KARAKTERISTIK BIOMETRIK DAN GENETIK SPESIES
IKAN LAIS (SILURIDAE) DAN BIOLOGI REPRODUKSI
Ompok miostoma
(Vaillant, 1902) DI SUNGAI MAHAKAM
KALIMANTAN TIMUR
JUSMALDI
SEKOLAH PASCA SARJANA
INSTITUT PERTANIAN BOGOR
INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa disertasi berjudul Karakteristik Biometrik dan Genetik Spesies Ikan Lais (Siluridae) dan Biologi Reproduksi Ompok miostoma (Vaillant, 1902) di Sungai Mahakam Kalimantan Timur adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam daftar pustaka di bagian akhir disertasi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Juli 2016
JUSMALDI. Karakteristik Biometrik dan Genetik Spesies Ikan Lais (Siluridae) dan Biologi Reproduksi Ompok miostoma (Vaillant, 1902) di Sungai Mahakam Kalimantan Timur. Dibimbing oleh DEDY DURYADI SOLIHIN, RIDWAN AFFANDI, MF RAHARDJO dan RUDHY GUSTIANO.
Lais adalah nama lokal di Sungai Mahakam Kalimantan Timur untuk kelompok ikan bersungut air tawar (catfish) famili Siluridae terutama genus Ompok dan Kryptopterus. Bagi masyarakat lokal, genus ini tertutama dimanfaatkan sebagai ikan konsumsi dan ikan hias. Kerusakan lingkungan dan penangkapan ikan yang tidak selektif yang dilakukan oleh para nelayan dikhawatirkan akan menurunkan populasi ikan lais dengan cepat, sementara informasi dasar biologi dan validitas spesies pada kedua genus ini di alam belum banyak diketahui. Untuk menjaga kelestarian dan pemanfaatan ikan lais berkelanjutan maka diperlukan konservasi dan pengelolaan yang baik. Tujuan dari penelitian ini adalah: 1) menentukan perbedaan karakter antara genus dan spesies
menggunakan biometrik serta menyediakan kunci identifikasi praktis. 2) menentukan identitas spesies ikan lais secara akurat menggunakan analisis
genetik. 3) mendeskripsikan penyebaran spasial dan temporal spesies ikan lais. 4) menyusun strategi dan pengelolaan perikanan untuk pemanfaatan berlanjutan sumber daya ikan khususnya spesies Ompok miostoma melalui kajian biologi reproduksi.
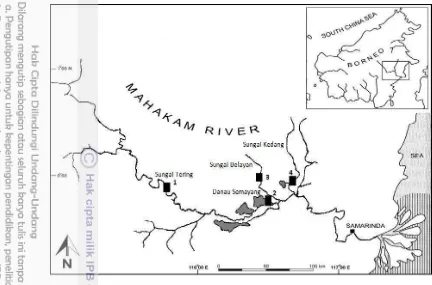
Pengumpulan sampel dan pengukuran kualitas perairan dilakukan setiap bulannya di empat lokasi di Sungai Mahakam selama satu tahun yang meliputi kawasan Sungai Mahakam Hulu di Sungai Tering (S 000 04' 03''; E 1150 40' 05'')
dan kawasan Sungai Mahakam Tengah di Danau Semayang (S 000 11' 09.8''; E 1160 27' 31.2''); Sungai Belayan (S 000 12' 18.2"; E 1160 31' 55.3"); Sungai
Kedang Muara Siran (S 000 07' 26.1''; E 1160 37' 56.4''). Identifikasi spesies, pengukuran karakter morfologis dan pengukuran parameter biologi reproduksi dilakukan di Laboratorium Biologi FMIPA Universitas Mulawarman Samarinda dan Museum Zoologi Bogor, sementara ektraksi DNA dan amplifikasi dilakukan di Laboratorium Molekular PSIH IPB Bogor. Perunutan DNA dilakukan di 1st BASE Malaysia.
Identifikasi, validasi dan distribusi spesies ikan lais dianalisis secara deskriptif. Untuk biometrik dilakukan menggunakan analisis komponen utama. Analisis karakter genetik, penanda genetik, jarak genetik dan konstruksi ulang pohon filogenetik dilakukan menggunakan software MEGA 6. Struktur populasi ikan lais berdasarkan rasio kelamin, ukuran, tingkat kematangan gonad, musim pemijahan dan tipe pemijahan dianalisis secara deskriptif. Hubungan panjang berat jantan dan betina, hubungan fekunditas dan ukuran ikan (panjang total dan berat) ditentukan menggunakan analisis regresi. Pola pertumbuhan, faktor kondisi, indeks kematangan gonad (IKG) dan ukuran ikan kali pertama matang gonad dianalisis dengan rumus yang telah ditentukan menggunakan program EXEL.
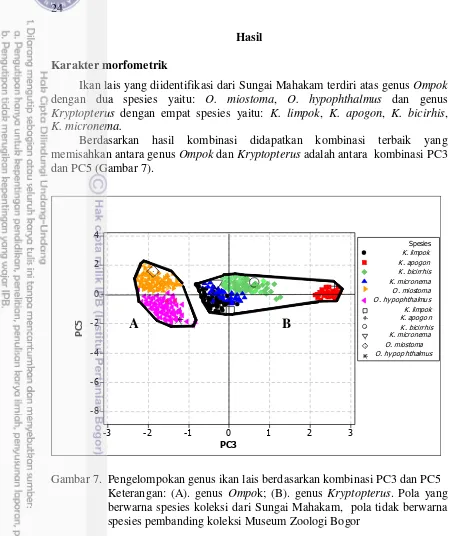
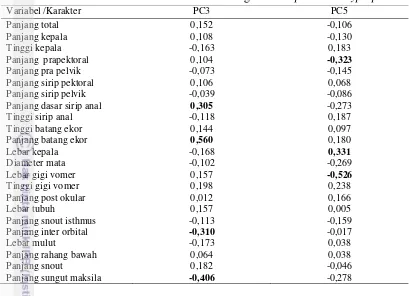
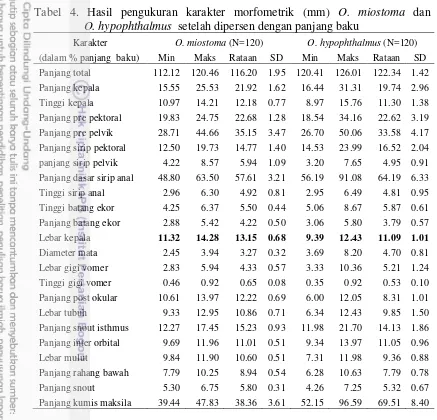
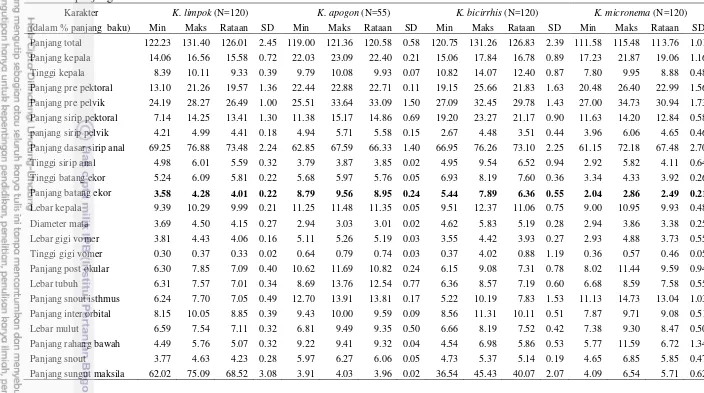
batang ekor, panjang sungut maksila, lebar kepala, panjang interorbital, panjang prepektoral, panjang dasar sirip anal, lebar gigi vomer, dan jumlah tapis insang. Antara spesies O. miostoma dan O. hypophthalmus dibedakan oleh perbedaan lebar kepala, panjang sungut maksila, bentuk gigi vomer, susunan gigi maksila, posisi noktah warna, sementara spesies K. limpok, K. apogon, K. micronema, K. bicirrhis dibedakan pada panjang batang ekor, panjang sungut maksila, bentuk gigi vomer, susunan gigi maksila, posisi noktah warna dan warna dorsal tubuh. Karakter biometrik dan deskripsi digunakan untuk menyediakan kunci identifikasi.
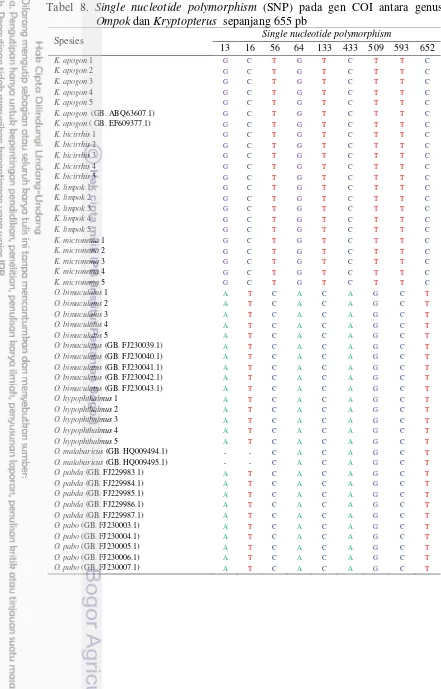
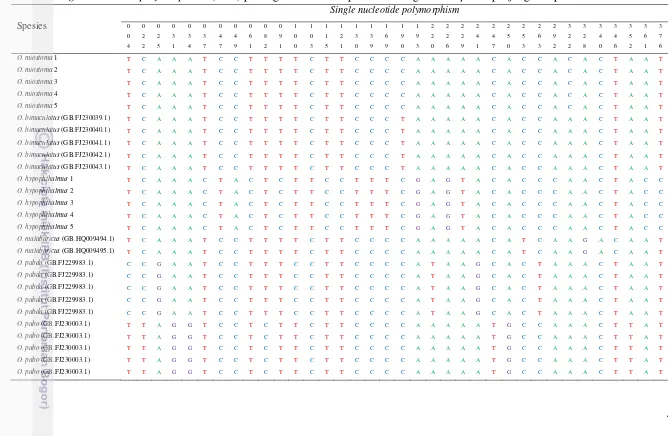
Perbedaan karakter genetik antara genus Ompok dan Kryptopterus berdasarkan gen COI sepanjang 655 pasang basa (pb) ditunjukkan oleh variasi pada kodon ketiga pada genus Ompok sebesar 22,75% dan genus Kryptopterus sebesar 12,22%. Antara genus Ompok dan Kryptopterus dibedakan oleh 9 single nucleotide polymorphism (SNP). Spesies O. miostoma dan O. hypophthalmus dibedakan oleh 22 SNP, sedangkan spesies K. limpok, K. apogon, K. micronema, K.bicirrhis dibedakan oleh 73 SNP. Rata-rata jarak genetik intraspesies ditemukan sebesar 0,15% ± 0,01%, antarspesies dalam genus Ompok sebesar 12,77 % ± 0,10%, antarspesies dalam genus Kryptopterus sebesar 10,99% ± 0,29% dan antargenus Ompok dan Kryptopterus sebesar 19,01% ± 1,68. Konstruksi ulang pohon filogenetik berdasarkan gen COI ditemukan enam kelompok spesies ikan lais di Sungai Mahakam.
