15 ABSTRACT
DWI FEBRIANTI. Effectiveness of intestinal shrimp probiotics in inhibiting growth of Vibrio harveyi in shrimp larvae Litopenaeus vannamei. Supervised by MUNTI YUHANA and WIDANARNI.
Vibriosis is one of infectious disease that often attacks the shrimp vaname culture. It caused by Vibrio harveyi. One of alternative in controlling the luminous disease is the application of probiotic bacteria which is considered as enviromental friendly treatment. This research was carried out to evaluate the effectiveness of four strains probiotic in inhibiting growth of Vibrio harveyi in shrimp larvae Litopenaeus vannamei. The study design used was the complete random design with six treatments: the addition of isolates F5, F17, F19, F43, positive control (the addition of V. harveyi MR 5339 Rf R), and the negative control (without addition of probiotics and V. harveyi MR 5339 Rf R). Based on the observations, the survival rate of shrimp post larvae treatment with four candidate probiotics have a significant difference survival rate (P <0,05) when compared with positive control (67,5±6,45 %). The survival rate of post larvae
treatment with candidate probiotics were ranging from 78,75±8,54 - 92,50±6,45 %. Daily growth rate of length and weight have no
significant differences in all treatments.
20
I. PENDAHULUAN
1.1 Latar Belakang
Udang vaname (Litopenaeus vannamei) merupakan salah satu komoditas ekspor unggulan Kementrian Kelautan dan Perikanan yang diharapkan mengalami peningkatan produksi sebesar 209% dalam kurun waktu 2009-2014 (KKP 2010) dari 201% target total peningkatan produksi udang Indonesia. Peningkatan produksi udang vaname diharapkan mencapai 16% pertahunnya. Produksi udang vaname pada tahun 2014 menurut KKP (2010) diproyeksikan sebesar 511 ton. Udang vaname dipilih sebagai komoditas unggulan karena memiliki beberapa kelebihan diantaranya banyak diminati oleh pasar lokal maupun internasional, lebih tahan terhadap penyakit jika dibandingkan dengan jenis udang putih lainnya, pertumbuhan relatif lebih cepat, serta memiliki toleransi yang cukup besar terhadap perubahan kualitas lingkungan (Atjo 2009).
Adanya perubahan kualitas lingkungan dan serangan penyakit menyebabkan timbulnya kendala dalam sistem budidaya udang vaname. Penyakit vibriosis merupakan salah satu jenis penyakit bakterial yang menyerang seluruh siklus hidup udang vaname dan telah menyebabkan tingginya tingkat kematian pada industri budidaya udang dunia (Heidarieh et al. 2010). Vibriosis merupakan salah satu jenis penyakit infeksius yang sering menyerang organisme akuatik air laut seperti ikan dan udang. Penyakit ini umumnya disebabkan oleh anggota genus Vibrio (Munn 1977) seperti Vibrio harveyi, Vibrio alginolyticus, Vibrio parahaemolyticus, dan Vibrio penaeicida (Longyant et al. 2008). Vibrio harveyi merupakan patogen dari genus Vibrio yang berasosisasi pada penyakit udang berpendar (Austin and Zhang 2006).
21 penyuntikan intramuskular, sedangkan pada konsentrasi >106-107 cfu/ml dapat menyebabkan kematian pada udang vaname (Litopenaeus vaname) melalui infeksi dengan teknik perendaman selama 2 jam (Saulnier et al. 2000).
Antibiotik merupakan salah satu bahan yang sering digunakan untuk pencegahan dan penanggulangan penyakit bakterial pada kegiatan budidaya udang. Penggunaan antibiotik seperti oxytetracycline, oxolinic acid, chloramphenicol dan furazolidone umumnya digunakan untuk menanggulangi penyakit vibriosis (Tendencia and de la Pena 2001). Menurut Serrano (2005) dalam Reboucas et al. (2011), patogen penyebab vibriosis yang terpapar antibiotik baik di dalam maupun di luar lingkungan hatchery pemeliharaan udang dapat menyebabkan timbulnya sifat resistensi melalui perpindahan materi genetik.
Salah satu alternatif pengendalian penyakit bakterial yang dapat dilakukan adalah dengan menggunakan bakteri probiotik. Beberapa mekanisme kerja bakteri probiotik dalam menghambat keberadaan bakteri patogen diantaranya menghasilkan senyawa penghambat, kompetisi memperoleh sumber energi, kompetisi tempat perlekatan, meningkatkan respon imun, serta memperbaiki kualitas air (Verschuere et al. 2000). Bakteri yang berhasil digunakan sebagai probiotik diantaranya berasal dari genus Vibrio, Bacillus spp., dan Thallasobacter utilis. Bakteri probiotik dapat diisolasi dari berbagai sumber diantaranya air
pemeliharaan udang ataupun saluran pencernaan udang itu sendiri (Guillan et al. 2004) ataupun terumbu karang (Sasanti 2008).
1.2 Tujuan
22 II. BAHAN DAN METODE
2.1 Seleksi Bakteri Probiotik
2.1.1 Karakterisasi morfologi dan fisiologis kandidat probiotik
Sebanyak 16 jenis bakteri hasil isolasi Ardiani (2011) ditumbuhkan pada media agar Sea Water Complete (SWC) lalu diinkubasi selama 24 jam pada suhu 28oC. Koloni yang tumbuh dilakukan serangkaian uji untuk mengetahui karakter morfologi dan fisiologis dari masing-masing isolat. Uji yang dilakukan meliputi (1) pewarnaan Gram dengan menggunakan larutan kristal violet, lugol, alkohol absolut, dan safranin (2) uji oksidatif-fermentatif dengan menggunakan media uji oksidatif-fermentatif, glukosa, dan parafin; (3) uji motilitas dengan menggunakan media Sulfide Indol Motility (SIM); (4) uji katalase dengan menggunakan
Hidrogen Peroksida (H2O2) 3%; dan (5) uji oksidase dengan menggunakan p-aminodhymethylalanin oxalat 1%.
2.1.2 Uji Aktivitas amilolitik dan proteolitik
Uji ini dilakukan untuk mengetahui aktivitas amilase dan protease dari masing-masing isolat melalui kemampuannya menghidrolisis karbohidrat dan protein. Bakteri kandidat probiotik ditumbuhkan pada media SWC yang telah ditambahkan tepung kanji sebanyak 2% untuk uji amilolitik. Kemampuan menghidrolisa karbohidrat ditandai dengan terbentuknya warna kuning terang di sekitar isolat sedangkan isolat yang tidak mampu menghidrolisa karbohidrat, area di sekitar isolat tetap berwarna gelap setelah penambahan reagen KI 1%. Sedangkan pada uji aktivitas proteolitik, bakteri kandidat probiotik ditumbuhkan pada media SWC yang telah ditambahkan susu skim sebanyak 2%. Kemampuan menghidrolisa protein ditandai dengan terbentuknya zona bening di sekitar isolat.
2.1.3 Uji sensitivitas antibiotik
23 pemeliharaan pascalarva. Jenis antibiotik yang digunakan adalah rifamfisin, streptomisin, kanamisin, dan penisilin G. Konsentrasi yang digunakan adalah 50 µg/ml dan 100 µg/ml. Kandidat probiotik ditumbuhkan pada media SWC agar yang telah mengandung antibiotik dengan konsentrasi 50 µg/ml dan 100 µg/ml menggunakan metode streak plate lalu diinkubasi selama 24 jam pada suhu 28oC. Bakteri yang sensitif terhadap antibiotik ditandai dengan tidak adanya pertumbuhan koloni pada media SWC agar yang telah mengandung antibiotik, sedangkan bakteri yang resisten akan tetap tumbuh pada media tersebut.
2.1.4 Metode kultur bersama
Kemampuan kandidat bakteri probiotik melawan bakteri V. harveyi MR 5339 Rf R diuji juga dengan menggunakan metode kultur bersama. Isolat bakteri kandidat probiotik dan bakteri V. harveyi MR 5339 Rf R yang telah berumur 24 jam diencerkan hingga mencapai kepadatan yang sama yakni mencapai 106 CFU/ml. Sebanyak 100 µl bakteri kandidat probiotik dan bakteri V. harveyi MR 5339 Rf R dimasukkan kedalam media SWC broth, lalu diinkubasi selama 24 jam pada shaker berkecepatan 140 rpm dengan suhu 29oC. Sebagai kontrol digunakan bakteri V. harveyi MR 5339 Rf R yang ditumbuhkan sendiri (tanpa bakteri kandidat probiotik) dengan kepadatan yang sama. Kemampuan bakteri kandidat probiotik menghambat pertumbuhan bakteri V. harveyi MR 5339 Rf R ditandai dengan kepadatan bakteri V. harveyi MR 5339 Rf R pada perlakuan kontrol yang lebih banyak dibandingkan dengan bakteri V. harveyi MR 5339 Rf R pada hasil kultur bersama.
2.1.5 Uji patogenitas bakteri kandidat probiotik
24 V. harveyi MR 5339 Rf R. Pemberian pakan dilakukan sebanyak 5 kali sehari dengan menggunakan pelet berjenis crumble. Pengamatan tingkat kelangsungan hidup dilakukan setiap hari selama 5 hari (Vijayan et al. 2006).
2.1.6 Pembuatan mutan bakteri kandidat probiotik
Isolat-isolat yang menghasilkan kelangsungan hidup terbaik pada uji patogenisitas, diberi penanda resisten streptomisin (St R) terlebih dahulu sebelum digunakan pada uji in vivo. Pemberian penanda streptomisin dilakukan dengan menumbuhkan isolat secara spontan pada media SWC agar yang mengandung streptomisin sebanyak 50 µg/ml media. Sedangkan patogen yang digunakan adalah V. harveyi MR 5339 Rf R yang ditumbuhkan secara spontan pada media SWC yang mengandung 50 µg/ml rifamfisin. Pemberian penanda ini dilakukan untuk memonitor keberadaan bakteri pada media pemeliharaan.
