi telur. Yoshihara et al. (1980) mengemukakan bahwa perilaku makan berkaitan erat dengan kandungan asam oksalat pada tanaman padi. Asam amino,
sukrosa juga diketahui merupakan stimulan makan WBC pada tanaman padi (Chen
2009).
Penggunaan varietas tahan dalam pengendalian hama dianggap cara yang
relatif murah dan ramah lingkungan. Supaya ketahanan terhadap WBC tidak
mudah dipatahkan, penggunaan varietas tahan dilakukan dengan pola pergiliran
tanaman. Pola pergiliran tanaman tersebut didasarkan atas perbedaan gen
ketahanan yang dikandungnya. Varietas tanpa gen ketahanan, rentan terhadap
semua biotipe WBC. Varietas dengan gen Bph 1 tahan terhadap WBC biotipe 1
tahan terhadap WBC biotipe 1 dan 2, tetapi rentan terhadap biotipe 3 dan 4
(Baehaki 2005). Ketahanan tanaman inang, dalam hal ini tanaman padi,
menimbulkan pengaruh pada respon WBC selanjutnya pada pertumbuhan dan
perkembangan WBC. WBC yang dikurung dengan varietas padi Mudgo
menyebabkan mortalitas tinggi, pertumbuhan lambat, ukuran tubuh kecil, dan
fekunditas yang rendah (Kazushige dan Pathak 1970). Komponen ketahanan pada
tanaman padi, misalnya yang terdapat pada Ratthu Heenati (RH) yaitu terletak
dalam floem, sedangkan ketahanan pada varietas IR46 menyebabkan
meningkatnya penusukkan oleh WBC pada permukaan tanaman padi, hal ini
dihubungkan dengan terdapatnya lilin dari permukaan tanaman (Woodhead dan
Padgham 1988). Hasil penelitian pada varietas padi dari Sri Lanka, diketahui
schaftosid dan isoschaftosid yang merupakan senyawa fenol pada floem padi dan
bersifat antifidan terhadap WBC. Zat tersebut lebih banyak terdapat pada padi
tahan dibanding padi rentan (Stevenson et al. 1996). Genetik hama merupakan faktor internal pengendali perkembangan biotipe WBC, apabila ditanam varietas
padi tahan terhadap satu biotipe secara terus-menerus, maka pada beberapa musim
berikutya WBC telah sanggup membentuk biotipe baru yang dapat mematahkan
ketahanan varietas padi tersebut.
Adaptasi WBC terhadap varietas tanaman padi ditunjukkan melalui
perubahan produksi embun madu, berat tubuh, kelangsungan hidup dan
reproduksi (Chen 2009). WBC juga dapat beradaptasi terhadap varietas tahan
dalam beberapa generasi melalui pemeliharaan terus-menerus di laboratorium.
Oleh karena itu, memahami faktor-faktor yang mendasari interaksi WBC dan
tanaman padi adalah sangat penting.
Perumusan Masalah
Penggunaan varietas tahan hama merupakan salah satu teknik pengendalian
yang digunakan karena relatif murah, mudah diterapkan, dan tidak mencemari
lingkungan. Adanya varietas padi tahan dianggap sebagai penyebab perubahan
kemampuan populasi WBC dalam menginfestasi tanaman padi. Di lapangan,
dengan adanya pertanaman padi yang mengandung gen tahan secara
terus-menerus dapat mempercepat timbulnya biotipe baru atau mempercepat terjadinya
dengan responnya terhadap berbagai varietas padi yang mengandung faktor
ketahanan yang berbeda, sehingga selalu menimbulkan masalah, baik dalam
pemuliaan varietas padi maupun penggunaan varietas secara luas.
Tanaman tahan secara genetik menghasilkan kondisi tanaman yang
menyebabkan kualitas pertumbuhan dan perkembangan hama tidak optimal.
Sifat-sifat yang perlu diamati pada mekanisme antisenosis, antara lain adalah preferensi
nimfa untuk berkoloni, preferensi imago untuk makan, dan preferensi imago
untuk oviposisi. Berdasarkan mekanisme antibiosis, respon serangga yang diamati
adalah uji embun madu, lama hidup nimfa dan waktu yang diperlukan hingga
mencapai dewasa. Selain itu, dilakukan pengamatan terhadap faktor-faktor
ketahanan dari tanaman padi yang diuji diantaranya analisa jaringan dan biokimia
tanaman padi dengan berbagai tingkat ketahanan yang berkaitan dengan preferensi
hinggap dan perilaku makan,
Kerangka Pemikiran
Fungsi tanaman inang bagi herbivora antara lain adalah sebagai tempat
makan, hidup, serta peletakan telur (Schoonhoven et al. 2005). Seleksi tanaman inang oleh herbivora dipengaruhi oleh zat kimia tanaman yang dinamakan
senyawa kimia sekunder (Fraenkel 1959). Kennedy (1965) menyatakan bahwa
seleksi tanaman inang adalah berdasarkan respon arthropoda terhadap zat nutrien
dan non-nutrien tumbuhan. Secara alami, tumbuhan memiliki ketahanan terhadap
herbivora. Faktor ketahanan inilah yang digunakan para pemulia untuk
mengembangkan padi tahan WBC. Di Indonesia pengembangan padi tahan
dilakukan oleh lembaga-lembaga penelitian, dan telah menghasilkan berbagai
varietas yang memiliki ketahanan terhadap WBC. Sebelum tahun 2006 pengujian
galur-biotipe Bph1, bph2, atau keturunan Bph1+, juga disiapkan varietas yang
memiliki ketahanan terhadap biotipe 3. Untuk mengatasi cepatnya kepatahan pada
varietas yang memiliki satu gen ketahanan, dikembangkan varietas-varietas yang
memiliki ketahanan terhadap lebih dari satu gen ketahanan (ketahanan horizontal),
varietas ini dianggap dapat mempertahankan ketahanan lebih panjang karena
tekanan seleksi yang terjadi tidak setinggi pada varietas dengan satu gen
ketahanan. Penggunaan varietas dengan ketahanan vertikal akan menyebabkan
baru dengan daya serang yang lebih kuat (Sharma et al. 2002). Keberhasilan varietas tahan dibatasi dengan timbulnya biotipe baru yang dapat hidup, berkembang, dan
selanjutnya dapat mematahkan varietas yang semula tahan (Bahagiawati dan
Samudra 1998). Keadaan ini perlu dihindari agar varietas tahan dapat bertahan
dalam waktu lama, salah satu upaya adalah dengan mengelola tanaman tahan yaitu
melakukan pergiliran tanaman tahan dengan mekanisme resistensi yang berbeda.
Tanaman tahan dengan mekanisme resistensi berbeda menimbulkan respon WBC
yang berbeda.
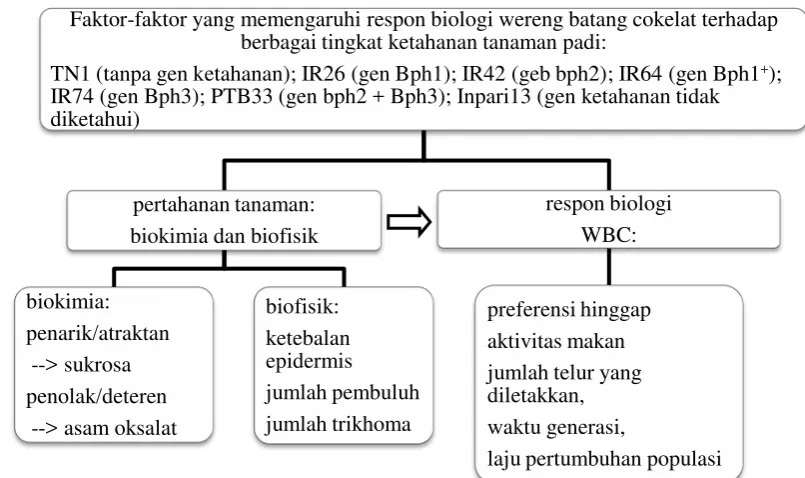
Gambar 1.1 Skema faktor-faktor yang mempengaruhi respon WBC terhadap ketahanan varietas tanaman padi. Keterangan: arah panah menunjukkan pengaruh faktor pertahanan tanaman padi terhadap respon biologi WBC
Ketahanan tanaman padi terhadap WBC ditentukan oleh berbagai gen dalam
bentuk pertahanan biokimia dan biofisik (Gambar 1.1), pada gilirannya ketahanan
ini akan mempengaruhi perilaku maupun metabolism WBC meliputi preferensi,
aktivitas makan, pertumbuhan dan perkembangan, serta jumlah telur yang
diletakkan, yang kemudian akan menentukan laju pertumbuhan populasi.
