BANTISTA ADIES KUNCAHYO
DEPARTEMEN KONSERVASI SUMBERDAYA HUTAN DAN EKOWISATA FAKULTAS KEHUTANAN
INSTITUT PERTANIAN BOGOR BOGOR
2015
SEBARAN SPASIAL DAN TIPE METAPOPULASI MACAN
DAHAN (
Neofelis diardi
Cuvier, 1823) DI TAMAN NASIONAL
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa skripsi berjudul Sebaran Spasial dan Tipe Metapopulasi Macan Dahan (Neofelis diardi Cuvier, 1823) di Taman Nasional Sebangau, Provinsi Kalimantan Tengah adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir skripsi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
ABSTRAK
BANTISTA ADIES KUNCAHYO. Sebaran Spasial dan Tipe Metapopulasi Macan Dahan (Neofelis diardi Cuvier, 1823) di Taman Nasional Sebangau, Provinsi Kalimantan Tengah. Dibimbing oleh HADI S. ALIKODRA dan HENDRA GUNAWAN.
Macan dahan (Neofelis diardi Cuvier, 1823) merupakan top-predator di hutan Taman Nasional Sebangau (TNS) yang keberadaannya semakin terancam akibat kehilangan dan fragmentasi habitat sehingga berpotensi menjadikan populasi N. diardi
terbagi ke dalam beberapa sub populasi atau yang dikenal dengan metapopulasi. Penelitian ini bertujuan untuk memperoleh informasi mengenai sebaran spasial N. diardi
di TNS dan memprediksi risiko kepunahan lokal melalui identifikasi tipe metapopulasi. Pengumpulan data dilakukan pada bulan Juni – November 2014 melalui metode studi literatur, wawancara, pengunduhan, orientasi lapang, serta kombinasi pengamatan langsung dan tidak langsung, seperti track-set, grid, camera trap, dan jejak (footprints, feses, claw, scratch, scrape, dan vokalisasi). Penelitian ini menunjukkan bahwa sebaran
N. diardi dipengaruhi oleh jarak dari sumber air (P = 0.002) dan potensi ganguan (P = 0.017), kerapatan tajuk hutan (canopy cover) (P = 0.04), kedalaman gambut (P = 0.002), tipe habitat di kawasan TNS (P = 0.011), serta keberadaan dan kelimpahan satwa mangsanya, namun faktor ketinggian tempat tidak memengaruh N. diardi (P = 0.064). Pola sebaran spasial N. diardi adalah mengelompok (x2hitung = 39.67, x
2
0.025 = 26.12) dengan tipe metapopulasi yang berbentuk patchy population akibat fragmentasi dari keberadaan jaringan kanal sehingga berisiko kecil terhadap kepunahan lokal, namun tipe metapopulasi dapat berubah mengikuti perubahan konektifitas habitat dan keberadaan ancaman baik terhadap N. diardi maupun habitatnya.
Kata kunci: Neofelis diardi, sebaran, metapopulasi, taman nasional sebangau
ABSTRACT
BANTISTA ADIES KUNCAHYO. Distribution Spatial and Type of Metapopulation The Sunda Clouded Leopard (Neofelis diardi Cuvier, 1823) in Sebangau National Park, Central Kalimantan Province. Supervised by HADI S. ALIKODRA and HENDRA GUNAWAN.
Sunda clouded leopard (Neofelis diardi Cuvier, 1823) is top predator in Sebangau National Park (SNP) forest which their existence are more threats caused by loss and fragmentation habitat in order that is potential to make N. diardi devided in to be several sub population or known as a metapopulation. This study aimed to obtain information about the spatial distribution of sunda clouded leopard in SNP and predict the risk of local extinction by identifying the type of metapopulation. Data collection was conducted from June until November 2014 by using study literature method, interviews, download, field orientation and combination of direct and indirect observation methods such as track set, grid, camera trap, and sign of animal (footprints, feces, claw,scratch, scrape, and voice). This study showed that distribution of N. diardi influenced by the distance from water sources (P = 0.002) and potential disturbance (P = 0.017), forest canopy cover (P = 0.04), peat depth (P = 0.002), and the type of habitat in SNP (P = 0.011) also the presence and abundance of preys, but only the altitude factor didn‟t affect toward distribution of N. diardi (P = 0.064). Spatial distribution patterns of N. diardi in SNP was clumped (x2hitung = 39.67, x20.025 = 26.12) with type of metapopulation was patchy population due to fragmentation from the existance of canal network so it had a low risk of local extinction, but the type of metapopulation could be changed by following the changes of habitat connectivity and the existence of threat to the N. diardi and their habitats.
Skripsi
sebagai salah satu syarat untuk memperoleh gelar Sarjana Kehutanan
pada
Departemen Konservasi Sumberdaya Hutan dan Ekowisata
SEBARAN SPASIAL DAN TIPE METAPOPULASI MACAN
DAHAN (
Neofelis diardi
Cuvier, 1823) DI TAMAN NASIONAL
SEBANGAU, PROVINSI KALIMANTAN TENGAH
DEPARTEMEN KONSERVASI SUMBERDAYA HUTAN DAN EKOWISATA FAKULTAS KEHUTANAN
INSTITUT PERTANIAN BOGOR BOGOR
2015
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah subhanahu wata‟ala atas
segala karunia-Nya sehingga karya ilmiah ini berhasil diselesaikan. Tema dalam penelitian yang dilaksanakan sejak bulan Juni 2014 ini adalah spesies macan dahan (Neofelis diardi) dengan judul Sebaran Spasial dan Tipe Metapopulasi Macan Dahan (Neofelis diardi Cuvier, 1823) di Taman Nasional Sebangau, Provinsi Kalimantan Tengah.
Terima kasih penulis ucapkan kepada Prof Dr Ir Hadi S. Alikodra, MS dan Dr Ir Hendra Gunawan, MSi selaku pembimbing yang selalu memberikan arahan, bimbingan, motivasi, ilmu, nasehat, perhatian, dan panutan selama penyelesaian skripsi ini serta Dr Ir Arzyana Sunkar, MSc selaku moderator yang sudah membantu selama seminar. Begitu juga dengan Prof Dr Ir Yanto Santosa, M dan Dr Erianto Indra Putra S.Hut, MSi yang telah memberikan saran, kritik, ilmu dan pengetahuan selama menguji di sidang komprehensif. Selain itu, penghargaan dan terima kasih juga penulis sampaikan kepada Ibu Rosenda Chandra Kasih, Bapak Okta Simon dan Adventus Panda beserta staff WWF-Kalimantan Tengah yang telah banyak membantu dalam berdiskusi, saran, akomodasi dan berbagai infomasi yang dibutuhkan selama kegiatan penelitian. Di samping itu, penulis ucapkan terima kasih dan penghargaan kepada Bapak Ir Adib Gunawan selaku Kepala Balai Taman Nasional Sebangau, Pak Suyoko, Pak Tatang, Mba Tinu, serta staff Balai TN Sebangau yang telah membantu selama penelitian. Penulis juga ucapkan terima kasih kepada Pak Pri, Pak Wanto, Pak Taufik (Opik), keluarga besar Camp Punggualas (Pak Dharma. Pak Pradino dan Bang Ajim), keluarga besar Camp Teluk Beruang (Pak Agus dan Pak Yani), Pak Nur, dan Bang Labay atas segala suka, duka serta pengalaman dan pelajaran yang sudah dibagikan selama di lapangan. Ungkapan terima kasih juga disampaikan kepada
ayah, ibu, kedua adik, serta seluruh keluarga atas segala do‟a dan kasih sayangnya.
Penulis juga sampaikan terima kasih untuk seluruh staff terutama kepada Mamang, Bibi, dan Babeh DKSHE atas segala bantuan, dukungan serta do‟a kepada penulis selama melakukan studi dan menyelesaikan skripsi ini. Terima kasih juga kepada seluruh teman-teman dan keluarga besar ANGGREK HITAM 46, serta Priyono Eka Pratiekno, teman-teman PKLP TN Merbabu, keluarga besar Krakatau dan Rakata (Mas eko, Mas Kuspri), Faiz, Bang Amrul dan Rumah Hijau atas segala
do‟a, dukungan, semangat, dan bantuan hingga selesai penyusunan skripsi ini. Semoga karya ilmiah ini bermanfaat.
