INDUKSI POLIPLOIDI ANGGREK BULAN (
Phalaenopsis
sp.)
MENGGUNAKAN KOLKISIN PADA ORGAN GENERATIF
DAN
PROTOCORM
TUBAGUS KIKI KAWAKIBI AZMI
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa tesis berjudul Induksi Poliploidi Anggrek Bulan (Phalaenopsis sp.) menggunakan Kolkisin pada Organ Generatif dan Protocorm adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Februari 2015
Tubagus Kiki Kawakibi Azmi
RINGKASAN
TUBAGUS KIKI KAWAKIBI AZMI. Induksi Poliploidi Anggrek Bulan (Phalaenopsis sp.) menggunakan Kolkisin pada Organ Generatif dan Protocorm. Dibimbing oleh DEWI SUKMA, SANDRA ARIFIN AZIZ dan MUHAMAD SYUKUR.
Spesies anggrek bulan (Phalaenopsis sp.) memiliki potensi yang besar untuk dikembangkan sebagai varietas anggrek bulan komersial, salah satunya melalui induksi poliploidi. Poliploid telah diketahui memberikan banyak manfaat dalam perbaikan karakter berbagai tanaman, terutama yang berhubungan dengan sifat gigantisme. Bunga anggrek bulan merupakan salah satu aspek penting yang sering menjadi perhatian pemulia dan petani anggrek. Salah satu karakter penting yang diinginkan dari anggrek bulan adalah ukuran bunga yang besar. Hal itulah yang melatarbelakangi usaha perbaikan karakter spesies anggrek bulan melalui induksi poliploidi menggunakan kolkisin.
Perbaikan karakter pada anggrek bulan diprioritaskan pada beberapa anggrek bulan spesies penting yang sering digunakan sebagai tetua persilangan, diantaranya adalah Phalaenopsis amabilis dan P. gigantea. Kedua spesies tersebut dapat ditemukan di Indonesia sehingga sangat potensial untuk dikembangkan. Umumnya P. amabilis digunakan sebagai tetua persilangan untuk menghasilkan hibrida dengan bunga besar putih, sedangkan P. gigantea digunakan sebagai tetua untuk menghasilkan hibrida novelti. Induksi poliploidi pada kedua spesies tersebut diarahkan untuk menghasilkan spesies dengan karakter bunga unggul yang dapat digunakan sebagai tetua persilangan ataupun spesies unggul komersial. Tujuan dari penelitian ini adalah mempelajari beberapa aspek penting dan menghasilkan teknik yang efektif dan efisien dalam poliploidisasi anggrek bulan menggunakan kolkisin. Tiga jenis organ generatif P. amabilis yaitu bunga setelah penyerbukan, kuncup bunga, dan spike, serta protocorm P. gigantea, digunakan pada percobaan yang terpisah secara pararel sebagai bahan untuk perlakuan kolkisin pada berbagai konsentrasi. Kisaran konsentrasi yang digunakan untuk perlakuan kolkisin pada organ generatif P . amabilis adalah 0-2000 mg L-1, sedangkan pada protocorm P. gigantea adalah 0-100 mg L-1.
Hasil percobaan menunjukkan bahwa konsentrasi kolkisin yang tinggi menghambat pekembangan ketiga jenis organ generatif P. amabilis dalam menghasilkan buah anggrek sampai pada perkecambahan biji. Regenerasi
sebesar 92.8% dari perlakuan kolkisin pada bunga setelah penyerbukan diperoleh pada konsentrasi 500 mg L-1, sedangkan dari perlakuan kolkisin pada kuncup bunga yang diserbuki sendiri, sebanyak 100.0% dari konsentrasi 1000 mg L-1. Pengujian jumlah kromosom dari ujung akar secara sitologi terhadap tipe planlet DP dari perlakuan kolkisin pada bunga setelah penyerbukan dengan konsentrasi 500 mg L-1 menunjukkan bahwa keseluruhan dari tipe planlet DP adalah planlet tetraploid (100.0%).
Diduga terdapat perbedaan mekanisme polipoidisasi dari setiap perlakuan kolkisin pada organ generatif. Perlakuan kolkisin pada bunga setelah penyerbukan menghasilkan planlet tetraploid melalui induksi poliploid pada proses pembentukan zigot ataupun embrio. Perlakuan kolkisin pada kuncup bunga diduga menghasilkan planlet poliploid melalui fertilisasi antara dua gamet poliploid. Kemungkinan pembentukan gamet jantan yang dihasilkan adalah gamet diploid atau gamet tetraploid, sedangkan kemungkinan gamet betina yang terbentuk adalah gamet diploid. Berdasarkan kondisi tersebut akan diperoleh planlet dengan dua kemungkinan tingkat ploidi, yaitu tetraploid melalui penggabungan gamet jantan dan betina yang diploid, atau planlet heksaploid melalui penggabungan gamet jantan tetraploid dan gamet betina diploid.
SUMMARY
TUBAGUS KIKI KAWAKIBI AZMI. Polyploidy Induction of Moth Orchid (Phalaenopsis sp.) Generative Organ and Protocorm using Colchicine. Supervised by DEWI SUKMA, SANDRA ARIFIN AZIZ and MUHAMAD SYUKUR.
Moth orchid species (Phalaenopsis sp.) is the foundation in developing moth orchid commercial varieties, one of which by polyploidization. Polyploid has contributed in many plant improvement which is related to especially gigantic character. Moth orchid flowers are generally one of important aspects that acquire a tremendous attention from orchid breeders and growers. One of its important character is size. This aspect was carried as a background in attempt to improve the size moth orchid species flower by polyploidy induction using colchicine.
Moth orchid species improvement is prioritized on some important species which often used as a parent in hybridization, such as Phalaenopsis amabilis and
P. gigantea. Both species are Indonesia native species so that they are very potential to be developed. In general P. amabilis is used as parental plant to generate white big flower hybrid, whereas P. gigantea is used as parental plant to generate novelty hybrids. Polyploidy induction on both species were headed to produce moth orchids with well flower characters for using as a parent in hybridization used or commercial moth orchid species.
This research aims were to study some important aspects and to obtain an effective and efficient technique in moth orchid polyploidization using colchicine. Three different kinds of P. amabilis generative organs which were pollinated flower, flower bud, and spike, and also P. gigantea protocorm were used in separated parallel experiments as material for colchicine treatments in various concentrations. The range of colchicine concentrations in P. amabilis generative organ treatment were 0-2000, and 0-100 mg L-1 in P. gigantea protocorm treatment.
The result showed that higher colchicine concentration inhibited P. amabilis
generative organs development to produce orchid pods and its seeds germination.
Phalaenopsis gigantea protocorm regeneration into seedling was also inhibited in higher colchicine concentration. There were two experiments which conducted till screening of polyploid putative seedling, which were colchicine treatment on pollinated flower and flower bud of P. amabilis, and showed its potential in obtaining polyploid seedlings. Seedling screening based on morphological characters, both in colchicine treatment on pollinated flower and flower bud, showed that polyploid putative seedling (PPS) significantly different compared to diploid control seedling for basal organ of the protocorm (BOP), leaf and root, and also stomatal density in colchicine treatment on pollinated flower. The highest number percentage of PPS in colchicine treatment on pollinated flower was 92.8% which obtained from 500 mg L-1 colchicine, and in flower bud was 100.0% which obtained from 1000 mg L-1 and by self pollinated. Cytological analysis on PPS obtained from colchicine treatment on pollinated flower with 500 mg L-1 concentration resulted that all of PPS were 100.0% tetraploid seedlings.
zygot formation or embryogenesis. While colchicine treatment on flower bud could be resulted polyploid seedling by fertlization of two gamete polyploids. Possibilities of male gametes polyploid formation were diploid or tetraploid, whereas female gametes were diploid. Based on some gametes polyploid formation possibilities, there would be two polyploid seedlings generated, which were tetraploid seedling which formed by fertilization between diploid male and female gametes, or hexaploid seedling which formed by fertilization between tetraploid male and diploid female gametes.
© Hak Cipta Milik IPB, Tahun 2015
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Pemuliaan dan Bioteknologi Tanaman
INDUKSI POLIPLOIDI ANGGREK BULAN (
Phalaenopsis
sp.)
MENGGUNAKAN KOLKISIN PADA ORGAN GENERATIF
DAN
PROTOCORM
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2015
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah SWT yang telah memberikan segala karunia dan rahmat-Nya sehingga penelitian dan karya ilmiah ini dapat diselesaikan. Penelitian dengan judul Induksi Poliploidi Anggrek Bulan (Phalaenopsis sp.) menggunakan Kolkisin pada Organ Generatif dan Protocorm
telah terlaksana dengan baik atas dukungan dari banyak pihak.
Ucapan terima kasih dan penghargaan yang sebesar-besarnya penulis sampaikan kepada:
1. Dr Dewi Sukma, SP MSi, Prof Dr Ir Sandra Arifin Aziz, MS, dan Prof Dr Muhamad Syukur, SP MSi yang telah banyak memberikan pengarahan serta bimbingan sejak awal penelitian hingga penulisan tesis.
2. Pihak Direktorat Jendral Pendidikan Tinggi selaku penyelenggara Beasiswa Unggulan DIKTI 2012 yang telah membiayai selama masa studi S2, serta dana hibah Penelitian Unggulan Perguruan Tinggi Desentralisasi tahun 2012-2014 yang diketuai oleh Prof Dr Ir Sandra Arifin Aziz, MS, yang telah mendukung pendanaan penelitian.
3. Kepada pihak-pihak yang telah banyak membantu penelitian: Bapak Slamet, Ibu Juariah selaku teknisi Laboratorium Kultur Jaringan Tanaman 1, Bapak Joko selaku teknisi Laboratorium Mikro Teknik, Bapak Edi selaku teknisi Angle House, serta rekan-rekan di Laboratorium yaitu Tika, Bu Tendy, Erick, Elin.
