Kajian efek sinergi antimikroba metabolit bakteri asam laktat dan monoasilgliserol minyak kelapa terhadap mikroba patogen pangan
Teks penuh
Gambar
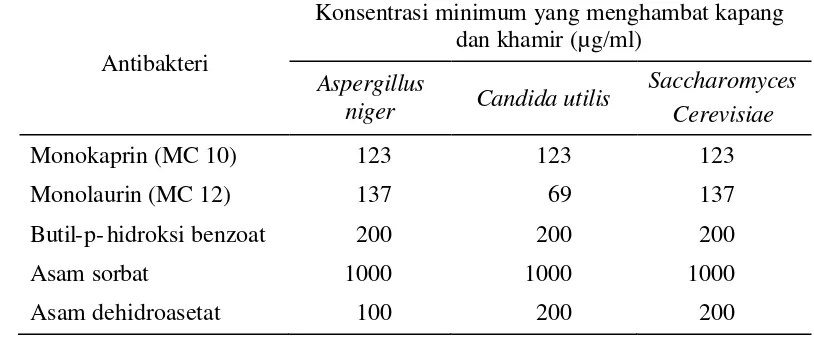
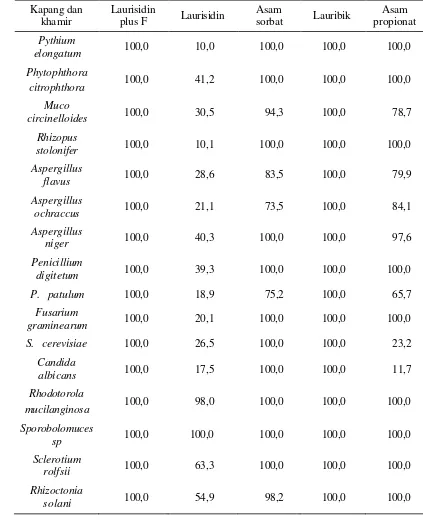
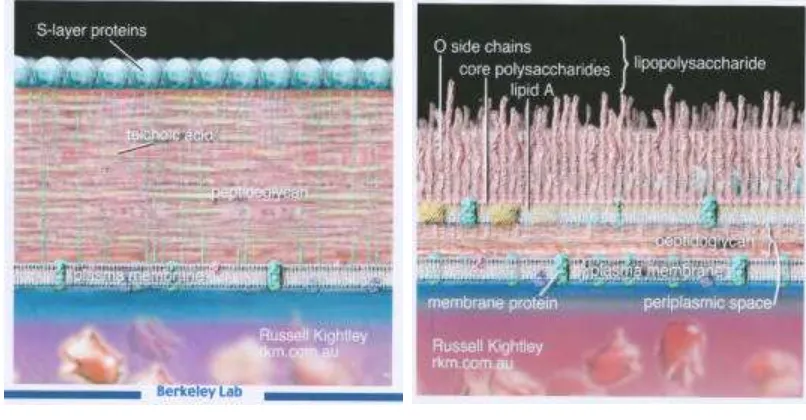

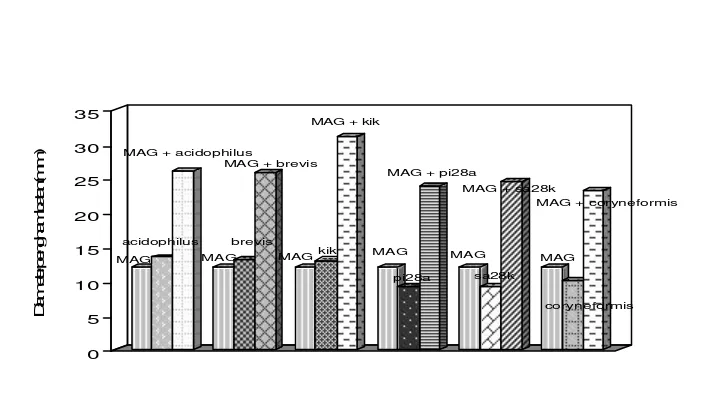
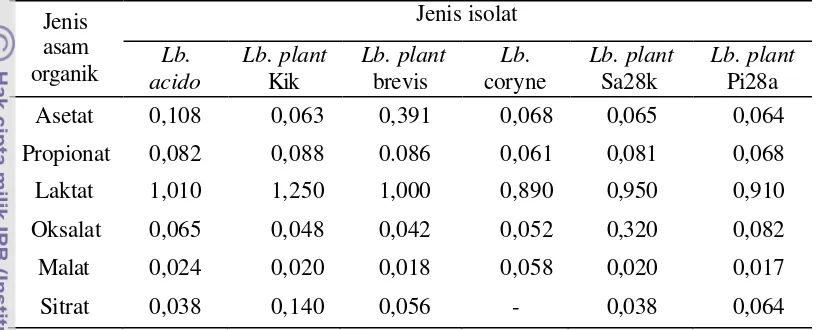
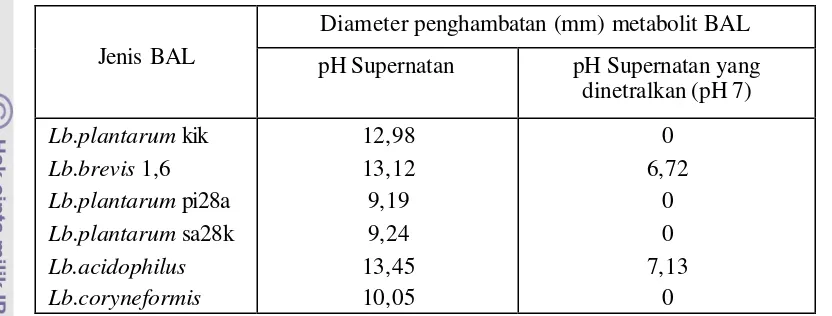
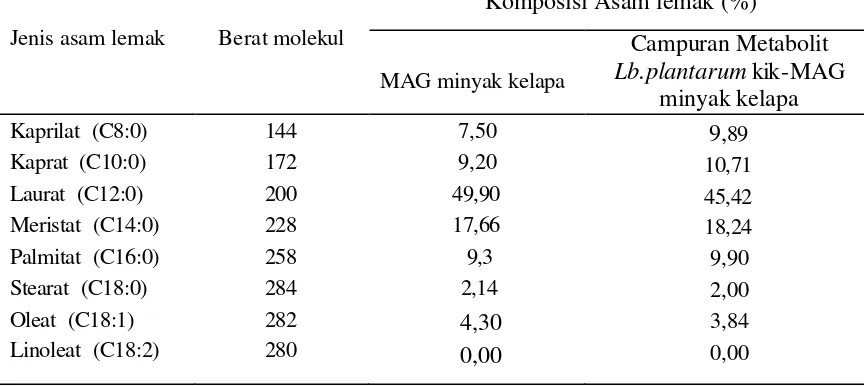
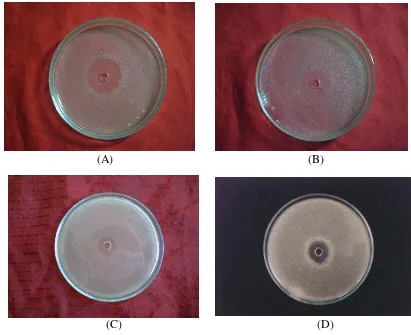
Dokumen terkait
Snakes and Ladders Terhadap Kompetensi Pengetahuan Matematika Siswa 104 Problem solving adalah model mengajar yang digunakan oleh guru dalam proses pembelajaran yang meliputi
Namun hasil penelitian berbeda yang dilakukan oleh Adi dan Ekaristi (2009) mengindikasikan bahwa banyak daerah otonom yang tidak memaksimalkan Pendapatan Asli Daerah dengan
Rumah Sakit Jiwa Daerah Dr. Amino Gondohutomo Semarang.. 69,4% dibandingkan pengetahuan tentang indikator kolaborasi lingkup praktek baik 41,2%, sedangkan praktek kolaborasi perawat
<edung harus mempunyai perlengkapan drainase untuk menyalurkan air hujan dari atap dan halaman atau pekarangan dengan pengerasan di dalam persil ke saluran air hujan kota
Mekanisme hambatan lainnya adalah produksi kitinase, pelekatan pada dinding sel cendawan, aktivitas peroksidase, induksi ketahanan (El Gouth et al., 2003), dan
buku panduan, brosur, leaflet), dan melalui forum sosialisasi langsung (pertemuan, rapat) serta media elektronik lainnya seperti radio. Visi dan Misi Program Pasca Sarjana Magister
Proses ini mulai dari setelah obat yang telah diberi etiket oleh petugas diletakkan kembali dalam kotak obat di meja pengerjaan obat, kemudian petugas penyerahan obat membawanya
[r]