Sebaran spesies berdasarkan lokasi penangkapan menunjukkan spesies O. miostoma dan K. apogon ditemukan pada semua lokasi, sedangkan spesies K. bicirrhis hanya ditemukan di kawasan Sungai Mahakam Hulu. Ikan lais di Sungai Mahakam didominasi oleh spesies O. miostoma, sedangkan spesies K. apogon ditemukan dalam jumlah yang sedikit. Kelimpahan relatif ikan lais di Sungai Mahakam Tengah lebih tinggi dibandingkan Sungai Mahakam Hulu. Distribusi spasial spesies ikan lais pada seluruh lokasi dipengaruhi oleh perbedaan kondisi habitat, sedangkan distribusi temproral dipengaruhi oleh musim.
Biologi reproduksi Ompok miostoma menunjukkan nisbah kelamin jantan dan betina pada saat matang gonad adalah 1 : 1,77. Panjang ikan kali pertama matang gonad pada jantan berkisar dari 191,05 - 202,60 mm dan betina berkisar dari 179,56 - 198,50 mm. Musim pemijahan terjadi pada bulan November sampai Januari (musim hujan) dan puncak pemijahan terjadi pada bulan Desember. Lokasi pemijahan berdasarkan pada persentase tertinggi matang gonad dan fekunditas total adalah di Danau Semayang. Berdasarkan pada sebaran diameter telur diketahui pola pemijahan serempak (total spawning). Rekomendasi strategi pengelolaan ikan lais di Sungai Mahakam, dapat ditentukan berdasarkan aspek biologi reproduksi meliputi: pembatasan penangkapan pada puncak musim pemijahan pada bulan Desember, ukuran O. miostoma jantan yang boleh ditangkap di atas 191,05 - 202,60 mm dan pada betina di atas 179,56 - 198,50 mm dan ukuran mata jaring yang disarankan di atas 2 Inci. Danau Semayang ditetapkan sebagai wilayah yang dikonservasi untuk pemijahan ikan lais. Pada tahap lebih lanjut diperlukan usaha domestikasi berdasarkan hasil penelitian.
JUSMALDI. Biometric Characteristics and Genetic Silurid Catfish and Reproduction Biology of Ompok miostoma (Vaillant, 1902) from the Mahakam River in East Kalimantan. Under direction of DEDY DURYADI SOLIHIN, RIDWAN AFFANDI, MF RAHARDJO and RUDHY GUSTIANO.
Lais is the local name in Mahakam River for the group of freshwater catfishes of the Siluridae family, belonging the genera Ompok and Kryptopterus. For local communities, these genera are mainly used as a food and ornamental fish. Degradation of environment and unselected fishing by the fishermen enable to reduce populations of lais fish rapidly. However, basic information on the biology and validity of the species in two genera is not clear. To preserve and sustainable use of lais group is required conservation and good management. The purpose of this research was: 1) to determine the differences in characteristics of lais both in the genera and species using biometric for providing practical key. 2) to determine the identity of lais species accurately using genetic analysis. 3) to describe the spatial and temporal patterns of the lais species. 4) to design strategies and fisheries management for the sustainable utilization of the fisheries resources especially for O. miostoma through the study of reproduction biology.
Collecting of samples and measurement of water quality were done every month in four locations in the Mahakam River for the whole year, covering the area of the upstream Mahakam River in Sungai Tering (S 000 04' 03''; E 1150 40' 05'') and the middle area in Semayang Lake (S 000 11' 09.8''; E 1160 27' 31.2''); Belayan River (S 000 12' 18.2"; E 1160 31' 55.3"); Kedang Muara Siran River (S 000 07' 26.1''; E 1160 37' 56.4''). Species identification, measuring of morphological characters and reproduction biology conducted in Biology Laboratory University of Mulawarman and Bogor Zoological Museum, while DNA extraction and amplification performed in Laboratory of Molecular PSIH IPB Bogor. DNA sequencing carried out in 1st BASE Malaysia.
Identification, validation and distribution species of lais fish were analyzed descriptively. For biometric determined by principal component analysis. Analysis of genetic characteristic, genetic variations, genetic distance and phylogenetic tree re-construction were done using MEGA program version 4. The structure of the lais fish population based on sex-ratio, size, gonad development stages, spawning period and spawning type were analyzed by descriptively. Length-weight relationship of male and female, fecundity and fish size (total length and weight) were determined using regretion analysis. Growth pattern, condition factor, gonado-somatic index (GSI), first size maturity were analysis with a formula using EXEL software.
Observed species found during study consisting of 2 genera and 6 species. The results of biometric characters showed that Ompok and Kryptopterus is distinguished by differences in presence of dorsal fin, caudal peduncle length, maxillary barbel length, head width, interorbital distance, prepectoral length, anal
and spot position. While K. limpok, K. apogon, K. micronema, K. bicirrhis differ in caudal peduncle length, maxillary barbel length, vomerine teeth shape, maxillary teeth structure, spot position and dorsal coloration. Biometric and descriptions characters are used to provide identification key.
Characteristics genetic differences between genus Ompok and Kryptopterus based on the 655 bp COI gene indicated that the variation of third codon in genus Ompok to 22.75% and genus Kryptopterus to 12.22%. Ompok and Kryptopterus
can be distinguished by 9 single nucleotide polymorphism (SNP). The species O. miostoma and O. hypophthalmus can be distinguished by 22 SNP, while K. limpok, K. apogon, K. micronema, K. bicirrhis can be distinguished individually by 73 SNP. The genetic distance interspecies was found reached 0,15% ± 0,01%, conspecifics in Ompok and Kryptopterus 12,77% ± 0,10% and 10,99% ± 0,29% respectively and congeneric species 19,01%. ± 1,68. Reconstruction of genetic tree based on the 655 bp COI gene was found six groups species lais fish from Mahakam River.
Distribution of the species based on fishing locations shows that O. miostoma and K. apogon were found in all location, while K. bicirrhis was
found only in the upstream Mahakam River. Lais fishes from the Mahakam River were dominated by O. miostoma, while K. apogon was found in lower number. The relative abundance of lais fish in the Middle Mahakam River is higher than the Upstream Mahakam River. The spatial distribution of lais fish in all location influenced by differences in habitat conditions, while the temporal distribution influenced by seasons.
Reproduction biology of Ompok miostoma shows that the sex ratio of male and female at gonad maturity stages were 1 : 1.77. The length at first maturity of male were ranging from 191.05 to 202.60 mm and female 179.56 to 198.50 mm. The spawning season was ranging from November to January (rainy seasons) and peak spawning occurs in December. Spawning location based on highest gonad maturity percentage and total fecundity occurred in Semayang Lake. Based on distribution of eggs diameter known spawning patterns simultaneously (total spawning). Recommended strategic management of lais fish in Mahakam River, could be determined by biology reproduction aspects include: limiting fishing during peak spawning season in December, the size of O. miostoma permitted fishing in male and female above 191.05 to 202.60 mm and 179.56 to 198.50 mm respectivly and mesh size recommended above 2 inches. Semayang Lake establishment as the protection zone for spawning lais fish. More over, an action
of domestication of the lais fish is required, based on the research’s result.
© Hak Cipta Milik IPB, Tahun 2016
Hak Cipta Dilindungi Undang - Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
KARAKTERISTIK BIOMETRIK DAN GENETIK SPESIES
IKAN LAIS (SILURIDAE) DAN BIOLOGI REPRODUKSI
Ompok miostoma
(Vaillant, 1902) DI SUNGAI MAHAKAM
KALIMANTAN TIMUR
JUSMALDI
Disertasi
Sebagai salah satu syarat untuk memperoleh gelar Doktor
pada
Program Studi Biosains Hewan
SEKOLAH PASCA SARJANA
INSTITUT PERTANIAN BOGOR
Penguji Luar Komisi pada Ujian Tertutup:
1. Dr Ir Dinar Tri Soelistyowati, DEA (Departemen Budidaya Perairan Fakultas Perikanan dan Ilmu Kelautan Institut Pertanian Bogor)
2. Dr Ir H Sudarto, MSc (Balai Penelitian dan Pengembangan Budidaya Ikan Hias Depok)
Penguji Luar Komisi pada Sidang Promosi Terbuka:
1. Prof Dr Ir Endi Setiadi Kartamihardja, MSc (Profesor Riset di Balai Penelitian Pemulihan dan Konservasi Sumber Daya Ikan, Pusat Penelitian Pengelolaan Perikanan dan Konservasi Sumber Daya Ikan, KKP Jatiluhur)
Penulis memanjatkan puji syukur kehadirat Allah SWT atas segala karunia, taufik dan hidayah-Nya sehingga penulisan disertasi berjudul Karakteristik Biometrik dan Genetik Spesies Ikan Lais (Siluridae) dan Biologi Reproduksi Ompok miostoma (Vaillant, 1902) di Sungai Mahakam Kalimantan Timur dapat diselesaikan dengan baik. Disertasi ini disusun sebagai salah satu syarat untuk memperoleh gelar Doktor di Program Studi Biosains Hewan, Sekolah Pasca Sarjana, Institut Pertanian Bogor.
