III. RESPON BIOLOGI WERENG BATANG COKELAT
TERHADAP FAKTOR BIOFISIK TUJUH VARIETAS
TANAMAN PADI
(Biological responses of brown planthopper to biophysical factors in seven rice varieties)
Abstrak
Faktor-faktor biofisik tanaman mempengaruhi perilaku serangga. Faktor ini dapat mempengaruhi aktivitas makan, peletakan telur atau sebagai tempat hidup WBC. Tujuan penelitian ini adalah untuk mempelajari respon biologi dalam hal preferensi hinggap dan peletakan telur pada tanaman padi. Penelitian dilaksanakan di Kebun Percobaan Balai Besar Penelitian Tanaman Padi, Muara, Bogor dan berlangsung selama November 2010 – Maret 2012. Pembuatan preparat jaringan dibuat di Lembaga Ilmu Pengetahuan Indonesia, Cibinong. Biotipe WBC yang diteliti adalah biotipe 2 dan 3, sedangkan varietas tanaman padi yang digunakan dalam penelitian ini antara lain TN1 (tanpa gen ketahanan), IR26 (Bph1), IR42 (bph2), IR64 (Bph1+), IR74 (Bph3), PTB33 (bph2+Bph3) and Inpari13 (gen ketahanan tidak diketahui). WBC menunjukkan reaksi non-preferen terhadap varietas tahan, termasuk Inpari13. Peletakan telur WBC biotipe 2 pada varietas rentan TN1 dan IR26 berbeda nyata dengan varietas tahan PTB33 dan IR74. WBC biotipe 3 meletakkan telur lebih banyak pada varietas rentan TN1 dan IR42 dan berbeda nyata dengan PTB33, IR74, IR64 dan Inpari13. Biotipe 3 memiliki ovipositor berukuran lebih besar dari biotipe 2. Jumlah trikhoma pada permukaan tanaman dan ketebalan jaringan epidermis batang tidak berpengaruh terhadap perilaku hinggap dan peletakan telur.
Kata kunci: respon biologi, WBC, padi, faktor biofisik
Abstract
Plant biophysical factors affect the behavior of insects. In the resistance mechanism of rice, antisenosis, these factors could affect feeding activity, oviposition or as shelter of brown planthopper. The aims of the research was to study the biological responses of brown planthopper on biophysical factors of rice varieties in terms of settling and oviposition preferences. The study was conducted in Research Station of the Indonesian Center for Rice Research, Muara- Bogor during November 2010-March 2012. Microtechnique of rice tissue were prepared in the Indonesian Institute of Science, Cibinong -Bogor. Rice varieties such as TN1 (no resistance gene), IR26 (Bph1), IR42 (bph2), IR64 (Bph1+), IR74 (Bph3), PTB33 (bph2+Bph3) and Inpari13 (unknown resistance gene) were used in this study. Adults showed same non-preference reactions to resistant varieties, included Inpari13 for settling. Oviposition of brown planthopper biotype 2 on susceptible varieties such as TN1 and IR26 was higher than on resistant varieties such as PTB33, IR74. Brown planthopper biotype 3 laid more eggs on susceptible TN1 and IR42 compared to PTB33, IR74, IR64 and Inpari13.varieties. Biotype 3
had larger ovipositor than biotype 2. Number of trichomes on leaf sheath and epidermis thickness were not associated with settling and oviposition preference of brown planthoppers.