Untuk memahami respon WBC terhadap tanaman padi, langkah-langkah penelitian
dibagi menjadi:
Faktor-faktor yang memengaruhi respon biologi wereng batang cokelat terhadap berbagai tingkat ketahanan tanaman padi:
TN1 (tanpa gen ketahanan); IR26 (gen Bph1); IR42 (geb bph2); IR64 (gen Bph1+);
1. Respon biologi WBC terhadap faktor biofisik tanaman padi pada 7 varietas padi
dengan berbagai tingkat ketahanan; pengamatan dilakukan terhadap preferensi
hinggap dan preferensi oviposisi.
2. Respon biologi WBC terhadap faktor biokimia tanaman padi pada 7 varietas
tanaman padi dengan berbagai tingkat ketahanan; pengamatan dilakukan terhadap
kemampuan hidup nimfa dan waktu yang diperlukan hingga mencapai dewasa,
tingkat mortalitas imago, waktu penggandaan populasi dan kesesuaian makan
melalui uji embun madu WBC.
Hipotesis
1. Faktor biofisik tanaman padi berpengaruh terhadap respon biologi WBC seperti
preferensi hinggap dan tempat oviposisi WBC.
2. Faktor biokimia tanaman padi berpengaruh terhadap respon biologi WBC yang
diamati melalui kemampuan hidup nimfa dan waktu yang diperlukan hingga
mencapai dewasa, tingkat mortalitas imago, waktu penggandaan populasi dan
kemampuan makan melalui uji embun madu WBC.
Tujuan Penelitian
1. Mengetahui respon biologi WBC terhadap faktor biofisik varietas tanaman
padi yaitu preferensi hinggap dan oviposisi WBC.
2. Mengetahui respon biologi WBC terhadap faktor biokimia varietas tanaman
padi yaitu kemampuan makan melalui uji embun madu WBC, kemampuan
hidup melalui analisis neraca kehidupan.
Manfaat Penelitian
Penelitian diharapkan dapat mengetahui respon biologi WBC terhadap faktor
biofisik dan biokimia beberapa varietas tanaman padi sehingga pengelolaan penggunaan varietas tahan dapat dilakukan. Hal tersebut dicapai melalui informasi
mengenai kebugaran WBC melalui hasil penelitian analisis fisiologi WBC dan
analisis jaringan dan biokimia jaringan batang tanaman inang. Informasi hasil penelitian ini diharapkan dapat melengkapi metode penapisan ketahanan varietas
II. TINJAUAN PUSTAKA
Biologi Wereng Batang Cokelat
Nimfa WBC dapat berkembang menjadi dua bentuk wereng dewasa. Bentuk
pertama adalah makroptera (bersayap panjang) yaitu WBC yang mempunyai
sayap depan dan sayap belakang normal. Bentuk kedua adalah brakhiptera
(bersayap kerdil) yaitu WBC dewasa yang mempunyai sayap depan dan sayap
belakang tumbuh tidak normal, terutama sayap belakang sangat rudimenter.
Serangga brakhiptera memiliki waktu preoviposisi lebih pendek yaitu 3.8 hari
dibanding makroptera yaitu 7.2 hari (Mochida dan Okada 1979).
WBC berkembangbiak secara seksual, masa pra-peneluran 3-4 hari untuk
brakhiptera dan 3-8 hari untuk makroptera. Telur biasanya diletakkan pada
jaringan pangkal pelepah daun, tetapi kalau populasinya tinggi telur diletakkan di
ujung pelepah dan tulang daun. Telur diletakkan berkelompok, satu kelompok
telur terdiri dari 3-21 butir. Satu ekor betina mampu meletakkan telur 100-500
butir. Telur menetas setelah 7-10 hari. Muncul wereng muda yang disebut nimfa
dengan masa hidup 12-15 hari yang terdiri dari 5 instar nimfa dan setelah fase ini
menjadi wereng dewasa (Baehaki dan Widiarta 2009).
Seleksi Tanaman Inang oleh WBC
Hubungan herbivora dan inangnya berkaitan dengan fungsi tanaman inang
yaitu sebagai tempat makan, hidup, serta peletakan telur tersebut bagi herbivora.
(Schoonhoven et al. 2005). Seleksi tanaman inang oleh herbivora dipengaruhi oleh faktor biofisik dan faktor biokimia tanaman (Smith 2005). Secara alami,
tumbuhan memiliki ketahanan terhadap herbivora. Ketahanan inilah yang
digunakan para pemulia untuk mengembangkan padi tahan WBC.
Kekhususan tanaman inang merupakan karakteristik wereng dari Famili
Delphacidae. Di lapangan, kelimpahan WBC pada awalnya sama diantara varietas
tanaman padi dan tanaman lain, hal ini menunjukkan bahwa WBC tidak
memperlihatkan preferensi hinggap (Cook dan Perfect 1985). Hasil penelitian
menunjukkan bahwa ketahanan tanaman terhadap WBC tergantung pada senyawa
kimia dalam floem karena penolakan terhadap tanaman terjadi setelah pengisapan
inang oleh WBC dapat disebabkan oleh kandungan nutrisi atau pertahanan kimia
tanaman. Seleksi tanaman inang berkaitan dengan senyawa kimia dalam floem,
dan rendahnya stimulan makan dalam tanaman padi (Cook dan Denno 1994).
Hasil penelitian pada varietas Mudgo (Bph1), asam silisik dan asam oksalat
mencegah berlanjutnya proses makan oleh WBC (Yoshihara et al.1980). Asam fenol pada tanaman tahan berkaitan dengan ketidakmampuan WBC untuk
menemukan dan mengisap cairan floem. Selain itu, WBC menolak varietas padi
yang mengandung sedikit asam amino esensial dalam floem (Sogawa 1982).
WBC yang dikurung dengan varietas padi Mudgo menyebabkan
pertumbuhan lambat, berukuran tubuh kecil, fekunditas yang rendah, dan
menyebabkan terjadinya mortalitas (Kazushige dan Pathak 1970). Terdapat dua
komponen utama ketahanan padi terhadap serangga hama WBC. Pertama pada
floem, termasuk yang terdapat pada varietas tahan Ratthu Heenati (RH).
Stevenson et al. (1996) mengemukakan bahwa terdapat ketahanan biokimia dalam floem varietas tahan Ratthu Heenati (RH), dengan adanya schaftosid,
isoschaftosid, dan apigenin-glycosides total (seluruhnya merupakan
C-glycosidic flavonoid) dalam konsentrasi yang tinggi. Mortalitas WBC meningkat
dengan meningkatnya konsentrasi schaftosid. Mekanisme yang berasal dari RH
diduga merupakan efek antifidan daripada efek toksik. Komponen ketahanan
kedua adalah ketahanan biofisik, yaitu pertahanan tanaman berupa karakteristik
permukaan tanaman (Panda dan Khush 1995). Pada varietas IR46 (Bph1),
kandungan lilin pada permukaan daun menghambat perilaku makan WBC, yaitu
WBC harus berkali-kali menusukkan stiletnya untuk mencapai floem (Stevenson
et al. 1996). Selain itu, asam silisik dikenal merupakan inhibitor makan yang kuat terhadap WBC, zat ini terletak pada sel-sel parenkim padi (Denno dan Roderick
1990). Varietas standar tahan PTB33 mengandung lipid permukaan lebih tinggi
dibandingkan varietas standar rentan TN1 dan mempengaruhi perilaku makan WBC,
yaitu menyebabkan lama pengisapan yang lebih pendek dan mobilitas yang lebih
Mekanisme Ketahanan Tanaman
Ada 4 strategi dasar yang digunakan tanaman untuk pertahanan dirinya guna
mengurangi kerusakan akibat serangan serangga herbivor, yaitu: 1) escape atau menghindari serangan serangga berdasarkan waktu atau tempat, misalnya tumbuh
pada tempat yang tidak mudah diakses oleh herbivor atau menghasilkan bahan
kimia penolak herbivor (repellent); 2) tanaman toleran terhadap herbivor dengan cara mengalihkan herbivor untuk makan bagian yang tidak penting bagi tanaman
atau mengembangkan kemampuan untuk melakukan pemulihan dari kerusakan
akibat serangan herbivor; 3) tanaman menarik datangnya musuh alami bagi
herbivor yang dapat melindungi tanaman tersebut dari serangan herbivor, dan
terakhir 4) tanaman melindungi dirinya sendiri melalui mekanisme pertahanan
kimia atau fisik seperti menghasilkan toksin yang dapat membunuh herbivor atau
dapat mengurangi kemampuan herbivor untuk mencerna tanaman itu yang sering
disebut dengan antibiosis (Painter 1951).