Penulis sangat menyadari bahwa proses penyelesaian penelitian dan penulisan disertasi ini tidak akan dapat berjalan lancar tanpa dukungan banyak pihak, sehingga pada kesempatan ini penulis mengucapkan terima kasih kepada Dr Ir Dedy Duryadi Solihin, DEA, Prof Dr Ir Ridwan Affandi, DEA, Prof Dr Ir MF Rahardjo, DEA dan Dr Ir Rudhy Gustiano, MSc sebagai komisi pembimbing atas waktu dan bimbingannya mulai dari penyusunan proposal, pelaksanaan penelitian hingga penulisan disertasi.
Terima kasih disampaikan kepada Kementrian Riset, Teknologi dan Pendidikan Tinggi, atas penyediaan beasiswa BPPS Doktoral sehingga penulis dapat memperdalam ilmu di Institut Pertanian Bogor. Terima kasih juga penulis sampaikan kepada pimpinan PT Gencraff atas izin pengunaan fasilitas Laboratorium Genetika di Jakarta.
Terima kasih dan penghargaan yang tinggi penulis haturkan kepada ibunda Mariana (Almarhum) dan ayahanda Juslim (Almarhum) atas doa, dukungan dan semangatnya selama ini. Kepada istri Adriany, SE dan anak tercinta Aura Anastacia Aldany, terima kasih atas semua doa, perhatian dan kesabarannya.
Terima kasih kepada rekan-rekan S3 Biosains Hewan angkatan 2011: Dr Andi Darmawan, MSi., FX Widadi Padmasari, MSi., Fitria Basalamah, MSc., Dr Kellyopas Krey, MSi., Maria Ulfah, MSc., RR Sricatur, MSi., dan Yuliadi Zamroni, MSi., atas kebersamaannya selama menempuh studi. Kepada rekan-rekan Laboratorium Molekuler Hewan Dr Nurlisa Butet, MSc., Dr Hari Prayogo, MSi., Dr Nungky, MSi., Dr Arini, MSi., Andi Haerul, MSi., Bahqiah, MSi., Syamsul, MSi., Jarulis, MSi.,Yuli, SSi., Della, SSi., Kamila, SSi. serta Pak Heri atas kebersamaan dan kerjasamanya selama penelitian. Secara khusus ucapan terimakasih diberikan kepada Ajeng Daniarsih, MSi., yang telah memberikan semangat dan banyak membantu dalam proses penyelesaian disertasi ini.
Akhir kata, semoga hasil penelitian ini dapat bermanfaat bagi pengembangan ilmu pengetahuan dan berkonstribusi bagi langkah pengelolaan perikanan air tawar. Demikian harapan penulis, terima kasih.
DAFTAR TABEL xiv
DAFTAR GAMBAR xv
DAFTAR LAMPIRAN xvii
1 PENDAHULUAN 1
Latar Belakang 1
Perumusan Masalah 3
Tujuan Penelitian 3
Manfaat Penelitian 3
Kebaruan (Novelty) 4
2 TINJAUAN PUSTAKA 5
3 BIOMETRIK DAN KUNCI IDENTIFIKASI IKAN LAIS GENUS Ompok DAN Kryptopterus (FAMILI SILURIDAE) ASAL SUNGAI MAHAKAM
KALIMANTAN TIMUR 20
Pendahuluan 20
Bahan dan Metode 21
Hasil 24
Pembahasan 29
Simpulan 37
4 KARAKTER GENETIKIKAN LAIS Ompok spp DAN Kryptopterus spp FAMILI SILURIDAE) ASAL SUNGAI MAHAKAM KALIMANTAN TIMUR BERDASARKAN GEN COI DNA MITOKONDRIA 38
Pendahuluan 38
Bahan dan Metode 39
Hasil 41
Pembahasan 51
Simpulan 53
5 DISTRIBUSI SPESIES IKAN LAIS (FAMILI SILURIDAE) DI SUNGAI MAHAKAM KALIMANTAN TIMUR 54
Pendahuluan 54
Bahan dan Metode 55
Hasil 56
Pembahasan 60
Simpulan 61
6 BIOLOGI REPRODUKSI IKAN LAIS Ompok miostoma (Vaillant, 1902) DI SUNGAI MAHAKAM KALIMANTAN TIMUR 62
Pendahuluan 62
Bahan dan Metode 63
Hasil 71
Pembahasan 93
8 SIMPULAN UMUM DAN SARAN 103
DAFTAR PUSTAKA 105
LAMPIRAN 114
RIWAYAT HIDUP 118
DAFTAR TABEL
1 Ciri-ciri morfologis ikan lais genus Ompok dan Kryptopterus 6
2 Karakteristik DNA inti dan DNA mitokondria 9
3 Nilai faktor koefisien PC3 dan PC5 dari genus Ompok dan
Kryptopterus 25
4 Hasil pengukuran karakter morfometrik (mm) O. miostoma dan
O. hypophthalmus setelah dipersenkan dengan panjang baku 26 5 Hasil pengukuran karakter morfometrik (mm) K. limpok,
K. apogon, K. bicirrhis dan K. micronema setelah dipersenkan
dengan panjang baku 27
6 Data meristik antar spesies dalam genus Ompok dan Kryptopterus 28 7 Karakterisasi nukleotida gen COI genus Ompok dan Kryptopterus
sepanjang 655 pb 42
8 Single nucleotide polymorphism (SNP) pada gen COI antara genus
Ompok dan Kryptopterus sepanjang 655 pb 44
9 Single nucleotide polymorphism (SNP) pada gen COI antar
spesies dalam genus Ompok sepanjang 655 pb 45
10 Single nucleotide polymorphism (SNP) pada gen COI antar
spesies dalam genus Kryptopterus sepanjang 655 pb 47 11 Jarak genetik ikan lais dari Sungai Mahakam ditambah data
GenBank berdasarkan runutan gen COI mitokondria 655 pb 49 12 Spesies ikan lais yang ditemukan di empat lokasi penelitian 56 13 Spesies dan kelimpahan ikan lais berdasarkan lokasi penelitian 57 14 Penentuan tingkat kematangan gonad ikan secara morfologis 65 15 Pengukuran parameter fisika-kimia perairan yang berperan dalam
reproduksi ikan 71
16 Jumlah, kisaran panjang total dan berat ikan lais O. miostoma
(Vaillant, 1902) selama satu tahun 72
17 Analisis hubungan panjang-berat ikan lais O. miostoma (Vaillant,
1902) di Sungai Mahakam 73
18 Faktor kondisi relatif bulanan ikan lais O. miostoma (Vaillant,
1902) jantan dan betina di lokasi Sungai Mahakam 74 19 Nisbah kelamin ikan lais O. miostoma (Vaillant, 1902) yang
tertangkap di lokasi Sungai Mahakam 75
20 Nisbah kelamin ikan lais O. miostoma (Vaillant, 1902) yang
O. miostoma (Vaillant, 1902) jantan secara morfologis dan
histologis 78
22 Penentuan tingkat kematangan gonad (TKG) ikan lais O. miostoma (Vaillant, 1902) betina secara morfologis dan
histologis 80
23 Tingkat kematangan gonad (TKG) ikan lais O. miostoma (Vaillant, 1902) jantan dan betina berdasarkan ukuran panjang
total (mm) dan berat tubuh (gram) 83
24 Indeks kematangan gonad (IKG) dan berat gonad ikan lais O. miostoma (Vaillant, 1902) jantan dan betina berdasarkan
TKG 85
25 Indeks kematangan gonad (IKG) ikan lais O. miostoma
(Vaillant, 1902) setiap bulan selama satu tahun penelitian 86 26 Fekunditas dan diameter telur ikan lais O. miostoma (Vaillant,
1902) di lokasi Sungai Mahakam 87
27 Persentase sebaran telur ikan lais O. miostoma (Vaillant, 1902)
berdasarkan tingkat kematangan gonad 89
28 Kondisi fisika dan kimia perairan di empat lokasi pengambilan
sampel selama satu tahun penelitian 91
DAFTAR GAMBAR
1 Peta jalan penelitian 4
2 Peta penyebaran famili Siluridae di Eropa dan Asia 5
3 Ilustrasi spesies ikan lais famili Siluridae 7
4 Organisasi genom mitokondria Silurus glanis 10
5 Lokasi pengambilan sampel ikan di Sungai Mahakam 22
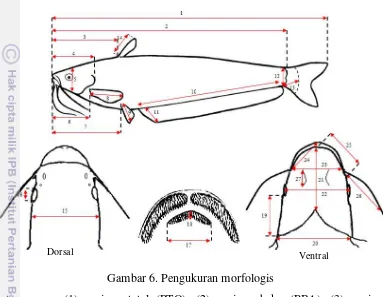
6 Pengukuran morfologis 23
7 Pengelompokan genus ikan lais berdasarkan kombinasi PC3
dan PC5 24
8 Kombinasi lebar kepala dan panjang batang ekor spesies pada
genus Ompok 25
9 Kombinasi lebar kepala dan panjang batang ekor spesies pada
genus Kryptopterus 28
10 Ompok miostoma (Vaillant, 1902) (panjang baku: 215 mm). (A) Noktah warna (N), (B) Bentuk vomer (V) dan susunan gigi
maksila (M) 31
11 Ompok hypophthalmus (Bleeker, 1846) (panjang baku: 187 mm). (A) Noktah warna (N), (B) Bentuk vomer (V) dan
susunan gigi maksila (M) 32
12 Kryptopterus limpok (Bleeker, 1852) (panjang baku 105 mm). (A) Noktah warna (N), (B) Bentuk vomer (V) dan susunan gigi
maksila (M) 33
13 Kryptopterus apogon (Bleeker, 1851) (panjang baku 208 mm). (A) warna dorsal (D) (B) Bentuk vomer (V) dan susunan gigi
mm). (A) Noktah warna (N), (B) Bentuk vomer (V) dan
susunan gigi maksila (M) 35
15 Kryptopterus micronema (Bleeker, 1846) (panjang baku 232 mm). (A) warna dorsal (D) (B) Bentuk vomer (V) dan susunan
gigi maksila (M) 36
16 Lokasi pengambilan sampel ikan di Sungai Mahakam 39
17 Pita hasil amplifikasi gen COI ikan lais 41
18 Skema posisi penempelan primer Fish F1 dan Fish R1 dengan
acuan gen COI utuh Silurus glanis (GB. AM398435.2) 42 19 Konstruksi ulang pohon filogenetik spesies ikan lais dalam
genus Ompok dan Kryptopterus berdasarkan model Kimura 2 parameter runutan nukleotida gen COI parsial 655 bp dengan
metode neigbour joining, bootstrap 1000 kali 50 20 Lokasi pengambilan sampel ikan di Sungai Mahakam 55 21 Fluktuasi jumlah individu spesies ikan lais yang tertangkap dari
bulan Januari hingga Desember di Danau Semayang 57 22 Fluktuasi jumlah individu spesies ikan lais yang tertangkap dari
bulan Januari hingga Desember di Sungai Belayan 57 23 Fluktuasi jumlah individu spesies ikan lais yang tertangkap dari
bulan Januari hingga Desember di Sungai Kedang Muara Siran 58 24 Fluktuasi jumlah individu spesies ikan lais yang tertangkap dari