Key words: biological response, brown planthopper, rice, biophysical factors
Pendahuluan
WBC merupakan hama tanaman padi pada ekosistem padi berpengairan di Indonesia. Beberapa strategi pengelolaan telah diajukan untuk mengendalikan WBC, di antaranya penggunaan tanaman tahan merupakan pilihan yang dianggap paling bersahabat dengan lingkungan dan mudah diimplementasikan petani. Upaya pengendalian hama WBC dengan menanam varietas tahan merupakan metode yang praktis, akan tetapi tantangannya adalah potensi berkembangnya biotipe baru WBC yang mampu mematahkan daya tahan varietas, sehingga varietas yang dulunya tahan akan berubah menjadi tidak tahan. Di Indonesia, terdapat perubahan biotipe WBC, diiringi dengan dilepasnya varietas tahan wereng yang mengandung gen ketahanan. Sehubungan dengan timbulnya biotipe 1 tahun 1972, varietas IR26 dilepas di Indonesia pada tahun 1975, dengan gen tahan Bph1 (Bph = Brown Plant Hopper), pada tahun 1976 muncul biotipe 2. Selanjutnya dilepas varietas IR42 pada tahun 1980 dengan gen tahan bph2 dan tahan biotipe 2. WBC yang semula tahan terhadap biotipe 2 ternyata bisa dipatahkan dengan muculnya biotipe 3 pada tahun 1981. Kemudian dilepas Varietas IR64 dilepas pada tahun 1986.mengandung gen tahan Bph1+
Keberhasilan hidup serangga untuk makan dan bertahan hidup diawali dengan perilaku serangga menemukan dan memakan tanaman inang yang kemudian diikuti dengan kesesuaian tanaman inang (Schoonhoven et al. 2005). Perilaku hinggap WBC, pada tanaman padi meliputi berjalan, berhenti, mengetukkan labium,
yang tahan terhadap serangan WBC biotipe 3. Varietas IR74 memiliki gen Bph3 dan tahan terhadap biotipe 1, 2, dan 3. PTB33 memiliki dua gen tahan yaitu bph2 dan Bph3, berasal dari India dan merupakan gen donor dalam pembentukan varietas tahan wereng (Baehaki 2008, Khush dan Virk 2005). Dalam program pemuliaan padi tahan, masih sedikit informasi mengenai identifikasi dan pemahaman mekanisme yang mendasari ketahanan tersebut serta respon WBC terhadap sifat ketahanan suatu varietas..
penyisipan alat mulut, mengecap, sekresi ludah, pengisapan, membentuk seludang ludah melanjutkan pengisapan, dan ekskresi embun madu (Sogawa 1982). Faktor-faktor yang menentukan pertumbuhan dan perkembangan serangga pada tanaman dikategorikan dalam dua kelompok yaitu respon serangga terhadap tanaman, dan karakter tanaman yang mempengaruhi respon serangga. Respon serangga termasuk orientasi, perilaku makan, pertumbuhan nimfa menjadi dewasa, lama hidup dewasa, banyaknya telur yang dihasilkan, peletakan telur, dan penetasan telur (Saxena dan Pathak 1979). Mekanisme penemuan tanaman inang seringkali dipengaruhi oleh penghalang fisik pada tanaman terhadap serangga untuk hinggap pada permukaan tanaman. Karakter morfologi tanaman merupakan salah satu kunci ketahanan tanaman terhadap serangga (Heinrichs 1992). Struktur morfologi atau faktor biofisik tanaman seperti bulu daun, lilin di permukaan daun, ketebalan jaringan dan kandungan alelokimia memungkinkan tanaman untuk mempengaruhi perilaku makan serangga (Saxena dan Pathak 1979). Faktor biofisik ini juga berkaitan dengan antisenosis tanaman dalam mengatasi perilaku makan dan peletakan telur oleh serangga herbivora. Penelitian ini bertujuan untuk mengkaji respon biologi WBC terhadap biofisik tanaman padi meliputi preferensi tempat hinggap/makan dan tempat oviposisi WBC, selain itu diamati pula karakteristik tanaman padi.
Bahan dan Metode Waktu dan Tempat
Penelitian dilakukan di laboratorium dan rumah kaca Kebun Percobaan Muara, Balai Besar Penelitian Tanaman Padi dan Laboratorium Fisiologi dan Toksikologi Departemen Proteksi, Fakultas Pertanian IPB di Bogor, pada bulan Nopember 2010 – Maret 2012. Pembuatan preparat tanaman padi dilakukan di Lembaga Ilmu Pengetahuan Indonesia di Cibinong.
Bahan Penelitian
Serangga uji yang digunakan dalam penelitian ini adalah WBC biotipe 2 yang dipelihara pada padi varietas IR26 dan biotipe 3 yang dipelihara pada varietas padi IR42.
Materi penelitian yang digunakan terdiri atas tujuh varietas tanaman padi, yaitu PTB33 (bph2, bph3), IR74 (Bph3), IR64 (Bph1+
Metode Penelitian
), IR42 (bph2), IR26 (Bph1), TN1 (tanpa gen ketahanan) dan Inpari13 (gen ketahanan tidak diketahui).
