Bismillahirrahmanirrahiim,
Dan seandainya pohon-pohon di bumi menjadi pena dun laut menjadi tinta, ditam- bahkan tujzch laut la@ sesrrdah keringnya, niscaya tidak akan habis-habisnya ditulis- kan ilmu Allah dan hikmahNya, sungg~ih Allah Maha Perkasa lagi Maha Bijaksana.
(QS.
Luqman : 27)Kupersembahkan untuk
KWJlAN
KONDISI
F
SELUEASE
DARl
LIMBAH KELWP
DAN
SABUT
1
S A F R I A N I
F 27. 1357
1 9 3 5
FAKULTAS TEKNOLOGl PERTANIAM
INSTITUT PERTARIIAN
BOGOR
< ; :'
*...
Safriani. F 27.1757. Kaj ian Kondisi .'Fermentasi Pada Produksi Selulase dari Limbah Kelapa Sawdt (Tandan Kosong dan Sabut) Oleh Neurospora sitophila. Dibawah bimbingan A. Aziz Darwis, Illah Sailah dan Tun Tedja Irawadi.
RINGKASAN
I.
Prodaksi minyak kelapa spwit pada tahun 1995 diperki- ',i
rakan sebesar 4.5 juta ton. Dari s$tu ton tandan buah se-
i
gar akan dihasi1kan:minyak sawit sebesar 0.21 ton dan inti
ss~,:it ssbanyak 0.05 ton, sedangkan sisanya berupa limbah
dalan bentuk tandan kosong, sabut dan cangkang biji yang
jun~lahnya masing-masing sebesar 23, 13.5 dan 5.5 persen.
Tandan kosong dan sabut kelapa sawit dapat digunakan
se'agai substrat untuk memproduksi selulase karena mengan-
dung holoselulosa dan lignin yang cukup tinggi yaitu
sebesar 65.28 persen dan 35.2 persen untuk tandan kosong
d a n 55 persen dan 21.92 persen untuk sabut kelapa sawit.
Produksi selulase antara lain dipengaruhi oleh kondisi
proses fermentasi. Parameter lingkungan yang berpengaruh
pada sintesa enzim antara lain pH, suhu, aerasi dan kekua-
tan ionik, sehingga diperlukan optimasi kondisi proses.
Penelitian ini bertujuan untuk menentukan tingkat suhu dan
aerasi serta lama fermentasi terbaik pada produksi selu-
lase dengan menggunakan Neurospora sitophila pada substrat
tandan kosong dan sabut kelapa sawit. Tandan kosonq dan
sabut kelapa sawit yang digunakan masing-masing beru-
sedangkan Neurospora sitophila yang digunakan adalah yang
telah berumur lima hari.
Sistem fermentasi yang digunakan pada penelitian ini
merupakan modifikasi dari sistem fermentasi Multiple Mini
Packed Bed (MPB) dengan ketebalan substrat 2 sentimeter.
Penentuan tingkat suhu dan aerasi terbaik dilakukan dengan
melakukan fermentasi pada suhu 25OC, 28OC dan 31°C dan la-
ju aerasi 0, 0.1, 0.3 dan 1 l/jam/g substrat. Untuk suhu
25OC inkubator diletakkan di dalam ruangan yang suhunya
terkontrol pada suhu tersebut dan untuk suhu 28OC diletak-
kan pada suhu kamar, sedangkan untuk suhu 31°C dilengkapi
dengan elemen pemanas dan thermostat yang diatur pada suhu
tersebut. Fermentasi dilakukan selama 10 hari, dimana
pengamatan dilakukan pada hari ke-2, 4 , 6, 8, dan 10.
Pada setiap kali pengamatan dilakukan analisa aktivitas
enzim yang dihasilkan yaitu FP-ase dan CMC-ase.
Aktivitas FP-ase dan CMC-ase tertinggi diperoleh pada
suhu 2S°C dan laju aerasi 1 l/jam/g substrat yaitu masing-
masing sebesar 2.04 IU/ml dan 69.26 IU/ml. Aktivitas FP-
ase tertinggi ini diperoleh pada hari kedelapan fermenta-
si, sedangkan aktivitas CMC-ase pada hari keenam fermenta-
si.
Hasil analisis statistik menunjukkan bahwa tingkat
suhu, aerasi dan lama fermentasi mempengaruhi aktivitas
.
KAJIAN KONDISI FERMENTASI PADA PRODUKSI SELULASE
DARI LIMBAH KELAPA SAWIT (TANDAN KOSONG DAN SABUT)
OLEH
Neurosporu
sitophilaOleh
SAFRIANI
F 27 1357
Sebagai salah satu syarat untuk memperoleh gelar SARJANA TEKNOLOGI PERTANIAN pada Jurusan Teknologi Industri Pertanian,
Fakultas Teknologi Pertanian, Institut Pertanian Bogor
1995
FAKULTAS TEKNOLOGI PERTANIAN
INSTITUT PERTANIAN BOGOR
INSTITUT PERTANIAN BOGOR FAKULTAS TEKNOLOGI PERTANIAN
KAJIAN KONDISI FERMENTAS1 PADA PRODUKSI SELULASE DARI L M B A H KELAPA SAWIT (TANDAN KOSONG DAN SABUT)
OLEH Nercrosporu sitophilu
SKRIPSI
Sebagai salah satu syarat untuk memperoleh gelar SARJANA TEKNOLOGI PERTAVIAN
pada Jurusan TEKNOLOGI NDUSTRI PERTANIAN
FAKULTAS TEKNOLOGI PERTAiTAN INSTITUT PERTANIAN BOGOR
Oleh
SAFRIANI F 27.1357
Dilahirkan pada tanggal 23 April 1971 di Banda Aceh
KATA PENGANTAR
Alhamdulillahirabbil'alarniin.
Puji syukur penulis panjatkan kepada Allah Swtyang telah melimpahkan rahmat dan karuniaNya sehingga penulis dapat menyelesaikan
skripsi ini.
Skripsi ini disusun sebagai salah satu syarat untuk memperoleh sarjana
Teknologi Pertanian pada jurusan Teknologi Industri Pertanian, Fakultas Teknologi
Pertanian, Institut Pertanian Bogor. Skripsi ini merupakan hasil penelitian yang dilaku-
kan dari bulan Oktober 1994 sampai bulan Februari 1995 di Laboratorium Terpadu
Analisis Kimia FMIPA IPB Bogor.
Pada kesempatan ini penulis menyampaikan terima kasih yang sebesar-besarnya
kepada Dr.Ir. A. Aziz Darwis, MSc., Dr.Ir. Illah Sailah, MS., dan Dr.Ir. Tun Tedja
Irawadi, MS., selaku dosen pembimbing yang telah memberikan bimbingan hingga se-
lesainya penyusunan skripsi ini serta semua pihak yang telah membantu penelitian
hingga selesainya skripsi ini. Ucapan terima kasih penulis sampaikan pula kepada
Ayah, Mamak dan adik-adik atas segala do'a restu, dorongan moril dan pengorbanan
yang diberikan, sehingga penulis dapat menyelesaikan tugas akhir ini.
Penulis menyadari sepenuhnya bahwa skripsi ini masih jauh dari sempurna.
Namun demikian, penulis berharap agar skripsi ini dapat bermanfaat bagi semua pihak
yang memerlukannya.
DAFTAR IS1
. . .
PEND+WLUAN 1
TINJAUAN PUSTAKA
. . .
4A
.
LIGNOSELULOSA. . .
4B
.
SELULASE. . .
8C
.
MIKROORGANISME PENGHASIL SELULASE. . .
11D
.
KONDISI FERMENTASI. . .
14BAXAN DAN METODA
. . .
21A
.
B A K U DAN ALAT. . .
211
.
Substrat. . .
212
.
Mikroorganisme. . .
213
.
Bahan Kimia. . .
214
.
Peralatan. . .
22B
.
METODA. . .
241
.
Persiapan Bahan Baku. . .
242
.
Analisis Komposisi Substrat. . .
243
.
Isolasi dan Persiapan Mikoorganisme. . .
24. . .
...
4.
Pernbuatan Media Fermentasi:
25 5.
Penentuan Suhu dan Tingkat Aerasi Terbaik.
25 C.
WLNCANGAN PERCOB AAN. . .
26. . .
IV
.
HASIL DAN PEMBAHAS AN. . .
30A
.
KOMPOSISI MEDIA FERMENTASI. . .
30B
.
KAPANG Neurospora sitopila. . .
31C
.
KONDISI LINGKUNGAN FERMENTASI. . .
331
.
Pengaruh Suhu terhadap Aktivitas Enzim. . . .
332
.
Pengaruh Aerasi terhadap Aktivitas Enzim..
40V
.
KESIMPULAN DAN SARAN. . .
52A
.
Kesimpulan. . .
52B
.
Saran. . .
53DAFTAR TABEL
Halaman
Tabel 1. Analisis Kimia dari Sabut dan Tandan Sawit. 5
DAFTAR GAMBAR
Halaman
[image:11.524.67.471.148.647.2]. . .
Gambar 1. Rumus Molekul Selulosa 5
Gambar 2. Mekanisme Hidrolisis Selulosa oleh Enzim Selulase (Montenecourt dan Eveleigh,
1979 dalam Enari, 1983)
. . .
9Gambar 3. Sistem Pembentukan Selulase (Mandels dan Weber, 1969 dalam Tun Tedja Irawadi
...
11. . .
Gambar 4. Pertumbuhan Kapang pada Substrat Padat 13Gambar 5. Skema Keterkaitan Kondisi Lingkungan de- ngan Parameter Lingkungan (Weiland, 1988) 16
Gambar 6. Inkubator Sistem Fermentasi MPB yang te- lah dimodifikasi
. . .
23Gambar 7. Skema Produksi Selulase pada Berbagai Tingkat Suhu dan Aerasi
...
27Gambar 8. Kapang Neurospora sitophila yang berumur Lima Hari
...
32Gambar 9. Aktivitas FP-ase pada Berbagai Tingkat Suhu
. . .
