INTERAKSI ANTARA BIAK SUSPENSI SEL GAHARU
(
Aquilaria malaccensis
Lam.) DAN
Fusarium
sp
.
DALAM
MENGHASILKAN SENYAWA SESKUITERPENA
ARYANI LEKSONOWATI
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa tesis berjudul “Interaksi antara Biak Suspensi Sel Gaharu (Aquilaria malaccensis Lam.) dan Fusarium sp. dalam Menghasilkan Senyawa Seskuiterpena” adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
RINGKASAN
ARYANI LEKSONOWATI. Interaksi antara Biak Suspensi Sel Gaharu (Aquilaria malaccensis Lam.) dan Fusarium sp. dalam Menghasilkan Senyawa Seskuiterpena. Dibimbing oleh DIAH RATNADEWI, WITJAKSONO dan ANDRIA AGUSTA.
Aquilaria malaccensis Lam. merupakan tumbuhan penghasil resin wangi atau gubal gaharu yang dimanfaatkan secara luas untuk industri parfum, dan obat-obatan. Resin wangi tersebut mengandung metabolit sekunder berupa seskuiterpena dan kromona. Gubal gaharu terbentuk sebagai respon dari masuknya patogen ke dalam jaringan yang terluka baik secara alami maupun disengaja. Tingginya permintaan pasar dan perburuan liar menyebabkan masalah serius dalam penurunan populasi spesies Aquilaria di alam, sehingga pada tahun 1995, CITES (Convention on International Trade in Endangered Species of Wild Fauna and Flora) menetapkan A. malaccensis masuk ke dalam Appendix II. Untuk mengatasi masalah tersebut dilakukan budi daya dan inokulasi buatan. Tetapi cara ini memiliki kelemahan karena memerlukan tanaman dalam jumlah besar, waktu yang lama dan konsistensi keberhasilan yang rendah. Kultur suspensi sel gaharu telah dipikirkan untuk menjadi salah satu solusi untuk memproduksi resin gaharu. Apabila berhasil, maka dapat menghemat lahan tanam, lebih cepat dan efisien. Penelitian ini bertujuan untuk mendapatkan metode inisiasi dan perbanyakan kalus remah serta mempelajari interaksi sel gaharu A. malaccensis dengan Fusarium sp. sebagai elisitor pada pembentukan senyawa seskuiterpena dalam sistem suspensi sel, khususnya interaksinya dengan Fusarium sp. strain 372.3 dan w1fus.sp.2.
Tunas in vitro berumur 2 bulan dari spesies gaharu A. malaccensis serta Fusarium sp. strain 372.3 dan strain w1fus.sp.2. digunakan sebagai bahan dalam penelitian ini. Tahapan penelitian meliputi inisiasi biak kalus gaharu, inisiasi dan pemeliharaan suspensi sel gaharu, perlakuan interaksi suspensi gaharu dan Fusarium sp. untuk menginduksi senyawa seskuiterpena.
Hasil percobaan menunjukkan bahwa kalus dapat diinduksi dari semua perlakuan auksin-sitokinin pada inokulum potongan daun dan ruas batang A. malaccensis. Kombinasi 1–2 mg/L 2,4-D dan BA dalam selang konsentrasi rendah (0.1–0.3 mg/L) dapat menginduksi pembentukan kalus yang tinggi (>80%) dan remah. Subkultur kalus ke medium dengan 0.5–1.0 mg/L 2,4-D mampu menghasilkan suspensi sel dengan struktur yang remah. Terbentuknya senyawa seskuiterpena pada sistem suspensi gaharu dipengaruhi faktor bentuk elisitor
cendawan, strain cendawan, umur suspensi dan lamanya waktu co-culture.
Interaksi 10% filtrat strain Fusarium sp. strain 372.3 dan suspensi gaharu umur 1 minggu dengan lama co-culture 1 bulan menginduksi terbentuknya senyawa seskuiterpena terbanyak dan spesifik pada gaharu di antaranya -eudesmol, elemol, dan -eudesmol. Perlakuan 10 dan 20% filtrat Fusarium sp. strain 372.3 dan w1fus.sp.2 dapat menghambat pertumbuhan sel, namun mampu menginduksi terbentuknya seskuiterpena lainnya yang spesifik terdapat pada gaharu yaitu dihidro--agarofuran, dan kariofilena oksida.
SUMMARY
ARYANI LEKSONOWATI. Interaction Between Cell Suspension Culture of Aquilaria malaccensis Lam. and Fusarium sp. in Producing Sesquiterpenoid Compounds. Supervised by DIAH RATNADEWI, WITJAKSONO dan ANDRIA AGUSTA.
Aquilaria malaccensis Lam. is a plant species producing fragrant woody material called “agarwood” commonly used as medicine and perfume. The fragrant compounds such as sesquiterpenoids and chromone derivatives are the major chemical components of agarwood. Agarwood is formed in response to pathogen infection through wounding occurred on stem, both naturally and artificially. High demand leading to illeggal logging has seriously affected in the decrease of Aquilaria species population in its natural habitat. A. malaccensis has been listed in Appendix II of the Convention on International Trade in Endangered Species of Wild Fauna and Flora (CITES) since 1995. Under these circumstances, conventional cultivation of agarwood trees and attempts to artificially induce agarwood on those trees are perfomed. But this method has some weakness since it requires cultivation on a large scale, long time and low consistency of success. Cell suspension culture may become an alternative method to produce the desired substances. Cell suspension culture is expected to be more effective because it can minimize the space, be more quickly and be more efficient. The aims of this research were to find a method of initiation and proliferation of friable calli; and to study interaction between A. malaccensis cells and Fusarium sp. as elicitors that influence sesquiterpenoids production in cell suspension culture, especially Fusarium sp. strain 372 and w1 fus. sp.2
Two-month old A. malaccensis planlet and Fusarium sp. strain 372 and w1 fus. sp.2 were used as materials in this research. This study was conducted in several steps, e.g. initiation of callus, initiation and maintenance of cell suspension culture, and treatment of A. malaccensis cell suspension culture with Fusarium sp. to induce sesquiterpenoid compounds.
The results showed that calli were established in almost all treatments of auxin-cytokinin on both leaf and internode explants. The combined treatment of 1–2 mg/L 2,4-D and 0.2–0.3 mg/L BA induced friable callus formation in more than 80% of cultures. Subculture of calli in medium with 0.5–1 mg/L 2,4-D induced friable calli. In the suspension cell system of A. malaccensis, some factors could affected sesquiterpenoid formation, such as types of elicitor, clones of fungi, the age of the suspension culture and the time course of co-culture. Most sesquiterpenoids were detected from one-week cell suspension culture which contained 10% filtrate of Fusarium sp. strain 372.3 and were harvested after one month of co-culture, such as -eudesmol, elemol, and -eudesmol. The treatments of 10 and 20% filtrate of Fusarium sp. strain 372.3 and w1fus.sp.2 inhibited the cell growth, but induced several specific agarwood compounds of sesquiterpenoid including dihydro--agarofuran, and caryophyllene oxide.
© Hak Cipta Milik IPB, tahun 2016
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Biologi Tumbuhan
INTERAKSI ANTARA BIAK SUSPENSI SEL GAHARU
(
Aquilaria malaccensis
Lam.) DAN
Fusarium
sp. DALAM
MENGHASILKAN SENYAWA SESKUITERPENA
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2016
Judul Tesis : Interaksi antara Biak Suspensi Sel Gaharu (Aquilaria malaccensis Lam.) dan Fusarium sp. dalam Menghasilkan Senyawa Seskuiterpena
Nama : Aryani Leksonowati NIM : G353120271
Disetujui oleh Komisi Pembimbing
Prof Dr Ir YM Diah Ratnadewi, DEA Ketua
Dr Ir Witjaksono, MSc Anggota
Dr Andria Agusta, MSc Anggota
Diketahui oleh
Ketua Program Studi Biologi Tumbuhan
Dr Ir Miftahudin, MSi
Dekan Sekolah Pascasarjana
Dr Ir Dahrul Syah, MScAgr
PRAKATA
Puji dan syukur penulis panjatkan ke hadirat Allah SWT atas segala karunia dan rahmatNya sehingga karya ilmiah yang berjudul “Interaksi antara Biak Suspensi Sel Gaharu (Aquilaria malaccensis Lam.) dan Fusarium sp. dalam Menghasilkan Senyawa Seskuiterpena” ini berhasil diselesaikan.
Penulis mengucapkan terima kasih kepada Prof Dr Ir YM Diah Ratnadewi, DEA, Dr Ir Witjaksono, MSc dan Dr Andria Agusta, MSc selaku pembimbing, serta Dr Ir Gayuh Rahayu selaku penguji luar komisi. Di samping itu, penulis ucapkan terima kasih kepada teman-teman di Laboratorium Biak Sel dan Jaringan Tumbuhan, Laboratorium Kimia Bahan Alam, Bidang Botani serta Laboratorium Kimia Analitik, InaCC, Pusat Penelitian Biologi LIPI, yang telah banyak membantu terlaksananya penelitian ini. Terima kasih kepada Kemenristek yang telah mendanai studi melalui program beasiswa Kemenristek tahun 2012-2014. Ungkapan terima kasih juga disampaikan kepada adik, saudara dan kerabat atas segala doa, serta seluruh pihak yang telah membantu yang tidak bisa disebutkan satu persatu.
Semoga karya ilmiah ini bermanfaat.
