DNA BARKODE
Spodoptera exigua
Hubn. POPULASI
DATARAN RENDAH DAN DATARAN TINGGI DI JAWA
ANDI TRISNANDI
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI SKRIPSI DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa skripsi berjudul DNA Barkode Spodoptera exigua Hubn. Populasi Dataran Rendah dan Dataran Tinggi di Jawa adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir skripsi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
ABSTRAK
ANDI TRISNANDI. DNA Barkode Spodoptera exigua Hubn. Populasi Dataran Rendah dan Dataran Tinggi di Jawa. Dibimbing oleh RIKA RAFFIUDIN dan I MADE SAMUDRA.
Spodoptera exigua menginfestasi tanaman bawang merah pada dataran rendah dan bawang daun pada dataran tinggi di Jawa. Berdasarkan morfologi ngengat S. exigua yang menginfestasi tanaman bawang pada dataran tinggi mempunyai ukuran morfologi lebih besar dibandingkan dataran rendah. Tujuan penelitian ini adalah untuk mempelajari variasi genetik pada perbedaan morfologi ngengat S. exigua berdasarkan analisis gen cythocrhome oxydase subunit 1 (COI) sebagai DNA Barkode. Sampel DNA S. exigua dikoleksi dari populasi dataran rendah (Cirebon dan Brebes) dan dataran tinggi (Bandung dan Cipanas) di Jawa dengan menggunakan perangkap berferomon dan kemudian dilakukan proses analisis DNA. Database sekuen gen COI ngengat S. exigua dibandingkan dengan S. exigua sekuen gen COI yang berasal dari S. exigua asal Florida dan Canada. Perbedaan haplotipe ini dapat berfungsi sebagai barkode untuk membedakan variasi di dalam spesies S. exigua di berbagai belahan dunia
Kata kunci: Gen COI, mitokondria, haplotipe, Spodoptera exigua, bawang merah
ABSTRACT
ANDI TRISNANDI. DNA Barcodes of Spodoptera exigua Hubn. in Lowlands and Highlands in Java. Supervised by RIKA RAFFIUDIN and I MADE SAMUDRA.
Spodoptera exigua infests shallot in lowlands and leek in highlands n Java. sequences of moth S. exigua from Japan, China, Thailand, Florida-USA, and Canada. This study obtained a total of 530 bp nucleotides of partial COI gene. Alignment of
Skripsi
sebagai salah satu syarat untuk memperoleh gelar Sarjana Sains
pada
Departemen Biologi
DNA BARKODE
Spodoptera exigua
Hubn. POPULASI
DATARAN RENDAH DAN DATARAN TINGGI DI JAWA
ANDI TRISNANDI
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM INSTITUT PERTANIAN BOGOR
Judul Skripsi : DNA Barkode Spodoptera exigua Hubn. Populasi Dataran Rendah dan Dataran Tinggi di Jawa
Nama : Andi Trisnandi NIM : G34090046
Disetujui oleh
Dr Ir Rika Raffiudin, MSi Pembimbing I
Dr Ir I Made Samudra, MSc Pembimbing II
Diketahui oleh
Dr Ir Iman Rusmana, MSi Ketua Departemen
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah subhanahu wa ta’ala atas segala karunia-Nya sehingga karya ilmiah ini berhasil diselesaikan. Tema yang dipilih dalam penelitian yang dilaksanakan sejak bulan Januari hingga juni 2013 ini ialah DNA barkode, dengan judul DNA barkode Spodoptera exigua di dataran rendah dan dataran tinggi di Jawa.
Terima kasih penulis ucapkan kepada Ibu Dr Ir Rika Raffiudin, MSi dan Bapak Dr Ir I Made Samudra, MSc selaku pembimbing. Di samping itu, penghargaan penulis sampaikan kepada Bapak Suwito dan Bapak Jusuf (staf Balai Besar Bioteknologi dan Pengembangan Sumberdaya Genetik Pertanian) yang telah membantu pelaksanaan penelitian ini. Ungkapan terima kasih juga disampaikan kepada ayah, ibu, dan seluruh keluarga atas segala doa, serta teman – teman Biologi 46, Ical, Kak Gina, Kak Eni, Kak Uce, dan semua pihak yang tidak dapat disebutkan seluruhnya atas segala bantuan dan dukungan kepada penulis
Semoga karya ilmiah ini bermanfaat.