Bogor, Agustus 2015
DAFTAR TABEL vii
DAFTAR GAMBAR vii
DAFTAR LAMPIRAN vii
PENDAHULUAN 1
Latar Belakang 1
Tujuan Penelitian 2
Manfaat Penelitian 2
METODE 3
Lokasi dan Waktu 3
Alat dan Bahan 3
Metode Pengumpulan Data 5
Analisis Data 7
HASIL DAN PEMBAHASAN 11
Kondisi Umum Lokasi Penelitian 11
Sebaran Neofelis diardi 11
Metapopulasi 39
Implikasi Pengelolaan 45
SIMPULAN DAN SARAN 46
Simpulan 46
Saran 46
DAFTAR PUSTAKA 47
DAFTAR TABEL
1 Rincian waktu penelitian 3
2 Kriteria nilai indeks Shannon-Wiener 8
3 Kriteria nilai kelimpahan 9
4 Komposisi sebaran Neofelis diardi terhadap kelas elevasi kawasan 18 5 Komposisi sebaran Neofelis diardi terhadap kelas “jarak dari sumber air”
20 6 Komposisi sebaran Neofelis diardi di masing-masing kelas “jarak dari
potensi gangguan” 23
7 Komposisi sebaran Neofelis diardi terhadap kelas kerapatan tajuk 24 8 Komposisi sebaran Neofelis diardi pada kelas kedalaman gambut 28 9 Komposisi sebaran Neofelis diardi terhadap kelas sub tipe habitat 29 10 Nilai berbagai indeks potensi satwa mangsa Neofelis diardi 33 11 Klasifikasi potensi satwa mangsa Neofelis diardi 34 12 Komposisi beberapa nilai indeks pada setiap lokasi camera trap 37
DAFTAR GAMBAR
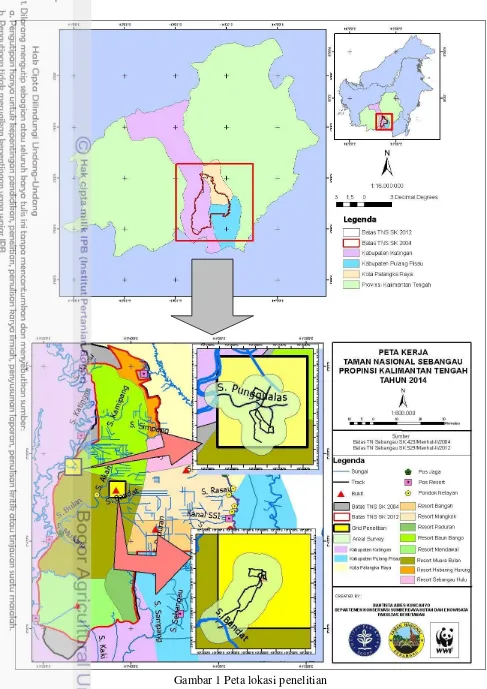
1 Peta lokasi penelitian 4
2 Pemasangan camera trap 6
3 Sketsa metode petak tunggal 7
4 Proses pengolahan peta altitude 7
5 Proses pengolahan peta "jarak dari sungai/sumber air" 8
6 Proses pengolahan peta tutupan lahan 10
7 Bentuk perjumpaan Neofelis diardi 12
8 Nilai RAI Neofelis diardi di lima stasiun camera trap 12
9 Bentuk aktivitas Neofelis diardi 13
10 Aktivitas penandaan teritori Neofelis diardi di berbagai tingkat
pertumbuhan(a) semai (b) tiang (c) pohon 14
11 Peta sebaran Neofelis diardi berdasarkan kelas ketinggian kawasan di area (a) Punggualas, (b) SSI, dan (c) Bukit Bulan 17 12 Sebaran Neofelis diarditerhadap kelas “jarak dari sumber air” di area (a)
SSI, (b) Punggualas, dan (c) Bukit Bulan 19
13 Peta sebaran Neofelis diardi terhadap potensi gangguan : (a.1) basecamp Punggualas, (a.2) basecamp SSI, dan (b) pondok Bukit Bulan 22 14 Peta sebaran Neofelis diardi terhadap kelas kerapatan tajuk hutan di area
(a) Bukit Bulan, (b) SSI, dan (c) Punggualas 25
15 Peta sebaran Neofelis diardi terhadap kelas kedalaman gambut di area
(a) Bukit Bulan, (b) SSI, dan (c) Punggualas 27
16 Peta sebaran Neofelis diardi terhadap kelas sub tipe habitat di area (a)
Bukit Bulan, (b) SSI, dan (c) Punggualas 30
17 Diagram kelas diameter tegakan tumbuhan di plot camera trap Neofelis
diardi 31
18 Diagram kelas tinggi total tegakan tumbuhan di plot camera trap
Neofelis diardi 32 20 Sarang burung di lubang pohon yang dirusak beruang madu 33 21 Sub populasi Neofelis diardi : (a) Bukit Bulan (b) Punggualas (c) SSI
(dokumentasi gambar c : Staff Balai TNS) 40
22 Kondisi sungai dan kanal di kawasan Taman Nasional Sebangau 41 23 Peta Metapopulasi Neofelis diardi di Taman Nasional Sebangau tipe
patchy population 42
24 Kebakaran hutan di kawasan Taman Nasional Sebangau (dokumentasi:
WWF-Kalteng) 43
25 Aktivitas perburuan (a) “kalong” dan (b) burung 43
26 Aktivitas illegal logging (dokumentasi: WWF-Kalteng) 44
DAFTAR LAMPIRAN
1 Identifikasi individu Neofelis diardi 52
2 Indikasi keberadaan (presence-absence) Neofelis diardi 54
3 Perhitungan uji korelasi chi-square 55
PENDAHULUAN
Latar Belakang
Macan dahan atau clouded leopard pernah dianggap sebagai satu spesies yaitu Neofelis nebulosa dengan beberapa sub spesies yang terdiri dari Neofelis nebulosa brachyurus (di Taiwan), Neofelis nebulosa diardi (di Sumatra dan Kalimantan), Neofelis nebulosa macrosceloides (di India, Nepal, Sikkim, dan Myanmar), serta Neofelis nebulosa nebulosa (di bagian Asia Tenggara, China, dan Hainan) (Kitchener et al. 2006). Berdasarkan analisis genetik pada tahun 2007 disimpulkan bahwa macan dahan terdiri dari dua spesies yaitu Neofelis nebulosa (tersebar di Mainland Asia beserta Taiwan) dan Neofelis diardi (tersebar di Kalimantan dan Sumatra) (ENS 2007 dan ScienceDaily 2007).
Macan dahan (Neofelis diardi, Cuvier 1823) merupakan satwa predator terbesar dan endemik di Pulau Kalimantan. Spesies tersebut berperan sebagai top-predator dalam rantai makanan di seluruh kawasan hutan Kalimantan sehingga keberadaannya sangat penting untuk menjaga keseimbangan ekosistem hutan. Selain itu, keberadaan N. diardi diduga berasosiasi dengan keberadaan hutan primer karena spesies tersebut bersifat arboreal sehingga membutuhkan strata tajuk yang lengkap untuk mendukung seluruh aktivitas hariannya. Menurut Indriyanto (2006) bahwa keseimbangan ekosistem hutan ditunjukkan oleh keseimbangan sistem komunitasnya yang dipengaruhi oleh stratifikasi tajuk dan canopy cover.
Ikan merupakan salah satu sumber pakan bagi N. diardi. Keberadaan ikan di suatu perairan dapat memberikan indikasi keseimbangan ekosistem perairan. Menurut Chahaya (2003), ikan yang hidup di suatu perairan dapat menjadi indikator biologis pencemaran air karena ikan mempunyai kemampuan dalam merespon keberadaan bahan pencemar. Oleh karena itu, keberadaan N. diardi diduga juga berasosiasi dengan ekosistem perairan di Kalimantan.
Pemerintah Indonesia telah memberikan perhatian terhadap kelestarian N. diardi dengan menetapkannya sebagai satwa dilindungi berdasarkan Peraturan Pemerintah nomor 7 tahun 1999 (Pemerintah Republik Indonesia 1999). IUCN (International Union for Conservation of Nature and Natural Resources) juga telah memasukkannya ke dalam red list IUCN tahun 2008 dengan kategori Vulnerable (rentan), sedangkan CITES (Convention International in Trade of Endangered Species of Flora and Fauna) pada tahun 2012 menetapkannya dalam Appendiks I yang berarti tidak boleh diperdagangkan (Hearn et al. 2008 dan CITES 2012). Ternyata, perhatian-perhatian tersebut belum cukup menjamin kelestarian satwa langka tersebut, bahkan ancaman terhadap spesies tersebut semakin meningkat, seperti perburuan N. diardi maupun satwa-satwa mangsanya, perdagangan ilegal, degradasi dan fragmentasi hutan beserta konversi hutan. Merduani (2013) melaporkan perdagangan macan dahan di daerah Sumatra Selatan dengan harga Rp 6.5 juta hingga Rp 10 juta per ekor.
penutupan hutan Pulau Kalimantan semakin berkurang sejak tahun 1990 hingga 2009 dengan kehilangan tutupan hutan mencapai 13.11 juta ha. Dengan perkataan lain laju kehilangan hutan di Kalimantan rata-rata 720 ribu ha per tahun, Provinsi Kalimantan Tengah merupakan wilayah yang paling banyak kehilangan hutan sejak tahun 2004 hingga 2008 (Jimmy 2011).
Taman Nasional Sebangau (TNS) merupakan kawasan konservasi yang sangat penting di Kalimantan terutama Provinsi Kalimantan Tengah karena merupakan hutan rawa gambut yang sangat tebal dengan kedalaman mencapai 3 – 26 m (Page et al. 1999). Selain itu, TNS juga sangat penting sebagai penyimpan karbon (carbon storage) dan pengatur tata air (hidrologi) serta habitat dari berbagai jenis bagi flora dan fauna endemik, terutama N. diardi yang berhasil didokumentasikan Cheyne et al. (2013) menggunakan camera trap. Kehilangan dan fragmentasi habitat merupakan ancaman utama bagi kelestarian spesies tersebut di TNS serta berpotensi menjadikan populasi N. diardi terbagi ke dalam beberapa sub populasi (populasi lokal) atau yang dikenal dengan metapopulasi (Akçakaya 2007 dan Gunawan 2010). Hal ini disebabkan sebelum menjadi taman nasional, kawasan tersebut merupakan hutan produksi yang dieksploitasi dan mengalami illegal logging hingga tahun 2006 (WWF-Indonesia Sebangau Project 2012).
Kelestarian N. diardi sangat tergantung pada upaya konservasi yang diterapkan, baik secara insitu maupun eksitu. Upaya-upaya tersebut perlu didukung oleh pengetahuan tentang berbagai informasi bio-ekologis spesies tersebut sehingga diperlukan berbagai penelitan untuk menggali informasi yang dibutuhkan sebagai bahan pertimbangan dalam melakukan berbagai upaya konservasi. Salah satu aspek penting yang perlu diketahui adalah sebaran spasial dan tipe metapopulasinya. Informasi sebaran individu macan dahan berguna untuk menetapkan wilayah-wilayah yang perlu dilindungi, sedangkan pengetahuan tentang metapopulasi dapat memberikan gambaran risiko kepunahan lokal di masa mendatang.