2.2 Uji in vivo Kandidat Probiotik
Isolat bakteri kandidat probiotik diinokulasikan pada media pemeliharaan hingga mencapai konsentrasi akhir 106 CFU/ml sehari setelah larva udang dimasukkan. Patogen V. harveyi MR 5339 Rf R diinokulasikan hingga mencapai kepadatan akhir 106 CFU/ml enam jam setelah pemberian kandidat probiotik. Percobaan dilakukan sebanyak empat kali ulangan termasuk kontrol positif (dengan inokulasi bakteri V. harveyi MR 5339 Rf R saja) dan kontrol negatif (tanpa penambahan bakteri kandidat probiotik maupun V. harveyi MR 5339 Rf R). Pengamatan dilakukan selama 14 hari meliputi tingkat kelangsungan hidup (SR), total populasi bakteri V. harveyi MR 5339 Rf R, total populasi bakteri kandidat probiotik, serta laju pertumbuhan panjang dan bobot harian.
2.3 Rancangan Penelitian dan Teknik Analisa
25 2.4 Parameter yang Diamati
2.4.1 Tingkat kelangsungan hidup pascalarva udang vaname
Tingkat kelangsungan hidup pascalarva udang vaname dihitung dengan menggunakan rumus sebagai berikut:
SR =
x 100% (Effendi 2004)
Keterangan:
SR = tingkat kelangsungan hidup (%)
Nt = jumlah pascalarva udang hidup pada akhir pengamatan No = jumlah pascalarva udang pada awal pengamatan
2.4.2 Pertumbuhan panjang dan bobot harian pascalarva udang vaname Pertumbuhan panjang dan bobot total diamati pada awal dan akhir penelitian. Pertumbuhan bobot pascalarva udang vaname dihitung berdasarkan pertambahan bobot berdasarkan rumus berikut :
SGR = { √
} (Effendie 1997)
Keterangan:
SGR = laju pertumbuhan harian bobot larva udang (%) t = lama waktu pemeliharaan larva udang (hari) Wt = bobot rata-rata akhir pascalarva udang (mg) Wo = bobot rata-rata awal pascalarva udang (mg)
Nilai pertumbuhan panjang harian (Daily increment in length) pascalarva udang vaname dihitung dengan menggunakan rumus sebagai berikut:
DIL = (Mai et al. 2000) DIL = pertumbuhan panjang harian
Lt = panjang rata-rata akhir pascalarva udang (mm) Lo = panjang rata-rata awal pascalarva udang (mm)
2.4.3 Populasi bakteri
27 III. HASIL DAN PEMBAHASAN
3.1 Karakterisasi Morfologi dan Fisiologis Kandidat Probiotik
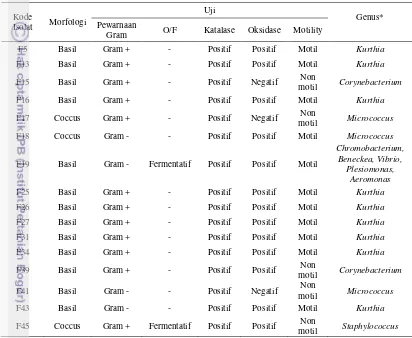
Hasil karakterisasi morfologi dan fisiologis yang dilakukan terhadap 16 jenis bakteri hasil isolasi Ardiani (2011) tertera pada tabel 1.
Tabel 1. Karakterisasi morfologi dan fisiologis kandidat probiotik
Kode
Isolat Morfologi
Uji
Genus* Pewarnaan
Gram O/F Katalase Oksidase Motility
F5 Basil Gram + - Positif Positif Motil Kurthia
F13 Basil Gram + - Positif Positif Motil Kurthia
F15 Basil Gram + - Positif Negatif Non
motil Corynebacterium F16 Basil Gram + - Positif Positif Motil Kurthia
F17 Coccus Gram + - Positif Negatif Non
motil Micrococcus F18 Coccus Gram - - Positif Positif Motil Micrococcus
F19 Basil Gram - Fermentatif Positif Positif Motil
Chromobacterium, Beneckea, Vibrio,
Plesiomonas, Aeromonas
F25 Basil Gram + - Positif Positif Motil Kurthia
F26 Basil Gram + - Positif Positif Motil Kurthia
F27 Basil Gram + - Positif Positif Motil Kurthia
F31 Basil Gram + - Positif Positif Motil Kurthia
F34 Basil Gram + - Positif Positif Motil Kurthia
F39 Basil Gram + - Positif Positif Non
motil Corynebacterium
F41 Basil Gram - - Positif Negatif Non
motil Micrococcus F43 Basil Gram - - Positif Positif Motil Kurthia
F45 Coccus Gram + Fermentatif Positif Positif Non
motil Staphylococcus *Mengacu pada metode identifikasi Cowan (1974)
** Hasil (-) menunjukkan tidak ada reaksi yang terjadi
28 F41, dan F45 bersifat tidak motil sedangkan isolat lainnya bersifat motil (mampu bergerak). Berdasarkan identifikasi tingkat genus yang mengacu pada metode Cowan (1974), isolat berhasil diidentifikasi sebagai anggota dari genus Kurthia, Corynebacterium, Micrococcus, Chromobacterium, Beneckea, Vibrio, Plesiomonas, Aeromonas, dan Staphylococcus.
3.2 Uji Aktivitas Amilolitik dan Proteolitik
Aktivitas amilase ditandai dengan terbentuknya zona berwarna kuning terang disekitar isolat sedangkan isolat yang tidak mampu menghidrolisa karbohidrat, area disekitar isolat tetap berwarna gelap setelah penambahan reagen KI. Sedangkan aktivitas protease ditandai dengan terbentuknya zona bening disekitar isolat. Hasil uji amilolitik dan proteolitik disajikan pada gambar 1 di bawah ini.
a. b.
Gambar 1. Hasil aktivitas (a) amilolitik dan (b) proteolitik
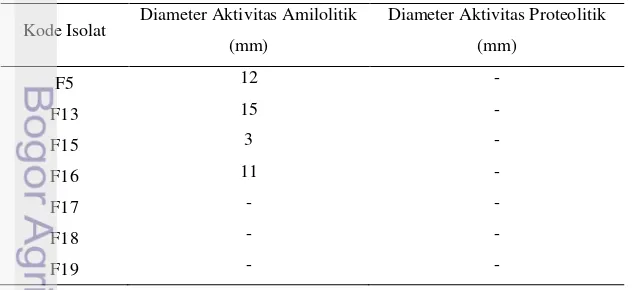
Hasil pengukuran diameter aktivitas amilolitik dan proteolitik dapat dilihat pada tabel 2.
Tabel 2. Diameter aktivitas amilolitik dan proteolitik kandidat probiotik
Kode Isolat Diameter Aktivitas Amilolitik (mm)
Diameter Aktivitas Proteolitik
(mm)
F5 12 -
F13 15 -
F15 3 -
F16 11 -
F17 - -
F18 - -
29 Tabel 2. Diameter aktivitas amilolitik dan proteolitik kandidat probiotik (lanjutan)
Kode Isolat Diameter Aktivitas Amilolitik (mm)
Diameter Aktivitas Proteolitik
(mm)
F25 6 -
F26 11 -
F27 10 -
F31 12 -
F34 - 15
F39 5 19
F41 - -
F43 - -
F45 - -
Berdasarkan hasil uji amilolitik isolat diketahui bahwa isolat yang memiliki aktivitas amilolitik terbesar berturut-turut terdapat pada isolat F13 dengan diameter zona amilolitik sebesar 15 mm, serta F5 dan F31 dengan diameter zona amilolitik sebesar 12 mm. Aktivitas proteolitik hanya terdapat pada isolat F34 dan F39. Aktivitas proteolitik terbesar terdapat pada isolat F39 dengan diameter zona sebesar 19 mm diikuti dengan isolat F34 dengan diameter zona sebesar 15 mm. Adanya kemampuan menghidrolisis karbohidrat dan protein menunjukkan isolat-isolat tersebut mampu memanfaatkan sumber energi berupa pati dan protein yang ditambahkan pada media menjadi sumber karbon.
Menurut Price dan Stevens (1996) dalam Yandri et al. (2008), protease merupakan enzim yang berfungsi memecah ikatan peptida untuk menghasilkan asam amino dan peptida sederhana lainnya. Enzim ini dapat diisolasi dari berbagai sumber seperti tanaman, hewan, dan mikroba (fungi dan bakteri). Protease atau disebut juga endopeptidase merupakan salah satu enzim pencernaan yang penting, termasuk di dalamnya tripsin dan kemotripsin yang bertanggung jawab terhadap hampir 60% dari proses pencernaan udang (Lemos et al. 2000).
3.3 Uji Sensitivitas Antibiotik
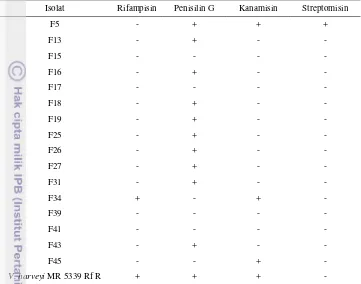
30 antibiotik. Konsentrasi antibiotik yang digunakan sebesar 50 µg/ml dan 100 µg/ml media. Hasil uji sensitivitas disajikan pada tabel di bawah ini.
Tabel 3. Hasil uji sensitivitas antibiotik pada konsentrasi 50 µg/ml
Isolat Rifampisin Penisilin G Kanamisin Streptomisin
F5 - + + +
F13 - + - -
F15 - - - -
F16 - + - -
F17 - - - -
F18 - + - -
F19 - + - -
F25 - + - -
F26 - + - -
F27 - + - -
F31 - + - -
F34 + - + -
F39 - - - -
F41 - - - -
F43 - + - -
F45 - - + -
V. harveyi MR 5339 Rf R + + + -
Keterangan: (-):sensitif; (+):resisten
Berdasarkan tabel 3, isolat F34 dan V. harveyi MR 5339 Rf R bersifat resisten terhadap antibiotik rifampisin. Isolat F15, F17, F34, F39, F41, F45 bersifat resisten terhadap penisilin G. Isolat F5, F34, F45 dan V. harveyi MR 5339 Rf R bersifat resisten terhadap kanamisin sedangkan semua isolat (kecuali isolat F5) bersifat sensitif terhadap streptomisin. Sifat resisten terhadap antibiotik pada bakteri dapat disebabkan oleh dua jenis mekanisme yaitu mutasi kromosom dan akuisisi plasmid. Mutasi pada kromosom tidak dapat ditransfer pada bakteri lain tetapi plasmid mampu mentransfer dengan cepat sifat resisten tersebut (Lewin 1992 dalam Schnick 2001). Sifat resistensi pada mikroorganisme juga diakibatkan oleh: (1) kemampuan menghasilkan enzim yang mampu menginaktivasi antibiotik; (2) adanya penambahan substansi kimia pada struktur kimia lain oleh
31 (3) perubahan struktur protein pada dinding sel; (4) berkurangnya permeabilitas dinding sel sehingga antibiotik tidak dapat masuk ke dalam sel (Guilfoile 2007).