34Gambar 10. Aktivitas CMC-ase pada Berbagai Tingkat Suhu
. . .
37Gambar 11. Aktivitas FP-ase pada Suhu 25O C dengan Berbagai Tingkat Aerasi
. . .
41Gambar 12. Aktivitas FP-ase pada Suhu 28O C dengan Berbagai Tingkat Aerasi
. . .
42 Gambar 13. Aktivitas FP-ase pada Suhu 31° C denganBerbagai Tingkat Aerasi
. . .
43Gambar 15
.
Aktivitas CMC-ase pada Suhu 28O C dengan Berbagai Tingkat Aerasi. . .
46Gambar 16
.
Aktivitas CMC-ase pada Suhu 31° C dengan Berbagai Tingkat Aerasi. . .
47Gambar 17
.
Aktivitas FP-ase pada Suhu 28O C dengan Aerasi sampai 2 l/jam/g. . .
4 8Gambar 18
.
Aktivitas CMC-ase pada Suhu 28O C dengan. . .
Aerasi sampai 2 l/jam/g 49
DAFTAR LAMPIRAN
Halaman
Lampiran 1. Bahan-bahan Kimia untuk Analisis
Proksimat dan Analisa Aktivitas Enzim 57
Lampiran 2. Prosedur Analisis
. . .
58Lampiran 3. Data Hasil Analisa Aktivitas FP-ase.. 66
Lampiran 4. Data Hasil Analisa Aktivitas CMC-ase. 67
Lampiran 5a. Analisis Ragam Aktivitas FP-ase
. . .
68Lampiran 5b. Hasil U j i Lanjut Duncan Aktivitas
FP-ase
. . .
68Lampiran 6a. Analisis Ragam Aktivitas CMC-ase
. . .
69Lampiran 6b. Hasil Uji Lanjut Duncan Aktivitas
Ind~nesia merupakan negara penghasil kelapa sawit ter-
hesar kedua di dunia. Usaha-usaha untuk meningkatkan pro-
duksi =inyak kelapa sawit terus dilakukan dengan jalan
...
I,,<-,..huka,.-.
lahan-lahan perkebunan baru. Sampai tahun 1993,Derdasarkan data PPKS (Pusat Penelitian Kelapa Sawit),
jumiah produksi minyak kelapa sawit atau Crude Palm Oil
(CPO) sekitar 3 276 000 ton. Pemerintah Indonesia telah
ner.targetkan bahwa pada tahun 1995 akan diproduksi minyak
sawit sebesar 4.5 juta ton (Darnoko, 1992).
Selana pengolahan kelapa sawit menjadi minyak sawit,
d a r I satu ton tandan buah segar (TBS) akan dihasilkan mi-
riyal.; sarait sebesar 0.21 ton dan inti sawit sebanyak 0.05
ton, sisanya merupakan limbah dalam bentuk tandan kosong,
sabut dan cangkang biji yang jumlahnya masing-masing se-
? - fi>.t.ar < 23, 13.5 dan 5.5 persen (Darnoko, 1992).
Usaha untuk memanfaatkan limbah padat kelapa sawit
sampai saat ini belum dilakukan secara intensif, kecuali
untuk inti sawit dan bungkilnya. Tandan kosong kelapa sa-
wit pads umumnya dibakar dalam incinerator dan abunya di-
manfaatkan untuk pupuk Kalium, sedangkan sabut dan cang-
fiang dirnanfaatkan sebagai bahan bakar boiler atau untuk
bahan pengeras jalan-jalan di sekitar perkebunan.
Konponen utama limbah padat kelapa sawit adalah selu-
iosa. Kandungan holoselulosa (selulosa dan hemiselulosa)
dan lignin pada lignoselulosa masing-masing berkisar
antara 62
-
64 persen dan 21-
23 persen (Sivalingan, 1983dalam Tun Tedja-Irawadi, 1991).
Lignoselulosa dapat dimanfaatkan sebagai substrat un-
i:uk memproduksi selulase. Enzim ini selanjutnya dapat
dimanfaatkan pada proses biokonversi selulosa. Penelitian
nengenai pemanfaatan lignoselulosa sebagai substrat pro-
duksi selulase telah banyak dilakukan.
Produksi selulase antara lain dipengaruhi oleh kondisi
proses fermentasi. Parameter kondisi yang berpengaruh
pada sintesa enzim adalah pH, suhu, aerasi dan kekuatan
ionik. Suhu optimal pertumbuhan kapang tidak sama dengan
suhu produksi enzim. Suhu untuk pertumbuhan biasanya le-
bih tinggi daripada suhu produksi enzim (Prior et al.,
1990).
Selama fermentasi berlangsung akan dihasilkan panas
yang cukup tinggi. Panas ini harus segera dihilangkan ka-
rena suhu akan mempengaruhi pertunasan spora, pertumbuhan,
pembentukan produk dan sporulasi kapang. Salah satu cara
yang dapat digunakan untuk menurunkan panas yaitu dengan
jalan mengalirkan udara steril ke dalam fermentor
(aerasi). Tingkat aerasi dipengaruhi oleh sifat mikroor-
ganisme yang digunakan, tingkat oksigen yang dibutuhkan
untuk sintesis produk, jumlah panas metabolik yang harus
Itat dimana C02 dan metabolit-metabolit lain yang mudah
menguap harus dihilangkan dan tingkat ruang udara yang
tersedia di dalam substrat (Lonsane et al., 1985).
Kadar air yang tinggi di dalam bahan sebagai akibat
dari suhu yang terlalu rendah, akan menyebabkan ruang ko-
song antar partikel dipenuhi oleh air sehingga cenderung
msnurunkan laju pindah massa O2 dan C02 dan laju pindah
panas. Sebaliknya, suhu yang terlalu tinggi akan mengaki-
batkan kurangnya kelembaban substrat dan tingginya panas
metabolik yang dihasilkan (Weiland, 1988)
.
Selain itupertimbangan ekonomis diperlukan untuk pengembangan ke
skala yang lebih besar. Oleh sebab itu suhu dan tingkat
aerasi yang tepat akan menghasilkan kondisi lingkungan
optimal untuk pertumbuhan kapang dan aktivitas enzim yang
dihasilkan.
Ruang lingkup penelitian ini meliputi perlakuan penda-
huluan yaitu pengeringan dan pengecilan ukuran tandan ko-
song dan sabut kelapa sawit, analisis komposisi substrat,
produksi selulase pada substrat dengan berbagai tingkat
suhu dan aerasi dengan menggunakan Neurospora sitophila,
dan analisis aktivitas enzim yang dihasilkan.
Penelitian ini bertujuan untuk menentukan kondisi su-
hu dan tingkat aerasi yang tepat serta lama fermentasi
optimal untuk produksi selulase menggunakan Neurospora
IT.
TINJAUAN
PUSTAKA
A.
LIGNOSELULOSA
Komponen utama limbah padat kelapa sawit adalah
selulosa, hemiselulosa dan lignin. Kandungan masing-
masing komponen tersebut di dalam lignoselulosa sebesar
4 : 3 : 3 (Tsao et al., 1978)
.
Sebagai contoh, kayulunak mengandung 42, 25 dan 28 persen selulosa, hemise-
lulosa dan lignin, sedangkan tongkol jagung mengandung
40, 36 dan 16 persen.
Menurut Tun Tedj a- Irawadi (1991) limbah padat ke-
lapa sawit rnengandung bagian karbohidrat dan lignin
yang cukup tinggi. Komposisi kandungan senyawa kimiawi
tandan kosong dan sabut kelapa sawit disajikan pada
Tabel 1.
Selulosa tidak pernah ditemukan dalam keadaan
murni di alam. Selulosa merupakan suatu linier homopo-
limes unit anhidroglukosa (2 000-10 000 unit glukosa)
yang diikat dengan ikatan 1,4 B- glikosidik.
Dua unit glukosa yang berdekatan akan berikatan
dengan cara melepaskan satu molekul air, yang terbentuk
dari gugus-gugus hidroksil pada atom C kesatu dan
keempat. Posisi beta dari grup-OH pada atom C akan
berhubungan dengan unit glukosa lain pada C1
-
C, dariTabel 1. Analisis kimia dari sabut dan tandan sawit (persen bahan kering)a
Bahan Lemak Protein selulosa Zignin Hemiselulosa
Sabut 6.95 6.94 28.28 27.86b 3 4 . 7 ~ ~ 29. 46C
Tandan 5.35. 4.45 32.55 2 ~ . 5 4 ~ 31.70~
a ) ~ u n Tedja Irawadi (1991)
b)~etoda Klason (Johnson, 1937) C)Metoda oksidasi permanganat d, NDF-ADF
Selulosa merupakan polimer glukosa yang homogen,
sedangkan molekul-molekul hemiselulosa merupakan poli-
mer dari pentosa (xilosa dan arabinosa, heksosa (mano-
sa) dan beberapa asam gula. Lignin sendiri merupakan
makromolekul polifenolik (Tsao et dl., 1978).
Terdapat dua jenis ikatan hidrogen pada struktur
selulosa, yaitu ikatan hidrogen intramolekuler yang
mempertahankan kekakuan rantai selulosa, dan ikatan
intermolekuler yang menyebabkan rantai selulosa saling
berikatan membentuk suatu mikrofibril (Achmadi, 1989).
Beberapa mikrofibril ini kemudian membentuk fibril dan
akhirnya menjadi serat selulosa. Struktur fibril dan
kuatnya ikatan hidrogen menyebabkan selulosa bersifat
[image:18.521.85.483.273.589.2]Unit S e l o b i o s i 1 . 0 3 nm
6
6 H2COH
v--'
0 0 0H ,/ H R a n t a i s e l u l o s a
,k..