Preferensi Tempat Hinggap
Penelitian preferensi tempat hinggap WBC, menggunakan metode Heinrichs et al. (1985). Benih varietas padi yang diuji disemai dalam kotak berukuran 60 cm x 40 cm x10 cm. Masing-masing benih berjarak 4 cm disemai berbentuk lingkaran berdiameter 40 cm. Setiap varietas diulang sebanyak 5 kali. Setelah varietas tanaman padi yang diuji berumur 3 minggu setelah semai, WBC betina imago sebanyak 105 ekor ditempatkan pada piring petri, kemudian diletakkan di tengah lingkaran tanaman. WBC dibiarkan bergerak dan memilih varietas padi untuk hinggap. Pada saat 2, 4, 8, 24, 48, dan 72 jam setelah pelepasan, WBC yang hinggap pada masing masing varietas diamati dan dihitung jumlahnya. Data hasil pengamatan diolah dengan menggunakan sidik ragam yang dilanjutkan dengan uji jarak berganda Duncan (α = 0.05) dengan menggunakan SAS v.9.
Preferensi Oviposisi WBC
Penelitian preferensi oviposisi oleh WBC menggunakan metode Heinrichs et al. (1985). Tujuh varietas tanaman uji disemai pada kotak penapisan berukuran 60 cm x 40 cm x10 cm. Setelah tanaman uji berumur 30 hari setelah semai, tunas tanaman padi dipotong hingga pangkal dan hingga tersisa 2 tunas per rumpun. WBC yang digunakan dalam penelitian berasal dari pemeliharaan di rumah kasa, yaitu biotipe 2 dipelihara pada varietas IR26, dan biotipe 3 dipelihara pada varietas IR42. Sepuluh ekor imago betina diinfestasikan pada setiap rumpun. Setiap varietas diulang 3 kali. Setelah 72 jam, tanaman padi dipotong pada pangkalnya, kemudian diperiksa di bawah mikroskop stereo untuk menghitung jumlah telur pada setiap varietas yang diuji. Banyak telur yang diletakkan pada setiap varietas dianalisis dengan menggunakan sidik ragam dan dilanjutkan dengan uji jarak berganda Duncan (α = 0.05) dengan menggunakan SAS v.9.
Luas Sentroid Ovipositor WBC
Analisis morfometri ovipositor WBC, dimulai dengan menyiapkan ovipositor untuk difoto dan kemudian dianalisis. Ovipositor dilepaskan dari abdomen betina dan diletakan di atas kaca obyek yang telah ditetesi pewarna kuku transparan. Pengambilan foto ovipositor dilakukan di laboratorium Jurusan Proteksi Tanaman. Kamera yang digunakan adalah kamera digital Nikon D100 dilengkapi satu paket lensa adapter (terdiri atas: BR-2, Bower 37-52, Bower 28-37, dan tabung krom), dihubungkan dengan lensa okuler Olympus PE 3,3 X 125, Mikroskop Olympus BX 41, dan lensa Objektif Olympus Plan 4X/0,01. Setiap foto ovipositor diberi label, kemudian seluruh foto yang akan dianalisis dikumpulkan menjadi satu folder. Pengukuran bagian luasan ovipositor (image) diukur dengan cara menetapkan titik-titik tertentu dari bagian tubuh yang kemudian diukur secara berurutan pada program TPS-util dan TPS-dig2 yang diunduh dari situs http://life.bio.sunysb.edu/morph/, metode ini merupakan modifikasi dari Tantowijoyo dan Hoffman (2010). Penentuan titik dilakukan pada skala perbesaran gambar 0,467 pada program tps.dig2. Hasil ploting setiap titik landmark secara otomatis berubah menjadi nilai angka di dalam koordinat sumbu x dan sumbu y dalam bentuk format data csv (data transformasi gambar ke nilai angka dalam program microsoft excel). Selanjutnya jarak titik pusat terhadap sumbu x (sentroid x) dan sumbu y (sentroid y) dihitung berturut-turut dengan merata-ratakan nilai x1, x2, x3, ..., xn dan y1, y2, y3,..., yn. Jarak setiap titik terhadap perpotongan sumbu x dan sumbu y (distanced) dihitung dengan mengukur jarak atau garis diagonal perpotongan sentroid x dan sentroid y. Luasan lansekap (bagian) tubuh yang diukur (sentroid size) dihitung dengan merata-ratakan setiap nilai distanced titik pengukuran. Luasan ini dapat ditransformasikan ke dalam ukuran mikrometer dengan membandingkan nilai sentroid size skala objektif mikrometer.
1.
Nilai hasil transformasi tersebut di atas dapat dihitung melalui program microsoft excel dengan rumus sebagai berikut:
Sentroid x = (x1+ x2+ x3 + ...xn 2. )/n Sentroid y = (y1+ y2+ y3 +...yn 3. )/n Distanced = SQRT ((xn - Sentroid x)^2 + ((yn 4.
