LAHAN PERTANIAN DI JAWA
TIRA SITI NUR AFIAH
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
TIRA SITI NUR AFIAH. Isolasi dan Karakterisasi Bateri Penghasil Acyl Homoserine Lactonase (AHL-laktonase) Asal Lahan Pertanian di Jawa. Dibimbing oleh IMAN RUSMANA dan ALINA AKHDIYA.
Quorum sensing (QS) adalah mekanisme komunikasi bakteri yang bergantung pada jumlah populasi bakteri tersebut dan akumulasi senyawa autoinducer (AI). Quorum sensing pada kelompok bakteri Gram negatif melibatkan senyawa acyl homoserine lactone (AHL) untuk mengatur beberapa ekspresi gen, termasuk gen virulensi. Senyawa anti-QS merupakan senyawa yang dapat mendegradasi AHL sehingga gen virulensi tidak dapat diekspresikan. Acyl homoserine lactonase (AHL-laktonase) yang disandikan oleh gen aiiA merupakan salah satu enzim yang dapat berperan sebagai anti-QS. Penelitian ini bertujuan untuk mengisolasi dan mengkarakterisasi bakteri penghasil AHL-laktonase yang berasal dari lahan pertanian di Jawa. Bioesei aktivitas degradasi AHL menggunakan bakteri indikator Chromobacterium violaceum. Verifikasi keberadaan AHL-laktonase pada isolat-isolat yang menunjukkan hasil bioesei positif dilakukan dengan amplifikasi gen aiiA. Dari 12 isolat bakteri yang menunjukkan aktivitas degradasi AHL, terdapat dua isolat yang memiliki gen aiiA yaitu isolat INT1c dan SGT3g. Kedua isolat tersebut memiliki bentuk sel batang, penataan sel berantai, berendospora, dan termasuk dalam kelompok bakteri Gram positif. Analisis sekuen gen aiiA menunjukkan bahwa gen aiiA isolat INT1c memiliki homologi dengan gen aiiA bakteri Bacillus cereus ATTCC 14579 sedangkan isolat SGT3g memiliki homologi dengan gen aiiA bakteri Bacillus thuringiensis BMB171. Identifikasi molekuler berdasarkan gen 16S rRNA menunjukkan bahwa isolat INT1c memiliki kemiripan terdekat dengan bakteri Bacillus cereus strain GM3 (98%) sedangkan isolat SGT3g memiliki kemiripan terdekat dengan bakteri Bacillus thuringiensis strain A1-1 (99%).
Kata kunci: anti-quorum sensing, AHL-laktonase, gen aiiA.
TIRA SITI NUR AFIAH. Isolation and Characterization of Acyl Homoserine Lactonase (AHL-lactonase) Producing Bacteria from Agricultural Field in Java. Under supervision of IMAN RUSMANA and ALINA AKHDIYA.
Quorum sensing (QS) is a bacterial communication mechanism correlating with the number of bacterial population and accumulation of autoinducer (AI). Quorum sensing in Gram-negative bacteria involves a AI group of acyl homoserine lactone (AHL) to regulate the expression of several genes, including virulence genes. Anti-quorum sensing compounds can degrade AHL so that virulence genes can not be expressed. Acyl homoserine lactonase (AHL-lactonase) encoded by the aiiA gene is one enzyme that can act as an anti-quorum sensing. This study aimed to isolate and characterize the AHL-lactonase producing bacteria isolated from agricultural field in Java. Activity of AHL degradation bioassay using Chromobacterium violaceum as a bacterial indicator. Verification the presence of AHL-laktonase was performed by gene aiiA amplification. As many as 12 bacterial isolates showed AHL degradation activity and two isolates had the aiiA gene i.e. INT1c and SGT3g isolates. These isolates had bacil cells with chain arrangement and endospores. They were grouped in Gram-positive bacteria. AiiA gene sequence analysis showed that aiiA gene of INT1c had homology with aiiA gene of Bacillus cereus ATTCC 14579 while the SGT3g isolate had homology with aiiA gene of Bacillus thuringiensis BMB171. Molecular identification based on 16S rRNA gene showed that INT1c isolate had the closest similarity with the Bacillus cereus strain GM3 (98%) while SGT3g isolate had the closest similarity with the Bacillus thuringiensis strains A1-1 (99%).
TIRA SITI NUR AFIAH
Skripsi
sebagai salah satu syarat untuk memperoleh gelar
Sarjana Sains pada
Departemen Biologi
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
NIM
: G34070022
Departemen : Biologi
Disetujui
Pembimbing I
Pembimbing II
Dr. Ir. Iman Rusmana, M.Si
Alina Akhdiya, M.Si
NIP. 196507201991031002
NIP. 196812082001122001
Diketahui
Ketua Departemen Biologi
Fakultas Matematika dan Ilmu Pengetahuan Alam Institut Pertanian Bogor
Dr. Ir. Ence Darmo Jaya Supena, M.S NIP.
196410021989031002
sehingga karya ilmiah ini dapat diselesaikan. Tema penelitian ini adalah anti-quorum sensing, dengan judul Isolasi dan Karakterisasi Bakteri Penghasil Acyl Homoserine Lactonase (AHL-Laktonase) Asal Lahan Pertanian di Jawa. Penelitian ini dilaksanakan pada bulan November 2010 hingga Mei 2011, bertempat di Laboratorium Mikrobiologi, Departemen Biologi, Institut Pertanian Bogor.
Penulis mengucapkan terima kasih kepada Bapak Dr. Ir. Iman Rusmana, M.Si dan Ibu Alina Akhdiya, M.Si selaku pembimbing. Ucapan terima kasih juga penulis sampaikan kepada Bapak, Almarhum Mama, serta seluruh keluarga, atas doa dan kasih sayangnya. Di samping itu, penulis mengucapkan terima kasih kepada Atun, Ka Fina, Bang Jo, Dwi, teman-teman yang tergabung dalam IR-Crew, teman yang telah bersedia mengambil sampel tanah, teman-teman seperjuangan: Bian, Inong, Iyut, Maya, Mayang, Nunu, Tri, dan teman-teman-teman-teman di laboratorium mikrobiologi yang telah membantu dalam penelitian ini.
Semoga karya ilmiah ini bermanfaat.
Bogor, Agustus 2011
Ai Djubaedah. Penulis merupakan anak keempat dari empat bersaudara. Tahun 2007 penulis lulus dari SMA Negeri 1 Sukanagara. Pada tahun yang sama, penulis lulus seleksi masuk IPB melalui jalur Undangan Seleksi Masuk IPB pada Departemen Biologi Fakultas Matematika dan Ilmu Pengetahuan Alam. Penulis mendapatkan beasiswa Peningkatan Prestasi Akademik (PPA) dari DIKTI pada tahun 2008-2011.
DAFTAR TABEL ... vi
DAFTAR GAMBAR ... vi
DAFTAR LAMPIRAN ... vi
PENDAHULUAN ... 1
Latar Belakang ... 1
Tujuan ... 1
METODE ... 1
Waktu dan Tempat Penelitian ... 1
Isolasi, Pemurnian, dan Penyimpanan Bakteri ... 1
Bioesei Aktivitas Degradasi AHL ... 2
Pengamatan Morfologi Sel dan Pewarnaan Gram ... 2
Amplifikasi Gen Penyandi AHL-Laktonase ... 2
Identifikasi Isolat ... 2
Sekuensing DNA dan Analisis Bioinformatika ... 2
Konstruksi Pohon Filogenetik ... 2
HASIL ... 2
Bioesei Aktivitas Degradasi AHL ... 2
Pengamatan Morfologi Sel dan Pewarnaan Gram ... 3
Amplifikasi Gen Penyandi AHL-Laktonase ... 4
Identifikasi Isolat ... 4
Konstruksi Pohon Filogenetik ... 5
PEMBAHASAN ... 5
SIMPULAN ... 6
SARAN ... 7
DAFTAR PUSTAKA ... 7
1 Isolat bakteri yang diperoleh dari lahan pertanian di Jawa dan aktivitas degradasi AHL ... 3
2 Diameter zona degradasi AHL pada 12 isolat ... 3
3 Karakteristik morfologi sel dan pewarnaan Gram isolat-isolat yang menunjukkan
aktivitas degradasi AHL ... 4
4 Hasil analisis sekuen gen aiiA isolat INT1c & SGT3g dengan menggunakan program BLAST-N ... 5
5 Hasil analisis sekuen gen aiiA isolat INT1c & SGT3g dengan menggunakan program
BLAST-X ... 5
6 Hasil analisis sekuen gen 16S rRNA isolat INT1c & SGT3g dengan menggunakan
program BLAST-N ... 5
DAFTAR GAMBAR
Halaman
1 Zona degradasi AHL 12 isolat terpilih ... 3
2 Visualisasi amplikon gen aiiA pada gel agarose 1% ... 4
3 Visualisasi amplikon 16S rRNA pada gel agarose 1% ... 4
4 Pohon filogenetik berdasarkan sekuen gen aiiA dari isolat INT1c dan SGT3g yang
dibandingkan dengan sekuen gen aiiA dari beberapa spesies bakteri lainnya ... 5
DAFTAR LAMPIRAN
Halaman
1 Komposisi media pertumbuhan yang digunakan ... 9
2 Hasil sekuensing dan analisis bioinformatik gen aiiA dan 16S rRNA dengan program
Bakteri fitopatogen merupakan bakteri yang menyebabkan infeksi atau penyakit pada tumbuhan. Pengendalian bakteri patogen dapat dilakukan dengan menggunakan senyawa antibiotik atau antimikrob. Masalah yang dihadapi dalam penggunaan antibiotik adalah timbulnya resistensi bakteri terhadap antibiotik tersebut. Hal ini mendorong dilakukannya berbagai penelitian tentang pengendalian bakteri patogen yang yang lebih aman dan tidak dikhawatirkan akan menimbulkan resistensi.
Defoirdt et al. (2004) menyatakan bahwa ekspresi gen virulensi pada beberapa bakteri patogen diregulasi oleh proses quorum sensing. Quorum sensing (QS) adalah mekanisme komunikasi bakteri sebagai suatu pengaturan ekspresi gen yang bergantung pada jumlah populasi bakteri tersebut dan akumulasi senyawa autoinducer (AI) (Rukayadi & Hwang 2009). QS terlibat dalam regulasi fungsi biologi yang penting seperti luminescence, produksi antibiotik, transfer plasmid, motilitas, virulensi, pembentukan biofilm dan berperan dalam regulasi ekspresi gen-gen patogen pada Pseudomonas solanacearum, Pseudomonas aeruginosa, dan beberapa spesies Erwinia (Dong et al. 2000; Zang & Dong 2004).
Proses QS melibatkan molekul kimia yang disebut autoinducer (AI). AI merupakan molekul sinyal yang disekresikan, diakumulasi, diserap kembali, dan dikenali oleh bakteri selama proses QS terjadi (Rukayadi & Hwang 2009). Salah satu jenis molekul AI adalah N-acylhomoserine lactone (AHL) yang dihasilkan oleh bakteri Gram negatif, serta digunakan untuk komunikasi dalam spesies yang sama (Rukayadi & Hwang 2009).