4. Orang tua tercinta Bapak Tb Ahmad Kurdi Yurani (Alm) dan Ibu Durrotul Bahiyah yang selalu menjadi inspirasi bagi penulis, serta keluarga besar yang selalu mendukung melalui do’a.
5. Seluruh teman-teman mahasiswa pascasarjana program studi Pemuliaan dan Bioteknologi Tanaman tahun 2012 yang telah membantu baik secara langsung ataupun tidak langsung melalui diskusi-diskusi.
Semoga karya ilmiah ini dapat memberikan kontribusi terhadap perkembangan ilmu pengetahuan dibidang pertanian secara umum, serta terhadap kemajuan pemuliaan anggrek bulan secara khusus.
Bogor, Februari 2015
DAFTAR ISI
DAFTAR TABEL vii
DAFTAR GAMBAR vii
DAFTAR LAMPIRAN viii
1 PENDAHULUAN 1
Latar Belakang 1
Perumusan Masalah 3
Tujuan Penelitian 3
Hipotesis 4
Manfaat Penelitian 4
Ruang Lingkup Penelitian 4
2 INDUKSI POLIPLOIDI PADA ANGGREK BULAN (Phalaenopsis amabilis
(L.) Blume) MELALUI APLIKASI KOLKISIN PADA BUNGA SETELAH
PENYERBUKAN 6
Pendahuluan 7
Bahan dan Metode 8
Hasil dan Pembahasan 10
Simpulan 20
3 POTENSI INDUKSI POLIPLOIDI BERDASARKAN MORFOLOGI ANGGREK BULAN (Phalaenopsis amabilis (L.) Blume) MELALUI
PERLAKUAN KOLKISIN PADA KUNCUP BUNGA 23
Pendahuluan 24
Bahan dan Metode 25
Hasil dan Pembahasan 26
Simpulan 34
4 PENGARUH PERLAKUAN KOLKISIN PADA PERKEMBANGAN SPIKE
ANGGREK BULAN (Phalaenopsis amabilis (L.) Blume) 37
Pendahuluan 38
Bahan dan Metode 39
Hasil dan Pembahasan 39
Simpulan 42
5 PENGARUH PERLAKUAN KOLKISIN PADA PERKEMBANGAN
PROTOCORMPhalaenopsis gigantea (L.) Blume) 43
Pendahuluan 44
Bahan dan Metode 45
Hasil dan Pembahasan 45
Simpulan 47
6 PEMBAHASAN UMUM 49
Simpulan 54
Saran 54
DAFTAR PUSTAKA 55
LAMPIRAN 59
RIWAYAT HIDUP 60
DAFTAR TABEL
1. Jumlah bunga perlakuan dan beberapa karakter buah yang dihasilkan
dari perlakuan kolkisin pada bunga setelah penyerbukan P. amabilis 12
2. Rata-rata jumlah daun, akar, protocorm like bodies (plbs) dan
persentase planlet membentuk plbs dari planlet hasil perlakuan kolkisin pada bunga setelah penyerbukan P. amabilis dan kontrol 16
3. Hasil uji kontras ortogonal pada ukuran basal organ of the protocorm
(BOP), daun, akar, dan stomata dari planlet hasil perlakuan kolkisin pada bunga setelah penyerbukan P. amabilis dan kontrol pada 16
minggu setelah subkultur (MSSk) 17
4. Hasil pengujian jumlah kromosom secara sitologi pada planlet hasil
perlakuan kolkisin pada bunga setelah penyerbukan 19
5. Jumlah perlakuan kuncup bunga dan beberapa karakter pada berbagai
konsentrasi kolkisin 26
6. Rata-rata jumlah daun dan akar dari planlet hasil perlakuan kolkisin
pada kuncup bunga P. amabilis dan kontrol 28
7. Persentase dan rata-rata protocorm like bodies (plbs) dari planlet hasil
perlakuan kolkisin pada kuncup bunga P. amabilis dan kontrol 29
8. Perbandingan beberapa karakter antara tipe planlet normal (N), tipe
planlet diduga poliploid (DP) hasil perlakuan kolkisin pada kuncup
bunga P. amabilis, dan kontrol pada 8 MSSk 32
9. Pengaruh kolkisin terhadap pertumbuhan dan perkembangan spike
membentuk bunga, serta beberapa karakterisitk buah 41
10. Pengaruh perlakuan kolkisin terhadap beberapa karakter pertumbuhan
dan perkembangan protocorm P. gigantea pada 14 minggu setelah
subkultur (MSSk) 47
11. Pengaruh perlakuan kolkisin terhadap rata-rata jumlah daun dan akar
planlet P. gigantea selama 14 minggu setelah subkultur (MSSk) 47
12. Perbandingan jenis material anggrek bulan untuk perlakuan kolkisin
terhadap kemungkinan tingkat ploidi dari planlet yang dihasilkan 53
DAFTAR GAMBAR
1. Bagan alir penelitian Induksi Poliploidi Anggrek Bulan (Phalaenopsis
sp.) Menggunakan Kolkisin pada Organ Generatif dan Protocorm 5
3. Bunga P. amabilis tiga hari setelah penyerbukan dan pembuangan sepal
dan petal (kastrasi) 9
4. Keragaan buah P. amabilis umur 8 minggu setelah perlakuan (MSPr)
dari perlakuan kolkisin pada bunga setelah penyerbukan pada berbagai
konsentrasi kolkisin 10
5. Pengaruh konsentrasi kolkisin terhadap persentase buah jadi hasil
perlakuan kolkisin pada bunga setelah penyerbukan P. amabilis 11
6. Perkecambahan biji anggrek P. amabilis menjadi protocorm dari
perlakuan kolkisin pada bunga setelah penyerbukan umur 8 minggu
setelah semai (MSS) 13
7. Planlet abnormal anggrek P. amabilis dari perlakuan kolkisin pada
bunga setelah penyerbukan 500 mg L-1 selama lima hari umur 16 MSSk 14
8. Perbandingan morfologi tipe planlet diduga poliploid (DP) dan tipe
planlet normal (N) hasil perlakuan kolkisin pada bunga setelah penyerbukan, dan planlet kontrol pada 16 minggu setelah subkutur
(MSSk) 16
9. Perbandingan basal organ of the protocorm (BOP), densitas dan ukuran
stomata, serta jumlah kromosom planlet poliploid dan tipe planlet normal (N) hasil perlakuan kolkisin pada bunga setelah penyerbukan P.
amabilis dengan konsentrasi 500 mg L-1 selama lima hari dan planlet
kontrol diploid 18
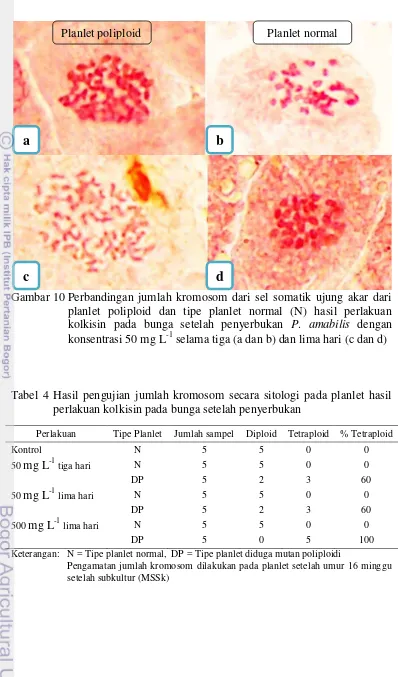
10. Perbandingan jumlah kromosom dari sel somatik ujung akar dari planlet
poliploid dan tipe planlet normal (N) hasil perlakuan kolkisin pada bunga setelah penyerbukan P. amabilis dengan konsentrasi 50 mg L-1
selama tiga dan lima hari 19
11. Mekanisme fertilisasi dan induksi poliploidi perlakuan kolkisin pada
bunga P. amabilis setelah penyerbukan 20
12. Kuncup bunga P. amabilis untuk perlakuan kolkisin 26 13. Pengaruh konsentrasi kolkisin terhadap persentase buah jadi perlakuan
kolkisin kuncup bunga P. amabilis 27
14. Basal organ of the protocorm (BOP) dan protocorm like bodies (plbs)
planlet hasil perlakuan kolkisin kuncup bunga P. amabilis pada konsentrasi 1000 mg L-1 yang diserbuki sendiri 30
15. Perbedaan morfologi antara tipe planlet diduga poliploid (DP) dan tipe
planlet normal (N) dari setiap perlakuan kolkisin pada kuncup bunga P.
amabilis dan kontrol pada 8 MSSk 30
16. Skema mekanisme-mekanisme dari sel induk polen dalam proses
pembentukan polen poliploid pada anggrek 34
17. Tiga kemungkinan mekanisme induksi poliploidi selama
mikrosporogenesis dari perlakuan kolkisin pada kuncup bunga P.
amabilis 35
18. Spike yang digunakan untuk perlakuan kolkisin 39
19. Pengaruh perlakuan kolkisin pada perkembangan spike P. amabilis
sekitar dua minggu setelah perlakuan kolkisin 40
20. Variasi ukuran buah P. amabilis yang dihasilkan dari perlakuan
kolkisin 50 mg L-1 pada spike dan kontrol 42
21. Protocorm P. gigantea umur sembilan bulan dari perkecambahan biji
22. Proliferasi protocorm like bodies (plbs) dari protocorm P. gigantea
perlakuan kolkisin 50 mg L-1 46
DAFTAR LAMPIRAN
1. Komposisi media dasar Knudson C 59
1
PENDAHULUAN
Latar Belakang
Anggrek bulan (Phalaenopsis sp.) adalah salah satu jenis anggrek yang menjadi komoditas penting di pasar dunia (Tang dan Chen 2007). Spesies anggrek bulan banyak ditemukan di Indonesia, dengan jumlah 21 (Puspitaningtyas dan Mursidawati 1999) dari 62 spesies yang teridentifikasi didunia (Christenson 2001). Spesies anggrek bulan umumnya memiliki karakter bunga dengan kualitas yang masih rendah dibandingkan anggrek bulan hibrida komersial, diantaranya bentuk kurang kompak, ukuran relatif kecil, serta sepal dan petal kurang tebal. Usaha perbaikan karakter tanaman dapat dilakukan salah satunya melalui induksi mutasi untuk menghasilkan variasi baru yang diikuti dengan seleksi. Induksi poliploidi merupakan salah satu metode induksi mutasi secara kimia yang dapat dimanfaatkan dalam menghasilkan variasi baru untuk tujuan perbaikan karakter bunga anggrek bulan spesies.
