IDENTIFIKASI KERAGAMAN GEN
ATP BINDING
CASSETTE
(ABCG2|
PstI
) PADA SAPI
FRIESIAN
HOLSTEIN
DENGAN METODE PCR-RFLP
WINDA TRISTIA NOVITASARI
DEPARTEMEN ILMU PRODUKSI DAN TEKNOLOGI PETERNAKAN FAKULTAS PETERNAKAN
INSTITUT PERTANIAN BOGOR BOGOR
iii
PERNYATAAN MENGENAI SKRIPSI DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa skripsi berjudul Identifikasi Keragaman Gen ATP Binding Cassette (ABCG2|PstI) pada Sapi Friesian Holstein dengan Metode PCR-RFLP adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir skripsi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, September 2013
Winda Tristia Novitasari
1
ABSTRAK
WINDA TRISTIA NOVITASARI. Identifikasi Keragaman Gen ATP Binding
Cassette (ABCG2|PstI) pada Sapi Friesian Holstein dengan Metode PCR-RFLP.
Dibimbing oleh CECE SUMANTRI dan ANNEKE ANGGRAENI.
Gen ATP Binding Cassette (ABCG2) berpengaruh pada produksi dan
komposisi susu. Reseptor yang disandi gen ABCG2 pada sel epitel alveoli kelenjar susu akan mengikat substrat spesifik seperti riboflavin, yang disekresikan secara aktif, sehingga susu kaya kandungan riboflavin. Penelitian ini bertujuan untuk mengidentifikasi keragaman genetik gen ABCG2 pada sapi Friesian
Holstein (FH) dengan metode Polymerase Chain Reaction-Restriction Fragment
Length Polymorphism (PCR-RFLP). Penelitian ini menggunakan sapi FH
sebanyak 224 ekor dari tujuh lokasi (BBIB Singasari, BIB Lembang, BPPTU Baturaden, BET Cipelang, BPPT-SP Cikole, KPSBU Pasir Kemis, dan Cilumber). Data dianalisis frekuensi alel, frekuensi genotipe, dan nilai PIC. Hasil genotyping
menghasilkan dua alel, yaitu alel A (292 pb) dan alel C (268 pb, 24 pb); serta tiga genotipe (AA, AC, dan CC). Secara keseluruhan, frekuensi genotipe homozigot AA ditemukan paling tinggi (0.964) vs heterozigot AC (0.031) dan homozigot CC (0.004); dengan frekuensi alel (A = 0.98; C = 0.02). Sapi FH pengamatan memiliki nilai PIC berkisar 0.000-0.189. Disimpulkan bahwa gen ABCG2|PstI
pada sapi FH pengamatan bersifat polimorfik, kecuali pada BBIB Singasari dan BET Cipelang bersifat monomorfik.
Kata kunci: Friesian Holstein, gen ABCG2, keragaman genetik, PCR-RFLP
ABSTRACT
WINDA TRISTIA NOVITASARI. Polymorphism Identification of the ATP
Binding Cassette (ABCG2|PstI) Gene in Holstein Friesian Cattle by PCR-RFLP
Method. Supervised by CECE SUMANTRI and ANNEKE ANGGRAENI.
ATP Binding Cassette (ABCG2) has effects on milk production and
composition. Reseptor which is encoded by the ABCG2 gene on alveoli epitel of mammary gland will bind its specific substrates, such as riboflavin, that are actively secreted, so milk will be rich of riboflavin. This study was aimed to identify the ABCG2 gene polymorphism in Holstein Friesian (HF) by Polymerase
Chain Reaction-Restriction Fragment Length Polymorphism (PCR-RFLP)
tested populations, the exception of monomorphic was in BBIB Singasari and BET Cipelang.
4
Skripsi
sebagai salah satu syarat untuk memperoleh gelar Sarjana Peternakan
pada
Departemen Ilmu Produksi dan Teknologi Peternakan
DEPARTEMEN ILMU PRODUKSI DAN TEKNOLOGI PETERNAKAN FAKULTAS PETERNAKAN
INSTITUT PERTANIAN BOGOR BOGOR
2013
WINDA TRISTIA NOVITASARI
IDENTIFIKASI KERAGAMAN GEN
ATP BINDING
Judul Skripsi : Identifikasi Keragaman Gen ATP Binding Cassette (ABCG2|PstI) pada Sapi Friesian Holstein dengan Metode PCR-RFLP
Nama : Winda Tristia Novitasari NIM : D14090011
Disetujui oleh
Prof Dr Ir Cece Sumantri, MAgrSc Pembimbing I
Ir Anneke Anggraeni, MSi PhD Pembimbing II
Diketahui oleh
Prof Dr Ir Cece Sumantri, MAgrSc Ketua Departemen
PRAKATA
Bismillahirrahmanirrahim. Puji dan syukur penulis panjatkan kepada Allah
Subhanahu Wa Ta’ala atas segala karunia-Nya, sehingga karya ilmiah ini berhasil diselesaikan. Penelitian ini berjudul Identifikasi Keragaman Gen ATP Binding
Cassette (ABCG2|PstI) pada Sapi Friesian Holstein dengan Metode PCR-RFLP.