Salah satu prinsip pengendalian virulensi bakteri fitopatogen dengan dasar QS adalah mencegah akumulasi AHL dengan mendegradasi molekul sinyal tersebut. Senyawa atau molekul yang dapat mendegradasi molekul AHL disebut senyawa anti-QS. Anti-QS memiliki kelebihan dalam mengontrol infeksi bakteri yaitu tidak mempengaruhi pertumbuhan, sehingga dapat menghindarkan timbulnya tekanan seleksi yang sering menghasilkan generasi patogen yang lebih resisten terhadap antibiotik (White & Finan 2009). Beberapa enzim pendegradasi AHL telah diidentifikasi dari beberapa
adalah enzim Acyl Homoserine Lactonase (AHL-laktonase) (Dong et al. 2000). Enzim ini disandikan oleh gen aiiA yang dimiliki oleh Bacillus dan mampu mendegradasi AHL dengan menghidrolisis ikatan lakton yang terdapat pada molekul AHL (Dong et al. 2000). AHL-laktonase sebagai salah satu senyawa anti-QS dapat digunakan sebagai alternatif dalam pengendalian bakteri fitopatogen.
Sebagai salah satu negara megabiodiversity, Indonesia mempunyai kekayaan dan keragaman sumberdaya mikroba yang sangat besar dan berpotensi sebagai penghasil AHL-laktonase. Oleh karena itu, isolasi dan karakterisasi bakteri penghasil AHL-laktonase merupakan langkah awal untuk untuk memanfaatkan potensi tersebut.
Tujuan
Penelitian ini bertujuan untuk mengisolasi dan mengkarakterisasi bakteri penghasil AHL-laktonase asal lahan pertanian di Jawa.
METODE
Waktu dan Tempat
Penelitian ini dilaksanakan mulai bulan November 2010 sampai dengan Mei 2011, bertempat di Laboratorium Mikrobiologi Departemen Biologi FMIPA IPB.
Isolasi, Pemurnian, dan Penyimpanan Bakteri
menunjukkan morfologi yang berbeda dimurnikan dengan metode cawan gores. Setelah murni, bakteri disimpan dalam agar miring.
Bioesei Aktivitas Degradasi AHL
Isolat-isolat murni ditumbuhkan pada media Luria broth (LB). Setelah 48 jam, kultur disentrifus selama 10 menit dengan kecepatan 10000 rpm. Sebanyak 100 μL supernatant diteteskan pada paper disk dan diletakkan pada permukaan media cawan Luria agar (LA) yang mengandung Chromobacterium violaceum 1%. Cawan tersebut kemudian diinkubasi pada suhu ruang selama 24 jam. Media LB steril digunakan sebagai kontrol. Paper disk yang dikelilingi oleh zona tidak berwarna ungu menunjukkan adanya aktivitas degradasi AHL.
Pengamatan Morfologi Sel dan Pewarnaan Gram
Isolat-isolat yang menunjukkan hasil bioesei positif diamati morfologi sel (bentuk sel, penataan sel, dan endospora) dan reaksi terhadap pewarnaan Gram.
Amplifikasi Gen Penyandi AHL-laktonase DNA bakteri diisolasi dengan metode Lazo (Lazo et al. 1987). Isolat-isolat yang menunjukkan hasil bioesei degradasi AHL positif diamplifikasi menggunakan primer spesifik untuk gen aiiA yaitu aiiAF (5’-ATC GGA TCC ATG ACA GTA AAG AAG CTT ATT TCG-3’, dan aiiAR (5’-GTC GAA TTC CTC AAC AAG ATA CTC CTA ATG ATGT-3’ (Dong et al. 2000). Campuran PCR terdiri dari 1.5 mM MgCl2, 0.2 mM dNTPs, 0.3 µM masing-masing primer, 0.02 U/µL Taq DNA polymerase, template DNA, ddH2O, dan buffer DNA polymerase. Amplifikasi dilakukan selama 30 siklus PCR dengan kondisi pra-denaturasi (94oC, 10 menit), denaturasi (94oC, 30 detik), annealing (52oC, 30 detik), elongasi (72oC, 1 menit), dan post-PCR (72oC, 5 menit) (Chan et al. 2007). Produk PCR dilarikan pada mini-gel elektroforesis (agarose 1%) pada tegangan listrik 50 volt selama 45 menit (Sambrook et al. 1989). Visualisasi di atas UV Transiluminator dilakukan dengan pewarnaan Ethidium Bromida.
Identifikasi Isolat
Isolat-isolat yang menunjukkan hasil positif pada bioesei degradasi AHL dan amplifikasi gen aiiA diidentifikasi secara molekuler (16S rRNA). Amplifikasi gen 16S rRNA menggunakan primer 63f (5’-CAG GCC TAA CAC ATG CAA GTC-3’) dan 1387r (5’-GGG CGG WGT GTA CAA GGC-3’) (Marchesi et al. 1998). Amplifikasi dilakukan selama 30 siklus PCR dengan kondisi pra-PCR (94oC, 2 menit), denaturasi (92oC, 30 detik), annealing (50oC, 1 menit), elongasi (72oC, 1 menit), dan post-PCR (75oC, 5 menit).
Sekuensing DNA dan Analisis Bioinformatika
Pengurutan DNA hasil amplifikasi dilakukan dengan memanfaatkan jasa perusahaan sekuensing 1st Base di Singapura. Sekuen yang diperoleh dijajarkan dengan data pada GenBank menggunakan program Basic Local Alignment Search Tool-Nucleotida (BLAST-N) dan Basic Local Alignment Search Tool Nucleotide 6-frame translation-protein (BLAST-X) dari situs National Center for Biotechnology Information (NCBI) melalui http://www.ncbi.nlm.nih.gov untuk mengetahui tingkat kemiripan gen aiiA dan 16S rRNA dari isolat yang dianalisa.
Konstruksi Pohon Filogenetik
Konstruksi pohon filogenetik dilakukan dengan metode Neighbor Joining (NJ) dengan bootstrap 1000x menggunakan program MEGA 5.0.
HASIL
Bioesei Aktivitas Degradasi AHL
Tabel 1 Isolat bakteri yang diperoleh dari lahan pertanian di Jawa dan aktivitas degradasi AHL Sampel
Jumlah isolat hasil isolasi
Jumlah isolat yang menunjukan ativitas
degradasi AHL
Asal Jenis Jumlah
Jawa barat:
Bogor Filosfer 3 12 0
Tanah 3 25 0
Cianjur Filosfer 7 30 2
Tanah 6 40 1
Indramayu Tanah 6 69 5
Pangandaran Tanah 2 22 0
Sukabumi Tanah 1 3 0
Jawa Timur:
Lamongan Tanah 1 3 0
Nganjuk Tanah 1 3 0
Pekalongan Tanah 2 8 1
Rembang Tanah 2 12 0
Sragen Tanah 3 34 3
Total 37 261 12
Gambar 1 Zona degradasi AHL 12 isolat terpilih.
Diameter zona degradasi AHL yang dihasilkan oleh 12 isolat berkisar antara 8 sampai 11 mm (Tabel 2). Isolat yang menunjukkan diameter zona degradasi AHL terbesar adalah isolat PKT2a dan SGT3g
yaitu sebesar 11 mm, sedangkan isolat CJF4a, INT1b, dan INT1f menunjukkan diameter zona degradasi AHL terkecil yaitu 8 mm.
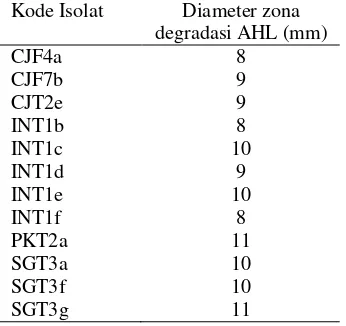
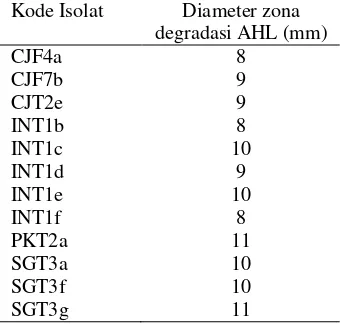
Tabel 2 Diameter zona degradasi AHL 12 isolat terpilih
Kode Isolat Diameter zona degradasi AHL (mm)
CJF4a 8
CJF7b 9
CJT2e 9
INT1b 8
INT1c 10
INT1d 9
INT1e 10
INT1f 8
PKT2a 11
SGT3a 10
SGT3f 10
SGT3g 11
Pengamatan Morfologi Sel dan Pewarnaan Gram
Tiga isolat yang menunjukkan bioesei aktivitas degradasi AHL positif termasuk dalam kelompok Gram negatif sedangkan sembilan isolat lainnya termasuk dalam kelompok Gram positif. Isolat INT1d dan INT1f memiliki bentuk sel bulat dengan penataan bergerombol sedangkan sepuluh isolat lainnya memiliki bentuk sel batang dengan penataan rantai. Tujuh isolat memiliki endospora, sedangkan lima isolat lainnya tidak.
CJF4a CJT2e
INT1c INT1d
INT1e PKT2a
SGT3a SGT3f SGT3g
Kontrol CJF7b
INT1b
~1.3 kb pewarnaan Gram isolat-isolat bakteri yang memiliki aktivitas degradasi AHL CJF4a Batang, rantai Positif, berendospora CJF7b Batang, rantai Positif, berendospora CJT2e Batang, rantai Negatif, tidak
berendospora INT1b Batang, rantai Positif, tidak
berendospora INT1c Batang, rantai Positif, berendospora INT1d Bulat,
bergerombol
Negatif, tidak berendospora INT1e Batang, rantai Positif, berendospora INT1f Bulat,
bergerombol
Positif, tidak berendospora PKT2a Batang, rantai Positif, berendospora SGT3a Batang, rantai Positif, berendospora SGT3f Batang, rantai Negatif, tidak
berendospora SGT3g Batang, rantai Positif, berendospora
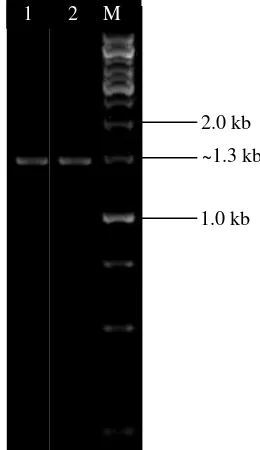
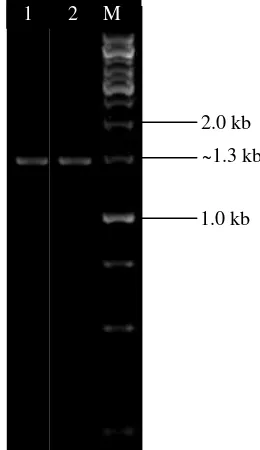
Amplifikasi Gen Penyandi AHL-laktonase Dari 12 isolat yang mampu mendegradasi AHL, dua isolat diantaranya (isolat SGT3g dan INT1c) menunjukkan hasil amplifikasi gen aiiA berukuran ~0.8 kb (Gambar 2).