DAFTAR ISI
DAFTAR TABEL xii
DAFTAR GAMBAR xii
DAFTAR LAMPIRAN xii
1 PENDAHULUAN 1
Latar Belakang 1
Tujuan Penelitian 2
Manfaat Penelitian 2
2 TINJAUAN PUSTAKA 2
Tumbuhan Gaharu 2
Pembentukan Senyawa Aromatik pada Gaharu 3
Produksi Metabolit Sekunder pada Gaharu Secara In Vitro 4
Jalur Biosintesis Senyawa Seskuiterpena 4
3 METODE 7
Waktu dan Tempat Penelitian 7
Bahan Penelitian 7
Prosedur Penelitian 8
Inisiasi Biak Kalus Gaharu 8
Inisiasi dan Pemeliharaan Biak Suspensi Sel Gaharu 10 Perlakuan Interaksi Suspensi Sel Gaharu dan Cendawan Fusarium sp.
untuk Induksi Pembentukan Seskuiterpena 10
Pemanenan dan Ekstraksi Seskuiterpena dari Suspensi Sel 11
Analisis Seskuiterpena dengan GC-MS 12
4 HASIL DAN PEMBAHASAN 12
Hasil 12
Pembahasan 26
5 SIMPULAN DAN SARAN 31
Simpulan 31
Saran 31
DAFTAR PUSTAKA 32
LAMPIRAN 37
DAFTAR TABEL
1 Perlakuan ZPT tunggal auksin serta kombinasi auksin-auksin dan auksin-sitokinin untuk induksi kalus A. malaccensis 9 2 Perlakuan interaksi suspensi sel gaharu dan cendawan Fusarium sp.
untuk induksi pembentukan seskuiterpena 20
3 Perbandingan jumlah senyawa seskuiterpena pada perlakuan interaksi, kontrol suspensi sel gaharu dan kayu A. malaccensis terinfeksi 22 4 Rata-rata jumlah ekstrak yang diperoleh dan jumlah seskuiterpena
terdeteksi pada perlakuan 10% dan 20% filtrat Fusarium sp. strain
372.3 dan w1fus.sp.2 24
DAFTAR GAMBAR
1 Jalur biosintesis senyawa seskuiterpena 6
2 Bagan alur penelitian dari induksi kalus remah gaharu hingga induksi
seskuiterpena 8
3 Perbedaan morfologi kalus dari eksplan potongan daun dan ruas batang
A. malaccensis 13
4 Pengaruh jenis dan konsentrasi auksin terhadap induksi kalus dari
eksplan potongan daun A. malaccensis. 14
5 Pengaruh konsentrasi auksin 2,4-D terhadap induksi kalus dari eksplan
ruas batang A. malaccensis. 14
6 Pengaruh kombinasi jenis dan konsentrasi auksin dan auksin-sitokinin terhadap kalus dari eksplan potongan daun A. malaccensis 16 7 Pengaruh kombinasi konsentrasi 2,4-D dan BA terhadap induksi kalus
dari eksplan ruas batang A. malaccensis 17
8 Morfologi kalus gaharu dalam medium cair 17
9 Kurva pertumbuhan sel gaharu 18
10 Kenampakan fisik kultur hasil interaksi suspensi sel gaharu umur 1 minggu dengan filtrat Fusarium sp. strain 372.3. 19
11 Kromatogram GC-MS dan struktur molekul 21
12 Pengaruh konsentrasi filtrat cendawan Fusarium sp. strain 372.3 dan w1fus.sp.2 pada umur suspensi sel A. malaccensis 1 minggu terhadap pertumbuhan suspensi sel selama masa co-culture 4 minggu 23 13 Konsentrasi relatif beberapa seskuiterpena terdeteksi per 25 mL
suspensi terhadap senyawa standar -eudesmol pada interaksi filtrat cendawan Fusarium sp. strain 372.3 dan w1fus.sp.2 dan suspensi sel A.
malaccensis 25
DAFTAR LAMPIRAN
1 Komposisi medium MS (Murashige & Skoog 1962)
38
2 Komposisi medium PDA (Potato Dextrose Agar) 39
4 Morfologi kalus yang terbentuk dari potongan daun dan ruas batang pada perlakuan kombinasi berbagai jenis dan konsentrasi auksin-auksin
dan auksin-sitokinin 41
5 Senyawa seskuiterpena yang terdeteksi GC-MS pada interaksi suspensi sel A. malaccensis dan cendawan Fusarium sp. strain 372.3 dan w1fus.sp.2 dengan variasi umur suspensi sel, lama co-culture dan jenis
elisitor 42
6 Kurva kalibrasi senyawa standar -eudesmol pada beberapa konsentrasi 46 7 Perhitungan kuantitatif beberapa senyawa seskuiterpena terdeteksi pada
perlakuan 10% filtrat Fusarium sp. strain 372.3, kontrol suspensi sel
dan kayu A. malaccensis terinfeksi 47
8 Kromatogram GC-MS pada perlakuan 10% dan 20% filtrat Fusarium
sp. strain 372.3 dan w1fus.sp.2 48
1
PENDAHULUAN
Latar Belakang
Tumbuhan gaharu Aquilaria malaccensis Lam. merupakan salah satu penghasil gubal gaharu yang dimanfaatkan secara luas untuk keperluan industri parfum, kosmetik, dan obat-obatan (Akter et al. 2013; Subasinghe et al. 2012). Masalah yang saat ini dihadapi adalah populasi tanaman gaharu yang semakin berkurang di alam. Hal ini disebabkan para pemburu gaharu tidak dapat mengenali dengan tepat mana tanaman yang sudah mengandung gubal gaharu dan tidak, sehingga mereka sering menebang pohon yang salah (Suhartono & Newton 2001). Sejak tahun 1995, tanaman gaharu A. malaccensis dimasukkan dalam daftar Appendix II CITES (Convention on International Trade in Endangered Species of Wild Fauna and Flora) sehingga perdagangannya dibatasi dengan kuota tertentu agar kepunahan spesies tanaman ini dapat dicegah (Barden et al. 2000). Kuota perdagangan direkomendasikan oleh Scientific Authority (LIPI), disahkan oleh Management Authority (Kementerian Lingkungan Hidup Kehutanan).
Gubal gaharu dihasilkan tanaman sebagai respon dari masuknya patogen ke dalam jaringan yang terluka. Luka pada tanaman berkayu dapat disebabkan secara alami, maupun secara sengaja (Ng et al. 1997). Gubal gaharu mengandung senyawa berupa resin berwarna coklat dan beraroma harum (Ng et al. 1997). Senyawa utama yang diketahui menyebabkan aroma wangi gaharu adalah seskuiterpena (golongan terpenoid) dan turunan feniletil kromona. Cendawan merupakan salah satu faktor penginduksi gubal gaharu, dan kelompok Fusarium merupakan jenis yang relatif sering ditemukan pada batang Aquilaria spp. (Isnaini et al. 2009; Budi et al. 2010; Nurbaya et al. 2014). Beberapa senyawa kimia juga dilaporkan dapat menjadi sinyal sekunder untuk menginduksi senyawa wangi gaharu seperti metil jasmonat dan asam salisilat (Nobuchi & Siripatanadilok 1991; Ito 2005; Okudera & Ito 2009).
Alternatif solusi untuk mengatasi masalah populasi gaharu di alam, masyarakat melakukan budi daya gaharu secara konvensional dan inokulasi buatan dengan cara mengebor batang tanaman penghasil gaharu untuk memasukkan inokulum patogenik ke dalamnya. Dari infeksi tersebut, akan terbentuk gubal yang mengandung resin wangi dan terakumulasi pada jaringan kayu. Senyawa wangi yang dihasilkan tersebut dapat diperoleh dengan mengekstraksi gubal gaharu secara langsung (Azah et al. 2008). Tetapi cara ini memerlukan budi daya tanaman dalam skala besar dan waktu yang lama serta konsistensi keberhasilan yang rendah. Cara lain yang dapat dipikirkan untuk diupayakan menjadi penghasil resin gaharu melalui metode kultur jaringan, seperti kultur kalus atau suspensi sel. Cara ini lebih efisien karena dapat menghemat luasan lahan tanam dan lebih cepat.
penyakit pada tumbuhan seperti busuk pangkal batang, tumor akar (root crown), penyakit pembuluh xilem, dan penyakit pasca panen (Wang & Jeffer 2000; Ploetz 2005). Melalui kultur suspensi sel, penggunaan Fusarium sp. sebagai elisitor diharapkan bisa lebih terkendali dan aman bagi lingkungan.
Tujuan Penelitian
Penelitian ini bertujuan untuk :
1. Mempelajari komposisi zat pengatur tumbuh untuk menginisiasi dan memelihara kalus remah A. malaccensis,
2. Mempelajari interaksi sel gaharu A. malaccensis dengan Fusarium sp., khususnya strain 372.3 dan strain w1fus.sp.2. sebagai elisitor pada pembentukan senyawa seskuiterpena dalam sistem suspensi sel.
Manfaat Penelitian
Manfaat dari penelitian ini adalah dengan mengetahui faktor-faktor yang berpengaruh dalam interaksi sel gaharu A. malaccensis dengan Fusarium sp. diharapkan manipulasi faktor tersebut dapat diterapkan untuk produksi resin gaharu di laboratorium. Diharapkan pula di masa yang akan datang dapat diperoleh teknologi ramah lingkungan terkait interaksi antara sel tumbuhan dan patogen dalam produksi senyawa seskuiterpena yang menjadi komponen resin gaharu, dan keberhasilan produksi resin in vitro diharapkan akan bermanfaat untuk mengatasi masalah penurunan populasi gaharu di alam akibat perburuan dan penebangan liar gaharu di hutan yang menggiring spesies tanaman ini menuju kepunahan.
2
TINJAUAN PUSTAKA
Tumbuhan Gaharu
Gaharu merupakan kayu yang mengandung resin berwarna gelap dan terbentuk sebagai akibat adanya infeksi patogen. Penghasil utama resin gaharu berasal dari genus Aquilaria dan Gyrinops, famili Thymelaeaceae. Gaharu memiliki beberapa nama umum/daerah seperti agarwood (Eropa), Malayan eagle wood, Malayan aloe wood (Inggris); calambac, calambour (Perancis); gaharu (Indonesia dan Malaysia); karas (Sunda); kekeras; kepang (Malaysia); oud (arab); agar (India); agaru (Tibet); dan jinkoh (Jepang) (PROSEA 1999).
ditemukan di Sumatera (Sibolangit, Bangka, Jambi, Riau, Sumatera Selatan), 2011). Pohon ini mulai memproduksi bunga dan biji sejak empat tahun (Akter et al. 2013).
Tumbuhan A. malaccensis termasuk pohon yang tumbuh cepat, dengan tinggi mencapai 20–40 m dan diameter 0,6–2,5 m. Tipe daunnya tunggal, berseling, elips oblong hingga lanset oblong, ukuran 7.5–12 cm x 2.5–5.5 cm, tangkai daun sepanjang 4–6 mm. Bunga tumbuh di ketiak daun, berbentuk payung, bercabang, bunga berkelopak tabung dengan ukuran 5–6 mm, dan berwarna hijau atau kuning tua (Suhartono & Newton 2001). Buah bulat telur dengan bagian basal lebih lonjong, ukuran 23 cm, daging buah tebal, jumlah biji 1–2 buah (PROSEA 1999).
Gaharu mengandung essens yang disebut sebagai minyak essens (essential oil) yang dibuat dengan ekstraksi atau penyulingan dari gubal gaharu. Minyak gaharu digunakan sebagai bahan pembuatan parfum, sedangkan serbuk atau abu gaharu digunakan sebagai bahan pembuatan dupa, aromaterapi, serta untuk keperluan upacara adat dan agama di India dan kawasan Asia Tenggara. Gaharu telah digunakan untuk tujuan medis hampir selama ribuan tahun di Ayuverda, Tibet, dan Asia Timur. Pengobatan tradisional Asia Timur menggunakannya sebagai obat rematik, obat gosok, obat perangsang, dan juga digunakan dalam produksi obat-obatan asma (Barden et al. 2000).