bulan Januari hingga Desember di Sungai Tering 58 25 Persentase kelimpahan total ikan lais di empat lokasi
penelitian 59
26 Lokasi pengambilan sampel ikan di Sungai Mahakam 63
27 Ikan lais Ompok miostoma (Vaillant, 1902) 64
28 Model regresi hubungan panjang dan berat tubuh ikan lais O. miostoma (Vaillant, 1902) jantan di Sungai Mahakam 72 29 Model regresi hubungan panjang dan berat tubuh ikan lais
O. miostoma (Vaillant, 1902) betina di Sungai Mahakam 73 30 Fluktuasi faktor kondisi relatif bulanan ikan lais O. miostoma
(Vaillant, 1902) jantan dan betina di lokasi Sungai Mahakam 74 31 Posisi gonad ikan jantan (a) dan betina (b) di dalam rongga
perut O. miostoma (Vaillant, 1902) 77
32 Struktur morfologis (a) dan histologis (b) gonad jantan
O. miostoma (Vaillant, 1902) 79
33 Struktur morfologis (a) dan histologis (b) gonad betina
O. miostoma (Vaillant, 1902) 81
34 Persentase tingkat kematangan gonad ikan lais O. miostoma (Vaillant, 1902) berdasarkan bulan penelitian di Sungai
Mahakam 82
35 Persentase TKG IV ikan lais O. miostoma (Vaillant, 1902)
berdasarkan lokasi penelitian di Sungai Mahakam 83 36 Hubungan faktor kondisi dengan tingkat kematangan gonad
O. miostoma (Vaillant, 1902) dengan curah hujan dan jumlah
hari hujan di lokasi Sungai Mahakam 86
38 Hubungan fekunditas total menurut panjang tubuh (mm) 87 39 Hubungan fekunditas total menurut berat tubuh (gram) 88 40 Fekunditas relatif menurut kelompok panjang tubuh 88 41 Fekunditas relatif menurut kelompok berat tubuh 89 42 Sebaran diameter telur ikan lais O. miostoma (Vaillant, 1902)
berdasarkan TKG III 90
43 Sebaran diameter telur ikan lais O. miostoma (Vaillant, 1902
berdasarkan TKG IV 90
44 Sebaran diameter telur ikan lais O. miostoma (Vaillant, 1902)
berdasarkan TKG V 90
45 Fluktuasi nilai parameter fisika kimia air di empat lokasi
penelitian selama satu tahun 92
DAFTAR LAMPIRAN
1 Lokasi penelitian di Sungai Mahakam 115
2 Contoh ukuran diameter telur 116
1 PENDAHULUAN
Latar Belakang
Sungai Mahakam merupakan sungai terpanjang di Provinsi Kalimantan Timur dan dikenal sebagai salah satu bagian dari perairan umum yang memegang peranan penting dalam menghasilkan ikan air tawar di Indonesia. Di perairan ini terdapat 174 jenis ikan air tawar dan salah satu jenis yang dominan adalah ikan lais (Kottelat 1994). Ikan lais adalah nama lokal yang diberikan oleh masyarakat di Kalimantan, ditujukan pada kelompok ikan air tawar bersungut (catfish) anggota famili Siluridae yang memiliki ciri morfologi tubuh mirip (Nelson 2009). Ikan ini umumnya dimanfaatkan sebagai ikan konsumsi dan ikan hias yang bernilai ekonomis. Menurut laporan Christensen (1992) setidaknya di perairan Sungai Mahakam terdapat 13 spesies ikan lais dan sebagian besar terdiri atas genus Ompok dan Kryptopterus.
Kerusakan lingkungan dan usaha penangkapan ikan yang tidak selektif terhadap ukuran yang dilakukan oleh para nelayan di perairan Sungai Mahakam dikhawatirkan akan menurunkan populasi ikan lais dimasa datang. Hal tersebut dapat dilihat dari semakin menurunnya hasil tangkapan nelayan dari tahun ke tahun. Berdasarkan data statistik kelautan dan perikanan menunjukkan produksi tangkapan ikan di perairan umum Provinsi Kalimantan Timur mengalami penurunan sebesar 1,72 % yaitu dari 44.108 ton pada tahun 2010 menjadi 43.348 ton pada tahun 2014 (KKP 2015). Penurunan produksi ini mengindikasikan mulai terjadinya penurunan stok ikan di Provinsi Kalimantan Timur khususnya di Sungai Mahakam, sementara di sisi lain informasi penting dan mendasar seperti biologi ikan lais yang merupakan salah satu tangkap utama di alam belum banyak diungkap (Ng 2003b).
Untuk menjaga kelestarian dan pemanfaatan berkelanjutan maka diperlukan usaha pengelolaan dan konservasi. Kebijakan pengelolaan dan konservasi memerlukan informasi ilmiah sebagai dasar pertimbangan pengelolaan meliputi jenis dan identitas spesies, karakteristik genetik dan morfologik, distribusi populasi, serta informasi biologi reproduksi.
Biometrik merupakan analisis morfometrik dan meristik yang lebih komprehensif diperlukan pada spesies yang sangat mirip dengan kerabat dekat. Morfometrik merupakan suatu metode untuk mengetahui perbedaan antar spesies dengan cara membandingkan dimensi ukuran morfologis ikan (Moyle dan Ceach 2004). Belum ada penelitian karakter morfometrik berdasarkan analisis nilai komponen utama karakter morfologis pada spesies ikan lais genus Ompok dan Kryptopterus asal Sungai Mahakam.
Analisis karakter genetik diperlukan untuk akurasi spesies terutama pada kasus spesies yang dekat hubungan kerabatnya. Gen sub unit 1 Cytochrome c oxidase (COI) DNA mitokondria merupakan salah satu marka molekular yang banyak digunakan sebagai DNA barcoding. Gen COI banyak digunakan sebagai DNA barcoding karena banyak bagian dari runutan nukleotidanya bersifat konservatif ditingkat spesies dan sedikit variasi, sehingga dapat dipakai untuk identifikasi, karakterisasi dan pengelompokan spesies pada berbagai taksa hewan termasuk ikan (Hebert et al. 2003, Ward et al. 2005). Selain itu marka genetik lain yang dipakai sebagai barcoding spesies adalah gen Cytochrome b. Penelitian identifikasi molekuler ikan lais asal Sungai Kampar Riau menunjukkan genus Ompok dan Kryptopterus berbeda pada asam amino ke-155 dari runutan gen Cytochrome b utuh yaitu Metionin atau Isoleusin pada genus Ompok, sedangkan Valin pada genus Kryptopterus (Elvyra dan Solihin 2007). Sementara penelitian identifikasi molekuler berdasarkan runutan gen parsial COI DNA mitokondria
yang dilaporkan pada spesies di luar Indonesia yaitu: O. pabo, O. pabda, O. bimaculatus, O. malabaricus (Lekshmi et al. 2010, Malakar et al. 2012) dan K. apogon (Ward dan Holmes 2007). Di samping itu, informasi runutan gen COI DNA mitokondria untuk ikan famili Siluridae sebagai DNA barcoding di dalam data GenBank hingga tahun 2015 masih sangat terbatas.
Penelitian berkaitan dengan distribusi ikan lais genus Kryptopterus dan Ompok di Sungai Mahakam sampai saat ini belum banyak dilakukan. Beberapa penelitian tentang distribusi ikan di Sungai Mahakam hanya sebatas pada lokasi tertentu dan belum mewakili keseluruhan perairan (Haryono 2006, Nasution et al. 2008). Informasi tentang distribusi spesies ikan lais secara temporal dan spasial di perairan Sungai Mahakam belum ada, informasi ini sangat penting bagi upaya konservasi dan pemetaan lokasi tangkapan ikan bagi nelayan setempat.
Informasi tentang siklus reproduksi ikan dan faktor-faktor lingkungan yang mempengaruhinya merupakan hal yang mendasar dalam biologi perikanan (Tomkiewicz et al. 2003). Beberapa informasi penting akan diperoleh dengan mempelajari aspek reproduksi ikan di antaranya adalah fekunditas yang berhubungan dengan rekruitmen, perkembangan gonad, ukuran pertama kali matang gonad, waktu pemijahan, dan tipe pemijahan. Informasi biologi reproduksi ini dapat digunakan sebagai dasar konservasi untuk menjamin keberadaan, ketersediaan dan kesinambungan ikan termasuk ekosistem, jenis dan genetik (PP. RI No. 60/ 2007).
Penelitian tentang biologi reproduksi ikan lais di Indonesia secara lengkap selama satu tahun baru dilakukan pada spesies O. hypophthalmus di Sungai Kampar Riau (Simanjuntak 2007, Elvyra 2009), sedangkan informasi biologi reproduksi ikan famili Silururidae di luar perairan Indonesia telah dilakukan terhadap tiga spesies yaitu: O. pabda, O. bimaculatus dan O. malabaricus di
et al. 2014, Malla dan Banik 2015). Penelitian biologi reproduksi pada berbagai spesies ikan air tawar bersungut (Catfishes) menunjukkan hasil yang berbeda tergantung pada spesies dan letak geografi perairan (Yalcin et al. 2001, Arthi et al. 2013), sehingga hal ini membutuhkan langkah pengelolaan yang berbeda pada masing-masing spesies dan letak georafi perairan. Sampai saat ini penelitian biologi reproduksi spesies O. miostoma sebagai spesies endemik ikan di perairan Sungai Mahakam belum pernah dilakukan.