Gambar 2 Visualisasi amplikon gen aiiA pada gel agarose 1% (M= 1 kb DNA ladder, Sumur 1= INT1c, Sumur 2= SGT3g).
Hasil analisis sekuen gen aiiA dengan menggunakan program BLAST-N menunjukkan bahwa gen aiiA isolat INT1c memiliki homologi dengan gen aiiA Bacillus cereus ATTCC 14579 dengan persen identitas sebesar 96% sedangkan isolat SGT3g memiliki homologi dengan gen aiiA Bacillus thuringiensis BMB171 dengan persen identitas sebesar 95% (Tabel 4). Analisis sekuen gen aiiA dengan program BLAST-X menunjukkan bahwa protein yang disandikan oleh gen aiiA pada kedua isolat memiliki homologi dengan struktur kristal AHL-laktonase pada B. thuringiensis (Tabel 5).
Identifikasi Isolat
Tabel 4 Hasil analisis sekuen gen aiiA INT1c & SGT3g dengan menggunakan program BLAST-N Kode isolat Sekuen bakteri yang homolog % identitas Nomor akses
INT1c Bacillus cereus ATCC 14579 96% AE016877.1
SGT3g Bacillus thuringiensis BMB171 95% CP001903.1
Tabel 5 Hasil analisis sekuen gen aiiA INT1c & SGT3g dengan menggunakan program BLAST-X
Kode isolat Sekuen bakteri yang homolog % identitas
Nomor akses INT1c Crystal Structure Of Quorum-Quenching
N-Acyl Homoserine Lactone Lactonase in Bacillus thuringiensis
82% 2BR6_A
SGT3g Crystal Structure Of Quorum-Quenching N-Acyl Homoserine Lactone Lactonase in Bacillus thuringiensis
98% 2BR6_A
Tabel 6 Hasil analisis sekuen gen 16S rRNA isolat INT1c & SGT3g dengan menggunakan program BLAST-N
Kode isolat Sekuen bakteri yang homolog %
identitas Nomor akses INT1c Bacillus cereus strain GM3 16S ribosomal
RNA gene, partial sequence
98% JF837192.1
SGT3g Bacillus thuringiensis strain A1-1 16S ribosomal RNA gene, partial sequence
99% JF496321.1
Konstruksi Pohon Filogenetik
Hasil analisis filogenetik gen aiiA isolat INT1c dan SGT3g yang dibandingkan dengan gen aiiA beberapa spesies bakteri di GenBank disajikan pada gambar 4.
Gambar 4 Pohon filogenetik berdasarkan sekuen gen aiiA dari isolat INT1c dan SGT3g yang dibandingkan dengan sekuen gen aiiA dari beberapa spesies bakteri lainnya.
PEMBAHASAN
Bakteri yang menghasilkan senyawa anti-quorum sensing dapat mendegradasi senyawa Acylhomoserine lactone (AHL). Bakteri ini dapat ditemukan pada habitat yang sama
dengan bakteri yang ber-quorum sensing (Chan et al. 2007). Keberadaan kedua jenis bakteri ini di alam menciptakan suatu keseimbangan ekosistem. Bakteri fitopatogen yang dapat ditemukan pada lahan pertanian mengindikasikan keberadaan bakteri penghasil senyawa anti-QS. Pada penelitian ini, eksplorasi bakteri filosfer dan tanah dari 10 sampel daun dan 27 sampel tanah yang berasal dari lahan pertanian di Jawa menghasilkan 12 isolat bakteri yang menunjukkan aktivitas degradasi AHL yang kuat.
Bioesei aktivitas degradasi AHL dilakukan untuk menyeleksi bakteri-bakteri yang menghasilkan senyawa pendegradasi AHL. Bakteri yang digunakan sebagai indikator terjadinya proses degradasi AHL adalah bakteri Chromobacterium violaceum. Bakteri C. violaceum termasuk dalam kelompok bakteri Gram negatif yang mudah ditemukan di tanah dan air, serta memproduksi pigmen ungu spesifik yang bernama violacein (McClean et al. 1997). Pigmen violacein diproduksi melalui proses QS. Penghambatan proses QS pada bakteri ini menyebabkan bakteri tidak dapat memproduksi pigmen violacein sehingga koloni bakteri tidak berwarna ungu.
Degradasi AHL dilakukan menggunakan supernatant yang diteteskan pada paper disk. Paper disk yang dikelilingi oleh zona tidak berwarna ungu menunjukkan adanya aktivitas
degradasi AHL oleh senyawa ekstraseluler yang dihasilkan oleh bakteri. Senyawa ekstraselular ini dapat berperan sebagai anti-QS.
Enzim yang dapat mendegradasi AHL dikelompokkan menjadi dua enzim utama yaitu AHL-laktonase dan AHL-acylase (Czajkowski & Jafra 2009). AHL-laktonase mampu menghidrolisis cincin lakton pada AHL dan termasuk ke dalam kelompok enzim superfamily metallolactamase (Liu et al. 2008). AHL-laktonase yang disandikan oleh gen aiiA merupakan anti-QS pertama yang diaplikasikan untuk mengontrol virulensi bakteri patogen (Dong et al. 2000).
Bioesei aktivitas degradasi AHL terhadap 260 isolat menunjukkan 12 isolat bakteri diantaranya mampu mendegradasi AHL. Paper disk kontrol dikelilingi oleh warna ungu yang disebabkan oleh pigmen violacein yang diproduksi bakteri C. violaceum melalui proses QS. Zona tidak berwarna ungu di sekeliling paper disk menunjukkan adanya suatu senyawa pada supernatant kultur yang dapat mencegah proses QS sehingga pigmen tersebut tidak dihasilkan. Diameter zona degradasi AHL yang dihasilkan oleh 12 isolat bervariasi. Diameter zona degradasi AHL terbesar dihasilkan oleh isolat PKT2a dan SGT3g yaitu sebesar 11 mm, sedangkan isolat CJF4a, INT1b, dan INT1f memiliki diameter zona degradasi AHL terkecil yaitu sebesar 8 mm. Hal ini menunjukan perbedaan aktivitas degradasi AHL oleh senyawa yang dihasilkan. Suhu dan pH optimum aktivitas AHL-laktonase adalah 200C dengan pH 8 (Chen et al. 2010). Aktivitas produksi enzim oleh bakteri juga dipengaruhi oleh beberapa faktor yaitu kandungan nutrisi, derajat keasaman media, tekanan osmotik, tingkat aerasi, suhu, dan kontrol terhadap kontaminasi selama inkubasi (Pandey et al. 2000).
Verifikasi keberadaan AHL-laktonase pada isolat-isolat yang menunjukkan hasil bioesei positif dilakukan dengan amplifikasi gen penyandi AHL-laktonase yaitu gen aiiA. Gen aiiA pertama kali ditemukan pada Bacillus sp. strain 240B1 (Dong et al. 2000). Homologi gen aiiA kemudian berhasil ditemukan pada beberapa strain bakteri seperti Bacillus thuringiensis, B. cereus, B. mycoides, dan B. amyloliquefaciens (Dong et al. 2002; Yin et al. 2010). Dari 12 isolat terpilih, dua isolat diantaranya menunjukkan amplifikasi gen aiiA yaitu isolat SGT3g dan INT1c. Amplifikasi gen aiiA pada isolat
INT1c dan SGT3g menghasilkan amplikon berukuran ~0.8 kb.
Analisis sekuen gen aiiA dengan program BLAST-X menunjukkan bahwa protein yang disandikan oleh gen aiiA pada isolat INT1c dan SGT3g mirip dengan struktur kristal AHL-laktonase pada B. thuringiensis. Tingkat kemiripan protein AHL-laktonase SGT3g sebesar 98% sedangkan tingkat kemiripan AHL-laktonase INT1c dengan AHL-laktonase B. thuringiensis hanya sebesar 82%. Hasil ini didukung oleh analisis dengan program BLAST-N dimana isolat SGT3g memiliki kemiripan 95% dengan gen aiiA pada B. thuringiensis BMB171 sedangkan gen aiiA isolat INT1c lebih mirip dengan gen aiiA pada Bacillus cereus ATCC 14579 (kemiripan 96%). Analisis filogenenetik gen aiiA isolat INT1c dan SGT3g menunjukkan bahwa isolat INT1c berkerabat dekat dengan B. cereus ATCC 14579 sedangkan isolat SGT3g berkerabat dekat dengan B. thuringiensis BMB171.
Identifikasi molekuler berdasarkan gen 16S rRNA menunjukkan bahwa isolat INT1c menunjukkan kemiripan 98% dengan dengan bakteri B. cereus strain GM3, sedangkan isolat SGT3g menunjukkan kemiripan 99% dengan bakteri B. thuringiensis strain A1-1. Bacillus cereus dan B. thuringiensis merupakan anggota genus Bacillus yang bersifat aerob atau anaerob fakultatif, termasuk dalam kelompok Gram positif, dan membetuk spora (Drobniewski 1993). Kedua bakteri tersebut telah dilaporkan memiliki gen penyandi AHL-laktonase yang homolog dengan gen aiiA pada Bacillus strain 240B1 (Dong et al. 2002; Dong et al. 2004). Dong et al. (2004) juga menyatakan bahwa B. thuringiensis mampu memproduksi enzim laktonase yang efektif untuk menginaktivasi autoinducer pada proses quorum sensing. Bacillus thuringiensis mampu mendegradasi AHL di lingkungannya sehingga dapat digunakan sebagai agen biokontrol terhadap virulensi bakteri yang dimediasi oleh sinyal AHL melalui quorum sensing (Dong et al. 2002).
SIMPULAN
penataan berantai, berendospora, dan termasuk dalam kelompok bakteri Gram positif. Identifikasi molekuler berdasarkan gen 16s rRNA menunjukkan bahwa isolat INT1c memiliki kemiripan terdekat dengan bakteri Bacillus cereus strain GM3 (98%) sedangkan isolat SGT3g memiliki kemiripan terdekat dengan bakteri Bacillus thuringiensis strain A1-1 (99%).
SARAN
Perlu dilakukan identifikasi molekuler gen pendegradasi AHL yang disandikan oleh gen selain aiiA pada sepuluh isolat terpilih lainnya.
DAFTAR PUSTAKA
Chan KG, Tiew SZ, Ng CC. 2007. Rapid isolation method of soil bacilli and screening of their quorum quenching activity. Asia Pasific J of Mol Biol and Biotechnol 15: 153-156.
Chen R, Zhou Z, Cao Y, Bai Y, Yao B. 2010. High yield expression of an AHL-lactonase from Bacillus sp. B546 in Pichia pastoris and its application to reduse Aeromonas hydrophia mortality in aquaculture. Microbial Cell Factories 9: 39-49.
Czajkowski R, Jafra S. 2009. Quenching of acyl-homoserine-dependent quorum sensing by enzymatic disruption of signal molecules. Acta Biochimica Polonica 56: 1-16.
Defoirdt T, Boon N, Bossier P, Verstraete W. 2004. Disruption of bacterial quorum sensing: an unexplored strategy to fight infections in aquaculture. Aquaculture 240: 69-88. quenching N-acyl homoserine lactonases from Bacillus species. Appl Environ Microbiol 68: 1754-1759.