DAFTAR ISI
DAFTAR TABEL vi
DAFTAR GAMBAR vi
DAFTAR LAMPIRAN vi
ABSTRAK ii
PENDAHULUAN 1
Latar Belakang 1
Tujuan Penelitian 2
Manfaat Penelitian 2
METODE PENELITIAN 2
Bahan 2
Ekstraksi dan Presipitasi DNA 2
Amplifikasi DNA 3
Elektroforesis dan Visualisasi DNA 4
Alignment DNA dan analisis filogeni 4
HASIL DAN PEMBAHASAN 4
Koleksi Sampel Ngengat Bawang S. exigua 4
Amplifikasi dan Alignment Sekuen DNA Gen COI 5
Analisis Jarak Genetik dan Konstruksi Filogeni Sekuen DNA Gen COI 9
SIMPULAN DAN SARAN 10
Simpulan 10
Saran 10
DAFTAR PUSTAKA 11
LAMPIRAN 13
RIWAYAT HIDUP 15
DAFTAR TABEL
1 Urutan primer untuk amplifikasi gen COI Spodoptera exigua (No.
Akses GenBank NC019622)... 3
2 Data hasil koleksi S. exigua yang digunakan dalam penelitian ... 4
3 Data sampel ingroup (S. exigua) dan outgroup (S. litura dan S. littoralis) yang digunakan untuk analisis gen COI pada penelitian ini 6
4 Jarak genetik berdasarkan sekuen DNA gen COI ingroup dan outgroup ... 9
DAFTAR GAMBAR
1 Ngengat S. exigua di dataran tinggi dan ngengat S. exigua di dataran rendah………... 52 Amplikon pita DNA gen COI S. exigua..……….6
3 Alignment DNA gen COI S. exigua……… 8
4 Konstruksi pohon filogeni 10
DAFTAR LAMPIRAN
1 Kromatogram sekuen gen COI DNA mitokondria ngengat S. exigua menggunakan primer forward COI-47F 12PENDAHULUAN
Latar Belakang
Ulat bawang Spodoptera exigua Hubn. (Lepidoptera: Noctuidae), merupakan salah satu spesies dari genus Spodoptera yang bersifat kosmopolit, dan polifag yang berasal dari Asia Tenggara. Persebaran S. exigua hampir berada diseluruh belahan bumi kecuali Amerika Selatan (Brown dan Drewhurst 1975). Ulat ini ditemukan di Amerika Utara pada tahun 1876, ketika itu ditemukan di Oregon dan Florida (Capinera 2001).
Persebaran S. exigua diberbagai belahan bumi dibantu oleh sifatnya yang dapat hidup di lebih dari satu tanaman inang. Beberapa tanaman yang menjadi inang dari ulat S. exigua adalah asparagus, kacang, brokoli, cabai, kembang kol, seledri, jagung, bawang, tomat, dan tembakau (Capinera 2001). Ulat S. exigua banyak menginfestasi tanaman bawang di Jawa, antara lain bawang merah dan bawang daun (Kalshoven 1981). Infestasi dari S. exigua dapat menyebabkan kehilangan hasil hingga 57% (Ditjen Hortikultura 2010). Infestasi ulat bawang ditandai dengan bercak putih transparan pada bagian epidermis daun karena daun bagian epidermis dalamnya dimakan.
Pada dataran tinggi, tanaman bawang yang dapat tumbuh dengan baik adalah bawang daun dan pada dataran rendah adalah bawang merah. Berdasarkan morfologi populasi ngengat S. exigua yang menginfestasi tanaman bawang pada dataran tinggi mempunyai ukuran morfologi lebih besar dibandingkan dataran rendah (Samudra 29 November 2012, komunikasi pribadi). Pada penelitian ini digunakan DNA barkode untuk melihat secara molekuler variasi genetik ulat bawang S. exigua Hubn. berdasarkan gen cytochrome oxydase subunit 1 (COI) dengan pengambilan sampel pada empat populasi berdasarkan ketinggian yaitu dua lokasi di dataran rendah (Cirebon dan Berebes) dan dua lokasi di dataran tinggi (Bandung dan Cipanas).