- Sentroid y)^2 Sentroid size = Sum (distanced 1, distanced 2, ... distanced n)/n
Faktor Biofisik Tanaman
Pengamatan karakter morfologi tanaman padi, dimulai dengan pembuatan preparat mikroskopis tanaman padi. Penelitian ini menggunakan metode metode infiltrasi parafin yang digunakan oleh Sass (1951) yang diadopsi oleh Sunarti et al. (2008). Setiap 1 (satu) cm bagian jaringan diisolasi dari tanaman uji, kemudian direndam dan disimpan sementara (tidak lebih dari 1 hari) dalam larutan 70% etanol teknis. Proses pelemasan jaringan batang biasanya dilakukan melalui perendaman jaringan terlebih dahulu ke dalam larutan 10% KOH di laboratorium selama beberapa jam sebelum difiksasi. Fiksasi jaringan dilakukan di dalam larutan FAA (campuran formalin dan asam asetat glasial) yang diletakkan dalam desikator vakum (wadah gelas hampa udara), selama minimal 24 jam. Setelah fiksasi, jaringan didehidrasi dengan cara memindahkannya ke dalam larutan etanol-silol bertingkat masing-masing selama 3 jam. Selanjutnya infiltrasi parafin ke dalam jaringan dilakukan secara perlahan dengan cara memindahkannya berturut-turut ke dalam larutan campuran silol-parafin bertingkat. Perbandingan antara silol dan parafin adalah 3:1, 1:1, dan 1:3, preparat diletakkan di dalam inkubator suhu ± 60 °C, masing-masing minimal selama 3 jam. Parafin yang digunakan memiliki titik leleh 56-58 °C. Selanjutnya parafin berisi jaringan dikeluarkan dari inkubator dan dibiarkan membeku. Proses ini disebut proses “embedding” dengan tujuan agar spesimen mudah dipotong oleh mikrotom. Jaringan kemudian dipotong dengan arah melintang dan membujur. Serial spesimen irisan jaringan diletakkan di atas permukaan objek gelas yang sebelumnya telah diolesi dengan glycerin. Spesimen ditetesi akuades dan diletakkan di atas hot plate. Untuk menghilangkan parafin, lekapan atau preparat direndam dalam larutan silol selama 2 x 3 menit. Selanjutnya dilakukan penghilangan molekul silol yang berada dalam jaringan dengan memasukkannya ke dalam larutan campuran silol-etanol bertingkat masing-masing selama 3 menit. Pewarnaan jaringan dilakukan dengan cara merendam lekapan ke dalam larutan safranin 1% dalam etanol setelah lekapan berada pada tahap perendaman etanol 70%. Untuk pewarnaan lanjut digunakan larutan pewarna fast green 2% dalam etanol absolut setelah lekapan melalui perendaman etanol bertingkat 70% dan 95%. Kemudian dilakukan dealkoholisasi dan infiltrasi silol ke dalam lekapan sebelum ditetesi bahan pengawet yaitu balsam canada. Setelah itu lekapan ditutup dengan
gelas objek. Pengukuran ketebalan epidermis diamati pada irisan membujur preparat tanaman.padi. Pertama-tama preparat yang telah dibuat, difoto dengan kamera Nikon™ dengan pembesaran 200x, lalu ketebalan diukur dengan program Motic Image Plus v.7. Jumlah trikhoma dihitung dengan cara menghitung banyak nya trikhoma pada irisan membujur sepanjang 1 (satu) cm preparat tanaman padi. Data dianalisis dengan analisis sidik ragam dan uji selang berganda Duncan.