Tujuan Penelitian
Penelitian ini bertujuan untuk memperoleh informasi sebaran spasial dan mengidentifikasi tipe metapopulasi macan dahan (Neofelis diardi) di Taman Nasional Sebangau.
Manfaat Penelitian
METODE
Lokasi dan Waktu
Penelitian dilakukan di Taman Nasional Sebangau (TNS) yang secara khusus di wilayah Danau Punggualas dan Bukit Bulan. Lokasi penelitian terletak di wilayah kerja SPTN III, Resort Baun Bango dan Muara Bulan. Secara umum kawasan TNS berada pada tiga wilayah Kabupaten/Kota, yaitu Pulang Pisau, Katingan, dan Kotamadya Palangka Raya. Kawasan Hutan Bukit Bulan dan Danau Punggualas yang merupakan lokasi penelitian terletak di Kabupaten Katingan yaitu Kecamatan Kamipang dan Mendawai. Secara geografis wilayah TNS dipisahkan oleh dua Daerah Aliran Sungai (DAS) yaitu DAS Katingan dan Sebangau. TNS terletak pada 1O54‟ – 3O08‟ LS dan 113O20‟ – 114O03‟ BT dengan luas ±568 700 ha, sedangkan wilayah penelitian mencakup areal seluas ±44.37 km2 (4 436.41 ha) (Gambar 1).
Penelitian ini dilakukan pada Bulan Juni – September 2014 yang terdiri dari lima periode dengan total waktu selama 63 hari dan rincian yang disajikan pada Tabel 1.
Tabel 1 Rincian waktu penelitian Periode
Penelitian Periode Waktu
Jumlah
Hari Kegiatan
I 13 – 20 Juni 8 Observasi Pendahuluan dan Wawancara II 6 Juli – 13 Agustus 39 Pemasangan Camera Trap dan Observasi III 19 – 28 Agustus 10 Pemantauan Camera Trap dan Observasi IV 23 – 28 September 6 Pemantauan Camera Trap dan Analisis
Titik Lokasi Presence-AbsenceNeofelis diardi berdasarakan Camera Trap
Alat dan Bahan
Metode Pengumpulan Data
Studi pendahuluan
Tahap ini dilakukan dengan studi literatur, orientasi lapang, pengunduhan, dan wawancara terhadap petugas lapangan dan staf Balai Taman Nasional Sebangau (TNS) serta WWF-Kalteng, masyarakat sekitar kawasan, buku, laporan, jurnal, dan media internet. Data yang dikumpulkan meliputi:
1. Studi literatur dan wawancara :
a. Kondisi umum lokasi penelitian, keberadaan (presence-absence) dan sebaran N. diardi beserta satwa mangsa, sumberdaya air, dan potensi gangguan
b. Data spasial seperti peta kedalaman gambut, kerapatan tajuk hutan (canopy cover), dan tipe habitat serta data spasial sungai dan kanal di Taman Nasional Sebangau
2. Pengunduhan (download) :
Data spasial seperti peta SRTM (Shuttle Radar Topography Mission), citra landsat TM (Thematic Mapper) 5 tahun 2010 path 118/row 61 & 62, dan peta Rupa Bumi Provinsi Kalimantan Tengah untuk mengolah peta tematik berupa penutupan lahan, ketinggian (elevasi) kawasan TNS, dan “jarak dari
sumber air”.
3. Orientasi lapang :
Lokasi penempatan grid, track-set dan pemasangan camera trap
Secara terperinci data pendukung mengenai N. diardi dan satwa mangsanya di dalam wawancara, yaitu :
1. Lokasi keberadaan dan penyebaran N. diardi
2. Kelimpahan secara deskriptif dari keberadaan N. diardi
3. Lokasi yang pernah dihuni macan dahan (catatan : tetapi sekarang sudah tidak dihuni)
4. Perburuan dan aktivitas yang memberikan tekanan terhadap macan dahan 5. Perburuan potensi satwa mangsa
6. Kelimpahan secara deskriptif dan bermacam spesies yang menjadi potensi mangsa
Observasi lapang
Identifikasi keberadaan (presence-absence) macan dahan
(padang rumput dan lokasi dengan vegetasi yang sedang berbuah), tempat kubangan, sungai/kanal keringyang ditinggal selama sebulan (Gambar 2).
Selain itu, setiap perjumpaan jejak dilakukan pengukuran untuk identifikasi individu. Setiap perjumpaan terhadap tanda keberadaan ditandai menggunakan GPS (Global Positioning System) dan dicatat titik koordinatnya.
Gambar 2 Pemasangan camera trap
Identifikasi daya dukung habitat dan potensi gangguan terhadap macan dahan Parameter yang dijadikan daya dukung habitat adalah potensi satwa mangsa, keadaan suhu udara, curah hujan, dan air. Secara umum seluruh parameter tersebut disurvey secara serentak bersamaan dengan pengamatan terhadap N. diardi di dalam jalur (track), meskipun sebelumnya juga telah diidentifikasi pada tahap studi pendahuluan.
Survei terhadap identifikasi potensi satwa mangsa juga menggunakan metode camera trap. Informasi yang dicatat adalah jenis dan jumlah satwa yang berpotensi sebagai mangsa serta lokasi keberadaan satwa tersebut. Selain itu, satwa mangsa diklasifikasikan ukuran berat tubuh berdasarkan penelitian Henschel (2008).
Identifikasi sumberdaya air dilakukan dengan mencatat jumlah dan lokasi keberadaan sumberdaya air beserta keterangan lain yang dapat mendukung informasi mengenai potensi sumberdaya air sebagai daya dukung habitat. Identifikasi bentuk tekanan terhadap habitat dilakukan dengan meninjau beberapa aspek seperti jenis atau bentuk tekanan dan lokasinya. Lokasi dari seluruh parameter yang dijumpai ditandai menggunakan GPS dan dicatat titik koordinatnya.
Kondisi habitat camera trap macan dahan
(gambar 3). Komponen pendukung habitat yang lain menggunakan metode penjelajahan di dalam dan sekitar petak tunggal.
Gambar 3 Sketsa metode petak tunggal
Analisis Data
Sebaran spasial
Hasil titik sebaran macan dahan yang telah di-input ke dalam GPS (Global Positioning System) dalam bentuk koordinat kemudian di-download ke dalam komputer. Selanjutnya, data diproses menggunakan software Arc GIS 9.3 dan ERDAS IMAGINE 9.1 melalui teknik tumpang susun (overlay) antara titik sebaran Neofelis diardi dengan beberapa jenis peta tematik kawasan Taman Nasional Sebangau (TNS), seperti tipe habitat, kerapatan tajuk permukaaan (canopy cover),
tingkat kedalaman gambut, ketinggian tempat (elevasi), “jarak dari sumber air” dan “jarak dari potensi tekanan (gangguan) terhadap habitat”.
Peta tematik ketinggian (elevasi) kawasan TNS memerlukan beberapa proses pengolahan yang ditunjukkan oleh gambar 4. Peta „jarak dari sumber air” diperoleh melalui pengolahan terhadap peta rupa bumi Provinsi Kalimantan Tengah beserta data spasial (fiture) terbaru yang berasal dari Balai TNS dengan melakukan proses buffer. Proses tersebut serupa dengan yang digunakan untuk
“jarak dari potensi gangguan habitat” yang disajikan dalam Gambar 5.
Gambar 4 Proses pengolahan peta altitude d
Gambar 5 Proses pengolahan peta "jarak dari sungai/sumber air" Potensi satwa mangsa
Indeks kekayaan jenis dihitung menggunakan metode Margalef dengan rumus sebagai berikut:
� = S−1
ln(N)
Simbol (S) adalah jumlah seluruh jenis, sedangkan (N) adalah jumlah individu seluruh jenis dan (Dmg) adalah nilai indeks Margalef (Ludwig and Reynold 1998).
Keanekaragaman jenis dihitung dengan indeks keanekaragaman
Shannon-Wiener (H‟) menggunakan rumus sebagai berikut:
�′ = − Pi ln Pi ;�� = �
�
Simbol (ni) adalah jumlah individu dalam setiap jenis, sedangkan (N) adalah total jumlah individu seluruh jenis (Ludwig and Reynold 1998). Selain itu, nilai indeks ini diklasifikasikan ke dalam beberapa kriteria (Tabel 2).
Tabel 2 Kriteria nilai indeks Shannon-Wiener Nilai Indeks
Shannon-Wiener Kriteria
>3 Keanekaragaman tinggi, penyebaran individu dalam setiap spesies tinggi, dan kestabilan komunitas tinggi
1 – 3 Keanekaragaman sedang, penyebaran individu dalam setiap spesies sedang, dan kestabilan komunitas sedang <1 Keanekaragaman rendah, penyebaran individu dalam
setiap spesies rendah, dan kestabilan komunitas rendah Sumber : (Fachrul 2008)
Indeks kelimpahan spesies satwa mangsa dapat menggunakan indeks Simpson dengan persamaan rumus (Magurran 2004) :
� = (��)2, �� = �
� 2
selanjutnya dikelompokkan sesuai kriteria (Liley and Robertson 2000 diacu dalam Jamaksari 2011) pada Tabel 3 :
Tabel 3 Kriteria nilai kelimpahan
Kategori Kelimpahan Nilai Kelimpahan Skala Ukuran
<0.1 1 Jarang
0.1 – 2.0 2 Tidak Umum
2.0 – 10.0 3 Sering
10.1 – 40.0 4 Umum
>40.0 5 Melimpah
Indeks kelimpahan relatif (RAI)
Indeks kelimpahan relaltif digunakan untuk menentukan ukuran keberhasilan camera trap maupun tingkat perjumpaan pada suatu areal/titik lokasi pemasangan yang dihutung menggunakan rumus sebagai berikut (Acrenaz et al. 2012):
��� = �
� ×
Simbol ( )adalah jumlah foto yang diperoleh (number of picture), sedangkan (�) adalah jumlah hari aktif pada camera trap (number of trap night). Konstanta (100) digunakan untuk menyetarakan satuan usaha hari aktif pada seluruh periode pemasangan (Lynam et al. 2000 diacu dalam Fata 2011).