Berdasarkan hal tersebut di atas maka pengelolaan sumberdaya perikanan berkelanjutan di Sungai Mahakam perlu secepatnya dilakukan untuk mencegah terjadinya penurunan populasi ikan lais. Usaha pengelolaan tersebut akan lebih terarah apabila informasi dasar mengenai ikan lais digali lebih dalam dan rinci. Informasi dasar yang dimaksud adalah jenis dan identitas spesies, karakteristik biometrik dan genetik, distribusi, serta pengetahuan biologi reproduksi dan habitatnya.
Perumusan Masalah
Meningkatnya permintaan konsumen terhadap ikan lais baik sebagai ikan konsumsi maupun sebagai ikan hias, membuat nelayan lebih mengandalkan hasil tangkapan dari alam sehingga pengadaannya di pasar ikan tidak memungkinkan berlangsung secara berkesinambungan dan dapat menimbulkan kekhawatiran terhadap penurunan populasi ikan ini dikemudian hari.
Peningkatan eksploitasi juga diiringi oleh kerusakan lingkungan yang terjadi di Sungai Mahakam dan kawasan sekitarnya, diperkirakan dapat membawa dampak buruk terhadap sumberdaya ikan lais. Di sisi lain informasi secara menyeluruh mengenai ikan lais terutama jenis dan identitas spesies, karakteristik biometrik dan genetik, distribusi, serta pengetahuan biologi reproduksi dan habitatnya masih sangat terbatas. Untuk itu perlu penelitian untuk mendapatkan informasi dasar guna pengelolaan secara berkesinambungan demi menjaga kelangsungan hidup ikan ini di habitatnya.
Tujuan Penelitian
Tujuan penelitian ini adalah untuk mendapatkan karakteristik pembeda antara genus dan spesies ikan lais secara biometrik dan morfologis serta menyusun kunci identifikasi praktis, menentukan identitas spesies yang akurat secara genetik, mendeskripsikan keberadaan spesies secara spasial dan temporal, menyusun strategi dan langkah pengelolaan yang tepat bagi keberlanjutan sumber daya ikan lais khususnya spesies Ompok miostoma melalui kajian biologi reproduksi.
Manfaat Penelitian
Kebaruan (Novelty)
Kebaruan (novelty) pada penelitian ini adalah :
1. Kunci identifikasi ikan lais asal Sungai Mahakam berdasarkan karakter biometrik dan morfologik.
2. Karakter molekuler berupa runutan gen COI ikan lais spesies: O. hypophthalmus, O. miostoma, K. apogon, K. bicirrhis, K. limpok dan
K. micronema sebagai dasar DNA barcoding spesies tersebut.
3. Informasi biologi reproduksi O. miostoma sebagai spesies endemik ikan famili Siluridae di perairan Sungai Mahakam Kalimantan Timur.
2 TINJAUAN PUSTAKA
Jenis dan distribusi ikan lais (famili Siluridae, ordo Siluriformes)
Jenis ikan bersungut air tawar anggota famili Siluridae umum dikenal dengan nama internasional sheat catfishes, sedangkan di Indonesia dikenal dengan nama ikan lais (Kalimantan) atau ikan silais (Sumatera). Ciri khusus famili ini tidak mempunyai sirip lemak, tidak mempunyai duri pada sirip punggung, dan sirip analnya sangat panjang (Kottelat et al. 1993). Di dalam ordo Siluriformes, ciri utama pembeda famili Siluridae dengan famili lainnya berdasarkan kepala dan badan yang lebih pipih, tulang punggung bersifat lentur, duri pada sirip dada lemah, beberapa genus tidak memiliki sirip punggung, satu atau dua pasang sungut pada rahang bawah dan rahang atas, sungut pada rahang atas biasanya lebih panjang, sirip anal sangat panjang terdiri dari 41 - 110 jari-jari lunak dan sirip perut kecil atau tidak ada (Diogo 2005, Nelson 2009).
Menurut Bornbusch (1995), famili Siluridae merupakan ikan air tawar bersungut paling beragam di Eurasia dengan jumlah 100 nominal spesies. Meskipun famili ini tersebar dari Eropa hingga Asia, tetapi keragaman spesies tertinggi ditemukan di kawasan Asia Tenggara (Gambar 2) Sebanyak 32 spesies diketahui dari Indonesia dan lima spesies diketahui dari Pakistan, Afghanistan dan India.
Gambar 2. Peta penyebaran famili Siluridae di Eropa dan Asia
Di Asia Tenggara, famili Siluridae terdapat di perairan tawar Indochina, Semenanjung Malaya hingga pulau-pulau di Indonesia, terutama ditemukan pada sungai-sungai yang bermuara ke arah Laut Cina Selatan dan Selat Malaka. Di Indonesia jenis ikan ini dapat ditemukan di Sumatera, Kalimantan dan Jawa, dengan sebaran utama di Sumatera dan Kalimantan (Kottelat et al. 1993).
et al. 2001) serta perairan di Brunei dan Serawak (Parenti dan Lim 2005). Total delapan genus dan 28 spesies ikan famili Siluridae telah diidentifikasi dari Kalimantan oleh Kottelat et al. (1993) yaitu: Belodontichthys dinema,
Ceratoglanis scleronema, Hemisilurus heterorhyncus, H. moolenburghi, K. apogon, K. bicirrhis, K. cryptopterus, K. platypogon, K. lais, K. limpok, K. lumholtzi, K. macrocephalus, K. micronema, K. minor, K. parvanalis, K. schilbeides, O. bimaculatus, O. borneensis, O. eugeneiatus,O. hypophthalmus,
O. leiacanthus, O. sabanus, O. weberi, Pterocryptis furnessi, Silurichthys haselti, S. phaoisoma, S. sanguineus, Wallago attu. Sebanyak 13 spesies ditemukan di DAS Mahakam termasuk K. apogon, K. bicirrhis, K. limpok, K. macrocephalus, K. micronema, O. miostoma, O. hypophthalmus, O. sabanus, O. jaynei, O. sp. nov., Wallago cf. leerii, W. miostoma dan Hemisilurodes sp. (Christensen 1992, Kottelat 1994).
Di Sungai Mahakam ikan lais umum dikenal masyarakat dengan nama lokal
lais lepok (O. miostoma), lais kembang (O. hypophthalmus), lais bentilap (K. apogon), lais jungkang (K. micronema) dan beberapa spesies lainnya hanya di
beri nama lais. Ikan lais genus Ompok mempunyai beberapa nama sinonim yaitu: Silurus, Callichrous, Pseudosilurus, Wallago, Silurichthys, Silurodes, Wallagonia dan Beladonicthys, sedangkan genus Kryptopterus mempunyai nama sinonim Cryptopterus, Kryptopterichthys, Phalacronotus, Micronema dan Hemisilurus (Froese dan Pauly 2015).
Ikan lais dari genus Kryptopterus dan Ompok memiliki ciri-ciri mofologis mirip dan hanya dibedakan dari kehadiran sirip punggung (Kottelat et al. 1993). Perbedaan ciri-ciri morfologi antara Ompok spp. dan Kryptopterus spp. (Tabel 1 dan Gambar 3).
Tabel 1. Ciri-ciri morfologis ikan lais genus Ompok dan Kryptopterus. Disarikan dari: (Weber dan de Beaufort 1913, Kottelat et al. 1993)
Karakter morfologis Ompok spp. Kryptopterus spp.
Bentuk tubuh Pipih (compressed)
memanjang tidak bersisik
Sangat pipih (strongly
compressed), memanjang tidak bersisik
Mulut Berbentuk miring (oblique) Berbentuk miring (oblique)
Rahang Rahang bawah lebih
panjang dari pada rahang atas
Rahang atas dan bawah seimbang
Sungut Sepasang pada rahang atas
dan sepasang pada rahang bawah
Sepasang pada rahang atas dan sepasang pada rahang bawah yang umumnya pendek atau rudimenter
Sirip punggung Tanpa duri (spina) terdiri
atas 3-4 jari-jari
Tidak ada (rudimenter)
Sirip dada Mempunyai duri Tanpa duri
Sirip perut Terdiri dari 7-8 jari-jari Terdiri dari 5-10 jari-jari
Sirip lemak (adipose) Tidak ada Tidak ada
Sirip dubur Panjang, bersambung atau
tidak bersambung dengan sirip ekor
Panjang, bersambung atau tidak bersambung dengan sirip ekor
Sirip ekor Bentuknya bercabang
(forked)
Bentuknya bercabang
a. Holotype Ompok miostoma b. Ompok hypophthalmus
c. Kryptopterus limpok d. Kryptopterus apogon
e. Kryptopterus bicirrhis f. Kryptoperus micronema
Gambar 3. Ilustrasi spesies ikan lais famili Siluridae. Sumber: (Bleeker 1858) Karakter morfologis ikan
Kajian karakter morfologi telah lama digunakan dalam biologi perikanan untuk mengukur jarak dan hubungan kekerabatan dalam pengkategorian variasi dalam taksonomi. Hal ini banyak membantu dalam menyediakan informasi untuk ragam dan kekayaan jenis ikan. Meskipun demikian pembatas utama dari karakter morfologi dalam tingkat intraspesies (ras) adalah variasi fenotip tidak selalu tepat di bawah kontrol genetik tapi dipengaruhi oleh perubahan lingkungan. Ragam fenotip dari ikan memungkinkan ikan dalam merespon secara adaptif perubahan dari lingkungan melalui modifikasi fisiologi dan kebiasaan (Turan 1999).
diameter mata (DMT), (17) lebar tulang vomer (LTV), (18) tinggi tulang vomer (TTV), (19) panjang postokular (PPO), (20) lebar tubuh (LTB), (21) panjang snout isthmus (PSI), (22) panjang interorbital (PIO), (23) lebar mulut (LBM), (24) panjang rahang bawah (PRB), (25) panjang snout (PSN), (26) panjang kumis maksila (PKM), (27) panjang kumis mandibula (PKD). Meristik adalah ciri yang berkaitan dengan jumlah bagian tubuh ikan seperti jumlah sisik pada garis rusuk, jumlah tapis insang, jumlah jari branchiostegal, pori-pori pada kepala, jumlah jari-jari keras dan lemah pada sirip dorsal, sirip pektoral, sirip pelvik, sirip anal, sirip kaudal (Affandi et al. 1992).