Hasil dan Pembahasan
Preferensi Tempat Hinggap dan Peletakan Telur oleh WBC pada Berbagai Varietas Padi
Seleksi tanaman inang oleh serangga, seperti halnya pada aphid, meliputi rangkaian proses meliputi aktivitas mencari, orientasi, kemudian di dalamnya termasuk hinggap, lalu melakukan seleksi tempat penusukan hingga penetrasi stilet (Schoonhoven et al. 2005). Pada penelitian ini dilakukan pengamatan terhadap preferensi hinggap WBC pada tanaman padi, dengan hasil disajikan pada Tabel 3.1
Tabel 3.1 Preferensi hinggap imago WBC pada berbagai varietas tanaman padi dan pada waktu yang berbeda setelah dilepaskan
Varietas Jumlah WBC hinggap pada tanaman padi setelah dilepaskan pada jam ke- (ekor)a
2 4 8 24 48 72
TN1 4.8a 5.3a 5.8a 6.2a 5.4a 5.5a
IR26 4.2a 4.3ab 5.1ab 4.9ab 4.5ab 4.5ab
IR42 3.6ab 3.4abc 3.5bc 3.5bc 3.6ab 3.7ab IR64 3.5ab 3.3abc 3.2bcd 3.3bc 3.6ab 3.2bc
IR74 1.6b 1.8cd 1.3de 1.1de 1.5cd 1.7cd
PTB33 1.6b 1.2d 0.9e 0.4e 0.4d 0.4d
Inpari13 1.6b 1.8cd 1.4de 1.8cde 1.3cd 1.1d a
Angka yang diikuti huruf yang sama pada kolom yang sama tidak berbeda nyata pada selang kepercayaan 5% dengan menggunakan uji selang berganda Duncan
Preferensi hinggap WBC pada 2 jam pertama setelah dilepaskan tidak berbeda nyata antara TN1, IR26, IR42, IR64. Tetapi varietas TN1 dan IR26 berbeda nyata dengan varietas tahan IR74, PTB33 dan Inpari13 (Tabel 3.1). Setelah empat jam, preferensi wereng memiliki kecenderungan untuk hinggap pada varietas yang rentan seperti TN1 yang tidak memiliki gen ketahanan dan
pada varietas inangnya yaitu IR42. Setelah 24 jam, terlihat preferensi dan varietas tahan berbeda nyata antara varietas IR74, PTB33 serta Inpari13. Pada awal pelepasan ke dalam kurungan, WBC hinggap pada tanaman padi secara acak, kemudian berangsur akan berpindah ke varietas yang lebih disukai yaitu varietas yang lebih rentan. Selama pengamatan, banyaknya WBC hinggap pada varietas TN1 dan IR26 lebih tinggi daripada varietas lainnya, kisaran WBC hinggap pada varietas TN1 selama pengamatan adalah 4.8 hingga 6.2 ekor/tanaman. Berdasarkan ketahanan varietas tanaman padi terhadap WBC, (Khush dan Virk 2005), IR26 dan IR64 tahan terhadap biotipe 3, kedua varietas ini sama-sama memiliki gen mayor Bph1. Preferensi hinggap WBC sama terhadap kedua varietas tersebut mulai pada 2 jam pertama hingga pengamatan 72 jam.
Karakteristik tanaman berupa stimulus fisik dapat mempengaruhi aktifitas serangga. Variasi ukuran daun, bentuk, warna, dan ada/tidaknya sekresi glandular mungkin dapat berperan dalam menentukan penerimaan serangga terhadap inangnya. Kondisi permukaan dan jaringan yang kuat dapat menjadi faktor pembatas dalam proses hinggap dan makan serangga. Karakter morfologi atau biofisik tanaman dikaitkan dengan mekanisme antisenosis selain karakter biokimia. Meski karakter biofisik dapat berpengaruh dalam menurunkan respon wereng, namun karakter biofisik tidak dianggap sebagai penyebab utama non-preferen oleh WBC, karena WBC dapat membedakan varietas tahan dan rentan yang secara morfologi identik (Soundararajan et al. 2005). WBC tidak menunjukkan perbedaan dalam preferensi terhadap varietas yang diuji, tetapi WBC, tidak akan menetap pada variets tahan untuk melakukan aktivitas makan secara terus menerus (Sogawa dan Pathak 1970). Woodhead dan Padgham (1988), mengemukakan bahwa aktivitas wereng pada permukaan tanaman padi dihubungkan dengan kandungan lilin pada bagian epikutikula tanaman. Varietas tahan mengandung lilin lebih tinggi.