OH3 2
I! OH
?(E
II OH [image:19.521.66.490.67.658.2]Ujung non r e d u k s i Ujung r e d u k s i
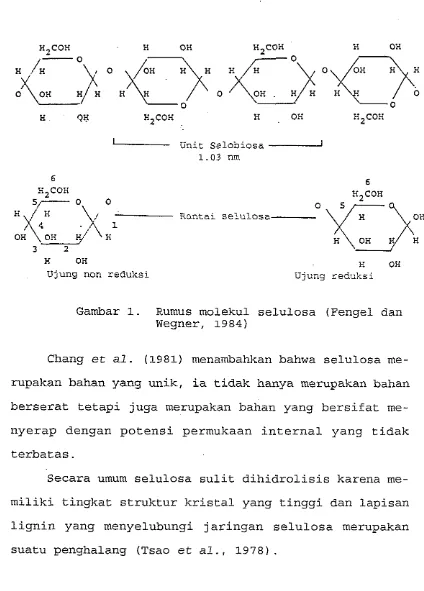
Gambar 1. Rumus molekul selulosa (Fengel dan Wegner, 1984)
Chang et al. (1981) menambahkan bahwa selulosa me-
rupakan bahan yang unik, ia tidak hanya merupakan bahan
berserat tetapi juga merupakan bahan yang bersifat me-
nyerap dengan potensi permukaan internal yang tidak
terbatas.
Secara umum selulosa sulit dihidrolisis karena me-
miliki tingkat struktur kristal yang tinggi dan lapisan
lignin yang menyelubungi jaringan selulosa merupakan
Bagian selulosa yang mudah dihidrolisis disebut
bagian amorf dari selulosa. Umumnya selulosa mengan-
dung 15 persen bagian amorf dan 85 persen bagian kris-
talin.
Hemiselulosa adalah polisakarida yang mempunyai
berat molekul yang lebih kecil daripada selulosa.
Molekul hemiselulosa lebih mudah menyerap air, bersifat
plastis dan mempunyai permukaan kontrol antar molekul
yang lebih luas dibandingkan dengan selulosa, sehingga
dapat memperbaiki ikatan antar serat pada pembuatan
kertas (Judoamidjojo et al., 1989)
.
Rantai utamahemiselulosa dapat hanya terdiri dari satu macam mono-
mer saja (homopolimer), misalnya xilan, atau dapat
terdiri dari dua atau lebih monomer (heteropolimer),
misalnya glukomanan (Fengel dan Wegener, 1984 di dalam
Tun Tedja-Irawadi, 1991).
Lignin merupakan senyawa polimer yang berikatan
dengan selulosa dan hemiselulosa pada jaringan tanaman.
Satuan penyusun lignin yaitu fenil propana yang tersub-
stitusi pada dua atau tiga posisi dalam cincin ben-
zennya. Lignin umumnya tidak pernah ditemui dalam
bentuk sederhana di antara polisakarida-polisakarida
dinding sel, tetapi selalu berikatan dengan polisakari-
da tersebut (Fengel dan Wegener, 1984 di dalam Tun
B.
SELULASE
Selulase merupakan golongan enzim yang mampu memu-
tus ikatan &-1.4 pada substrat selulosa, selodekstrin,
selobiosa dan turunan selulosa lainnya. Enzim ini
menurut Gong dan Tsao (1979) terdiri atas tiga kelompok
besar enzim dalam kompleks selulase,
a.
Endoglukanase (1,4-&-D-glukan, 4-glukanohidrolase, atau endo-B-1,4-glukanase atau &-1,4-glukanohidro-lase; EC.3.2.2.4) atau umumnya dikenal dengan nama
CMC-ase atau Cx selulase,
b. Selobiohidrolase (1,4-i3-D-glukan selobiohidrolase
atau ekso-IS-1,4-glukanase atau fS-l,4-glukan selobio-
hidrolase; EC 3.2.1.91) atau umumnya dikenal sebagai
Avicelase atau C1 selulase, dan
c.
fS-glukosidase (&-D-glukosida glukohidrolase atau 15-1,4-glukosidase; EC.3.2.1.21).
Ketiga enzim ini bekerja sama menghidrolisis se-
lulosa yang tidak larut menjadi glukosa sehingga akti-
vitas gabungan ketiga enzim dapat diukur dengan meman-
tau jumlah glukosa yang dihasilkan (Gong dan Tsao,
1979). Mekanisme hidrolisis selulosa oleh selulase da-
daerah kristalin vv.w daerah amorf
?
I
-
2
a ~ndo-glikanase (EG)
~elobiohidrolase . . (CBH)
1
"T
EG atau
CBH
3.
d 0
-
glukosidaseSelooligosakarida
,
-
Glukosa [image:22.518.77.486.55.493.2]dan selobiosa
-
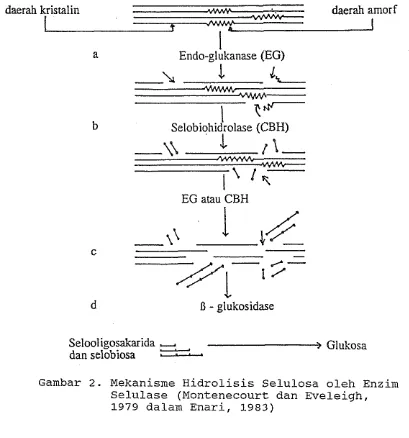
Gambar 2. Mekanisme Hidrolisis Selulosa oleh Enzim Selulase (Montenecourt dan Eveleigh, 1979 dalam Enari, 1983)
Endoglukanase (Cx) memiliki kemampuan menghidro-
lisis selulosa secara acak, menghasilkan selodekstrin,
selobiosa dan glukosa. 'Aksi dari enzim ini berupa pe-
nurunan viskositas dari substrat dapat larut. Endoglu-
kanase ini menyerang bagian amorf dari serat selulosa
sehingga membuka jalan bagi enzim selobiohidrolase
(C1).
Selobiohidrolase yang dikenal sebagai komponen C1,
oleh T. reesei. Enzim ini bekerja dengan cara melepas-
kan unit-unit selobiosa dari ujung non-reduksi rantai
selulosa. Produk hidrolisis utamanya adalah selobiosa.
J3-glukosidase adalah enzim yang dapat menghidrolisis
selobiosa dan selooligomer-selooligomer pendek lainnya
menjadi glukosa.
Selulase merupakan enzim ekstraselular yaitu enzim
yang bekerja menghidrolisis substrat-substrat yang be-
rat molekulnya besar. Pada mikroorganisme enzim ini
umumnya berfungsi memproduksi nutrisi dari polimer-
polimer biologi yang terdapat di sekeliling sel (Frost
dan Moss, 1987)
.
Selulosa merupakan penginduksi universal dalam
sintesis selulase. Penelitian yang dilakukan oleh be-
berapa peneliti menunjukkan bahwa sejumlah kecil enzim
selulase diproduksi oleh sel dan dilepaskan ke medium
pertumbuhan. Enzim ini kemudian menghidrolisis selulo-
sa menjadi selobiosa yang akan bertindak sebagai peng-
induksi (Gong dan Tsao, 1979).
Selobiosa merupakan penginduksi enzim selulase
yang baik bagi sebagian besar kapang, misalnya Neuro-
spora crassa, Chaetomium celluloliticum (Leisola et
a1
.
,
1985 di dalam Tun Tedja, 1991).
Selobiosa dapatmemacu pertumbuhan kapang dan produksi enzim, khususnya
apabila kapang tumbuh pada kondisi sub-optimal yang
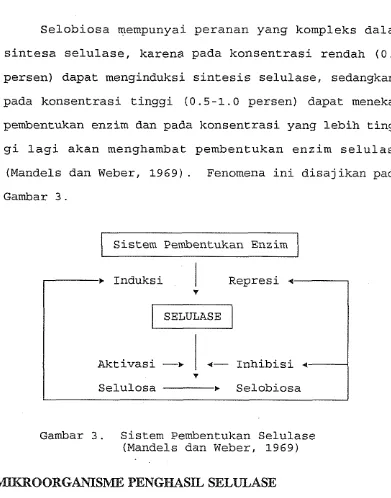
Selobiosa mempunyai peranan yang kompleks dalam
sintesa selulase, karena pada konsentrasi rendah (0.1
persen) dapat menginduksi sintesis selulase, sedangkan
pada konsentrasi tinggi (0.5-1.0 persen) dapat menekan
pembentukan enzim dan pada konsentrasi yang lebih ting-
gi lagi akan menghambat pembentukan enzim selulase
[image:24.521.75.466.61.559.2](Mandels dan Weber, 1969). Fenomena ini disajikan pada
Gambar 3 .
I
Sistem Pembentukan EnzimI
t Induksi
I
Represi 4v
Aktivasi -t 4- Inhibisi 4
1
Selulosa b Selobiosa-1
Gambar 3. Sistem Pembentukan Selulase (Mandels dan Weber, 1969)
6.
OORGANISME PENGNASUL SELULASE
Menurut Judoamidjojo et al. (1989)
,
sejumlah besarbakteri dan kapang mampu menghidrolisis selulosa sampai
tahapan tertentu. Mikroba tersebut menggunakan selulo-
Beberapa bakteri tidak menghasilkan enzim selulo-
litik dalam jumlah banyak, tetapi masih tetap digunakan
karena kemampuannya untuk tumbuh pada bahan-bahan
selulosik. Salah satu bakteri yang telah dipelajari
secara intensif yai,& Cellulomonas uda (Srinivasan,
1 9 7 5 dalam Enari, 1983). Selain itu juga dijumpai
jenis-jenis bakteri lain yang dapat menghasilkan selu-
lase yaitu Pseudomonas, Ruminococcus, Bacteroides, dan
Clos tridium. Clostridium ini merupakan bakteri pengha-
sil selulase terbaik diantara jenis bakteri lainnya
(Zeikus, 1980 dalam Enari, 1983)
.
Kemampuan untuk menghasilkan enzim selulolitik
ekstraselular banyak dijumpai pada golongan kapang,
diantaranya Trichoderma reesei
,
T. viride, Aspergillusoryzae, A. niger, dan Phanerochae ta chrysosporium.
Penelitian ini menggunakan N. sitophila sebagai
inokulan. N. sitophila merupakan kapang yang termasuk
dalam Subdivisi Eumycophyta, Klas Ascomycetes, Ordo
Sphriales dan Famili Sordoriaceae. Kapang ini mudah
menyebar dan berkembang biak secara cepat terutama
dengan cara aseksual, dan biasanya ditemukan pada
tingkat konidia. Spora seksualnya jarang ditemui
karena hanya dibuat dalam jumlah sedikit (Alexopoulus
dan Mims, 1979).