DNA mitokondria ikan
Mitokondria merupakan organel sel penghasil energi yang terdapat di dalam sitoplasma. DNA mitokondria (mtDNA) memiliki sejumlah sifat genetik khas yang membedakannya dari genom inti. Genom mitokondria berbentuk sirkuler, beruntai ganda, memiliki panjang sekitar 16,5 kb yang mengandung basa guanine (G) dan cytosine (C) berkisar antara 32 - 45,6%. Kedua basa tersebut menyebar tidak merata di antara kedua untai DNA. Distribusi asimetris nukleotida menimbulkan heavy strand (untai berat) dan light strand (untai ringan) ketika molekul mtDNA dipisahkan dalam gradien basa CsCl. Heavy strand atau untai H berisi lebih banyak nukleotida guanine (G) yang mempunyai berat molekul terbesar di antara keempat nukleotida, sedangkan Light strand atau untai L berisi lebih sedikit basa guanine (G) tetapi mengandung banyak basa nukleotida Adenine (A) dan Thymine (T) (Taylor dan Turnbull 2005).
MtDNA merupakan DNA yang padat gen dan tidak mempunyai intron, berukuran sebesar kurang lebih 16.569 pasang basa (pb) yang membentuk 37 gen terdiri dari 13 polipeptida untuk protein kompleks rantai respirasi, 22 tRNA dan 2 rRNA (12S dan 16S) yang berfungsi dalam proses sintesis protein mitokondria (Andersons et al. 1981). Ketiga belas polipeptida meliputi tujuh subunit (ND1, 2, 3, 4, 4L, 5 dan 6) dari kompleks I rantai respirasi, satu subunit (Cytochrome b) dari kompleks III, tiga subunit (Cytochrome c oxidase subunit I, II, III) dari kompleks IV dan dua subunit (ATP ase 6 dan ATP ase 8) dari kompleks V (Attardi et al. 1987). Terdapat satu daerah pengontrol (control region) yang tidak menyandi (non coding region), disebut sebagai displacement loop (D-loop). Daerah D-loop merupakan struktur beruntai tiga (triple stranded), terdiri dari 1.122 pb, mempunyai titik asal replikasi rantai H (OH) dan 2 promotor utama
untuk untai H dan L (PH dan PL). Transkripsi MtDNA diinisiasi dari dua promotor
yang ada di daerah D-loop yaitu PH dan PL, transkripsi berjalan mengelilingi
mtDNA sehingga membentuk RNA. D-loop merupakan daerah kontrol utama ekspresi mtDNA, dan berfungsi sebagai promotor utama transkripsi (Clayton 1991).
Jumlah molekul mtDNA di dalam sel sangat bervariasi. Rata-rata terdapat 4 - 5 salinan molekul mtDNA per mitokondria. Setiap sel dapat berisi ratusan mitokondria, bahkan dapat sampai beberapa ribu mitokondria seperti pada kasus sel telur (ovum). Namun diperkirakan rata-rata terdapat sekitar 500 mtDNA di sebagian besar sel. Hal tersebut menjadikan keberhasilan amplifikasi mtDNA dalam setiap sel lebih besar (relatif terhadap penanda DNA nukleus) dengan sampel biologis yang mungkin telah rusak karena panas atau kelembaban (Butler 2005). Perbandingan karakteristik dasar DNA inti dan DNA mitokondria (Tabel 2).
Tabel 2. Karakteristik DNA inti dan DNA mitokondria
Karakteristik DNA inti DNA mitokondria
Ukuran genom ~ 3,2 milyar pb ~ 16.569 pb
Total DNA/sel 99,75% 0,25%
Jumlah kopi/sel 2 (1 alel setiap tetua) Dapat >1000 Struktur Linier; dikemas dalam
kromosom
Sirkuler Diturunkan dari Ibu dan bapak Ibu
Pasangan kromosom Diploid Haploid
Rekombinasi generasi Ya Tidak
Replikasi Ya Ya
Keunikan Unik untuk setiap
individu
(kecuali kembar identik)
Unik untuk setiap individu
(relatif sama dengan ibu)
Laju mutasi Rendah ± 5 - 10 kali lipat dari
DNA inti Sumber: (Butler 2005)
DNA mitokondria bersifat unik dan berbeda dengan DNA inti karena memiliki laju mutasi yang tinggi yaitu sekitar 5 - 10 kali DNA inti (Butler 2005). Tingginya laju mutasi mt DNA disebabkan oleh enzim DNA polimerase alfa yang digunakan dalam proses replikasi mtDNA tidak memiliki mekanisme reparasi (perbaikan) yang efisien (prooffreading) sehingga tidak dapat mengoreksi kesalahan-kesalahan selama proses replikasi (Watson et al. 1997).
MtDNA mempunyai beberapa kelebihan yang menjadikannya banyak digunakan untuk mengidentifikasi keanekaragaman genetik dan populasi. Beberapa kelebihan tersebut adalah (1) memiliki ukuran yang kompak dan relatif kecil (16.000 – 20.000 pasang basa), tidak sekomplek DNA inti sehingga dapat dipelajari sebagai satu kesatuan yang utuh; (2) berevolusi lebih cepat dan dapat dibandingkan dengan jelas perbedaan antara populasi dan hubungan kekerabatannya; (3) hanya sel telur yang menyumbangkan material mitokondria sehingga mtDNA hanya diturunkan dari induk betina; dan (4) bagian-bagian dari genom mitokondria berevolusi dengan laju yang berbeda, sehingga dapat berguna untuk studi sistematika dan penelusuran kesamaan asal-usul (Solihin 1994).
sistematika dan biogeografi. Umumnya, gen-gen yang terkonservasi dengan baik (berevolusi lambat) dapat dijadikan dasar penelusuran asal-usul atau filogeni. Sebaliknya, gen-gen yang tidak terkonservasi dengan baik (berevolusi cepat) dapat digunakan untuk perbandingan galur-galur baru (Solihin 1994).
Pada ordo Siluriformes genom mtDNA lengkap telah dilaporkan sebanyak tujuh spesies yaitu: Pseudobagrus tokiensis (kode akses GB: AB054127) (famili Bagridae), Corydoras rabauti (kode akses GB: AB054128) (famili Callichthyidae), Ictalurus punctatus (kode akses GB: AF482987) (famili Ictaluridae), Cranoglanis bouderius (kode akses GB: AY898626) (famili Cranoglanididae), Pangasianodon gigas (kode akses GB: AY762971) (famili Pangasiidae), Liobagrus obesus (kode akses GB: DQ321752) (famili Amblycipitidae), dan Silurus glanis (kode akses GB: AM398435) (famili Siluridae) (Vittas et al. 2011). Namun sampai saat ini genom mtDNA lengkap famili Siluridae pada genus Ompok dan Kryptopterus masih belum tersedia (Gen Bank 2015).
Genom mtDNA lengkap European catfish Silurus glanis (famili Siluridae)
memiliki panjang 16.526 pb, terdiri dari 37 gen yaitu: 13 gen penyandi protein, 2 RNAs ribosomal (rRNA), 22 tRNAs dan satu daerah kontrol bukan penyandi protein (non coding) (Gambar 4).
Gambar 4. Organisasi genom mitokondria Silurus glanis
Keterangan: gen untuk protein dan rRNA ditampilkan dengan singkatan standar, gen untuk tRNA ditampilkan dengan huruf tunggal untuk Asam Amino yang
sesuai. Sumber: (Vittas et al. 2011).
Heavy (H) Strand
Untai H atau untai berat mtDNA terdiri atas 2 RNAs ribosomal (12S rRNA dan 16S rRNA); 12 gen penyandi protein masing-masing NADH dehidrogenase (ND1, ND2, ND3, ND4, ND5, ND4L), sitokrom c oksidase (COI, COII, COIII), sitokrom b (Cyt b), ATPase (ATP6, ATP8); dan 14 tRNA masing-masing tRNA fenil alanin (tRNAPhe), valin (tRNAVal), leusin (tRNALeu), isoleusin (tRNAIle), metionin (tRNAMet), triptofan (tRNATrp), asam aspartat (tRNAAsp), lisin (tRNALys), glisin (tRNAGly), arginin (tRNAArg), histidin (tRNAHis), serin (tRNASer), leusin (tRNALeu) dan treonin (tRNAThr), pada untai L atau untai ringan mtDNA mengandung sisanya yaitu 1 gen penyandi protein NADH dehidrogenase 6 (ND6); dan 8 tRNA yaitu tRNA glutamin (tRNAGln), alanin (tRNAAla), asparagin (tRNAAsn), sistein (tRNACys), tirosin (tRNATyr), serin (tRNASer), asam glutamat (tRNAGlu), dan prolin (tRNAPro) (Vittas et al. 2011).
Gen COI sebagai DNA barcoding
Gen COI merupakan salah satu gen yang terdapat dalam genom mitokondria. Dalam studi molekuler, gen COI digunakan sebagai penanda genetik untuk mempelajari karakteristik genetik antar spesies. Gen COI juga dapat digunakan sebagai DNA barcodingkarena sedikit sekali delesi dan insersi dalam sekuennya, serta variasi juga sedikit ditingkat spesies (Hebert et al. 2003). Dengan demikian gen COI ini dapat digunakan untuk merekonstruksi filogenetik pada cabang evolusi tingkat spesies. Gen ini telah berhasil mengidentifikasi maupun membedakan spesies berbagai invertebrata dan vertebrata, dari Lepidoptera sampai Aves dan dalam sebaran geografi berbeda (Hajibabaei et al. 2007). Penelitian keragaman genetik menggunakan gen COI juga telah dilakukan untuk identifikasi spesies pada berbagai ikan air laut dan air tawar termasuk silurid catfish (Hebert et al. 2003, Hardman 2004, Ward et al. 2005, Lakra et al. 2011, Malakar et al. 2012).
Gen COI memiliki kelebihan untuk digunakan dalam mengakurasi spesies dibandingkan dengan gen Cyt b. Salah satu kelebihan gen COI sebagai penanda analisis filogeni adalah asam amino pada fragmen COI jarang mengalami substitusi. Namun, basa-basa pada triple kodonnya terutama pada kodon ketiga masih berubah dan perubahannya bersifat silent (Lynch dan Jarrell 1993).
identifikasi spesies dan golongan taksonominya (spesies, famili), 4) harus cukup kuat, dengan lokasi primer yang terkonservasi, dan tingkat amplifikasi dan sekuensing DNA yang dapat dipercaya, 5) sasaran daerah DNA harus cukup pendek untuk memudahkan amplifikasi DNA yang terdegradasi.