Kondisi vegetasi
Analisis vegetasi dilakukan dengan mengelompokkan beberapa karakteristik vegetasi seperti tinggi total (Tt), Tinggi bebas cabang (Tbc), dan diameter (Dbh) tegakan sesuai dengan penelitian Purwadi (2010).
Pola sebaran dan hubungan Neofelis diardi dengan daya dukung habitat Pola sebaran yang dibentuk macan dahan kalimantan di kawasan TNS dianalisis menggunakan Indeks Dispersal (ID) dengan rumusan sebagai berikut:
Keterangan :
Selanjutnya, nilai ID akan dilakukan uji Chi-Square untuk memperoleh bentuk pola sebaran spasial dengan persamaan
Penggunaan Uji Chi-Square untuk N<30 dan N adalah jumlah kontak dengan satwa. Bentuk pola sebaran spasial disesuikan terhadap perbandingan antara nilai Χ2hitung dan Χ2tabel dengan kriteria seperti :
1. Jika Χ2hitung > Χ20.025 , maka pola sebaran berbentuk kelompok
Χ2= ID (N-1) ID : Indeks Dispersal
S2 : Keragaman Contoh X : rataan contoh
ID = S 2
2. Jika Χ20.025 ≤ Χ2hitung ≤Χ20.975 , maka pola sebaran berbentuk acak 3. Jika Χ2hitung < Χ20.975 , maka pola sebaran berbentuk seragam/sistematik
Analisis mengenai hubungan antara keberadaan Neofelis diardi dengan berbagai daya dukung habitat yang berbentuk data spasial seperti peta-peta tematik (hasil overlay) dilakukan uji korelasi menggunakan Chi-Square dengan bantuan software MINITAB 14. Apabila Χ2hitung < Χ2tabel atau (P < 0.05), maka terima H1 dengan hipotesis yang dibangun adalah
H0 : tidak ada hubungan yang signifikan antara keberadaan Neofelis diardi dengan faktor (ketinggian kawasan, kerapatan tajuk hutan, tingkat
kedalaman gambut, tipe habitat, “jarak dari sumber air‟, dan “jarak dari potensi gangguan”)
H1 : ada hubungan yang signifikan antara keberadaan Neofelis diardi dengan faktor (ketinggian kawasan, kerapatan tajuk hutan, tingkat
kedalaman gambut, tipe habitat, “jarak dari sumber air‟, dan “jarak dari potensi gangguan”)
Metapopulasi
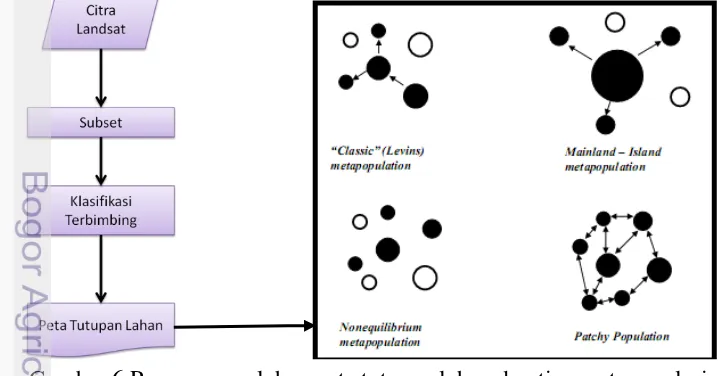
Analisis mengenai metapopulasi dilakukan untuk mengidentifikasi tipe-tipe metapopulasi. Tipe metapopulasi akan dimodelkan sesuai dengan kondisi keberadaan N. diardi yang di-overlay (tumpang susun) dengan peta tutupan lahan TNS. Peta penutupan lahan diperoleh melalui hasil pengolahan terhadap Citra Landsat seperti yang ditunjukkan oleh Gambar 6.
Hasil interpretasi tersebut akan dipelajari terkait dengan berbagai peluang konektifitas N. diardi yang berada di tiap patch tersebut melalui permodelan yang digambarkan oleh Hanski dan Simberloff (1997) beserta Harrison dan Taylor (1997) diacu dalam Gunawan (2010) seperti classic metapopulation, mainland-island metapopulation, nonequilibrium metapopulation, dan patchy population.
HASIL DAN PEMBAHASAN
Kondisi Umum Lokasi Penelitian
Kawasan Taman Nasional Sebangau (TNS) sebelumnya merupakan kawasan Hutan Produksi (HP) dan Hutan Produksi yang dapat dikonversi (HPK). Status TNS ditunjuk berdasarkan Surat Keputusan Menteri Kehutanan Nomor SK.423/Menhut-II/2004 tanggal 19 Oktober 2004 dengan luas ±568 700 ha yang merupakan hasil penggabungan dari ±510 250 ha HP dan ±58 450 ha HPK. Ketika masih berstatus hutan produksi kawasan TNS pernah mengalami kebakaran besar sebanyak empat kali yaitu pada tahun 1992, 1994, 1997, dan 2002 yang dipicu oleh kondisi gambut yang rusak akibat illegal logging dan pemanfaatan lahan yang menyebabkan adanya parit atau kanal buatan serta akibat terjadinya pengeringan (BTNS 2013). Kawasan TNS menurut sistem klasifikasi Koppen memiliki tipe iklim Af dengan delapan bulan basah dan tidak mempunyai bulan kering yang nyata (BTNS 2007). Jumlah curah hujan pada bulan terkering lebih dari 60 mm dengan variasi pada kisaran 2 000 – 4 000 mm sehingga dikategorikan bercurah hujan tinggi (BTNS 2013). Kawasan TNS tersusun atas formasi endapan alluvium/endapan permukaan (Qa) dan batuan sedimen tanah muka (Tq). Endapan permukaan yang terbentuk berasal dari endapan alluvium sungai, endapan gambut dan/atau bahan organik. Adapun bahan organik yang menyusun kawasan TNS terdiri dari endapan bahan organik, liat, dan pasir. Bahan organik tersebut terbagi ke dalam tiga kelompok berdasarkan tingkat kematangan, yaitu fibric, hemic, dan safric. Perbedaan tingkat kematangan dan ketebalan tanah memberikan pengaruh terhadap fisiografi kawasan TNS yang akan tampak seperti suatu dataran rawa (floodplain), tanggul sungai (levee), dan satuan lahan kubah gambut baik gambut ombrogen ataupun oligotrofik.
Sebaran Neofelis diardi
Wilting (2007) pernah berjumpa langsung dengan N. diardi di Tabin Wildlife Reserve, Sabah Malaysia.
Gambar 7 Bentuk perjumpaan Neofelis diardi
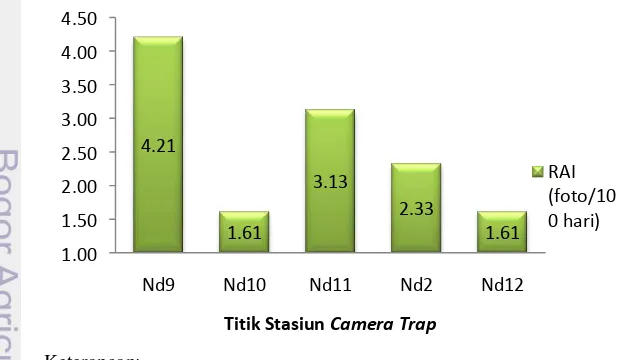
Kesulitan untuk menemukan N. diardi juga ditunjukkan oleh nilai RAI (Relatife Abundance Index)/tingkat perjumpaan dari pemasangan camera trap di 14 titik stasiun dengan jumlah hari aktif kamera 30 – 62 hari. Tingkat perjumpaan N. diardi bernilai rataan sebesar 1.37 foto/100 hari dengan rentang nilai 1.61 – 4.21 foto/100 hari. Perbedaan nilai RAI dipengaruhi oleh beberapa faktor eksternal dan internal dalam penelitian. Beberapa faktor internal yang berpengaruh adalah penggunaan jumlah dan periode pemasangan camera trap, penemuan lokasi strategis pemasangan kamera (penemuan tanda baru ataupun lama), dan perilaku maupun kemampuan satwa (ukuran home range dan daily movement), sedangkan faktor eksternalnya meliputi kondisi habitat satwa (waktu musiman, substrat, dan daya dukung habitat) serta intensitas dan kualias keberadaan ancaman bagi satwaliar. RAI N. diardi di kelima stasiun camera trap dapat dilihat pada Gambar 8.
Gambar 8 Nilai RAI Neofelis diardi di lima stasiun camera trap
41.67%
Tingkat perjumpaan untuk N. diardi lebih rendah dibandingkan oleh Panthera pardus di Gabon Tengah (Afrika) yang mempunyai nilai RAI dengan rentang 0 – 6.3 foto/100 hari dengan 87 unit camera trap dan periode pemasangan 39 – 62 hari (Henschel 2008). Selain itu, Fata (2011) memperoleh nilai RAI N. diardi di Pulau Sumatera yang lebih tinggi dengan nilai 4.75 dan 7.85 foto/100 hari di dua lokasi kamera, sedangkan rataan RAI Panthera tigris sumatrae yang bernilai 0.93 foto/100 hari dengan rentang 0 – 7.85 foto/100 hari dari penggunaan 28 unit camera trap dan lama periode pemasangan 18 – 59 hari lebih rendah dari RAI N. diardi selama penelitian. Menurut Acrenaz et al. (2012) nilai RAI ini bukan merupakan ukuran kelimpahan spesies dalam suatu populasi, namun lebih cenderung menunjukkan tingkat kemudahan dalam menemukan keberadaan suatu spesies sehingga dapat dikatakan bahwa RAI merupakan indikator ukuran keberhasilan trapping. Oleh karena itu, kelima stasiun lokasi camera trap dapat menjadi pertimbangan dalam memasang camera trap bagi N. diardi di kawasan TNS.