DNA barkode adalah suatu sistem baru yang menjadi salah satu alternatif pelengkap atau komplemen yang dapat memperkuat identifikasi morfologi secara cepat dan akurat (Lahaye et al. 2008). DNA barkode dirancang untuk digunakan untuk mengidentifikasi hampir semua spesies hewan (Hebert et al. 2003). DNA mitokondria bersifat haploid, tidak ada rekombinan, memiliki jumlah kopi yang banyak, dan diturunkan secara maternal (maternal inherited) (Harrison 1989).
2
.
Tujuan Penelitian
Penelitian ini bertujuan untuk mempelajari variasi genetik gen COI Spodoptera exigua Hubn. pada empat populasi di dataran rendah (Brebes dan Cirebon) dan dataran tinggi (Bandung dan Cipanas) di Jawa untuk digunakan sebagai database DNA barkode.
Manfaat Penelitian
Penelitian ini bermanfaat untuk digunakan sebagai database dan sebagai penciri molekuler dari spesies Spodoptera exigua asal Jawa.
METODE PENELITIAN
Penelitian ini dilaksanakan dari bulan Januari sampai dengan Juni 2013 bagian Fungsi Hayati dan Perilaku Hewan Departemen Biologi, Institut Pertanian Bogor.
Bahan
Bahan yang digunakan dalam penelitian ialah DNA yang diekstraksi pada bagian toraks ngengat S. exigua. Sampel diambil dari daerah dataran rendah (Brebes dan Cirebon) dan dataran tinggi (Bandung dan Cipanas). Pengambilan sampel dilakukan dengan menggunakan perangkap berferomon. Feromon exi (yang diperoleh dari BB Biogen), yaitu feromon analog S. exigua betina yang berguna untuk menarik S. exigua jantan. Sampel yang didapatkan kemudian dimasukkan ke dalam tabung 1.5 ml yang berisi alkohol absolut. dalam jaringan dengan pergantian TE dilakukan sebanyak 3 kali.
3 isopropanol 600 µl dan disimpan semalam pada suhu -4 °C.
Tahapan berikutnya ialah presipitasi DNA dilakukan dengan sentrifugasi 11 300 g (30 menit) pada suhu 4 °C untuk mendapatkan endapan DNA. Isopropanol kemudian dibuang dan ditambahkan alkohol 70% (v/v) lalu disentrifugasi 11 300 g (10 menit) pada suhu 4 °C. Setelah itu alkohol 70% dibuang dan pelet DNA dikeringkan dengan cara divakum 650 mm/Hg (30 menit) sampai kandungan alkohol hilang. Selanjutnya endapan DNA murni hasil ekstraksi ditambahkan 20 µl TE (0.5 mM) kemudian disimpan dalam suhu -4 °C untuk dilanjutkan ke tahapan analisis berikutnya.
Amplifikasi DNA
DNA diamplifikasi secara in vitro dengan teknik Polymerase Chain Reaction (PCR) menggunakan mesin thermocycler (ESCO SWIFT MAXI-BLC1). Gen target yang diamplifikasi adalah gen COI, dengan menggunakan primer forward COI-47F dan reverse-916R (Tabel 1). Sekuen kedua primer ini adalah hasil modifikasi dari primer forward COI-45F dan reverse-914R (Nagoshi et al. 2010), yaitu dengan penambahan dua basa pada ujung 3’ (Tabel 1). Modifikasi primer dilakukan karena hasil pita DNA dari gen target yang dihasilkan dengan menggunakan primer dari Nagoshi menghasilkan pita yang tipis dan dengan suhu annealing yang rendah yaitu 48 °C Primer tersebut mengamplifikasi sekuen elongation pada suhu 72°C selama 1 menit, post-elongation selama 2 menit
4
Elektroforesis dan Visualisasi DNA
Hasil amplifikasi DNA kemudian dimigrasikan sebanyak 1 µl dengan poliakrilamid gel elektroforesis (PAGE) 6% menggunakan buffer 1x TBE (Tris 0.5 M, asam borat 0.65 M, EDTA 0.002 M). Visualisasi DNA dengan metode pewarnaan perak nitrat (Byun et al. 2009).