Berdasarkan hasil penelitian Malakar et al. (2012), ukuran gen COI parsial
DNA mitokondria pada tiga spesies Ompok (O. pabda, O. pabo, dan O. bimaculatus) berasal dari India yang diamplifikasi menggunakan primer Fish
F1 dan Fish R1 adalah sepanjang 655 pb. Pada spesies O. pabda diperoleh situs terkonservasi 651 pb (99,3%), situs bervariasi 4 pb (0,61%), dan situs parsimoni informatif 4 pb (0,61%); pada spesies O. pabo terkonservasi 651 pb (99,3%), situs bervariasi 4 pb (0,61%), dan situs parsimoni informatif 4 pb (0,61%); pada spesies O. bimaculatus diperoleh situs terkonservasi 650 pb (99, 2%), situs bervariasi 5 pb (0,76%) dan situs parsimoni informatif 3 pb (0,45%).
Secara keseluruhan rata-rata jarak genetik intraspesies dalam penelitian yang dilakukan oleh Malakar et al. (2012) adalah 0,35%, sedangkan rata-rata jarak genetik interspesies adalah 13,1%. Lebih lanjut dikatakan rata-rata jarak genetik interspesies tertinggi ditemukan antara O. pabda dan O. pabo sebesar 15,3% , sedangkan rata-rata jarak genetik interspesies terendah ditemukan antara O. pabo dan O. bimaculatus sebesar 10,2%.
Biologi reproduksi ikan
Reproduksi merupakan aspek yang penting dalam pengelolaan suatu sumber daya perairan. Keberhasilan suatu spesies ikan dalam daur hidupnya ditentukan dari kemampuan anggotanya untuk bereproduksi pada lingkungan yang berfluktuasi untuk menjaga keberadaan populasinya (Moyle dan Cech 2004). Reproduksi pada ikan merupakan suatu cara yang dilakukan oleh ikan untuk mendapatkan keturunan dan menjamin keberlangsungan hidup spesiesnya serta merupakan tahapan penting dalam siklus hidupnya (Effendi 2002).
Faktor-faktor lingkungan yang mengontrol siklus reproduksi ikan di perairan terdiri atas faktor fisika, kimia dan biologi. Pada ikan yang hidup di daerah tropis, faktor fisika yang mengontrol siklus reproduksi adalah arus, suhu dan substrat. Faktor kimia adalah gas-gas terlarut, pH, nitrogen dan metabolitnya serta zat buangan berbahaya bagi kehidupan ikan di perairan. Faktor biologi yang mengontrol siklus reproduksi ikan dibagi menjadi faktor biologi internal dan eksternal. Faktor biologi internal terdiri atas faktor fisiologi individu dan respon terhadap berbagai faktor lingkungan, sedangkan faktor biologi eksternal meliputi patogen, predator dan kompetisi sesama spesies ikan tertentu atau dengan spesies lain (Bye 1984).
Reproduksi pada ikan berkaitan erat dengan kemampuan untuk bertelur (fekunditas) dan organ reproduksi utama (gonad) sebagai alat reproduksi seksualnya. Menurut Nikolsky (1963), aspek biologi reproduksi terdiri atas faktor kondisi, nisbah kelamin, tingkat kematangan gonad, indeks kematangan gonad, fekunditas, sebaran diameter telur, tipe pemijahan, waktu pemijahan, ukuran ikan pertama kali matang gonad (IKG) dan lokasi memijah.
Umumnya strategi reproduksi ditujukan untuk terjaminnya keamanan tempat meletakkan telur, mencari waktu yang tepat untuk ketersediaan makanan yang maksimum untuk anak-anak ikan nantinya dan menghindari pemangsaan oleh predator terhadap anak-anak ikan (Welcomme 1979).
Induk ikan yang sudah matang gonad bermigrasi (beruaya) dari sungai menuju daerah rawa banjiran, terutama hutan rawa yang banyak ditumbuhi tanaman dengan substrat keras, seperti pohon-pohon yang sudah mati sebagai tempat menempelkan telur. Penelitian yang dilakukan oleh Pellokila (2009) terhadap biologi reproduksi ikan betok (Anabas testudineus Bloch, 1792) di rawa banjiran DAS Sungai Mahakam menemukan bahwa ikan betok jantan dan betina mencapai TKG III dan IV terjadi pada bulan November - Januari di setiap stasiun penelitian, sehingga diduga bahwa dalam jangka waktu tersebut ikan betok sudah memasuki musim pemijahan. Persentase jumlah ikan jantan dan betina tertinggi yang telah matang gonad (TKG III dan IV) terdapat di stasiun rawa dan yang terendah terdapat di stasiun sungai pada setiap bulan penelitian. Puncak pemijahan ikan betok diduga terjadi pada bulan Desember, saat kenaikan permukaan air tertinggi akibat curah hujan yang cenderung lebih tinggi dibandingkan bulan-bulan penelitian lainnya.
A. Faktor kondisi
Faktor kondisi adalah keadaan yang menyatakan kemontokan ikan secara kualitas dan perhitungannya didasarkan pada panjang dan berat ikan (Royce 1972). Faktor kondisi atau indek ponderal sering disebut sebagai faktor K merupakan hal penting dari pertumbuhan ikan, karena faktor kondisi dapat digunakan untuk menganalisis populasi. Faktor kondisi menunjukkan keadaan baik dari ikan dilihat dari segi kapasitas fisik untuk survival dan reproduksi.
Beragamnya faktor kondisi disebabkan oleh pengaruh makanan, umur, jenis kelamin dan kematangan gonad (Effendie 2002). Berdasarkan hasil penelitian Simanjuntak (2007), ikan silais (O. hypophthalmus) memiliki nilai faktor kondisi relatif berkisar antara 0,70 – 2,51, faktor kondisi ikan jantan berkisar antara 0,70 - 2,51 dan ikan betina berkisar antara 0,73 – 1,34. Selanjutnya Makri (2011) menyatakan faktor kondisi ikan lais (K. limpok) jantan dan betina bervariasi setiap bulan, faktor kondisi ikan lais jantan mempunyai kisaran antara 1,0 – 1,03 dan ikan lais betina berkisar antara 1,01 – 1,02 dan faktor kondisi tertinggi ikan lais jantan maupun betina terjadi pada bulan Juli. Wotton dan Potts (1984) menyatakan bahwa pertumbuhan ovarium dan pemijahan memiliki kaitan yang erat dengan penurunan pertumbuhan somatik atau faktor kondisi. Penurunan ini setara dengan besarnya energi yang diperlukan untuk memproduksi ovum.
B. Nisbah kelamin
Nisbah kelamin diduga mempunyai keterkaitan dengan habitat suatu spesies ikan. Nikolsky (1969) menyatakan bahwa perbandingan kelamin dapat berubah menjelang dan selama musim pemijahan. Ikan jantan lebih banyak mengalami perubahan nisbah kelamin secara teratur dalam ruaya pemijahannya. Pada awalnya jumlah ikan jantan lebih banyak dibandingkan dengan ikan betina, kemudian nisbah kelamin berubah menjadi 1:1 diikuti dengan dominasi ikan betina.
Perbandingan nisbah kelamin 1 : 1 yaitu 50% jantan dan 50% betina merupakan kondisi ideal untuk mempertahankan spesies. Namun pada kenyataannya di alam, perbandingan nisbah kelamin tidaklah mutlak. Hal ini dipengaruhi oleh pola distribusi yang disebabkan oleh ketersediaan makanan, kepadatan populasi, dan keseimbangan rantai makanan (Effendie 2002). Selain itu juga dipengaruhi oleh adanya pola tingkah laku ikan, perbedaan laju mortalitas, dan pertumbuhan (Turkmen et al. 2002). Penyimpangan dari kondisi ideal tersebut disebabkan oleh faktor tingkah laku ikan itu sendiri, perbedaan laju mortalitas dan pertumbuhannya. Keseimbangan rasio kelamin dapat berubah menjelang pemijahan. Pada waktu melakukan ruaya pemijahan, populasi ikan didominasi oleh ikan jantan, kemudian menjelang pemijahan populasi ikan jantan dan betina dalam kondisi seimbang, lalu didominasi oleh ikan betina (Agus 2008). Komposisi jantan dan betina dalam populasi merupakan faktor penting untuk kelestarian populasi.
Perbandingan jenis kelamin dapat digunakan untuk menduga keberhasilan pemijahan, yaitu dengan melihat perimbangan jumlah ikan jantan dan ikan betina di suatu perairan, juga berpengaruh terhadap produksi, rekruitmen, dan konservasi sumber daya ikan tersebut (Effendie 2002). Untuk mempertahankan keberlangsungan spesies, perbandingan hewan jantan dan betina diharapkan seimbang. Rasio jantan lebih tinggi dapat mengganggu kelestarian spesies dengan asumsi bahwa peluang jantan untuk melakukan perkawinan dan menghasilkan keturunan akan lebih rendah karena jumlah hewan betina yang terdapat dalam populasi tersebut lebih sedikit. Gangguan pada kelestarian populasi ini kemungkinan dapat lebih buruk jika terjadi penangkapan spesies tertentu saja oleh manusia (Candramila dan Junardi 2012).
jenis kelamin jantan dalam populasi karena berpengaruh pada perkembangan seksual dan diferensiasi seks (Shang et al. 2006).
C. Tingkat kematangan gonad
Kematangan gonad suatu individu sangat menentukan reproduksi dari individu ikan tersebut. Pembuahan dapat terjadi apabila gonadnya sudah benar-benar matang sempurna. Ikan yang telah dewasa ditandai dengan kematangan gonad dan didukung dengan ukuran panjang serta bobotnya. Pada saat ikan mulai berkembang, gonad betina (telur) mulai terlihat berwarna kuning dan memenuhi rongga tubuh saat memasuki tahap matang, sedangkan gonad jantan (testis) akan berwarna putih pada saat matang (Royce 1972).
