CYTOTAXONOMY
PTERIS ENSIFORMIS
BURM.f.
MUHAMMAD EFENDI
THE GRADUATE SCHOOL
BOGOR AGRICULTURAL UNIVERSITY BOGOR
STATEMENT OF RESEARCH ORIGINALITY AND
INFORMATION SOURCE
This is to verify that my thesis entitled: Cytotaxonomy Pteris ensiformis Burm.f. is true represent result of my research and have never been publicized. All information and data used have been expressed clearly and can be checked its truth. I devolved copyright of my thesis to Bogor Agricultural University.
RINGKASAN
MUHAMMAD EFENDI. Sitotaksonomi Pteris ensiformis Burm.f. Dibimbing oleh TATIK CHIKMAWATI dan DEDY DARNAEDI.
Penelitian tentang sitotaksonomi Pteris ensiformis Burm.f. dari berbagai daerah di Indonesia telah dilakukan. Variasi infraspesifik pada P. ensiformis dan hubungan intraspesies dari P. ensiformis dapat digunakan untuk memperjelas batasan infraspesies dari P. ensiformis itu sendiri. Penelitian ini bertujuan untuk mengkaji variasi morfologi, sitologi dan tipe reproduksi pada P. ensiformis dan mengkaji hubungan antara tipe reproduksi, tingkatan ploidi dengan variasi morfologi pada P. ensiformis.
Penelitian ini dilakukan sejak September 2012 sampai Juni 2013. Pengamatan morfologi dilakukan pada 75 individu yang telah dikoleksi dari Gorontalo, Lombok dan berbagai tempat di Jawa dan ditambah dengan spesimen dari Herbarium Bogoriense (BO). Pengataman sitologi dilakukan dengan cara memeriksa kromosom somatik pada ujung akar dengan menggunakan metode squash. Analisis tipe reproduksi dilakukan dengan cara menghitung jumlah spora pada setiap sporangium.
Sebuah rekaman data sitologi terbaru pada P. ensiformis var. victoriae telah ditemukan dari Indonesia yaitu 2n=58 dengan tipe diploid seksual dari Gorontalo, Sulawesi dan 2n=87 dengan tipe triploid dari Pulau Lombok dan Bogor Jawa Barat. Sitotipe diploid var. victoriae memiliki habitus yang lebih kecil dibanding dengan sitotipe triploid var. victoriae. Sitotipe tetraploid seksual (2n=116) var. ensiformis yang telah dikoleksi dari berbagai tempat di Jawa menambah data sito-reproduksi dari var. ensiformis di Indonesia. Berdasarkan data morfologi, P. ensiformis secara konsisten dapat dikelompokkan ke dalam dua kelompok yaitu P. ensiformis var. ensiformis dan P. ensiformis var. victoriae. Ke depannya, penelitian untuk memperjelas hubungan antara varietas dalam P. ensiformis sangat direkomendasikan.
SUMMARY
MUHAMMAD EFENDI. Cytotaxonomy Pteris ensiformis Burm.f. Supervised by TATIK CHIKMAWATI and DEDY DARNAEDI.
The cytotaxonomy of Pteris ensiformis Burm.f. from several places in Indonesia have been studied. The information about infraspesific variation of P. ensiformis and the relationships within P. ensiformis can be used to clarify the intraspecies delimitation of P. ensiformis. The objectives of this study were to examine morphological, cytological and mode reproduction of P. ensiformis and to clarified the correlation between mode of reproduction, ploidy level and morphological variation in P. ensiformis. Therefore, the aim of this study was to examined the intraspesific variation of P. ensiformis in Indonesia.
This study was conducted from September 2012 to June 2013. The morphological character was examined from 60 collections of living plants originated from Gorontalo, Lombok and various places in Java, added the specimen from Herbarium Bogoriense (BO). Cytological observations were conducted by observing somatic chromosome of root tips using squash methods. Modes of reproduction were determined by counting the number of spores of each sporangium.