Tabel 3.2 Preferensi peletakan telur WBC pada berbagai varietas tanaman padi
Varietas
Banyaknya telur yang diletakkan (butir/induk)a± SD Biotipe 2 Biotipe 3 TN1 25.77 ± 4.49 a 36.07 ± 3.13 a IR26 21.53 ± 5.33 a 23.03 ± 4.70 c IR42 9.53 ± 2.29 b 30.10 ± 2.20 b IR64 10.87 ± 1.74 b 15.43 ± 4.74 d IR74 8.77 ± 2.15 b 13.93 ± 3.87 d PTB33 5.63 ± 2.11 b 7.07 ± 0.72 e Inpari13 9.97 ± 2.44 b 12.73 ± 2.29 d a
Angka yang diikuti huruf yang sama pada kolom yang sama tidak berbeda nyata pada selang kepercayaan 5% dengan menggunakan uji selang berganda Duncan
Preferensi peletakan telur atau oviposisi WBC pada berbagai tingkat ketahanan tanaman padi menunjukkan variasi baik oleh wereng biotipe 2 maupun biotipe 3 (Tabel 3.2). Kedua biotipe WBC menunjukkan pola oviposisi yang serupa pada varietas standar tahan PTB33 (jumlah telur yang diletakkan terendah) dan varietas rentan TN1 (jumlah telur yang diletakkan tertinggi), juga untuk varietas IR74, IR64, dan IR26. Pada varietas IR64, peletakan telur oleh WBC biotipe 2 adalah rendah, juga pada varietas IR42, IR74, PTB33 dan Inpari13 (Tabel 3.2). Menurut Khush dan Virk (2005), IR64 memiliki ketahanan sedang atau moderate resistance terhadap biotipe 2, sedangkan IR42, IR74 dan PTB33 bereaksi tahan terhadap WBC biotipe 2. Inpari13 yang merupakan varietas introduksi dari Vietnam dan belum diketahui gen ketahanannya, bereaksi tahan terhadap WBC biotipe 2 ini. Jadi ada faktor resisten pada varietas IR74, PTB33 dan Inpari13 terhadap respon peneluran WBC. WBC biotipe 3 meletakkan telur terbanyak pada TN1 yaitu sebanyak 36.07 butir diikuti oleh IR42, IR26, IR64, IR74, Inpari13 dan PTB33. Varietas IR42 yang memiliki gen bph2 bereaksi rentan terhadap biotipe 3, sedangkan varietas IR64 (Bph1+), IR74 (Bph3) dan PTB33 (bph2, Bph3) bereaksi tahan, juga Inpari13. Tingginya peneluran biotipe 3 pada varietas IR42 disebabkan varietas ini merupakan tanaman inang bagi perbanyakan IR42 di laboratorium, sehingga biotipe 3 telah beradaptasi dengan baik pada varietas ini. Menurut Khush dan Virk (2005), IR26 bereaksi tahan terhadap biotipe 3, tetapi tingginya peneluran WBC biotipe 3 pada IR26 dalam
penelitian ini menunjukkan bahwa WBC biotipe 3 dapat mengatasi ketahanan IR26.
WBC meletakkan telurnya dengan cara menusukkan ovipositor ke dalam jaringan tanaman padi. Berdasarkan penelitian Hattori dan Sogawa (2002), terdapat tiga urutan perilaku peletakan telur WBC setelah penusukan ovipositor, yaitu menggerakan ovipositor seperti menggergaji, pengeluaran telur, dan menarik keluar ovipositor. Gerakan seperti menggergaji inilah yang menyebabkan kumpulan telur WBC kumpulan seperti deretan buah pisang.
Luas Sentroid Ovipositor WBC
Ovipositor berfungsi untuk menyisipkan telur ke dalam jaringan pelepah daun padi. Berdasarkan pengukuran morfometri pada ovipositor WBC adalah sebagai berikut (Tabel 3.3):
Tabel 3.3 Luas ovipositor WBC betina biotipe 2 dan biotipe 3 berdasarkan morfometri ovipositor
Populasi WBC Rerata luas sentroid ovipositor ± SD
Uji t
Biotipe 2 291.11 ± 25.31 0,0022*
Biotipe 3 315.61 ± 17.61
Keterangan: tanda * menunjukkan perbedaan nyata luas ovipositor antara kedua biotipe WBC
Hasil analisis morfometri menunjukkan bahwa ovipositor WBC biotipe 3 memiliki luas sentroid ovipositor lebih besar dibandingkan ovipositor WBC biotipe 2. Hasil penelitian terhadap preferensi peneluran (Tabel 3.2) menunjukkan peneluran oleh biotipe 3 lebih banyak daripada biotipe 2, terutama peneluran oleh biotipe 3 pada IR42. Kemungkinannya adalah biotipe 3 telah beradaptasi dengan baik pada IR42 yang merupakan tanaman inangnya. Morfologi ovipositor diduga berkaitan dengan kemampuan menusukkan ovipositor ke dalam jaringan tanaman (Chapman 1998). Pada jangkrik tanah, panjang ovipositor berkaitan dengan kemampuan untuk meletakan telur ke dalam pasir (Reale dan Roff 2002).