Tingkat kematangan gonad (TKG) merupakan tahap perkembangan gonad sebelum dan sesudah ikan memijah (Effendie 1979). Perkembangan gonad merupakan bagian dari proses reproduksi. Terjadinya perkembangan gonad ini sebagai akibat adanya proses vitellogenesis, yaitu proses akumulasi di dalam sel telur (Effendie 2002). Pencatatan perubahan atau tahap-tahap kematangan gonad diperlukan untuk mengetahui perbandingan ikan-ikan yang akan melakukan reproduksi atau tidak. Berdasarkan tahap kematangan gonad juga dapat diketahui kapan ikan akan memijah, baru memijah atau sudah memijah. Pendugaan puncak pemijahan dapat dilakukan berdasarkan persentase jumlah ikan yang matang gonad pada suatu waktu (Sulistiono et al. 2006).
Faktor utama yang memengaruhi kematangan gonad ikan antara lain suhu, dan makanan selain faktor keberadaan hormon (Tang dan Affandi 2000). Penelitian Ernawati (2002) menemukan implan hormon dapat mempercepat kematangan gonad ikan jambal siam (Pangasius hypopthalmus). Dalam reproduksi, sebelum pemijahan ukuran gonad akan mencapai puncaknya pada waktu ikan akan memijah, kemudian menurun dengan cepat selama pemijahan berlangsung sampai selesai (Effendie 1979). Parameter fisika juga dapat berpengaruh terhadap pemijahan ikan, misalnya posisi matahari dan fase bulan yang dapat memengaruhi pemijahan misalnya pada ikan giru (Amphiprion percula) (Yusuf et al. 2001).
Lagler et al. (1977) menyatakan bahwa perkembangan gonad terdiri atas dua tahap yaitu tahap pertumbuhan gonad hingga mencapai dewasa dan tahap pematangan gonad. Tahap pertama dimulai sejak ikan baru menetas hingga mencapai dewasa kelamin dilanjutkan dengan tahap pematangan seksual dan terus berlangsung sampai reproduksi berakhir. Selama proses perkembangan alat kelamin, ovarium dan testis akan mengalami serangkaian perubahan morfologis dan histologis. Selanjutnya dikatakan bahwa ada dua faktor yang memengaruhi saat kali pertama ikan matang gonad yaitu: faktor dalam dan faktor luar. Faktor dalam antara lain: perbedaan spesies, umur, ukuran serta sifat-sifat fisiologi dari ikan tersebut seperti kemampuan adaptasi terhadap lingkungan, sedangkan faktor luar yang memengaruhinya yaitu: makanan, suhu, arus, dan adanya individu yang berlainan jenis kelamin dan tempat berpijah yang sama.
betina di perairan Tripura India mulai matang gonad pada ukuran 17,0 cm dan jantan 16,5 cm (Gupta et al. 2014, Malla dan Banik 2015). Di Indonesia, Elvira
(2009) menemukan rata-rata panjang dan berat tubuh ikan lais betina (O. hypophthalmus) matang gonad (TKG IV) adalah 24,9 ± 1,57 cm dan berat
74,26 ± 12,40 gram. Sementara itu, rata-rata ikan lais jantan matang gonad adalah pada ukuran panjang 25,9 ± 1,88 cm dan berat 79,80 ± 20,49 gram.
Hormon juga memengaruhi perkembangan gonad secara langsung maupun tidak langsung. Tang dan Affandi (2000) menyatakan bahwa aktifitas hormon gonadotropin memengaruhi pematanggan oosit secara tidak langsung. Keberadaan ikan TKG III dan IV di suatu daerah mengindikasikan bahwa adanya ikan yang memijah di daerah tersebut.
D. Indeks kematangan gonad (IKG)
Indek kematangan gonad (IKG) merupakan nilai perbandingan antara berat gonad dengan berat tubuh ikan yang dinyatakan dengan angka persen. IKG juga dapat menggambarkan ukuran ikan pada saat memijah. IKG memiliki nilai maksimum pada waktu akan terjadi pemijahan, dan kisaran IKG ikan betina cenderung lebih besar dibandingkan dengan kisaran IKG pada ikan jantan (Effendi 2002). Gupta et al. (2014) mendapatkan nilai kisaran IKG O. pabda di Sungai Gomti India pada betina 0,5 - 8,3 dan jantan 0,1 - 1,8.
Pada tingkat kematangan gonad yang sama, IKG ikan jantan akan berbeda dengan IKG ikan betina. Umumnya IKG ikan betina akan lebih besar dari pada IKG ikan jantan dan rata-rata IKG cendrung meningkat ketika ikan mencapai dewasa dan menurun setelah memijah dan selama fase istirahat. Sebagai contoh, rata-rata maksimum IKG O. bimaculatus selama priode pemijahan di perairan Tripura India pada ikan betina 15,58 lebih besar dari ikan jantan 2,19 (Malla dan Banik 2015).
E. Fekunditas
Fekunditas merupakan jumlah telur masak sebelum dikeluarkan pada waktu ikan memijah (Effendie 2002). Menurut Nikolsky (1969), fekunditas individu adalah jumlah telur dari generasi tahun itu yang dikeluarkan pada tahun itu pula. Royce (1972) mengemukakan bahwa fekunditas total diartikan sebagai jumlah telur yang dihasilkan oleh ikan selama hidupnya, sedangkan fekunditas relatif adalah jumlah telur persatuan bobot atau panjang.
sedangkan spesies yang fekunditasnya kecil biasanya melindungi telurnya dari pemangsa atau menempelkan telurnya pada tanaman atau habitat lainnya (Nikolsky 1963).
Fekunditas ikan dapat ditentukan dengan metode gravimetrik yaitu suatu metode dengan cara mengukur berat gonad. Contoh ikan dibandingkan dengan berat gonad total, kemudian dikalikan dengan jumlah telur contoh. Menurut Elvira (2009), nilai fekunditas rata-rata pada O. hypophthalmus di Sungai Kampar Riau adalah sebesar 6.635 butir telur dengan diameter telur 0,77 mm.
F. Diameter telur dan tipe pemijahan
Diameter telur adalah garis tengah atau ukuran panjang suatu telur yang diukur dengan mikrometer berskala yang sudah ditera. Ukuran diameter telur dipakai untuk menentukan kualitas telur (Effendie 2002). Telur yang berukuran besar akan menghasilkan larva ikan yang berukuran lebih besar dibanding dengan larva ikan dengan telur yang lebih kecil. Perkembangan diameter pada telur yang semakin meningkat mengindikasikan meningkatnya tingkat kematangan gonad.
Masa pemijahan setiap spesies ikan berbeda-beda, ada pemijahan yang berlangsung singkat (total spawning), tetapi banyak pula dalam waktu yang panjang (partial spawning) ada pada ikan yang berlangsung sampai beberapa hari. Semakin meningkat tingkat kematangan, garis tengah telur yang ada dalam ovarium semakin besar pula (Effendie 1979). Lama pemijahan dapat diduga dari frekuensi ukuran diameter telur. Ovarium yang mengandung telur matang berukuran sama, menunjukkan waktu pemijahan yang pendek, sebaliknya waktu pemijahan yang panjang dan terus menerus ditandai dengan banyaknya ukuran telur yang berbeda di dalam ovarium. Pada spesies O. bimaculatus dari India mempunyai diameter telur berkisar dari 0,827 – 1,358 mm dan tipe pemijahan terjadi secara bertahap selama priode bulan Mai hingga Agustus (Malla dan Banik 2015).
Pemijahan ikan lais O. hypophthalmus di Sungai Kampar Riau berlangsung sebelum terjadinya banjir maksimum. Hal ini merupakan strategi adaptasi ikan lais untuk mendapatkan keuntungan dari sumber daya makanan alami yang tersedia bagi juvenil ikan. Beberapa bulan kemudian ketika musim banjir besar tiba, anak-anak ikan lais sudah dapat mengkonsumsi anak-anak spesies ikan lain yang baru menetas (Simanjuntak 2007).
Gambaran umum perairan di Sungai Mahakam
Daerah aliran sungai (DAS) Mahakam merupakan salah satu kawasan di Kalimantan Timur yang memiliki luas 8,2 juta hektar atau sekitar 41% dari luas wilayah Provinsi Kalimantan Timur. DAS Mahakam dengan luas 77.095.460 ha meliputi Kabupaten Kutai Barat, Kutai Timur, Malinau, Kutai Kertanegara dan Kota Samarinda. Bahkan daerah tangkapan airnya tidak hanya di Provinsi Kalimantan Timur, namun juga di Provinsi Kalimantan Tengah dan diduga sebagian kecil di Serawak yang merupakan negara bagian Malaysia.
Belayan (10.350 km), subDAS Kedang Kepala dan Kedang Rantau (20.190 km), dan subDAS Mahakam Hilir (6.910 km) (Fakhrudin dan Kosman 1997).
DAS Mahakam dengan seluruh subDASnya berperan penting bagi kehidupan masyarakat dan berfungsi strategis secara ekonomis maupun ekologis (hidrologis). Secara ekonomis, DAS Mahakam telah dimanfaatkan sebagai jalur transportasi kapal penumpang dan kapal pengangkut barang, industri yang berkaitan dengan basil hutan (kehutanan), pertambangan (batu bara, emas, dan minyak bumi), pertanian dan perkebunan (tanaman pangan dan kelapa sawit), budidaya perikanan dan sumber bahan baku air bersih, serta untuk pariwisata (ekowisata). Selain itu, pinggiran Sungai Mahakam (kanan-kiri) dimanfaatkan sebagai permukiman dan usaha ekonomi penduduk setempat. DAS Mahakam juga berfungsi secara ekologis dan berperan penting dalam pengaturan hidrologi, yaitu sebagai pengendali banjir dengan berfungsinya tiga danau (Danau Jempang, Semayang, dan Melintang) sebagai tempat resapan air yang dapat menciptakan keseimbangan lingkungan antara ekosistem danau dan aliran sungai secara fisik (daerah pengaliran sungai). SubDAS Danau Semayang, Melintang, dan Jempang berperan penting dalam pengendalian banjir yang terjadi pada Sungai Mahakam. Ketiga danau tersebut akan berfungsi sebagai daerah resapan atau penghambat lajunya air menuju outlet ke Sungai Mahakam. Danau Semayang dan Melintang merupakan danau dangkal yang mempunyai fluktuasi tinggi permukaan air yang dipengaruhi oleh musim (curah hujan) dan merupakan tipe danau paparan banjir (flood plain). Selama