Oleh karena itu suatu varietas tanaman dapat disebut tahan apabila: (1)
memiliki sifat-sifat yang memungkinkan tanaman itu menghindar, atau pulih
kembali dari serangan hama pada keadaan yang akan mengakibatkan kerusakan
pada varietas lain yang tidak tahan; (2) memiliki sifat-sifat genetik yang dapat
mengurangi tingkat kerusakan yang disebabkan oleh serangan hama; (3) memiliki
sekumpulan sifat genetik yang dapat diwariskan, yang dapat mengurangi
kemungkinan hama untuk menggunakan tanaman tersebut sebagai inang; atau (4)
mampu menghasilkan produk yang lebih banyak dan lebih baik dibandingkan
dengan varietas lain pada tingkat populasi hama yang sama (Sumarno 1992).
Painter (1951) membagi mekanisme ketahanan tanaman terhadap serangga
hama ke dalam 3 bentuk, yaitu:
a). Ketidaksukaan (non-preferences) yang kemudian istilah tersebut diganti dengan antisenosis atau menolak kehadiran serangga pada tanaman. Bentuk
mekanisme ketahanan antisenosis dibagi dalam dua kelompok, yaitu:
antisenosis kimiawi, menolak kerana adanya senyawa allelokimia dan
antisenosis fisik, menolak karena adanya struktur atau sifat morfologi
b). Antibiosis yaitu sifat ketahanan yang memberi pengaruh fisiologis yang
merugikan pada serangga, yang merupakan akibat dari serangga yang makan
dan mencerna jaringan atau cairan tanaman tertentu. Gejala-gejala akibat
antibiosis pada serangga diantaranya adalah: kematian larva atau pradewasa,
pengurangan laju pertumbuhan, peningkatan mortalitas pupa, kegagalan pupa
menjadi dewasa, imago tidak normal dan fekunditas serta fertilitas rendah,
masa hidup serangga berkurang, terjadi malformasi, kegagalan mengumpulkan
cadangan makanan dan kegagalan hibernasi, perilaku gelisah dan abnormalitas
lainnya. Menurut Kogan dan Ortman (1978) gejala-gejala abnormal tersebut
terjadi diakibatkan oleh beberapa hal, antara lain: adanya metabolit toksik
pada jaringan tanaman seperti alkaloid, glukosid dan quinon, tidak ada atau
kurang tersedianya unsur nutrisi utama bagi serangga, ketidakseimbangan
perbandingan unsur-unsur nutrisi yang tersedia, adanya antimetabolit yang
menghalangi ketersediaan beberapa unsur nutrisi bagi serangga, dan adanya
enzim-enzim yang mampu menghalangi proses pencernaan makanan dan
pemanfaatan unsur nutrisi oleh serangga. Contoh beberapa kasus antibiosis,
antara lain: pengurangan kadar asparagin (Chen 2009) pada varietas yang
tahan terhadap WBC.
c). Toleran merupakan respon tanaman terhadap serangga, sehingga beberapa ahli
tidak memasukannya dalam ketahanan. Beberapa faktor yang mengakibatkan
tanaman toleran terhadap serangan hama, adalah: kekuatan tanaman secara
umum, pertumbuhan kembali jaringan tanaman yang rusak, ketegaran batang
dan ketahanan terhadap rebah, produksi cabang tambahan, pemanfaatan lebih
efisien oleh serangga dan kompensasi lateral oleh tanaman tetangganya.
Sifat-sifat Ketahanan Tanaman
Ketahanan tanaman inang terhadap hama, dapat bersifat genetik, yaitu sifat
tahan yang diatur oleh sifat-sifat genetik yang dapat diwariskan, dan ketahanan
ekologi, yaitu ketahanan tanaman yang disebabkan oleh pengaruh faktor
Ketahanan Genetik
Ketahanan genetik juga dapat dibedakan menjadi beberapa tipe: (1)
ketahanan vertikal, ketahanan hanya terhadap satu biotipe hama, dan biasanya
bersifat sangat tahan tetapi mudah patah oleh munculnya biotipe baru; (2)
ketahanan horizontal atau ketahanan umum, ketahanan terhadap banyak biotipe
hama dengan derajat ketahanan yang tidak tinggi (agak tahan); dan (3) ketahanan
ganda, memiliki sifat tahan terhadap beberapa jenis hama.
Varietas-varietas padi yang dilepas di Indonesia, sebagian besar tidak
diketahui gen ketahanannya. Varietas Ciherang dilepas tahun 2000 misalnya,
merupakan hasil persilangan IR18349-53-1-3-1-3/IR19661-131-3-1 dengan IR64.
Berdasarkan hasil pengujian penapisan, varietas ini tahan terhadap biotipe 2 dan
agak tahan terhadap biotipe 3. Padi-padi hibrida yang memiliki potensi hasil
tinggi, namun belum disertai dengan sifat ketahanan terhadap WBC biotipe 3
(Suprihatno et al. 2010).
Tipe ketahanan vertikal dikendalikan oleh gen tunggal (monogenik) atau oleh beberapa gen (oligogenik ) dan hanya efektif terhadap biotipe hama tertentu. Secara umum sifat ketahanan vertikal mempunyai ciri-ciri : (1) biasanya
diwariskan oleh gen tunggal atau hanya sejumlah kecil gen, (2) relatif mudah
diidentifikasi dan banyak dipakai dalam program perbaikan ketahanan genetik, (3)
biasanya dikaitkan dengan hipotesis “gen for gen” , (4) menghasilkan ketahanan genetik tingkat tinggi, tidak jarang mencapai imunitas, tetapi jika timbul biotipe
baru maka ketahanan ini akan mudah patah dan biasanya tanaman menjadi sangat
rentan terhadap biotipe tersebut, dan (5) biasanya menunda awal terjadinya
epidemi, tetapi apabila terjadi epidemi maka kerentanannya tidak akan berbeda
dengan kultivar yang rentan (Sutopo dan Saleh, 1992).
Tipe ketahanan horizontal disebut juga ketahanan kuantitatif. Tanaman yang
memiliki ketahanan demikian masih menunjukkan sedikit kepekaan terhadap
hama tetapi memiliki kemampuan untuk memperlambat laju perkembangan
epidemi. Secara teoritis, ketahanan horisontal efektif untuk semua biotipe suatu
hama. Oleh karena itu, ketahanan ini umumnya sulit dipatahkan meskipun muncul
biotipe baru dengan daya serang yang lebih tinggi. Varietas dengan tipe ketahanan
minor ke dalam suatu varietas dengan karakter agronomik yang unggul melalui
pemuliaan konvensional maupun non-konvesional.
Ketahanan Ekologi
Ketahanan ekologi atau ketahanan terlihat (apparent resistance) atau ketahanan palsu (pseudo resistance) dikendalikan oleh keadaan lingkungan. Ketahanan ekologi ini tidak diturunkan dan tergantung dari kekuatan tekanan dari
lingkungan.