Gaharu termasuk salah satu komoditas hutan bukan kayu (HHBK) yang bernilai tinggi dan banyak diburu untuk diambil gubalnya. Kayu yang belum terisi resin bersifat lunak, ringan, pucat, mudah rusak, sedangkan kayu yang sudah berisi resin berwarna gelap, berat dan keras. Badan Standardisasi Nasional atau BSN (2011) membedakan kualitas gaharu menjadi tiga tingkatan yaitu gubal, kamedangan dan serbuk gaharu berdasarkan warna, bobot dan aromanya. Gubal gaharu dibagi menjadi double super, super A, super B, super tanggung A, dan super tanggung B. Kemedangan dibagi menjadi sabah, kemedangan A, kemedangan B, tanggung C, kemedangan hijau, dan kemedangan putih.
Pembentukan Senyawa Aromatik pada Gaharu
Mekanisme terbentuknya gubal gaharu secara fisiologis dimulai dari masuknya mikroorganisme patogenik ke dalam jaringan kayu. Kemudian cendawan menyerang dan masuk sampai ke pembuluh batang. Infeksi cendawan ke dalam jaringan tanaman dianggap sebagai benda asing sehingga sel tumbuhan akan menghasilkan senyawa fitoaleksin yang berfungsi sebagai pertahanan terhadap infeksi patogen (Isa 2008). Awalnya, dampak infeksi dapat terlihat dengan adanya lapisan berwarna kecoklatan pada jaringan kayu. Infeksi cendawan ini kemudian perlahan menyebar dan membentuk lapisan-lapisan pada batang yang disebut gubal. Gubal yang terbentuk juga semakin banyak dengan menyebarnya infeksi. Gubal ini mengandung resin yang wangi dan terakumulasi pada jaringan kayu. Akibat akumulasi resin, serat-serat kayu lebih padat dan warna serat berubah dari putih menjadi kecoklatan, kemudian kehitaman seiring dengan meningkatnya konsentrasi resin tersebut (Rahayu & Situmorang 2006).
Ada banyak sekali jenis cendawan yang menginfeksi gaharu. Ng et al. (1997) melaporkan cendawan yang mungkin menginfeksi dalam pembentukan gaharu adalah Epicoccum granulatum atau Cytosphaera mangiferae. Sementara itu, Budi et al. (2010) telah mengumpulkan sejumlah isolat yang memiliki potensi untuk menginduksi gaharu pada sampel kayu gaharu dari berbagai daerah seperti Bangka, Bogor, Bohorok, Gorontalo, Kalimantan Timur, Lampung, Mentawai, Papua, dan Sukabumi. Hasil identifikasi koleksi isolat tersebut didapat spesies antara lain Fusarium solani (Mart.), Appell dan Walenw, F. tricinctum (corda) Sacc, F. sambucinum Fuckel, dan F. moniliformae. Mohamed et al. (2010) melaporkan bahwa isolat cendawan yang dikumpulkan dari pohon gaharu yang berasal dari hutan alami di Malaysia Barat termasuk ke dalam kelompok Cunninghamella, Curvularia, Fusarium dan Trichoderma. Nagajothiet al. (2016) juga mengumpulkan 17 kultur cendawan dari pohon gaharu yang berasal dari berbagai tempat di India dan setelah diidentifikasi cendawan tersebut termasuk ke dalam kelompok Aspergillus, Penicillium, Fusarium, Lasiodiploidia dan Chaetomium.
Dari populasi spesies gaharu di alam, hanya 10% yang kemungkinan dapat terinfeksi jamur dan menghasilkan gubal gaharu secara alami (umur pohon di atas 20 tahun, dan diameter kayu 40 cm). Secara tradisional, masyarakat hanya memanen pohon-pohon terinfeksi tanpa menanamnya kembali. Namun banyaknya permintaan telah menyebabkan kelangkaan spesies akibat pemanenan pohon baik yang sakit maupun sehat secara berlebihan (Isa 2008). Oleh karena itu, masyarakat mulai membudidayakan gaharu dan melakukan inokulasi buatan untuk memperoleh gubal yang berisi resin wangi.
Produksi Metabolit Sekunder Pada Gaharu secara In Vitro
Aquilaria malaccensis (Ama 7, Ama 13), dan A. microcarpa (Ami 2064) menunjukkan adanya interaksi dengan cendawan Acremonium dalam hal pembentukan senyawa wangi gaharu pada media miskin nutrisi dan pH berbeda (Isnaini et al. 2009). Kultur kalus gaharu yang ditumbuhkan pada media semipadat diduga juga menghasilkan senyawa metabolit sekunder jika diinduksi cendawan patogen penginduksi resin, seperti Fusarium sp.
Kultur suspensi sel tanaman sangat berguna untuk mempelajari biosintesis dari metabolit sekunder. Menurut Ito et al. (2005), penambahan senyawa metil jasmonat (MeJA) pada kultur suspensi Aquilaria sinensis dapat menginduksi terbentuknya senyawa wangi setelah 7 hari inkubasi. Berdasarkan analisis GC-MS, teridentifikasi beberapa senyawa seskuiterpena seperti -guaiena, -guaiena dan -humulena. Komposisi senyawa seskuiterpena dapat berubah berdasarkan lamanya waktu inkubasi. Suspensi sel yang dipanen setelah 36 jam dan 7 hari pemberian MeJA, kemudian diekstrak selnya dan dianalisis dengan GC-MS menunjukkan rasio -guaiena, -guaiena dan -humulena yang berbeda, dimana senyawa -humulena lebih dominan pada masa inkubasi 36 jam.
Qi et al. (2005) mempelajari bahwa 2-(2-feniletil) kromona merupakan komponen utama yang bertanggung jawab pada kualitas gaharu. Percobaan dilakukan menggunakan ekstrak kasar Melanotus flavolivens dengan pelarut yang berbeda untuk memperoleh produksi 2-(2-feniletil) kromona dalam kultur suspensi sel A. sinensis. Dari analisis LC-MS dapat dideteksi empat macam 2-(2-feniletil) kromona yaitu 2-(2-feniletil) kromona (1), 6,7-dimetoksi-2-[2-(4′-metoksifenil)etil] kromona (2), 6-metoksi-2-[2-(4′-metoksifenil)etil] kromona (3) dan 6-metoksi -2-(2-feniletil) kromona (4).
Hingga saat ini, berbagai strategi terus dikembangkan untuk meningkatkan produksi metabolit sekunder pada kultur sel tanaman. Metabolit sekunder biasanya terakumulasi dalam jaringan tanaman dalam kondisi tertentu. Untuk memaksimalkan produksi dan akumulasi metabolit sekunder dalam sel atau jaringan, dapat dilakukan dengan cara antara lain memanipulasi parameter lingkungan dan medium, memilih strain yang unggul, memberikan suplai prekursor, serta elisitasi (Mulabagal & Tsay 2004).
Jalur Biosintesis Senyawa Seskuiterpena
Senyawa seskuiterpena merupakan salah satu bagian dari terpenoid. Pada tumbuhan tingkat tinggi (Gambar 1), dua jalur sintesis terpenoid terlokalisasi di kompartemen intraseluler yang berbeda, melibatkan biosintesis isopentenyl diphosphate (IPP) dan isomernya dimethylallyl diphosphate (DMAPP). Dalam sitosol, IPP berasal dari jalur asam mevalonat (MVA) yang dimulai dengan kondensasi asetil-CoA, sedangkan di plastida, IPP dihasilkan dari piruvat dan gliseraldehida 3-fosfat (GA3P) melalui jalur methylerythritol phosphate (MEP).
Gambar 1. Jalur biosintesis terpenoid (Yu & Utsumi 2009)
menghasilkan rantai karbon bercabang sebagaimana ditemukan pada asam mevanolat. Reaksi-reaksi berikutnya ialah fosforilasi, eliminasi asam fosfat dan dekarboksilasi menghasilkan IPP yang selanjutnya berisomerisasi menjadi DMAPP oleh enzim isomerase. IPP sebagai unit isoprena aktif bergabung pada bagian kepala ke ekor dengan DMAPP. Untuk jalur sintesis plastida, prekursornya adalah piruvat dan GA3P.
Seskuiterpena merupakan golongan terbesar dari terpenoid. Tersusun dari 3 macam unit isoprena dengan 15 atom karbon. Seskuiterpena merupakan komponen utama minyak atsiri dan berperan penting dalam memberi aroma pada buah dan bunga. Seskuiterpena dipilih berdasarkan kerangka karbon dasarnya, yang umum adalah asiklik, monosiklik dan bisiklik. Beberapa jenis seskuiterpena antara lain kariofilena, farnesena, kurkumena, bisabolol.
3
METODE
Waktu dan Tempat Penelitian
Penelitian dilaksanakan dari bulan Februari 2014 sampai April 2016, bertempat di Laboratorium Biak Sel dan Jaringan Tumbuhan dan Laboratorium Kimia Bahan Alam, Pusat Penelitian Biologi, LIPI Cibinong, Bogor. Analisis senyawa seskuiterpena dengan GC-MS dilakukan di Indonesian Culture Collection (InaCC), Pusat Penelitian Biologi, LIPI Cibinong, Bogor.
Bahan Penelitian
Prosedur Penelitian
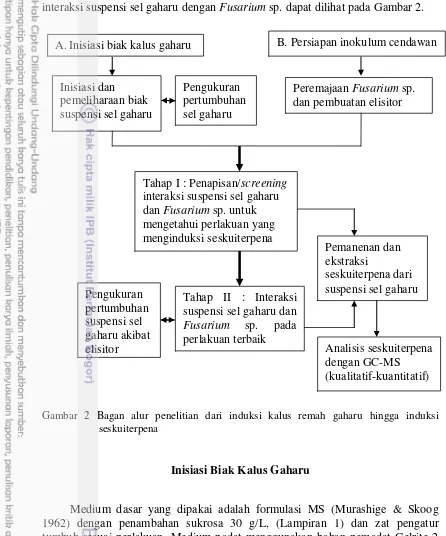
Prosedur penelitian dari inisiasi biak kalus remah gaharu hingga tahap interaksi suspensi sel gaharu dengan Fusarium sp. dapat dilihat pada Gambar 2.