New cytological record of P. ensiformis var. victoriae have been found from Indonesia i.e 2n=58 (diploid sexual) from Gorontalo, Celebes and 2n=87 (triploid) from Lombok Island and Bogor West Java. The diploid sexual of var. victoriae has smaller in habitus than that of the triploid races. Cytotype tetraploid sexual (2n=116) of var. ensiformis, collected from various places in Java is an important information of cyto-reproduction data of var. ensiformis in Indonesia. Based on morphological data, P. ensiformis consistenly can be classified into two varieties namely P. ensiformis var. ensiformis and P. ensiformis var. victoriae. However, futher study to clarify their relationship among varieties is strongly recommended.
.
Copy right© 2014, Bogor Agricultural University
All right reversed by Law
It is prohibited to cites all parts of this thesis without referring to and mention the source
a. Citation only permitted for the sake of education, research, sciencific writing, critical writing or reviewing scientific problems.
b. Citation does not inflict the name and honor of Bogor Agricultural University
Thesis
Submitted as partial fulfillment requirement for The Master Degree in Plant Taxonomy
CYTOTAXONOMY
PTERIS
ENSIFORMIS
BURM.f.
THE GRADUATE SCHOOL
BOGOR AGRICULTURAL UNIVERSITY BOGOR
2014
Tittle : Cytotaxonomy Pteris ensiformis Burm.f. Names : Muhammad Efendi
Reg. Number : G353110261
Approved by, Supervisor committee
Dr. Ir. Tatik Chikmawati, M.Si (Chairman)
Prof. Dr. Dedy Darnaedi, M.Sc (Member)
Head of Plant Biology Graduate Program
Dr. Ir. Miftahudin, M.Si
Dean of Graduate School
Dr. Ir. Dahrul Syah, MSc.Agr
PREFACE
A study of cytoreproduction in fern species with the title “Cytotaxonomy Pteris ensiformis Burm.f.” was conducted from September 2012 to June 2013. Field study was carried out at Gorontalo, Lombok and several places in Java i.e Bogor, Sukabumi, Subang, Banyumas, Banjarnegara, Wonogiri and Pacitan. The morphological and reproduction mode observations were conducted at the Plant Taxonomy Laboratory of Biology Department, Bogor Agricultural University. While, cytological observation and specimen identifications were conducted at Laboratory of Morphological, Anatomical and Cytological of Herbarium Bogoriense (BO).
A part of this study was presented on The 9th International Flora Malesiana Symposium in Bogor, 27th – 31st Agust 2013 with the tittle “ A New Cytotype of Pteris ensiformis var. victoriae from Malesian Region” and the manuscript of the article was submitted to Reinwardtia Journal.
I greatly appreciate to my supervisors: Dr. Ir. Tatik Chikmawati, M.Si and Prof. Dr. Dedy Darnaedi, M.Sc for their advice, guidance and encouragements throughout this study. I wish to thank Dr. Sunaryo, Ujang Hapidz, Widoyanti, Eka, and Drs. Erwin from Herbarium Bogoriense for helping during laboratory and photograph work. I also thanks to my friends from Plant Biology for their support and discussion.
Finally, my grateful goes to my dear father (Harun who has passed away 25 years ago) and my mother (Ibu Ngatini), my brother (Kang Ibik) and my sister (Mbak Rom) and especially my little family (Bahiyah and our child Athifatuzzahra) and Mr. Bunyamin’s family for their moral supports.
I hope this thesis will be usefull.