Genetik hama merupakan faktor internal pengendali perkembangan WBC,
sehingga apabila ditanam varietas padi tahan terhadap satu biotipe secara
terus-menerus, maka pada beberapa musim saja WBC telah sanggup membentuk
biotipe baru yang lebih ganas. Oleh karena itu dikembangkan varietas padi tahan
terhadap lebih dari satu biotipe untuk menekan timbulnya biotipe baru, misalnya
Inpari13 yang baru dilepas tahun 2009, berdasarkan hasil penapisan, varietas ini
tahan terhadap biotipe 1, 2 dan 3. Penggunaan varietas tahan dalam pengendalian
hama dianggap cara yang relatif murah dan mudah untuk diterapkan petani, serta
ramah lingkungan. Penggunaan varietas tahan dilakukan dengan pola pergiliran
tanaman. Pola pergiliran tanaman tersebut didasarkan atas gen ketahanan yang
berbeda yang memberikan respon berbeda terhadap WBC. Varietas TN1 yaitu
varietas tanpa gen tahan, rentan terhadap semua biotipe WBC. Varietas dengan
gen Bph 1 tahan terhadap WBC biotipe 1 dan 3, tetapi rentan terhadap biotipe 2
dan 4. Varietas dengan gen Bph 2 tahan terhadap WBC biotipe 1 dan 2, tetapi
rentan terhadap biotipe 3 dan 4 (Baehaki 2005).
Ketahanan Tanaman Padi terhadap Wereng Batang Cokelat sebagai Dasar Pembentukan Perakitan Varietas Padi Tahan
Di Indonesia pengembangan padi tahan WBC dilakukan oleh
lembaga-lembaga penelitian dan telah menghasilkan berbagai varietas yang memiliki sifat
ketahanan terhadap WBC. Sebelum tahun 2006 pengujian galur-biotipe bph2,
Bph1 atau keturunan Bph1+, juga disiapkan varietas yang memiliki ketahanan
terhadap biotipe 3. Untuk mengatasi cepatnya kepatahan terhadap serangan WBC
pada varietas yang memiliki satu gen ketahanan, dikembangkan varietas-varietas
memiliki ketahanan lebih lama karena tekanan seleksi yang terjadi tidak setinggi
pada varietas dengan satu gen ketahanan.
Dalam pengembangan varietas tahan WBC, proses uji penapisan yang
digunakan saat ini adalah standard seedbox screening test (SSST). Benih dari galur yang diuji ditanam pada kotak berukuran 60 cm x 40 cm x10 cm. Varietas
standar tahan dan standar rentan ditanam dalam kotak yang sama, kemudian
diinfestasi oleh nimfa instar 2 per benih. Ketika tanaman standar rentan TN1 mati,
dilakukan skoring terhadap kerusakan tanaman (Velusamy et al. 1986). Metode ini hanya dapat mengevaluasi respon makan nimfa terhadap galur yang diuji.
WBC dapat beradaptasi dan mematahkan ketahanan varietas yang semula
bereaksi tahan. Penanaman varietas yang sama secara terus-menerus, pemakaian
insektisida yang kurang bijaksana, dan teknik budidaya yang kurang baik akan
mendorong munculnya biotipe baru WBC (Harahap et al. 1987, Hanarida 1998, Soewito et al. 1995).
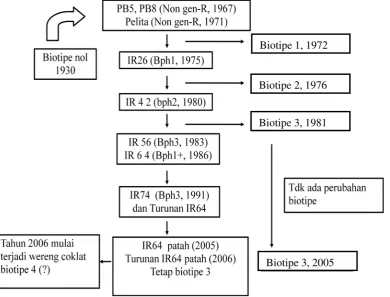
Saat ini di Indonesia diketahui terdapat WBC biotipe 1, 2, 3, dan 4.
Intensifikasi pertanian dengan menggunakan varietas padi lokal, memunculkan
biotipe 1 pada tahun 1972. Kestabilan WBC biotipe nol bertahan selama 41 tahun
sebelum menjadi WBC biotipe 1. Untuk mengatasi biotipe 1, dilepas varietas
IR26 yang dilepas IRRI tahun 1973 diintroduksikan ke Indonesia pada tahun
1975. IR26 dengan gen tahan Bph1 (Bph = Brown Plant Hopper) dapat dipatahkan pada tahun 1976, muncul biotipe 2. Selanjutnya dilepas varietas IR42
pada tahun 1980 dengan gen tahan bph2. Varietas yang tahan terhadap biotipe 2
ternyata bisa dipatahkan dengan munculnya biotipe 3 pada tahun 1981.
Pertanaman monokultur varietas IR56 (Bph3) secara terus-menerus
mengakibatkan gen tahan Bph3 patah kembali. Perubahan WBC biotipe 1 ke
WBC biotipe 2 terjadi hanya dalam waktu 4 tahun, dan perubahan WBC biotipe 2
ke WBC biotipe 3 hanya dalam kurun waktu 5 tahun. Kemudian dilepas Varietas
IR 64 mengandung gen tahan Bph1+ yang tahan terhadap serangan WBC biotipe 3
pada tahun 1986. Varietas IR64, seperti varietas IR26 memiliki gen ketahanan
mayor yang sama yaitu Bph1, namun IR64 bertahan lebih lama dibanding IR26.
Hal tersebut disebabkan IR64 memiliki satu atau lebih gen ketahanan minor,
biotipe 3 tersebut bertahan hingga 2005, berarti sudah 25 tahun sejak timbulnya
WBC biotipe 2 masih tetap didominasi WBC biotipe 3, namun pada 2006 mulai
dilaporkan adanya serangan WBC, yang diduga biotipe 4 di Asahan, Sumatera
Utara (Baehaki 2008, Khush dan Virk 2005). Perkembangan penggunaan varietas
tahan dan perkembangan biotipe WBC dipaparkan pada Gambar 2.1. Hasil uji
biotipe WBC di Indonesia oleh Baehaki dan Munawar (2007) dikemukakan
bahwa di Jawa Barat, Jawa Tengah, Jawa Timur dan Sumatera Selatan adalah
biotipe 3. WBC yang berasal dari Maros, Sulawesi Selatan adalah campuran
biotipe 2 dan 3, sedangkan WBC di Asahan, Sumatra Utara adalah campuran
biotipe 2, 3, dan 4.
Gambar 2.1 Perkembangan biotipe dan varietas tahan WBC di Indonesia (Baehaki
2008)
Definisi ketahanan tanaman terhadap serangga yang disampaikan para ahli
beragam sesuai sudut pandang mereka, antara lain: Painter (1951) mendefinisikan
ketahanan tanaman merupakan sifat-sifat tanaman yang dapat diturunkan dan pada
tingkat serangan hama yang sama dapat berproduksi lebih baik, serta dapat
mempengaruhi tingkat kerusakan oleh serangga. Beck (1965) mengemukakan Biotipe 1, 1972
Biotipe 2, 1976
Biotipe 3, 1981
bahwa ketahanan tanaman adalah semua ciri dan sifat tanaman yang
memungkinkan tanaman terhindar dan/atau menekan pertumbuhan dan
perkembangan serangga. Senada dengan Painter (1951), Teetes (1996)
menyatakan bahwa dalam praktek pertanian, ketahanan tanaman berarti
kemampuan tanaman untuk berproduksi lebih baik dibandingkan tanaman lain
dengan tingkat populasi hama yang sama.
Dalam penerapan tanaman tahan diharapkan suatu tanaman tahan dapat
bertahan lama atau tidak mudah dipatahkan. Dalam prakteknya, tanaman tahan
merupakan salah satu komponen PHT dan dalam penggunaan tanaman tahan perlu
dikelola sehingga tidak mudah dipatahkan. Hal ini dapat dilakukan apabila
dipahami respon WBC terhadap tanaman tahan yang merupakan tujuan penelitian
III. RESPON BIOLOGI WERENG BATANG COKELAT
TERHADAP FAKTOR BIOFISIK TUJUH VARIETAS
TANAMAN PADI
(Biological responses of brown planthopper to biophysical factors in seven rice varieties)
Abstrak
Faktor-faktor biofisik tanaman mempengaruhi perilaku serangga. Faktor ini dapat mempengaruhi aktivitas makan, peletakan telur atau sebagai tempat hidup WBC. Tujuan penelitian ini adalah untuk mempelajari respon biologi dalam hal preferensi hinggap dan peletakan telur pada tanaman padi. Penelitian dilaksanakan di Kebun Percobaan Balai Besar Penelitian Tanaman Padi, Muara, Bogor dan berlangsung selama November 2010 – Maret 2012. Pembuatan preparat jaringan dibuat di Lembaga Ilmu Pengetahuan Indonesia, Cibinong. Biotipe WBC yang diteliti adalah biotipe 2 dan 3, sedangkan varietas tanaman padi yang digunakan dalam penelitian ini antara lain TN1 (tanpa gen ketahanan), IR26 (Bph1), IR42 (bph2), IR64 (Bph1+), IR74 (Bph3), PTB33 (bph2+Bph3) and Inpari13 (gen ketahanan tidak diketahui). WBC menunjukkan reaksi non-preferen terhadap varietas tahan, termasuk Inpari13. Peletakan telur WBC biotipe 2 pada varietas rentan TN1 dan IR26 berbeda nyata dengan varietas tahan PTB33 dan IR74. WBC biotipe 3 meletakkan telur lebih banyak pada varietas rentan TN1 dan IR42 dan berbeda nyata dengan PTB33, IR74, IR64 dan Inpari13. Biotipe 3 memiliki ovipositor berukuran lebih besar dari biotipe 2. Jumlah trikhoma pada permukaan tanaman dan ketebalan jaringan epidermis batang tidak berpengaruh terhadap perilaku hinggap dan peletakan telur.