Terima kasih penulis ucapkan kepada Prof Dr Ir Cece Sumantri, MAgrSc dan Ir Anneke Anggraeni, MSi PhD selaku dosen pembimbing atas bimbingannya selama pelaksanaan penelitian dan penulisan skripsi. Penulis juga mengucapkan terima kasih kepada Dr Jakaria, SPt MSi, Ir Anita S Tjakraadidjaja, MRurSc, dan Bapak Edit Lesa Aditia, SPt MSc selaku komisi penguji dalam ujian sidang sarjana, yang telah memberikan banyak saran dan masukan dalam perbaikan penulisan skripsi ini, serta Dr Ir Asnath M Fuah, MS selaku dosen pembimbing akademik. Ucapan terima kasih penulis sampaikan pula kepada kak Eryk, Ferdy, Irine, Isyana atas segala bentuk arahan teknis selama pelaksanaan penelitian di Laboratorium Genetika dan Molekuler Ternak. Ungkapan terima kasih juga disampaikan penulis kepada kedua orang tua, Bapak Sutrisno dan Ibu Laswiwinarsih, seluruh keluarga besar atas segala doa dan kasih sayangnya, serta sahabat, dan teman-teman Golden Ranch 46.
Bogor, September 2013
8
DAFTAR ISI
DAFTAR ISI vii
DAFTAR TABEL viii
DAFTAR GAMBAR viii
PENDAHULUAN 1
Latar Belakang 1
Tujuan Penelitian 1
Ruang Lingkup Penelitian 1
METODE 2
Waktu dan Tempat Penelitian 2
Bahan 2
Alat 2
Prosedur 3
Ekstraksi DNA 3
Polymerase Chain Reaction (PCR) 3
Elektroforesis 3
Restriction Fragment Length Polymorphisms (RFLP) 4
Genotyping 4
Analisis Data 4 HASIL DAN PEMBAHASAN 5 Amplifikasi Gen ABCG2|PstI 5 Keragaman Gen ABCG2|PstI 6 Frekuensi Genotipe dan Alel Gen ABCG2|PstI 8 Derajat Polimorfisme Gen ABCG2|PstI 10
SIMPULAN DAN SARAN 10
DAFTAR TABEL
1 Frekuensi genotipe dan alel gen ABCG2|PstI pada sapi FH 8 2 Keragaman gen ABCG2 pada beberapa bangsa sapi FH 9 3 Nilai PIC gen ABCG2|PstI pada sapi Friesian Holstein 10
DAFTAR GAMBAR
1 Amplifikasi gen ABCG2 pada gel agarose 1.5 % 5 2 Posisi penempelan primer forward dan reverse pada sekuen gen
PENDAHULUAN
Latar Belakang
Sapi Friesian Holstein (FH) merupakan bangsa sapi perah jenis Bos taurus
yang umum dipelihara di Indonesia, karena mempunyai kemampuan untuk memproduksi susu dalam jumlah tinggi. Rataan produksi susu sapi FH di daerah asalnya saat ini sekitar 6 900-7 600 liter/tahun (DEFRA 2012), sedangkan di Indonesia hanya sekitar 1 700-2 300 liter/tahun (Dirjen PKH 2011). Jumlah produksi dan komposisi susu dipengaruhi oleh faktor genotipe, lingkungan, dan interaksi antara keduanya. Sifat ekonomis tersebut pada dasarnya merupakan sifat kuantitatif yang dikontrol oleh banyak gen. Salah satu upaya meningkatkan kemampuan genetik dalam hal jumlah produksi dan komposisi susu dapat melalui seleksi terhadap kandidat gen-gen major yang mengontrol sifat terkait. Pemanfaatan teknologi molekuler memungkinkan untuk dilakukannya seleksi pada tingkat DNA, yang diharapkan akan meningkatkan respon seleksi. Identifikasi terhadap keragaman gen yang terkait dengan sifat yang ingin diperbaiki merupakan langkah awal untuk dapat mengetahui individu ternak yang memiliki genotipe dengan ekspresi keunggulan sifat tersebut.
Gen ATP Binding Cassette, subfamily G, member 2 (ABCG2) yang terletak
pada kisaran wilayah 49 ± 0.5 cM dalam susunan QTL (Quantitative Traits Loci) dari kromosom 6 (BTA6), berada dalam wilayah yang sama dengan gen PPARGC1 dan SPP1 (Ogorevc et al. 2009), diketahui sebagai salah satu gen yang berpengaruh besar terhadap sifat susu (Ron et al. 2001; Olsen et al. 2005). Gen ABCG2|PstI memiliki dua alel, yaitu alel A dan alel C. Adanya perubahan alel A menjadi alel C menyebabkan terjadinya perubahan sintesis asam amino tyrosine
menjadi serine pada posisi kodon ke-581 (Y581S) di exon 14. Alel A berpengaruh pada produksi susu yang rendah dengan persentase lemak dan protein susu yang tinggi, sebaliknya alel C berpengaruh pada produksi susu yang tinggi dan persentase lemak dan protein susu yang rendah (Cohen-Zinder et al. 2005; Olsen
et al. 2007). Dengan demikian identifikasi keragaman gen ABCG2 pada sapi
perah FH di Indonesia dapat digunakan sebagai langkah awal dalam upaya perbaikan sifat produksi susu. Oleh karena itu diperlukan adanya penelitian mengenai keragaman genetik gen ABCG2 yang terkait dengan sifat produksi dan komposisi susu (protein dan lemak) pada sapi perah Friesian Holstein di Indonesia.