Alignment DNA dan Analisis Filogeni
Hasil Hasil amplifikasi DNA disekuen dengan dikirim kepada perusahaan jasa sekuen. Runutan nukleotida yang diperoleh diedit secara manual berdasarkan kromatogram menggunakan software BioEdit Sequence Alignment dan Genetyx Vs 4.01. Runutan – runutan nukleotida yang telah diedit lalu di alignment menggunakan program Clustal X. Analisis variasi nukleotida dan analisis filogeni menggunakan program MEGA versi 5 (Tamura et al. 2011). Proses pengeditan diawali dengan proses menghilangkan basa yang menumpuk pada hasil kromatogram primer forward dan reverse pada ujung 5’ dan ujung
3’ (Lampiran 1,2). Kemudian dilakukan alignment basa hasil kromatogram primer forward dan reverse per individu. Hasil DNA yang telah sejajar kemudian disejajarkan dan dibandingkan dengan database DNA gen COI dari complete genome mitochondria S. exigua dari Cina (NC019622), Jepang (AB733674), Thailand (FN908004), Canada (GU094753) dan Florida. Sebagai outgroup dalam analisis filogeni digunakan S. litura (HM756093) dan S. littoralis (HM756074). Analisis filogeni gen COI dilakukan dengan metode Neighbour Joining (NJ) dengan bootstrap 1000 x.
HASIL DAN PEMBAHASAN
Koleksi Sampel Ngengat Bawang S. exigua
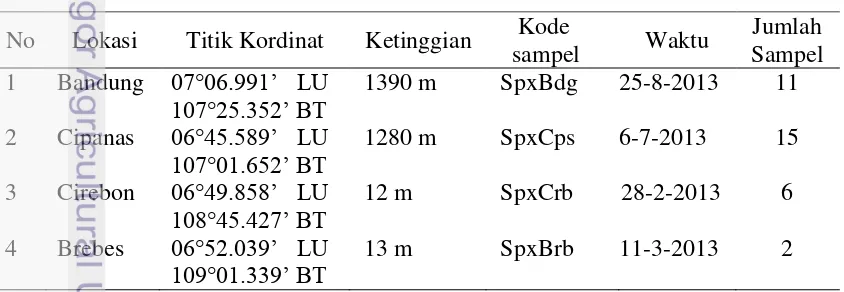
Sampel ngengat bawang S. exigua dikoleksi berdasarkan ketinggian yaitu pada daerah dataran rendah (Cirebon dan Brebes) dan dataran tinggi di Jawa (Bandung dan Cipanas) (Tabel 2). Sampel dikoleksi dengan menggunakan perangkap berferomon exi
5
(a) (b)
Sampel dewasa S. exigua yang dikoleksi dengan perangkap berferomon exi adalah pada fase ngengat (imago). Spodoptera exigua mempunyai siklus hidup yang terdiri atas fase telur, larva, pupa, dan imago (Kalshoven 1981). Telur tersusun atas kelompok telur yang berjumlah 300-600 telur per kelompok. Telur biasanya diletakkan oleh imago betina pada bagian bawah daun di tanaman bawang. Fase telur terjadi selama 2-3 hari. Larva merupakan fase terjadinya infestasi S. exigua pada tanaman bawang. Pada kondisi normal larva terdiri atas 5 instar, perbedaan instar dapat terlihat pada garis yang terdapat dibagian tubuh dorsal larva. Semakin gelap garis yang terbentuk menunjukan larva berada pada instar akhir. Fase larva terjadi selama 11-16 hari. abu-abu. Fase ngengat terjadi selama 3-7 hari (Capinera 2001).