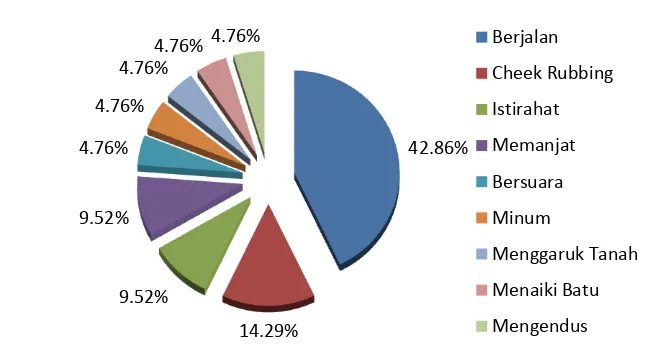
Indikasi keberadaan N. diardi di TNS menunjukkan beberapa aktivitas yang berbeda-beda. Bentuk aktivitas terbanyak yang ditemukan adalah berjalan sebanyak 9 aktivitas (42.86%), sedangkan menggesekkan pipi (cheek rubbed) ditemukan sebanyak 3 aktivitas (14.29%) (Gambar 9). Cheek rubbed adalah salah satu aktivitas perilaku teritorial dan komunikasi antar individu dari N. diardi yang dilakukan pada batang tegakan. Aktivitas tersebut ditemukan di tiga lokasi camera trap yang berbeda dan substrat yang menjadi objek penandaan (marking) terdiri dari tiga tingkat pertumbuhan yang juga berbeda, yaitu semai, tiang (genus Artocarpus) dan pohon (genus Agathis). Gambar 10 menunjukkan bentuk penandaan wilayah N. diardi di tipe habitat Tall Interior Forest (Punggualas) dan Hill Forest (Bukit Bulan).
Gambar 9 Bentuk aktivitas Neofelis diardi
MacDonald (1985) diacu dalam MacDonald and Loveridge (2010) menjelaskan cheek rubbed termasuk bentuk marking Felidae (facial scent glands) terutama dalam aspek komunikasi melalui bau (smells). Selain itu, bagian pipi (cheek) Felidae terdapat whiskers yang berperan sebagai indera peraba sehingga ketika N. diardi melakukan aktivitas ini, maka cairan (fluid) yang berasal dari kantung kelenjar tidak hanya ditinggalkan sebagai tanda scent marking, melainkan
juga menghantarkan syaraf dengan cepat. Oleh karena itu, satwa ini juga mampu mengenali maupun menjelaskan kondisi lingkungan sekitar, bahkan mendeteksi dan berinteraksi dengan mangsa yang potensial. MacDonald and Loveridge (2010) menegaskan kondisi ini dipengaruhi oleh cairan dari kantung kelenjar yang kaya akan suplai syaraf sehingga apapun yang tersentuh dapat cepat terdeteksi, sedangkan kemampuan mendeteksi mangsa berasal dari kombinasi terhadap visuo-tactile sense. Kebanyakan Felidae berburu menggunakan indera penglihatan maupun suara untuk mendeteksi lokasi mangsa, namun beberapa Felidae menggunakan indera penciuman untuk mendeteksi mangsa seperti yang dilakukan oleh ocelots (Leopardus pardalis) (Kitchener 1991 diacu dalam MacDonald and Loveridge 2010).
(a)
(b) (c)
Famili Moraceae mempunyai ciri batang yang bergetah sangat rekat (Dixon 2011). Famili Araucariaceae pada Genus Agathis mempunyai ciri kulit kayu yang mengandung resin melimpah dengan sifat resin yang mudah mengering dan rekat (Heung-Kyu et al. 2011). Sifat kedua getah yang rekat diindikasikan akan memberikan pengaruh bau (scent) yang ditinggalkan oleh N. diardi sehingga scent akan lebih merekat pada substrat (tumbuhan) yang ditandai dan tumbuhan tersebut termasuk sebagai catnip. Menurut MacDonald and Loveridge (2010) Felidae memilih tempat untuk aktivitas marking secara hati-hati. Selain itu, Todd (1962) beserta Palen and Goddard (1966) diacu dalam MacDonald and Loveridge (2010) juga menambahkan terdapat interaksi antara Felidae dengan tumbuhan akibat pengaruh catnip sehingga satwa tertarik untuk merespon dengan beberapa aktivitas seperti menggesekkan bagian tubuh pipi, dagu, dan badan (cheek, chin and body rubbing) atau bahkan berguling (rolling). Perilaku ini serupa dengan yang ditunjukkan oleh kucing domestik.
Informasi mengenai aktivitas teritorial dari macan dahan secara umum sangat terbatas sebagian besar hanya diperoleh dari lembaga konservasi eksitu yang menurut Fletchall (2000) salah satu bentuk perilaku marking N. nebulosa adalah cheek marking yang cenderung memanfaatkan indera penciuman sebagai bentuk komunikasi sehingga terdapat tanda yang ditinggalkan melalui kelenjar sebaceous (sebaceous glands) pada bagian pipi dan kondisi tersebut lebih menonjol pada pejantan dibandingkan oleh betina. Berdasarkan dua kejadian marking (foto b dan c gambar 10) di lokasi penelitian diketahui bahwa N. diardi berjenis kelamin jantan, sedangkan satu kejadian lagi (foto a gambar 10) tidak dapat diketahui karena trap tidak memperoleh penampakan seluruh bagian tubuh satwa. Wilting (2007) juga menemukan aktivitas teritori N. diardi di alam berupa cheek rubbed dan urinisasi.
persentase tumpang tindih antar pejantan N. nebulosa bernilai 31.4% dan 47.5% yang menurut Crawshaw and Quigley (1991) kemungkinan terjadi karena derajat toleransi sosial yang tinggi atau akibat kondisi habitat yang kompleks sehingga memberikan kesulitan dalam mempertahankan teritori. Gambaran mengenai wilayah jelajah mungkin samar di dalam realitas sosial Felidae karena kejadian tumpang tindih berhubungan dengan susunan ruang (spatial organization) yang dibatasi oleh skala waktu, sedangkan susunan ruang tersebut bersifat dinamis (Doncaster and MacDonald 1991 diacu dalam MacDonald and Loveridge 2010). Selain itu, Crawshaw and Quigley (1991) beserta Grassman (1999) menyebutkan kejadian tumpang tindih antar pejantan juga terjadi pada beberapa spesies Felidae seperti jaguar (Panthera onca) dan leopard (Panthera pardus).
Sebaran menurut ketinggian tempat (elevasi)
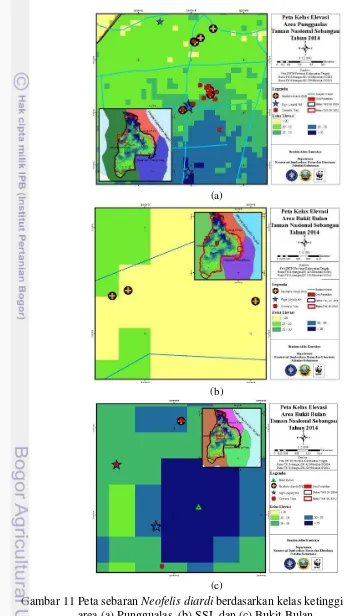
Topografi Taman Nasional Sebangau cenderung datar dan bergelombang, meskipun terdapat wilayah perbukitan dengan puncak tertinggi 190 m dpl. Gandasasmita et al. (2006) menjelaskan penurunan tanah pada ekosistem rawa pasang surut air dapat terjadi secara tidak berangsur-angsur atau mendadak dengan jarak perubahan yang pendek dan kecenderungan pola permukaan yang ditunjukkan oleh ekosistem rawa gambut yaitu semakin meninggi menuju daerah kubah gambut (peat dome). Oleh karena itu, ditemukan wilayah dengan ketinggian tempat di atas permukaan laut yang sama, meskipun di dua tipe habitat yang berbeda seperti hutan gambut di Punggualas dan hutan perbukitan di Bukit Bulan (Gambar 11).
Ketinggian kawasan hutan TNS dibagi menjadi lima kriteria kelas dengan selang 5 m dpl, yaitu sangat rendah, rendah, sedang, tinggi, dan sangat tinggi. Nilai kelas 0 – 20 m dpl dijadikan sebagai kriteria sangat rendah karena menurut Noor (2001) Hutan Sebangau merupakan gambut dataran tinggi dengan ketinggian 10 – 20 m dpl, sedangkan nilai kelas 35 – 190 m dpl sebagai kriteria sangat tinggi karena menurut BTNS (2007) wilayah perbukitan di kawasan TNS berada di ketinggian > 35 m dpl. Hampir di setiap kriteria kelas ditemukan N. diardi, meskipun ditemukan sebaran terbanyak di kriteria kelas rendah dengan delapan perjumpaan (44.45%). Oleh karena itu, keberadaan N. diardi diindikasikan tidak berhubungan dengan faktor ketinggian tempat di atas permukaan laut. Komposisi sebaran N. diardi terhadap setiap kriteria kelas ketinggian kawasan dapat dilihat pada Tabel 4.
Tidak adanya hubungan tersebut disebabkan oleh perbedaan ketinggian yang seragam (tidak jauh berbeda). Lakitan (2002) menjelaskan perubahan ketinggian tempat setiap kenaikan 100 mdpl memengaruhi suhu udara yang menurun 0.6oC. Selain itu, macan dahan dapat hidup di berbagai ketinggian tempat (elevasi) mulai dari habitat dengan ketinggian terendah seperti mangrove, pantai, rawa pantai dan gambut hingga wilayah perbukitan dan pegunungan, bahkan di wilayah dengan ketinggian 3 000 m dpl (Rabinowitz 1988, Chiang 2007, Hearn et al. 2008, Hancock 2012).