Gambar 2 Bagan alur penelitian dari induksi kalus remah gaharu hingga induksi seskuiterpena
Inisiasi Biak Kalus Gaharu
Medium dasar yang dipakai adalah formulasi MS (Murashige & Skoog 1962) dengan penambahan sukrosa 30 g/L, (Lampiran 1) dan zat pengatur tumbuh sesuai perlakuan. Medium padat menggunakan bahan pemadat Gelrite 2 g/L, sedangkan untuk medium cair tanpa menggunakan bahan pemadat. Sebelum ditambahkan Gelrite, medium tumbuh diatur pHnya menjadi 5.7–5.8 dengan menambahkan larutan penyangga 0,1 N HCl atau KOH. Setelah itu diautoklaf pada suhu 121 oC dan tekanan 15 Psi selama 20 menit. Medium kemudian dibawa ke Laminair Air Flow Cabinet (LAFC) dan dituang secara aseptik ke Petri dish steril berukuran 90 mm × 20 mm, masing-masing sebanyak 25 mL. Medium
Peremajaan Fusarium sp. dan pembuatan elisitor
B. Persiapan inokulum cendawan A.Inisiasi biak kalus gaharu
tumbuh disimpan dalam lemari medium minimal sehari sebelum dipakai pada ruangan bersuhu 25 °C.
Sumber eksplan yang digunakan dalam percobaan ini adalah planlet gaharu A. malaccensis yang dipelihara secara in vitro (koleksi Laboratorium Biak Sel dan Jaringan, Pusat Penelitian Biologi, LIPI) berumur 2 bulan pada subkultur terakhir. Helai daun pada posisi kedua sampai dengan keempat dari pucuk dipotong pada kedua sisinya, dengan ukuran sekitar 0.5 x 0.8 cm2. Potongan daun ini ditanam pada medium perlakuan dan tiap perlakuan terdiri dari 3–4 ulangan (Petri dish), dan masing-masing Petri dish diisi 14 eksplan. Untuk mendapatkan potongan eksplan ruas batang, tunas gaharu ditumbuhkan terlebih dahulu pada medium MS ditambah 0.5 mg/L Giberelin (GA) untuk menginduksi pemanjangan ruasnya. Ruas batang yang dipakai untuk bahan percobaan berukuran panjang sekitar 0.5– 0.8 cm, setiap perlakuannya terdiri dari 3–5 ulangan (Petri dish), dan masing-masing Petri dish diisi 5 eksplan.
Induksi kalus A. malaccensis dipelajari dengan menggunakan zat pengatur tumbuh (ZPT) auksin 4-amino-3,5,6-trichloropyridine-2-carboxylic acid (Picloram), 2,4-dichlorophenoxyacetic acid (2,4-D), indole-3-acetic acid (IAA), indole-3-butyric acid (IBA), 1-naphthaleneacetic acid (NAA) serta sitokinin benzyl adenine (BA) baik tunggal maupun kombinasi pada eksplan potongan daun dan ruas batang. Perlakuan ZPT yang diberikan dapat dilihat pada Tabel 1.
Tabel 1 Perlakuan ZPT tunggal auksin serta kombinasi auksin dan auksin-sitokinin untuk induksi kalus A. malaccensis
Parameter persentase hidup, pembentukan kalus, penutupan kalus, serta morfologi kalus diamati pada umur 1 bulan setelah inisiasi. Persentase hidup dihitung dari jumlah eksplan yang hidup dibandingkan total eksplan yang ditanam. Persentase kalus dihitung dari jumlah eksplan yang membentuk kalus dibandingkan dengan total eksplan yang ditanam, sedangkan persentase penutupan kalus dihitung dari luas kalus yang telah menutup eksplan dibandingkan luas eksplan daun atau ruas batang. Data ditampilkandalam bentuk rata-rata dan standar error (SE). Salah satu kombinasi formulasi medium dan ZPT yang menghasilkan pertumbuhan kalus remah dan tumbuh cepat dengan tingkat Kombinasi Eksplan Jenis dan konsentrasi ZPT
Auksin tunggal Potongan daun Picloram (0, 0.1, 0,5, 1, dan 5 mg/L)
2,4-D (0, 0.1, 0.5, 1, dan 5 mg/L)
IBA (0, 5, 10, dan 15 mg/L)
IAA (0, 5, dan 15 mg/L)
Ruas batang 2,4-D (0, 0.1, 0.5, 1, dan 5 mg/L)
Auksin-auksin Potongan daun 2,4-D (1 dan 2 mg/L) + NAA (0, 0.1, 0.5, dan 1 mg/L)
Auksin-sitokinin Potongan daun 2,4-D (1 dan 2 mg/L) + BA (0, 0.1, 0.2, dan 0.3 mg/L)
NAA (0, 0.1, 0.5, dan 1 mg/L) + BA (0.1 dan 0.2 mg/L)
IBA (0.5, 1, 5, 10, dan 15 mg/L) + BA (0, 0.1, 0.2, 0.3 mg/L)
induksi kalus yang tinggi dipakai sebagai bahan untuk perlakuan suspensi sel gaharu. Inisiasi dan proliferasi kalus dilakukan di ruang gelap dengan suhu sekitar 25 oC.
Inisiasi dan Pemeliharaan Biak Suspensi Sel Gaharu
Sekitar 0,3 g (0,20–0,38 mL) kalus remah yang terbentuk dari eksplan yang tumbuh pada medium padat ditransfer ke medium cair untuk membuat kultur suspensi sel. Perlakuan yang digunakan adalah auksin 2,4-D (4 ulangan) dan Picloram (3 ulangan) secara tunggal, dengan konsentrasi masing-masing 0.1, 0.5, 1, 2, dan 5 mg/L. Agregat kalus dicacah sampai ukuran kecil (sekitar 1 mm) lalu dimasukkan ke dalam 25 mL medium pada labu Erlenmeyer 100 mL kemudian digoyang di atas shaker dengan kecepatan 120 rpm agar menghasilkan suspensi yang homogen. Ruang kultur diatur suhunya sekitar 22–25 oC. Pertumbuhan biak suspensi sel diukur tiap minggu dari minggu ke-1 sampai minggu terakhir (fase stationary) dengan cara memindahkan secara aseptik seluruh sel suspensi ke tabung sentrifugasi steril berukuran 50 mL, kemudian diukur volume sel yang mengendap. Data ditampilkan dalam bentuk rata-rata dan standar error (SE). Pemisahan sel-sel dari agregat besar dan kecil dilakukan dengan menyaring suspensi sel dengan stainless steel filter berukuran pori 1.5 mm pada saat subkultur. Sel-sel yang lolos pada saringan tersebut yang digunakan sebagai inokulum untuk disubkultur berikutnya. Subkultur dilakukan setiap 2 minggu dengan menggunakan medium terbaik.
Perlakuan Interaksi Suspensi Sel Gaharu dan Cendawan Fusarium sp. untuk Induksi Pembentukan Seskuiterpena
Tahap I : Penapisan/screening
Cendawan yang dipakai untuk percobaan ini berfungsi sebagai elisitor. Perlakuan terdiri dari :
1) Cendawan Fusarium sp. strain 372.3 dan strain w1 fus. sp.2 dan bentuk elisitornya yaitu : a) filtrat dari suspensi cendawan hidup (filtrat segar), b) filtrat dari biak suspensi cendawan diautoklaf, c) campuran filtrat segar dan filtrat diautoklaf, d) hifa diautoklaf, dan e) hifa hidup.
2) Umur suspensi sel gaharu saat co-culture yaitu 1, 2, 3 dan 4 minggu. 3) Lama waktu co-culture yaitu mulai dari 0–1, 1–2, dan 2–3 bulan.
Kontrol yang digunakan yaitu kultur suspensi gaharu tanpa perlakuan sebagai kontrol negatif, kultur suspensi gaharu ditambah 100 M asam salisilat dengan lama co-culture 1–2 bulan sebagai kontrol positif, serta kontrol elisitor hifa/filtrat Fusarium sp. strain 372.3 dan strain w1 fus. sp.2 tanpa suspensi gaharu. Kontrol gubal A. malaccensis yang diketahui mengandung resin wangi dipakai sebagai pembanding.
Brooth). Biak yang berumur 10 hari di medium PDB tersebut akan dipakai sebagai sumber filtrat maupun hifa yang dicampurkan dalam medium perlakuan suspensi sel gaharu. Filtrat segar dibuat dengan cara memisahkan hifa hidup dari medium suspensinya. Setelah itu medium disentrifusi dengan kecepatan 3000 rpm selama 20 menit untuk mengendapkan potongan sisa hifa, spora dan lain-lain. Selanjutnya supernatan disaring dengan filter milipore berukuran 0.2 m. Filtrat ini dipakai sebagai perlakuan filtrat segar yang dicampurkan ke medium yang telah diautoklaf dengan konsentrasi 10%. Filtrat diautoklaf diperoleh dari biak suspensi cendawan yang sudah diautoklaf, selanjutnya tahapannya sama dengan penyaringan filtrat segar. Filtrat campuran terdiri dari 5% filtrat segar dan 5% filtrat diautoklaf. Hifa diautoklaf berasal dari sisa hifa yang dipisahkan dari mediumnya pada pembuatan filtrat segar, lalu diautoklaf dan dicampurkan pada medium sebagai perlakuan sebanyak 0.5–1 g per 25 mL medium di labu Erlenmeyer. Untuk perlakuan hifa hidup, yang dipakai adalah potongan medium dengan hifa cendawan berukuran 0.5 cm2 dari kultur berumur 7 hari di medium PDA, lalu dicampurkan ke medium yang berisi suspensi gaharu.
Semua perlakuan menggunakan medium dasar MS dengan penambahan ZPT terbaik pada pembuatan suspensi sel, dengan sel sebanyak 0.3–0.5 g setiap volume 25 mL/labu Erlenmeyer 100 ml kemudian digoyang di atas shaker dengan kecepatan 120 rpm. Ruang kultur diatur suhunya sekitar 22–25 oC. Dari semua perlakuan interaksi ini, suspensi sel dipanen pada berbagai lama waktu co-culture untuk dianalisis kandungan senyawa seskuiterpenanya secara kualitatif dengan GC-MS. Dari hasil analisis perlakuan kualitatif sebelumnya, dipilih beberapa senyawa seskuiterpena pada perlakuan terbaik untuk dihitung secara kuantitatif dengan menggunakan senyawa standar.
Tahap II. Interaksi suspensi sel gaharu dan Fusarium sp. pada perlakuan terbaik Perlakuan kualitatif terbaik tersebut juga akan diulang untuk diuji kandungan seskuiterpenanya secara kuantitatif. Masing-masing perlakuan dibuat sebanyak 6 ulangan, yaitu 3 ulangan untuk dianalisis kandungan seskuiterpena dan 3 ulangan lainnya untuk diukur pertumbuhan selnya. Pertumbuhan biak sel gaharu pada perlakuan diamati setiap minggu hingga panen. Setelah itu ditimbang bobot basah kalusnya. Data ditampilkan dalam bentuk rata-rata dan standar error (SE). Pengukuran kandungan seskuiterpena secara kualitatif dan kuantitatif dilakukan dengan gas chromatography-mass spectrometry (GC-MS).