Spesies anggrek bulan berpotensi untuk dikembangkan melalui induksi poliploidi. Beberapa diantaranya adalah Phalaenopsis amabilis dan Phalaenopsis gigantea. Berdasarkan Christenson (2001), P. amabilis merupakan spesies anggrek bulan dengan ukuran bunga terbesar dalam genusnya. Petal P. amabilis
lebar dan kondisi bunganya dapat mekar secara penuh. Spesies ini banyak digunakan sebagai dasar dalam persilangan untuk menghasilkan anggrek bulan hibrida dengan bunga besar dan bulat sampai saat ini. Phalaenopsis gigantea
2
Teknik dalam poliploidisasi pada anggrek bulan dan anggrek jenis lain umumnya dilakukan melalui perlakuan menggunakan agen anti-mitosis yang ditargetkan pada sel-sel somatik dari jaringan meristematik, seperti protocorm
(Griesbach 1981; Chaicharoen dan Saejew 1981; Sarathum et al. 2010; Miguel dan Leonhardt 2011; Kerdsuwan dan Te-chato 2012; dan Atichart 2013) dan planlet (Vichiato et al. 2007 dan Rahayu 2014). Diantara percobaan-percobaan tersebut, hasil tertinggi diperoleh oleh Kerdsuwan dan Te-chato (2012), dengan persentase poliploid sebesar 60%, namun hasil tersebut juga berasosiasi dengan rendahnya persentase hidup yang hanya sebesar 26%. Induksi poliploidi pada jaringan meristematik memiliki peluang yang tinggi untuk terjadinya kimera, yaitu dalam bentuk mixoploid. Kondisi tersebut memungkinkan terjadinya perubahan yang mengarah pada recovery menjadi kondisi asalnya diploid (diplontic selection). Eigsti dan Dustin (1957) menyatakan bahwa dalam program pemuliaan tanaman yang menggunakan poliploid, kimera dan mixoploid
merupakan masalah yang penting. Induksi poliploidi yang menghasilkan kimera telah dilaporkan oleh Nilanthi et al. (2011) pada tanaman Echinacea purpurea
menggunakan eksplan tunas dan Lam et al. (2014) pada Acacia menggunakan biji. Zang et al. (2008) menemukan bahwa persentase kimera lebih rendah dengan perlakuan kolkisin yang lebih lama pada tanaman Phlox, perlakuan selama 30 hari menghasilkan miksoploid 27.8%, sedangkan selama 10 hari menghasilkan 52.6%.
Permasalahan-permasalahan dalam induksi poliploidi perlu diatasi dengan menggunakan alternatif-alternatif metode ataupun teknik yang tepat. Alternatif metode yang dapat digunakan salah satunya adalah melalui pembentukan gamet poliploid (unreduced gamet). Polen poliploid pada anggrek diploid dapat terbentuk secara spontan dengan persentase yang sangat kecil, sebagaimana yang dilaporkan oleh Teoh (1984) pada Spathoglottis plicata dan Calanthe rubens, yaitu masing-masing secara berurutan sebesar 0.63 dan 5.01%. Peningkatan peluang pembentukan gamet poliploid dapat ditingkatkan melalui induksi menggunakan agen anti-mitosis pada organ generatif muda. Informasi tentang induksi gamet poliploid pada anggrek bulan dan anggrek jenis lain masih sangat terbatas dan belum ada laporan terkait keberhasilannya. Terdapat satu laporan yang dilakukan oleh Nakasone (1960) melalui perlakuan kolkisin pada spike Vanda Miss Joaquin dan tidak diperoleh individu poliploid. Tanpa melihat peluang keberhasilannya yang kecil, percobaan induksi gamet poliploid memiliki nilai tambah yang besar dalam peningkatan program pemuliaan anggrek bulan. Peluang keberhasilannya yang kecil sangat mungkin untuk diatasi melalui teknik-teknik baru, seperti penggunaan organ generatif muda pada beberapa fase perkembangan sebagai material untuk perlakuan induksi menggunakan agen anti-mitosis. Teknik baru yang digunakan dalam penelitian ini terletak pada penggunaan bunga setelah penyerbukan (pollinated flower) sebagai bahan perlakuan kolkisin. Organ generatif lain yang digunakan adalah kuncup bunga dan
3
protocorm, serta untuk mendapatkan teknik yang efektif dan efisien dalam menghasilkan anggrek bulan poliploid dalam jumlah banyak.
Perumusan Masalah
Poliploidi merupakan salah satu bentuk mutasi jumlah kromosom yang banyak dimanfaatkan untuk perbaikan karakter penting anggrek bulan. Kompensasi dari perubahan poliploid adalah sifat gigantisme yang sering diminati para penggemar anggrek, khususnya pada bunga. Namun, permasalahan utama yang sering dihadapi dalam memperoleh mutan poliploid, salah satunya adalah kimera seperti mixoploid. Kondisi tersebut merupakan masalah besar karena dapat menghasilkan perubahan poliploid yang sementara akibat diplontic selection.
Selain permasalahan yang telah disebutkan sebelumnya, metode yang efektif dalam menghasilkan anggrek bulan poliploid juga masih terbatas, khususnya terkait dengan variasi penggunaan jenis material dan tekniknya. Penggunaan jenis material sebagai target perlakuan agen anti-mitosis merupakan salah satu aspek yang sangat penting sebagai penentu keberhasilan induksi poliploidi. Pertimbangan dalam penggunaan material didasarkan pada seberapa besar tingkat sel dari suatu jaringan yang belum terdiferensiasi, semakin kecil tingkat diferensiasinya akan meminimalisir hasil poliploid yang bersifat kimera. Sebagian besar jenis material yang digunakan dalam induksi poliploidi pada anggrek adalah protocorm. Kelemahan dalam penggunaan material ini adalah tingginya persentase kematian akibat toksisitas mutagen, sehingga hanya sedikit hasil poliploid yang diperoleh. Selain itu masih terdapat peluang hasil perubahan yang bersifat kimera. Pendekatan metode yang dapat dilakukan untuk mengurangi atau mengeliminasi permasalahan dalam induksi poliploidi salah satunya melalui induksi gamet poliploid. Jenis material yang digunakan dalam induksi gamet poliploid adalah organ generatif yang diberi perlakuan menggunakan agen anti-mitosis seperti kolkisin. Penggabungan gamet poliploid antara jantan dan betina melalui fertilisasi akan menghasilkan individu poliploid yang stabil sampai pada generasi berikutnya.
Tujuan Penelitian
1. Mempelajari pengaruh kolkisin terhadap perkembangan tiga jenis organ generatif P. amabilis, yaitu bunga setelah penyerbukan, kuncup bunga dan
spike, serta terhadap protocorm P. gigantea
2. Mempelajari potensi induksi poliploidi berdasarkan karakter morfologi melalui perlakuan kolkisin pada organ generatif P. amabilis
3. Mendapatkan planlet poliploid melalui perlakuan kolkisin pada organ generatif P. amabilis berdasarkan uji sitologi
4
Hipotesis
1. Kolkisin berpengaruh terhadap perkembangan tiga jenis organ generatif P. amabilis, yaitu bunga setelah penyerbukan, kuncup bunga dan spike dalam membentuk buah serta terhadap kemampuan regenerasi protocorm P. gigantea
2. Terdapat karakter morfologi yang berbeda dari planlet hasil perlakuan kolkisin pada organ generatif P. amabilis
3. Panlet poliploid dapat diperoleh melalui perlakuan kolkisin pada bunga setelah penyerbukan dan kuncup bunga dari P. amabilis
4. Teknik perlakuan kolkisin pada organ generatif P. amabilis berpengaruh terhadap keberhasilan induksi poliploidi
Manfaat Penelitian
Inovasi dalam induksi poliploidi yang menggunakan organ generatif untuk menghasilkan sel gamet poliploid akan menjadi dasar untuk pengembangan secara lebih cepat varian-varian baru anggrek bulan melalui penyerbukan sendiri atau silang. Efisiensi waktu proses pemuliaan anggrek bulan untuk menghasilkan
progeny triploid melalui persilangan dengan gamet haploid. Anggrek bulan poliploid dari biji yang dihasilkan dalam jumlah banyak memfasilitasi tahapan seleksi dengan lebih baik untuk memperoleh calon varietas unggul.