CONTENTS
LIST OF TABLE vi
LIST OF FIGURE vi
1 INTRODUCTION 1
Background 1
Objectives 2
2 METHODS 2
Times and location 2
Plant Materials 3
Methods 3
Data analysis 4
3 RESULT AND DISCUSSION 5
Morphological variations 6
Somatic chromosome number and mode of reproduction 8
Phenetic analysis 13
4 CONCLUSSION 14
REFERENCES 15
LIST OF TABLE
1 Ecological data of Pteris ensiformis 5
2 Morphological characters of Pteris ensiformis 6
3 Spore sizes of Pteris ensiformis 8
4 Cytological and reproduction modes of Pteris ensiformis in various
regions in Indonesia 9
5 Stomata sizes of Pteris ensiformis 11
LIST OF FIGURE
1 Study sites. 1. Pelabuhan Ratu Sukabumi 2. Bogor 3. Subang 4. Banyumas 5. Banjarnegara 6. Wonogiri 7. Pacitan 8. Mataram, Lombok
9. Gorontalo. 2
2 Morphological character of Pteris ensiformis. A. Sterile leave; B.
Fertile leave. 4
3 Natural habitat of Pteris ensiformis A. P. ensiformis var. victoriae B-E.
P. ensiformis var. ensiformis 5
4 Variation of pinnae colour. A. Green to dark green; B. Green to
yellowish; C. Variegated pinnae. 7
5 Variation in ratio of lamina and stipe length (L/S). A. L/S > 1; B.
L/S=1; C. L/S<1.. 7
6 Spores of Pteris ensiformis. A. Cytotype diploid sexual of var. victoriae; B. Cytotype tetraploid sexual of var. ensiformis; C. Cytotype
tetraploid sexual of var. ensiformis. 8
7 Somatic chromosome number of Pteris ensiformis; A. P. ensiformis var. victoriae, 2n=58; B. P. ensiformis var. victoriae, 2n=87; C. P.
ensiformis var. ensiformis, 2n=116. 10
8 Three cytotypes of Pteris ensiformis. A. P. ensiformis var. ensiformis tetraploid sexual type; B. P. ensiformis var. victoriae triploid type; C. P.
ensiformis var. victoriae diploid type. 11
9 Kariotype of P. ensiformis. A. P. ensiformis var. victoriae; diploid sexual type; B. P. ensiformis var. ensiformis; tetraploid sexual type
(Bar=3 µm). 12
10 Dendogram of Pteris ensiformis based on morphological character. E= var. ensiformis that chromosome number still unknown; ET= var. ensiformis tetraploid races; VT= var. victoriae triploid races; VD= var.
victoriae diploid races. 13
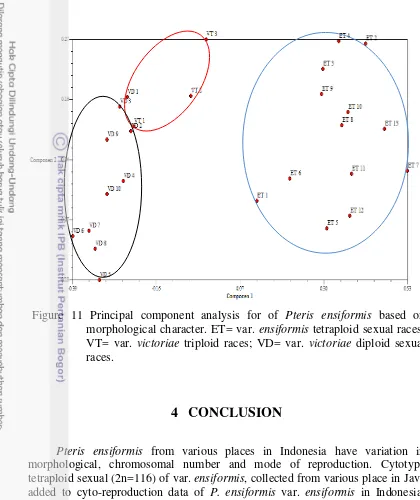
11 Principal component analysis for of Pteris ensiformis based on morphological character; ET= var. ensiformis tetraploid races; VT= var. victoriae triploid races; VD= var. victoriae diploid sexual races. 14
LIST OF APPENDIXES
1 Matric coding based on morphological character
24
1
INTRODUCTION
Background
Pteris L. is a large fern genus belong to the family Pteridaceae. This genus consists of about 300 species in the world. It is a cosmopolitan fern genus which predominantly distributed in all tropical and subtropical countries. It has diverse habitat although most species occur in forest, frequenly at secondary forests in opening or along rocky stream banks (Holltum 1966). Backer and Posthumus (1939) reported 19 species found in Java and de Winter and Amoroso (2003) reported 80 species of Pteris L. found in South-east Asia.
Pteris ensiformis Burm.f. is a member of genus Pteris L. that has wide distribution. P. ensiformis naturally occurs in Ceylon, India, China, throughout South-east Asia, Northen Australia and Polinesia. This species grows terrestrially in diverse habitat, such as in shady places, under rock, in crevices or old wet walls. P. ensiformis is easily distingushed from its distinctly dimorphic leaves. Sterile leaf is oblong with serrate in the margin whereas fertile leaf is simple with sori a long leaf margin and protected by false indusia (Holttum 1966).