Penelitian-penelitian mengenai kemampuan Neuro-
kapang yaitu memproduksi selulase dan &-glukosidase
juga telah banyak dilakukan. Media yang digunakan
beraneka ragam diantaranya adalah tongkol jagung dan
bagas tebu (Oguntimein et dl., 1992). Banerjee et dl.
(1994) juga menggunakan Neurospora si tophila pada
produksi protein kapang dengan limbah jagung sebagai
sumber karbonnya.
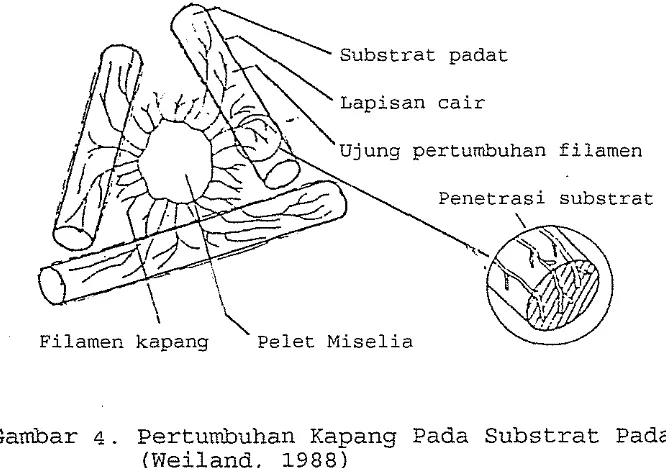
[image:26.521.121.454.397.631.2]Pertumbuhan kapang pada substrat padat dapat di-
gambarkan seperti akar di dalam tanah (Gambar
a
).
Bagian miselia memenuhi ruang antar partikel padat,
tetapi selama tidak ada air bebas dan nutrien dalam
Lapisan c a i r
a
' ~ j u n g pertumbuhan filarnen-
Filamen kapang \ ~ e l e t M i s e l i aruang ini, laju pertumbuhan akan tergantung pada kemam-
puan bagian miselia mencapai partikel substrat beri-
kutnya yang tersedia. Keterbatasan geometris secara
parsial dapat diatasi dengan memilih ukuran inokulum
yang cukup besar dimana bagian miselia atau spora dapat
pada awalnya membentuk koloni pada sebagian besar
partikel substrat.
D.
KONDISI FERMENTASI
Fermentasi media padat adalah proses fermentasi
yang substratnya tidak dapat larut dan tidak mengandung
air bebas, tetapi cukup mengandung air untuk keperluan
mikroba (Chahal, 1985)
.
Fermentasi media padat secaraalami berlangsung pada medium dengan kadar air yang
berkisar antara 60 - 80 persen, karena pada keadaan ini
medium mengandung air yang cukup untuk pertumbuhan
mikroba (Aidoo et d l . , 1982)
.
Menurut Maggy-T. Suhartono (1989)
,
f ermentasi me-dia padat pada produksi enzim umurnnya memberikan hasil
lebih baik karena jumlah substrat yang tersedia lebih
banyak yaitu padatan sekitar 20-50 persen. Selain
lebih banyak, enzim yang dihasilkan biasanya beragam.
Cara fermentasi media padat disukai untuk menghasilkan
berbagai enzim ekstraseluler. Enzim ini menghidrolisis
polimer substrat fermentasi menj adi nutrisi yang dapat
Fermentasi media padat mempunyai keuntungan dan
kerugian. Menurut Smith (1990) keuntungannya antara
lain substrat yang digunakan sederhana dengan komponen
alami yang lebih murah dibandingkan dengan komponen
sintesis yang mahal. Kontaminasi mikroorganisme ren-
dah, sehingga seringkali tidak perlu sterilisasi serta
proses hilir yang mudah. Persyaratan aerasi dapat di-
penuhi dengan mudah melalui difusi gas atau aerasi.
Hasil produksi tinggi dan kebutuhan energi rendah di-
bandingkan dengan bioreaktor tangki berpengaduk. Frost
dan Moss ( 198 7 ) menambahkan bahwa dengan menggunakan
fermentasi media padat kondisi tumbuh kapang lebih men-
dekati keadaan yang biasa dijumpai di alam dan inokula-
si dapat langsung dari spora, tanpa melalui media ino-
kulum.
Sementara kerugian fermentasi media padat dianta-
ranya proses terbatas hanya untuk pertumbuhan kapang
saja, yaitu mikroba yang dapat mentolerir rendahnya ka-
dar air (Frost dan Moss, 1987). Timbulnya panas hasil
metabolisme dalam proses berskala besar dapat menimbul-
kan masalah. Demikian pula pemantauan proses misalnya
tingkat kadar air, biomassa, kadar O2 dan C02, sukar
dilaksanakan dengan akurat serta rancangan bioreaktor
belum sempurna dan laju pertumbuhan mikroorganisme le-
Produktivitas proses biologis secara umum ditentu-
kan oleh jenis mikroorganisme yang digunakan dan kondi-
si lingkungan selama jalannya fermentasi. Kondisi
lingkungan sangat tergantung pada jenis reaktor yang
digunakan, parameter proses yang terkontrol, morfologi
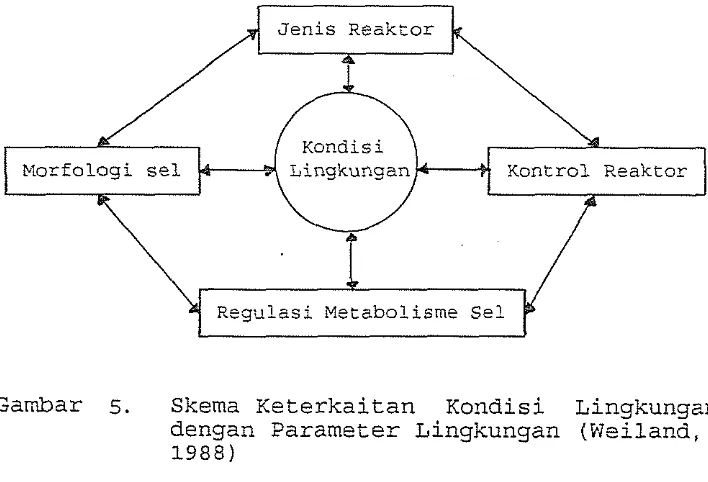
sel dan regulasi metabolisme sel (Gambar 5.).
Pada fermentasi padat semua parameter sangat ber-
hubungan satu sama lain, yang membatasi optimasi kondi-
si proses. Faktor-faktor yang paling membatasi fermen-
tasi media padat adalah sukarnya mengatur kelembaban,
suhu dan pH serta tidak memadainya pindah massa perse-
diaan O2 dan penghilangan C02 (Weiland, 1988).
[image:29.524.119.473.406.647.2]E.lorfoloai sel Kontrol Reaktor
Kondisi lingkungan seperti suhu, pH, aktivitas
air, tingkat oksigen serta konsentrasi nutrien dan pro-
duk mempengaruhi pertumbuhan mikrobial dan pembentukan
produk. Pada kultur terendam teraduk
,
kontrol ling-kungan relatif sederhana karena kehomogenan suspensi
sel-sel mikrobial dan kehomogenan larutan nutrien dan
produk dalam fasa cair. Suhu optimum untuk berbagai
sistem fermentasi padat mikrobial berbeda-beda dimana
suhu optimum untuk pertumbuhan tidak selalu sama dengan
pembentukan produk. Selanjutnya menurut Reid (1989)
dalam Prior et al. (1980) respon dari mikroorganisme
yang berbeda yang digunakan dalam delignifikasi biolo-
gis terhadap suhu bervariasi. Xebanyakan kapang (White
rot) adalah mesofil dengan suhu optimum 15-35O~, meski-
pun beberapa seperti Phanerochaeta chrysosporium meru-
pakan termofilik moderat.
Prior et a2. (1980) menuliskan bahwa pada beberapa
spesies, selektivitas jerami gandum yang didelignifika-
si menurun pada saat suhu meningkat dari 20°c menjadi
30°c. Selain itu juga dilaporkan bahwa suhu yang di-
naikkan (30°c) meningkatkan degradasi oleh Trametes
hirsuta. Berbeda dengan kapang white rot, Sporotrichum
pulverulentum dan Dichomitus squalens, laju degradasi-
nya meningkat dengan meningkatnya suhu dari 2 5 O ~ menja-
Suhu juga mempengaruhi proses-proses yang dikenda-
likan oleh difusi. Difusi molekuler ditunjukkan oleh
energi aktivasi sekitar 6 kkal/mol. Sebenarnya, ketika
tumbuh atau pembentukan produk menunjukkan ketergan-
tungan kurang dari 10 kkal, ha1 ini merupakan alasan
menduga keterbatasan difusi pada proses (Wang et al.,
1979).
Konsentrasi substrat yang tinggi per unit volume
yang merupakan ciri dari fermentasi media padat, menye-
babkan pembentukan panas mikrobial per unit volume
akan lebih besar daripada fermentasi cair. (Prior et
al., 1990)
.
Pembentukan panas total berkisar dari 419sampai 2 617 kj/kg bahan padatan terfermentasi dila-
porkan pada beberapa proses koji untuk biomassa, enzim
atau produksi asam organik (Aidoo e t al., 1982).
Panas yang dihasilkan harus dihilangkan, karena
suhu mempengaruhi pertunasan spora, pertumbuhan, pem-
bentukan produk dan sporulasi (Lonsane et al., 1985).
Metoda-metoda yang berbeda, seperti memasukkan udara
dalam jumlah besar ke dalam fermentor, menutup bagian
atas drum dengan lap yang dibasahi air, menempatkan
fermentor dalam ruang yang suhunya terkontrol, sirkula-
si air dalam jacket di sekeliling fermentor, atau me-
rendam fermentor dalam penangas air yang suhunya ter-
kontrol, digunakan untuk menghilangkan panas dari
25-32O~, yaitu selang suhu yang digunakan pada fermen-
tasi media padat.