Sifat resistensi ini dapat dipindahkan baik secara horizontal maupun vertikal (Madigan et al. 2003 dalam Reboucas et al. 2011). Baticados et al. (1990) dalam Tendencia and de la Pena (2001) melaporkan bahwa patogen penyebab luminous disease seperti Vibrio harveyi dan Vibrio splendidus yang diisolasi dari larva udang resisten terhadap erythromycin, kanamycin, pencillin G, dan streptomycin.
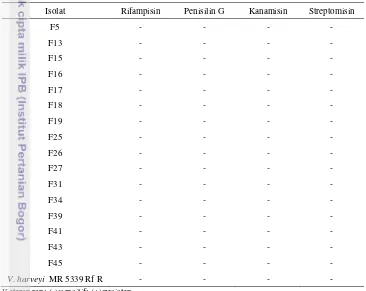
Tabel 4. Hasil uji sensitivitas antibiotik pada konsentrasi 100 µg/ml
Isolat Rifampisin Penisilin G Kanamisin Streptomisin
F5 - - - -
F13 - - - -
F15 - - - -
F16 - - - -
F17 - - - -
F18 - - - -
F19 - - - -
F25 - - - -
F26 - - - -
F27 - - - -
F31 - - - -
F34 - - - -
F39 - - - -
F41 - - - -
F43 - - - -
F45 - - - -
V. harveyi MR 5339 Rf R - - - -
Keterangan: (-):sensitif; (+):resisten
32 3.4 Metode Kultur Bersama
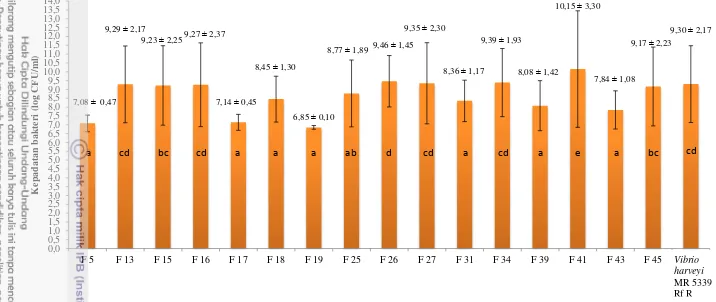
Uji in vitro kultur bersama dilakukan untuk mengetahui kemampuan bakteri kandidat dalam menghambat pertumbuhan bakteri patogen (V. harveyi MR 5339 Rf R). Uji ini dilakukan dengan membandingkan jumlah kepadatan bakteri V. harveyi MR 5339 Rf R yang tumbuh di tabung kontrol dengan tabung kultur bersama.
33 Gambar 2. Penghambatan V. harveyi MR 5339Rf R oleh kandidat probiotik pada uji in vitro metode kultur bersama
7,08 ± 0,47 9,29 ± 2,17
9,23 ± 2,259,27 ± 2,37
7,14 ± 0,45
8,45 ± 1,30
6,85 ± 0,10
8,77 ± 1,89 9,46 ± 1,45 9,35 ± 2,30
8,36 ± 1,17
9,39 ± 1,93
8,08 ± 1,42
10,15 ± 3,30
7,84 ± 1,08
9,17 ± 2,23
9,30 ± 2,17
0,0 0,5 1,0 1,5 2,0 2,5 3,0 3,5 4,0 4,5 5,0 5,5 6,0 6,5 7,0 7,5 8,0 8,5 9,0 9,5 10,0 10,5 11,0 11,5 12,0 12,5 13,0 13,5 14,0
F 5 F 13 F 15 F 16 F 17 F 18 F 19 F 25 F 26 F 27 F 31 F 34 F 39 F 41 F 43 F 45 Vibrio
harveyi K epa da ta n ba k ter i (lo g CF U/m l) Vibrio harveyi MR 5339 Rf R
ab a bc
cd bc cd a a a d cd a e cd
34 Tidak semua jenis bakteri mampu memproduksi inhibitory metabolites (senyawa penghambat) pada media agar karena terdapat kemungkinan produksi inhibitory metabolites akan lebih banyak jika kultur dilakukan pada media broth. Adanya perbedaan komposisi medium juga mempengaruhi produksi senyawa penghambat (Verschuere et al. 2000). Hal tersebut membuktikan bahwa penghambatan pertumbuhan populasi tidak selalu dapat dilihat melalui metode zona bening (Kirby Bauer Method).
Lactid Acid Bacteria (LAB) yang diisolasi dari saluran pencernaan mampu memproduksi komponen bakteriosin yang mampu menghambat pertumbuhan mikroorganisme lainnya dan secara tidak langsung akan mempengaruhi kompetisi dalam memperoleh energi di lingkungannya. LAB yang ditemukan dalam saluran pencernaan organisme akuatik seperti ikan mampu meningkatkan status kesehatan organisme tersebut (Verschuere et al. 2000). Sasanti (2005) melaporkan bahwa dari 110 isolat hasil isolasi dari terumbu karang, hanya 10 isolat saja yang potensial sebagai kandidat probiotik. Lima diantaranya terbukti mampu menghambat pertumbuhan Vibrio harveyi MR 5339 Rf R walaupun tidak menunjukkan adanya zona hambat pada uji in vitro.
Untuk mengetahui tingkat kepadatan bakteri yang dikultur sebelum masuk ketahapan kultur bersama, dilakukan pengukuran kepadatan bakteri dengan menggunakan spektrofotometer. Kepadatan bakteri diketahui dengan mengukur nilai optical density (OD) dari populasi sel yang dikultur selama 24 jam. Pengukuran OD dilakukan pada panjang gelombang 600 nm (lampiran 1).
3.5 Uji Patogenisitas Bakteri Kandidat Probiotik
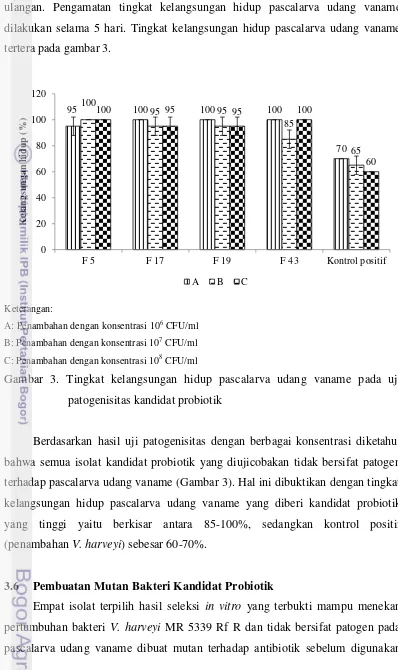
35 ulangan. Pengamatan tingkat kelangsungan hidup pascalarva udang vaname dilakukan selama 5 hari. Tingkat kelangsungan hidup pascalarva udang vaname tertera pada gambar 3.
Keterangan:
A: Penambahan dengan konsentrasi 106 CFU/ml
B: Penambahan dengan konsentrasi 107 CFU/ml
C: Penambahan dengan konsentrasi 108 CFU/ml
Gambar 3. Tingkat kelangsungan hidup pascalarva udang vaname pada uji patogenisitas kandidat probiotik
Berdasarkan hasil uji patogenisitas dengan berbagai konsentrasi diketahui bahwa semua isolat kandidat probiotik yang diujicobakan tidak bersifat patogen terhadap pascalarva udang vaname (Gambar 3). Hal ini dibuktikan dengan tingkat kelangsungan hidup pascalarva udang vaname yang diberi kandidat probiotik yang tinggi yaitu berkisar antara 85-100%, sedangkan kontrol positif (penambahan V. harveyi) sebesar 60-70%.
3.6 Pembuatan Mutan Bakteri Kandidat Probiotik
Empat isolat terpilih hasil seleksi in vitro yang terbukti mampu menekan pertumbuhan bakteri V. harveyi MR 5339 Rf R dan tidak bersifat patogen pada pascalarva udang vaname dibuat mutan terhadap antibiotik sebelum digunakan
95 100 100 100
70 100
95 95
85
65
100 95 95 100
60
0 20 40 60 80 100 120
F 5 F 17 F 19 F 43 Kontrol positif
K
ela
ng
sun
g
a
n hid
up
(
%)
36 pada uji in vivo. Empat isolat yaitu F5, F17, F19, dan F43 dibuat mutan secara spontan dengan cara menyebar biakan cair isolat pada media SWC agar yang mengandung streptomisin 50 µg/ml. Isolat yang tumbuh (resisten) pada media SWC yang mengandung streptomisin diambil sebagai inokulan untuk uji in vivo.
Bentuk dan ciri fisik isolat mutan tidak berbeda dengan isolat awalnya. Isolat F5 memiliki ciri berbentuk bulat, tepian rata, elevasi cembung, dan koloni berwarna putih susu (Gambar 4a). Hasil identifikasi genus merujuk pada metode identifikasi Cowan (1974) menunjukkan isolat F5 tergolong anggota genus Kurthia. Isolat F17 memiliki ciri koloni berbentuk bulat, tepian rata, elevasi cembung, berwarna putih, dan teridentifikasi sebagai anggota genus Micrococcus (Gambar 4b). Isolat F19 memiliki ciri koloni berbentuk bulat, berwarna putih susu, tepian rata, berelevasi cembung, dan teridentifikasi sebagai anggota genus Chromobacterium, Beneckea, Vibrio, Plesiomonas, dan Aeromonas. Isolat F43 memiliki ciri koloni berwarna putih, berbentuk bulat kecil, tepian rata, berelevasi cembung dan teridentifikasi sebagai anggota genus Kurthia.
a. b.
c. d.
37 3.7 Uji in vivo Kandidat Probiotik
Empat isolat hasil uji in vitro yaitu isolat F5, F17, F19, dan F43 diuji kemampuannya dalam menghambat pertumbuhan V. harveyi MR 5339 Rf R pada pascalarva (PL 26) udang vaname. Pengamatan yang dilakukan pada uji in vivo meliputi survival rate (tingkat kelangsungan hidup), laju pertumbuhan harian, total populasi V. harveyi MR 5339 Rf R, dan total populasi probiotik.