Kata kunci: respon biologi, WBC, padi, faktor biofisik
Abstract
had larger ovipositor than biotype 2. Number of trichomes on leaf sheath and epidermis thickness were not associated with settling and oviposition preference of brown planthoppers.
Key words: biological response, brown planthopper, rice, biophysical factors
Pendahuluan
WBC merupakan hama tanaman padi pada ekosistem padi berpengairan di
Indonesia. Beberapa strategi pengelolaan telah diajukan untuk mengendalikan
WBC, di antaranya penggunaan tanaman tahan merupakan pilihan yang dianggap
paling bersahabat dengan lingkungan dan mudah diimplementasikan petani.
Upaya pengendalian hama WBC dengan menanam varietas tahan merupakan
metode yang praktis, akan tetapi tantangannya adalah potensi berkembangnya
biotipe baru WBC yang mampu mematahkan daya tahan varietas, sehingga
varietas yang dulunya tahan akan berubah menjadi tidak tahan. Di Indonesia,
terdapat perubahan biotipe WBC, diiringi dengan dilepasnya varietas tahan
wereng yang mengandung gen ketahanan. Sehubungan dengan timbulnya biotipe
1 tahun 1972, varietas IR26 dilepas di Indonesia pada tahun 1975, dengan gen
tahan Bph1 (Bph = Brown Plant Hopper), pada tahun 1976 muncul biotipe 2. Selanjutnya dilepas varietas IR42 pada tahun 1980 dengan gen tahan bph2 dan
tahan biotipe 2. WBC yang semula tahan terhadap biotipe 2 ternyata bisa
dipatahkan dengan muculnya biotipe 3 pada tahun 1981. Kemudian dilepas
Varietas IR64 dilepas pada tahun 1986.mengandung gen tahan Bph1+
Keberhasilan hidup serangga untuk makan dan bertahan hidup diawali dengan
perilaku serangga menemukan dan memakan tanaman inang yang kemudian diikuti
dengan kesesuaian tanaman inang (Schoonhoven et al. 2005). Perilaku hinggap WBC, pada tanaman padi meliputi berjalan, berhenti, mengetukkan labium,
yang tahan
terhadap serangan WBC biotipe 3. Varietas IR74 memiliki gen Bph3 dan tahan
terhadap biotipe 1, 2, dan 3. PTB33 memiliki dua gen tahan yaitu bph2 dan Bph3,
berasal dari India dan merupakan gen donor dalam pembentukan varietas tahan
wereng (Baehaki 2008, Khush dan Virk 2005). Dalam program pemuliaan padi
tahan, masih sedikit informasi mengenai identifikasi dan pemahaman mekanisme
yang mendasari ketahanan tersebut serta respon WBC terhadap sifat ketahanan
penyisipan alat mulut, mengecap, sekresi ludah, pengisapan, membentuk seludang
ludah melanjutkan pengisapan, dan ekskresi embun madu (Sogawa 1982).
Faktor-faktor yang menentukan pertumbuhan dan perkembangan serangga pada tanaman
dikategorikan dalam dua kelompok yaitu respon serangga terhadap tanaman, dan
karakter tanaman yang mempengaruhi respon serangga. Respon serangga
termasuk orientasi, perilaku makan, pertumbuhan nimfa menjadi dewasa, lama
hidup dewasa, banyaknya telur yang dihasilkan, peletakan telur, dan penetasan
telur (Saxena dan Pathak 1979). Mekanisme penemuan tanaman inang seringkali
dipengaruhi oleh penghalang fisik pada tanaman terhadap serangga untuk hinggap
pada permukaan tanaman. Karakter morfologi tanaman merupakan salah satu
kunci ketahanan tanaman terhadap serangga (Heinrichs 1992). Struktur morfologi
atau faktor biofisik tanaman seperti bulu daun, lilin di permukaan daun, ketebalan
jaringan dan kandungan alelokimia memungkinkan tanaman untuk mempengaruhi
perilaku makan serangga (Saxena dan Pathak 1979). Faktor biofisik ini juga
berkaitan dengan antisenosis tanaman dalam mengatasi perilaku makan dan
peletakan telur oleh serangga herbivora. Penelitian ini bertujuan untuk mengkaji
respon biologi WBC terhadap biofisik tanaman padi meliputi preferensi tempat
hinggap/makan dan tempat oviposisi WBC, selain itu diamati pula karakteristik
tanaman padi.
Bahan dan Metode
Waktu dan Tempat
Penelitian dilakukan di laboratorium dan rumah kaca Kebun Percobaan
Muara, Balai Besar Penelitian Tanaman Padi dan Laboratorium Fisiologi dan
Toksikologi Departemen Proteksi, Fakultas Pertanian IPB di Bogor, pada bulan
Nopember 2010 – Maret 2012. Pembuatan preparat tanaman padi dilakukan di
Lembaga Ilmu Pengetahuan Indonesia di Cibinong.
Bahan Penelitian
Serangga uji yang digunakan dalam penelitian ini adalah WBC biotipe 2
yang dipelihara pada padi varietas IR26 dan biotipe 3 yang dipelihara pada
Materi penelitian yang digunakan terdiri atas tujuh varietas tanaman padi,
yaitu PTB33 (bph2, bph3), IR74 (Bph3), IR64 (Bph1+
Metode Penelitian
), IR42 (bph2), IR26
(Bph1), TN1 (tanpa gen ketahanan) dan Inpari13 (gen ketahanan tidak diketahui).
Preferensi Tempat Hinggap
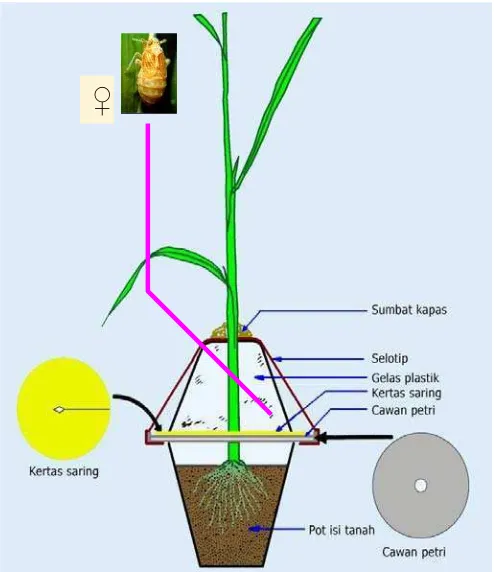
Penelitian preferensi tempat hinggap WBC, menggunakan metode Heinrichs
et al. (1985). Benih varietas padi yang diuji disemai dalam kotak berukuran 60 cm x 40 cm x10 cm. Masing-masing benih berjarak 4 cm disemai berbentuk lingkaran
berdiameter 40 cm. Setiap varietas diulang sebanyak 5 kali. Setelah varietas tanaman
padi yang diuji berumur 3 minggu setelah semai, WBC betina imago sebanyak 105
ekor ditempatkan pada piring petri, kemudian diletakkan di tengah lingkaran
tanaman. WBC dibiarkan bergerak dan memilih varietas padi untuk hinggap. Pada
saat 2, 4, 8, 24, 48, dan 72 jam setelah pelepasan, WBC yang hinggap pada masing
masing varietas diamati dan dihitung jumlahnya. Data hasil pengamatan diolah
dengan menggunakan sidik ragam yang dilanjutkan dengan uji jarak berganda Duncan (α = 0.05) dengan menggunakan SAS v.9.