(2002) juga menjelaskan bahwa jumlah jenis satwa semakin menurun mengikuti ketinggian tempat. Kondisi inilah yang menyebabkan sebaran N. diardi di kriteria rendah ditemukan dengan frekuensi terbanyak.
(a)
(b)
(c)
Tabel 4 Komposisi sebaran Neofelis diardi terhadap kelas elevasi kawasan terdapat hubungan yang signifikan dari kedua parameter. Kondisi ini sesuai karena N. diardi dapat beradaptasi dan hidup di berbagai ketinggian. Selain itu, perbedaan ketinggian di kawasan hutan TNS yang seragam sehingga bentuk permukaannya cenderung datar. Oleh karena itu, ketinggian kawasan di ekosistem rawa gambut TNS tidak direkomendasikan untuk dipergunakan sebagai parameter dalam monitoring N. diardi.
Sebaran menurut jarak dari sumber air
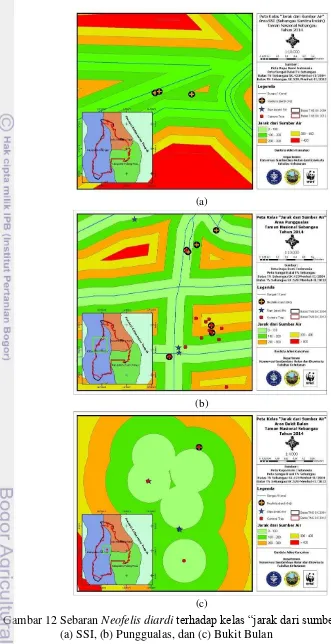
Sumber air di kawasan TNS dipengaruhi oleh dua faktor, yaitu hujan dan/atau pasang surut air sungai. Kondisi ini ditunjukkan oleh lokasi di daerah Sebangau Sanitra Indah (SSI) dan Punggualas sebagai kawasan yang dipengaruhi kedua faktor tersebut karena terletak di dekat sungai dan berupa kawasan bergambut, sedangkan di Bukit Bulan hanya bergantung pada kejadian hujan karena berada di daerah pedalaman. Oleh karena itu, sumber air di kawasan Bukit Bulan berbentuk kubangan-kubangan air, sedangkan air pasang surut dari Sungai Bulan hanya mempengaruhi sumber air di sekitar kawasan bukit yang merupakan kawasan bergambut. Peta sebaran N. diarditerhadap kelas “jarak dari sumber air” menunjukkan perbedaan karakteristik sumber air yang dijadikan indikator pada masing-masing kawasan (Gambar 12).
Kriteria kelas “jarak dari sumber air” dibagi menjadi lima, yaitu sangat
dekat, dekat, sedang, jauh, dan sangat jauh dengan selang jarak 100 m setiap kelas. Perjumpaan terbanyak N. diardi berada di kriteria kelas sangat dekat dengan sebelas perjumpaan (61.11%), sedangkan proporsi sebaran di kelas lainnya adalah 5,56% untuk kedua kelas sedang dan jauh, serta 27.78% di kelas dekat. Oleh karena itu, keberadaan N. diardi diindikasikan berhubungan dengan jarak dari sumber air. Komposisi sebaran N. diardi di masing-masing kelas “jarak dari
(a)
(b)
(c)
Tabel 5 Komposisi sebaran Neofelis diarditerhadap kelas “jarak dari sumber air” along small stream) direkomendasikan sebagai lokasi untuk melakukan observasi terhadap predator tersebut. Selain itu, keberadaan N. diardi juga ditemukan di daerah kanal Laboratorium Alam Hutan Gambut (LAHG) CIMTROP, TNS (Cheyne et al. 2013), bahkan Lynam et al. (2001) diacu dalam Chiang (2007) menyebutkan salah satu lokasi camera trapping yang berhasil menangkap keberadaan N. nebulosa di Thailand adalah dasar sungai yang mengering (dry stream beds). Grassman (2003) juga menemukan lokasi wilayah jelajah N. nebulosa yang berada di daerah aliran sungai utama (major stream). Pemasangan satu camera trap di parit Punggualas yang sudah mulai mengering juga menemukan keberadaan N. diardi.
Air merupakan sumber penghidupan bagi makhluk hidup terutama untuk satwa karena berperan sebagai sumberdaya minumnya. Selain itu, air juga dibutuhkan satwaliar untuk menjaga suhu, kelembapan, dan kebersihan tubuh yang umumnya dilakukan satwa mamalia besar dengan aktivitas berkubang, bahkan air merupakan habitat bagi selurub satwa aquatic seperti ikan yang potensial menjadi mangsa karnivora. Ikan berpotensi sebagai satwa mangsa bagi N. diardi terutama di kawasan Punggualas karena beberapa jenis ikan di daerah tersebut berukuran besar dengan berat tubuh mencapai 2.75 kg (small prey) dan hidup di bagian tepi sungai (Lampiran). Kondisi ini juga didukung oleh kemampuan N. nebulosa yang mampu memegang mangsa (grabbed) seperti halnya yang dilakukan oleh fishing cats (Prionailurus viverrinus) untuk memperoleh ikan di dalam air (Grzimeks 1975). Kebutuhan terhadap sumberdaya air untuk minum diindikasikan oleh penemuan N. diardi di area Sebangau Sanitra Indah (SSI) yang terjadi pada Bulan Mei 2008 ketika curah hujan menurun.
Kondisi vegetasi di sekitar sumber air juga mendukung N. diardi dalam berburu karena umumnya vegetasi rapat oleh rerumputan maupun semak belukar yang lebat. Ismail (2001) menambahkan Rerumputan/sedges (Cyperaceae) dan paku-pakuan (ferns) banyak ditemukan di daerah pinggir sungai, terlebih akan lebih melimpah pada daerah bekas terbakar seperti yang terjadi pada daerah muara Sungai Punggualas. Selain itu, Manshur (2011) juga menyebutkan habitat semak belukar dapat ditemukan di daerah pinggiran sungai. Menurut Gunawan and Alikodra (2013), salah satu strategi predator berada di sekitar sumber air terutama satwa Pantherinae seperti macan tutul adalah untuk menunggu potensi satwa-satwa mangsa yang datang mendekati sumber sehingga ketika sedang beristirahat di dalam semak dan rumput dapat segera menyergap satwa mangsa tersebut.
Hasil uji Chi-square menunjukkan nilai (P = 0.002) atau (P < 0.05) yang menjelaskan ada hubungan antara keberadaan N. diardi dengan faktor jarak dari sumber air seperti sungai dan kubangan air dengan signifikan. Hasil tersebut juga menjelaskan N. diardi cenderung berada di sekitar sumber air sehingga satwa dapat menjadi indikator biologis bagi kondisi sumber air di suatu habitat. Oleh karena itu, faktor jarak dari sumber air dapat dijadikan pertimbangan untuk monitoring N. diardi di kawasan TNS.
Sebaran menurut jarak dari potensi gangguan
Alikodra (2002) menyatakan salah satu komponen habitat dari faktor biotik adalah komunitas biotik pada suatu habitat, sedangkan manusia merupakan bagian dari komunitas biotik tersebut. Salah satu pengaruh negatif dari manusia dikenal sebagai ancaman atau gangguan, baik aktif maupun pasif. Basecamp/stasiun penelitian dan pondok mampu berpotensi sebagai gangguan terhadap satwaliar, bahkan juga dapat bersifat aktif maupun pasif. Perburuan merupakan salah satu bentuk ancaman yang aktif, sedangkan kehadiran manusia saja merupakan ancaman pasif karena umumnya satwa akan menghindar dari kehadiran manusia. Hasil observasi tidak menunjukkan bentuk ancaman aktif yaitu perburuan terhadap N. diardi, sedangkan ancaman pasif yang diidentifikasi berupa keberadaan pondok masyarakat ataupun basecamp (stasiun penelitian).
Keberadaan pondok masyarakat sebagian besar tersebar di dalam kawasan hutan seperti yang terdapat pada kawasan Punggualas dan Sungai Bulan, termasuk juga Bukit Bulan. Pondok-pondok tersebut bersifat sementara karena dibuat masyarakat selama berburu burung di dalam kawasan sehingga tidak dijadikan parameter di dalam penelitian seperti di kawasan Punggualas. Penggunaan parameter pondok dilakukan di kawasan Bukit Bulan karena lokasinya terkonsentrasi selama penelitian, sedangkan pondok lain di kawasan Bukit Bulan ditemukan rusak. Gambar 13 menunjukkan peta sebaran N. diardi terhadap kelas
“jarak dari potensi gangguan”.
Kriteria kelas “jarak dari potensi gangguan” dikelompokkan menjadi lima
pada masing-masing kelas “jarak dari potensi gangguan” dapat dilihat pada Tabel 6.
(a.1)
(a.2)
(b)
Tabel 6 Komposisi sebaran Neofelis diardi di masing-masing kelas “jarak dari
Berdasarkan penjelasan mengenai bentuk perjumpaan dalam setiap sebaran, N. diardi banyak dijumpai secara tidak langsung. Kondisi ini merupakan salah satu indikasi bahwa N. diardi menghindari manusia. Hal ini dibuktikan oleh sebaran terbanyak N. diardi yang berada di kelas sangat jauh dari potensi gangguan. Sifat elusive predator tersebut juga dialami selama penelitian, yaitu N. diardi tidak kembali ke tempat yang sebelumnya ditemukan jejak (footprints) dengan selang waktu hanya satu hari. Cheyne et al. (2013) juga menemukan N. diardi jantan dengan jarak 700 m dari basecamp CIMTROP di Taman Nasional Sebangau (TNS). Selain itu, fenomena seperti itu juga ditemukan oleh Fata (2011) yang menjelaskan keberadaan Panthera tigris sumatrae lebih banyak dijumpai lokasi yang jauh dari letak permukiman dan aktivitas manusia dengan kelas jarak 15 – 20 km, bahkan Yanti (2011) menemukan keberadaan Panthera pardus melas lebih banyak di lokasi yang jauh dari permukiman masyarakat dengan jarak > 1 km.