Berdasarkan hasil pengamatan diketahui bahwa tingkat kelangsungan hidup pascalarva udang vaname yang diberi keempat kandidat probiotik memiliki perbedaan yang signifikan (P<0,05) jika dibandingkan dengan kontrol positif (67,5%). Namun jika dibandingkan dengan kontrol negatif tidak menunjukkan perbedaan yang signifikan. Tingkat kelangsungan hidup pascalarva udang vaname yang diberi kandidat probiotik berkisar antara 78,75-92,50% sedangkan kontrol positif sebesar 67,5% (Gambar 5).
Keterangan:
A: F5 + V. harveyi MR 5339 Rf R C: F19 + V. harveyi MR 5339 Rf R E: V. harveyi MR 5339 Rf R
B: F17 + V. harveyi MR 5339 Rf R D: F43 + V. harveyi MR 5339 Rf R F: Kontrol negatif
Gambar 5. Tingkat kelangsungan hidup pascalarva udang vaname yang diberi bakteri kandidat probiotik pada uji in vivo
Pemberian bakteri probiotik mampu meningkatkan kelangsungan hidup larva udang vaname secara signifikan dapat disebabkan oleh adanya kompetisi dalam mendapatkan sumber energi. Menurut Smith and Davey (1993) dalam Verschuere et al. (2000) melaporkan bahwa Pseudomonas fluorescent F19 mampu menghambat pertumbuhan Aeromonas salmonicida pada media
92,50 ± 6,45
86,25 ± 4,79
78,75 ± 8,54 82,50 ± 6,45
67,50 ± 6,45
83,75 ± 8,54
0 20 40 60 80 100 120
A B C D E F
Sur v iv a l r a te ( %) Perlakuan
38 pemeliharaan Atlantic salmon. Mekanisme penghambatannya diketahui melalui kompetisi dalam memperoleh zat besi. Adanya produksi senyawa penghambat juga turut memberikan pengaruh dalam proses penghambatan pertumbuhan patogen oleh bakteri probiotik. Senyawa penghambat yang dimaksud dapat berupa antibiotik, senyawa yang bersifat bakteriostatik maupun bakteriosida, lysozyme, ataupun protease. Senyawa-senyawa tersebut diketahui mampu menghambat pertumbuhan mikroorganisme lain. Adanya kompetisi dalam memperoleh energi dan produksi senyawa penghambat inilah yang diduga mampu menekan pertumbuhan bakteri patogen.
Rengpipat et al. (2000) juga melaporkan bahwa penambahan Bacillus S11 mampu meningkatkan kelangsungan hidup pada larva udang windu (Penaeus monodon). Hasil penelitian Vaseeharan and Ramasamy (2003) menunjukkan bahwa tingkat kematian udang windu yang tidak diberi probotik Bacillus subtilis BT23 mencapai 50% pada hari ke-9 dan mencapai 100% pada hari ke-17.
Probiotik umumnya diaplikasikan pada media atau kolam pemeliharaan sebagai bentuk tindakan preventif terhadap serangan penyakit serta aktivator regenerasi nutrien (Yasuda dan Taga 1980 dalam Verschuere et al. 2000). Beberapa kelompok bakteri diketahui sebagai agen pengontrol biologis dalam akuakultur diantaranya kelompok Lactid Acid Bacteria (genus Lactobacillus dan Carnobacterium), anggota genus Vibrio (Vibrio alginolyticus), anggota genus Bacillus, serta anggota genus Pseudomonas. Menurut Verschuere et al. (2000),
mekanisme kerja probiotik diantaranya adalah adanya kemampuan memproduksi senyawa inhibitor, kompetisi dalam memperoleh energi, kompetisi tempat pelekatan, meningkatkan respon imun, dan memperbaiki kualitas air.
39 meningkatkan pertumbuhan dan tingkat kelangsungan hidup juvenile udang putih terhadap serangan Vibrio harveyi dan WSSV (White Spot Syndrome Virus).
Laju pertumbuhan harian bobot pascalarva udang vaname yang diberi keempat isolat (F5, F17, F19, dan F45) probiotik tidak menunjukkan perbedaan yang signifikan jika dibandingkan dengan kontrol negatif dan kontrol positif. Laju pertumbuhan pascalarva udang yang diberi kandidat probiotik berkisar antara 11,70-14,33 %, sedangkan laju pertumbuhan pada kontrol negatif dan positif berturut-turut adalah 11,78% dan 12,45% (Gambar 6).
Keterangan:
A: F5 + V. harveyi MR 5339 Rf R C: F19 + V. harveyi MR 5339 Rf R E: V. harveyi MR 5339 Rf R
B: F17 + V. harveyi MR 5339 Rf R D: F43 + V. harveyi MR 5339 Rf R F: Kontrol negatif
Gambar 6. Laju pertumbuhan harian bobot pascalarva udang vaname yang diberi bakteri kandidat probiotik pada uji in vivo
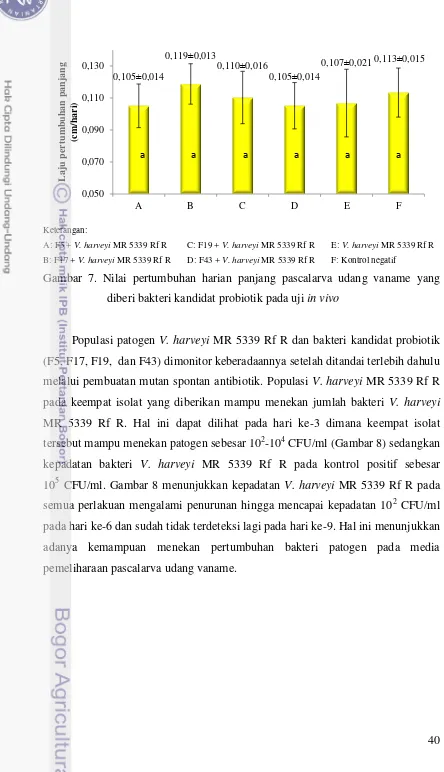
Nilai pertumbuhan harian panjang pascalarva yang diberi kandidat probiotik juga tidak menunjukkan perbedaan yang nyata terhadap kontrol negatif dan kontrol positif (P>0,05). Nilai pertumbuhan harian panjang pascalarva yang diberi kandidat probiotik berkisar antara 0,105-0,119 cm/hari sedangkan nilai pertumbuhan harian panjang kontrol positif dan kontrol negatif berturut-turut adalah 0,107 dan 0,113 cm/hari (Gambar 7).
12,39 ± 3,45
14,33 ± 0,31
11,70 ± 1,56
13,21 ± 2,23 12,45 ± 3,98
11,78 ± 1,52
0 2 4 6 8 10 12 14 16 18
A B C D E F
P
er
tum
bu
ha
n
bo
bo
t
(%)
Perlakuan
40
Keterangan:
A: F5 + V. harveyi MR 5339 Rf R C: F19 + V. harveyi MR 5339 Rf R E: V. harveyi MR 5339 Rf R
B: F17 + V. harveyi MR 5339 Rf R D: F43 + V. harveyi MR 5339 Rf R F: Kontrol negatif
Gambar 7. Nilai pertumbuhan harian panjang pascalarva udang vaname yang diberi bakteri kandidat probiotik pada uji in vivo
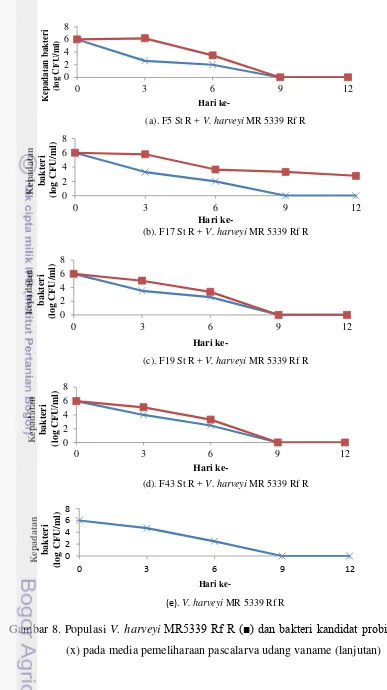
Populasi patogen V. harveyi MR 5339 Rf R dan bakteri kandidat probiotik (F5, F17, F19, dan F43) dimonitor keberadaannya setelah ditandai terlebih dahulu melalui pembuatan mutan spontan antibiotik. Populasi V. harveyi MR 5339 Rf R pada keempat isolat yang diberikan mampu menekan jumlah bakteri V. harveyi MR 5339 Rf R. Hal ini dapat dilihat pada hari ke-3 dimana keempat isolat tersebut mampu menekan patogen sebesar 102-104 CFU/ml (Gambar 8) sedangkan
kepadatan bakteri V. harveyi MR 5339 Rf R pada kontrol positif sebesar 105 CFU/ml. Gambar 8 menunjukkan kepadatan V. harveyi MR 5339 Rf R pada
semua perlakuan mengalami penurunan hingga mencapai kepadatan 102 CFU/ml pada hari ke-6 dan sudah tidak terdeteksi lagi pada hari ke-9. Hal ini menunjukkan adanya kemampuan menekan pertumbuhan bakteri patogen pada media pemeliharaan pascalarva udang vaname.