Faktor Biofisik Tanaman Padi pada Berbagai Varietas
Pengamatan terhadap faktor biofisik yang diamati yaitu banyaknya trikhoma dan ketebalan epidermis pada irisan membujur pelepah daun berumur 30 hari setiap varietas disajikan pada Tabel 3.4.
Tabel 3.4 Jumlah trikhoma dan tebal epidermis pada pelepah daun berbagai varietas tanaman padi
Varietas Rerata jumlah trikhoma ± SD (/cm)
Rerata tebal epidermis ± SD (µ) a a TN1 6.5 ± 2.68 a 23.14 ± 2.90 b IR26 7.9 ± 3.38 a 24.33 ± 2.72 ab IR42 7.6 ± 3.44 a 24.57 ± 2.31 ab IR64 8.4 ± 4.19 a 25.59 ± 2.33 a IR74 8.6 ± 3.13 a 25.38 ± 2.34 ab PTB33 8.5 ± 3.57 a 26.27 ± 2.06 a Inpari13 7.8 ± 2.39 a 24.25 ± 1.31 ab a
Angka yang diikuti huruf yang sama pada kolom yang sama tidak berbeda nyata pada selang kepercayaan 5% dengan menggunakan uji selang berganda Duncan
Hasil pengamatan terhadap jumlah trikhoma, tidak terdapat perbedaan nyata antar varietas tanaman padi. Perbedaan dalam perilaku WBC pada berbagai varietas tidak terkait dengan jumlah trikhoma tetapi mungkin berkaitan dengan komposisi kimia permukaan daun (Nugaliyadde dan Wilkins 2012). Hasil penelitian Nwilene et al. (2009) mengenai komponen antisenosis pada tanaman padi, diantaranya disebutkan bahwa kepadatan trikhoma pada daun padi tidak berkaitan dengan ketahanan terhadap African rice gall midge, Orseolia oryzivora.
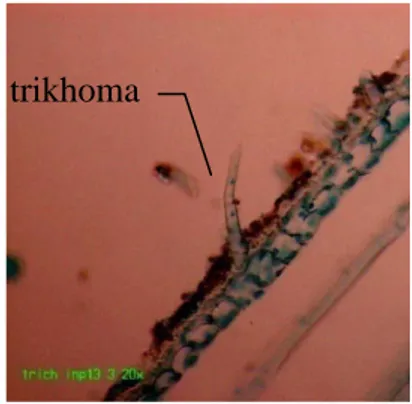
Melalui pengamatan pada struktur trikhoma, terdapat trikhoma dengan bentuk memanjang seperti pada varietas Inpari13 (Gambar 3.1). Jadi dalam mempertimbangkan trikhoma sebagai faktor resistensi hendaknya diobservasi jumlah trikhoma per unit luas area. Selain itu bentuk dan panjang trikhoma perlu diamati.
Tebal epidermis varietas tahan PTB33 (26.27µ) berbeda nyata dengan varietas rentan TN1 (23.14µ), tetapi tebal epidermis varietas tanaman padi IR26, IR42, IR74, IR64 dan Inpari13 tidak berbeda nyata dengan PTB33. Hal ini menunjukkan perbedaan gen ketahanan terhadap WBC dalam varietas tanaman padi tidak berkaitan dengan ketebalan epidermis. PTB33 yang mengandung gen ketahanan bph2 dan Bph3, memiliki tebal epidermis yang tidak berbeda dengan
IR26 (gen Bph1). IR64 dan PTB33 mempunyai tebal epidermis berbeda dengan varietas standar rentan TNI.
Gambar 3.1 Trikhoma pada permukaan pelepah daun varietas Inpari13 berdasarkan irisan membujur (perbesaran 200x)
Faktor-faktor biofisik tanaman yang berperan dalam pertahanan tanaman terhadap herbivor diantaranya trikhoma lilin permukaan, ketebalan jaringan (Smith 2005). Dalam penelitian ini yang diamati adalah trikhoma dan ketebalan jaringan, dari hasil penelitian diduga gen ketahanan varietas padi tidak berkaitan dengan banyaknya trikhoma dan ketebalan epidermis, hanya PTB33 yang menunjukkan perbedaan ketebalan dengan varietas standar rentan TN1.