Tujuan Penelitian
Penelitian ini bertujuan untuk mengidentifikasi keragaman gen ABCG2|PstI
pada sapi perah Friesian Holstein (FH) dengan metode Polymerase Chain
Reaction-Restriction Fragment Length Polymorphism (PCR-RFLP).
Ruang Lingkup Penelitian
2
(BPPT-SP Cikole), dan Peternakan Rakyat (KPSBU Pasar Kemis dan Cilumber). Keragaman gen diidentifikasi menggunakan metode PCR-RFLP dengan enzim restriksi PstI.
METODE
Waktu dan Tempat Penelitian
Penelitian dilaksanakan selama empat bulan yaitu pada bulan Januari 2013 sampai April 2013. Penelitian dilaksanakan di Laboratorium Genetika Molekuler Ternak, Bagian Pemuliaan dan Genetika Ternak, Departemen Ilmu Produksi dan Teknologi Peternakan, Fakultas Peternakan, Institut Pertanian Bogor.
Bahan
Sampel penelitian ini menggunakan hasil ekstraksi DNA sapi FH, koleksi Laboratorium Genetika dan Molekuler Ternak, Fakultas Peternakan, IPB. Sampel darah sapi FH bersumber dari UPT Nasional (BBIB Singasari = 26 jantan, BIB Lembang = 8 jantan, BPPTU Baturaden = 112 betina, dan BET Cipelang = 18 betina), UPT Daerah (BPPT-SP Cikole = 17 betina), dan Peternakan Rakyat (KPSBU Pasir Kemis = 21 betina dan Cilumber = 22 betina).
Ekstraksi DNA dilakukan dengan menggunakan bahan-bahan meliputi sampel darah/semen, DW, 1xSTE, 10% SDS, Proteinase-K (5mg/ml), NaCl 5 M,
phenol, CIAA, EtOH 70%, buffer TE 80%.
Bahan-bahan untuk proses PCR meliputi primer forward, primer reverse, dNTPs, MgCl2, 10xBuffer, dan Taq polymerase, destilated water. Primer yang
digunakan mengacu pada hasil modifikasi Komisarek dan Dorynek (2009) yaitu
forward: 5’-AAC AGC CTC AGC TCC AGA GAG ATA T-3’ dan reverse: 3’
-CGG TGA CAG ATA AGG AGA ACA TAC T-5’.
Proses elektroforesis menggunakan gel agarose 1.5%, 0.5xTBE, Ethidium Bromide, loading dye, dan marker 100 pb. Bahan-bahan untuk RFLP meliputi enzim restriksi PstI, buffer O, dan destilated water.
Alat
Peralatan yang digunakan dikelompokkan sesuai dengan urutan proses kerja. Pertama, elektroforesis meliputi mikropipet beserta tip, vortex, sentrifuge, inkubator, rotary mixer, freezer. Kedua, proses PCR meliputi mikropipet, tip, tabung 1.5 ml, tabung 0.2 ml, rak tabung, vortex, sentrifuge, dan AB Applied Biosystem GeneAmp* PCR System 9700/Eppendorf. Ketiga, proses elektroforesis (timbangan digital, gelas kimia, sudip, cetakan gel, sisir, gelas ukur, cawan,
microwave, mikropipet, tip, magnetic stirrer, power supply 100 volt, dan uv
transilluminator). Keempat, proses Restriction Fragment Length Polymorphism
(RFLP) meliputi mikropipet, tip, tabung 1.5 ml, tabung 0.5 ml, rak tabung, vortex,
Prosedur
Ekstraksi DNA
DNA diisolasi dengan metode phenol chloroform mengacu pada Sambrook
et al. (1989). Sampel darah sebanyak 200 µl ditambah dengan 1 000 µl DW (destilated water), disentrifuse pada 8000 rpm selama lima menit, supernatan dibuang. Selanjutnya untuk melisiskan sel, degradasi protein dan bahan organik dilakukan dengan penambahan 1xSTE 350 µl, 10% SDS 40 µl, dan 10 µl Proteinase K (5 mg/ml) serta dikocok (tilting) dalam inkubator dengan suhu 55 °C selama dua jam. Pemurnian sel dilakukan dengan menambahkan 40 µl NaCl 5 M, 400 µl phenol, dan 400 µl CIAA (24:1) serta dikocok pada suhu ruang selama satu jam. Fase DNA dipisahkan dari fase phenol dengan sentrifugasi pada 12 000 rpm selama lima menit. Setelah itu, supernatan dipindah ke tabung baru, ditambahkan 40 µl NaCl 5 M dan 800 µl EtOH, dihomogenkan, dan dilakukan freezing over
night. Campuran tersebut selanjutnya disentrifugasi pada 12 000 rpm selama lima
menit, supernatan dibuang, serta ditambah dengan 800 µl EtOH 70%. Tahap tersebut diulangi, selanjutnya diuapkan. DNA yang berupa benang putih dilarutkan dengan 100 µl buffer TE 80%.