0,105±0,014
0,119±0,013
0,110±0,016
0,105±0,014
0,107±0,021 0,113±0,015
0,050 0,070 0,090 0,110 0,130
A B C D E F
L
a
ju per
tum
bu
ha
n
pa
nja
ng
(cm
/ha
ri)
41 (a). F5 St R + V. harveyi MR 5339 Rf R
(b). F17 St R + V. harveyi MR 5339 Rf R
(c). F19 St R + V. harveyi MR 5339 Rf R
(d). F43 St R + V. harveyi MR 5339 Rf R
(e). V. harveyi MR 5339 Rf R
Gambar 8. Populasi V. harveyi MR5339 Rf R (■) dan bakteri kandidat probiotik (x) pada media pemeliharaan pascalarva udang vaname (lanjutan) 0
2 4 6 8
0 3 6 9 12
K ep a d a tan b a k teri (l o g CF U/ m l) Hari ke- 0 2 4 6 8
0 3 6 9 12
K epa da ta n ba k ter i ( lo g CF U/m l) Hari ke- 0 2 4 6 8
0 3 6 9 12
k epa da ta n ba k ter i (lo g CF U/m l) Hari ke- 0 2 4 6 8
0 3 6 9 12
K epa da ta n ba k ter i ( lo g CFU/m l) Hari ke- 0 2 4 6 8
0 3 6 9 12
42 Adanya peningkatan kelangsungan hidup pada pascalarva udang disebabkan oleh adanya penekanan terhadap pertumbuhan V. harveyi MR 5339 Rf R. Penekanan pertumbuhan populasi patogen ini ditunjukkan dengan berkurangnya jumlah bakteri patogen pada media pemeliharaan yang diberi kandidat probiotik jika dibandingkan dengan kontrol positif (Gambar 9). Populasi V. harveyi MR 5339 Rf R pada kontrol positif mencapai 105 CFU/ml pada hari ke-3, ini lebih tinggi daripada populasi V. harveyi MR 5339 Rf R pada perlakuan penambahan probiotik yang mencapai kepadatan 102-104 CFU/ml. Penekanan populasi patogen terbaik terdapat pada perlakuan penambahan isolat F5 yakni mampu mengeliminasi patogen sebesar 103 CFU/ml pada hari ke-3. Kemampuan penekanan V. harveyi MR 5339 Rf R oleh isolat F5 diikuti pula oleh isolat F19, F17, dan F43. Kemampuan menghambat ini diduga menyebabkan tingginya tingkat kelangsungan hidup pascalarva udang vaname pada perlakuan penambahan isolat F5 (Gambar 9).
Keterangan:
A: F5 + V. harveyi MR 5339 Rf R C: F19 + V. harveyi MR 5339 Rf R E: V. harveyi MR 5339 Rf R
B: F17 + V. harveyi MR 5339 Rf R D: F43 + V. harveyi MR 5339 Rf R F: Kontrol negatif
Gambar 9. Tren populasi Vibrio harveyi MR 5339 Rf R pada berbagai perlakuan
Penambahan bakteri probiotik diawal pemeliharaan saja menyebabkan populasi kandidat probiotik menurun hingga hari ke-12 pengamatan (Gambar 10). Isolat F5, F19, dan F43 mengalami penurunan populasi hingga tidak terdeteksi sama sekali pada hari ke-9. Sedangkan isolat F17 mampu bertahan pada media pemeliharaan hingga hari ke-12 pengamatan. Isolat F5 dan F19 memiliki populasi
0 1 2 3 4 5 6 7
0 3 6 9 12
43 yang lebih tinggi dari pada isolat F17 dan F43 pada hari ke-3 pengamatan. Lebih tingginya kepadatan isolat F5 pada hari ke-3 berkolerasi dengan kemampuannya menghambat pertumbuhan Vibrio harveyi MR 5339 Rf R (Gambar 9). Hubungan ini juga terdapat pada perlakuan penambahan isolat F17 dan F43.
Keterangan:
A: F5 + V. harveyi MR 5339 Rf R C: F19 + V. harveyi MR 5339 Rf R E: V. harveyi MR 5339 Rf R
B: F17 + V. harveyi MR 5339 Rf R D: F43 + V. harveyi MR 5339 Rf R F: Kontrol negatif
Gambar 10. Tren populasi kandidat probiotik St R pada berbagai perlakuan
Vibriosis pada udang merupakan penyakit bakterial yang disebabkan oleh V. harveyi. Penyakit ini bersifat oportunis dimana akan menyerang pada saat sistem imun inang lemah (Defoirdt et al. 2008). Serangan vibriosis akan menyebabkan larva menjadi abnormal diantaranya saluran pencernaan menjadi kosong, hepatopankreas menyusut dan munculnya bintik-bintik melanisasi pada permukaan tubuh, serta kolonisasi bakteri pada permukaan kutikula larva (Brock and Main 1994). Patogen ini juga dilaporkan sebagai agen penyebab penyakit pada kuda laut (Hipocampus sp.), skin ucleration (bisul) pada teripang (Holothuria scabra) di Madagaskar, dan luminous disease pada larva phyllosoma (Jasus verreauxi). Gejala pada teripang yang terserang patogen ini berupa bintik putih dan kematian akan terjadi selama tiga hari (Austin and Zhang 2006).
Tingkat patogenisitas Vibrio harveyi berhubungan dengan mekanisme quorum sensing. Quorum sensing merupakan mekanisme komunikasi antar sel melalui produksi molekul-molekul tertentu (Defoirdt et al. 2007) atau mekanisme regulasi gen dimana bakteri mengkoordinasikan ekspresi gen tertentu dalam
0 1 2 3 4 5 6 7
0 3 6 9 12
45 IV. KESIMPULAN DAN SARAN
4.1. Kesimpulan
Keempat isolat (F5, F17, F19, dan F43) hasil isolasi dari usus udang vaname efektif dalam menghambat pertumbuhan Vibrio harveyi MR 5339 Rf R. Hal ini dibuktikan dengan nilai kelangsungan hidup post larva udang vaname yang diberi kandidat probiotik menunjukkan hasil yang berbeda nyata (P<0,05) antara yang diberi kandidat probiotik (78,75-92,50%) dengan kontrol positif sebesar 67,5%.
4.2. Saran
EFEKTIVITAS PROBIOTIK ASAL USUS UDANG DALAM
MENGHAMBAT PERTUMBUHAN Vibrio harveyi PADA LARVA UDANG VANAME Litopenaeus vannamei
DWI FEBRIANTI
DEPARTEMEN BUDIDAYA PERAIRAN FAKULTAS PERIKANAN DAN ILMU KELAUTAN
14 ABSTRAK
DWI FEBRIANTI. Efektivitas probiotik asal usus udang dalam menghambat pertumbuhan Vibrio harveyi pada larva udang vaname Litopenaeus vannamei. Dibimbing oleh MUNTI YUHANA dan WIDANARNI.
Vibriosis merupakan salah satu jenis penyakit infeksius yang sering menyerang udang vaname yang salah satunya disebabkan oleh Vibrio harveyi. Salah satu alternatif pengendalian penyakit bakterial yang dapat dilakukan adalah dengan menggunakan bakteri probiotik. Penelitian ini bertujuan mempelajari efektifitas empat strain strain probiotik dalam menghambat pertumbuhan Vibrio harveyi pada larva udang vaname Litopenaeus vannamei. Rancangan penelitian yang digunakan adalah Rancangan Acak Lengkap (RAL) dengan enam perlakuan yaitu penambahan isolat F5, F17, F19, F43, kontrol positif (Vibrio harveyi MR 5339 Rf R), dan kontrol negatif (tanpa penambahan bakteri probiotik maupun Vibrio harveyi MR 5339 Rf R) dan empat ulangan. Berdasarkan hasil pengamatan diketahui bahwa tingkat kelangsungan hidup pascalarva udang vaname yang diberi keempat kandidat probiotik memiliki perbedaan yang signifikan (P<0,05) jika dibandingkan dengan kontrol positif (67,5±6,45 %). Tingkat kelangsungan hidup pascalarva udang vaname yang diberi kandidat probiotik berkisar antara 78,75±8,54 - 92,50±6,45 %. Laju pertumbuhan harian panjang dan bobot tidak memiliki perbedaan yang signifikan pada semua perlakuan.
15 ABSTRACT
DWI FEBRIANTI. Effectiveness of intestinal shrimp probiotics in inhibiting growth of Vibrio harveyi in shrimp larvae Litopenaeus vannamei. Supervised by MUNTI YUHANA and WIDANARNI.
Vibriosis is one of infectious disease that often attacks the shrimp vaname culture. It caused by Vibrio harveyi. One of alternative in controlling the luminous disease is the application of probiotic bacteria which is considered as enviromental friendly treatment. This research was carried out to evaluate the effectiveness of four strains probiotic in inhibiting growth of Vibrio harveyi in shrimp larvae Litopenaeus vannamei. The study design used was the complete random design with six treatments: the addition of isolates F5, F17, F19, F43, positive control (the addition of V. harveyi MR 5339 Rf R), and the negative control (without addition of probiotics and V. harveyi MR 5339 Rf R). Based on the observations, the survival rate of shrimp post larvae treatment with four candidate probiotics have a significant difference survival rate (P <0,05) when compared with positive control (67,5±6,45 %). The survival rate of post larvae
treatment with candidate probiotics were ranging from 78,75±8,54 - 92,50±6,45 %. Daily growth rate of length and weight have no
significant differences in all treatments.
9 EFEKTIVITAS PROBIOTIK ASAL USUS UDANG DALAM
MENGHAMBAT PERTUMBUHAN Vibrio harveyi PADA LARVA UDANG VANAME Litopenaeus vannamei
DWI FEBRIANTI
SKRIPSI
sebagai salah satu syarat untuk memperoleh gelar Sarjana Perikanan pada Program Studi Teknologi dan Manajemen Perikanan Budidaya
Departemen Budidaya Perairan, Fakultas Perikanan dan Ilmu Kelautan,
Institut Pertanian Bogor
DEPARTEMEN BUDIDAYA PERAIRAN FAKULTAS PERIKANAN DAN ILMU KELAUTAN
10 PERNYATAAN MENGENAI SKRIPSI
DAN SUMBER INFORMASI
Dengan ini menyatakan bahwa skripsi yang berjudul:
EFEKTIVITAS PROBIOTIK ASAL USUS UDANG DALAM
MENGHAMBAT PERTUMBUHAN Vibrio harveyi PADA LARVA UDANG VANAME Litopenaeus vannamei
adalah benar merupakan hasil karya yang belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Semua sumber data dan informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir skripsi ini.