Aerasi bahan fermentasi pada fermentasi skala la-
boratorium dan skala besar dicapai dengan mengalirkan
udara steril bertekanan. Tinykat aerasi optimal yang
diberikan dipengaruhi oleh sifat mikroorganisme yang
digunakan, tingkat oksigen yang dibutuhkan untuk sinte-
sis produk, jumlah panas metabolik yang harus dihilang-
kan dari bahan, ketebalan lapisan substrat, tingkat C02
dan metabolit-metabolit lain yang mudah menguap harus
dihilangkan, dan tingkat ruang udara yang tersedia di
dalam substrat (Lonsane et al., 1985)
.
Berdasarkanpenelitian Abdullah et al. (1985) laju aerasi 0.12
l/jam/g substrat dengan ketebalan substrat 1
-
2 senti-meter memberikan hasil terbaik pada fermentasi jerami
gandum oleh Chaetomium cellulolyticum.
Lebih jauh Prior et al. (1980) menjelaskan bahwa
aerasi memenuhi empat fungsi yaitu mempertahankan kon-
disi aerobik, untuk penghilangan C02, mengatur suhu
substrat dan mengatur kadar air.
Kadar air merupakan parameter kunci untuk mengatur
dan mengoptimasi proses fermentasi padat, karena kadar
air substrat mempengaruhi pertumbuhan kapang, aktivitas
pembentukan produk. Lebih lanjut kadar air dalam pori-
111. BAHAN DAN NIETODA
A.
BAHAN
DAN ALAT1. Substrat
Substrat yang dipakai dalam penelitian ini ada-
lah tandan kosong dan sabut kelapa sawit yang diper-
oleh dari pabrik pengolahan kelapa sawit perkebunan
Kertajaya, Banten, Jawa Barat. Substrat tersebut
sebelum digunakan terlebih dahulu dikeringkan di
panas matahari selama satu hari dan digiling.
2. Mikroorganisme
Mikroorganisme yanq digunakan adalah Neurospora
sitophila yang diperoleh dari hasil isolasi kapang
tersebut yang terdapat pada tandan kosong kelapa
sawit yang telah 3 hari di tempat penumpukan.
3. Bahan Kimia
Bahan kimia yang digunakan adalah larutan nu-
trisi yaitu yang digunakan oleh Tun Tedja-Irawadi
(1991) dan telah dimodifikasi berdasarkan hasil pe-
nelitian Ikatrinasari (1995). Larutan nutrisi ter-
sebut terdiri dari 12.25 g (NH4)2S04, 17.5 g KH2P04,
Urea yang semuanya dimasukkan ke dalam labu ukur 250
ml dan diencerkan sampai tanda tera. Pada setiap
gram substrat ditambahkan 2 ml larutan nutrisi.
Untuk ekstraksi digunakan Tween-80 0.1 persen,
sedangkan bahan-bahan untuk pengujian aktivitas FP-
ase dan CMC-ase, dan bahan-bahan untuk analisis
proksimat disajikan pada Lampiran 1.
4 . Peralatan
Alat-alat yang digunakan antara lain Sentrifuse
merek Clements 2000, Spektrofotometer Double Beam
COLLEMAN-124, autoklaf, Finn Pippete Diluter kapasi-
tas 1 ml dengan tingkat pengenceran maksimum 200
kali, pH-meter, penggiling, pengayak, Water Bath,
inkubator goyang, elemen pemanas solder, thermostat,
Aerator dan pengatur laju udara.
Bioreaktor yang digunakan berupa kotak yang
terbuat dari logam. Di dalam kotak tersebut terda-
pat rak tempat tabung-tabung berisi substrat dile-
takkan. Kotak tersebut dilengkapi dengan aerator
dan pengatur laju aerasi (Flow meter). Laju aerasi
ditentukan dengan mengkonversi satuan Flow meter ke
satuan l/jam/g substrat. Untuk suhu 25OC kotak ter-
sebut diletakkan di dalam ruangan yang suhunya ter-
kontrol pada suhu tersebut dan untuk suhu 2S°C di-
kotak dilengkapi dengan elemen pemanas dan thermo-
stat yang diatur pada suhu tersebut. Gambar biore-
[image:36.524.169.416.191.439.2]aktor ini disajikan pada Gambar 6.
Gambar 6. Inkubator Sistem Fermentasi MPB yang telah dimodifikasi
Alat-alat gelas yang digunakan diantaranya
pipet dengan berbagai ukuran volume, labu ukur,
erlenmeyer, gelas ukur dan tabung reaksi, sedangkan
alat-alat plastik berupa tabung-tabung plastik dan
B. METODA PENELITIAN
I. Persiapan Bahan Baku
Tandan kosong dan sabut kelapa sawit dikering-
kan pada panas matahari selama satu h.ari, digiling
lalu diayak. Tandan kosong kelapa sawit diayak
dengan ayakan 1 0 mesh, sedangkan sabut kelapa sawit
diayak dengan ayakan 30 mesh. Hasil penggilingan
yang diperoleh diukur kadar airnya dan disimpan di
tempat kering.
2. Analisis Komposisi Substrat
Substrat yang berukuran 2 0 mesh dianalisis kom-
posisi kimiawinya. Analisis yang dilakukan adalah
analisis kadar air, protein, lemak, serat kasar,
abu, lignin, selulosa, hemiselulosa dan mineral.
Prosedur analisis disajikan pada Lampiran 2.
3. Isolasi dan Persiapan Mikroorganisme
Kapang diisolasi dari tandan kosong kelapa sa-
wit yang telah 3 hari di tempat penumpukan. Selan-
jutnya ditumbuhkan pada media PDA sekitar 3
-
4 haridan pemurnian dilakukan tiga tahap. Sebelum diguna-
kan kapang ditumbuhkan kembali di media agar miring
selama 5 hari. Kemudian dalam setiap tabung agar
garam fisiologis. Suspensi merupakan inokulan yang
siap digunakan. Pada penelitian ini digunakan 1 ml
suspensi inokulan yang mengandung 0.3 - 1.6 x lo7
spora
.
4. Pembuatan Media Fermentasi
Tandan kosong dan sabut kelapa sawit yang sudah
digiling masing-masing berukuran 10 dan 30 mesh di-
campur dengan perbandingan 1 : 2. Kemudian substrat
ditimbang sebanyak 10 gram. Selanjutnya setelah
diberi larutan nutrisi disterilisasi pada tekanan 1
atm, 121°C selama 30 menit. Substrat diinokulasi
dengan 1 ml suspensi Neurospora sitophila serta
difermentasi selama 10 hari. Analisis aktivitas FP-
ase dan CMC-ase (Lampiran 2) dilakukan pada hari ke-
2, 4, 6, 8 dan 10.
5. Penentuan Suhu dan Tingkat Aerasi yang Terbaik
Sistem fermentasi yang digunakan merupakan
modifikasi dari sistem Multiple Mini Packed Bed.
Fermentasi dilakukan dengan memasukkan substrat yang
telah disterilisasi dan diinokulasi ke dalam tabung-
tabung plastik yang telah dilubangi di seluruh
permukaan tabung dengan jumlah lubang yang sama
untuk setiap tabung. Ketebalan substrat yang digu-
dari satu perlakuan untuk satu kali pengambilan
contoh, untuk memudahkan pengambilan contoh.
Tabung-tabung yang telah berisi substrat terse-
but diletakkan pada rak dan kemudian dimasukkan ke
dalam kotak-kotak logam. Kotak-kotak tersebut se-
lanjutnya ditempatkan di dalam ruangan yang suhunya
terkontrol pada 25OC. Untuk suhu 2S°C ditempatkan
pada suhu kamar, dan untuk suhu 31°C ditambahkan
elemen pemanas dan thermostat pada kotak logam dan
diatur pada tingkat suhu tersebut. Untuk masing-
masing suhu diberi aerasi dengan mengalirkan udara
lembab steril sebesar 0, 0.1, 0.3 dan 1.0 l/jam/gram
[image:39.521.91.482.490.643.2]substrat. Skema produksi enzim dapat dilihat pada
Gambar 7.
C . RANCANGAN PERCOBAAN
Rancangan percobaan yang digunakan adalah rancang-
an acak lengkap faktorial dengan tiga faktor (Sudjana,
1989). Perlakuan yang diberikan yaitu tingkat suhu
(A), tingkat aerasi (B) dan lama fermentasi (C).
Tingkat suhu dilakukan dengan tiga taraf (25OC, 2S°C,
dan 31°C) dan aerasi dengan empat taraf (0, 0.1, 0.3,
dan 1.0 l/jam/g substrat). Tiap perlakuan dilakukan
Campuran t a n d a n ( 1 0 mesh dan S a b u t ( 3 0 mesh)
Penambahan L a r u t a n N u t r i s i ( 1 0 % b / v )
I n o k u l a s i dengan N. si-
t o p h i l a berumur 5 h a r i
F e r m e n t a s i p a d a suhu 2 5 , 28, 31°c, Aerasi 0 , 0 . 1 , 0 . 3 , d a n 1 . 0 l / j / g s e l a m a 10 h a r i (pengamatan pada h a r i ke-2, 4 , 6 , 8 d a n 1 0
E k s t r a k s i enzim dengan Tween-80 0.1% p a d a se- t i a p k a l i pengamatan
A n a l i s a A k t i v i t a s Enzim
[image:40.521.166.386.86.561.2]I
Model umum rancangan percobaan tersebut adalah
sebagai berikut :
Yijk = /L
+
Ai+
Bj+
Ck+
(AB). .