Ruang Lingkup Penelitian
Penelitian yang dilakukan meliputi beberapa kegiatan sebagai suatu tahapan yang berbeda untuk mencapai tujuan akhir yang diharapkan. Beberapa tahapan dalam penelitian ini adalah: Percobaan 1. Induksi poliploidi pada anggrek bulan (Phalaenopsis amabilis (L.) Blume) melalui aplikasi kolkisin pada bunga setelah penyerbukan. Percobaan 2. Potensi induksi poliploidi anggrek bulan (Phalaenopsis amabilis (L.) Blume) melalui perlakuan kolkisin pada kuncup bunga. Percobaan 3. Pengaruh perlakuan kolkisin pada perkembangan spike
5
Gambar 1 Bagan alir penelitian Induksi Poliploidi Anggrek Bulan (Phalaenopsis sp.) Menggunakan Kolkisin pada Organ Generatif dan
Protocorm
Lapang
Induksi poliploidi organ generatif
Buah anggrek
In vitro
1.Bunga setelah penyerbukan
Induksi poliploidi protocorm
4.Protocorm
Konfirmasi jumlah ploidi secara sitologi
Planlet poliploid Perlakuan kolkisin
Pengecambahan biji in vitro Planlet 3.Spike
2.Kuncup bunga
Screening planlet diduga poliploid secara morfologi
2
INDUKSI POLIPLOIDI PADA ANGGREK BULAN
(
Phalaenopsis amabilis
(L.) Blume) MELALUI APLIKASI
KOLKISIN PADA BUNGA SETELAH PENYERBUKAN
Abstrak
Aplikasi kolkisin pada bunga setelah penyerbukan Phalaenopsis amabilis
bertujuan untuk mengetahui pengaruh kolkisin terhadap pembentukan buah dan keberhasilan induksi planlet poliploid yang dihasilkan dari buah. Konsentrasi kolkisin yang digunakan dalam percobaan adalah 0, 50, 500, 1000, dan 2000 mg L-1, dan durasi penyungkupan dengan aluminium foil selama perlakuan kolkisin adalah dua dan tiga hari tersarang pada konsentrasi kolkisin (Nested Design). Hasil percobaan menunjukkan bahwa konsentrasi kolkisin yang tinggi menghambat panjang buah dan persentase jumlah buah. Skrining terhadap planlet yang dihasilkan dari buah asal perlakuan kolkisin telah menghasilkan tipe planlet normal (N) dan tipe planlet diduga poliploid (DP). Tipe planlet DP dari semua perlakuan kolkisin berbeda nyata dengan kontrol untuk karakter panjang dan lebar
basal organ of the protocorm (BOP), panjang daun, serta panjang dan diameter akar. Pengujian jumlah kromosom secara sitologi terhadap tipe planlet DP menunjukkan bahwa 60.0% dari masing-masing perlakuan kolkisin 50 mg L-1 selama tiga hari dan lima hari, dan 100.0% dari perlakuan kolkisin 500 mg L-1 selama lima hari adalah planlet tetraploid. Hasil tersebut membuktikan keberhasilan induksi poliploidi pada fase perkembangan zigot ataupun embrio pada P. amabilis.
Kata kunci: jumlah kromosom, konsentrasi kolkisin, pembentukan buah, poliploidisasi
POLYPLOIDY INDUCTION OF MOTH ORCHID (
Phalaenopsis
amabilis
(L.) Blume) BY COLCHICINE TREATMENT ON
POLLINATED FLOWER
Abstract
7
to control (0 mg L-1) for characters of basal organ of the protocorm (BOP) length and width, leaf length, and root length and diameter. The PPS chromosome number analysis revealed that 50 mg L-1 with three or five days colchicine application and 500 mg L-1 with five days colchicine application produced 60.0 and 100.0% tetraploid mutant seedlings, respectively. This result indicated that polyploidy induction occurred during zygot or embryo development.
Keywords: chromosome number, colchicine concentration, pod formation, polyploidization
Pendahuluan
Salah satu spesies anggrek bulan yang memiliki potensi untuk dikembangkan sebagai varietas komersial adalah Phalaenopsis amabilis. Spesies anggrek bulan ini tersebar di Indonesia sampai bagian selatan Filipina, serta Papua Nugini bagian timur dan Queensland, Australia (Christenson 2001). Di Indonesia,
P. amabilis yang berasal dari Cianjur Selatan, Jawa Barat, tumbuh diwilayah dataran rendah dan sangat potensial sebagai dasar untuk pengembangan varietas anggrek bulan dataran rendah. Perbaikan terhadap beberapa karakter bunganya diperlukan agar mampu bersaing dengan anggrek bulan varietas komersial, terutama bentuk dan ukurannya. Induksi poliploidi diharapkan mampu menjadi solusi untuk pengembangan P. amabilis. Menurut Griesbach (1985) perbaikan karakter perlu dilakukan pada anggrek bulan spesies yang penting seperti P. amabilis, yang merupakan salah satu spesies yang sering digunakan sebagai tetua persilangan untuk menghasilkan hibrida-hibrida dengan karakter bunga putih standar besar (the standard big flower). Poliploidi memiliki peran yang penting dalam perbaikan tanaman hibrida ataupun spesies. Sektor florikultura merupakan yang paling diuntungkan karena poliploidi dapat meningkatkan keragaman genetik, ukuran bunga serta memperbaiki bentuknya.
Meskipun memiliki manfaat yang sangat besar dalam perbaikan anggrek, teknik ataupun protokol induksi poliploidi yang efektif dan mampu menghasilkan individu poliploid dalam jumlah banyak masih belum tersedia atau sangat terbatas. Efektivitas induksi poliploidi salah satunya ditentukan oleh penggunaan jenis bahan tanaman untuk perlakuan kolkisin. Penggunaan bunga setelah penyerbukan sebagai bahan untuk perlakuan kolkisin pada anggrek belum pernah dilakukan. Potensi penggunaan bunga setelah penyerbukan sebagai bahan untuk induksi poliploidi sangat besar karena yang menjadi target adalah zigot ataupun embrio yang terbentuk setelah terjadi fertilisasi. Zigot terdiri dari satu sel yang kemudian akan mengalami pembelahan mitosis membentuk embrio, jika terjadi penggandaan kromosom selama proses pembelahan mitosis tersebut maka akan dihasilkan biji anggrek yang bersifat poliploid. Kemungkinan terjadinya poliploid yang bersifat kimera sangat kecil karena yang terinduksi adalah zigot ataupun embrio yang memiliki jumlah sel yang masih sedikit.
8
atau telofase, dimana kromosom tersebar dan tidak mengarah pada dua kutub yang berlawanan. Benang gelendong (spindle fiber) merupakan substrat yang dipengaruhi oleh aktivitas kolkisin. Molekul kolkisin bereaksi dengan molekuler sistem dari benang gelendong dianggap sebagai dasar hubungan anatara keduanya. Mekasnisme penghambatan atau destruksi terhadap benang gelendong akan terlihat sebagai reaksi kuantitatif, karena konsentrasi kolkisin merupakan faktor yang kritikal (Eigsti dan Dustin 1957).
Keunggulan lain dalam induksi poliploidi pada bunga setelah penyerbukan adalah dapat dihasilkannya planlet poliploid dalam jumlah yang banyak. Buah anggrek memiliki biji dalam jumlah yang sangat banyak, dengan kisaran mulai dari 1300 sampai 4 000 000 biji per buah (Arditti 1992). Selain itu, penggunaan bunga dapat dimanipulasi melalui penyerbukan silang ataupun sendiri untuk meningkatkan keragaman planlet poliploid yang dihasilkan. Bunga setelah penyerbukan dari P. amabilis digunakan dalam penelitian ini sebagai bahan untuk perlakuan kolkisin. Penelitian ini bertujuan untuk mempelajari pengaruh kokisin terhadap bunga setelah penyerbukan serta untuk mendapatkan planlet poliploid dalam jumlah banyak melalui induksi poliploidi pada zigot ataupun embrio.
Bahan dan Metode
Penelitian ini dilakukan di tempat penangkar anggrek di kawasan Puncak, Kabupaten Bogor, dan di Laboratorium Kultur Jaringan Tanaman Departemen Agronomi dan Hortikultura, Fakultas Pertanian IPB pada bulan Februari 2013 sampai Juli 2014. Bahan tanaman yang digunakan adalah P. amabilis yang sedang berbunga asal Jawa Barat. Bahan lain adalah kolkisin, komposisi media Knudson C, HCl 1 N, asam asetat glasial, dan orcein 45%. Alat yang digunakan adalah aluminium foil, kapas, dan mikroskop cahaya.
Bunga yang sedang mekar dilakukan penyerbukan sendiri buatan (selfing). Kastrasi atau pembuangan sepal dan petal, pembungkusan bunga dengan kapas, dan perlakuan larutan kolkisin dilakukan pada hari ketiga setelah penyerbukan buatan. Konsentrasi larutan kolkisin yang digunakan adalah 0, 50, 500, 1000, dan 2000 mg L-1. Bunga yang telah diberi perlakuan kolkisin disungkup menggunakan aluminium foil dengan durasi tiga dan lima hari. Penelitian disusun secara RAL, dengan durasi penyungkupan tersarang pada setiap taraf kolkisin. Setiap perlakuan kolkisin dilakukan pada empat tanaman (empat ulangan). Proses penyerbukan bunga P. amabilis disajikan pada Gambar 2 dan bunga setelah penyerbukan dan kastrasi pada Gambar 3.
9 portable dan uji lanjut Duncan untuk rata-rata jumlah daun dan akar. Analisis karakter-karakter tipe planlet diduga mutan poliploidi dan tipe planlet normal dari setiap perlakuan kolkisin seperti: panjang dan lebar basal organ of the protocorm
(BOP), daun dan stomata, serta panjang dan diameter akar dilakukan dengan Kontras Ortogonal. Pengujian jumlah kromosom secara sitologi pada ujung akar planlet dilakukan setelah umur 16 minggu setelah subkultur (MSSk) dari media pembesaran.