Keberadaan N. diardi di dekat sumber gangguan diduga karena mengikuti satwa mangsanya. Lokasi basecamp di Punggualas terletak di pinggir Sungai Punggualas yang menjadi tempat berkumpul beberapa jenis satwa mangsa seperti Macaca fascicularis dan Macaca nemestrina, bahkan sungai tersebut juga merupakan habitat bagi berbagai jenis ikan yang juga menjadi mangsa N. diardi. Di samping itu, Sus barbatus, Muntiacus atherodes, dan Tragulus javanicus juga kerap ditemukan dekat pondok di Bukit Bulan. Kasus serupa juga pernah dilaporkan di area kerja CIMTROP, bahkan N. diardi memasuki bangunan camp yang kemudian ditangkap untuk dipindahkan dan dilepasliarkan kembali.
hutan untuk bersiap masuk ke habitat padang rumput (grassland), bahkan juga ditemukan menggunakan habitat bekas kebun masyarakat.
Hasil uji Chi-square menunjukkan adanya hubungan yang signifikan antara keberadaan N. diardi terhadap jarak dari potensi gangguan seperti pondok ataupun basecamp dengan nilai (P = 0.017) sehingga (P < 0.05). Kondisi ini juga menjelaskan N. diardi rentan berpengaruh terhadap keberadaan dan aktivitas manusia. Oleh karena itu, keberadaan potensi gangguan dapat menjadi pertimbangan untuk monitoring N. diardi di kawasan TNS.
Sebaran menurut kerapatan tajuk hutan (canopy cover)
Kondisi tajuk hutan di kawasan TNS bervariasi yang diduga akibat eksploitasi yang terjadi sebelum menjadi kawasan konservasi, namun secara umum masih tergolong rapat. Menurut Pusat Penelitian Biologi LIPI (2007) bagian hutan primer di TNS merupakan vegetasi sekunder bekas tebangan yang kondisinya relatif baik karena tajuknya yang masih rapat dan terdiri dari tiga strata. Gambar 14 merupakan peta sebaran N. dardi terhadap berbagai kondisi kerapatan tajuk di kawasan TNS terutama daerah SSI, Punggualas, dan Bukit Bulan.
Kriteria kelas kerapatan penutupan tajuk (canopy cover) dibagi menjadi lima, yaitu tidak bertajuk, renggang, sedang, rapat, dan sangat rapat. Sebaran terbanyak N. diardi berada di kriteria kelas sangat rapat dengan 12 perjumpaan (66.66%), sedangkan proporsi sebaran di kelas lainnya meliputi 5.56% untuk kelas tidak bertajuk dan renggang serta 22.22% untuk kelas sedang. Oleh karena itu, diduga terdapat hubungan antara keberadaan N. diardi terhadap faktor kerapatan tajuk dan keberadaan hutan primer. Komposisi sebaran N. diardi pada masing-masing kelas kerapatan tajuk dapat dilihat di Tabel 7.
Tabel 7 Komposisi sebaran Neofelis diardi terhadap kelas kerapatan tajuk Kriteria Kelas
menemukan N. nebulosa betina menggunakan hutan tertutup (closed forest) di hutan semi evergreen.
(a)
(b)
(c)
Keberadaan N. diardi di kelas tidak bertajuk berhubungan dengan tipe habitat yang digunakan karena di kelas tersebut merupakan tipe habitat lahan terbuka di Sebangau Sanitra Indah (SSI) yaitu padang rumput bekas lahan terbakar. Sebaran di kelas sedang juga berhubungan dengan kondisi habitat di pinggir Sungai Punggualas yang berupa semak belukar dan ditumbuhi rerumputan. Grassman et al. (2005) juga menemukan keberadaan N. nebulosa menggunakan daerah terbuka berupa padang rumput. Selain itu, masing-masing sebaran di kedua kelas juga berhubungan dengan keberadaan satwa mangsanya.
Satwa arboreal seperti N. diardi membutuhkan kondisi kerapatan penutupan tajuk hutan yang baik untuk menunjang segala aktivtas dan kebutuhan kesehariannya, bukan hanya sebagai penyedia mangsa, namun juga sebagai tempat beristirahat atau bersarang. Chiang (2007) juga mengatakan N. nebulosa memanfaatkan pepohonan untuk beristirahat dan berburu. Menurut Grzimeks (1975) N. nebulosa beristirahat dan tidur di pohon dengan membangun sarang. N. nebulosa beraktivtas secara arboreal dengan proporsi 54% dibandingkan dengan hidup secara terestrial (Rabinowitz 1988). Selain itu, Griffiths (1993) diacu dalam Chiang (2007) menemukan kecenderungan N. nebulosa memangsa satwa primata yang ditunjukkan oleh hasil analisis fesesnya yaitu empat dari tujuh scat adalah primata.
Hasil uji Chi-square bernilai (P = 0.04) atau (P < 0.05) yang berarti ada hubungan yang signifikan antara keberadaan N. diardi dengan kondisi kerapatan penutupan tajuk hutan (canopy cover). Kecenderungannya N. diardi memilih habitat dengan penutupan tajuk yang rapat. Dengan demikian, parameter penutupan tajuk dapat dijadikan pertimbangan dalam monitoring N. diardi. Selain itu, spesies tersebut juga dapat dijadikan pertimbangan sebagai indikator bagi kondisi hutan yang masih baik.
Sebaran menurut kedalaman gambut
Sebaran tingkat kedalaman gambut di TNS mempunyai dua pola, yaitu keberadaan depresi (gambut dalam) di tengah-tengah dan/atau menyamping mengikuti bentuk pulau delta. Menurut Gandasasmita et al. (2006) kondisi depresi pada lahan gambut tidak selalu di tengah-tengah, namun seringkali menyamping mengikuti bentuk pulau delta. Pola depresi menyamping dijumpai di daerah Punggulas dan SSI bagian hilir menuju muara Sungai Sebangau. Kondisi tersebut ditunjukkan oleh Gambar 15 yang merupakan peta sebaran Neofelis diardi terhadap kelas tingkat kedalaman gambut.
(a)
(b)
(c)
Tabel 8 Komposisi sebaran Neofelis diardi pada kelas kedalaman gambut
Gambut dalam cenderung mempunyai tingkat kesuburan lahan yang rendah sehingga gambut bersifat oligotrofik (Noor 2001 dan Gandasasmita et al. 2006). Selain itu, lahan di gambut dalam mempunyai tingkat kemasaman yang tinggi sehingga air bersifat sangat masam dan kerapatan lindak juga sangat labil karena umumnya daerah tersebut merupakan bagian kubah gambut (peat-dome) serta merupakan gambut ombrogen (Andriesse 2003 dan Noor 2001). Driesse and Soepraptohardjo (1974) diacu dalam Noor (2001) juga menambahkan semakin dalam gambut, maka kandungan abu semakin rendah, unsur Ca dan Mg menurun, serta tanah bereaksi menjadi lebih masam. Oleh karena itu, hanya vegetasi tertentu yang tumbuh di gambut dalam. Menurut Anderson (1961) diacu dalam Whitmore (1984), bentuk vegetasi di daerah kubah gambut umumnya kerdil dengan bentuk tipe hutan padang keruntum yang berupa semak-semak dengan ketinggian setidaknya mencapai 2 m ataupun tipe hutan tiang dengan ciri vegetasi yang kecil, seragam, pendek, dan sangat rapat.
Sebaran N. diardi di kelas kedalaman ekstrem diindikasikan oleh bentuk pemanfaatan untuk keperluan yang lebih secretive ataupun sebagai cover seperti bereproduksi, menyimpan mangsa, berlindung, bersembunyi, dan merawat anak. Kondisi ini dikarenakan daerah tersebut sulit dimasuki oleh manusia maupun satwa besar terestrial. Di samping itu, sifat N. diardi yang adaptif terhadap berbagai tipe habitat dan hidup secara arboreal sehingga untuk mencapai daerah gambut dalam dapat memanfaatkan berbagai vegetasi yang sangat rapat. Gunawan (2010) juga menjelaskan pemanfaatan berbagai tipe habitat oleh Panthera pardus melas di Jawa Tengah dan menunjukkan bahwa vegetasi dengan tumbuhan bawah yang mencapai tinggi 2 m dan rapat dimanfaatkan satwa tersebut sebagai cover untuk bersembunyi dan berlindung. Gunawan et al. (2012) menambahkan karakteristik habitat yang dijadikan tempat berlindung Panthera pardus melas adalah hutan bervegetasi lebat dan sulit diakses oleh manusia.
tinggi, Kandungan mineral tersebut mengandung unsur yang dibutuhkan oleh satwa herbivora sehingga tampak juga adanya hubungan dengan ketersediaan satwa mangsa N. diardi.
Hasil uji Chi-square bernilai (P = 0.002) atau (P < 0.05) yang berarti adanya hubungan yang signifikan antara keberadaan N. diardi dengan tingkat kedalaman gambut. Kondisi ini menunjukkan adanya kecenderungan untuk memilih salah satu kriteria kelas kedalaman gambut. Oleh karena itu, tingkat kedalaman gambut dapat dipertimbangkan untuk monitoring N. diardi di kawasan TNS.
