Stabilitas Transgen Dan Ekspressi Gen B11 Pada Tanaman Tembakau Transgenik Generasi T3.
Teks penuh
Gambar
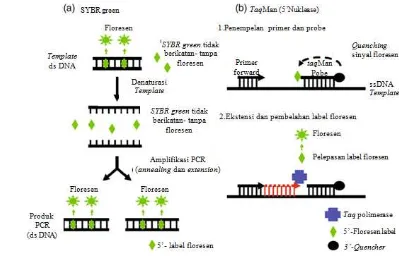
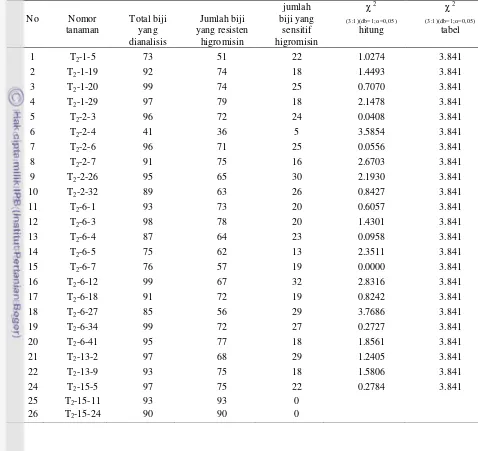
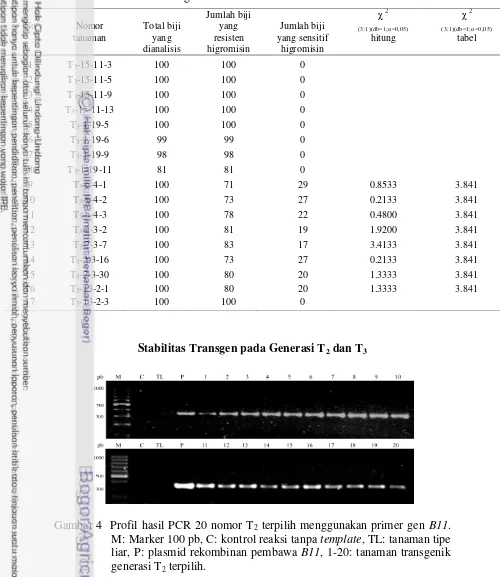
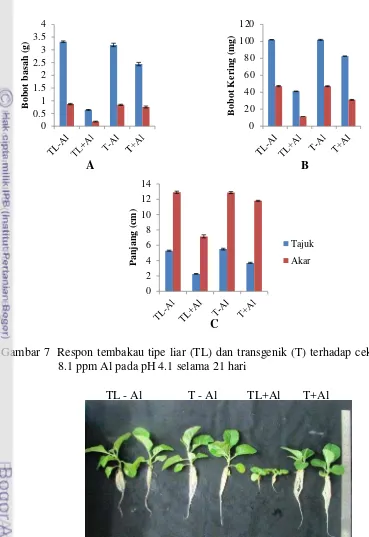
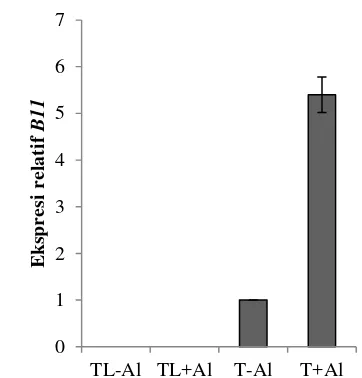
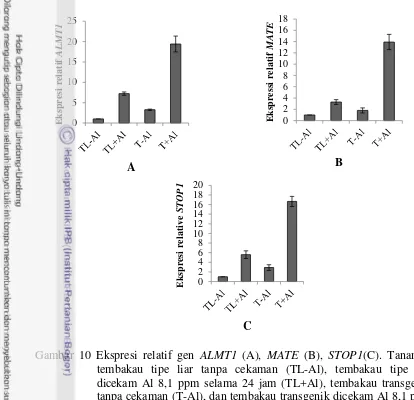
Dokumen terkait
Hasil penelitian menunjukkan galur tanaman transgenik pembawa gen RB memiliki tinggi tanaman dan jumlah daun yang seragam antar galur, namun berbeda dalam jumlah
Pada tingkat konsentrasi 100 mg/L dari 364 benih padi yang terdiri dari 115 tanaman nontransgenik dan 249 tanaman transgenik terseleksi, namun terjadi gejala
Efikasi gen RB pada umbi tanaman transgenik, setelah 10 hari diinokulasi dengan isolat Lembang, Pangalengan, dan Galunggung, menunjukkan bahwa gen RB tidak mampu
Efikasi gen RB pada umbi tanaman transgenik, setelah 10 hari diinokulasi dengan isolat Lembang, Pangalengan, dan Galunggung, menunjukkan bahwa gen RB tidak mampu
Penelitian ini bertujuan: (1) melakukan persilang- an untuk memasukkan gen ketahanan (RB) terhadap hawar daun dari tanaman kentang transgenik Katah- din SP904 dan SP951 ke
Selain itu, ekspresi ZmLEA14tv pada tanaman model dikotil seperti tembakau secara signifikan meningkatkan kemampuan pemulihan, sedangkan peningkatan jagung
Penelitian ini bertujuan: (1) melakukan persilang- an untuk memasukkan gen ketahanan (RB) terhadap hawar daun dari tanaman kentang transgenik Katah- din SP904 dan SP951 ke
Hasil penelitian yang diukur pada pengamatan terhadap tanaman uji adalah (a) Kualitas total DNA dari tanaman kacang tanah transgenik dan non transgenik dan (b) Hasil PCR berupa