Pemanenan dan Ekstraksi Seskuiterpena dari Suspensi Sel
disimpan di freezer -20 oC sebelum dianalisis dengan GC-MS. Hal yang sama juga dilakukan untuk mengekstraksi suspensi sel kontrol dan semua perlakuan.
Analisis Seskuiterpena dengan GC-MS
Ekstrak yang diperoleh kemudian dianalisis senyawa seskuiterpenanya dengan metode GC-MS merek Shimadzu tipe QP2010 Ultra berdasarkan metode yang pernah dilakukan Okudera & Ito (2009). Kondisi GC-MS: kolom kapiler Rtx-5MS (30 m x 0.25 mm); gas pembawa helium; laju alir gas 2.72 mL/menit. Kolom diprogram dengan suhu awal 50 °C dan suhu akhir 250 °C. Kenaikan suhu dua tahap, pada tahap awal suhu kolom dibuat konstan 50 °C selama 3 menit dan kemudian dinaikkan sampai suhu 150 °C dengan kecepatan kenaikan suhu 5 °C/menit, selanjutnya suhu dinaikkan sampai 250 °C dengan kecepatan kenaikan suhu 3 °C/menit yang dipertahankan selama 4 menit. Suhu injektor diprogram konstan pada 250 °C, sedangkan volume sampel yang diinjeksikan adalah 5 µL. Setiap puncak dari kromatogram yang dihasilkan kemudian diidentifikasi massa dan fragmen-fragmen massanya dengan membandingkan fragmen massa dari senyawa yang telah diketahui menggunakan bank data dari National Institute Standard of Technology (NIST Library), Mainlib, dan Wiley.
Beberapa senyawa seskuiterpena yang terbentuk dipilih untuk dilakukan analisis secara kuantitatif. Senyawa standar dibuat larutan stoknya terlebih dahulu, lalu dibuat pengencerannya sebanyak 5 seri dari konsentrasi terendah hingga tertinggi, kemudian dianalisis dengan GC-MS pada kondisi yang sama dengan sampel. Luas area pada puncak kromatogram yang diperoleh dari masing-masing konsentrasi digunakan untuk menentukan kurva kalibrasi senyawa standar. Senyawa seskuiterpena pada sampel yang sama dengan senyawa standar dapat dihitung konsentrasinya menggunakan kurva kalibrasi tersebut, sedangkan seskuiterpena lain yang tidak ada standarnya, dapat dibandingkan konsentrasi relatifnya terhadap senyawa standar yang tersedia.
4
HASIL DAN PEMBAHASAN
Hasil
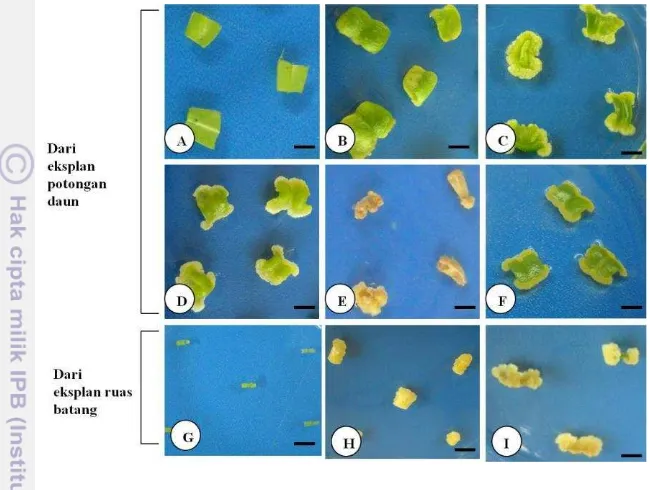
Inisiasi dan Morfologi Kalus dari Daun dan Ruas Batang
membentuk kalus kompak berwarna kuning (Gambar 3C), ada pula yang berupa kalus kapas berwarna putih (Gambar 3D). Sebagian eksplan ada yang mengalami pencoklatan dan mati (Gambar 3E). Beberapa medium perlakuan menginduksi kalus yang kecil-kecil, cukup remah berwarna kekuningan (Gambar 3F dan 3I).
Gambar 3 Perbedaan morfologi kalus dari eksplan potongan daun dan ruas batang A. malaccensis. (A-F) eksplan potongan daun; (B) pembentukan kalus pada tahap awal; (C) kalus kompak; (D) kalus kapas; (E) kalus mencoklat; (F) kalus remah; (G-I) eksplan ruas batang; (H) ruas batang mulai membengkak; (I) kalus dari ruas batang; bar = 5 mm
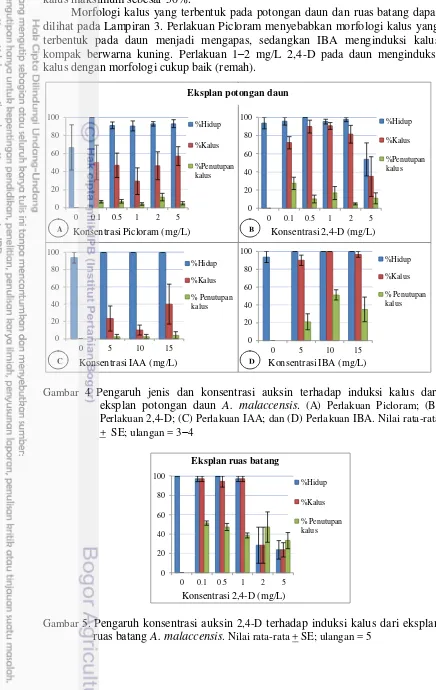
Pengaruh Pemberian Auksin Tunggal pada berbagai Konsentrasi
Pengaruh auksin terhadap frekuensi pembentukan kalus dan morfologinya pada eksplan potongan daun dan ruas batang dapat dilihat pada Gambar 3. Semua eksplan dapat hidup pada medium dengan penambahan auksin maupun tidak tetapi tanpa penambahan auksin, eksplan tidak dapat membentuk kalus. Perlakuan Picloram dan IAA, meskipun daya hidupnya tinggi tetapi hanya menginduksi kalus dengan persentase rendah (<60%) dengan penutupan kalus <20% (Gambar 4A dan 4C). Perlakuan 2,4-D memberikan daya hidup yang tinggi pada daun (90– 100%) dengan induksi kalus sekitar 70–90%, kecuali pada 5 mg/L 2,4-D hanya 53% yang bertahan hidup dengan persentase kalus dua pertiganya (35%). Daya hidup dan persentase kalus maksimum pada 0,5 dan 1 mg/L 2,4-D, dengan penutupan kalus 10–16% (Gambar 4B). ZPT IBA mampu menginduksi 100% eksplan daun untuk hidup dan membentuk kalus sebesar 96–100%. Persentase penutupan kalus maksimum pada potongan daun diperoleh pada 10 mg/L IBA sebesar 51% (Gambar 4D).
30% (Gambar 5). ZPT 0.1 mg/L 2,4-D mampu membentuk persentase penutupan kalus maksimum sebesar 50%.
Morfologi kalus yang terbentuk pada potongan daun dan ruas batang dapat dilihat pada Lampiran 3. Perlakuan Picloram menyebabkan morfologi kalus yang terbentuk pada daun menjadi mengapas, sedangkan IBA menginduksi kalus kompak berwarna kuning. Perlakuan 1–2 mg/L 2,4-D pada daun menginduksi kalus dengan morfologi cukup baik (remah).
Gambar 4 Pengaruh jenis dan konsentrasi auksin terhadap induksi kalus dari eksplan potongan daun A. malaccensis. (A) Perlakuan Picloram; (B) Perlakuan 2,4-D; (C) Perlakuan IAA; dan (D) Perlakuan IBA. Nilai rata-rata + SE; ulangan = 34
Gambar 5. Pengaruh konsentrasi auksin 2,4-D terhadap induksi kalus dari eksplan ruas batang A. malaccensis. Nilai rata-rata + SE; ulangan = 5
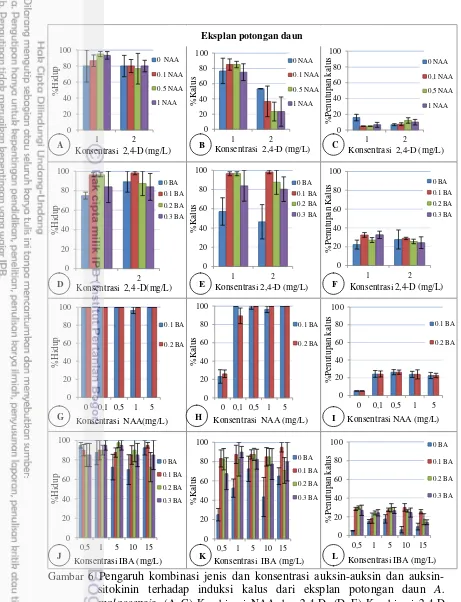
Perlakuan Kombinasi Auksin-Auksin dan Auksin-Sitokinin
Pengaruh kombinasi auksin-auksin serta auksin-sitokinin pada eksplan daun disajikan pada Gambar 6. Potongan daun dan ruas batang dapat hidup dengan persentase cukup tinggi (>70%) dan mampu membentuk kalus pada semua kombinasi ZPT tumbuh yang diberikan. Pada kombinasi 2,4-D dan NAA (Gambar 6A–6C), pemberian NAA pada konsentrasi 1 mg/L 2,4-D sedikit meningkatkan persentase eksplan membentuk kalus, tetapi pada konsentrasi 2 mg/L 2,4-D rata-ratanya menurun hingga kurang dari 50%. Sementara itu, untuk parameter persentase penutupan kalus pada kombinasi ini hanya mencapai 12%.
Pada kombinasi 2,4-D dan BA (Gambar 6D–6F), pemberian BA sangat baik untuk meningkatkan daya hidup potongan daun hingga lebih dari 80%. Penambahan konsentrasi 0.1 dan 0.2 mg/L BA pada 1 mg/L 2,4-D serta 0.1 mg/L BA pada 1 mg/L 2,4-D dapat menginduksi kalus hingga hampir 100%, sedangkan untuk persentase penutupan kalusnya tidak berbeda antarperlakuan dalam kombinasi 2,4-D dan BA (25–32%).