Berdasarkan morfologi ngengat S. exigua yang menginfestasi tanaman bawang, populasi ngengat S. exigua pada dataran tinggi mempunyai ukuran morfologi lebih besar dibandingkan dengan dataran rendah (Samudra 29 November 2012, komunikasi pribadi). Kemudian dilakukan pengukuran morfologi pada panjang sayap ngengat S. exigua pada satu individu di dataran tinggi dan dataran morfologi ngengat S. exigua di dataran tinggi dan dataran rendah.
Amplifikasi dan Alignment Sekuen DNA Gen COI
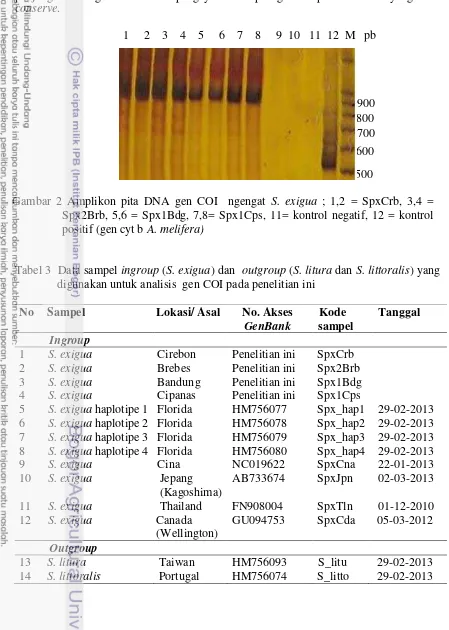
Amplikon gen COI pada ke-empat sampel yaitu, S. exigua Cirebon, Brebes, Bandung dan Cipanas memperlihatkan ukuran pita DNA yang sama panjang. Hasil amplifikasi gen COI dengan primer forward COI-47 F dan reverse COI-916 R menunjukan pita DNA ± 900 pb (Gambar 2). Kemudian alignment sekuen dilakukan antara sekuen gen COI ingroup (SpxCrb, Spx2Brb,
6
Spx1Bdg, Spx1Cps, Spx haplotipe Florida, Spx Cina, Spx Jepang, Spx Thailand serta Spx Canada) dan outgroup (S. litura dan S. littoralis) (Tabel 3). Hasil dari alignment antara sekuen gen COI ingroup dan outgroup dihasilkan panjang sekuen sebesar 530 pb. Terdapat pemotongan sekuen gen COI dari 900 pb menjadi 530 pb. Pemotongan sekuen dilakukan untuk menyesuaikan dengan panjang sekuen gen COI dari Jepang yaitu 530 pb agar didapatkan daerah yang conserve.
Tabel 3 Data sampel ingroup (S. exigua) dan outgroup (S. litura dan S. littoralis) yang digunakan untuk analisis gen COI pada penelitian ini
No Sampel Lokasi/ Asal No. Akses
5 S. exigua haplotipe 1 Florida HM756077 Spx_hap1 29-02-2013 6 S. exigua haplotipe 2 Florida HM756078 Spx_hap2 29-02-2013 7 S. exigua haplotipe 3 Florida HM756079 Spx_hap3 29-02-2013 8 S. exigua haplotipe 4 Florida HM756080 Spx_hap4 29-02-2013
9 S. exigua Cina NC019622 SpxCna 22-01-2013
10 S. exigua Jepang
(Kagoshima)
AB733674 SpxJpn 02-03-2013
11 S. exigua Thailand FN908004 SpxTln 01-12-2010
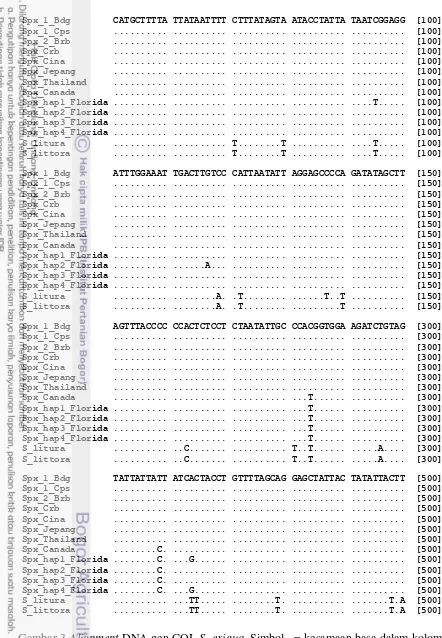
7 Berdasarkan hasil alignment sampel S. exigua di Jawa dari penelitian ini baik antara sampel di dataran rendah dan dataran tinggi tidak memperlihatkan adanya variasi nukleotida. Dengan demikian, data gen COI S. exigua dari hasil penelitian ini dapat menjadi ciri gen COI untuk S. exigua asal Jawa. Penggunaan gen COI pada penelitian ini merupakan gen yang digunakan pada analisis dengan DNA Barkode. DNA barkode merupakan suatu cara untuk mengidentifikasi spesies secara cepat dan akurat dengan menggunakan DNA pendek yang distandardisasi (Hebert et al. 2005). Pemilihan gen COI di dalam analisis menggunakan DNA barkode karena gen COI yang digunakan relatif pendek yaitu 648 pb, bersifat conserve, relatif