+
(AC) ik+
(BC) jk1 3
+
(ABC)ijk+
eijkdimana :
'ijk = Respon pengamatan pengaruh taraf ke-i faktor A, taraf ke-j faktor B, taraf ke-k faktor C
pada ulangan ke-1
P = rata-rata umum
Ai = pengaruh perlakuan suhu ; i = 1,
...,
3 Bj = pengaruh perlakuan aerasi ; j = 1,...,
4 Ck = pengaruh hari pengamatan ; k = 1,...,
5(AB) i j = pengaruh interaksi suhu dan aerasi
(AC)ik = pengaruh interaksi suhu dan hari
(BC) jk = pengaruh interaksi aerasi dan hari
(ABC)ijk = pengaruh interaksi suhu, aerasi dan hari
ei jk = efek unit eksperimen karena perlakuan ke-i, j dan k
Hipotesis yang diuji pada penelitian ini adalah
hipotesis no1 (Ho) dimana semua perlakuan tidak mempe-
ngaruhi aktivitas enzim yang dihasilkan, dan hipotesis
alternatif (HI) apabila ada minimal satu dari perlakuan
Uji lanjut yang digunakan untuk menentukan perla-
kuan terbaik adalah uji wilayah berganda Duncan (Steel
dan Torrie, 1991)
.
D. N'AKTU DAN TEMPAT PENELITIAN
Penelitian dilakukan dari bulan Oktober 1994
sampai bulan Februari 1995 di Laboratorium analisis
IV. HASIL DAN PEhlBAHASAN
A. KOMPOSISI MEDIA F'ERMENTASI
Substrat yang digunakan berupa tandan kosong dan
sabut kelapa sawit. Analisis komposisi substrat yang
dilakukan terhadap tandan kosong dan sabut kelapa sawit
dimaksudkan untuk mengetahui kandungan substrat yang
digunakan sehingga penambahan larutan nutrisi dapat
disesuaikan
.
Hasil analisis komposisi substrat secaralengkap disajikan pada Tabel 2 . Terbukti bahwa kandun-
gan selulosa substrat cukup tinggi (tandan kosong 3 9 . 6 3
persen dan sabut 3 1 . 8 2 persen),
Tabel 2. Analisis Komposisi Substrat ( % bahan kering)
Jenis Analisis Tandan Kosong Sabut
Kadar air
( % I
Lemak ( % )
Protein 1%)
Mineral
K ( % )
ca
( % I
M9 ( 8 )
Mn
( % I
Zn Na Fe
Se u os
j
~ i g n i n ~ Hemiselulosa Serat kasar Kadar abu [image:43.518.75.474.389.660.2]sehingga substrat ini cukup baik untuk digunakan dalam
memproduksi selulase.
Kandungan lignin pada tandan kosong lebih tinggi
jika dibandingkan dengan sabut kelapa sawit. Kandungan
tersebut masing-masing adalah 35.2 persen untuk tandan
kosong dan 21.92 persen untuk sabut kelapa sawit.
Perbedaan kandungan lignin ini dijadikan dasar dalam
penentuan perbandingan penggunaan tandan kosong dan
sabut kelapa sawit. Substrat yang digunakan dengan
perbandingan satu bagian tandan kosong dan dua bagian
sabut kelapa sawit. Hal ini sesuai dengan hasil pene-
litian Rasmiati (1995), dimana tandan kosong dan sabut
kelapa sawit dengan perbandingan 1 : 2 yang digunakan
sebagai substrat pada produksi selulase oleh Neurospora
sitophila menghasilkan selulase dengan aktivitas ter-
tinggi.
B.
KAPANG
Neilrosporn sitophilaNeurospora sitophila yang ditumbuhkan pada media
agar miring pada awal masa pertumbuhannya ditandai
dengan tumbuhnya hifa dan membentuk miselium yang
berwarna putih. Setelah berumur satu hari muncul spora
yang berwarna oranye kemerahan. Warna merah hilang dan
jumlah spora semakin banyak dengan bertambahnya umur
kapang, dan pada hari kelima kapang berwarna oranye
yang digunakan pada produksi selulase. Umur kapang ini
disesuaikan dengan hasil penelitian Tun Tedja-Irawadi
(1991), dimana kapang yang berumur lima hari mampu
menghasilkan enzim dengan aktivitas tertinggi.
Kapang yang berumur lima hari telah mengandung
spora yang banyak sehingga pada awalnya bagian miselium
atau spora ini dapat membentuk koloni pada sebagian
besar substrat.
Pertumbuhan N. sitophila pada media agar miring
terlihat cenderung mendekati mulut tabung, hanya seba-
[image:45.521.132.464.330.618.2]gian kecil yang tumbuh pada agar miring (Gambar 8).
Hal ini membuktikan bahwa kapanq tersebut merupakan
jenis kapanq aerobik yaitu jenis kapang yang membutuh-
kan oksigen dalam pertumbuhannya.
1. Pengaruh Suhu Terhadap ~ktivitas Enzim
Suhu yanq digunakan pada penelitian ini adalah
25OC, 2S°C dan 31°C. Perlakuan terbaik dilihat dari
aktivitas selulase yang dihasilkan.
a. Aktivitas FP-ase
Substrat yang digunakan untuk penentuan ak-
tivitas FP-ase pada penelitian ini adalah kertas
sarinq Whatman No.1, dan produk yanq terukur ada-
lah glukosa. Substrat ini merupakan selulosa ti-
dak larut, sehingga dibutuhkan waktu reaksi yang
cukup lama agar enzim dapat berdifusi ke dalam
serat selulosa, yaitu selama 1 jam. Nilai ak-
tivitas FP-ase dinyatakan dalam satuan IU (Inter-
national Unit) per mililiter enzim kasar.
Aktivitas FP-ase pada berbagai tingkat suhu
disajikan pada Gambar 9. Pada Gambar 9 dapat
dilihat bahwa aktivitas FP-ase yang paling ren-
dah diperoleh pada suhu 25OC (1.34 IU/ml)
.
Suhumenurun dengan cepat, sehingga aktivitas enzim
yang diperoleh juga menurun (Maggy-T. Suhartono).
Laju aerasi yang diberikan juga dapat menurunkan
suhu fermentasi (Prior et al., 1 9 9 0 ) , sehingga
proses tidak berada pada kondisi optimal pertum-
buhan atau pembentukan produk.
Kadar air yang tinggi pada suhu rendah
[image:47.518.167.446.310.560.2]menyebabkan ruang kosong antar partikel dipenuhi
oleh air, sehingga fase gas akan terdorong keluar
(Weiland, 1988). Kondisi ini menghambat pertum-
buhan kapang aerobik yang membutuhkan oksigen da-
lam pertumbuhannya. Kondisi ini juga tidak meng-
untungkan karena memudahkan kontaminasi oleh ni-
kroba. Relative Humidity (RH) yang terukur pada
suhu ini adalah sekitar 90 persen.
Aktivitas FP-ase tertinggi (2.04 IU/ml) di-
peroleh pada suhu 2S°C. Pada suhu ini RH yang
terukur adalah 90 persen. Aerasi yang diberikan
juga membantu menghilangkan sebagian panas yang
dihasilkan sehingga suhu substrat dapat diperta-
hankan (Lonsane et al., 1985).
Jika proses penurunan panas tidak mampu
mengimbangi panas yang dihasilkan selama fermen-
tasi, maka suhu fermentasi akan meningkat.
Peningkatan suhu ini akan mengakibatkan kerusakan
struktur protein dan DNA yang memegang peranan
penting dalam metabolisme dan pertumbuhan sel
(Maggy-T. Suhartono), dan ha1 ini tentu saja akan
mempengaruhi aktivitas enzim yang dihasilkan.
Hal ini terbukti pada Gambar 9., dimana aktivi-
tas FP-ase yang dihasilkan pada suhu 31°C (1.70
IU/ml) lebih rendah jika dibandingkan dengan pada
Secara umum, pada ketiga tingkat suhu yang
diberikan, pada awal fermentasi aktivitas enzim
masih sangat rendah. Aktivitas enzim terus me-
ningkat sejalan dengan bertambahnya waktu fermen-
tasi dan menurun pada hari ke sepuluh. Hal ini
mengikuti pola pertumbuhan mikroorganisme yang
mengalami beberapa fase pertumbuhan yaitu fase
adaptasi, fase pertumbuhan logaritmik, fase
pertumbuhan lambat, fase pertumbuhan statis dan
f ase kematian. Menurut Maggy-T. Suhartono
(1989), organisme pembentuk spora biasanya mem-
produksi enzim pada fase pasca eksponensial.
Jadi dapat diduga bahwa pada saat aktivitas enzim
yang dihasilkan tinggi, maka kapang telah berada
pada fase tersebut.
Pada suhu 25OC aktivitas enzim pada awal
fermentasi masih sangat rendah (0.08 IU/ml),
kemudian meningkat dan mencapai puncaknya pada
hari keenam fermentasi, selanjutnya terus menurun
sampai hari terakhir fermentasi. Hari ke delapan
fermentasi pada suhu 2S°C memberikan nilai ak-
tivitas tertinggi (2.04 IU/ml). Akan tetapi pada
suhu 31°C aktivitas tertinggi (1.70 IU/ml) diper-
menurun drastis pada hari keenam fermentasi, un-
tuk kemudian naik lagi dan menurun pada hari ter-
akhir fermentasi.
b. Aktivitas CMC-ase
Aktivitas CMC-ase pada berbagai suhu dapat
dilihat pada Gambar 10. Dari gambar tersebut
terlihat bahwa aktivitas tertinggi CMC-ase seba-
gaimana pada FP-ase diperoleh pada suhu 2S°C
(69.26 IU/ml), sedangkan aktivitas paling rendah
diperoleh pada suhu 31°C (40.17 IU/ml).
2 4 6 8 10
[image:50.524.129.463.246.651.2]Lomo Fermentosi (Hori)
Pada suhu 25OC aktivitas CMC-ase tertinggi
(55.08 IU/ml) diperoleh setelah hari keenam
fermentasi, demikian pula halnya dengan suhu
28OC. Sedangkan pada suhu 31°C aktivitas ter-
tinggi tersebut diperoleh pada hari keempat
fermentasi. Pada hari keenam suhu tersebut
terjadi penurunan aktivitas enzim, untuk kemudian
naik lagi pada hari kedelapan. Fenomena yang
terjadi dapat dijelaskan sebagai berikut.