Polymerase Chain Reaction (PCR)
Ekstraksi sampel DNA sebanyak 2 µl menggunakan mikropipet dimasukkan ke dalam tabung 0.2 ml. Kemudian ditambahkan 13 µl PCR mix (0.1 µl primer
forward dan reverse; 0.1 µl dNTPs; 0.5 µl MgCl2; 1.5 µl 10xbuffer; 0.05 µl Taq
polymerase; 10.75 µl destilated water). Sampel selanjutnya disentrifuse,
kemudian dimasukkan ke dalam mesin thermocycler (AB Applied Biosystem GeneAmp* PCR System 9700/Eppendorf).
Kondisi PCR yang digunakan meliputi predenaturasi (95 °C, 5 menit), denaturasi (95 °C, 30 detik), annealing (58 °C, 45 detik), ekstensi (72 °C, 1 menit), ekstensi akhir (72 °C, 5 menit), dengan jumlah siklus PCR sebanyak 35 siklus.
Elektroforesis
Elektroforesis diawali dengan pembuatan gel agarose 1.5%. Gel agarose dibuat dengan melarutkan agarose (0.45 g untuk cetakan besar atau 0.3 g untuk cetakan kecil) dengan 0,5xTBE (30 ml untuk cetakan besar atau 20 ml untuk cetakan kecil). Larutan agarose kemudian dipanaskan dalam microwave (suhu
medium high selama ± 5 menit). Setelah itu, larutan agarose diaduk dengan
magnetic stirrer dan ditambahkan Ethidium Bromide (2.5 µl untuk cetakan besar
atau 2 µl untuk cetakan kecil). Larutan agarose didiamkan hingga berubah bentuk menjadi gel.
4
Restriction Fragments Length Polymorphism (RFLP)
Sampel hasil PCR sebanyak 5 µl dimasukkan ke dalam tabung 0,5 ml menggunakan mikropipet. Sebanyak 2 µl RE mix (0.7 µl buffer O; 0.3 µl PstI; 1
µl destilated water) selanjutnya ditambahkan ke dalam sampel. Sampel
disentrifuse dan diinkubasi dalam inkubator pada suhu 37 °C selama 16 jam. Sampel hasil PCR-RFLP dielektroforesis pada gel agarose 2% dengan tegangan 100 volt. Hasil elektroforesis divisualisasi dengan uv transilluminator.
Genotyping
Genotyping dilakukan dengan menarik lurus fragmen pita DNA sampel ke
arah marker, selanjutnya dibandingkan terhadap marker untuk mengetahui panjang fragmen. Diperoleh genotipe AA apabila terdapat satu fragmen pita DNA (292 pb), genotipe AC dengan tiga fragmen pita DNA (292 pb, 268 pb, dan 24 pb), genotipe CC dengan dua fragmen pita DNA (268 pb dan 24 pb).
Analisis Data
Data hasil genotyping gen ABCG2|PstI dihitung mengenai frekuensi genotipe, frekuensi alel, dan nilai PIC. Selanjutnya data dianalisis dan dibandingkan antarlokasi sumber sampel sapi FH pengamatan.
Frekuensi Alel dan Genotipe
Frekuensi genotipe merupakan rasio dari jumlah suatu genotipe terhadap jumlah populasi yang diamati. Keragaman genotipe pada masing-masing individu ternak dapat ditentukan berdasarkan fragmen pita-pita yang ditemukan. Frekuensi genotipe (Xii) diukur dengan menggunakan rumus Nei dan Kumar (2000) sebagai
berikut:
Frekuensi alel merupakan frekuensi relatif dari suatu alel dalam populasi atau jumlah suatu alel terhadap jumlah keseluruhan alel yang terdapat dalam suatu populasi. Frekuensi alel (Xi) dari gen ABCG2 dihitung berdasarkan rumus Nei
dan Kumar (2000) sebagai berikut:
Keterangan:
Χii : frekuensi genotipe ke-ii
Χi : frekuensi alel ke-i
Polymorphic Information Content (PIC)
Kegunaan dari marker genetik untuk analisa garis keturunan bergantung pada seberapa sering marker tersebut ditemukan dalam kondisi polimorfik. Derajat polimorfisme dapat diukur dengan menghitung nilai PIC (Buchanan dan Thue 1998). Perhitungan nilai PIC mempertimbangkan jumlah alel yang dimiliki oleh lokus marker dan frekuensi dari alel-alel tersebut (Botstein et al. 1980).
Keterangan:
k : jumlah alel pi : frekuensi alel ke-i pj : frekuensi alel ke-j
HASIL DAN PEMBAHASAN
Amplifikasi Gen ABCG2|PstI
Amplifikasi gen ABCG2 pada penelitian ini dilakukan dengan mengguna-kan metode PCR. PCR merupakan reaksi perbanyakan jumlah molekul DNA secara in vitro di dalam mesin thermocycler dengan bantuan oligonukleotida rantai pendek dari primer forward dan reverse, yang memiliki susunan basa nukleotida berkomplemen dengan DNA target; serta enzim Taq polymerase, yang berperan dalam sintesis molekul DNA baru (Muladno 2010). Hasil amplifikasi gen ABCG2 menggunakan marker 100 pb pada gel agarose 1.5% ditampilkan dalam Gambar 1.