stabil tidak mudah mengalami perubahan-perubahan dengan gen-gen mitokondria yang lainnya, dan gen COI terdapat dalam jumlah kopi yang banyak (Hebert et al. 2003). Beberapa penelitian lain menunjukan bahwa gen COI dapat mengidentifikasi spesies kompleks. Pada contoh lain, Hausman et al. (2011) menggunakan DNA barkode untuk monitoring keragaman dan identifikasi spesies kupu-kupu dan ngengat (Lepidoptera) di German. Hampir (99%) dari 957 spesies diidentifikasi dengan menggunakan sekuen DNA barkode.
Gen COI diturunkan secara meternal, sehingga tidak adanya variasi nukleotida pada gen COI pada populasi S. exigua yang terdapat di Jawa disebabkan asam amino biasanya terjadi pada perubahan nukleotida pada posisi kodon pertama dan kedua, sedangkan posisi ketiga jarang menyebabkan terjadinya perubahan. Peluang terjadinya mutasi sinonim pada nukleotida kodon pertama 5%, pada kodon ketiga sebesar 72% dan pada kodon kedua selalu menyebabkan asam amino berubah (Nei 1987).
Selanjutnya, berdasarkan alignment antara sekuen gen COI hasil penelitian ini dengan sekuen gen COI asal negara Asia lain seperti Thailand, Jepang, dan Cina juga tidak memperlihatkan adanya variasi nukleotida, kecuali satu pada populasi Cina (data tidak dipublikasikan). Hasil alignment antara sekuen Asia dengan sekuen Florida dan Canada menunjukan variasi nukleotida gen COI yang tinggi (Gambar 3). Variasi nukleotida yang tinggi banyak terdapat pada sekuen gen COI asal Florida dan Canada. Variasi nukleotida terdapat pada basa ke 95, 117, 284, 459 dan 464. Pada basa ke 95 terjadi mutasi titik C–T pada kodon ke-2 sehingga mengubah asam amino serina menjadi leusina, basa ke-117 mutasi titik G–A pada kodon ke 3 tidak menyebabkan asam amino berubah yaitu leusina, nukleotida ke-284 terjadi mutasi titik C–T mengubah asam amino treonina menjadi metionina, nukleotida ke-459 terjadi mutasi titik T–C pada kodon ke-3 menyebabkan asam amino tidak berubah yaitu tirosina dan pada basa ke-464 mutasi titik A–G pada kodon ke-2 yaitu histidin menjadi arginin.
8
Spx_1_Bdg CATGCTTTTA TTATAATTTT CTTTATAGTA ATACCTATTA TAATCGGAGG [100]
Spx_1_Cps ... ... ... ... ... [100]
Spx_1_Bdg ATTTGGAAAT TGACTTGTCC CATTAATATT AGGAGCCCCA GATATAGCTT [150] Spx_1_Cps ... ... ... ... ... [150]
Spx_1_Bdg AGTTTACCCC CCACTCTCCT CTAATATTGC CCACGGTGGA AGATCTGTAG [300] Spx_1_Cps ... ... ... ... ... [300]
9 Berdasarkan distribusi sebaran S. exigua secara geografis di dunia, daerah distribusi S. exigua meningkat sebanyak 34 negara dari 67 negara sejak tahun 1972 (Zheng 2011). Meningkatnya distribusi S. exigua dipengaruhi oleh adanya strategi adaptasi terhadap musim dan juga tanaman inang yang beragam.