Based on the morphological characters, P. ensiformis was consist of three varieties i.e var. ensiformis Burmann, var. victoriae Backer and var. rheophilla Kato, Darnaedi et Iwatsuki. P. ensiformis var. ensiformis has full green leave, while var. victoriae has variegated leave (Holttum 1966). P. ensiformis var. rheophilla has flexible leaf axes with serrate in leaf margine (Kato et al. 1991).
The diversity and distribution of fern was related to the environmental factors, i.e soil, light, rain, wind, temperature, elevation and also biogeography (Holttum 1966). Kato et al. (1991) reported that rheophyte ferns have morphological and physiological adaptation to the specific habitat, included root-system, leaf axes and leaf form. Fern rheophytes also have stronger habitus than dryland one, as shown for Diplazium subsinuatum var. rheophillum. Darnaedi (1987) reported that mode of rproduction and ploidy level have strong correlation with morphological character and size of leaf as demonstrated by Dryopteris sparsa growing in Mt. Gede- Pangrango. Perwati (2009) reported that the higher level of the plant ploidy caused bigger stomata and spore size of Adiantum raddianum. Ploidy levels presumable related to the elevation and temperature of plant habitat (Perwati 2009). Since P. ensiformis distributed in wide areas, intraspesific variation was also found in this species.
4
length (g), pinnae set number, pinnae density, pinnae colour, pinnae margin form, ratio of stipe size and lamina length, ratio of lamina and leave length, and ratio of lowest pinnae and lamina length (Figure 2).
Cytological observation
Cytological observation were carried out by observing somatic chromosome of root tips using squash method (Manton 1950) modified by Darnaedi (1991). Root tips were pretreated in 0.002 M 8-hydroxyquinolin for 24 hours at 20OC, and then they were fixed in 45% asetat acid for 10 minutes. The fixed root was macerated in a mixture of 45% asetat acid and 1 N HCl (1:3) for 3-4 minutes at 60oC then stained with 2% arceto orcein solution. The number of chromosomes were counted during observation under a microscope Olympus CX31 with 100X objective lens. Photograph were taken using Nikon Camera in Herbarium Bogoriense.
Reproduction mode observation
Reproduction modes were determined by counting the number of spores within each sporangium from 75 individuals. Five to 10 sporangia were observed for each individuals. Sporangium with 64 normal spores were treated as sexual reproduction type, while the sporangium with 32 spores treated as apogamous one (Walker 1962).
Data analysis
The qualitative and quantitative data of morphological character were scoring as multivariate data. Simple matching (SM) index was used to analyze the coefficient of similarity. Unweight pair group with aritmethical average (UPGMA) was used for clustering analysis. A dendogram were obtained using TREE. Then, morphological data of known cytotype was analyzed using matrix plot. All data analyses were performed using NTSys PC versi 21 programme (Rohlf 1998).
3
RESULT AND DISCUSSION
Ecology and distribution of Pteris ensiformis
6
Morphological variation of Pteris ensiformis
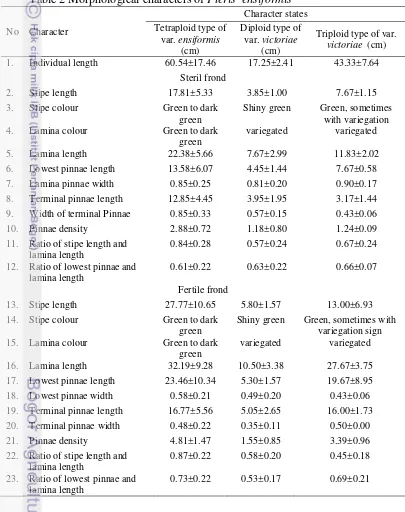
Pteris ensiformis showed variation in morphological characters i.e. individual length, stipe length, stipe colour, lamina colour, lamina length, ratio of lamina length and stipe length, pinnae density and ratio of lowest pinnae and lamina length (Table 2). The morphological variation of P. ensiformis may related to environmental and genetic factors.