Preferensi Oviposisi WBC
Penelitian preferensi oviposisi oleh WBC menggunakan metode Heinrichs et al. (1985). Tujuh varietas tanaman uji disemai pada kotak penapisan berukuran 60 cm x 40 cm x10 cm. Setelah tanaman uji berumur 30 hari setelah semai, tunas
tanaman padi dipotong hingga pangkal dan hingga tersisa 2 tunas per rumpun. WBC
yang digunakan dalam penelitian berasal dari pemeliharaan di rumah kasa, yaitu
biotipe 2 dipelihara pada varietas IR26, dan biotipe 3 dipelihara pada varietas IR42.
Sepuluh ekor imago betina diinfestasikan pada setiap rumpun. Setiap varietas
diulang 3 kali. Setelah 72 jam, tanaman padi dipotong pada pangkalnya, kemudian
diperiksa di bawah mikroskop stereo untuk menghitung jumlah telur pada setiap
varietas yang diuji. Banyak telur yang diletakkan pada setiap varietas dianalisis
Luas Sentroid Ovipositor WBC
Analisis morfometri ovipositor WBC, dimulai dengan menyiapkan
ovipositor untuk difoto dan kemudian dianalisis. Ovipositor dilepaskan dari
abdomen betina dan diletakan di atas kaca obyek yang telah ditetesi pewarna kuku
transparan. Pengambilan foto ovipositor dilakukan di laboratorium Jurusan
Proteksi Tanaman. Kamera yang digunakan adalah kamera digital Nikon D100
dilengkapi satu paket lensa adapter (terdiri atas: BR-2, Bower 37-52, Bower
28-37, dan tabung krom), dihubungkan dengan lensa okuler Olympus PE 3,3 X 125,
Mikroskop Olympus BX 41, dan lensa Objektif Olympus Plan 4X/0,01. Setiap
foto ovipositor diberi label, kemudian seluruh foto yang akan dianalisis
dikumpulkan menjadi satu folder. Pengukuran bagian luasan ovipositor (image) diukur dengan cara menetapkan titik-titik tertentu dari bagian tubuh yang
kemudian diukur secara berurutan pada program TPS-util dan TPS-dig2 yang
diunduh dari situs
modifikasi dari Tantowijoyo dan Hoffman (2010). Penentuan titik dilakukan pada
skala perbesaran gambar 0,467 pada program tps.dig2. Hasil ploting setiap titik
landmark secara otomatis berubah menjadi nilai angka di dalam koordinat sumbu x dan sumbu y dalam bentuk format data csv (data transformasi gambar ke nilai angka dalam program microsoft excel). Selanjutnya jarak titik pusat terhadap sumbu x (sentroid x) dan sumbu y (sentroid y) dihitung berturut-turut dengan merata-ratakan nilai x1, x2, x3, ..., xn dan y1, y2, y3,..., yn. Jarak setiap titik
terhadap perpotongan sumbu x dan sumbu y (distanced) dihitung dengan mengukur jarak atau garis diagonal perpotongan sentroid x dan sentroid y. Luasan lansekap (bagian) tubuh yang diukur (sentroid size) dihitung dengan merata-ratakan setiap nilai distanced titik pengukuran. Luasan ini dapat ditransformasikan ke dalam ukuran mikrometer dengan membandingkan nilai sentroid size skala objektif mikrometer.
1.
Nilai hasil transformasi tersebut di atas dapat dihitung melalui program
microsoft excel dengan rumus sebagai berikut: Sentroid x = (x1+ x2+ x3 + ...xn Distanced = SQRT ((xn - Sentroid x)^2 + ((yn 4.
Faktor Biofisik Tanaman
Pengamatan karakter morfologi tanaman padi, dimulai dengan pembuatan
preparat mikroskopis tanaman padi. Penelitian ini menggunakan metode metode
infiltrasi parafin yang digunakan oleh Sass (1951) yang diadopsi oleh Sunarti et al. (2008).
Setiap 1 (satu) cm bagian jaringan diisolasi dari tanaman uji, kemudian direndam
dan disimpan sementara (tidak lebih dari 1 hari) dalam larutan 70% etanol teknis.
Proses pelemasan jaringan batang biasanya dilakukan melalui perendaman jaringan
terlebih dahulu ke dalam larutan 10% KOH di laboratorium selama beberapa jam
sebelum difiksasi. Fiksasi jaringan dilakukan di dalam larutan FAA (campuran
formalin dan asam asetat glasial) yang diletakkan dalam desikator vakum (wadah
gelas hampa udara), selama minimal 24 jam. Setelah fiksasi, jaringan didehidrasi
dengan cara memindahkannya ke dalam larutan etanol-silol bertingkat
masing-masing selama 3 jam. Selanjutnya infiltrasi parafin ke dalam jaringan dilakukan
secara perlahan dengan cara memindahkannya berturut-turut ke dalam larutan
campuran silol-parafin bertingkat. Perbandingan antara silol dan parafin adalah 3:1, 1:1, dan 1:3, preparat diletakkan di dalam inkubator suhu ± 60 °C, masing-masing
minimal selama 3 jam. Parafin yang digunakan memiliki titik leleh 56-58 °C.
Selanjutnya parafin berisi jaringan dikeluarkan dari inkubator dan dibiarkan
membeku. Proses ini disebut proses “embedding” dengan tujuan agar spesimen
mudah dipotong oleh mikrotom. Jaringan kemudian dipotong dengan arah melintang
dan membujur. Serial spesimen irisan jaringan diletakkan di atas permukaan objek
gelas yang sebelumnya telah diolesi dengan glycerin. Spesimen ditetesi akuades dan
diletakkan di atas hot plate. Untuk menghilangkan parafin, lekapan atau preparat direndam dalam larutan silol selama 2 x 3 menit. Selanjutnya dilakukan
penghilangan molekul silol yang berada dalam jaringan dengan memasukkannya ke
dalam larutan campuran silol-etanol bertingkat masing-masing selama 3 menit.
Pewarnaan jaringan dilakukan dengan cara merendam lekapan ke dalam larutan
safranin 1% dalam etanol setelah lekapan berada pada tahap perendaman etanol
70%. Untuk pewarnaan lanjut digunakan larutan pewarna fast green 2% dalam
etanol absolut setelah lekapan melalui perendaman etanol bertingkat 70% dan 95%.
Kemudian dilakukan dealkoholisasi dan infiltrasi silol ke dalam lekapan sebelum
gelas objek. Pengukuran ketebalan epidermis diamati pada irisan membujur preparat
tanaman.padi. Pertama-tama preparat yang telah dibuat, difoto dengan kamera
Nikon™ dengan pembesaran 200x, lalu ketebalan diukur dengan program Motic
Image Plus v.7. Jumlah trikhoma dihitung dengan cara menghitung banyak nya
trikhoma pada irisan membujur sepanjang 1 (satu) cm preparat tanaman padi. Data
dianalisis dengan analisis sidik ragam dan uji selang berganda Duncan.