Pada awal fermentasi, glukosa diperlukan un-
tuk pertumbuhan kapang dan produksi enzim. Kemu-
dian produksi glukosa meningkat sejalan dengan
berjalannya traktu fermentasi. Tersedianya gula
pereduksi yang cukup banyak pada substrat, menye-
babkan selulase tidak aktif. Kondisi ini dikenal
sebagai efek represi katabolit. Selulase kembali
aktif memproduksi gula pereduksi pada saat perse-
diaan gula pereduksi telah habis (sedikit)
.
Halini sesuai dengan penjelasan Mandels dan Weber
(1969) di dalam Tun Tedja-Irawadi (1991) bahwa
konsentrasi yang tinggi dari selobiosa atau
sumber karbon yang dapat cepat di metabolisme
seperti glukosa atau gliserol dapat menghambat
pembentukan selulase.
Suhu mempengaruhi aktivitas enzim yang di-
suhu optimal pertumbuhan kapang dan produksi en-
zim cenderung menurunkan aktivitas enzim yang
dihasilkan. Suhu yang terlalu rendah menyebabkan
metabolisme sel menurun dengan cepat, sebaliknya
suhu yang tinggi menyebabkan kerusakan protein
dan DNA (Maggy-T. Suhartono).
Hasil penelitian yang dilakukan oleh Abdul-
lah et dl., (1985) juga menunjukkan hasil yang
tidak jauh berbeda yaitu dari empat tingkat suhu
yang diberikan (30°C, 35OC, 37OC dan 40°C) pada
fermentasi jerami gandum dengan menggunakan
Chaetomium cellulolyticum, suhu 37OC memberikan
hasil yang terbaik. Artinya suhu yang terlalu
rendah atau suhu yang terlalu tinggi tidak mem-
berikan hasil yang memuaskan.
Pada kondisi suhu tinggi, jika kadar air
yang ditambahkan tidak mampu mengimbangi kadar
air yang menguap akibat panas yang dihasilkan
oleh proses metabolisme sel, maka akan terjadi
kekeringan pada substrat. Hal ini dapat dilihat
pada suhu 31°C, dimana RH yang terukur adalah 65
persen. Pada keadaan ini laju alir udara yang
besar mengakibatkan laju penguapan air yang
tinggi pula, sehingga terjadi kekeringan sub-
penetrasi enzim ke dalam partikel-partikel sub-
strat, sehingga mengurangi aktivitas enzim dan
mengurangi kemudahan mencapai nutrien, karena
rendahnya pengembangan substrat dan mendorong
terjadinya sporulasi dini (Weiland, 1988).
Dari hasil uji lanjut yang dilakukan diper-
oleh aktivitas FP-ase dan CMC-ase tertinggi pada
suhu 28OC. Lama fermentasi terbaik diperoleh
pada hari kedelapan untuk FP-ase dan hari keenam
untuk CMC-ase (Lampiran 5b dan Lampiran 6b).
2. Pengaruh Aerasi Terhadap Aktivitas Enzim
Aerasi yang diberikan pada penelitian ini ada-
lah aliran udara lembab steril dengan laju 0.1, 0.3
dan 1 l/jam/g substrat serta tanpa aerasi sebagai
kontrol.
a. Aktivitas FP-ase
Aktivitas FP-ase tertinggi pada suhu 25OC
sebagaimana dapat dilihat pada Gambar 11. diper-
oleh pada laju aerasi 0.1 l/jam/g substrat (1.34
IU/ml)
.
Pada suhu 28°C (Gambar 2 . ) nilaiaktivitas FP-ase tertinggi (2.04 IU/ml) diperoleh
pada kondisi tanpa aerasi memberikan nilai ak-
tivitas yang rendah (0.98 IU/ml). Hal ini menun-
jukkan bahwa aerasi memang berpengaruh terhadap
aktivitas enzim yang dihasilkan. Laju aerasi
yang dibutuhkan untuk suhu 28OC (1 l/ jam/g sub-
strat) lebih tinggi daripada untuk suhu 25OC (0.1
l/jam/g substrat). Hal ini diduga karena pada
suhu 28OC panas metabolik yang dihasilkan lebih
Lama Fermentasi (hari)
-Tanpa Aerasi -+ Aerasi 0.1 l/jam/g
[image:54.514.122.481.302.616.2]++Aerasi 0.3 l/jarn/g + Aerasi 1 l/jarn/g
Lama Ferrnentasi (hari)
-Tanpa
Aerasi
-Aerasi
0.1l/jam/g
[image:55.521.128.476.49.438.2]"Aerasi
0.3l/jam/g "Aerasi
1i/jam/g
Gambar 12. Aktivitas FP-ase pada Suhu 2S°C dengan Berbagai Tingkat Aerasi
tinggi, o l e h s e b a b i t u d i p e r l u k a n l a j u a e r a s i y a n g lebih
tinggi untuk menghilangkan sebagian panas terse-
but.
Panas yang dihasilkan selama fermentasi
harus dihilangkan, oleh sebab itu pengaliran
udara lembab steril ke dalam fermentor diperlukan
untuk menjaga kondisi optimal. Jadi laju aerasi
1 l/jam/g substrat ini diduga mampu menghilangkan
Berbeda dengan kedua tingkat suhu di atas,
pada suhu 31°C (Gambar 13), nilai FP-ase terendah
diperoleh pada laju aerasi 1 l/jam/g substrat
(0.81 IU/ml), sedangkan nilai tertinggi diperoleh
pada aerasi 0.3 l/jam/g substrat (1.70 IU/ml).
Pada suhu 31°C, aerasi yang terlalu tinggi akan
menyebabkan kekeringan pada substrat, karena
kadar air yang menguap tidak dapat diimbangi oleh
kadar air yang disuplai ke media yang akhirnya
Lama Fermentasi (hari)
[image:56.518.106.488.62.655.2]1
-Tanpa Aerasi - Aerasi 0.1 M a i dakan menghambat pertumbuhan kapang dan produksi
enzim. Kondisi ini berhubungan dengan RH yang
terukur pada suhu ini (65 persen), sehingga laju
aerasi yang tinggi akan memperbesar laju pengua-
pan air dengan tingginya suhu yang diberikan.
Sama halnya dengan nilai aktivitas FP-ase,
pada suhu 25OC dan 28OC nilai aktivitas CMC-ase
(Gambar 14 dan 15.) tertinggi diperoleh berturut-
turut pada aerasi 0.1 l/ jam/g substrat (55.08
IU/ml) dan 1 l/jam/g substrat (69.26 IU/ml)
.
Sedangkan pada suhu 31°C (Gambar 16), nilai
tertinggi CMC-ase diperoleh pada aerasi 0.3
l/jam/g substrat (40.17 IU/ml). Perbedaan laju
aerasi optimal pada ketiga tingkat suhu tersebut
dapat dijelaskan sebagai berikut.
Tingkat pindah panas total diduga dibatasi
oleh tingkat pindah panas intrapartikel dan
interpartikel, tingkat dimana panas ditransfer
dari permukaan partikel ke Ease gas dan tingkat
dimana panas dalam fase gas dikeluarkan. Tingkat
pindah panas dari padatan dan fase gas dapat le-
bih ditingkatkan dengan mengalirkan gas dalam
Lama Fermentasi (hari)
-
Tanpa Aerasi-
Aerasi 0.1 l/jarn/g [image:58.514.114.462.54.452.2]*
Aerasi 0.3 l/jarn/g " Aerasi 1 l/jarn/gGambar 14. Aktivitas CMC-ase pada Suhu 25OC dengan Berbagai Tingkat Aerasi
Meskipun fungsi utama aerasi selama kultiva-
si padat aerobik adalah untuk mensuplai O2 untuk
pertumbuhan sel dan untuk menghilangkan C02, juga
membantu pindah panas dan lembab antara fase
padat dan gas (Prior et al., 1990).
Peningkatan laju aerasi dapat digunakan
untuk menurunkan suhu substrat (Prior et al.,
1990). Oleh sebab itu pada suhu 2 5 O C , pemberian
aerasi yang besar akan menurunkan suhu fermentasi
-- Tanpa Aerasi - Aerasi 0.1 l/jam/g
[image:59.514.132.473.75.459.2]*Aerasi 0.3 l/jam/g + Aerasi 1 l/jam/g
Gambar 15. Aktivitas CMC-ase pada Suhu 2a°C dengan Berbagai Tingkat Aerasi
suhu 28OC, dengan pemberian aerasi yang lebih
besar akan menghilangkan sebagian panas yang
dihasilkan, sehingga suhu fermentasi akan tetap
berada pada selang suhu optimum produksi enzim.
Peningkatan tingkat aerasi sampai 2 l/jam/g
pada suhu 28OC cenderung menurunkan aktivitas en-
zim (Gambar 17 dan 18). Laju aerasi yang terla-
lu tinggi juga menjadi masalah, jika lebih banyak
0 2 4 6 8 1 0 Lama Ferrnentasi (hari)
Gambar 16. Aktivitas CMC-ase pada Suhu 31° dengan Berbagai Tingkat Aerasi
-
Tanpa Aerasi - Aerasi 0.1 l/jam/g XAerasi 0.3 l/jam/g + Aerasi 1 i/jam/gmedium. Hal ini dapat menyebabkan kekurangan
air untuk pertumbuhan mikrobial yang sering
menghambat induksi, tetapi tidak menghalangi
sporulasi jamur (Weiland, 1988). Oleh sebab itu
pada suhu 31°C berdasarkan hasil penelitian ini,
laju aerasi yang tinggi cenderung menurunkan
aktivitas enzim. Dari hasil uji lanjut yang
dilakukan diperoleh aktivitas tertinggi pada laju
kondisi tanpa aerasi memberikan nilai alctivitas
terendah (Lampiran 5b dan 6b).