Kombinasi BA dan NAA menyebabkan tingginya daya hidup dan induksi kalus yang terbentuk pada eksplan (89–100%), dengan penutupan kalus sekitar 19–36% (Gambar 6G–6I). Potongan daun yang diberi perlakuan NAA tanpa BA dapat hidup 100%, tetapi tingkat induksi kalusnya rendah (<30%). Untuk kombinasi IBA dan BA (Gambar 6J–6L), daya hidup eksplan sekitar 70–100%. Tanpa penambahan BA, persentase kalusnya tidak terlalu tinggi, hanya mencapai 20–70%. Penambahan BA meningkatkan persentase kalusnya menjadi 6795%, dengan penutupan kalus 14–30%.
Pengaruh kombinasi auksin-auksin serta auksin-sitokinin pada eksplan ruas batang disajikan pada Gambar 7. Eksplan ruas batang juga memiliki daya hidup yang tinggi antara 86–100% (Gambar 7A) dengan kemampuan membentuk kalus 90100%, kecuali pada 2 mg/L 2,4-D yang diberi 0,3 mg/L 2,4-D hanya 50% (Gambar 7B). Persentase penutupan kalus pada eksplan ruas batang sekitar 31– 66% (Gambar 7C), lebih tinggi dibanding pada eksplan potongan daun.
auksin-Gambar 7. Pengaruh kombinasi konsentrasi 2,4-D dan BA terhadap induksi kalus dari eksplan ruas batang A.malaccensis. (A) Persentase hidup; (B) Persentase kalus; dan (C) Persentase penutupan kalus. Nilai rata-rata + SE; ulangan = 3
Inisiasi dan Pemeliharaan Biak Suspensi Sel A. malaccensis
Kultur suspensi dibuat dari kalus remah yang tumbuh dari eksplan daun. Kalus gaharu yang dipindahkan ke medium cair lama kelamaan akan membesar (Gambar 8A). Pengocokan yang terus-menerus menyebabkan terbentuknya sel-sel yang lepas-lepas, sehingga membentuk sekumpulan sel yang halus jika dilihat secara langsung (Gambar 8B).
Gambar 8. Morfologi kalus gaharu dalam medium cair. (A) Kalus yang membentuk gumpalan dengan diameter sekitar 2–5 mm; dan (B) Kalus sudah mulai membentuk kumpulan sel yang halus
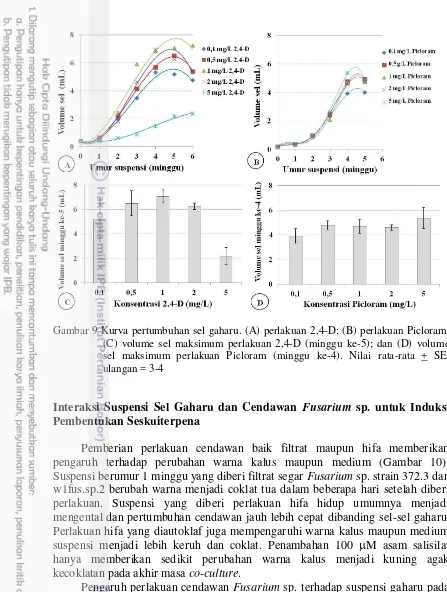
Pada perlakuan 2,4-D pertumbuhan suspensi gaharu membentuk kurva sigmoid pada semua konsentrasi, kecuali 5 mg/L (Gambar 9A). Sementara itu, penambahan Picloram pada medium memberikan respon pertumbuhan kalus berupa kurva sigmoid juga, tetapi volume selnya lebih rendah dan masa kulturnya lebih pendek (5 minggu) dibanding perlakuan 2,4-D yang berumur 6 minggu (Gambar 9B). Pertumbuhan sel maksimum pada perlakuan 2,4-D diperoleh pada konsentrasi 1 mg/L dengan volume sel 7 mL pada minggu ke-5 (Gambar 9C), sedangkan pada perlakuan Picloram respon maksimum volume selnya tidak berbeda nyata pada semua konsentrasi (Gambar 9D). Suspensi sel terbaik dengan kumpulan sel yang halus yaitu pada 0,5 dan 1 mg/L dipilih dan disubkultur di medium MS ditambah 1 mg/L 2,4-D pada fase linear.
Gambar 9 Kurva pertumbuhan sel gaharu. (A) perlakuan 2,4-D; (B) perlakuan Picloram; (C) volume sel maksimum perlakuan 2,4-D (minggu ke-5); dan (D) volume sel maksimum perlakuan Picloram (minggu ke-4). Nilai rata-rata + SE; ulangan = 3-4
Interaksi Suspensi Sel Gaharu dan Cendawan Fusarium sp. untuk Induksi Pembentukan Seskuiterpena
Pemberian perlakuan cendawan baik filtrat maupun hifa memberikan pengaruh terhadap perubahan warna kalus maupun medium (Gambar 10). Suspensi berumur 1 minggu yang diberi filtrat segar Fusarium sp. strain 372.3 dan w1fus.sp.2 berubah warna menjadi coklat tua dalam beberapa hari setelah diberi perlakuan. Suspensi yang diberi perlakuan hifa hidup umumnya menjadi mengental dan pertumbuhan cendawan jauh lebih cepat dibanding sel-sel gaharu. Perlakuan hifa yang diautoklaf juga mempengaruhi warna kalus maupun medium suspensi menjadi lebih keruh dan coklat. Penambahan 100 M asam salisilat hanya memberikan sedikit perubahan warna kalus menjadi kuning agak kecoklatan pada akhir masa co-culture.
karena hanya terdeteksi 1 senyawa tersebut. Perlakuan hifa hidup Fusarium sp. strain 372.3 pada umur suspensi 3 minggu dengan lama co-culture 2.5 bulan juga hanya menginduksi 1 senyawa seskuiterpena. Perlakuan hifa Fusarium sp. strain 372.3 yang diautoklaf menginduksi terbentuknya bau wangi gaharu dengan 1 senyawa seskuiterpena terdeteksi pada umur suspensi 4 minggu saat perlakuan dengan lama co-culture 1 bulan. Dibandingkan strain 372.3, tampaknya strain w1fus.sp.2 memiliki daya induksi yang rendah, karena hanya terbentuk 1 senyawa seskuiterpena pada pemberian perlakuan di umur suspensi 3 minggu dan lama co-culture 1 bulan. Baik kontrol suspensi gaharu, kontrol 100 M asam salisilat maupun hifa hidup Fusarium sp. strain w1fus.sp.2 masing-masing dapat pula menginduksi terbentuk senyawa seskuiterpena meskipun hanya sedikit yang terdeteksi. Pada kontrol filtrat ataupun hifa diautoklaf pada kedua strain cendawan sama sekali tidak terdeteksi senyawa seskuiterpena.
Gambar 10 Kenampakan fisik kultur hasil interaksi suspensi sel gaharu umur 1 minggu dengan filtrat Fusarium sp. strain 372.3. (A) Kontrol suspensi gaharu; (B) 10% filtrat segar; (C) 10% filtrat diautoklaf; dan (D) 5% filtrat segar + 5% filtrat diautoklaf
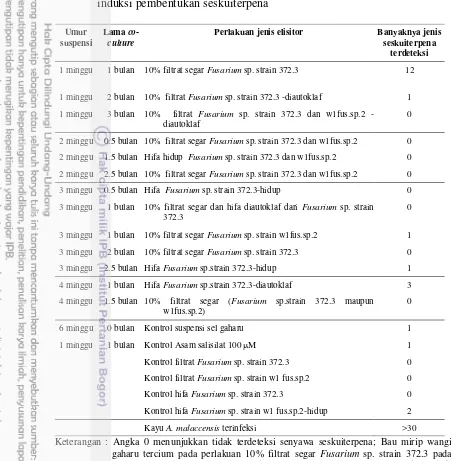
Tabel 2 Perlakuan interaksi suspensi sel gaharu dan cendawan Fusarium sp. untuk induksi pembentukan seskuiterpena
Keterangan : Angka 0 menunjukkan tidak terdeteksi senyawa seskuiterpena; Bau mirip wangi gaharu tercium pada perlakuan 10% filtrat segar Fusarium sp. strain 372.3 pada umur suspensi 1 minggu dan waktu co-culture 1 bulan serta perlakuan hifa Fusarium sp.strain 372.3-diautoklaf pada umur suspensi 4 minggu dan lama co-culture 1 bulan; Volume sampel yang diinjeksikan = 5 L
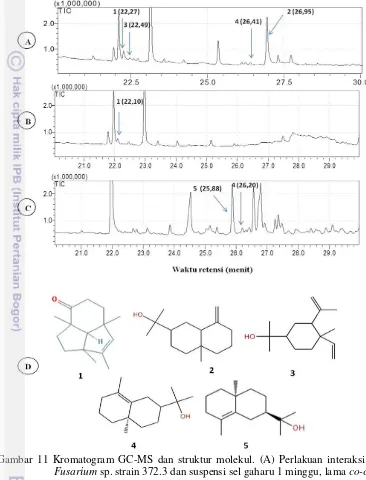
Kromatogram hasil analisis GC-MS pada perlakuan filtrat terbaik, kontrol suspensi dan kayu gaharu disajikan pada Gambar 11. Pada perlakuan filtrat terbaik (Gambar 11A), muncul peak di waktu retensi 26.9 menit yang diidentifikasi sebagai -eudesmol (identifikasi berdasarkan perbandingan dengan data base dan senyawa standar). Sekuiterpena lain yang muncul yaitu 1s,4R,7R,11R-1,3,4,7-Tetrametiltrisiklo [5.3.1.0(4,11)] undek-2-en-8-one, elemol dan -eudesmol dengan peak yang lebih kecil. Pada kontrol suspensi (Gambar 11B), peak senyawa 1s,4R,7R,11R-1,3,4,7-Tetrametiltrisiklo [5.3.1.0(4,11)] undek-2-en-8-on masih muncul. Kayu A. malaccensis terinfeksi berasal dari kayu yang sudah diketahui mengandung resin wangi (Gambar 11C). Pada kayu, banyak
Umur suspensi
Lama co-culture
Perlakuan jenis elisitor Banyaknya jenis
seskuiterpena
4 minggu 1 bulan Hifa Fusarium sp.strain 372.3-diautoklaf 3
4 minggu 1.5 bulan 10% filtrat segar (Fusarium sp.strain 372.3 maupun w1fus.sp.2)
0
6 minggu 0 bulan Kontrol suspensi sel gaharu 1
1 minggu 1 bulan Kontrol Asam salisilat 100 M 1
Kontrol filtrat Fusarium sp. strain372.3 0
Kontrol filtrat Fusarium sp. strainw1 fus.sp.2 0
Kontrol hifa Fusarium sp. strain372.3 0
Kontrol hifa Fusarium sp. strainw1 fus.sp.2-hidup 2
sekali terdeteksi senyawa seskuiterpena di antaranya -eudesmol dan 8-epi- -eudesmol tetapi tidak ada satupun yang mirip senyawa 1s,4R,7R,11R-1,3,4,7-Tetrametiltrisiklo [5.3.1.0(4,11)] undek-2-en-8-on, sedangkan untuk senyawa -eudesmol munculnya peak di waktu retensi yang berbeda dengan senyawa standar -eudesmol. Struktur molekul senyawa seskuiterpena yang terdeteksi dapat dilihat pada Gambar 11D.