Dong YH, Zhang XF, Xu JL, Zhang LH. 2004. Insecticidal B. thuringiensis silences Erwinia carotovora virulence by a new form of microbial antagonism, signal interference. Appl Environ Microbiol 70: 954-960. Drobniewski FA. 1993. Bacillus cereus and
related species. Clinical Microbiol Reviews 6: 324-338.
Lazo GR, Roffey R, Gabriel DW. 1987. Conservation of plasmid DNA sequences and pathovar identification of strain Xanthomonas campestris. Pytopathology 77: 1461-1467.
Liu D, et al. 2008. Mechanism of the quorum-quenching lactonase (AiiA) from Bacillus thuringiensis. 1. Product-bound strctures. Biochemistry 47: 7706-7714.
Marchesi JR, et al. 1998. Design and evaluation of useful bacterium-spesific PCR primer that amplify genes coding for bacterial 16S rRNA. Appl Environ Microbiol 64: 795-799.
McClean KH, et al. 1997. Quorum sensing and Chromobacterium violaceum: exploitation of violacein production and inhibition for the detection of N-acylhomoserine lactones. Microbiol 143: 3703-3711.
Pandey A, et al. 2000. Advances in microbial amylases. Biotechnol Appl Biochem 31: 135-152.
Rukayadi Y, Hwang JK. 2009. Pencegahan quorum sensing: suatu pendekatan baru dalam mengatasi infeksi bakteri. Medicinus 22: 22-27.
Sambrook J, Fritsch EF, Maniatis T. 1989. Molecular Cloning: a Laboratory Manual. Ed ke-2. New York: Cold Spring Harbor Laboratory.
White CE, Finan TM. 2009. Quorum sensing in Agrobacterium tumefaciens: chance or necessity?. J Bacteriol 191: 1123-1125.
Yin XT, et al. 2010. Isolation and characterization of an AHL-lactonase gene from Bacillus amyloliquefaciens. World J Microb Biotechnol 26:1361 – 1367.
Lampiran 1 Komposisi media pertumbuhan yang digunakan A. Media nutrien agar (NA)
- Agar 1.5%
- Nutrient broth 0.8%
B. Media King’s B agar
- Agar 1.5%
- Pepton 2%
- K2HPO4 0.15% - MgSO4.4H2O 0.15% - Gliserol 1.5%
C. Media luria agar (LA)
- Agar 1.5%
- NaCl 1%
- Tripton 1%
Lampiran 2 Hasil sekuensing dan analisis bioinformatika gen aiiA dan 16S rRNA dengan program BLAST-N dan BLAST X
A. Urutan nukleotida gen aiiA isolat INT1c
1 AAAAACACACAAACCTTACGTACTATCAGTTAAACGAAGATAGTTCAAAAAAAGTTTAT 70 71 AATTTATTTAAAAAAATTATTTAAACGAAAAAGTCATGAGCACGCGAACTCATGACTTTT 130 131 TGCACTATATATACTCAGGGAACACTCTACAACCCTTTTCCTGCTCTATATCATGACCAA 190 191 AGAAAACAATTGGTTTCTCTTTCGCCACAACTCCTTTTAAACGTTTAATTGAAGATAAAG 250 251 CTAATTCTGGATCAAATCCTGCGTTAAACGGCACTTCATCTTCAAAATTCTCTTTCGTGT 310 311 ACGATGCATCAATTGTTAATAAAACTGAGCCGGATTGCTCCGTCTCCAAATGAATAGCGA 370 371 CTGATAGGCCTGGAGAATGAATTCCCTGGCGCGAGTATACAATTAATTGAACACCTGGTA 430 431 CCACTTCATAATCCCCTTCAATAATTTTGTAGTTCAAATGCGGTAATATACATTCTTTCA 490 491 TATATTCTTCTCTATG 495
B. Analisis sekuen gen aiiA isolat INT1c dengan program BLAST-N
> gb|AE016877.1| Bacillus cereus ATCC 14579, complete genome
Length=5411809
Features in this part of subject sequence:
hypothetical protein
N-acyl homoserine lactone hydrolase
Score = 798 bits (432), Expect = 0.0
Identities = 470/485 (97%), Gaps = 15/485 (3%) Strand=Plus/Plus
Query 11 AAACCTTACGTACTATCAGTTAAACGAAGATAGTTCAAAAAAAGTTTATAATTTATTTAA 70 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 3409374 AAACCTTACGTACTATCAGTTAAACGAAGATAGTTCAAAAAAAGTTTATAATTTATTTAA 3409433
Query 71 AAAAATTATTTAAACGAAAAAGTCATGAGCACGCGAACTCATGACTTTTTGCACTATATA 130 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 3409434 AAAAATTATTTAAACGAAAAAGTCATGAGCACGCGAACTCATGACTTTTTGCACTATATA 3409493
Query 131 TACTCAGGGAACACTCTACAACCCTTTTCCTGCTCTATATCATGACCAAAGAAAACAATT 190 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 3409494 TACTCAGGGAACACTCTACAACCCTTTTCCTGCTCTATATCATGACCAAAGAAAACAATT 3409553
Query 191 GGTTTCTCTTTCGCCACAACTCCTTTTAAACGTTTAATTGAAGATAAAGCTAATTCTGGA 250 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 3409554 GGTTTCTCTTTCGCCACAACTCCTTTTAAACGTTTAATTGAAGATAAAGCTAATTCTGGA 3409613
Query 251 TCAAATCCTGCGTTAAACGGCACTTCATCTTCAAAATTCTCTTTCGTGTACGATGCATCA 310 |||||||||||| || ||||||||||||||||||||||||||||||||||||||||||| Sbjct 3409614 TCAAATCCTGCG--AA-CGGCACTTCATCTTCAAAATTCTCTTTCGTGTACGATGCATCA 3409670
Query 311 ATTGTTAATAAAACTGAGCCGGATTGCTCCGTCTCCAAATGAATAGCGACTGATAGGCCT 370 ||||||||||||||||||||||||||||||||||| || ||||||||||||||| ||||| Sbjct 3409671 ATTGTTAATAAAACTGAGCCGGATTGCTCCGTCTC-AA-TGAATAGCGACTGAT-GGCCT 3409727
Query 371 GGAGAATGAATTCCCTGGCGCGAGTATACAATTAATTGAACACCTGGTACCACTTCATAA 430 ||||||||| || ||||| |||||||| ||||||||||||||||||||||||||| Sbjct 3409728 GGAGAATGA---CC-TGGCG----TATACAAT-AATTGAACACCTGGTACCACTTCATAA 3409778
Query 431 TCCCCTTCAATAATTTTGTAGTTCAAATGCGGTAATATACATTCTTTCATATATTCTTCT 490 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 3409779 TCCCCTTCAATAATTTTGTAGTTCAAATGCGGTAATATACATTCTTTCATATATTCTTCT 3409838
Query 491 CTATG 495 |||||
C. Urutan nukleotida gen aiiA isolat SGT3g
1 CGGGATCCCATGACCAGTAAAAGAAGGCTTTTATTTTCCCCCCCCCCCC 49 50 TCCCAGCAAGGTCCGTTGTATTGGTTAGATCAATTCTTTCTGTTAATTAGTACACTCCGC 109 110 GCCGGGGGAATTTTATTGAACTTTACCTGTATGGTGTTATCTTTTTGGAGACAGAAGAGG 169 170 GGCCTATTTTAGTAGATACAGGTATGCCAGAAAGTGCAGTTAATAATGAAGGGATTTTTA 229 230 ACGGTACATTTGTTGAAGGACAGATTTTACCGAAAATGACTGAAGAAGATAGAATCGTGA 289 290 ATATATTAAAGCGTGTAGGGTATGAGCCGGACGACC-TTT-TATATTATTAGTTCTCACT 347 348 TACATTTTGATCATGCAGGAGGAAACGGTGCTTTTACAAATACACCGATTATTGTGCAGC 407 408 GAGCGGAATATGAGGCAGCACTTCATAGAGAAGAATATATGAAAGAATGTATATTACCGC 467 468 ATTTGAACTACAAAATTATTGAAGGGGATTATGAAGTGGTACCAGGTGTTCAATTATTGT 527 528 ATACGCCAGGTCATTCTCCAGGCCATCAGTCGTTATTCATTGAGACGGAGCAATCCGGTT 587 588 CAGTTTTATTAACAATTGATGCATCGTACACGAAAGAGAATTTTGAAGATGAAGTGCCGT 647 648 TCGCAGGATTTGATCCAGAATTAGCTTTATCTTCAATCAAACGCTTAAAAGAAGTTGTGA 707 708 CAAAAGAGAAATCGATTGTTTTCTTTGGTCATGATATAGAGCAGGAAAAAGGGTTGTAGA 767 768 GTGTTCCCGGAGTATATATAGTGCAAAAAGTCATGAGTTCGCGTGCTCATGACTTTTTCG 827 828 TTTAAATAATTTTTTTTAAATAAATTATAAACTTTTTTTCAAACTATCTTCCGTTTAACT 887 888 GGATAGTACGTAAAGTTTTTACATCAATTAAGGAGTATCCTTG 930
D. Analisis sekuen gen aiiA isolat SGT3g dengan program BLAST-N
> gb|CP001903.1| Bacillus thuringiensis BMB171, complete genome
Length=5330088
Features in this part of subject sequence:
hypothetical protein
N-acyl homoserine lactone hydrolase
Score = 1404 bits (760), Expect = 0.0
Identities = 846/883 (96%), Gaps = 24/883 (3%) Strand=Plus/Minus
Query 50 TCCCAGCAAGGTCCGTTGTATTGGTTAGATCAATTCTTTCTGTTAATTAGTACACTCCGC 109 ||||||| |||| ||||||| | |||||||| |||| ||||||||| | ||||||| ||| Sbjct 3334673 TCCCAGC-AGGT-CGTTGTA-T-GTTAGATC-ATTC-TTCTGTTAA-TGGTACACT-CGC 3334622
Query 110 GCCGGGGGAATTTTATTGAACTTTACCTGTATGGTGTTATCTTTTTGGAGACAGAAGAGG 169 ||| |||||| |||||||||| ||||||||||||||||||| |||||||||||||||||| Sbjct 3334621 GCC-GGGGAA-TTTATTGAAC-TTACCTGTATGGTGTTATC-TTTTGGAGACAGAAGAGG 3334566
Query 170 GGCCTATTTTAGTAGATACAGGTATGCCAGAAAGTGCAGTTAATAATGAAGGGATTTTTA 229 ||||||||||||||||||||||||||||||||||||||||||||||||||||| |||||| Sbjct 3334565 GGCCTATTTTAGTAGATACAGGTATGCCAGAAAGTGCAGTTAATAATGAAGGGCTTTTTA 3334506
Query 230 ACGGTACATTTGTTGAAGGACAGATTTTACCGAAAATGACTGAAGAAGATAGAATCGTGA 289 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 3334505 ACGGTACATTTGTTGAAGGACAGATTTTACCGAAAATGACTGAAGAAGATAGAATCGTGA 3334446