Hasil dan Pembahasan
Preferensi Tempat Hinggap dan Peletakan Telur oleh WBC pada Berbagai Varietas Padi
Seleksi tanaman inang oleh serangga, seperti halnya pada aphid, meliputi
rangkaian proses meliputi aktivitas mencari, orientasi, kemudian di dalamnya
termasuk hinggap, lalu melakukan seleksi tempat penusukan hingga penetrasi
stilet (Schoonhoven et al. 2005). Pada penelitian ini dilakukan pengamatan terhadap preferensi hinggap WBC pada tanaman padi, dengan hasil disajikan pada
Tabel 3.1
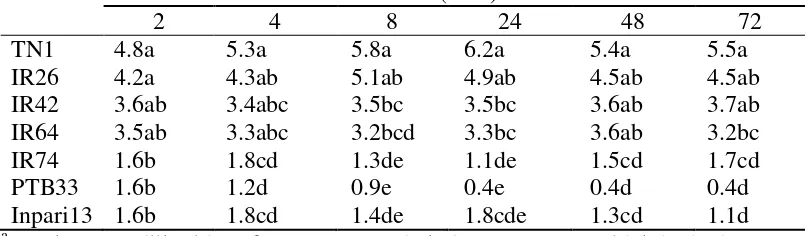
Tabel 3.1 Preferensi hinggap imago WBC pada berbagai varietas tanaman padi dan pada waktu yang berbeda setelah dilepaskan
Varietas Jumlah WBC hinggap pada tanaman padi setelah dilepaskan pada jam ke- (ekor)a
2 4 8 24 48 72
TN1 4.8a 5.3a 5.8a 6.2a 5.4a 5.5a
IR26 4.2a 4.3ab 5.1ab 4.9ab 4.5ab 4.5ab IR42 3.6ab 3.4abc 3.5bc 3.5bc 3.6ab 3.7ab IR64 3.5ab 3.3abc 3.2bcd 3.3bc 3.6ab 3.2bc IR74 1.6b 1.8cd 1.3de 1.1de 1.5cd 1.7cd PTB33 1.6b 1.2d 0.9e 0.4e 0.4d 0.4d Inpari13 1.6b 1.8cd 1.4de 1.8cde 1.3cd 1.1d a
Angka yang diikuti huruf yang sama pada kolom yang sama tidak berbeda nyata pada selang kepercayaan 5% dengan menggunakan uji selang berganda Duncan
Preferensi hinggap WBC pada 2 jam pertama setelah dilepaskan tidak
berbeda nyata antara TN1, IR26, IR42, IR64. Tetapi varietas TN1 dan IR26
berbeda nyata dengan varietas tahan IR74, PTB33 dan Inpari13 (Tabel 3.1).
Setelah empat jam, preferensi wereng memiliki kecenderungan untuk hinggap
pada varietas inangnya yaitu IR42. Setelah 24 jam, terlihat preferensi dan varietas
tahan berbeda nyata antara varietas IR74, PTB33 serta Inpari13. Pada awal
pelepasan ke dalam kurungan, WBC hinggap pada tanaman padi secara acak,
kemudian berangsur akan berpindah ke varietas yang lebih disukai yaitu varietas
yang lebih rentan. Selama pengamatan, banyaknya WBC hinggap pada varietas
TN1 dan IR26 lebih tinggi daripada varietas lainnya, kisaran WBC hinggap pada
varietas TN1 selama pengamatan adalah 4.8 hingga 6.2 ekor/tanaman.
Berdasarkan ketahanan varietas tanaman padi terhadap WBC, (Khush dan Virk
2005), IR26 dan IR64 tahan terhadap biotipe 3, kedua varietas ini sama-sama
memiliki gen mayor Bph1. Preferensi hinggap WBC sama terhadap kedua varietas
tersebut mulai pada 2 jam pertama hingga pengamatan 72 jam.
Karakteristik tanaman berupa stimulus fisik dapat mempengaruhi aktifitas
serangga. Variasi ukuran daun, bentuk, warna, dan ada/tidaknya sekresi glandular
mungkin dapat berperan dalam menentukan penerimaan serangga terhadap
inangnya. Kondisi permukaan dan jaringan yang kuat dapat menjadi faktor
pembatas dalam proses hinggap dan makan serangga. Karakter morfologi atau
biofisik tanaman dikaitkan dengan mekanisme antisenosis selain karakter
biokimia. Meski karakter biofisik dapat berpengaruh dalam menurunkan respon
wereng, namun karakter biofisik tidak dianggap sebagai penyebab utama
non-preferen oleh WBC, karena WBC dapat membedakan varietas tahan dan rentan
yang secara morfologi identik (Soundararajan et al. 2005). WBC tidak
menunjukkan perbedaan dalam preferensi terhadap varietas yang diuji, tetapi
WBC, tidak akan menetap pada variets tahan untuk melakukan aktivitas makan
secara terus menerus (Sogawa dan Pathak 1970). Woodhead dan Padgham (1988),
mengemukakan bahwa aktivitas wereng pada permukaan tanaman padi
dihubungkan dengan kandungan lilin pada bagian epikutikula tanaman. Varietas
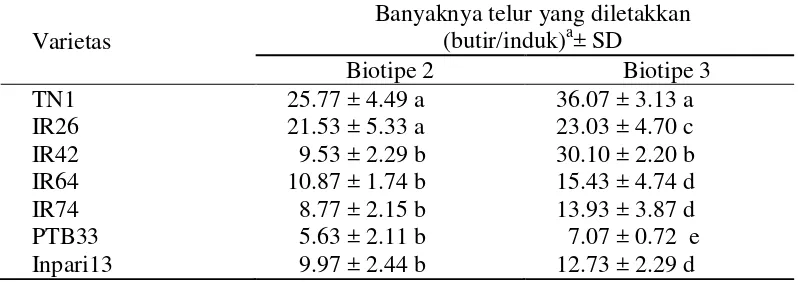
Tabel 3.2 Preferensi peletakan telur WBC pada berbagai varietas tanaman padi
Varietas
Banyaknya telur yang diletakkan (butir/induk)a± SD
Angka yang diikuti huruf yang sama pada kolom yang sama tidak berbeda nyata pada selang kepercayaan 5% dengan menggunakan uji selang berganda Duncan
Preferensi peletakan telur atau oviposisi WBC pada berbagai tingkat
ketahanan tanaman padi menunjukkan variasi baik oleh wereng biotipe 2 maupun
biotipe 3 (Tabel 3.2). Kedua biotipe WBC menunjukkan pola oviposisi yang
serupa pada varietas standar tahan PTB33 (jumlah telur yang diletakkan terendah)
dan varietas rentan TN1 (jumlah telur yang diletakkan tertinggi), juga untuk
varietas IR74, IR64, dan IR26. Pada varietas IR64, peletakan telur oleh WBC
biotipe 2 adalah rendah, juga pada varietas IR42, IR74, PTB33 dan Inpari13
(Tabel 3.2). Menurut Khush dan Virk (2005), IR64 memiliki ketahanan sedang
atau moderate resistance terhadap biotipe 2, sedangkan IR42, IR74 dan PTB33 bereaksi tahan terhadap WBC biotipe 2. Inpari13 yang merupakan varietas
introduksi dari Vietnam dan belum diketahui gen ketahanannya, bereaksi tahan
terhadap WBC biotipe 2 ini. Jadi ada faktor resisten pada varietas IR74, PTB33
dan Inpari13 terhadap respon peneluran WBC. WBC biotipe 3 meletakkan telur
terbanyak pada TN1 yaitu sebanyak 36.07 butir diikuti oleh IR42, IR26, IR64,
IR74, Inpari13 dan PTB33. Varietas IR42 yang memiliki gen bph2 bereaksi
rentan terhadap biotipe 3, sedangkan varietas IR64 (Bph1+), IR74 (Bph3) dan
PTB33 (bph2, Bph3) bereaksi tahan, juga Inpari13. Tingginya peneluran biotipe 3
pada varietas IR42 disebabkan varietas ini merupakan tanaman inang bagi
perbanyakan IR42 di laboratorium, sehingga biotipe 3 telah beradaptasi dengan
baik pada varietas ini. Menurut Khush dan Virk (2005), IR26 bereaksi tahan
penelitian ini menunjukkan bahwa WBC biotipe 3 dapat mengatasi ketahanan
IR26.
WBC meletakkan telurnya dengan cara menusukkan ovipositor ke dalam
jaringan tanaman padi. Berdasarkan penelitian Hattori dan Sogawa (2002),
terdapat tiga urutan perilaku peletakan telur WBC setelah penusukan ovipositor,
yaitu menggerakan ovipositor seperti menggergaji, pengeluaran telur, dan menarik
keluar ovipositor. Gerakan seperti menggergaji inilah yang menyebabkan
kumpulan telur WBC kumpulan seperti deretan buah pisang.