Hal ini juga sesuai dengan penelitian Abdul-
lah et a l . (1985) yang menggunakan lima tingkat aerasi (0, 0.06, 0.12, 0.24. 1.2 l/jam/g sub-
strat) dan suhu (30°C, 35OC, 37OC dan 40°C) pada
optimasi fermentasi jerami gandum menggunakan
Chaetomium c e l l u l o l y t i c u m menghasilkan kondisi terbaik pada laju aerasi 0.12 l/jam/g substrat
Lama Ferrnentasi (hari)
-
T a n p a Aerasi
-
Aerasi
0.1Iljamlg
+ A e r a s i
0.3l/jam/g + A e r a s i 1 l/jam/g
[image:61.518.119.481.120.650.2]*
Aerasi 2 l/jam/g
Lama F e r m e n t a s i ( h a r i )
-
Tanpa Aerasi - Aerasi 0.1 l/jam/g++ Aerasi 0.3 l/jam/g + Aerasi 1 l/jam/g
[image:62.521.120.460.68.423.2]*Aerasi 2 l/jam/g
Gambar 18. Aktivitas CMC-ase pada Suhu 28OC dengan Aerasi sampai 2 l/jam/g
dan hasil terendah diperoleh pada laju aerasi 1.2
l/jam/g substrat. Artinya bahwa pada suhu ting-
gi, dengan laju aerasi yang tinggi memberikan
hasil yang rendah pula.
Pada penelitian ini digunakan tandan kosong
10 mesh, sehingga diharapkan ruang kosong antar
partikel akan meningkat. Jika ruang kosong cukup
tinggi, pengadukan kontinyu dan aerasi tidak
tinggi, pengadukan kontinyu dan aerasi tidak
selalu dibutuhkan, karena ruang kosong mungkin
mengandung O2 yang cukup untuk metabolisme per-
tumbuhan sel. Namun, untuk melepaskan C02 yang
terperangkap tetap dibutuhkan aerasi atau penga-
dukan substrat (Weiland, 1988).
Secara keseluruhan dapat dilihat bahwa akti-
vitas enzim pada awal fermentasi sangat rendah,
kemudian meningkat antara hari keempat sampai
hari kedelapan untuk selanjutnya menurun dengan
semakin lamanya waktu fermentasi dan akhirnya
mengalami kematian. Hal ini dapat dimengerti
karena pada awal fermentasi, kapang masih berada
pada masa adaptasi, kemudian berada pada fase
logaritmik, untuk kemudian mengalami fase sta-
sioner dimana sudah banyak kapang yang mati.
Aktivitas CMC-ase tertinggi diperoleh pada
hari keenam fermentasi, sedangkan aktivitas FP-
ase tertinggi diperoleh pada hari kedelapan fer-
mentasi
.
Hal ini disebabkan karena CMC-aselebih dulu bekerja dengan menyerang bagian amorf
dari selulosa, sedangkan FP-ase membutuhkan
waktu yang lebih lama untuk menyerang struktur
kristal dari selulosa (Enari, 1983).
Berdasarkan analisis statistik, interaksi
terendah pada suhu 31°C dengan aerasi 1 l/ jam/g
(1.43 IU/ml)
.
Sedangkan CMC-ase tertinggidiperoleh pada suhu 2S°C dengan tingkat aerasi I
l/jam/g (69.26 IU/ml) dan nilai terendah pada
suhu 31°C dan aerasi 0.3 l/jam/ (40.17 IU/ml).
Nilai aktivitas FP-ase tertinggi dari semua
perlakuan adalah 2.042 IU/ml, sedangkan CMC-ase
sebesar 69.26 IU/ml yang diperoleh pada suhu 28OC
V. KESIMPULAN DAN SARAN
A. KESIIMPULAN
Tandan kosong dan sabut kelapa sawit merupakan
substrat yang potensial pada produksi selulase karena
mengandung holoselulosa dan lignin yang cukup tinggi
yaitu sebesar 65.28 persen dan 35.2 persen untuk tandan
kosong dan 55 persen dan 21.92 persen untuk sabut
kelapa sawit.
Suhu mempengaruhi aktivitas selulase yang dihasil-
kan. Suhu yang terlalu rendah dan melebihi suhu opti-
mal pertumbuhan kapang dan pembentukan produk cenderung
menurunkan aktivitas enzim. Suhu optimal yang dipero-
leh pada penelitian ini adalah 2SoC.
Aerasi yang diberikan selama proses fermentasi
membantu mempertahankan kondisi aerobik yang dibutuhkan
kapang selama pertumbuhannya dan membantu menurunkan
panas yang dihasilkan selama proses fermentasi, sehing-
ga dapat menghasilkan enzim dengan aktivitas yang
tinggi. Laju aerasi yang diberikan pada penelitian ini
berbeda-beda tergantung pada suhu proses fermentasi.
Dari penelitian ini diperoleh aktivitas tertinggi pada
laju aerasi 1 l/jam/g substrat dan suhu 2S°C.
FP-ase membutuhkan waktu yang lebih lama diban-
FP-ase tertinggi diperoleh pada hari kedelapan fermen-
tasi, sedangkan CMC-ase pada hari keenam.
Secara statistik, faktor suhu, aerasi dan lama
Lama fermentasi terbaik untuk memproduksi CMC-ase
adalah 6 hari dan 8 hari untuk memproduksi FP-ase.
Perlakuan yang menghasilkan aktivitas FP-ase dan CMC-
ase tertinggi adalah suhu 28°C dan tingkat aerasi 1
l/jam/g yaitu masing-masing sebesar 2.04 IU/ml dan
69.26 IU/ml.
B. SARAN
Pada penelitian ini belum semua faktor lingkungan
fermentasi diamati, sehingga perlu dilakukan penelitian
mengenai ketebalan substrat maupun pH selama fermenta-
si. penelitian ini masih dilakukan pada skala labora-
torium jadi perlu dilakukan penelitian dalam skala yang
lebih besar.
Selain itu juga diperlukan penelitian mengenai op-
timasi proses hilir untuk meningkatkan rendemen enzim
yang dihasilkan dengan aktivitas yang tinggi pula.
Aplikasi penggunaan selulase yang dihasilkan untuk
menghidrolisis limbah-limbah pertanian dapat dilakukan
pada penelitian selanjutnya, disamping penelitian untuk
memanfaatkan biomassa yang dihasilkan sebagai pakan
DAFTAR PUSTAKA
AOAC. 1980. Official Method of Analytic of Association of Official Analytic Chemist. Association of Offi- cial Analytic Chemist, Washington DC.
Achmadi, S. 1989. Kimia kayu. Diktat PAU Ilmu Hayati. Institut Pertanian Bogor, Bogor.
Aidoo, K.E., R. Hendry dan B.J.B. Wood. 1982. Solid Substrat Fermentation. Di dalam Advances in Applied Microbiology. Academic Press, Inc. 28 : 201-203.
Alexopoulus, G.J. dan C.W. Mims. 1979. Introductory 8Mycology. John Wiley and Sons, New York.
Banerjee, U.C., Y. Chisti dan M. Moo-young. 1994. Pro- tein Enrichment of Corn Stover Using Neurospora sitophila. Simp. Bioproducts Processing. University of Waterloo, Canada.
Chahal, D.S. 1985. Solid State Fermentation with Tricho- derma reesei for Cellulase Production. Appl. Envi- ron. Microbiol. 49(1) : 205-210.
Chang, M. M, T.Y.C. Chon dan G.t. Tsao. 1981. Structure Pretreatment and Hidrolysis Cellulose. Adv. Bio- chem. Eng. Vol 20.
Darnoko. 1992. Potensi Pemanfaatan Limbah Lignoselulosa Kelapa Sawit melalui Biokonversi. Berita Pen. Perke- bunan, Medan
.
Darwis, A.A. dan E. Sukara. 1990. Isolasi, Purifikasi dan Karakterisasi Enzim. PAU IPB, Bogor.
Enari, T.M. 1983. Microbial Cellulases. Di dalam W.M. Fogarty (ed.) Microbial Enzymes and Biotechnology. Appl. Science, London.
Frost, G.M. dan D.A. Moss. 1987. Production of Enzymes by Fermentation. Di dalam Biotechnology Vol. 7a. VHC
,
Germany.Goering, H.K., Van Soest, P.J. 1970. Forage Analysis Agriculture Handbook No. 379 Ag. Res. Service, USDA.
Annual Report on Fermentation Process. Academic Press, New York.
Ikatrinasari, Z.F. 1995. P e n g g u n a a n N i t r o g e n d a n M i n e r a l M i k r o
dalam Produksi Selulase pada Medium Limbah Padat Kelapa Sawit oleh Neurospora sitophila. Skripsi. FATETA IPB, Bogor.
IRRI. 1972. Laboratory Manual for Physiologycal Studies of Rice the Internationa1,Rice. Research Institute
Losbanos, Philiphines.
Judoamidjojo, R.M., E. Gumbira Sa'id dan L. Hartoto. 1989. Biokonversi. PAU Bioteknologi IPB, Bogor.
Lonsane, B.K., N.P. Ghildyal, S. Budiatman dan S.V. Ramak- rishna. 1985. Engineering Aspects of solid State Fermentation. J. Enzyme Microb. Tech, 7:258
-
265.Maggy-T. Suhartono. 1989. Enzim dan Bioteknologi. PAU IPB, Bogor.
Mandels, M. and E.T. Reese. 1957. Induction of Cellulase in Trichoderma viride as Influenced by Carbon Sources and Metals. J. Bacteriol. 73:269.
Mandels, M. 1982. Cellulases. Di dalam Tsao, G. Annual Report on Fermentation Processes. Vol. 5. Academic Press, New York.
Miller, G.L. 1959. The Use of Dinitrosalicylic Acid Reagent for Determination of Reducing Sugar. Anal. Chem. 31 (3) : 426-428.
Oguntimein, G.O. Vlach dan M. Moo-young. 1991. Produc- tion of Cellulolytic Enzymes by Neurospora sitophila Grown on Cellulosic Material. Bioresourch. Tech. 39
: 277-283
Prior, B.A., J.C. Du Preez dan P.W. Rein. 1990. Environ- mental Parameters. Di dalam. W.