Gambar 3 Bunga P. amabilis tiga hari setelah penyerbukan dan pembuangan sepal dan petal (kastrasi)
Gambar 2 Tahapan penyerbukan sendiri buatan pada anggrek bulan
Pengambilan polinia Polinia dipindah
dari rostellum
Polinia dimasukan ke dalam column
10
Hasil dan Pembahasan
Terdapat beberapa perubahan karakteristik bunga pada tiga hari setelah penyerbukan sendiri secara buatan, yaitu stigma (column) mulai menutup. Bunga tersebut mampu berkembang menjadi buah pada kontrol dan setiap perlakuan kolkisin. Namun, terdapat beberapa buah dari perlakuan kolkisin yang tidak berkembang dan mengalami kerontokan atau kering sebelum mencapai umur panen sekitar 24 MSP. Kondisi buah anggrek pada berbagai konsentrasi dan durasi perlakuan kokisin disajikan pada Gambar 4.
Perlakuan kolkisin dengan konsentrasi yang lebih tinggi menghambat panjang buah P. amabilis pada 16 minggu setelah perlakuan (MSPr). Beberapa buah rontok sebelum mencapai umur panen, seperti pada perlakuan kolkisin 1000 dan 2000 mg L-1. Nakasone (1960) melaporkan pengaruh perlakuan kolkisin terhadap kerontokan bunga anggrek pada konsentrasi 1000 sampai 20000 mg L-1. Wu et al. (2007) juga menemukan bahwa perlakuan kolkisin pada tunas bunga muda dari Lilium (non anggrek) dengan konsentrasi kolkisin 2000 mg L-1 menyebabkan kerontokan bunga hingga 81.30% pada varietas Con.Amore.
Gambar 4 Keragaan buah P. amabilis umur 8 minggu setelah perlakuan (MSPr) dari perlakuan kolkisin pada bunga setelah penyerbukan pada berbagai konsentrasi kolkisin: a. 0, b. 50, c. 500, d. 1000, dan e. 2000 mg L-1
11 Hasil analisis ragam menunjukkan bahwa persentase jumlah buah tidak dipengaruhi oleh durasi penyungkupan, tetapi secara nyata dipengaruhi oleh konsentrasi kolkisin. Pengaruh konsentrasi kolkisin terhadap persentase buah yang dihasilkan dari setiap perlakuan disajikan dalam Gambar 5. Konsentrasi kolkisin yang lebih tinggi menurunkan persentase jumlah buah. Kondisi tersebut menunjukkan bahwa kolkisin pada konsentrasi yang tinggi, khususnya 1000 dan 2000 mg L-1, bersifat toksik terhadap jaringan tanaman. Toksisitas kolkisin tersebut dapat menyebabkan sel-sel pada jaringan mati sehingga buah anggrek yang terbentuk menjadi kering dan rontok. Semua buah P. amabilis dari kontrol (sebelas buah) menghasilkan biji yang bisa berkecambah menjadi protocorm
secara in vitro pada media Knudson C. Buah yang diperoleh dari perlakuan kolkisin tidak semuanya mampu menghasilkan biji yang bisa berkecambah menjadi protocorm, hanya sembilan buah (36.00%) dari 25 buah asal perlakuan kolkisin yang menghasilkan protocorm.
Buah yang mampu menghasilkan protocorm diantaranya diperoleh dari perlakuan kolkisin 50 mg L-1 selama tiga dan lima hari, 500 mg L-1 selama lima hari, dan 1000 mg L-1 selama tiga hari (Tabel 1). Perlakuan kolkisin pada bunga setelah penyerbukan dengan konsentrasi yang lebih tinggi telah menurunkan persentase buah yang mampu menghasilkan protocorm. Perlakuan kolkisin 500 mg L-1 selama tiga hari, 1000 mg L-1 selama lima hari, dan 2000 mg L-1 selama tiga dan lima hari, bahkan tidak menghasilkan satupun buah dengan biji yang mampu berkecambah menjadi protocorm sampai umur 12 minggu setelah semai (MSS). Perkecambahan biji anggrek umumnya ditandai dengan perkembangan biji menjadi berbentuk bulat dan berwarna hijau, namun biji anggrek bulan dari perlakuan-perlakuan tersebut tidak menunjukkan perubahan yang mengarah pada perkecambahan menjadi protocorm. Kondisi tersebut dapat disebabkan oleh dua kemungkinan, yaitu tidak terbentuk biji didalam buah dan atau terbentuk biji didalam buah namun tidak viabel.
Gambar 5 Pengaruh konsentrasi kolkisin terhadap persentase buah jadi hasil perlakuan kolkisin pada bunga setelah penyerbukan P. amabilis. Huruf berbeda menunjukkan perbedaan nyata berdasarkan uji Duncan pada taraf 5%.
a
ab ab
bc c 0 20 40 60 80 100 120
0 50 500 1000 2000
P er sent a se jum la h bu a h ja di ( %)
12
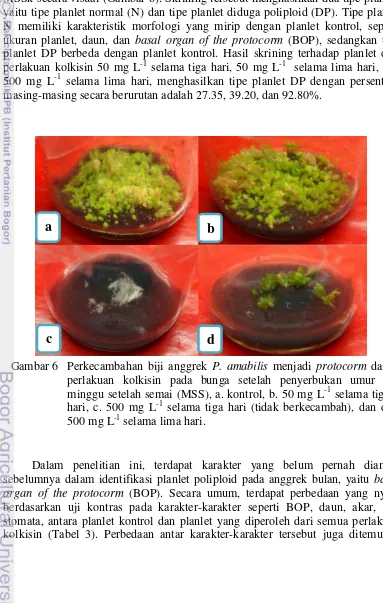
Terdapat perbedaan kisaran jumlah protocorm per buah dari setiap perlakuan kolkisin pada bunga setelah penyerbukan. Kisaran jumlah protocorm
perbuah tersebut dikelompokan menjadi tiga kategori, yaitu banyak (protocorm>250), sedang (250>protocorm>50), dan sedikit (protocorm<50). Perkecambahan biji membentuk protocorm disajikan pada Gambar 6. Panjang buah yang lebih kecil dari kontrol, pada perlakuan kolkisin 500 mg L-1 selama lima hari dan 1000 mg L-1 selama tiga hari menghasilkan kisaran jumlah
protocorm perbuah pada kategori sedang dan sedikit, hanya tiga protocorm saja dari perlakuan 1000 mg L-1 selama tiga hari. Perlakuan kolkisin 50 mg L-1 selama tiga dan lima hari yang menghasilkan ukuran buah setara dengan kontrol, memiliki kisaran jumlah protocorm perbuah pada kategori banyak. Nakasone (1960) menemukan bahwa perlakuan kolkisin lebih dari empat hari pada biji beberapa jenis anggrek menyebabkan gangguan pada perkecambahannya. Hasil yang sama juga dilaporkan pada tanaman lain oleh Takamura dan Miyajima (1996) pada benih Cyclamen, dan Liu et al. (2007) pada benih Platanus acerifolia.
Planlet P. amabilis memperlihatkan pertumbuhan yang baik dan mampu menghasilkan daun dan akar selama inkubasi di ruang kultur sampai 16 MSSk. Selain itu, planlet juga membentuk protocorm like bodies (plbs). Semua planlet tumbuh normal kecuali pada perlakuan kolkisin 500 mg L-1 lima hari, yaitu terdapat satu protocorm abnormal (Gambar 7). Protocorm abnormal tersebut tidak menghasilkan daun sampai akhir pengamatan. Penelitian Rahayu (2014) juga memperoleh planlet abnormal dari perlakuan kolkisin pada protocorm P. amabilis
dan P. amboinensis, planlet menjadi lebih pendek dan daun lebih tebal dari diploidnya. Sarathum et al. (2010) juga menemukan planlet sitokimera dengan morfologi yang abnormal.
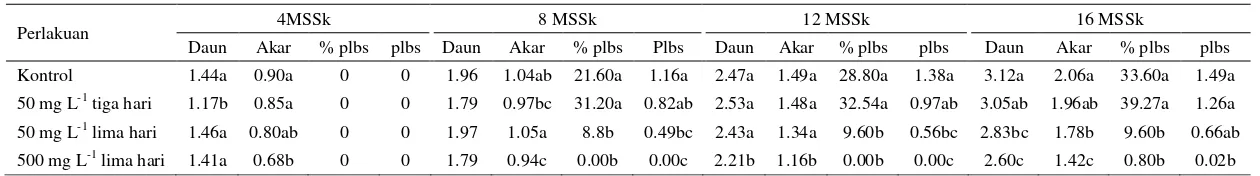
Planlet P. amabilis menunjukkan perbedaan nyata pada rata-rata jumlah daun, akar, plbs, dan persentase planlet membentuk plbs baik masing-masing Tabel 1 Jumlah bunga pada perlakuan dan beberapa karakter buah yang dihasilkan
dari perlakuan kolkisin pada bunga setelah penyerbukan P. amabilis
Kolkisin
(mg L-1) Durasi
Jumlah bunga diberi perlakuan Jumlah buah(a) Rata-rata panjang buah (cm) Jumlah buah menghasilkan protocorm(a)
Kisaran jumlah protocorm per
buah
0 3 Hari 4 4/4 (100.00) 5.12 4 (100.00) Banyak (protocorm>250)
0 5 Hari 7 7/7 (100.00) 4.92 7 (100.00) Banyak (protocorm>250)
50 3 Hari 5 3/5 (60.00) 5.26 3 (100.00) Banyak (protocorm>250)
50 5 Hari 7 4/7 (57.14) 4.52 3 (75.00) Banyak (protocorm>250)
500 3 Hari 7 5/7 (71.42) 3.52 0 (0.00) Tidak ada
500 5 Hari 7 6/7 (85.71) 3.68 2 (33.33) Sedang (250>protocorm>50)
1000 3 Hari 7 4/7 (57.14) 2.80 1 (25.00) Sedikit (protocorm<50)
1000 5 Hari 7 2/7 (28.57) 2.60 0 (0.00) Tidak ada
2000 3 Hari 5 1/5 (20.00) 2.00 0 (0.00) Tidak ada
2000 5 Hari 4 0/4 (0.00) 0.00 0 (0.00) Tidak ada
Keterangan: Rata-rata panjang buah diamati sekitar 16 MSP
Pengamatan pembentukan protocorm dilakukan pada 8 MSS
(a)
13 perlakuan kolkisin ataupun dengan kontrol (Tabel 2). Planlet dari perlakuan kolkisin 500 mg L-1 selama lima hari pada 16 MSSk memiliki nilai yang lebih kecil untuk rata-rata jumlah daun dan akar yaitu masing-masing adalah 0.83 dan 0.68 kali dari planlet kontrol. Planlet ini juga menghasilkan rata-rata dan persentase planlet membentuk plbs yang paling sedikit diantara perlakuan lain dan kontrol.