Query 290 ATATATTAAAGCGTGTAGGGTATGAGCCGGACGACC-TTT-TATATTATTAGTTCTCACT 347 |||||||||||||||||||||||||||||||||||| ||| ||||||||||||||||||| Sbjct 3334445 ATATATTAAAGCGTGTAGGGTATGAGCCGGACGACCTTTTATATATTATTAGTTCTCACT 3334386
Query 348 TACATTTTGATCATGCAGGAGGAAACGGTGCTTTTACAAATACACCGATTATTGTGCAGC 407 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 3334385 TACATTTTGATCATGCAGGAGGAAACGGTGCTTTTACAAATACACCGATTATTGTGCAGC 3334326
Query 408 GAGCGGAATATGAGGCAGCACTTCATAGAGAAGAATATATGAAAGAATGTATATTACCGC 467 || ||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 3334325 GAACGGAATATGAGGCAGCACTTCATAGAGAAGAATATATGAAAGAATGTATATTACCGC 3334266
Query 528 ATACGCCAGGTCATTCTCCAGGCCATCAGTCGTTATTCATTGAGACGGAGCAATCCGGTT 587 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 3334205 ATACGCCAGGTCATTCTCCAGGCCATCAGTCGTTATTCATTGAGACGGAGCAATCCGGTT 3334146
Query 588 CAGTTTTATTAACAATTGATGCATCGTACACGAAAGAGAATTTTGAAGATGAAGTGCCGT 647 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 3334145 CAGTTTTATTAACAATTGATGCATCGTACACGAAAGAGAATTTTGAAGATGAAGTGCCGT 3334086
Query 648 TCGCAGGATTTGATCCAGAATTAGCTTTATCTTCAATCAAACGCTTAAAAGAAGTTGTGA 707 ||||||||||||||||||||||||||||||||||||| ||||| ||||||| ||||||| Sbjct 3334085 TCGCAGGATTTGATCCAGAATTAGCTTTATCTTCAATTAAACGTTTAAAAGGAGTTGTGG 3334026
Query 708 CAAAAGAGAAATCGATTGTTTTCTTTGGTCATGATATAGAGCAGGAAAAAGGGTTGTAGA 767 | ||||||||| | ||||||||||||||||||||||||||||||||||| |||||||||| Sbjct 3334025 CGAAAGAGAAACCAATTGTTTTCTTTGGTCATGATATAGAGCAGGAAAA-GGGTTGTAGA 3333967
Query 768 GTGTTCCCGGAGTATATATAGTGCAAAAAGTCATGAGTTCGCGTGCTCATGACTTTTTCG 827 |||||||| ||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 3333966 GTGTTCCCTGAGTATATATAGTGCAAAAAGTCATGAGTTCGCGTGCTCATGACTTTTTCG 3333907
Query 828 TTTAAATAATTTTTTTTAAATAAATTATAAACTTTTTTTCAAACTATCTTCCGTTTAACT 887 |||||||||||||||| |||||||||||||||||||||| || |||||||| |||||||| Sbjct 3333906 TTTAAATAATTTTTTT-AAATAAATTATAAACTTTTTTTGAA-CTATCTTC-GTTTAACT 3333850
Query 888 GGATAGTACGTAAAGTTTTTACATCAATTAAGGAGTATCCTTG 930 | ||||||||||| ||||| |||||| | | |||||||| |||
Sbjct 3333849 G-ATAGTACGTAA-GTTTT-ACATCA-TCA-GGAGTATC-TTG 3333813
E. Analisis sekuen gen aiiA isolat INT1c dengan program BLAST-X
> pdb|2BR6|A Chain A, Crystal Structure Of
Quorum-Quenching N-Acyl Homoserine Lactone Lactonase
Score = 197 bits (500), Expect = 3e-51
Identities = 102/123 (83%), Positives = 108/123 (88%), Gaps = 5/123 (4%)
Frame = -1
Query 495
HREEYMKECILPHLNYKIIEGDYEVVPGVQLIVYSRQGIHSPGLSVAIHLETEQSGSVLL 316 HREEYMKECILPHLNYKIIEGDYEVVPGVQL+ HSPG ++ ++TEQSGSVLL
Sbjct 135 HREEYMKECILPHLNYKIIEGDYEVVPGVQLLYTPG---HSPGHQ-SLFIKTEQSGSVLL 190
Query 315
TIDASYTKENFEDEVPFNAGFDPELALSSIKRLKGVVAKEKPIVFFGHDIEQEKGCRVFP 136 TIDASYTKENFEDEVPF AGFDPELALSSIKRLK VV KEKPI+FFGHDIEQEK CRVFP
Sbjct 191
TIDASYTKENFEDEVPF-AGFDPELALSSIKRLKEVVKKEKPIIFFGHDIEQEKSCRVFP 249
F. Analisis sekuen gen aiiA isolat INT1c dengan program BLAST-X
> pdb|2BR6|A Chain A, Crystal Structure Of
Quorum-Quenching N-Acyl Homoserine Lactone Lactonase
pdb|2BTN|A Chain A, Crystal Structure And Catalytic
Mechanism Of The Quorum-
Quenching N-Acyl Homoserine Lactone Hydrolase Length=252
Sort alignments for this subject sequence by:
E value
Score Percent identity
Query
start position Subject start position
Score = 285 bits (728), Expect(4) = 1e-109
Identities = 138/143 (97%), Positives = 140/143 (98%), Gaps = 0/143 (0%)
Frame = +2
Query 329
YIISSHLHFDHAGGNGAFTNTPIIVQRAEYEAALHREEYMKECILPHLNYKIIEGDYEVV 508 YIISSHLHFDHAGGNGAFTNTPIIVQR
EYEAALHREEYMKECILPHLNYKIIEGDYEVV Sbjct 101
YIISSHLHFDHAGGNGAFTNTPIIVQRTEYEAALHREEYMKECILPHLNYKIIEGDYEVV 160
Query 509
PGVQLLYTPGHSPGHQSLFIETEQSGSVLLTIDASYTKENFEDEVPFAGFDPELALSSIK 688
PGVQLLYTPGHSPGHQSLFI+TEQSGSVLLTIDASYTKENFEDEVPFAGFDPELALSSIK Sbjct 161
PGVQLLYTPGHSPGHQSLFIKTEQSGSVLLTIDASYTKENFEDEVPFAGFDPELALSSIK 220
Query 689 RLKEVVTKEKSIVFFGHDIEQEK 757 RLKEVV KEK I+FFGHDIEQEK Sbjct 221 RLKEVVKKEKPIIFFGHDIEQEK 243
Score = 123 bits (308), Expect(4) = 1e-109
Identities = 59/60 (98%), Positives = 60/60 (100%), Gaps = 0/60 (0%)
Frame = +1
Query 154
LETEEGPILVDTGMPESAVNNEGIFNGTFVEGQILPKMTEEDRIVNILKRVGYEPDDLLY 333
LETEEGPILVDTGMPESAVNNEG+FNGTFVEGQILPKMTEEDRIVNILKRVGYEPDDLLY Sbjct 42
LETEEGPILVDTGMPESAVNNEGLFNGTFVEGQILPKMTEEDRIVNILKRVGYEPDDLLY 101
Score = 25.4 bits (54), Expect(4) = 1e-109
Identities = 9/12 (75%), Positives = 11/12 (92%), Gaps = 0/12 (0%)
Query 750 RKKGCRVFPEYI 785 ++K CRVFPEYI Sbjct 241 QEKSCRVFPEYI 252
Score = 24.6 bits (52), Expect(4) = 1e-109
Identities = 14/33 (42%), Positives = 16/33 (48%), Gaps = 7/33 (21%)
Frame = +3
Query 90 C*LVHSA---PGEFY*TLPVWCYLFGDRRG 170 C L HS+ PG+ LPVWCYL G Sbjct 16 CMLDHSSVNSALTPGKLL-NLPVWCYLLETEEG 47
G. Urutan nukleotida gen 16S rRNA isolat INT1c
1 GGAGAATGATAAGAGCTTGCTCTTATGAAGTTAGCGGCGGACGGGTGAGTAACACGTGGGT 61 62 AACCTGCCCATAAGACTGGGATAACTCCGGGAAACCGGGGCTAATACCGGATAACATTTT 121 122 GAACCGCATGGTTCGAAATTGAAAGGCGGCTTCGGCTGTCACTTATGGATGGACCCGCGT 181 182 CGCATTAGCTAGTTGGTGAGGTAACGGCTCACCAAGGCAACGATGCGTAGCCGACCTGAG 241 242 AGGGTGATCGGCCACACTGGGACTGAGACACGGCCCAGACTCCCTACGGGGAAGGCAGCC 301 302 AGTAGGGAATCTTCCGCAATGGAACGATAAGTTCTGACGGAGCAACCGCCGGCGTGAGTG 361 362 ATGAAGGCCTTTCGGGTTCGTAAAACTCTGTTGTTTAGGGAAAGAACAAGTGCTTAGTTG 421 422 AATAAGCTGGCACCTTTGACGGTACCTAACCAGAAAGCCACGGCTAACTACGTGCCAGCA 481 482 GCCGCGGTAATACGTAGGTGGCAAGCGTTATCCGGAATTATTGGGCGTAAAGCGCGCGCA 541 542 GGTGGTTTCTTAAGTCTGATGTGAAAGCCCACGGCTCAACCGTGGAGGGTCATTGGAAAC 601 602 TGGGGAGACTTGAGTGCAGAAGAGGAAAGTGGAATTCCATGTGTAGCGGTGAAATGCGTA 661 662 GAGATATGGAGGAACACCAGTGGCGAAGGCGACTTTCTGGTCTGTAACTGACACTGAGGC 721 722 GCGAAAGCGTGGGGAGCAAACAGGATTAGATACCCTGGTAGTCCACGCCGTAAACGATGA 781 782 GTGCTAAGTGTTAGAGGGTTTCCGCCCTTTAGTGCTGAAGTTAACGCATTAAGCACTCCG 841 813 GTGCTAAGTGTTAGAGGGTTTCCGCCCTTTAGTGCTGAAGTTAACGCATTAAGCACTCCG 872 842 CCTGGGGAGTACGGCCGCAAGGCTGAAACTCAAAGGAATTGACGGGGGCCCGCACAAGCG 901 902 GTGGAGCATGTGGTTTAATTCGAAGCAACGCGAAGAACCTTACCAGGTCTTGACATCCTC 961 962 TGAAAACCCTAGAGATAGGGCTTCTCCTTCGGGAGCAGAGTGACAGGTGGTGCATGGTTG 1021 1022 TCGTCAGCTCGTGTCGTGAGATGTTGGGTTAAGTCCCGCAACGAGCGCAACCCTTGATCT 1081 1082 TAGTTGCCATCATTAAGTTGGGCACTCTAAGGTGACTGCCGGTGACAAACCGGAGGAAGG 1141 1142 TGGGGATGACGTCAAATCATCATGCCCCTTATGACCTGGGCTACACACGTGCTACAATGG 1201 1202 ACGGTACAAAGAGCTGCAAGACCGCGAGGTGGAGCTAATCTCATAAAACCGTTCTCAGTT 1261 1262 CGGATTGTAGGCTGCAACTCGCCTACATGAAGCTGGAATCGCTAGTAATCGCGGATCAGCA 1322
H. Analisis sekuen gen 16S rRNA isolat INT1c dengan program BLAST-N
> gb|JF837192.1| Bacillus cereus strain GM3 16S ribosomal RNA gene, partial
sequence Length=1363
Score = 2324 bits (1258), Expect = 0.0
Identities = 1303/1321 (99%), Gaps = 18/1321 (1%) Strand=Plus/Plus
Query 4 GAAT-GA-TAAGAGCTTGCTCTTATGAAGTTAGCGGCGGACGGGTGAGTAACACGTGGGT 61 |||| || |||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 49 GAATGGATTAAGAGCTTGCTCTTATGAAGTTAGCGGCGGACGGGTGAGTAACACGTGGGT 108
Query 62 AACCTGCCCATAAGACTGGGATAACTCCGGGAAACCGGGGCTAATACCGGATAACATTTT 121 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 109 AACCTGCCCATAAGACTGGGATAACTCCGGGAAACCGGGGCTAATACCGGATAACATTTT 168