Bogor, November 2011
11 PENGESAHAN
Judul Skripsi : Efektivitas probiotik asal usus udang dalam menghambat pertumbuhan Vibrio harveyi pada larva udang
vaname Litopenaeus vannamei Nama Mahasiswa : Dwi Febrianti
Nomor Pokok : C14070067
Disetujui
Pembimbing I Pembimbing II
Dr. Munti Yuhana, S.Pi., M.Si. Dr. Ir. Widanarni, M.Si. NIP. 19691220 199403 2 002 NIP. 19670927 199403 2 001
Diketahui,
Ketua Departemen Budidaya Perairan
Dr. Ir. Odang Carman, M.Sc. NIP. 19591222 198601 1 001
12 KATA PENGANTAR
Puji syukur penulis panjatkan kepada Allah SWT karena atas rahmat dan hidayah-Nya lah penulis dapat menyelesaikan karya ilmiah ini dengan baik. Penelitian ini dilaksanakan pada bulan Maret hingga Agustus 2011 bertempat di Laboratorium Kesehatan Ikan, Departemen Budidaya Perairan, Fakultas Perikanan dan Ilmu Kelautan, Institut Pertanian Bogor. Penelitian ini berjudul
“Efektivitas Probiotik Asal Usus Udang dalam Menghambat Pertumbuhan Vibrio harveyi pada Larva Udang Vaname Litopenaeus vannamei”.
Penulis mengucapkan terimakasih kepada: Pemerintah Provinsi Jambi khususnya Dinas Pendidikan Provinsi Jambi yang telah memberikan Beasiswa kepada penulis hingga penulis bisa menyelesaikan studi di Institut Pertanian Bogor; Dr. Munti Yuhana, S.Pi., M.Si., dan Dr. Ir. Widanarni, M.Si. selaku dosen pembimbing yang telah banyak memberikan bimbingan, masukan, dan motivasi kepada penulis; Kedua orang tua dan seluruh keluarga besar atas doa dan dukungannya; Rekan-rekan Laboratorium Kesehatan Ikan, Pak Ranta, Kak Rahman, Kak Ulvi, Kak Fariq, Ghita, Dama, Ika, dan Ririn atas semangat dan bantuannya; serta Rekan-rekan BDP 44 atas semangat dan kebersamaannya.
Semoga tulisan ini dapat memberikan manfaat bagi penulis dan semua pembaca.
Bogor, November 2011
13 RIWAYAT HIDUP
Penulis dilahirkan di Kuala Tungkal pada tanggal 14 Februari 1990 dari pasangan Bapak Cholil Sadikin dan Ibu Herlina. Penulis merupakan anak kedua dari tiga bersaudara.
Penulis menyelesaikan pendidikan formal di SDN 3/V Kuala Tungkal (2001), SMPN 1 Kuala Tungkal (2004), dan SMAN 1 Kuala Tungkal (2007). Penulis masuk IPB melalu jalur Beasiswa Utusan Daerah (BUD) pada tahun 2007 dan mengambil program studi Teknologi dan Manajemen Perikanan Budidaya pada Fakultas Perikanan dan Ilmu Kelautan dengan kompetensi minor Kewirausahaan Agribisnis pada Fakultas Ekonomi dan Manajemen.
Selama mengikuti perkuliahan penulis aktif menjadi pengurus Himpunan Mahasiswa Akuakultur pada divisi Sosial Kemasyarakatan (2009/2010) serta divisi Publikasi dan Pengembangan Sumber Daya (2010/2011). Selain itu penulis juga aktif pada organisasi ekstra kampus sebagai anggota Himpunan Mahasiswa Jambi (HIMAJA). Penulis juga pernah menjadi asisten pada beberapa mata kuliah yaitu Ikhtiologi (2009/2010), Teknologi Penangangan dan Transportasi Hasil Perairan (2010/2011), Dasar-dasar Mikrobiologi Akuatik (2010/2011), Penyakit Organisme Akuatik (2010/2011), dan Mikrobiologi Akuakultur (2011/2012). Penulis merupakan salah satu anggota penerima dana DIKTI pada PKM bidang
pengabdian masyarakat (2010) yang berjudul “Pelatihan produksi ikan hias
dengan metode induced breeding di Kampung Setu, Desa Parigi Mekar, Kecamatan Ciseeng, Kabupaten Bogor”.
16 DAFTAR ISI
Halaman DAFTAR TABEL ... ii DAFTAR GAMBAR ... iii DAFTAR LAMPIRAN ... iv I. PENDAHULUAN ... 1
1.1 Latar Belakang ... 1 1.2 Tujuan... 2 II. BAHAN DAN METODE ... 3 2.1 Seleksi Bakteri Probiotik ... 3
2.1.1 Karakterisasi morfologi dan fisiologis kandidat
probiotik ... 3 2.1.2 Uji aktivitas amilolitik dan proteolitik ... 3 2.1.3 Uji sensitivitas antibiotik ... 3 2.1.4 Metode kultur bersama ... 4 2.1.5 Uji patogenisitas bakteri kandidat probiotik ... 4 2.1.6 Pembuatan mutan bakteri kandidat probiotik ... 5 2.2 Uji in vivo Kandidat Probiotik ... 5 2.3 Rancangan Penelitian dan Teknik Analisa ... 5 2.4 Parameter yang Diamati ... 5
2.4.1 Tingkat kelangsungan hidup pascalarva udang
vaname ... 5 2.4.2 Pertumbuhan harian panjang dan bobot
harian pascalarva udang vaname ... 6 2.4.3 Populasi bakteri ... 6 III. HASIL DAN PEMBAHASAN ... 8
3.1 Karakterisasi Morfologi dan Fisiologis Kandidat
17 DAFTAR TABEL
Halaman 1. Karakterisasi morfologi dan fisiologis kandidat probiotik ... 7 2. Diameter aktivitas amilolitik dan proteolitik kandidat
18 DAFTAR GAMBAR
Halaman 1. Hasil aktivitas amilolitik dan proteolitik ... 8 2. Penghambatan V. harveyi MR 5339Rf R oleh kandidat
probiotik pada uji in vitro metode kultur bersama ... 13
3. Tingkat kelangsungan hidup pascalarva udang vaname
pada uji patogenisitas kandidat probiotik ... 15 4. Penampilan mutasi spontan isolat F5, F17, F19 dan F43
pada media SWC agar ... 16 5. Tingkat kelangsungan hidup pascalarva udang vaname
yang diberi bakteri kandidat probiotik pada uji in vivo ... 17 6. Laju pertumbuhan harian bobot pascalarva udang vaname
yang diberi bakteri kandidat probiotik pada uji in vivo ... 19 7. Nilai pertumbuhan harian panjang pascalarva udang vaname
yang diberi bakteri kandidat probiotik pada uji in vivo ... 20 8. Populasi Vibrio harveyi MR5339 Rf R dan bakteri kandidat
probiotik pada media pemeliharaan pascalarva udang vaname 21 9. Tren populasi Vibrio harveyi MR5339 Rf R pada berbagai
19 DAFTAR LAMPIRAN
Halaman 1. Grafik hubungan nilai optical density (OD) dan kepadatan
bakteri ... 30 2. Nilai parameter kualitas air pascalarva udang vaname selama
pemeliharaan pada uji in vivo ... 32 3. Uji homogenisitas laju pertumbuhan harian (LPH) udang
vaname ... 33 4. Analisis statistik kultur bersama kandidat probiotik dan
V. harveyi MR 5339 Rf R ... 34 5. Nilai kelangsungan hidup pascalarva udang vaname pada
uji patogenisitas kandidat probiotik ... 35 6. Analisis statistik tingkat kelangsungan hidup pascalarva udang
vaname pada uji patogenisitas kandidat probiotik ... 37 7. Analisa statistik tingkat kelangsungan hidup pascalarva
udang vaname hasil uji in vivo ... 39 8. Analisa statistik laju pertumbuhan harian (LPH) panjang
dan berat pascalarva udang vaname hasil uji in vivo ... 40 9. Laju pertumbuhan harian (LPH) panjang pascalarva udang
vaname hasil uji in vivo ... 41 10. Laju pertumbuhan harian (LPH) bobot pascalarva udang
[image:39.595.47.497.53.827.2]20
I. PENDAHULUAN
1.1 Latar Belakang
Udang vaname (Litopenaeus vannamei) merupakan salah satu komoditas ekspor unggulan Kementrian Kelautan dan Perikanan yang diharapkan mengalami peningkatan produksi sebesar 209% dalam kurun waktu 2009-2014 (KKP 2010) dari 201% target total peningkatan produksi udang Indonesia. Peningkatan produksi udang vaname diharapkan mencapai 16% pertahunnya. Produksi udang vaname pada tahun 2014 menurut KKP (2010) diproyeksikan sebesar 511 ton. Udang vaname dipilih sebagai komoditas unggulan karena memiliki beberapa kelebihan diantaranya banyak diminati oleh pasar lokal maupun internasional, lebih tahan terhadap penyakit jika dibandingkan dengan jenis udang putih lainnya, pertumbuhan relatif lebih cepat, serta memiliki toleransi yang cukup besar terhadap perubahan kualitas lingkungan (Atjo 2009).
Adanya perubahan kualitas lingkungan dan serangan penyakit menyebabkan timbulnya kendala dalam sistem budidaya udang vaname. Penyakit vibriosis merupakan salah satu jenis penyakit bakterial yang menyerang seluruh siklus hidup udang vaname dan telah menyebabkan tingginya tingkat kematian pada industri budidaya udang dunia (Heidarieh et al. 2010). Vibriosis merupakan salah satu jenis penyakit infeksius yang sering menyerang organisme akuatik air laut seperti ikan dan udang. Penyakit ini umumnya disebabkan oleh anggota genus Vibrio (Munn 1977) seperti Vibrio harveyi, Vibrio alginolyticus, Vibrio parahaemolyticus, dan Vibrio penaeicida (Longyant et al. 2008). Vibrio harveyi merupakan patogen dari genus Vibrio yang berasosisasi pada penyakit udang berpendar (Austin and Zhang 2006).
21 penyuntikan intramuskular, sedangkan pada konsentrasi >106-107 cfu/ml dapat menyebabkan kematian pada udang vaname (Litopenaeus vaname) melalui infeksi dengan teknik perendaman selama 2 jam (Saulnier et al. 2000).
Antibiotik merupakan salah satu bahan yang sering digunakan untuk pencegahan dan penanggulangan penyakit bakterial pada kegiatan budidaya udang. Penggunaan antibiotik seperti oxytetracycline, oxolinic acid, chloramphenicol dan furazolidone umumnya digunakan untuk menanggulangi penyakit vibriosis (Tendencia and de la Pena 2001). Menurut Serrano (2005) dalam Reboucas et al. (2011), patogen penyebab vibriosis yang terpapar antibiotik baik di dalam maupun di luar lingkungan hatchery pemeliharaan udang dapat menyebabkan timbulnya sifat resistensi melalui perpindahan materi genetik.