Gambar 11 Kromatogram GC-MS dan struktur molekul. (A) Perlakuan interaksi filtrat Fusarium sp. strain 372.3 dan suspensi sel gaharu 1 minggu, lama co-culture 1 bulan; (B) Perlakuan kontrol suspensi sel gaharu 1.5 bulan; (C) Kayu gubal A.malaccensis; dan (D) struktur molekul senyawa yang terdeteksi. 1 = senyawa 1s,4R,7R,11R-1,3,4,7-Tetrametiltrisiklo [5.3.1.0(4,11)] undek-2-en-8-on (C15H22O); 2 = -eudesmol (C15H26O2); 3 = Elemol (C15H26O2); 4 =
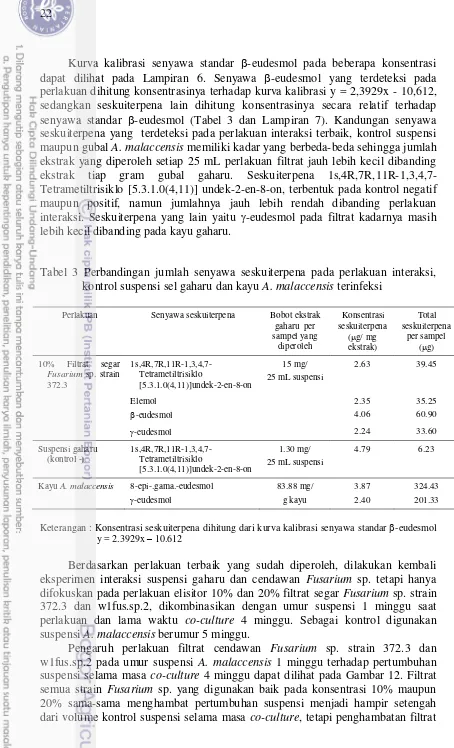
Kurva kalibrasi senyawa standar -eudesmol pada beberapa konsentrasi dapat dilihat pada Lampiran 6. Senyawa -eudesmol yang terdeteksi pada perlakuan dihitung konsentrasinya terhadap kurva kalibrasi y = 2,3929x - 10,612, sedangkan seskuiterpena lain dihitung konsentrasinya secara relatif terhadap senyawa standar -eudesmol (Tabel 3 dan Lampiran 7). Kandungan senyawa seskuiterpena yang terdeteksi pada perlakuan interaksi terbaik, kontrol suspensi maupun gubal A. malaccensis memiliki kadar yang berbeda-beda sehingga jumlah ekstrak yang diperoleh setiap 25 mL perlakuan filtrat jauh lebih kecil dibanding ekstrak tiap gram gubal gaharu. Seskuiterpena 1s,4R,7R,11R-1,3,4,7-Tetrametiltrisiklo [5.3.1.0(4,11)] undek-2-en-8-on, terbentuk pada kontrol negatif maupun positif, namun jumlahnya jauh lebih rendah dibanding perlakuan interaksi. Seskuiterpena yang lain yaitu -eudesmol pada filtrat kadarnya masih lebih kecil dibanding pada kayu gaharu.
Tabel 3 Perbandingan jumlah senyawa seskuiterpena pada perlakuan interaksi, kontrol suspensi sel gaharu dan kayu A. malaccensis terinfeksi
Perlakuan Senyawa seskuiterpena Bobot ekstrak gaharu per
Kayu A. malaccensis 8-epi-.gama.-eudesmol
-eudesmol
Keterangan : Konsentrasi seskuiterpena dihitung dari kurva kalibrasi senyawa standar -eudesmol y = 2.3929x – 10.612
Berdasarkan perlakuan terbaik yang sudah diperoleh, dilakukan kembali eksperimen interaksi suspensi gaharu dan cendawan Fusarium sp. tetapi hanya difokuskan pada perlakuan elisitor 10% dan 20% filtrat segar Fusarium sp. strain 372.3 dan w1fus.sp.2, dikombinasikan dengan umur suspensi 1 minggu saat perlakuan dan lama waktu co-culture 4 minggu. Sebagai kontrol digunakan suspensi A. malaccensis berumur 5 minggu.
Fusarium strain 372.3 lebih besar dibandingkan strain w1fus.sp.2. Konsentrasi 20% pada filtrat Fusarium sp. strain 372.3 menyebabkan penghambatan pertumbuhan yang lebih tinggi dibanding 10% filtrat yang sama maupun filtrat dari strain w1fus.sp.2. Pada saat panen, bobot basah terendah diperoleh pada 20% filtrat strain 372.3. strain w1fus.sp.2; dan 20% C2 = 20% filtrat Fusarium sp. strain w1fus.sp.2. Nilai rata-rata + SE; ulangan = 3
Perlakuan filtrat yang diberikan selama masa co-culture tidak semuanya menimbulkan aroma wangi pada kultur suspensi. Aroma wangi yang tercium pada perlakuan filtrat Fusarium sp. strain 372.3 agak berbeda dari strain w1fus.sp.2, namun tidak ada satupun yang mirip wangi gaharu. Pada saat pemanenan, dari masing-masing perlakuan dipilih tiga sampel terutama yang tercium aroma wangi untuk dianalisis kandungan seskuiterpenanya dengan GC-MS. Rata-rata jumlah ekstrak yang diperoleh pada masing-masing perlakuan disajikan pada Tabel 4.
Pada perlakuan filtrat Fusarium strain 372.3, peningkatan konsentrasi berpengaruh terhadap penurunan bobot ekstrak yang diperoleh tetapi tidak berpengaruh pada banyaknya seskuiterpena terdeteksi. Sementara itu, bobot ekstrak pada perlakuan 10% filtrat strain w1fus.sp.2 lebih besar dibanding pada konsentrasi 20%, tetapi pada konsentrasi 20% jumlah seskuiterpenanya lebih banyak dibanding konsentrasi 10%. Pada kontrol suspensi lima minggu hanya diperoleh ekstrak sebanyak 1.5 mg dengan jumlah seskuiterpena terdeteksi yang sangat sedikit.
Lama co-culture (minggu)
Tabel 4 Rata-rata jumlah ekstrak yang diperoleh dan jumlah seskuiterpena terdeteksi pada perlakuan 10% dan 20% filtrat Fusarium spp strain 372.3 dan w1fus.sp.2
Keterangan : Nilai rata-rata+SE; ulangan=3
Jenis seskuiterpena terdeteksi beserta kromatogram hasil analisis GC-MS pada perlakuan 10% dan 20% filtrat Fusarium sp. strain 372.3 dan w1fus.sp.2 dapat dilihat pada Lampiran 8 dan 9. Hasil analisis tersebut menunjukkan bahwa tidak ditemukan senyawa -eudesmol dengan peak yang waktu retensinya sama dengan senyawa standar -eudesmol seperti pada eksperimen sebelumnya, tetapi terdeteksi jenis seskuiterpena lain. Secara umum, jenis seskuiterpena yang ditemukan sangat variatif baik antar ulangan dalam satu perlakuan maupun antar perlakuan filtrat. Banyak dari seskuiterpena yang hanya ditemukan pada satu atau dua ulangan saja dalam satu perlakuan. Dari keseluruhan seskuiterpena yang terdeteksi dipilih yang sering muncul dan merupakan senyawa spesifik pada gubal gaharu yaitu dihidro--agarofuran, kariofilena oksida, -elemena, cis-Z-. .-bisabolena, (-)--panasinsena dan ginsenol.
Konsentrasi relatif seskuiterpena terpilih terhadap senyawa standar -eudesmol dapat dilihat pada Gambar 13. Senyawa dihidro--agarofuran dan (-)- -panasinsena hanya muncul pada perlakuan filtrat Fusarium sp. strain 372.3 dengan konsentrasi relatif 1015 g, sedangkan senyawa ginsenol hanya ditemukan pada perlakuan filtrat Fusarium sp. strain w1fus.sp.2 sebesar 423 g. Senyawa -elemena terdapat di hampir semua ulangan pada kedua strain Fusarium sp. tetapi pada strain 372.3 konsentrasinya lebih tinggi dibanding strain w1fus.sp.2. Senyawa kariofilena oksida umumnya muncul pada perlakuan Fusarium spp strain 372.3 dengan konsentrasi relatif >10 g, sedangkan cis-Z-. .-bisabolena umumnya muncul pada perlakuan strain w1fus.sp.2 dengan konsentrasi relatif <10 g.
Perlakuan Bobot ekstrak gaharu yang diperoleh per 25 mL suspensi (mg)
Banyaknya jenis seskuiterpena
terdeteksi
10% filtrat Fusarium sp. strain 372.3 (C1) 5.67 + 2.29 11 + 3.00
20% filtrat Fusarium sp. strain 372.3 (C1) 4.93 + 1.09 11 + 3.48
10% filtrat Fusarium sp. strain w1fus.sp.2 (C2) 3.33 + 1.42 7 + 1.00
20% filtrat Fusarium sp. strain w1fus.sp.2 (C2) 0.80 + 0.45 14 + 2.40
Gambar 13 Konsentrasi relatif beberapa seskuiterpena terdeteksi per 25 mL suspensi terhadap senyawa standar -eudesmol hasil analisis GC-MS pada interaksi filtrat cendawan Fusarium sp. strain 372.3 dan w1fus.sp.2 dan suspensi A. malaccensis. (A) Dihidro--Agarofuran; (B) Kariofilena oksida; (C) -Elemena; (D) cis-Z--Bisabolena; (E) (-)--Panasinsena; dan (F) Ginsenol. Km5 = kontrol suspensi gaharu umur 5 minggu; C1 10% = 10% filtrat Fusarium sp. strain 372.3; C1 20% = 20% filtrat Fusarium sp. strain 372.3; C2 10% = 10% filtrat Fusarium sp. strain w1fus.sp.2; dan C2 20% = 20% filtrat Fusarium sp. strain w1fus.sp.2. Nilai rata-rata; ulangan = 3
Pembahasan
Kalus gaharu dapat diinisiasi dari potongan daun ataupun ruas batang planlet A. malaccensis yang diberi perlakuan ZPT auksin tunggal maupun kombinasi auksin-auksin atau auksin-sitokinin. Ekplan dari planlet gaharu memiliki kemampuan proliferasi kalus yang baik, karena berdasarkan semua perlakuan auksin dan sitokinin yang dicoba, potongan daun dan ruas batang umumnya memiliki daya hidup >80% dengan tingkat pembentukan kalus >50%. Perlakuan auksin Picloram dan IAA, tidak direkomendasikan untuk menginduksi kalus gaharu karena daya hidupnya cukup rendah (<50%). Dari semua ZPT yang digunakan, pemberian IBA tunggal sangat efektif dalam menginduksi kalus dari segi persentase kalus maupun penutupan kalus, sedangkan 2,4-D lebih baik dalam menginduksi morfologi kalus menjadi cukup remah meskipun persentase kalus dan penutupan kalusnya kurang dari IBA. Pada kombinasi BA dan 2,4-D, adanya penambahan BA dalam konsentrasi rendah (0.1–0.3 mg/L) pada 1–2 mg/L 2,4-D lebih efektif memacu terbentuknya kalus dari segi persentase maupun keremahan kalus. Eksplan potongan daun dan ruas batang, keduanya dapat digunakan untuk menginduksi kalus dengan persentase yang tinggi dengan tipe remah. Persentase pembentukan dan penutupan kalus pada eksplan ruas batang lebih tinggi dibanding eksplan potongan daun pada perlakuan kombinasi BA dan 2,4-D. Hal ini dikarenakan pada ekplan ruas batang terdapat pembuluh xilem dan floem yang mengalirkan unsur hara dan hasil fotosintesis, serta kandungan auksin endogen yang lebih tinggi dibanding eksplan potongan daun.