Query 122 GAACCGCATGGTTCGAAATTGAAAGGCGGCTTCGGCTGTCACTTATGGATGGACCCGCGT 181 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 169 GAACCGCATGGTTCGAAATTGAAAGGCGGCTTCGGCTGTCACTTATGGATGGACCCGCGT 228
Query 182 CGCATTAGCTAGTTGGTGAGGTAACGGCTCACCAAGGCAACGATGCGTAGCCGACCTGAG 241 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 229 CGCATTAGCTAGTTGGTGAGGTAACGGCTCACCAAGGCAACGATGCGTAGCCGACCTGAG 288
Sbjct 289 AGGGTGATCGGCCACACTGGGACTGAGACACGGCCCAGACT-CCTAC-GGG-AGGCAG-C 344
Query 302 AGTAGGGAATCTTCCGCAATGGAACGATAAGTTCTGACGGAGCAACCGCCGGCGTGAGTG 361 |||||||||||||||||||||| |||| ||| ||||||||||||| |||| ||||||||| Sbjct 345 AGTAGGGAATCTTCCGCAATGG-ACGA-AAG-TCTGACGGAGCAA-CGCC-GCGTGAGTG 399
Query 362 ATGAAGGCCTTTCGGGTTCGTAAAACTCTGTTGTTTAGGGAAAGAACAAGTGCTTAGTTG 421 ||||||| |||||||| |||||||||||||||| |||||| |||||||||||| |||||| Sbjct 400 ATGAAGG-CTTTCGGG-TCGTAAAACTCTGTTG-TTAGGG-AAGAACAAGTGC-TAGTTG 454
Query 422 AATAAGCTGGCACCTTTGACGGTACCTAACCAGAAAGCCACGGCTAACTACGTGCCAGCA 481 |||||||||||||| ||||||||||||||||||||||||||||||||||||||||||||| Sbjct 455 AATAAGCTGGCACC-TTGACGGTACCTAACCAGAAAGCCACGGCTAACTACGTGCCAGCA 513
Query 482 GCCGCGGTAATACGTAGGTGGCAAGCGTTATCCGGAATTATTGGGCGTAAAGCGCGCGCA 541 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 514 GCCGCGGTAATACGTAGGTGGCAAGCGTTATCCGGAATTATTGGGCGTAAAGCGCGCGCA 573
Query 542 GGTGGTTTCTTAAGTCTGATGTGAAAGCCCACGGCTCAACCGTGGAGGGTCATTGGAAAC 601 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 574 GGTGGTTTCTTAAGTCTGATGTGAAAGCCCACGGCTCAACCGTGGAGGGTCATTGGAAAC 633
Query 602 TGGGGAGACTTGAGTGCAGAAGAGGAAAGTGGAATTCCATGTGTAGCGGTGAAATGCGTA 661 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 634 T-GGGAGACTTGAGTGCAGAAGAGGAAAGTGGAATTCCATGTGTAGCGGTGAAATGCGTA 692
Query 662 GAGATATGGAGGAACACCAGTGGCGAAGGCGACTTTCTGGTCTGTAACTGACACTGAGGC 721 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 693 GAGATATGGAGGAACACCAGTGGCGAAGGCGACTTTCTGGTCTGTAACTGACACTGAGGC 752
Query 722 GCGAAAGCGTGGGGAGCAAACAGGATTAGATACCCTGGTAGTCCACGCCGTAAACGATGA 781 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 753 GCGAAAGCGTGGGGAGCAAACAGGATTAGATACCCTGGTAGTCCACGCCGTAAACGATGA 812
Query 782 GTGCTAAGTGTTAGAGGGTTTCCGCCCTTTAGTGCTGAAGTTAACGCATTAAGCACTCCG 841 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 813 GTGCTAAGTGTTAGAGGGTTTCCGCCCTTTAGTGCTGAAGTTAACGCATTAAGCACTCCG 872
Query 842 CCTGGGGAGTACGGCCGCAAGGCTGAAACTCAAAGGAATTGACGGGGGCCCGCACAAGCG 901 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 873 CCTGGGGAGTACGGCCGCAAGGCTGAAACTCAAAGGAATTGACGGGGGCCCGCACAAGCG 932
Query 902 GTGGAGCATGTGGTTTAATTCGAAGCAACGCGAAGAACCTTACCAGGTCTTGACATCCTC 961 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 933 GTGGAGCATGTGGTTTAATTCGAAGCAACGCGAAGAACCTTACCAGGTCTTGACATCCTC 992
Query 962 TGAAAACCCTAGAGATAGGGCTTCTCCTTCGGGAGCAGAGTGACAGGTGGTGCATGGTTG 1021 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 993 TGAAAACCCTAGAGATAGGGCTTCTCCTTCGGGAGCAGAGTGACAGGTGGTGCATGGTTG 1052
Query 1022 TCGTCAGCTCGTGTCGTGAGATGTTGGGTTAAGTCCCGCAACGAGCGCAACCCTTGATCT 1081 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 1053 TCGTCAGCTCGTGTCGTGAGATGTTGGGTTAAGTCCCGCAACGAGCGCAACCCTTGATCT 1112
Query 1082 TAGTTGCCATCATTAAGTTGGGCACTCTAAGGTGACTGCCGGTGACAAACCGGAGGAAGG 1141 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 1113 TAGTTGCCATCATTAAGTTGGGCACTCTAAGGTGACTGCCGGTGACAAACCGGAGGAAGG 1172
Query 1142 TGGGGATGACGTCAAATCATCATGCCCCTTATGACCTGGGCTACACACGTGCTACAATGG 1201 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 1173 TGGGGATGACGTCAAATCATCATGCCCCTTATGACCTGGGCTACACACGTGCTACAATGG 1232
Query 1202 ACGGTACAAAGAGCTGCAAGACCGCGAGGTGGAGCTAATCTCATAAAACCGTTCTCAGTT 1261 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 1233 ACGGTACAAAGAGCTGCAAGACCGCGAGGTGGAGCTAATCTCATAAAACCGTTCTCAGTT 1292
Query 1262 CGGATTGTAGGCTGCAACTCGCCTACATGAAGCTGGAATCGCTAGTAATCGCGGATCAGC 1321 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 1293 CGGATTGTAGGCTGCAACTCGCCTACATGAAGCTGGAATCGCTAGTAATCGCGGATCAGC 1352
I. Urutan nukleotida gen 16S rRNA isolat SGT3g
1 GGGGGGGAAGGTTAAGAGCTTGCTCTTATGAAGTTAGCGGCGGACGGGTGAGTAACACG 71 72 TGGGTAACCTGCCCATAAGACTGGGATAACTCCGGGAAACCGGGGCTAATACCGGATAAC 131 132 ATTTTGAACCGCATGGTTCGAAATTGAAAGGCGGCTTCGGCTGTCACTTATGGATGGACC 191 192 CGCGTCGCATTAGCTAGTTGGTGAGGTAACGGCTCACCAAGGCAACGATGCGTAGCCGAC 251 252 CTGAGAGGGTGATCGGCCACACTGGGACTGAGACACGGCCCAGACTCCTACGGGAGGCAG 311 312 CAGTAGGGAATCTTCCGCAATGGACGAAAGTCTGACGGAGCAACGCCGCGTGAGTGATGA 371 372 AGGCTTTCGGGTCGTAAAACTCTGTTGTTAGGGAAGAACAAGTGCTAGTTGAATAAGCTG 431 432 GCACCTTGACGGTACCTAACCAGAAAGCCACGGCTAACTACGTGCCAGCAGCCGCGGTAA 491 492 TACGTAGGTGGCAAGCGTTATCCGGAATTATTGGGCGTAAAGCGCGCGCAGGTGGTTTCT 551 552 GAAAGCCCACGGCTCAACCGTGGAGGGTCATTGGAAACTGGGAGACTTGAGTGCAGAAGA 611 612 GGAAAGTGGAATTCCATGTGTAGCGGTGAAATGCGTAGAGATATGGAGGAACACCAGTGG 671 672 TAAGTCTGATGTCGAAGGCGACTTTCTGGTCTGTAACTGACACTGAGGCGCGAAAGCGTG 731 732 GGGAGCAAACAGGATTAGATACCCTGGTAGTCCACGCCGTAAACGATGAGTGCTAAGTGT 791 792 TAGAGGGTTTCCGCCCTTTAGTGCTGAAGTTAACGCATTAAGCACTCCGCCTGGGGAGTA 851 852 CGGCCGCAAGGCTGAAACTCAAAGGAATTGACGGGGGCCCGCACAAGCGGTGGAGCATGT 911 912 GGTTTAATTCGAAGCAACGCGAAGAACCTTACCAGGTCTTGACATCCTCTGAAAACCCTA 971 972 GAGATAGGGCTTCTCCTTCGGGAGCAGAGTGACAGGTGGTGCATGGTTGTCGTCAGCTCG 1031 1032 TGTCGTGAGATGTTGGGTTAAGTCCCGCAACGAGCGCAACCCTTGATCTTAGTTGCCATC 1091 1040 ATTAAGTTGGGCTTGGGTTAAGTCCCGCAACGAGCGCAACCCTTGATCTTAGTTGCCATC 1099 1092 ATTAAGTTGGGCACTCTAAGGTGACTGCCGGTGACAAACCGGAAGAAGGTGGGGATGACG 1151 1152 TCAAATCATCATGCCCCTTAAGACCTGGGCTACCCACGTGCTACAATGGACGGTTCAAAA 1211 1212 AGCTGCAAGACCGCGAGGTGGAGCTAATCTCATAAAACCGTTCTCAGTTCGGATTGTAGG 1269 1270 CTGCAACTCGCCTACATGAAGCTGGAATCGCTAGTAATCGCGNTCAGCAGCCGC 1313
J. Analisis sekuen gen 16S rRNA isolat SGT3g dengan program BLAST-N
> gb|JF496321.1| Bacillus thuringiensis strain A1-1 16S ribosomal RNA gene,
partial sequence Length=1411
Score = 2364 bits (1280), Expect = 0.0
Identities = 1296/1304 (99%), Gaps = 2/1304 (0%) Strand=Plus/Plus
Query 12 TTAAGAGCTTGCTCTTATGAAGTTAGCGGCGGACGGGTGAGTAACACGTGGGTAACCTGC 71 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 20 TTAAGAGCTTGCTCTTATGAAGTTAGCGGCGGACGGGTGAGTAACACGTGGGTAACCTGC 79
Query 72 CCATAAGACTGGGATAACTCCGGGAAACCGGGGCTAATACCGGATAACATTTTGAACCGC 131 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 80 CCATAAGACTGGGATAACTCCGGGAAACCGGGGCTAATACCGGATAACATTTTGAACCGC 139
Query 132 ATGGTTCGAAATTGAAAGGCGGCTTCGGCTGTCACTTATGGATGGACCCGCGTCGCATTA 191 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 140 ATGGTTCGAAATTGAAAGGCGGCTTCGGCTGTCACTTATGGATGGACCCGCGTCGCATTA 199
Query 192 GCTAGTTGGTGAGGTAACGGCTCACCAAGGCAACGATGCGTAGCCGACCTGAGAGGGTGA 251 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 200 GCTAGTTGGTGAGGTAACGGCTCACCAAGGCAACGATGCGTAGCCGACCTGAGAGGGTGA 259
Query 252 TCGGCCACACTGGGACTGAGACACGGCCCAGACTCCTACGGGAGGCAGCAGTAGGGAATC 311 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 260 TCGGCCACACTGGGACTGAGACACGGCCCAGACTCCTACGGGAGGCAGCAGTAGGGAATC 319
Query 312 TTCCGCAATGGACGAAAGTCTGACGGAGCAACGCCGCGTGAGTGATGAAGGCTTTCGGGT 371 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 320 TTCCGCAATGGACGAAAGTCTGACGGAGCAACGCCGCGTGAGTGATGAAGGCTTTCGGGT 379