Table 2 Morphological characters of Pteris ensiformis No Character
1. Individual length 60.54±17.46 17.25±2.41 43.33±7.64
Steril frond
2. Stipe length 17.81±5.33 3.85±1.00 7.67±1.15
3. Stipe colour Green to dark
green
Shiny green Green, sometimes with variegation
4. Lamina colour Green to dark
green
variegated variegated
5. Lamina length 22.38±5.66 7.67±2.99 11.83±2.02
6. Lowest pinnae length 13.58±6.07 4.45±1.44 7.67±0.58
7. Lamina pinnae width 0.85±0.25 0.81±0.20 0.90±0.17
8. Terminal pinnae length 12.85±4.45 3.95±1.95 3.17±1.44
9. Width of terminal Pinnae 0.85±0.33 0.57±0.15 0.43±0.06
10. Pinnae density 2.88±0.72 1.18±0.80 1.24±0.09
11. Ratio of stipe length and lamina length
0.84±0.28 0.57±0.24 0.67±0.24
12. Ratio of lowest pinnae and lamina length
0.61±0.22 0.63±0.22 0.66±0.07
Fertile frond
13. Stipe length 27.77±10.65 5.80±1.57 13.00±6.93
14. Stipe colour Green to dark
green
Shiny green Green, sometimes with variegation sign
15. Lamina colour Green to dark
green
variegated variegated
16. Lamina length 32.19±9.28 10.50±3.38 27.67±3.75
17. Lowest pinnae length 23.46±10.34 5.30±1.57 19.67±8.95
18. Lowest pinnae width 0.58±0.21 0.49±0.20 0.43±0.06
19. Terminal pinnae length 16.77±5.56 5.05±2.65 16.00±1.73
20. Terminal pinnae width 0.48±0.22 0.35±0.11 0.50±0.00
21. Pinnae density 4.81±1.47 1.55±0.85 3.39±0.96
22. Ratio of stipe length and lamina length
0.87±0.22 0.58±0.20 0.45±0.18
23. Ratio of lowest pinnae and lamina length
0.73±0.22 0.53±0.17 0.69±0.21
9
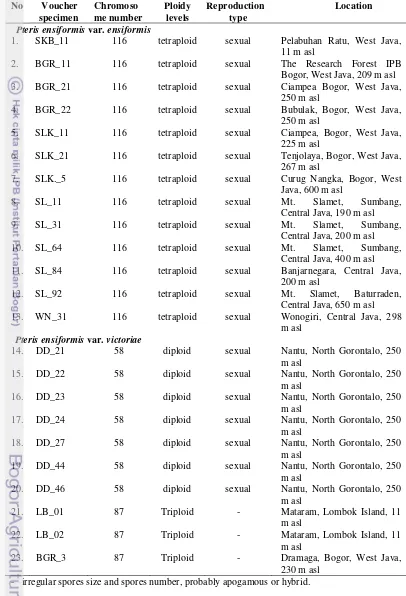
Table 4 Cytological and reproduction modes of P. ensiformis in various regions in Indonesia
Pteris ensiformis var. ensiformis
1. SKB_11 116 tetraploid sexual Pelabuhan Ratu, West Java,
6. SLK_21 116 tetraploid sexual Tenjolaya, Bogor, West Java,
267 m asl
11. SL_84 116 tetraploid sexual Banjarnegara, Central Java,
200 m asl
12. SL_92 116 tetraploid sexual Mt. Slamet, Baturraden,
Central Java, 650 m asl
13. WN_31 116 tetraploid sexual Wonogiri, Central Java, 298
m asl Pteris ensiformis var. victoriae
14
Figure 11 Principal component analysis for of Pteris ensiformis based on morphological character. ET= var. ensiformis tetraploid sexual races; VT= var. victoriae triploid races; VD= var. victoriae diploid sexual races.