Gambar 1 Amplifikasi gen ABCG2 sepanjang 292 pb pada gel agarose 1.5 %. M (marker); 1-7 (sampel)
M 1 2 3 4 5 6 7
500 pb 400 pb
300 pb 200 pb
100 pb
6
Amplifikasi gen ABCG2|PstI menunjukkan bahwa panjang pita yang diperoleh adalah 292 pb. Hal ini sesuai dengan penelitian Komisarek dan Dorynek (2009) yang menyebutkan bahwa panjang fragmen hasil amplifikasi gen ABCG2 adalah 292 pb. Tingkat keberhasilan amplifikasi gen ABCG2 mencapai 100% pada seluruh populasi, kecuali pada BBIB Singasari (81.25%) dan BPPTU Baturaden (90.32%). Adapun berdasarkan keseluruhan sampel, tingkat keberhasilan amplifikasi sebesar 92.56%.
Beberapa faktor yang menentukan tingkat keberhasilan PCR, antara lain suhu annealing, kualitas DNA hasil ekstraksi, dan komposisi bahan-bahan penyusun mix PCR yang digunakan. Beberapa hal yang penting untuk diperhatikan agar diperoleh hasil PCR yang optimal yaitu suhu annealing, konsentrasi Mg2+, konsentrasi primer, dan konsentrasi DNA target (Viljoen et al.
2005). Adapun posisi penempelan primer pada sekuen gen ABCG2|PstI
(GenBank Nomor Akses AJ871176) ditampilkan pada Gambar 2.
Keragaman Gen ABCG2|PstI
Analisis keragaman dilakukan setelah DNA hasil amplifikasi dipotong dengan menggunakan enzim restriksi. PstI merupakan enzim restriksi yang memiliki titik potong spesifik yaitu 5’ CTGCA|G 3’. Primer reverse yang digunakan pada penelitian ini telah dimodifikasi oleh Komisarek dan Dorynek (2009) dengan tujuan agar diperoleh titik potong oleh enzim PstI. Teknik introduksi sekuen primer ke dalam bagian dari situs restriksi dikenal dengan istilah PIRA (Primer Introduced Restriction Analysis). Teknik PIRA-PCR umumnya digunakan untuk mendeteksi keberadaan single nucleotide
polymorphisms (SNP). Untuk membuat rekayasa RFLP, ketidakcocokan
(mismatch) berupa basa tunggal (single base) biasanya diintroduksikan pada
ujung 3’ dari primer yang dekat dengan titik mutasi (Ke et al. 2001). Hasil
pemotongan enzim PstI menghasilkan tiga fragmen dengan panjang berbeda yaitu 292 pb (alel A) serta 268 pb dan 24 pb (alel C). Dengan demikian suatu individu
forward
62281 tattataaat atatttagat ccctaaacag cctcagctcc agagagatat acacatacaa 62341 agaggatagg gtaagtcttc agagtagtgg aaagaatttc aaagaagagt tgtccttagc 62401 agtattcacg agactgtcag ggacttaaag aggctatttg ctagacggca ccagatctga
Terjadi mutasi pada situs pemotongan enzim restriksi PstI (ctgca|g) pada posisi 62569 (Exon 14) Alel A : 5’...ATACGGCTATGCAGTATGTT...3’ sintesis AA tyrosine
Alel C : 5’...ATACGGCTCTGCAGTATGTT...3’ sintesis AA serine : Posisi penempelan primer
_____ : Situs pemotongan enzim restriksi PstI
dikatakan memiliki genotipe AA apabila proses restriksi menghasilkan satu fragmen pita (292 pb), genotipe AC jika tiga fragmen pita (292 pb, 268 pb, dan 24 pb), dan genotipe CC jika dua fragmen pita (268 pb dan 24 pb). Hasil genotiping gen ABCG2|PstI pada gel agarose 2% dapat dilihat pada Gambar 3.
Gambar 3 mengilustrasikan untuk masing-masing genotipe, panjang fragmen yang diperoleh sesuai dengan penelitian Komisarek dan Dorynek (2009). Panjang fragmen 24 pb tidak ditemukan pada genotipe AC dan CC. Hal ini disebabkan karena fragmen tersebut telah mengalami running off saat elektroforesis. Molekul DNA yang lebih kecil memiliki kecepatan bermigrasi ke kutub positif lebih cepat dibandingkan molekul DNA yang lebih besar saat elektroforesis (Muladno 2010). Namun demikian, adanya fragmen 292 pb dan 268 pb pada genotipe AC; serta fragmen 268 pb pada genotipe CC sudah cukup untuk mengidentifikasi kedua genotipe tersebut. Identifikasi keragaman gen ABCG2 menunjukkan bahwa ketiga genotipe (AA, AC, dan CC) ditemukan pada 224 sapi FH pengamatan. Hal ini berbeda dengan penelitian yang dilakukan oleh Komisarek dan Dorynek (2009) menggunakan 453 ekor Polish Holstein; Yildrim dan Sahin (2010) dengan 207 ekor Turkish Holstein. Hasil penelitian keduanya tidak menemukan adanya individu dengan genotipe homozigot CC.