Sebaran menurut sub tipe habitat
Kriteria kelas berdasarkan tipe habitat dibagi menjadi lima yaitu lahan terbuka (Open Land), Mixed Swamp Forest (MSF), Tall Interior Forest (TIF), Low Pole Forest (LPF), dan hutan perbukitan (Hill Forest). Sebaran N. diardi terbanyak dijumpai di tipe habitat TIF dengan 12 perjumpaan (66.66%), sedangkan tiga perjumpaan (16.67%) masing-masing di hutan terbuka dan perbukitan. Oleh karena itu, sebaran N. diardi di TNS diduga berhubungan dengan tipe habitat TNS. Gambar 16 menunjukkan peta sebaran N. diardi pada berbagai tipe habitat di TNS. Komposisi sebaran N. diardi terhadap masing-masing tipe habitat dapat dilihat pada Tabel 9.
Purwadi (2010) menjelaskan jenis tumbuhan di tipe habitat TIF berjumlah lebih banyak dibandingkan yang ditemukan di MSF dan LPF. Suyanti (2007) juga menambahkan tipe habitat TIF mempunyai rata-rata tinggi tegakan tertinggi dibandingkan dengan MSF dan LPF dengan nilai 28.92 m yang juga termasuk kelas tinggi. Tipe habitat TIF dan MSF mempunyai strukur tajuk (kanopi) yang lebih beragam dan berbeda dibandingkan LPF sehingga TIF diduga sebagai tipe habitat dengan tingkat kesuburan yang baik. Selain itu, Alikodra (2002) menjelaskan bahwa kelimpahan jenis satwaliar berhubungan dengan kesuburan maupun keragaman habitat seperti kelimpahan tumbuhan yaitu semakin melimpah tumbuhannya maka semakin melimpah pula satwaliarnya. Oleh karena itu, tipe habitat TIF diduga merupakan habitat yang mampu mendukung kehidupan N. diardi.
(a)
(b)
(c)
N. diardi diduga memilih jalur pergerakannya. Berdasarkan hasil analisis terhadap struktur vegetasi di tiga lokasi camera trap yang digunakan sebagai jalur N. diardi, ditemukan bahwa diameter, tinggi total, dan tinggi bebas cabang tegakan menjadi pertimbangan pemilihan oleh N. diardi untuk pergerakan hariannya sebagai satwa arboreal. Ketiga aspek tersebut dibagi menjadi tiga kelas seperti penelitian Purwadi (2010). Di Taman Nasional Sebangau kondisi vegetasi hutan rawa gambutnya memiliki karakteristik yang sangat berbeda dengan ekosistem hutan lainnya. Galdikas (1984) diacu dalam Purwadi (2010) juga menjelaskan tegakan di TN Sebangau mempunyai pertumbuhan yang sangat terbatas.
Gambar 17 Diagram kelas diameter tegakan tumbuhan di plot camera trap Neofelis diardi
Gambar 18 Diagram kelas tinggi total tegakan tumbuhan di plot camera trap Neofelis diardi
Keterangan : Klasifikasi Kelas (Purwadi 2010)
CTP : Kode titik lokasi pemasangan camera trap di area Punggualas Pendek : Kelas tinggi total tegakan < 12 m
Sedang : Kelas tinggi total tegakan 12 – 19 m Tinggi : Kelas tinggi total tegakan > 19 m
Keterangan : Klasifikasi Kelas (Purwadi 2010)
CTP : Kode titik lokasi pemasangan camera trap di area Punggualas Kecil : Kelas diameter tegakan < 12.52 cm
Kondisi diameter di ketiga lokasi terbagi dalam kelas diameter kecil (< 12.52 cm), sedang (12.52 – 21.87 cm), dan besar (> 21.87 cm) dengan rentang nilai sebesar 7.1 – 54.08 cm dan rataan 21.43 cm. N. diardi cenderung memilih lokasi dengan karakteristik diameter yang didominasi oleh kelas diameter besar dan kecil (Gambar 17). Diamater yang besar sangat kuat untuk menjadi tumpuan tubuh N. diardi ketika beraktivitas secara arboreal.
Tinggi total tegakan terbagi dalam kelas pendek (< 12 m), sedang (12 – 19 m), dan tinggi ( > 19 m) dengan rentang nilai 6.8 m dan 33 m serta rataan sebesar 13.19 m. N. diardi cenderung memilih lokasi dengan tinggi total tegakan yang didominasi oleh kelas ketinggian pendek (Gambar 18). Kelas pendek yang mendominasi diduga dapat dimanfaatkan oleh N. diardi untuk memperoleh mangsa yang berada di tajuk dari permukaan tanah (terestrial) maupun dari atas tajuk yang lebih tinggi karena jumlah tegakan yang tidak mendominasi.
Gambar 19 Diagram kelas tinggi bebas cabang tumbuhan di plot camera trap Neofelis diardi
Kelas Tinggi Bebas Cabang (TBC) terbagi dalam kelas pendek ( < 8 m), sedang (8 – 14 m), dan.tinggi ( > 14 m) dengan rentang nilai antara 3.5 – 26.5 m dan rataan sebesar 8.99 m. N. diardi cenderung memilih lokasi dengan tinggi bebas cabang yang didominasi oleh kelas pendek dan sedang (Gambar 19). Grassman (2003) menemukan N. nebulosa berada pada cabang pohon yang besar dan rendah dengan ketinggian mencapai ±3 m untuk istirahat. Selain itu, menurut catatan Chiang (2007) N. nebulosa juga menyimpan sisa mangsa di cabang pohon dengan ketinggian 4 m. Selain itu, struktur vegetasi berperan penting sebagai cover bagi satwa predator (Bailey 1984 dan Shaw 1985 diacu dalam Gunawan 2010).
Beberapa kesamaan komponen pelengkap habitat ditemukan di ketiga lokasi camera trap yang digunakan sebagai jalur N. diardi, yaitu keberadaan tanda gesekan tubuh satwa seperti babi berjenggot (Sus barbatus) dan sarang orangutan (Pongo pygmaeus), bahkan sarang orangutan juga ditemukan dekat dengan lokasi penemuan tanda claw panjat N. diardi yang berjarak 3 – 7 m. Kedua satwa
Keterangan : Klasifikasi Kelas (Purwadi 2010)
CTP : Kode titik lokasi pemasangan camera trap di area Punggualas Pendek : Kelas tinggi bebas cabang tegakan < 8 m
tersebut potensial menjadi mangsa N. diardi terutama anak (infant) dan remaja (juvenile) (Payne et al. 2000 dan Wilting 2007). Kasus pemangsaan terhadap orangutan belum ditemukan bukti pendukungnya. Kondisi ini juga terjadi pada kasus pemangsaan macan tutul (Panthera pardus) terhadap gorilla (Gorilla gorilla) di Afrika yang sejauh ini belum ditemukan kejadian langsung, meskipun Henschel (2008) telah memperoleh hasil identifikasi komponen mangsa dari feses Panthera pardus yaitu berupa bagian tubuh dari gorilla. Kedua kejadian tersebut hanya dapat terjadi pada satwa yang berumur muda dan sedang jauh dari pengawasan induknya atau satwa yang sakit, terluka, dan mati setelah perkelahian. Pemanfaatan sarang orangutan juga diduga untuk beristirahat maupun tidur seperti halnya pemanfaatan lubang/liang pohon yang terbentuk oleh beruang madu (Helarctos malayanus). Salah satu bentuk pemanfaatan lubang/liang pohon juga ditunjukkan oleh burung untuk bersarang (Gambar 20). Prater (1965) juga menambahkan N. nebulosa muda tercatat berada di lubang pohon. Menurut Grzimeks (1975) N. nebulosa beristirahat dan tidur di pohon, juga disertai membangun sarang.
Gambar 20 Sarang burung di lubang pohon yang dirusak beruang madu Potensi satwa mangsa (preys) di habitat TIF mempunyai kekayaan dan keanekaragaman jenis yang lebih tinggi dibandingkan dengan tipe habitat Hill Forest (Tabel 10). Potensi satwa mangsa di Lokasi TIF (Punggualas) ditemukan sebanyak 24 jenis, sedangkan di Bukit Bulan 10 jenis (Tabel 11). Babi berjenggot (Sus barbatus) merupakan mangsa dengan kelas kelimpahan umum dan melimpah di kedua tipe habitat yaitu 27.96 (Punggualas) dan 42.62 (Bukit Bulan).
Tabel 10 Nilai berbagai indeks potensi satwa mangsa Neofelis diardi
No Lokasi Tipe Habitat Dmg H‟
Tabel 11 Klasifikasi potensi satwa mangsa Neofelis diardi
2 Owa ungko Hylobates agilis albybarbis
Keterangan : Klasifikasi (Liley & Robertson 2000 diacu dalam Jamaksari 2011) J : Jarang U : Umum S : Sering TU : Tidak Umum M : Melimpah
Callosciurus baluensis, Nannosciurus melanotis, dan Lhopura erythrophthalma. Menurut Griffiths (1993) diacu dalam Chiang (2007) N. nebulosa cenderung memilih mangsa terutama jenis primata yang diketahui melalui fecal analysis dengan hasil 4 dari 7 bagian scats adalah primata. Selain itu, Chiang (2007) juga menyebutkan laporan dari penduduk asli di Taiwan yang menjelaskan N. nebulosa senang memakan monyet taiwan (Macaca cyclopis), sedangkan Lhopura swinhoii potensial menjadi mangsa. Grasmaan et al. (2005) menambahkan bajing tanah Indochina (Menetes berdmorei) merupakan mangsa terbanyak yang ditemukan dalam feses N. nebulosa .
Tabel 11 Klasifikasi potensi satwa mangsa Neofelis diardi (lanjutan) No Spesies
19 Kucing batu Pardofelis marmorata Keterangan : Klasifikasi (Liley & Robertson 2000 diacu dalam Jamaksari 2011)
J : Jarang U : Umum S : Sering TU : Tidak Umum M : Melimpah