Kesimpulan
Dalam proses seleksi tanaman inang, WBC menunjukkan reaksi non-preferen terhadap varietas tahan PTB33, IR74, IR64, dan Inpari13. WBC biotipe 2 lebih banyak meletakkan telur pada varietas rentan TN1 dan IR26 dibandingkan pada varietas tahan PTB33 dan IR74. WBC biotipe 3 juga meletakkan telur lebih banyak pada TN1 dan IR42 daripada PTB33, IR74, IR64 dan Inpari13. Besar ovipositor WBC juga berperan dalam merespon ketahanan tanaman padi, yaitu WBC biotipe 3 memiliki luas sentroid ovipositor lebih besar dari biotipe 2. Ketebalan sel epidermis dan jumlah trikhoma pada pelepah daun tidak berperan dalam mekanisme ketahanan tanaman padi terhadap WBC, yaitu dalam hal perilaku hinggap dan peletakan telur.
Daftar Pustaka
Baehaki SE. 2008. Perkembangan wereng batang cokelat Biotipe 4 http://www.litbang.deptan.go.id/artikel/one/171/pdf/perkembangan%20Wer eng%20Cokelat%20Biotipe%204.pdf [10 Juli 2008].
Chapman RF. 1998. The Insect Structure and Function. Ed ke 4. Cambridge (UK): Cambridge University Press.
Hattori M, Sogawa K. 2002 Oviposition behavior of the rice brown planthopper, Nilaparvata lugens (Stål), and its electronic monitoring. Journal of Insect Behavior 15 (2): 283-293.
Heinrichs EA, Medrano FG, Rapusas HR.1985. Genetic Evaluation for Insect Resistance in Rice. Los Banos (Philippines): IRRI.
Heinrichs EA. 1992. Rice insects: the role of host plant resistance in integrated management systems. Kor J Appl Entomol 31(3):256-275.
Khush GS, Virk PS. 2005. IR Varieties and Their Impact. Los Baños (Philippines): IRRI.
Nwilene FE, Okhidevbie O, Agunbiade TA. 2009. An antisenosis component of rice resistance to African rice gall midge, Orselia oryzivora. IRRN.
Nugaliyadde L, Wilkins RM. Influence of surface lipid of some rice varieties on the feeding behavior of Nilaparvata lugens. http://www.goviya.lk/agri_learning/Paddy/Paddy_Research/Paddy_pdf/P10. pdf [10 Agustus 2012].
Panda N, Khush GS. 1995. Host Plant Resistance to Insects. Oxon: CAB International.
Reale D, Roff DA. 2002. Quantitative genetics of oviposition behaviour and interactions among oviposition traits in the sand cricket. Animal Behaviour 64: 397–406.
Sass, J.E. 1951. Botanichal Microtechnique. 2nd
Saxena RC, Pathak MD. 1979. Factors governing susceptibility and resistance of certain rice varieties to the brown planthopper. In: Brown planthopper: Threat of Rice Production in Asia. Los Banos (Philippines): IRRI. hlm 303-317.
Ed. Iowa: The Iowa State University Press.
Schoonhoven LM, van Loon JJA, Dicke M. 2005. Insect-Plant Biology. Second Edition. NewYork: Oxford University Press.
Smith CM. 2005. Plant Resistance to Arthtropods – Molecular and Conventional Approaches., Netherlands: Springer.
Sogawa K. 1982. The rice brown planthopper: Feeding physiology and host plant interactions. Ann Rev Entomol 27: 49-73.
Sogawa K, Pathak MD. 1970. Menchanism of brown planthopper resistance in Mudgo variety of rice. Appl Entomol Zool 5: 145–158.
Soundararajan RP, Gunathilagaraj K, Chitra N, Maheswaran M, Kadirvel P. 2005. Mechanism and genetics of resistance to brown planthopper, Nilaparvata lugens in rice, Oryza sativa L. – a review. Agric Rev 26 (2): 79 – 91.
Sunarti, S., Rugayah, dan E.F. Tihurua. Studi anatomi dan jenis-jenis Averrhoa di Indonesia untuk mempertegas status taksonominya. Berita Biologi 9 (3): 253-257.
Tantowijoyo W, Hoffman AA. 2010. Variation in morphological characters of two invasive leafminers, Liriomyza huidobrensis and L. sativae, across a tropical elevation gradient. Journal of Insect Science: 11 (69): 1-16.
Woodhead S, Padgham DE. 1988. The effect of plant surface characteristics on resistance of rice to the brown planthopper, Nilaparvata lugens. Entomol Exp Appl 47: 15-22.