Berdasarkan 224 ekor sapi FH pengamatan, genotipe homozigot AA paling banyak ditemukan. Hal menarik yang ditemukan dalam penelitian ini yaitu bahwa jumlah genotipe AC hanya sedikit ditemui pada lima lokasi, yaitu BIB Lembang (1 ekor), BPPT-SP Cikole (2 ekor), KPSBU Pasar Kemis (1 ekor), BPPTU Baturaden (2 ekor), dan Cilumber (1 ekor), bahkan genotipe homozigot CC hanya ditemui di lokasi BPPT-SP Cikole (1 ekor). Dengan demikian keragaman gen ABCG2 terendah ditemukan pada lokasi BBIB Singasari dan BET Cipelang karena hanya terdapat satu jenis genotipe yaitu genotipe homozigot AA. Sebaliknya, keragaman gen tertinggi ditemukan pada lokasi BPPT-SP Cikole karena terdapat tiga genotipe yaitu AA, AC, dan CC.
8
Frekuensi Genotipe dan Alel Gen ABCG2|PstI
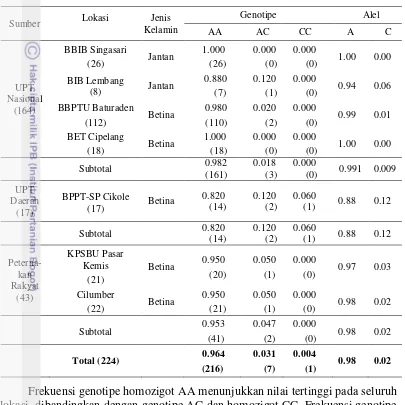
Nilai frekuensi genotipe dan alel sapi FH pengamatan di masing-masing lokasi ditampilkan pada Tabel 1.
Frekuensi genotipe homozigot AA menunjukkan nilai tertinggi pada seluruh lokasi, dibandingkan dengan genotipe AC dan homozigot CC. Frekuensi genotipe AA memiliki kisaran 0.82-1.00. Frekuensi genotipe AA bernilai 1.00 ditemukan pada BBIB Singasari dan BET Cipelang, menunjukkan bahwa pada keseluruhan sapi FH pengamatan bergenotipe AA. Sementara itu, frekuensi genotipe CC bernilai 0.00 pada seluruh lokasi, kecuali di BPPT-SP Cikole (0.06). Nilai frekuensi genotipe 0.00 tersebut menunjukkan bahwa pada sapi FH pengamatan tidak ditemukan adanya individu bergenotipe CC. Secara keseluruhan dari 224 sapi FH yang diamati, frekuensi genotipe AA > AC > CC (AA = 0.964; AC = 0.031; dan CC = 0.004). Hasil penelitian Komisarek dan Dorynek (2009) memperoleh nilai frekuensi genotipe AA dan AC masing-masing sebesar 0.985 dan 0.015 serta tidak ditemui individu bergenotipe CC.
Berdasarkan Tabel 1, diketahui bahwa nilai frekuensi alel A pada sapi FH di BBIB Singasari bernilai 1.00 yang menunjukkan bahwa semua ternak tersebut bergenotipe AA, sedangkan pada sapi FH di BIB Lembang diperoleh frekuensi Tabel 1 Frekuensi genotipe dan alel gen ABCG2|PstI pada sapi FH
alel A sebesar 0.94 dan frekuensi alel C sebesar 0.06. Pada sapi perah FH betina di BET Cipelang diperoleh frekuensi alel A sebesar 1.00, sedangkan pada keempat lokasi lainnya diperoleh nilai frekuensi alel A dengan kisaran 0.88-0.99. Untuk keseluruhan sapi FH pengamatan, menunjukkan bahwa frekuensi total alel A sebesar 0.98 dan alel C sebesar 0.02. Dengan demikian, frekuensi alel A sangat tinggi jika dibandingkan dengan alel C. Hal ini sesuai dengan studi pada sapi FH lainnya, dengan frekuensi alel A = 0.99 dan alel C = 0.01 (Komisarek dan Dorynek 2009); frekuensi alel A = 0.98 dan alel C = 0.02 (Yildrim dan Elif 2010); frekuensi alel A = 0.80 dan alel C = 0.20 (Cohen-Zinder et al. 2005).