Pembentukan kalus pada eksplan, secara fisiologi dipengaruhi oleh perubahan genetik pada sel tanaman oleh auksin. Sel yang merespon auksin akan menyebabkan dediferensiasi dan memacu pembelahan sel (George 1993). Tiap jenis auksin terikat pada reseptor yang berbeda-beda supaya terjadi perubahan metabolik yang mengarah pada penguatan sinyal sehingga menyebabkan respon tumbuhan terhadap ZPT tersebut. Selain itu, aplikasi konsentrasi auksin yang berbeda-beda sangat diperlukan karena setiap ZPT memiliki konsentrasi optimal yang berbeda untuk dapat menginduksi pertumbuhan kalus secara maksimal (Pierik 1997). IAA dan IBA merupakan auksin alami yang disintesis oleh tumbuhan, efektif untuk iniasisi akar adventif, tetapi tidak terlalu efektif dalam pembentukan kalus, sedangkan 2,4-D yang merupakan auksin sintetik yang berfungsi sebaliknya, memiliki pengaruh kuat dalam membentuk kalus (George 1993). Auksin 2,4-D merupakan herbisida selektif, diserap melalui daun dan ditranslokasikan ke jaringan meristem tanaman dapat meningkatkan laju pertumbuhan pada konsentrasi yang optimal (Song 2014).
penambahan 10-6M 2,4-D dan 10-6M BA (Okudera dan Ito 2009), sedangkan kombinasi 2 mg/L 2,4-D dan 0.1 mg/L kinetin dapat menginduksi kalus dari daun A. malaccensis hingga 70-73% dengan biomassa yang tinggi dan tumbuh cepat (Saikia et al., 2013). Khairunisa (2013) dan Zalikha (2013) melaporkan bahwa kalus dari eksplan lamina A. beccariana dan daun A. malaccensis dapat diinduksi dari medium MS ditambah 2 mg/L 2,4-D dan 0,5 mg/L BAP, sedangkan Jayaraman et al. (2014) memperoleh kalus remah dari eksplan daun yang berasal dari persemaian dengan biomassa tertinggi (berat kering 93.3 mg) melalui kombinasi hormon 1.1 μM NAA dan β.β μM BAP.
minggu ke-2 sampai ke-3 di medium MS ditambah 1 mg/L 2,4-D. Penyaringan dengan diameter <1.5 mm pada saat subkultur akan membantu mendapatkan kultur dengan kumpulan sel yang halus dan homogen.
Pada sistem suspensi gaharu in vitro, induksi dengan elisitor berupa faktor biotik atau abiotik merupakan prasyarat untuk memaksimalkan produksi dan akumulasi metabolit sekunder pada jaringan (Mulabagal & Tsay 2004). Dalam hal ini, filtrat maupun hifa berfungsi sebagai elisitor biotik yang dapat meningkatkan sintesis senyawa metabolit sekunder (Patel & Krishnamurthy 2013). Elisitor merupakan molekul sinyal yang dapat menggerakkan proses transduksi sinyal secara cepat yang mengarah pada aktivasi dan ekspresi gen yang terlibat di dalam biosintesis metabolit sekunder (Zhao et al. 2005; Wang & Wu 2013).Elisitor bisa berupa molekul yang dilepaskan secara endogen oleh tumbuhan ataupun molekul yang diekskresikan patogen pada saat terjadinya luka.
Filtrat cendawan yang diberikan tidak membunuh sel gaharu. Umur suspensi gaharu yang tepat pada saat perlakuan elisitor juga berpengaruh terhadap terbentuk atau tidaknya seskuiterpena. Semakin muda kultur suspensi sel pada saat co-culture, maka semakin banyak sel baru yang tumbuh dan terpengaruh senyawa toksin pada filtrat. Pada saat co-culture diharapkan terjadi proses induksi dan diferensiasi pada sel-sel yang baru terbentuk. Khayrunnisa (1999) melaporkan bahwa elisitor filtrat kasar Acremonium sp. dapat menginduksi lebih banyak senyawa wangi gaharu dibandingkan jenis elisitor lain (dinding sel maupun sitoplasmanya) pada kultur kalus Aquilaria crassna. Pada filtrat yang diautoklaf, kemungkinan toksin dalam filtrat rusak akibat pemanasan sehingga daya stimulasi terhadap sel untuk membentuk seskuiterpena menjadi berkurang. Senyawa toksin yang disekresikan secara ekstraseluler oleh cendawan dapat mengganggu permeabilitas dinding sel tanaman sehingga menyebabkan kerusakan struktur membran. Umumnya toksin merupakan senyawa sekunder berbobot molekul rendah yang dikeluarkan secara ekstraseluler oleh cendawan. Kelompok Fusarium menghasilkan beberapa jenis toksin seperti enniatin, fumonisin, sambutoksin, dan trikotesen (Kim et al. 1995; Hermann et al. 1996; Seo et al. 1996; Kang & Buchenenauer 2002; Langevin et al. 2004).
Mekanisme induksi seskuiterpena saat co-culture kemungkinan tidak disebabkan induksi asam salisilat karena senyawa yang dihasilkan hanya sedikit. Asam salisilat merupakan sinyal utama dalam mengekspresikan gen ketahanan yang mengatur regulasi resistensi terhadap cendawan dan bakteri pathogen (Patel & Krishnamurthy 2013). Pada elisitor berupa hifa cendawan, induksi seskuiterpena dapat terjadi karena kontak sel gaharu dan dinding sel cendawan. Dinding sel cendawan relatif stabil terhadap panas, jadi meskipun hifa diautoklaf, masih dapat menginduksi terbentuknya seskuiterpena. Komponen dinding sel utama cendawan berupa polisakarida glukan, berperan penting dalam patogenitas cendawan pada tumbuhan sehingga dapat merangsang inang tumbuhan yang terinfeksi membentuk fitoaleksin (Pitson et al. 1997).
sehingga konsentrasinya menjadi lebih lebih tinggi dibanding yang tidak diberi perlakuan. Seskuiterpena -eudesmol, elemol dan -eudesmol hanya terdeteksi pada perlakuan filtrat namun tidak ada pada kontrol suspensi, berarti senyawa tersebut merupakan hasil interaksi sel gaharu dan Fusarium sp. Rijal (2011) melaporkan bahwa senyawa cis--bisabolena masih terdeteksi pada kalus A. malaccensis umur 3 minggu yang tidak diberi perlakuan cendawan Fusarium sp.
Kandungan senyawa seskuiterpena pada tiap jenis gaharu baik di alam maupun in vitro berbeda-beda. Berdasarkan laporan Jong et al. (2014), senyawa -eudesmol serta -elemol diketahui terbentuk pada ekstrak jenis gaharu A. malaccensis di alam dengan kualitas A dan tanaman induk di alam setelah 6 bulan perlukaan, sedangkan dari sampel minyak gaharu (Aquilaria sp.) diperoleh persentase relatif senyawa 10-Epi--eudesmol kualitas A (high quality) sebesar 8.1–10.9% dan kualitas B (unknown quality) sebesar 7.1–20.6% (Azah et al. 2014). Senyawa -eudesmol teridentifikasi pada sampel kayu dan minyak gaharu Malaysia kualitas C yang berasal dari Terengganu sebesar 5.74% (Azah et al. 2008).
Perlakuan 10% dan 20% filtrat kedua strain Fusarium sp. yang diberikan mampu menghambat pertumbuhan sel gaharu tetapi tidak sampai tahap mematikan sehingga sel-sel gaharu masih dapat menimbulkan aroma wangi sebagai respon pertahanan terhadap toksin dalam filtrat. Senyawa seskuiterpena banyak ditemukan pada suspensi yang diberi filtrat. Dalam hal ini, pengurangan produk metabolisme primer berupa pertumbuhan sel bergeser menjadi biosintesis metabolisme sekunder sedangkan pada kontrol suspensi, pertumbuhan hanya mengarah untuk menghasilkan produk metabolisme primer saja sehingga biosintesis metabolisme sekunder hanya sedikit (Bourgaud et al. 2001). Di alam, interaksi cendawan patogen akan menyebabkan terjadinya perubahan fisiologis pada tanaman yang berdampak terhadap terjadinya perubahan visual pada sel, jaringan, atau organ tanaman. Putri (2011) melaporkan bahwa Fusarium sp. IPBCC. 08.569 memiliki patogenitas terhadap pohon gaharu (Aquilaria sp.) dalam proses pembentukan gaharu ditingkat sel dan organ dengan respon berupa akumulasi senyawa terpenoid yang ditemukan pada parenkima jejari, included phloem, unsur trakea xilem, dan empelur. Perubahan yang terjadi pada tingkat sel menyebabkan terjadinya perubahan warna kayu yang terjadi meluas ke luar daerah infeksi dan terbentuknya aroma wangi. Jayaraman & Mohammed (2015) melaporkan bahwa terjadi penurunan bobot kalus pada kultur suspensi dari daun A. malaccensis yang diberi 8 mg ekstrak kasar Trichoderma, tetapi justru muncul aroma yang agak mirip wangi gaharu setelah 20 hari masa co-culture dan setelah dianalisis GC-MS terdeteksi ada seskuiterpena 8-epi-gama eudesmol dan -guaiena.