Luas Sentroid Ovipositor WBC
Ovipositor berfungsi untuk menyisipkan telur ke dalam jaringan pelepah
daun padi. Berdasarkan pengukuran morfometri pada ovipositor WBC adalah
sebagai berikut (Tabel 3.3):
Tabel 3.3 Luas ovipositor WBC betina biotipe 2 dan biotipe 3 berdasarkan morfometri ovipositor
Populasi WBC Rerata luas sentroid ovipositor ± SD
Uji t
Biotipe 2 291.11 ± 25.31 0,0022*
Biotipe 3 315.61 ± 17.61
Keterangan: tanda * menunjukkan perbedaan nyata luas ovipositor antara kedua biotipe WBC
Hasil analisis morfometri menunjukkan bahwa ovipositor WBC biotipe 3
memiliki luas sentroid ovipositor lebih besar dibandingkan ovipositor WBC
biotipe 2. Hasil penelitian terhadap preferensi peneluran (Tabel 3.2) menunjukkan
peneluran oleh biotipe 3 lebih banyak daripada biotipe 2, terutama peneluran oleh
biotipe 3 pada IR42. Kemungkinannya adalah biotipe 3 telah beradaptasi dengan
baik pada IR42 yang merupakan tanaman inangnya. Morfologi ovipositor diduga
berkaitan dengan kemampuan menusukkan ovipositor ke dalam jaringan tanaman
(Chapman 1998). Pada jangkrik tanah, panjang ovipositor berkaitan dengan
Faktor Biofisik Tanaman Padi pada Berbagai Varietas
Pengamatan terhadap faktor biofisik yang diamati yaitu banyaknya trikhoma
dan ketebalan epidermis pada irisan membujur pelepah daun berumur 30 hari
setiap varietas disajikan pada Tabel 3.4.
Tabel 3.4 Jumlah trikhoma dan tebal epidermis pada pelepah daun berbagai varietas tanaman padi
Varietas Rerata jumlah trikhoma ± SD (/cm)
Angka yang diikuti huruf yang sama pada kolom yang sama tidak berbeda nyata pada selang kepercayaan 5% dengan menggunakan uji selang berganda Duncan
Hasil pengamatan terhadap jumlah trikhoma, tidak terdapat perbedaan nyata
antar varietas tanaman padi. Perbedaan dalam perilaku WBC pada berbagai
varietas tidak terkait dengan jumlah trikhoma tetapi mungkin berkaitan dengan
komposisi kimia permukaan daun (Nugaliyadde dan Wilkins 2012). Hasil
penelitian Nwilene et al. (2009) mengenai komponen antisenosis pada tanaman padi, diantaranya disebutkan bahwa kepadatan trikhoma pada daun padi tidak
berkaitan dengan ketahanan terhadap African rice gall midge, Orseolia oryzivora.
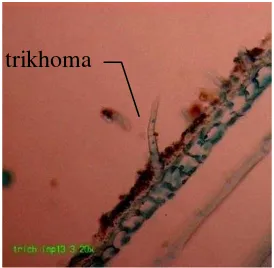
Melalui pengamatan pada struktur trikhoma, terdapat trikhoma dengan
bentuk memanjang seperti pada varietas Inpari13 (Gambar 3.1). Jadi dalam
mempertimbangkan trikhoma sebagai faktor resistensi hendaknya diobservasi
jumlah trikhoma per unit luas area. Selain itu bentuk dan panjang trikhoma perlu
diamati.
Tebal epidermis varietas tahan PTB33 (26.27µ) berbeda nyata dengan
varietas rentan TN1 (23.14µ), tetapi tebal epidermis varietas tanaman padi IR26,
IR42, IR74, IR64 dan Inpari13 tidak berbeda nyata dengan PTB33. Hal ini
menunjukkan perbedaan gen ketahanan terhadap WBC dalam varietas tanaman
padi tidak berkaitan dengan ketebalan epidermis. PTB33 yang mengandung gen
IR26 (gen Bph1). IR64 dan PTB33 mempunyai tebal epidermis berbeda dengan
varietas standar rentan TNI.
Gambar 3.1 Trikhoma pada permukaan pelepah daun varietas Inpari13 berdasarkan irisan membujur (perbesaran 200x)
Faktor-faktor biofisik tanaman yang berperan dalam pertahanan tanaman
terhadap herbivor diantaranya trikhoma lilin permukaan, ketebalan jaringan
(Smith 2005). Dalam penelitian ini yang diamati adalah trikhoma dan ketebalan
jaringan, dari hasil penelitian diduga gen ketahanan varietas padi tidak berkaitan
dengan banyaknya trikhoma dan ketebalan epidermis, hanya PTB33 yang
menunjukkan perbedaan ketebalan dengan varietas standar rentan TN1.
Kesimpulan
Dalam proses seleksi tanaman inang, WBC menunjukkan reaksi
non-preferen terhadap varietas tahan PTB33, IR74, IR64, dan Inpari13. WBC biotipe 2
lebih banyak meletakkan telur pada varietas rentan TN1 dan IR26 dibandingkan
pada varietas tahan PTB33 dan IR74. WBC biotipe 3 juga meletakkan telur lebih
banyak pada TN1 dan IR42 daripada PTB33, IR74, IR64 dan Inpari13. Besar
ovipositor WBC juga berperan dalam merespon ketahanan tanaman padi, yaitu
WBC biotipe 3 memiliki luas sentroid ovipositor lebih besar dari biotipe 2.
Ketebalan sel epidermis dan jumlah trikhoma pada pelepah daun tidak berperan
dalam mekanisme ketahanan tanaman padi terhadap WBC, yaitu dalam hal
Daftar Pustaka
Baehaki SE. 2008. Perkembangan wereng batang cokelat Biotipe 4 http://www.litbang.deptan.go.id/artikel/one/171/pdf/perkembangan%20Wer eng%20Cokelat%20Biotipe%204.pdf [10 Juli 2008].
Chapman RF. 1998. The Insect Structure and Function. Ed ke 4. Cambridge (UK): Cambridge University Press.
Hattori M, Sogawa K. 2002 Oviposition behavior of the rice brown planthopper,
Nilaparvata lugens (Stål), and its electronic monitoring. Journal of Insect Behavior 15 (2): 283-293.
Heinrichs EA, Medrano FG, Rapusas HR.1985. Genetic Evaluation for Insect Resistance in Rice. Los Banos (Philippines): IRRI.
Heinrichs EA. 1992. Rice insects: the role of host plant resistance in integrated management systems. Kor J Appl Entomol 31(3):256-275.
Khush GS, Virk PS. 2005. IR Varieties and Their Impact. Los Baños (Philippines): IRRI.
Nwilene FE, Okhidevbie O, Agunbiade TA. 2009. An antisenosis component of rice resistance to African rice gall midge, Orselia oryzivora.IRRN.
Nugaliyadde L, Wilkins RM. Influence of surface lipid of some rice varieties on the feeding behavior of Nilaparvata lugens.
http://www.goviya.lk/agri_learning/Paddy/Paddy_Research/Paddy_pdf/P10. pdf [10 Agustus 2012].
Panda N, Khush GS. 1995. Host Plant Resistance to Insects. Oxon: CAB International.
Reale D, Roff DA. 2002. Quantitative genetics of oviposition behaviour and interactions among oviposition traits in the sand cricket. Animal Behaviour
64: 397–406.
Sass, J.E. 1951. Botanichal Microtechnique. 2nd
Saxena RC, Pathak MD. 1979. Factors governing susceptibility and resistance of certain rice varieties to the brown planthopper. In: Brown planthopper: Threat of Rice Production in Asia. Los Banos (Philippines): IRRI. hlm 303-317.
Ed. Iowa: The Iowa State University Press.
Schoonhoven LM, van Loon JJA, Dicke M. 2005. Insect-Plant Biology. Second Edition. NewYork: Oxford University Press.
Sogawa K. 1982. The rice brown planthopper: Feeding physiology and host plant interactions. Ann Rev Entomol 27: 49-73.
Sogawa K, Pathak MD. 1970. Menchanism of brown planthopper resistance in Mudgo variety of rice. Appl Entomol Zool 5: 145–158.
Soundararajan RP, Gunathilagaraj K, Chitra N, Maheswaran M, Kadirvel P. 2005. Mechanism and genetics of resistance to brown planthopper, Nilaparvata lugens in rice, Oryza sativa L. – a review. Agric Rev 26 (2): 79 – 91.
Sunarti, S., Rugayah, dan E.F. Tihurua. Studi anatomi dan jenis-jenis Averrhoa di Indonesia untuk mempertegas status taksonominya. Berita Biologi 9 (3): 253-257.
Tantowijoyo W, Hoffman AA. 2010. Variation in morphological characters of two invasive leafminers, Liriomyza huidobrensis and L. sativae, across a tropical elevation gradient. Journal of Insect Science: 11 (69): 1-16.