Skrining terhadap planlet diduga poliploid dilakukan dengan cara membandingkan planlet kontrol dengan planlet hasil perlakuan kolkisin pada 16 MSSk secara visual (Gambar 8). Skrining tersebut menghasilkan dua tipe planlet, yaitu tipe planlet normal (N) dan tipe planlet diduga poliploid (DP). Tipe planlet N memiliki karakteristik morfologi yang mirip dengan planlet kontrol, seperti ukuran planlet, daun, dan basal organ of the protocorm (BOP), sedangkan tipe planlet DP berbeda dengan planlet kontrol. Hasil skrining terhadap planlet dari perlakuan kolkisin 50 mg L-1 selama tiga hari, 50 mg L-1 selama lima hari, dan 500 mg L-1 selama lima hari, menghasilkan tipe planlet DP dengan persentase masing-masing secara berurutan adalah 27.35, 39.20, dan 92.80%.
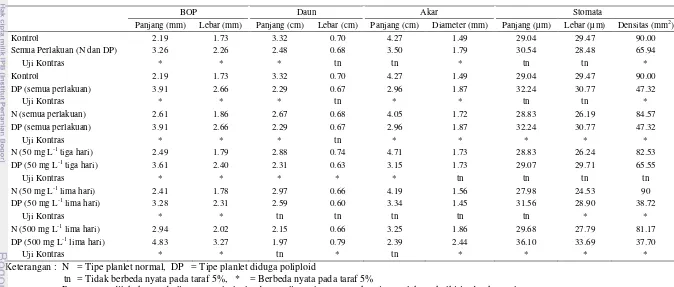
Dalam penelitian ini, terdapat karakter yang belum pernah diamati sebelumnya dalam identifikasi planlet poliploid pada anggrek bulan, yaitu basal organ of the protocorm (BOP). Secara umum, terdapat perbedaan yang nyata berdasarkan uji kontras pada karakter-karakter seperti BOP, daun, akar, dan stomata, antara planlet kontrol dan planlet yang diperoleh dari semua perlakuan kolkisin (Tabel 3). Perbedaan antar karakter-karakter tersebut juga ditemukan
Gambar 6 Perkecambahan biji anggrek P. amabilis menjadi protocorm dari perlakuan kolkisin pada bunga setelah penyerbukan umur 8 minggu setelah semai (MSS), a. kontrol, b. 50 mg L-1 selama tiga hari, c. 500 mg L-1 selama tiga hari (tidak berkecambah), dan d. 500 mg L-1 selama lima hari.
a
c
d
[image:31.595.110.493.204.807.2]14
pada tipe planlet N dan tipe planlet DP dari semua perlakuan kolkisin. Analisis terhadap semua perlakuan kolkisin menunjukkan bahwa tipe planlet DP memiliki panjang dan lebar BOP yang lebih besar sekitar 1.5 kali dibandingkan tipe planlet N. Hasil yang sama juga diperoleh pada masing-masing perlakuan kolkisin 50 mg L-1 selama tiga hari, 50 mg L-1 selama lima hari, dan 500 mg L-1 selama lima hari. Panjang dan lebar stomata tipe planlet DP berbeda nyata dengan nilai yang lebih besar dari tipe planlet N, pada semua perlakuan kolkisin dan perlakuan kolkisin 500 mg L-1 selama lima hari. Panjang dan lebar stomata dengan nilai yang lebih besar telah dilaporkan juga pada planlet tetraploid anggrek, yaitu oleh Chaicharoen dan Saejew (1981), Chen et al. (2009), Kerdsuwan dan Te-chato (2012), dan Rahayu (2014), begitu juga pada tanaman non anggrek seperti
Anthurium oleh Chen et al. (2011a) dan jeruk siam madu (Citrus nobilis) oleh Yulianti et al. (2014).
Karakter densitas stomata dari tipe planlet DP pada semua perlakuan kolkisin memiliki perbedaan yang nyata dengan nilai yang lebih kecil, yaitu 0.52 kali dari planlet kontrol, begitu juga ketika dibandingkan dengan tipe planlet N, yaitu 0.55 lebih kecil. Densitas stomata dengan nilai yang lebih kecil pada anggrek tetraploid juga dilaporkan oleh Chen et al (2009), yaitu pada P. aphrodite
dan P. hieroglyphica, begitu juga oleh Yang et al (2006) pada Vitis vinifera (non anggrek). Berbeda dengan Kerdsuwan dan Te-chato (2012), dimana densitas stomata Rhynchostylis tetraploid memiliki nilai yang lebih tinggi dari diploid.
[image:32.595.85.486.105.674.2]Tipe planlet N dan tipe planlet DP dari setiap perlakuan kolkisin diuji jumlah kromosomnya secara sitologi menggunakan mikroskop cahaya dengan perbesaran 100x. Berdasarkan hasil pengujian sitologi, semua tipe planlet N dari setiap perlakuan kolkisin adalah 100.0% diploid (2n=2x=38). Hasil analisis sitologi juga menunjukkan bahwa perlakuan kolkisin 50 mg L-1 selama tiga dan
15 lima hari menghasilkan persentase planlet poliploid masing-masing sebesar 60.0%, dari keseluruhan tipe planlet DP yang diuji. Hasil pengujian tertinggi diperoleh dari perlakuan kolkisin 500 mg L-1 selama lima hari, dengan semua tipe planlet DP teridentifikasi sebagai planlet poliploid (100.0%) (Tabel 4). Tingkat ploidi dari planlet poliploid yang diperoleh secara keseluruhan menunjukkan kondisi tetraploid. Tiga planlet yang dihasilkan dari perlakuan kolkisin 1000 mg L-1 tidak menunjukkan poliploid setelah diuji sitologi. Perbandingan BOP, ukuran dan densitas stomata, dan jumlah kromosom disajikan pada Gambar 9. Penelitian yang hampir sama, yaitu perlakuan kolkisin pada spike telah dilakukan oleh Nakasone (1960), namun tidak diperoleh anggrek poliploid. Induksi poliploidi dengan kolkisin pada protocorm anggrek yang dilakukan oleh Chaicharoen dan Saejew (1981) dan Griesbach (1981) menghasilkan sekitar 50.0% mutan tetraploid. Sarathum et al. (2010) dan Kerdsuwan dan Te-chato (2012) menghasilkan persentase mutan tetraploid anggrek masing-masing sebesar 43.0 dan 60.0%. Induksi poliploidi pada tanaman lain diperoleh 50.0% poliploid pada Platycodon grandiflorus (Wu et al. 2011).
Proses penyerbukan diawali dengan jatuhnya polinia ke permukaan stigma yang berlendir didalam column. Polen yang berada didalam polinia kemudian berkecambah dan membentuk tabung polen. Sebagian besar polen anggrek memiliki satu inti vegetatif dan satu sel gamet (Arditti 1992). Tabung polen yang membawa inti vegetatif dan sel gamet jantan kemudian masuk kedalam embryo sac melalui microphyl. Embryo sac memiliki delapan inti yang terdiri dari tiga sel antipodal, dua inti polar, dua sel sinergid, dan satu sel gamet betina. Fertilisasi kemudian terjadi melalui peleburan antara sel gamet jantan dan gamet betina yang berada didekat microphyl, sehingga membentuk satu sel zigot. Sel gamet jantan lain dari polen kemudian membuahi dua inti polar didalam embryo sac yang kemudian mengalami proliferasi membentuk endosperm. Biji anggrek tidak memiliki endosperm, akibat proses peleburan tiga inti yang terdiri dari dua inti polar dan satu sel gamet jantan (inti generatif) mengalami degenerasi (Niimoto dan Sagawa 1962). Zigot akan mengalami pembelahan mitosis sampai terbentuk embrio yang masak.