Query 372 CGTAAAACTCTGTTGTTAGGGAAGAACAAGTGCTAGTTGAATAAGCTGGCACCTTGACGG 431 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 380 CGTAAAACTCTGTTGTTAGGGAAGAACAAGTGCTAGTTGAATAAGCTGGCACCTTGACGG 439
Query 432 TACCTAACCAGAAAGCCACGGCTAACTACGTGCCAGCAGCCGCGGTAATACGTAGGTGGC 491 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 440 TACCTAACCAGAAAGCCACGGCTAACTACGTGCCAGCAGCCGCGGTAATACGTAGGTGGC 499
Query 552 GAAAGCCCACGGCTCAACCGTGGAGGGTCATTGGAAACTGGGAGACTTGAGTGCAGAAGA 611 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 560 GAAAGCCCACGGCTCAACCGTGGAGGGTCATTGGAAACTGGGAGACTTGAGTGCAGAAGA 619
Query 612 GGAAAGTGGAATTCCATGTGTAGCGGTGAAATGCGTAGAGATATGGAGGAACACCAGTGG 671 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 620 GGAAAGTGGAATTCCATGTGTAGCGGTGAAATGCGTAGAGATATGGAGGAACACCAGTGG 679
Query 672 CGAAGGCGACTTTCTGGTCTGTAACTGACACTGAGGCGCGAAAGCGTGGGGAGCAAACAG 731 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 680 CGAAGGCGACTTTCTGGTCTGTAACTGACACTGAGGCGCGAAAGCGTGGGGAGCAAACAG 739
Query 732 GATTAGATACCCTGGTAGTCCACGCCGTAAACGATGAGTGCTAAGTGTTAGAGGGTTTCC 791 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 740 GATTAGATACCCTGGTAGTCCACGCCGTAAACGATGAGTGCTAAGTGTTAGAGGGTTTCC 799
Query 792 GCCCTTTAGTGCTGAAGTTAACGCATTAAGCACTCCGCCTGGGGAGTACGGCCGCAAGGC 851 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 800 GCCCTTTAGTGCTGAAGTTAACGCATTAAGCACTCCGCCTGGGGAGTACGGCCGCAAGGC 859
Query 852 TGAAACTCAAAGGAATTGACGGGGGCCCGCACAAGCGGTGGAGCATGTGGTTTAATTCGA 911 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 860 TGAAACTCAAAGGAATTGACGGGGGCCCGCACAAGCGGTGGAGCATGTGGTTTAATTCGA 919
Query 912 AGCAACGCGAAGAACCTTACCAGGTCTTGACATCCTCTGAAAACCCTAGAGATAGGGCTT 971 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 920 AGCAACGCGAAGAACCTTACCAGGTCTTGACATCCTCTGAAAACCCTAGAGATAGGGCTT 979
Query 972 CTCCTTCGGGAGCAGAGTGACAGGTGGTGCATGGTTGTCGTCAGCTCGTGTCGTGAGATG 1031 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 980 CTCCTTCGGGAGCAGAGTGACAGGTGGTGCATGGTTGTCGTCAGCTCGTGTCGTGAGATG 1039
Query 1032 TTGGGTTAAGTCCCGCAACGAGCGCAACCCTTGATCTTAGTTGCCATCATTAAGTTGGGC 1091 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 1040 TTGGGTTAAGTCCCGCAACGAGCGCAACCCTTGATCTTAGTTGCCATCATTAAGTTGGGC 1099
Query 1092 ACTCTAAGGTGACTGCCGGTGACAAACCGGAAGAAGGTGGGGATGACGTCAAATCATCAT 1151 ||||||||||||||||||||||||||||||| |||||||||||||||||||||||||||| Sbjct 1100 ACTCTAAGGTGACTGCCGGTGACAAACCGGAGGAAGGTGGGGATGACGTCAAATCATCAT 1159
Query 1152 GCCCCTTAAGACCTGGGCTACCCACGTGCTACAATGGACGGTTCAAAAAGCTGCAAGACC 1211 |||||||| |||||||||||| |||||||||||||||||||| |||| |||||||||||| Sbjct 1160 GCCCCTTATGACCTGGGCTACACACGTGCTACAATGGACGGTACAAAGAGCTGCAAGACC 1219
Query 1212 GCGAGGTGGAGCTAATCTCATAAAACCGTTCTCAGTTCGGATTGTAGGCTGCAACTCGCC 1271 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 1220 GCGAGGTGGAGCTAATCTCATAAAACCGTTCTCAGTTCGGATTGTAGGCTGCAACTCGCC 1279
TIRA SITI NUR AFIAH. Isolasi dan Karakterisasi Bateri Penghasil Acyl Homoserine Lactonase (AHL-laktonase) Asal Lahan Pertanian di Jawa. Dibimbing oleh IMAN RUSMANA dan ALINA AKHDIYA.
Quorum sensing (QS) adalah mekanisme komunikasi bakteri yang bergantung pada jumlah populasi bakteri tersebut dan akumulasi senyawa autoinducer (AI). Quorum sensing pada kelompok bakteri Gram negatif melibatkan senyawa acyl homoserine lactone (AHL) untuk mengatur beberapa ekspresi gen, termasuk gen virulensi. Senyawa anti-QS merupakan senyawa yang dapat mendegradasi AHL sehingga gen virulensi tidak dapat diekspresikan. Acyl homoserine lactonase (AHL-laktonase) yang disandikan oleh gen aiiA merupakan salah satu enzim yang dapat berperan sebagai anti-QS. Penelitian ini bertujuan untuk mengisolasi dan mengkarakterisasi bakteri penghasil AHL-laktonase yang berasal dari lahan pertanian di Jawa. Bioesei aktivitas degradasi AHL menggunakan bakteri indikator Chromobacterium violaceum. Verifikasi keberadaan AHL-laktonase pada isolat-isolat yang menunjukkan hasil bioesei positif dilakukan dengan amplifikasi gen aiiA. Dari 12 isolat bakteri yang menunjukkan aktivitas degradasi AHL, terdapat dua isolat yang memiliki gen aiiA yaitu isolat INT1c dan SGT3g. Kedua isolat tersebut memiliki bentuk sel batang, penataan sel berantai, berendospora, dan termasuk dalam kelompok bakteri Gram positif. Analisis sekuen gen aiiA menunjukkan bahwa gen aiiA isolat INT1c memiliki homologi dengan gen aiiA bakteri Bacillus cereus ATTCC 14579 sedangkan isolat SGT3g memiliki homologi dengan gen aiiA bakteri Bacillus thuringiensis BMB171. Identifikasi molekuler berdasarkan gen 16S rRNA menunjukkan bahwa isolat INT1c memiliki kemiripan terdekat dengan bakteri Bacillus cereus strain GM3 (98%) sedangkan isolat SGT3g memiliki kemiripan terdekat dengan bakteri Bacillus thuringiensis strain A1-1 (99%).
Kata kunci: anti-quorum sensing, AHL-laktonase, gen aiiA.
TIRA SITI NUR AFIAH. Isolation and Characterization of Acyl Homoserine Lactonase (AHL-lactonase) Producing Bacteria from Agricultural Field in Java. Under supervision of IMAN RUSMANA and ALINA AKHDIYA.
Quorum sensing (QS) is a bacterial communication mechanism correlating with the number of bacterial population and accumulation of autoinducer (AI). Quorum sensing in Gram-negative bacteria involves a AI group of acyl homoserine lactone (AHL) to regulate the expression of several genes, including virulence genes. Anti-quorum sensing compounds can degrade AHL so that virulence genes can not be expressed. Acyl homoserine lactonase (AHL-lactonase) encoded by the aiiA gene is one enzyme that can act as an anti-quorum sensing. This study aimed to isolate and characterize the AHL-lactonase producing bacteria isolated from agricultural field in Java. Activity of AHL degradation bioassay using Chromobacterium violaceum as a bacterial indicator. Verification the presence of AHL-laktonase was performed by gene aiiA amplification. As many as 12 bacterial isolates showed AHL degradation activity and two isolates had the aiiA gene i.e. INT1c and SGT3g isolates. These isolates had bacil cells with chain arrangement and endospores. They were grouped in Gram-positive bacteria. AiiA gene sequence analysis showed that aiiA gene of INT1c had homology with aiiA gene of Bacillus cereus ATTCC 14579 while the SGT3g isolate had homology with aiiA gene of Bacillus thuringiensis BMB171. Molecular identification based on 16S rRNA gene showed that INT1c isolate had the closest similarity with the Bacillus cereus strain GM3 (98%) while SGT3g isolate had the closest similarity with the Bacillus thuringiensis strains A1-1 (99%).
Bakteri fitopatogen merupakan bakteri yang menyebabkan infeksi atau penyakit pada tumbuhan. Pengendalian bakteri patogen dapat dilakukan dengan menggunakan senyawa antibiotik atau antimikrob. Masalah yang dihadapi dalam penggunaan antibiotik adalah timbulnya resistensi bakteri terhadap antibiotik tersebut. Hal ini mendorong dilakukannya berbagai penelitian tentang pengendalian bakteri patogen yang yang lebih aman dan tidak dikhawatirkan akan menimbulkan resistensi.