Analisis Jarak Genetik dan Konstruksi Filogeni Sekuen DNA Gen COI Analisis jarak genetik dilakukan berdasarkan sekuen DNA gen COI ingroup (S. exigua) dan outgroup (S. littura dan S. littoralis). Jarak genetik tertinggi dihasilkan dari sekuen gen COI intraspesies antara S. exigua haplotipe 1 Floridadengan S. exigua asal Asia sebesar 0.008 (0.8%) dan jarak genetik terendah dihasilkan dari sekuen DNA gen COI kelompok asia sebesar 0.000 (0%) (Tabel 4). Kemudian, jarak genetik gen COI spesies S. exigua pada penelitian ini yang dikoleksi dari populasi dataran rendah dan dataran tinggi di Jawa tidak menunjukan jarak genetik yang berbeda yaitu 0.000 (0%).
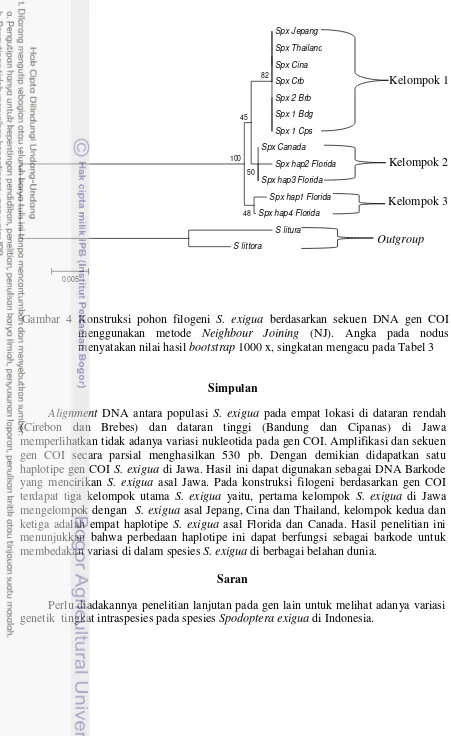
Hasil konstruksi filogeni berdasarkan gen COI (Gambar 4) menunjukan terdapat tiga kelompok utama S. exigua yaitu, pertama kelompok S. exigua di Jawa mengelompok dengan S. exigua asal Jepang, Cina, dan Thailand dengan nilai bootstrap 82 %. Kelompok kedua adalah dua haplotipe (haplotipe 2 dan 3) S. exigua asal Florida dan Canada dengan nilai bootstrap 50%. Kelompok ketiga adalah dua haplotipe lain dari sampel S. exigua dari Florida (haplotipe 1 dan 4) dengan nilai bootstrap 48%. Konstruksi filogeni berdasarkan sekuen DNA gen COI memperlihatkan pengelompokkan yang terpisah dengan nilai bootstrap 100 % antara sekuen ingroup (S. exigua dari berbagai lokasi) serta outgroup yaitu S. littura dan S. littoralis (Nagoshi et al. 2010).
Tabel 4 Jarak genetik berdasarkan sekuen DNA gen COI ingroup dan outgroup
10 memperlihatkan tidak adanya variasi nukleotida pada gen COI. Amplifikasi dan sekuen gen COI secara parsial menghasilkan 530 pb. Dengan demikian didapatkan satu haplotipe gen COI S. exigua di Jawa. Hasil ini dapat digunakan sebagai DNA Barkode yang mencirikan S. exigua asal Jawa. Pada konstruksi filogeni berdasarkan gen COI terdapat tiga kelompok utama S. exigua yaitu, pertama kelompok S. exigua di Jawa mengelompok dengan S. exigua asal Jepang, Cina dan Thailand, kelompok kedua dan ketiga adalah empat haplotipe S. exigua asal Florida dan Canada. Hasil penelitian ini menunjukkan bahwa perbedaan haplotipe ini dapat berfungsi sebagai barkode untuk membedakan variasi di dalam spesies S. exigua di berbagai belahan dunia.