16
Tabel 2 Rata-rata jumlah daun, akar, protocorm like bodies (plbs) dan persentase planlet membentuk plbs dari planlet hasil perlakuan kolkisin pada bunga setelah penyerbukan P. amabilis dan kontrol
Perlakuan 4MSSk 8 MSSk 12 MSSk 16 MSSk
Daun Akar % plbs plbs Daun Akar % plbs Plbs Daun Akar % plbs plbs Daun Akar % plbs plbs
Kontrol 1.44a 0.90a 0 0 1.96 1.04ab 21.60a 1.16a 2.47a 1.49a 28.80a 1.38a 3.12a 2.06a 33.60a 1.49a
50 mg L-1 tiga hari 1.17b 0.85a 0 0 1.79 0.97bc 31.20a 0.82ab 2.53a 1.48a 32.54a 0.97ab 3.05ab 1.96ab 39.27a 1.26a
50 mg L-1 lima hari 1.46a 0.80ab 0 0 1.97 1.05a 8.8b 0.49bc 2.43a 1.34a 9.60b 0.56bc 2.83bc 1.78b 9.60b 0.66ab
500 mg L-1 lima hari 1.41a 0.68b 0 0 1.79 0.94c 0.00b 0.00c 2.21b 1.16b 0.00b 0.00c 2.60c 1.42c 0.80b 0.02b
Keterangan: Angka yang diikuti huruf berbeda pada kolom yang sama menunjukkan perbedaan nyata berdasarkan uji Duncan pada taraf 5%. Pengamatan dilakukan pada 125 sampel planlet setiap perlakuan kolkisin dan kontrol, kecuali perlakuan kolkisin 50 mg L-1 tiga hari yang terdapat 117 sampel planlet mulai 8 MSSk
Gambar 8 Perbandingan morfologi tipe planlet diduga poliploid (DP) dan tipe planlet normal (N) hasil perlakuan kolkisin pada bunga setelah penyerbukan, dan planlet kontrol pada 16 minggu setelah subkutur (MSSk)
[image:34.842.239.606.292.474.2]17
Tabel 3 Hasil uji kontras ortogonal pada ukuran basal organ of the protocorm (BOP), daun, akar, dan stomata dari planlet hasil perlakuan kolkisin pada bunga setelah penyerbukan P. amabilis dan kontrol pada 16 minggu setelah subkultur (MSSk)
BOP Daun Akar Stomata
Panjang (mm) Lebar (mm) Panjang (cm) Lebar (cm) Panjang (cm) Diameter (mm) Panjang (µm) Lebar (µm) Densitas (mm2)
Kontrol 2.19 1.73 3.32 0.70 4.27 1.49 29.04 29.47 90.00
Semua Perlakuan (N dan DP) 3.26 2.26 2.48 0.68 3.50 1.79 30.54 28.48 65.94
Uji Kontras * * * tn tn * tn tn *
Kontrol 2.19 1.73 3.32 0.70 4.27 1.49 29.04 29.47 90.00
DP (semua perlakuan) 3.91 2.66 2.29 0.67 2.96 1.87 32.24 30.77 47.32
Uji Kontras * * * tn * * tn tn *
N (semua perlakuan) 2.61 1.86 2.67 0.68 4.05 1.72 28.83 26.19 84.57
DP (semua perlakuan) 3.91 2.66 2.29 0.67 2.96 1.87 32.24 30.77 47.32
Uji Kontras * * * tn * * * * *
N (50 mg L-1 tiga hari) 2.49 1.79 2.88 0.74 4.71 1.73 28.83 26.24 82.53
DP (50 mg L-1 tiga hari) 3.61 2.40 2.31 0.63 3.15 1.73 29.07 29.71 65.55
Uji Kontras * * * * * tn tn tn tn
N (50 mg L-1 lima hari) 2.41 1.78 2.97 0.66 4.19 1.56 27.98 24.53 90
DP (50 mg L-1 lima hari) 3.28 2.31 2.59 0.60 3.34 1.45 31.56 28.90 38.72
Uji Kontras * * tn tn tn tn tn * *
N (500 mg L-1 lima hari) 2.94 2.02 2.15 0.66 3.25 1.86 29.68 27.79 81.17
DP (500 mg L-1 lima hari) 4.83 3.27 1.97 0.79 2.39 2.44 36.10 33.69 37.70
Uji Kontras * * tn * tn * * * *
Keterangan : N = Tipe planlet normal, DP = Tipe planlet diduga poliploid
tn = Tidak berbeda nyata pada taraf 5%, * = Berbeda nyata pada taraf 5%
Pengamatan dilakukan pada lima sampel planlet dengan lima ulangan untuk setiap perlakuan kolkisin dan kontrol
18
[image:36.595.87.477.75.716.2]19
Tabel 4 Hasil pengujian jumlah kromosom secara sitologi pada planlet hasil perlakuan kolkisin pada bunga setelah penyerbukan
Perlakuan Tipe Planlet Jumlah sampel Diploid Tetraploid % Tetraploid
Kontrol N 5 5 0 0
50 mg L-1 tiga hari N 5 5 0 0
DP 5 2 3 60
50 mg L-1 lima hari N 5 5 0 0
DP 5 2 3 60
500 mg L-1 lima hari N 5 5 0 0
DP 5 0 5 100
Keterangan: N = Tipe planlet normal, DP = Tipe planlet diduga mutan poliploidi
[image:37.595.116.507.57.370.2]Pengamatan jumlah kromosom dilakukan pada planlet setelah umur 16 minggu setelah subkultur (MSSk)
Gambar 10 Perbandingan jumlah kromosom dari sel somatik ujung akar dari planlet poliploid dan tipe planlet normal (N) hasil perlakuan kolkisin pada bunga setelah penyerbukan P. amabilis dengan konsentrasi 50 mg L-1 selama tiga (a dan b) dan lima hari (c dan d)
a
c
Planlet poliploid Planlet normal
b
[image:37.595.110.508.70.747.2]20
Gambar 11 Mekanisme fertilisasi dan induksi poliploidi perlakuan kolkisin pada bunga P. amabilis setelah penyerbukan. Tahap I menunjukkan tabung polen mulai memasuki embryo sac melalui
microphyl. Tahap II inti vegetatif dan dua gamet jantan masuk embrio sac untuk memulai fertilisasi ganda, diikuti degenerasi inti vegetatif, dua sel sinergid, dan tiga sel antipodal. Tahap III fertilisasi terjadi antara sel gamet betina dan gamet jantan membentuk zigot (satu sel), dan induksi poliploidi oleh kolkisin mulai terjadi. Tahap IV zigot membelah sebanyak empat kali dan membentuk pro embrio (16 sel). Tahap V embrio masak mulai terbentuk. Tahap I sampai IV berlangsung mulai tiga hari sampai delapan hari setelah penyerbukan, dan perlakuan kolkisin berada pada rentang lima sampai delapan hari setelah penyerbukan
I II
III IV
V
AP = sel antipodal IP = inti polar S = sel sinergid GB = sel gamet betina IV = inti vegetatif GJ = gamet jantan IPGJ = penggabungan
dua inti polar dengan satu gamet jantan
ZG = zigot
Fase globular
Fase hati
Fase torpedo
Fase kotiledonari Kolkisin
K
o
lki
21 Menurut hasil penelitian yang dilakukan oleh Niimoto dan Sagawa (1962),
ovarium dari bunga anggrek bulan relatif belum berkembang dan bakal biji (ovul) masih belum terdiferensiasi ketika bunga memasuki masa anthesis. Penyerbukan dan pertumbuhan tabung polen menstimulasi pertumbuhan plasenta dan diferensiasi sampai akhirnya membentuk sel induk megaspora. Sel induk megaspora akan mengalami pembelahan meiosis sebanyak dua kali hingga akhirnya membentuk embryo sac dengan delapan inti. Proses pembentukan
embryo sac tersebut membutuhkan waktu 60-65 hari setelah penyerbukan (HSP) pada P. pucherima. Fertilisasi baru terjadi pada 65-70 HSP dan pembelahan pertama zigot terjadi pada 75-80 HSP.
Jika proses fertilisasi baru terjadi sekitar dua bulan setelah penyerbukan, maka pemberian perlakuan kolkisin pada bunga setelah penyerbukan yang dilakukan selama jangka waktu enam sampai delapan hari akan menginduksi sel-sel somatik pada saat proses diferensiasi ovarium terjadi. Akibatnya yang terinduksi menjadi poliploid adalah sel-sel somatik yang akan menjadi sel induk megaspora, sehingga akan menghasilkan poliploid pada sel gamet betinanya saja menjadi gamet diploid. Sel gamet jantan dari polinia yang sudah berada di permukaan stigma didalam column sudah tidak mengalami pembelahan meiosis lagi, sehingga tidak terjadi penggandaan kromosom akibat pengaruh kolkisin. Jika hal tersebut yang terjadi maka tidak akan diperoleh planlet yang tetraploid, tetapi hasilnya akan berbentuk planlet triploid karena hasil penggabungan gamet jantan haploid dan gamet betina diploid. Sebagaimana yang diperoleh oleh Wu et al.
(2007) pada tanaman Lilium (non anggrek), penelitian induksi poliploidi melalui perlakuan kolkisin pada tunas bunga muda yang disilangkan dengan bunga kontrol (tanaman diploid), hasilnya diperoleh progeni triploid sebesar 18.0%. Berdasarkan fakta yang diperoleh dalam penelitian ini, dimana planlet yang diperoleh adalah tetraploid, maka diduga kuat bahwa bagian yang terinduksi menjadi poliploid adalah zigot ataupun embrio yang sudah terbentuk minimal enam hari setelah penyerbukan pada P. amabilis. Proses stimulasi pembentukan
embryo sac oleh polen sampai terjadi fertilisasi pada P. amabilis berlangsung selama enam hari setelah penyerbukan. Namun untuk memastikan kemungkinan hal tersebut diperlukan pengamatan mikroskopis pada beberapa waktu terhadap bunga setelah penyerbukan.
Poliploidi dapat terjadi dalam bentuk endopoliploid melalui proses endoreduplikasi. Endoreduplikasi merupakan proses duplikasi kromosom yang terjadi di dalam membran inti, tanpa adanya pembelahan mitosis yang lengkap maupun parsial. Pembentukan poliploid melalui proses endoreduplikasi merupakan mekanisme yang paling umum terjadi secara alami pada tanaman (Joubes dan Chevalier 2000). Chen et al. (2011b) menemukan adanya endopoliploid pada pada jaringan yang berbeda dari tahap perkembangan
protocorm dan juga planlet P. aphrodite. Tingkat endopoliploid yang tinggi ditemukan pada jaringan yang telah mengalami diferensiasi, sedangkan jaringan meristematik dibagian apikal memiliki tingkat endopoliploid yang rendah. Keberadaan endopoliploid yang secara alami pada proses perkembangan
22
kromosom yang diperoleh dari perlakuan kolkisin pada bunga setelah penyerbukan P. amabilis adalah bukan berasal dari endoreduplikasi. Penggandaan kromosom yang diperoleh dari hasil endoreduplikasi akan menghasilkan jumlah kromosom yang sama dengan diploidnya, namun jumlah lengan kromosom meningkat sehingga menghasilkan kromosom politen.