Defoirdt et al. (2004) menyatakan bahwa ekspresi gen virulensi pada beberapa bakteri patogen diregulasi oleh proses quorum sensing. Quorum sensing (QS) adalah mekanisme komunikasi bakteri sebagai suatu pengaturan ekspresi gen yang bergantung pada jumlah populasi bakteri tersebut dan akumulasi senyawa autoinducer (AI) (Rukayadi & Hwang 2009). QS terlibat dalam regulasi fungsi biologi yang penting seperti luminescence, produksi antibiotik, transfer plasmid, motilitas, virulensi, pembentukan biofilm dan berperan dalam regulasi ekspresi gen-gen patogen pada Pseudomonas solanacearum, Pseudomonas aeruginosa, dan beberapa spesies Erwinia (Dong et al. 2000; Zang & Dong 2004).
Proses QS melibatkan molekul kimia yang disebut autoinducer (AI). AI merupakan molekul sinyal yang disekresikan, diakumulasi, diserap kembali, dan dikenali oleh bakteri selama proses QS terjadi (Rukayadi & Hwang 2009). Salah satu jenis molekul AI adalah N-acylhomoserine lactone (AHL) yang dihasilkan oleh bakteri Gram negatif, serta digunakan untuk komunikasi dalam spesies yang sama (Rukayadi & Hwang 2009).
Salah satu prinsip pengendalian virulensi bakteri fitopatogen dengan dasar QS adalah mencegah akumulasi AHL dengan mendegradasi molekul sinyal tersebut. Senyawa atau molekul yang dapat mendegradasi molekul AHL disebut senyawa anti-QS. Anti-QS memiliki kelebihan dalam mengontrol infeksi bakteri yaitu tidak mempengaruhi pertumbuhan, sehingga dapat menghindarkan timbulnya tekanan seleksi yang sering menghasilkan generasi patogen yang lebih resisten terhadap antibiotik (White & Finan 2009). Beberapa enzim pendegradasi AHL telah diidentifikasi dari beberapa
adalah enzim Acyl Homoserine Lactonase (AHL-laktonase) (Dong et al. 2000). Enzim ini disandikan oleh gen aiiA yang dimiliki oleh Bacillus dan mampu mendegradasi AHL dengan menghidrolisis ikatan lakton yang terdapat pada molekul AHL (Dong et al. 2000). AHL-laktonase sebagai salah satu senyawa anti-QS dapat digunakan sebagai alternatif dalam pengendalian bakteri fitopatogen.
Sebagai salah satu negara megabiodiversity, Indonesia mempunyai kekayaan dan keragaman sumberdaya mikroba yang sangat besar dan berpotensi sebagai penghasil AHL-laktonase. Oleh karena itu, isolasi dan karakterisasi bakteri penghasil AHL-laktonase merupakan langkah awal untuk untuk memanfaatkan potensi tersebut.
Tujuan
Penelitian ini bertujuan untuk mengisolasi dan mengkarakterisasi bakteri penghasil AHL-laktonase asal lahan pertanian di Jawa.
METODE
Waktu dan Tempat
Penelitian ini dilaksanakan mulai bulan November 2010 sampai dengan Mei 2011, bertempat di Laboratorium Mikrobiologi Departemen Biologi FMIPA IPB.
Isolasi, Pemurnian, dan Penyimpanan Bakteri
Bakteri fitopatogen merupakan bakteri yang menyebabkan infeksi atau penyakit pada tumbuhan. Pengendalian bakteri patogen dapat dilakukan dengan menggunakan senyawa antibiotik atau antimikrob. Masalah yang dihadapi dalam penggunaan antibiotik adalah timbulnya resistensi bakteri terhadap antibiotik tersebut. Hal ini mendorong dilakukannya berbagai penelitian tentang pengendalian bakteri patogen yang yang lebih aman dan tidak dikhawatirkan akan menimbulkan resistensi.
Defoirdt et al. (2004) menyatakan bahwa ekspresi gen virulensi pada beberapa bakteri patogen diregulasi oleh proses quorum sensing. Quorum sensing (QS) adalah mekanisme komunikasi bakteri sebagai suatu pengaturan ekspresi gen yang bergantung pada jumlah populasi bakteri tersebut dan akumulasi senyawa autoinducer (AI) (Rukayadi & Hwang 2009). QS terlibat dalam regulasi fungsi biologi yang penting seperti luminescence, produksi antibiotik, transfer plasmid, motilitas, virulensi, pembentukan biofilm dan berperan dalam regulasi ekspresi gen-gen patogen pada Pseudomonas solanacearum, Pseudomonas aeruginosa, dan beberapa spesies Erwinia (Dong et al. 2000; Zang & Dong 2004).
Proses QS melibatkan molekul kimia yang disebut autoinducer (AI). AI merupakan molekul sinyal yang disekresikan, diakumulasi, diserap kembali, dan dikenali oleh bakteri selama proses QS terjadi (Rukayadi & Hwang 2009). Salah satu jenis molekul AI adalah N-acylhomoserine lactone (AHL) yang dihasilkan oleh bakteri Gram negatif, serta digunakan untuk komunikasi dalam spesies yang sama (Rukayadi & Hwang 2009).
Salah satu prinsip pengendalian virulensi bakteri fitopatogen dengan dasar QS adalah mencegah akumulasi AHL dengan mendegradasi molekul sinyal tersebut. Senyawa atau molekul yang dapat mendegradasi molekul AHL disebut senyawa anti-QS. Anti-QS memiliki kelebihan dalam mengontrol infeksi bakteri yaitu tidak mempengaruhi pertumbuhan, sehingga dapat menghindarkan timbulnya tekanan seleksi yang sering menghasilkan generasi patogen yang lebih resisten terhadap antibiotik (White & Finan 2009). Beberapa enzim pendegradasi AHL telah diidentifikasi dari beberapa
adalah enzim Acyl Homoserine Lactonase (AHL-laktonase) (Dong et al. 2000). Enzim ini disandikan oleh gen aiiA yang dimiliki oleh Bacillus dan mampu mendegradasi AHL dengan menghidrolisis ikatan lakton yang terdapat pada molekul AHL (Dong et al. 2000). AHL-laktonase sebagai salah satu senyawa anti-QS dapat digunakan sebagai alternatif dalam pengendalian bakteri fitopatogen.
Sebagai salah satu negara megabiodiversity, Indonesia mempunyai kekayaan dan keragaman sumberdaya mikroba yang sangat besar dan berpotensi sebagai penghasil AHL-laktonase. Oleh karena itu, isolasi dan karakterisasi bakteri penghasil AHL-laktonase merupakan langkah awal untuk untuk memanfaatkan potensi tersebut.
Tujuan
Penelitian ini bertujuan untuk mengisolasi dan mengkarakterisasi bakteri penghasil AHL-laktonase asal lahan pertanian di Jawa.
METODE
Waktu dan Tempat
Penelitian ini dilaksanakan mulai bulan November 2010 sampai dengan Mei 2011, bertempat di Laboratorium Mikrobiologi Departemen Biologi FMIPA IPB.
Isolasi, Pemurnian, dan Penyimpanan Bakteri
menunjukkan morfologi yang berbeda dimurnikan dengan metode cawan gores. Setelah murni, bakteri disimpan dalam agar miring.
Bioesei Aktivitas Degradasi AHL
Isolat-isolat murni ditumbuhkan pada media Luria broth (LB). Setelah 48 jam, kultur disentrifus selama 10 menit dengan kecepatan 10000 rpm. Sebanyak 100 μL supernatant diteteskan pada paper disk dan diletakkan pada permukaan media cawan Luria agar (LA) yang mengandung Chromobacterium violaceum 1%. Cawan tersebut kemudian diinkubasi pada suhu ruang selama 24 jam. Media LB steril digunakan sebagai kontrol. Paper disk yang dikelilingi oleh zona tidak berwarna ungu menunjukkan adanya aktivitas degradasi AHL.
Pengamatan Morfologi Sel dan Pewarnaan Gram
Isolat-isolat yang menunjukkan hasil bioesei positif diamati morfologi sel (bentuk sel, penataan sel, dan endospora) dan reaksi terhadap pewarnaan Gram.
Amplifikasi Gen Penyandi AHL-laktonase DNA bakteri diisolasi dengan metode Lazo (Lazo et al. 1987). Isolat-isolat yang menunjukkan hasil bioesei degradasi AHL positif diamplifikasi menggunakan primer spesifik untuk gen aiiA yaitu aiiAF (5’-ATC GGA TCC ATG ACA GTA AAG AAG CTT ATT TCG-3’, dan aiiAR (5’-GTC GAA TTC CTC AAC AAG ATA CTC CTA ATG ATGT-3’ (Dong et al. 2000). Campuran PCR terdiri dari 1.5 mM MgCl2, 0.2 mM dNTPs, 0.3 µM masing-masing primer, 0.02 U/µL Taq DNA polymerase, template DNA, ddH2O, dan buffer DNA polymerase. Amplifikasi dilakukan selama 30 siklus PCR dengan kondisi pra-denaturasi (94oC, 10 menit), denaturasi (94oC, 30 detik), annealing (52oC, 30 detik), elongasi (72oC, 1 menit), dan post-PCR (72oC, 5 menit) (Chan et al. 2007). Produk PCR dilarikan pada mini-gel elektroforesis (agarose 1%) pada tegangan listrik 50 volt selama 45 menit (Sambrook et al. 1989). Visualisasi di atas UV Transiluminator dilakukan dengan pewarnaan Ethidium Bromida.
Identifikasi Isolat
Isolat-isolat yang menunjukkan hasil positif pada bioesei degradasi AHL dan amplifikasi gen aiiA diidentifikasi secara molekuler (16S rRNA). Amplifikasi gen 16S rRNA menggunakan primer 63f (5’-CAG GCC TAA CAC ATG CAA GTC-3’) dan 1387r (5’-GGG CGG WGT GTA CAA GGC-3’) (Marchesi et al. 1998). Amplifikasi dilakukan selama 30 siklus PCR dengan kondisi pra-PCR (94oC, 2 menit), denaturasi (92oC, 30 detik), annealing (50oC, 1 menit), elongasi (72oC, 1 menit), dan post-PCR (75oC, 5 menit).
Sekuensing DNA dan Analisis Bioinformatika
Pengurutan DNA hasil amplifikasi dilakukan dengan memanfaatkan jasa perusahaan sekuensing 1st Base di Singapura. Sekuen yang diperoleh dijajarkan dengan data pada GenBank menggunakan program Basic Local Alignment Search Tool-Nucleotida (BLAST-N) dan Basic Local Alignment Search Tool Nucleotide 6-frame translation-protein (BLAST-X) dari situs National Center for Biotechnology Information (NCBI) melalui http://www.ncbi.nlm.nih.gov untuk mengetahui tingkat kemiripan gen aiiA dan 16S rRNA dari isolat yang dianalisa.
Konstruksi Pohon Filogenetik
Konstruksi pohon filogenetik dilakukan dengan metode Neighbor Joining (NJ) dengan bootstrap 1000x menggunakan program MEGA 5.0.
HASIL
Bioesei Aktivitas Degradasi AHL