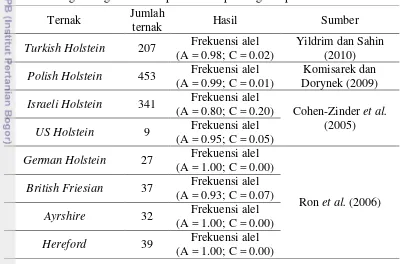
Ron et al. (2006) dalam penelitiannya pada gen ABCG2 dengan 35 bangsa
sapi perah (3 Bos indicus dan 32 Bos taurus), menjelaskan bahwa alel A ditemukan paling banyak pada seluruh bangsa dan tidak ditemukan alel C pada bangsa Bos indicus. Sebaliknya, alel C hanya ditemukan pada 12 bangsa Bos
taurus, dengan kisaran frekuensi alel A sebesar 0.80-0.99. Keragaman gen
ABCG2 pada beberapa bangsa sapi perah di dunia dapat dilihat pada Tabel 2. Tabel 2 Keragaman gen ABCG2 pada beberapa bangsa sapi FH di dunia
Tabel di atas menunjukkan bahwa frekuensi alel gen ABCG2|PstI yang diperoleh pada hasil penelitian sapi Friesian Holstein pengamatan tidak jauh berbeda dengan frekuensi alel yang terdapat pada beberapa bangsa sapi perah di dunia. Dengan demikian dapat disimpulkan bahwa pada gen ABCG2, alel A memiliki frekuensi lebih tinggi dibandingkan alel C, sehingga seleksi terhadap alel A akan lebih terbatas untuk dilakukan.
Tingkat keragaman suatu gen dalam suatu populasi dapat dilihat berdasarkan nilai dari frekuensi alel (Nei 1987). Suatu alel dapat dikatakan bersifat polimorfik apabila frekuensi alel sama dengan atau kurang dari 0.99 (Nei 1987). Adapun suatu populasi dapat dikatakan beragam apabila terdapat dua atau lebih alel dalam suatu lokus dengan frekuensi yang cukup (Falconer dan Mackay 1996). Dengan demikian disimpulkan bahwa gen ABCG2 pada keseluruhan
Ternak Jumlah
ternak Hasil Sumber
Turkish Holstein 207 Frekuensi alel
(A = 0.98; C = 0.02)
Yildrim dan Sahin (2010)
Polish Holstein 453 Frekuensi alel
(A = 0.99; C = 0.01)
Komisarek dan Dorynek (2009)
Israeli Holstein 341 Frekuensi alel
(A = 0.80; C = 0.20) Cohen-Zinder et al.
(2005)
US Holstein 9 Frekuensi alel
(A = 0.95; C = 0.05)
German Holstein 27 Frekuensi alel
(A = 1.00; C = 0.00)
Ron et al. (2006)
British Friesian 37 Frekuensi alel
(A = 0.93; C = 0.07)
Ayrshire 32 Frekuensi alel
(A = 1.00; C = 0.00)
Hereford 39 Frekuensi alel
10
populasi bersifat polimorfik karena memiliki frekuensi alel kurang dari atau sama dengan 0.99 (0.88-0.99), kecuali pada lokasi di BBIB Singasari dan BET Cipelang yang bersifat monomorfik (frekuensi alel sama dengan 1.00).
Derajat Polimorfisme Gen ABCG2|PstI
Derajat polimorfisme suatu gen dapat diketahui dengan menghitung nilai PIC. Nilai PIC dari gen ABCG2|PstI ditampilkan pada Tabel 3.
Tabel 3. Nilai PIC gen ABCG2|PstI pada sapi Friesian Holstein
Peternakan Rakyat KPSBU Pasir Kemis 0.056
Cilumber 0.038
PIC menggambarkan nilai heterozigositas yang tekoreksi oleh informasi perkawinan sebagian (partially informative mating). Nilai PIC memiliki kisaran 0-1. Nilai PIC sama dengan nol diperoleh apabila hanya ditemukan satu alel pada marker genetik, sedangkan diperoleh nilai PIC sama dengan satu (PIC = 1) apabila terdapat jumlah alel yang tak terhingga (Hildebrand et al. 1992). Apabila suatu gen hanya memiliki dua alel maka akan dihasilkan nilai PIC maksimum sebesar 0.375 (Hildebrand et al. 1992).
Nilai PIC sama dengan nol ditemukan pada lokasi BBIB Singasari dan BET Cipelang, menunjukkan bahwa pada lokasi tersebut gen ABCG2|PstI bersifat monomorfik karena hanya ditemukan satu alel, yaitu alel A. Sementara di lokasi lainnya, derajat polimorfisme tertinggi dari gen ABCG2|PstI ditemukan di lokasi BPPT SP Cikole (PIC = 0.189). Secara keseluruhan derajat polimorfisme gen ABCG2|PstI memiliki kisaran 0.020-0.189.
SIMPULAN DAN SARAN
Simpulan
Saran
Penambahan sampel sapi FH perlu dilakukan untuk memperoleh data lebih akurat mengenai kondisi keragaman gen ABCG2 pada sapi FH di Indonesia. Selain itu diperlukan penelitian lanjutan untuk mengetahui hubungan antara keragaman gen ABCG2|PstI terhadap sifat produksi dan komposisi susu sapi FH serta kemampuan gen ABCG2 sebagai multidrugs transporter.
DAFTAR PUSTAKA
Botstein D, White RL, Skolnick M, Davis RW. 1980.Construction of a genetic linkage map in man using restriction fragment length polymorphisms. Am. J. Hum. Genet. 32: 314–331.