Salah satu alternatif pengendalian penyakit bakterial yang dapat dilakukan adalah dengan menggunakan bakteri probiotik. Beberapa mekanisme kerja bakteri probiotik dalam menghambat keberadaan bakteri patogen diantaranya menghasilkan senyawa penghambat, kompetisi memperoleh sumber energi, kompetisi tempat perlekatan, meningkatkan respon imun, serta memperbaiki kualitas air (Verschuere et al. 2000). Bakteri yang berhasil digunakan sebagai probiotik diantaranya berasal dari genus Vibrio, Bacillus spp., dan Thallasobacter utilis. Bakteri probiotik dapat diisolasi dari berbagai sumber diantaranya air
pemeliharaan udang ataupun saluran pencernaan udang itu sendiri (Guillan et al. 2004) ataupun terumbu karang (Sasanti 2008).
1.2 Tujuan
22 II. BAHAN DAN METODE
2.1 Seleksi Bakteri Probiotik
2.1.1 Karakterisasi morfologi dan fisiologis kandidat probiotik
Sebanyak 16 jenis bakteri hasil isolasi Ardiani (2011) ditumbuhkan pada media agar Sea Water Complete (SWC) lalu diinkubasi selama 24 jam pada suhu 28oC. Koloni yang tumbuh dilakukan serangkaian uji untuk mengetahui karakter morfologi dan fisiologis dari masing-masing isolat. Uji yang dilakukan meliputi (1) pewarnaan Gram dengan menggunakan larutan kristal violet, lugol, alkohol absolut, dan safranin (2) uji oksidatif-fermentatif dengan menggunakan media uji oksidatif-fermentatif, glukosa, dan parafin; (3) uji motilitas dengan menggunakan media Sulfide Indol Motility (SIM); (4) uji katalase dengan menggunakan
Hidrogen Peroksida (H2O2) 3%; dan (5) uji oksidase dengan menggunakan p-aminodhymethylalanin oxalat 1%.
2.1.2 Uji Aktivitas amilolitik dan proteolitik
Uji ini dilakukan untuk mengetahui aktivitas amilase dan protease dari masing-masing isolat melalui kemampuannya menghidrolisis karbohidrat dan protein. Bakteri kandidat probiotik ditumbuhkan pada media SWC yang telah ditambahkan tepung kanji sebanyak 2% untuk uji amilolitik. Kemampuan menghidrolisa karbohidrat ditandai dengan terbentuknya warna kuning terang di sekitar isolat sedangkan isolat yang tidak mampu menghidrolisa karbohidrat, area di sekitar isolat tetap berwarna gelap setelah penambahan reagen KI 1%. Sedangkan pada uji aktivitas proteolitik, bakteri kandidat probiotik ditumbuhkan pada media SWC yang telah ditambahkan susu skim sebanyak 2%. Kemampuan menghidrolisa protein ditandai dengan terbentuknya zona bening di sekitar isolat.
2.1.3 Uji sensitivitas antibiotik
23 pemeliharaan pascalarva. Jenis antibiotik yang digunakan adalah rifamfisin, streptomisin, kanamisin, dan penisilin G. Konsentrasi yang digunakan adalah 50 µg/ml dan 100 µg/ml. Kandidat probiotik ditumbuhkan pada media SWC agar yang telah mengandung antibiotik dengan konsentrasi 50 µg/ml dan 100 µg/ml menggunakan metode streak plate lalu diinkubasi selama 24 jam pada suhu 28oC. Bakteri yang sensitif terhadap antibiotik ditandai dengan tidak adanya pertumbuhan koloni pada media SWC agar yang telah mengandung antibiotik, sedangkan bakteri yang resisten akan tetap tumbuh pada media tersebut.
2.1.4 Metode kultur bersama
Kemampuan kandidat bakteri probiotik melawan bakteri V. harveyi MR 5339 Rf R diuji juga dengan menggunakan metode kultur bersama. Isolat bakteri kandidat probiotik dan bakteri V. harveyi MR 5339 Rf R yang telah berumur 24 jam diencerkan hingga mencapai kepadatan yang sama yakni mencapai 106 CFU/ml. Sebanyak 100 µl bakteri kandidat probiotik dan bakteri V. harveyi MR 5339 Rf R dimasukkan kedalam media SWC broth, lalu diinkubasi selama 24 jam pada shaker berkecepatan 140 rpm dengan suhu 29oC. Sebagai kontrol digunakan bakteri V. harveyi MR 5339 Rf R yang ditumbuhkan sendiri (tanpa bakteri kandidat probiotik) dengan kepadatan yang sama. Kemampuan bakteri kandidat probiotik menghambat pertumbuhan bakteri V. harveyi MR 5339 Rf R ditandai dengan kepadatan bakteri V. harveyi MR 5339 Rf R pada perlakuan kontrol yang lebih banyak dibandingkan dengan bakteri V. harveyi MR 5339 Rf R pada hasil kultur bersama.
2.1.5 Uji patogenitas bakteri kandidat probiotik
24 V. harveyi MR 5339 Rf R. Pemberian pakan dilakukan sebanyak 5 kali sehari dengan menggunakan pelet berjenis crumble. Pengamatan tingkat kelangsungan hidup dilakukan setiap hari selama 5 hari (Vijayan et al. 2006).
2.1.6 Pembuatan mutan bakteri kandidat probiotik
Isolat-isolat yang menghasilkan kelangsungan hidup terbaik pada uji patogenisitas, diberi penanda resisten streptomisin (St R) terlebih dahulu sebelum digunakan pada uji in vivo. Pemberian penanda streptomisin dilakukan dengan menumbuhkan isolat secara spontan pada media SWC agar yang mengandung streptomisin sebanyak 50 µg/ml media. Sedangkan patogen yang digunakan adalah V. harveyi MR 5339 Rf R yang ditumbuhkan secara spontan pada media SWC yang mengandung 50 µg/ml rifamfisin. Pemberian penanda ini dilakukan untuk memonitor keberadaan bakteri pada media pemeliharaan.
2.2 Uji in vivo Kandidat Probiotik
Isolat bakteri kandidat probiotik diinokulasikan pada media pemeliharaan hingga mencapai konsentrasi akhir 106 CFU/ml sehari setelah larva udang dimasukkan. Patogen V. harveyi MR 5339 Rf R diinokulasikan hingga mencapai kepadatan akhir 106 CFU/ml enam jam setelah pemberian kandidat probiotik. Percobaan dilakukan sebanyak empat kali ulangan termasuk kontrol positif (dengan inokulasi bakteri V. harveyi MR 5339 Rf R saja) dan kontrol negatif (tanpa penambahan bakteri kandidat probiotik maupun V. harveyi MR 5339 Rf R). Pengamatan dilakukan selama 14 hari meliputi tingkat kelangsungan hidup (SR), total populasi bakteri V. harveyi MR 5339 Rf R, total populasi bakteri kandidat probiotik, serta laju pertumbuhan panjang dan bobot harian.
2.3 Rancangan Penelitian dan Teknik Analisa
25 2.4 Parameter yang Diamati
2.4.1 Tingkat kelangsungan hidup pascalarva udang vaname
Tingkat kelangsungan hidup pascalarva udang vaname dihitung dengan menggunakan rumus sebagai berikut:
SR =
x 100% (Effendi 2004)
Keterangan:
SR = tingkat kelangsungan hidup (%)
Nt = jumlah pascalarva udang hidup pada akhir pengamatan No = jumlah pascalarva udang pada awal pengamatan
2.4.2 Pertumbuhan panjang dan bobot harian pascalarva udang vaname Pertumbuhan panjang dan bobot total diamati pada awal dan akhir penelitian. Pertumbuhan bobot pascalarva udang vaname dihitung berdasarkan pertambahan bobot berdasarkan rumus berikut :
SGR = { √
} (Effendie 1997)
Keterangan:
SGR = laju pertumbuhan harian bobot larva udang (%) t = lama waktu pemeliharaan larva udang (hari) Wt = bobot rata-rata akhir pascalarva udang (mg) Wo = bobot rata-rata awal pascalarva udang (mg)
Nilai pertumbuhan panjang harian (Daily increment in length) pascalarva udang vaname dihitung dengan menggunakan rumus sebagai berikut:
DIL = (Mai et al. 2000) DIL = pertumbuhan panjang harian
Lt = panjang rata-rata akhir pascalarva udang (mm) Lo = panjang rata-rata awal pascalarva udang (mm)
2.4.3 Populasi bakteri
27 III. HASIL DAN PEMBAHASAN
3.1 Karakterisasi Morfologi dan Fisiologis Kandidat Probiotik
[image:47.595.112.524.207.545.2]Hasil karakterisasi morfologi dan fisiologis yang dilakukan terhadap 16 jenis bakteri hasil isolasi Ardiani (2011) tertera pada tabel 1.
Tabel 1. Karakterisasi morfologi dan fisiologis kandidat probiotik
Kode
Isolat Morfologi
Uji
Genus* Pewarnaan
Gram O/F Katalase Oksidase Motility
F5 Basil Gram + - Positif Positif Motil Kurthia
F13 Basil Gram + - Positif Positif Motil Kurthia
F15 Basil Gram + - Positif Negatif Non
motil Corynebacterium F16 Basil Gram + - Positif Positif Motil Kurthia
F17 Coccus Gram + - Positif Negatif Non
motil Micrococcus F18 Coccus Gram - - Positif Positif Motil Micrococcus
F19 Basil Gram - Fermentatif Positif Positif Motil
Chromobacterium, Beneckea, Vibrio,
Plesiomonas, Aeromonas
F25 Basil Gram + - Positif Positif Motil Kurthia
F26 Basil Gram + - Positif Positif Motil Kurthia
F27 Basil Gram + - Positif Positif Motil Kurthia
F31 Basil Gram + - Positif Positif Motil Kurthia
F34 Basil Gram + - Positif Positif Motil Kurthia
F39 Basil Gram + - Positif Positif Non
motil Corynebacterium
F41 Basil Gram - - Positif Negatif Non
motil Micrococcus F43 Basil Gram - - Positif Positif Motil Kurthia
F45 Coccus Gram + Fermentatif Positif Positif Non
motil Staphylococcus *Mengacu pada metode identifikasi Cowan (1974)
** Hasil (-) menunjukkan tidak ada reaksi yang terjadi
28 F41, dan F45 bersifat tidak motil sedangkan isolat lainnya bersifat motil (mampu bergerak). Berdasarkan identifikasi tingkat genus yang mengacu pada metode Cowan (1974), isolat berhasil diidentifikasi sebagai anggota dari genus Kurthia, Corynebacterium, Micrococcus, Chromobacterium, Beneckea, Vibrio, Plesiomonas, Aeromonas, dan Staphylococcus.