Simpulan
3
POTENSI INDUKSI POLIPLOIDI ANGGREK BULAN
(
Phalaenopsis amabilis
(L.) Blume) MELALUI PERLAKUAN
KOLKISIN PADA KUNCUP BUNGA
Abstrak
Pembentukan sel gamet poliploid merupakan salah satu metode poliploidisasi yang memiliki banyak manfaat karena menggunakan organ generatif. Beberapa manfaat tersebut adalah dapat menghasilkan progeni-progeni dengan tingkat ploidi yang beragam dalam waktu bersamaan, melalui penyerbukan sendiri dan silang. Aplikasi kolkisin pada kuncup bunga Phalaenopsis amabilis bertujuan untuk mempelajari pengaruh kolkisin terhadap perkembangan kuncup bunga dan potensi keberhasilan induksi poliploidi melalui penyerbukan sendiri dan silang. Konsentrasi kolkisin dalam percobaan adalah 0, 50, 500, 1000, dan 2000 mg L-1, dan durasi penyungkupan selama perlakuan kolkisin adalah tiga hari. Hasil percobaan menunjukkan bahwa konsentrasi kolkisin yang tinggi menghambat perkembangan kuncup bunga. Skrining terhadap planlet hasil perlakuan kolkisin menunjukkan potensi dalam menghasilkan planlet poliploid, terlihat dari perbedaan yang nyata secara morfologi, yaitu tipe planlet normal (N) dan tipe planlet diduga poliploid (DP). Tipe planlet DP dari semua perlakuan kolkisin berbeda nyata dengan kontrol untuk karakter panjang dan lebar basal organ of the protocorm (BOP), panjang dan lebar daun, serta panjang dan diameter akar. Kata kunci: anggrek, konsentrasi kolkisin, kuncup bunga, planlet poliploidi,
poliploidisasi
POTENTIAL OF POLYPLOIDY INDUCTION OF MOTH
ORCHID (
Phalaenopsis amabilis
(L.) Blume) USING
COLCHICINE ON FLOWER BUD
Abstract
24
(PPS). The probable polyploid seedlings of all colchicine treatments were significantly different as compared to control (0 mg L-1) for length and width of basal organ of the protocorm (BOP), leaf length and width, and root length and diameter.
Keywords: colchicine concentration, flower bud, orchid, polyploidization, polyploidy seedling
Pendahuluan
Metode poliploidisasi pada anggrek umumnya dilakukan melalui perlakuan menggunakan agen anti-mitosis yang ditargetkan pada sel-sel somatik dari jaringan meristematik, seperti protocorm dan planlet. Metode tersebut mengarah pada terbentuknya tanaman dari sel-sel somatik yang telah mengganda kromosomnya. Induksi poliploidi pada anggrek yang menggunakan organ generatif muda seperti kuncup bunga sebagai bahan perlakuan agen anti-mitosis sangat jarang dilakukan. Organ generatif muda sangat potensial digunakan untuk induksi poliploidi melalui proses pembentukan sel gamet jantan dan betina yang bersifat poliploid.
Pembentukan sel gamet terjadi melalui proses sporogenesis dan gametogenesis. Selama proses pembentukan sel gamet tersebut, terjadi pembelahan meiosis dan juga mitosis. Pemberian perlakuan kolkisin dapat mempengaruhi proses pembentukan sel gamet sehingga dapat mengakibatkan perubahan yang mengarah pada peningkatan tingkat ploidi. Menurut Teoh (1984), sel gamet poliploid pada anggrek diploid dapat terbentuk secara alami dengan persentase yang sangat kecil, seperti pada anggrek Spathoglottis plicata yang menghasilkan polen poliploid sebesar 0.63%. Perlakuan agen anti-mitosis seperti kolkisin pada organ generatif muda dapat meningkatkan persentase sel gamet poliploid.
Perlakuan kolkisin pada kuncup bunga anggrek memiliki beberapa keunggulan, diantaranya adalah dapat menghasilkan tingkat ploidi yang bervariasi. Kondisi tersebut dapat terjadi karena kolkisin akan menghasilkan sel gamet poliploid dengan tingkat yang berbeda bergantung pada tahapan dari proses pembentukan sel gamet, apakah ketika proses meiosis atau mitosis. Penggabungan antara sel gamet jantan dan betina yang bersifat poliploid akan menghasilkan progeni-progeni poliploid. Selain itu, keunggulan lain dari progeni atau planlet poliploid yang dihasilkan melalui perlakuan kolkisin pada kuncup bunga merupakan poliploid yang sifatnya solid. Berbeda dengan perlakuan kolkisin pada
25 Induksi sel gamet poliploid sangat potensial untuk diterapkan pada pemuliaan anggrek bulan dalam rangka menghasilkan varietas anggrek bulan poliploid dengan karakter bunga yang diinginkan. Penelitian induksi poliploidi yang menggunakan kuncup bunga P. amabilis bertujuan untuk mempelajari pengaruh kolkisin pada perkembangan kuncup bunga dan potensinya dalam menghasilkan planlet poliploid berdasarkan karakter morfologi.
Bahan dan Metode
Penelitian ini dilakukan di tempat penangkar anggrek di kawasan Puncak, Kabupaten Bogor, dan di Laboratorium Kultur Jaringan Tanaman Departemen Agronomi dan Hortikultura, Fakultas Pertanian IPB pada bulan Februari 2013 sampai Juli 2014. Bahan tanaman yang digunakan adalah P. amabilis yang sedang berbunga pada fase kuncup bunga asal Cianjur Selatan, Jawa Barat. Kuncup bunga dengan diameter sekitar 2 cm (Gambar 12) dibalut dengan kapas dan diberikan perlakuan kolkisin menggunakan kuas kecil. Konsentrasi larutan kolkisin yang digunakan adalah 0, 50, 500, 1000, dan 2000 mg L-1. Kuncup bunga yang telah diberi perlakuan kolkisin disungkup menggunakan aluminium foil selama tiga hari. Penelitian disusun berdasarkan Rancangan Acak Lengkap (RAL) dengan perlakuan kolkisin. Perlakuan kolkisin dilakukan pada 1-2 kuncup bunga pertanaman dengan empat ulangan, sehingga terdapat 20 tanaman sebagai satuan percobaan. Kuncup bunga dari perlakuan kolkisin yang berkembang dan telah mekar sempurna kemudian diserbuk sendiri dan silang secara buatan agar diperoleh buah anggrek. Penyerbukan silang dilakukan pada sebagian bunga dari perlakuan kolkisin 500 dan 1000 mg L-1, dengan menggunakan polen dari kontrol. Semua buah anggrek dipanen sekitar 24 minggu setelah penyerbukan (MSP).
Biji dari setiap buah anggrek kemudian disemai secara in vitro pada media Knudson C sampai diperoleh planlet umur 20 minggu setelah semai (MSS). Perlakuan kolkisin 0, 50, 500, dan 1000 mg L-1 yang menghasilkan planlet disubkultur pada media pendewasaan, yaitu media Hyponex. Sebanyak 125 planlet dipilih acak dari setiap perlakuan kolkisin, kecuali pada perlakuan dengan konsentrasi 1000 mg L-1 yang diserbuki sendiri karena hanya menghasilkan 50 planlet. Penelitian disusun secara RAL faktor tunggal dengan lima ulangan, setiap ulangan terdiri dari lima botol kultur, sedangkan pada perlakuan kolkisin 1000 mg L-1 yang disebuki sendiri terdiri dari dua botol kultur, sehingga terdapat 135 satuan percobaan. Dalam setiap botol terdiri dari lima planlet sehingga terdapat 675 planlet sebagai satuan amatan. Skrining planlet diduga poliploid dilakukan secara visual dengan membandingkan planlet hasil perlakuan kolkisin dengan kontrol berdasarkan morfologi pada umur 8 minggu setelah subkutur (MSSk). Karakter yang dibandingkan diantaranya adalah ukuran planlet, daun, dan basal organ of the protocorm (BOP).
26
Hasil dan Pembahasan
Kuncup bunga yang diberi perlakuan kolkisin dan kontrol dapat berkembang menjadi bunga yang mekar setelah sekitar 4-6 minggu setelah perlakuan (MSPr) kolkisin. Beberapa kuncup bunga dari setiap perlakuan kolkisin kering dan rontok. Persentase kuncup bunga hidup sampai menjadi bunga mekar sama dengan persentase jumlah buah pada setiap perlakuan kolkisin (Tabel 5). Konsentrasi kolkisin berpengaruh nyata terhadap persentase jumlah buah (Gambar 13). Konsentrasi kolkisin yang lebih tinggi dari 50 mg L-1 hanya menghasilkan persentase jumlah buah 37.5%, yaitu pada konsentrasi 500 dan 1000 mg L-1, dan 0.0% pada konsentrasi 2000 mg L-1. Perlakuan kolkisin dengan konsentrasi 2000 mg L-1 sangat toksik terhadap kuncup bunga, semua kuncup bunga rontok pada 1-2 MSPr. Menurut Nakasone (1960), perlakuan kolkisin pada spike dari Vanda
Miss Joaquin dengan konsentrasi 1000 sampai 20000 mg L-1 menyebabkan kerontokan kuncup bunga. Pengaruh kolkisin pada kerontokan kuncup bunga juga dilaporkan oleh Wu et al. (2007) pada tanaman Lilium (non anggrek) dengan konsentrasi kolkisin 2000 mg L-1.
Tabel 5 Jumlah perlakuan kuncup bunga dan beberapa karakter pada berbagai konsentrasi kolkisin
Kolkisin (mg L-1)
Jumlah kuncup bunga diberi
perlakuan
Jumlah buah(b)
Jumlah buah menghasilkan