Saran
Perlu diadakannya penelitian lanjutan pada gen lain untuk melihat adanya variasi genetik tingkat intraspesies pada spesies Spodopteraexigua di Indonesia.
Gambar 4 Konstruksi pohon filogeni S. exigua berdasarkan sekuen DNA gen COI menggunakan metode Neighbour Joining (NJ). Angka pada nodus menyatakan nilai hasil bootstrap 1000 x, singkatan mengacu pada Tabel 3
11
DAFTAR PUSTAKA
Brown ES, Drewhurst CF. 1975. The genus Spodoptera (Lepidoptera, Noctuidae) in Africa and the Near East. Bull of Entomol Res. 65: 221-262
Byun SO, Fang Q, Zhou H, Hickford JGH. 2009. An efeective method for silver-staining DNA in large numbers of polyacrylamide gels. Anal Biochem. 385: 174-175.
Capinera JL. 2001. Handbook of Vegetable Pests. San Diego (US): Academic press. [Ditjen Hortikultura] Direktorat Jenderal Hortikultura. 2010. Statistik Produksi
Hortikultura Tahun 2009. Jakarta (ID): Dirjen Hortikultura Pasar Minggu.
Hajibabaei M, Janzen DH, Burns JM, Hallwachs W, Hebert PDN. 2006.DNA barkode distinguish species of tropical Lepidoptera. PNAS 103: 968-971.
Harrison RG. 1989. Animal mitochondrial DNA as a genetic marker in population and evolutionary biology. Tre Ecol Evol. 4: 6-11.
Hebert PDN, Cywin SA, Ball SL, Ward JR. 2003. Biological idntifications through DNA barcodes. Proc R Soc. 270: 313-321.
Hausmann A, Haszprunar G, Segerer AH, Speidel W, Behounek G, Hebert PDN. 2011. Now DNA Barcoded : the butterflies dan large moths of Germany. Spixiana 34: 47-58.
Hebert PDN, Gregory TR. 2005. The promise of DNA barcoding for taxonomy. Syst Biol. 54: 852-859.
Kalshoven LGE. 1981. The Pests of Crops In Indonesia. Jakarta (ID): PT. Ichtiar Baru. Lahaye R, Michelle VDB, Bogarin D, Warner J, Pupulin F, Gigot G, Maurin O, Duthoit S, Barraclough TG, Savolainen V. DNA barcoding the floras of biodiversity hotspots. PNAS 105: 2923-2928.
Nagoshi RN, Brambila J, Meager RL. 2010. Use of DNA barcodes to identify invasive armyworm Spodoptera spesies in Florida. J Ins Sci. 11: 1536-2442.
Nei M. 1987. Molecular evolutionary genetics. New York (US): Columbia University Press.
Raffiudin R, Crozier. 2007. Phylogenetic analysis of honeybee behavioural evolution. Mol Phylogen Evol 48: 543-552.
Sambrook J, Fritsch EF, Maniatis T. 1989. Molecular Cloning: A Laboratorium ManualSecond Edition. New York: Cold Spring Harbor Laboratory Press.
Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S. 2011. MEGA5: Molecular evolutionary Genetics Analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol Biol Evol. 28:2731-2739.
12
Ward RD, Zemlak TS, Innes BH, Last PR, Hebert PDN. 2005. DNA barcoding Australia fish spesies. Phil Trans R Soc B 360: 1847- 1857.
Zheng XL, Cong XP, Wang XP, Lei CL. 2011. A Review of Geographic Distribution, Overwintering and Migration in Spodoptera exigua Hübner (Lepidoptera: Noctuidae). J Ento Mol Res Soc.13: 39-48.
13
LAMPIRAN
14
Lampiran 2 Kromatogram sekuen gen COI DNA Mitokondria ngengat S. exigua menggunakan primer reverse COI-916R
15