3.2 Uji Aktivitas Amilolitik dan Proteolitik
Aktivitas amilase ditandai dengan terbentuknya zona berwarna kuning terang disekitar isolat sedangkan isolat yang tidak mampu menghidrolisa karbohidrat, area disekitar isolat tetap berwarna gelap setelah penambahan reagen KI. Sedangkan aktivitas protease ditandai dengan terbentuknya zona bening disekitar isolat. Hasil uji amilolitik dan proteolitik disajikan pada gambar 1 di bawah ini.
[image:48.595.85.510.57.730.2]
a. b.
Gambar 1. Hasil aktivitas (a) amilolitik dan (b) proteolitik
Hasil pengukuran diameter aktivitas amilolitik dan proteolitik dapat dilihat pada tabel 2.
Tabel 2. Diameter aktivitas amilolitik dan proteolitik kandidat probiotik
Kode Isolat Diameter Aktivitas Amilolitik (mm)
Diameter Aktivitas Proteolitik
(mm)
F5 12 -
F13 15 -
F15 3 -
F16 11 -
F17 - -
F18 - -
[image:48.595.113.430.611.756.2]29 Tabel 2. Diameter aktivitas amilolitik dan proteolitik kandidat probiotik (lanjutan)
Kode Isolat Diameter Aktivitas Amilolitik (mm)
Diameter Aktivitas Proteolitik
(mm)
F25 6 -
F26 11 -
F27 10 -
F31 12 -
F34 - 15
F39 5 19
F41 - -
F43 - -
F45 - -
Berdasarkan hasil uji amilolitik isolat diketahui bahwa isolat yang memiliki aktivitas amilolitik terbesar berturut-turut terdapat pada isolat F13 dengan diameter zona amilolitik sebesar 15 mm, serta F5 dan F31 dengan diameter zona amilolitik sebesar 12 mm. Aktivitas proteolitik hanya terdapat pada isolat F34 dan F39. Aktivitas proteolitik terbesar terdapat pada isolat F39 dengan diameter zona sebesar 19 mm diikuti dengan isolat F34 dengan diameter zona sebesar 15 mm. Adanya kemampuan menghidrolisis karbohidrat dan protein menunjukkan isolat-isolat tersebut mampu memanfaatkan sumber energi berupa pati dan protein yang ditambahkan pada media menjadi sumber karbon.
Menurut Price dan Stevens (1996) dalam Yandri et al. (2008), protease merupakan enzim yang berfungsi memecah ikatan peptida untuk menghasilkan asam amino dan peptida sederhana lainnya. Enzim ini dapat diisolasi dari berbagai sumber seperti tanaman, hewan, dan mikroba (fungi dan bakteri). Protease atau disebut juga endopeptidase merupakan salah satu enzim pencernaan yang penting, termasuk di dalamnya tripsin dan kemotripsin yang bertanggung jawab terhadap hampir 60% dari proses pencernaan udang (Lemos et al. 2000).
3.3 Uji Sensitivitas Antibiotik
30 antibiotik. Konsentrasi antibiotik yang digunakan sebesar 50 µg/ml dan 100 µg/ml media. Hasil uji sensitivitas disajikan pada tabel di bawah ini.
Tabel 3. Hasil uji sensitivitas antibiotik pada konsentrasi 50 µg/ml
Isolat Rifampisin Penisilin G Kanamisin Streptomisin
F5 - + + +
F13 - + - -
F15 - - - -
F16 - + - -
F17 - - - -
F18 - + - -
F19 - + - -
F25 - + - -
F26 - + - -
F27 - + - -
F31 - + - -
F34 + - + -
F39 - - - -
F41 - - - -
F43 - + - -
F45 - - + -
V. harveyi MR 5339 Rf R + + + -
Keterangan: (-):sensitif; (+):resisten
Berdasarkan tabel 3, isolat F34 dan V. harveyi MR 5339 Rf R bersifat resisten terhadap antibiotik rifampisin. Isolat F15, F17, F34, F39, F41, F45 bersifat resisten terhadap penisilin G. Isolat F5, F34, F45 dan V. harveyi MR 5339 Rf R bersifat resisten terhadap kanamisin sedangkan semua isolat (kecuali isolat F5) bersifat sensitif terhadap streptomisin. Sifat resisten terhadap antibiotik pada bakteri dapat disebabkan oleh dua jenis mekanisme yaitu mutasi kromosom dan akuisisi plasmid. Mutasi pada kromosom tidak dapat ditransfer pada bakteri lain tetapi plasmid mampu mentransfer dengan cepat sifat resisten tersebut (Lewin 1992 dalam Schnick 2001). Sifat resistensi pada mikroorganisme juga diakibatkan oleh: (1) kemampuan menghasilkan enzim yang mampu menginaktivasi antibiotik; (2) adanya penambahan substansi kimia pada struktur kimia lain oleh
31 (3) perubahan struktur protein pada dinding sel; (4) berkurangnya permeabilitas dinding sel sehingga antibiotik tidak dapat masuk ke dalam sel (Guilfoile 2007).
[image:51.595.111.477.269.560.2]Sifat resistensi ini dapat dipindahkan baik secara horizontal maupun vertikal (Madigan et al. 2003 dalam Reboucas et al. 2011). Baticados et al. (1990) dalam Tendencia and de la Pena (2001) melaporkan bahwa patogen penyebab luminous disease seperti Vibrio harveyi dan Vibrio splendidus yang diisolasi dari larva udang resisten terhadap erythromycin, kanamycin, pencillin G, dan streptomycin.
Tabel 4. Hasil uji sensitivitas antibiotik pada konsentrasi 100 µg/ml
Isolat Rifampisin Penisilin G Kanamisin Streptomisin
F5 - - - -
F13 - - - -
F15 - - - -
F16 - - - -
F17 - - - -
F18 - - - -
F19 - - - -
F25 - - - -
F26 - - - -
F27 - - - -
F31 - - - -
F34 - - - -
F39 - - - -
F41 - - - -
F43 - - - -
F45 - - - -
V. harveyi MR 5339 Rf R - - - -
Keterangan: (-):sensitif; (+):resisten
32 3.4 Metode Kultur Bersama
Uji in vitro kultur bersama dilakukan untuk mengetahui kemampuan bakteri kandidat dalam menghambat pertumbuhan bakteri patogen (V. harveyi MR 5339 Rf R). Uji ini dilakukan dengan membandingkan jumlah kepadatan bakteri V. harveyi MR 5339 Rf R yang tumbuh di tabung kontrol dengan tabung kultur bersama.
33 Gambar 2. Penghambatan V. harveyi MR 5339Rf R oleh kandidat probiotik pada uji in vitro metode kultur bersama
7,08 ± 0,47 9,29 ± 2,17
9,23 ± 2,259,27 ± 2,37
7,14 ± 0,45
8,45 ± 1,30
6,85 ± 0,10
8,77 ± 1,89 9,46 ± 1,45 9,35 ± 2,30
8,36 ± 1,17
9,39 ± 1,93
8,08 ± 1,42
10,15 ± 3,30
7,84 ± 1,08
9,17 ± 2,23
9,30 ± 2,17
0,0 0,5 1,0 1,5 2,0 2,5 3,0 3,5 4,0 4,5 5,0 5,5 6,0 6,5 7,0 7,5 8,0 8,5 9,0 9,5 10,0 10,5 11,0 11,5 12,0 12,5 13,0 13,5 14,0
F 5 F 13 F 15 F 16 F 17 F 18 F 19 F 25 F 26 F 27 F 31 F 34 F 39 F 41 F 43 F 45 Vibrio
harveyi K epa da ta n ba k ter i (lo g CF U/m l) Vibrio harveyi MR 5339 Rf R
ab a bc
cd bc cd a a a d cd a e cd
34 Tidak semua jenis bakteri mampu memproduksi inhibitory metabolites (senyawa penghambat) pada media agar karena terdapat kemungkinan produksi inhibitory metabolites akan lebih banyak jika kultur dilakukan pada media broth. Adanya perbedaan komposisi medium juga mempengaruhi produksi senyawa penghambat (Verschuere et al. 2000). Hal tersebut membuktikan bahwa penghambatan pertumbuhan populasi tidak selalu dapat dilihat melalui metode zona bening (Kirby Bauer Method).
Lactid Acid Bacteria (LAB) yang diisolasi dari saluran pencernaan mampu memproduksi komponen bakteriosin yang mampu menghambat pertumbuhan mikroorganisme lainnya dan secara tidak langsung akan mempengaruhi kompetisi dalam memperoleh energi di lingkungannya. LAB yang ditemukan dalam saluran pencernaan organisme akuatik seperti ikan mampu meningkatkan status kesehatan organisme tersebut (Verschuere et al. 2000). Sasanti (2005) melaporkan bahwa dari 110 isolat hasil isolasi dari terumbu karang, hanya 10 isolat saja yang potensial sebagai kandidat probiotik. Lima diantaranya terbukti mampu menghambat pertumbuhan Vibrio harveyi MR 5339 Rf R walaupun tidak menunjukkan adanya zona hambat pada uji in vitro.
Untuk mengetahui tingkat kepadatan bakteri yang dikultur sebelum masuk ketahapan kultur bersama, dilakukan pengukuran kepadatan bakteri dengan menggunakan spektrofotometer. Kepadatan bakteri diketahui dengan mengukur nilai optical density (OD) dari populasi sel yang dikultur selama 24 jam. Pengukuran OD dilakukan pada panjang gelombang 600 nm (lampiran 1).
3.5 Uji Patogenisitas Bakteri Kandidat Probiotik
35 ulangan. Pengamatan tingkat kelangsungan hidup pascalarva udang vaname dilakukan selama 5 hari. Tingkat kelangsungan hidup pascalarva udang vaname tertera pada gambar 3.
Keterangan:
A: Penambah