Buchanan FC, Thue TD. 1998. Intrabreed polymorphic information content of microsatellites in cattle and sheep. [Internet]. [diunduh 2013 Agustus 23]; Canada (CAN): Can. J. Anim. Sci. 78:425-428. Tersedia pada:
http://pubs.aic.ca/doi/pdfplus/10.4141/A98-002.
Cohen-Zinder M, Serioussi E, Larkin DM, Loor JJ, Evertvan der Wind A, Lee JH, Drackley JK, Band MR, Hernandez AG, Shani M et al. 2005. Identification of a missense mutation in the bovine ABCG2 gene with a major effect on the QTL on chromosome 6 affecting milk yield and composition in Holstein cattle. Genome Res. 15 (7):936-944.doi:10.1101/gr.3806705.
[DEFRA]. Department for Environment, Food and Rural Affairs. 2012.
Agriculture in the United Kingdom. Northern Ireland (UK): Department of
Agriculture and Rural Development.
[Dirjen PKH]. Direktorat Jenderal Peternakan dan Kesehatan Hewan. 2011.
Statistik Peternakan dan Kesehatan Hewan. Jakarta (ID): Kementerian
Pertanian Republik Indonesia.
Falconer DS, Mackay TFC. 1996. Introduction to Quantitive Genetics. Ed ke-4. New York (US): Longman.
Hildebrand CE, Torney DC, Wagner RP. 1992. Informativeness of polymorphic DNA markers. Los Alamos Sci. 20:100-102.
Ke Xiayi, Collins A, Ye S. 2001. PIRA PCR designer for restriction analysis of single nucleotide polymorphism. Bioinformatics Appl. Note. 17(9):838-839. Komisarek J, Dorynek Z. 2009. Effect of ABCG2, PPARGC1A, OLR1 and SCD
gene polymorphism on estimated breeding values for functional and production traits in Polish Holstein-Friesian bulls. J Appl Genet. 50(2):125-132.doi: 10.1007/BF03195663.
Muladno. 2010. Seputar Teknologi Rekayasa Genetika. Ed ke-2. Bogor (ID): Pustaka Wirausaha Muda & USESE Foundation.
Nei M. 1987. Molecular Evolutionary Genetics. New York (USA): Columbia University Press.
Nei M, Kumar, S. 2000. Molecular, Evolution and Phylogenetics. New York (US): Oxford University Press.
12
Olsen HG, Lien S, Gautier M, Nilsen H, Roseth A, Berg PR, Sundsaasen KK, Svendsen M, Meuwissen THE. 2005. Mapping of a milk production quantitative trait locus to a 420-kb region on bovine chromosome 6. Amerika (US): Genetics. 169(1):275-283 .doi: 10.1534/genetics.104.031559.
Olsen HG, Nilsen H, Hayes B, Berg PR, Svendsen M, Lien S, Meuwissen THE. 2007. Genetic support for a quantitative trait nucleotide in the ABCG2 gene affecting milk composition of dairy cattle. BMC Genet.8:32.doi: 10.1186/1471-2156-8-32.
Ron M, Cohen-Zinder M, Peter C, Weller JI, Erhardt G. 2006. A polymorphism in ABCG2 in Bos indicus and Bos taurus cattle breeds. J Dairy Sci. 89(12):4921-4923.doi:10.3168/jds.S0022-0302(06)72542-5.
Ron M, Kliger D, Feldmesser E, Seroussi E, Ezra E, Weller JI. 2001. Multiple quantitative trait locus analysis of bovine chromosome 6 in the Israeli Holstein population by a daughter design. [Internet]. [diunduh 2013 Juli 15]; Amerika (US): Genetics. 159(2):727-735. Tersedia pada:
http://www.ncbi.nlm.nih.gov/pmc.articles/PMC1461848/.
Sambrook, J, Fritsch F, Miniatis T. 1989. Molecular Cloning Laboratory Manual. 3rd Edition. New York (US): Cold Spring Harbor Laboratory Press.
Viljoen GJ, Nel LH, Crowther JR. 2005. Molecular Diagnostic PCR Handbook. Netherland (NL): Springer.
Yildrim M, Sahin E. 2010. ABCG2 gene polymorphism in Holstein cows of Turkey: research article. Kafkas Univ Vet Fak Derg [Internet]. [diunduh 2012 Nov 7]; Turki (TR). Kafkas Univ Vet Fak Derg. 16 (3): 473-476. Tersedia pada: http://vetdergi.kafkas.edu.tr/extdocs/2010 3/473 476.pdf.
RIWAYAT HIDUP
Penulis merupakan anak pertama dari dua bersaudara, dilahirkan di Rembang pada 1 November 1991 dari pasangan Bapak Sutrisno dan Ibu Laswiwinarsih. Penulis menempuh pendidikan di SMP Negeri 1 Lasem pada 2003-2006 dan SMA Negeri 1 Rembang pada 2006-2009 dan diterima di Institut Pertanian Bogor (IPB) melalui jalur Undangan Seleksi Mahasiswa IPB (USMI) pada 2009 dengan Mayor Ilmu Produksi dan Teknologi Peternakan dengan Minor Nutrisi Ternak, Fakultas Peternakan.