4
CONCLUSION
15
REFERENCES
Abraham A. 1962. Studies on the cytology and phylogeni of the Pteridophytes VII observation on one hundred spesies of South India fern. J Ind Bot Soc 41: 344-348.
Aldasoro JJ, Cabezas F, Aedo C. 2004. Diversity and distribution of fern in Sub-Saharan Africa, Madagaskar and some islands of the South Atlantic. J Pteris (Pteridaceae). J Bot 2012:1-7.
Darnaedi D. 1987. Sitotaksonomi Dryopteris sparsa di Taman Nasional Gunung Gede-Pangrango. Floribunda 1(2):5-8.
Darnaedi D. 1991. Informasi kromosom : Pelatihan Sitogenetika Tumbuhan. Bogor (ID) : Herbarium BO Puslitbang Biologi. hlm 1-8.
Darnaedi D, Kato M, Iwatsuki K. Five new or ill-defined species related to Dryopteris sparsa (Dryopteridaceae). J Jap Bot. 64(10): 299-312.
de Winter WP, Amoroso VB. 2003. Plant Resources of South-east Asia Cryptogams : Ferns and Fern Allies. Bogor (ID): PROSEA. hlm 166-170. Gifford E, Foster AS. 1989. Morphology and Evolution Plants. New York (US):
W.H Freeman & Co. hlm. 176-177.
Holttum RE. 1966. A Revised Flora of Malaya. Volume II. Singapura (SG): Government Printing Off. hlm 393-412.
Kato M, Darnaedi D, Iwatsuki K. 1991. Fern rheophytes of Borneo. J Fac Sci Univ Tokyo III : 37-56.
Klekowski EJ. 1979. Sexual and subsexual systems in homosporous Pterydophytes: a new hypothesis. Amer J Bot 60: 535-544.
Mahabale TS, Kamble SY. 1981. Cytology of fern and other Pteridophytes of Western India. Proc. Indian natn. Sci. Acad. B47 (2): 260-278.
Manton I. 1950. Problems Cytology and Evolution in the Pteridophyta. New York (US): Cambridge Univ Pr. hlm:158-208.
Mendez, M.M., L.C. Gazarini, M.L. Rodrigues. 2001. Aclimation of Myrtus communis to contrasting Mediteranean light environments-effects on structure and chemical composition of foliage and plant water relation. Environ Exper Bot 45(2): 165-178.
Munir A. 2003. Keanekaragaman jenis tumbuhan paku di Taman Nasional Rawa Aopa Watumohai Sulawesi Tenggara [thesis]. Bogor (ID): Sekolah Pascasarjana IPB.
Perwati LK. 2009. Analisis derajat ploidi dan pengaruhnya terhadap variasi ukuran stomata dan spora pada Adiantum raddianum. Bioma 11(2):39-44. Praptosuwiryo TN, Darnaedi D. 1995. Survai kromosom tumbuhan paku liar di
16
Praptosuwiryo TN, Darnaedi D. 2008. Cytological observation on fern genus Pteris in Bogor Botanic Garden. Bul Kebun Raya 11: 15-24.
Rohlf FJ. 1998. NTSys-PC. Numerical Taxonomy and Multivariate Analysis. Version 2.0. User Guide. New York (US): Applied Biostatistic Inc. hlm 105-110.
Rugayah, Retnowati A, Windadri FI, Hidayat A. 2004. Pengumpulan Data Taksonomi. Di dalam : Rugayah, Widjaja EA, Praptiwi, editor. Pedoman Pengumpulan Data Keanekaragaman Flora. Bogor (ID) : Puslit Biologi LIPI. hlm 5-42.
Sharpe JM, Mehltreter K. 2010. Ecological Insights from Fern Population Dynamic. In : Mehltreter K, Walker LR, Sharpe JM, editor. Fern Ecology. New York (US): Cambridge Univ Pr. Pg 61-110.
Walker TG. 1962. Cytology and evolution in the fern genus Pteris L. Evolution 16:27-4.