Keragaman genetik dan adaptabilitas gandum (Triticum aestivum L ) introduksi di lingkungan tropis
Teks penuh
Gambar
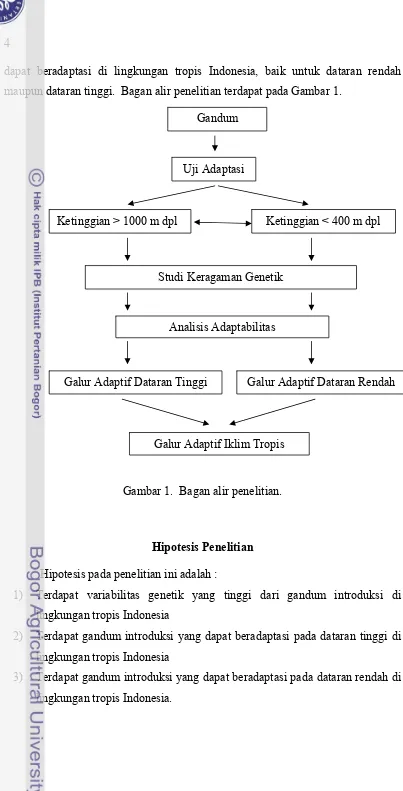



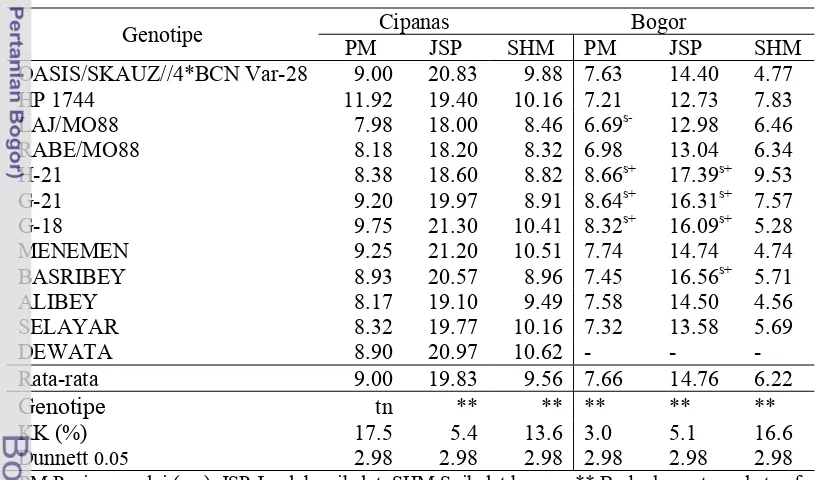
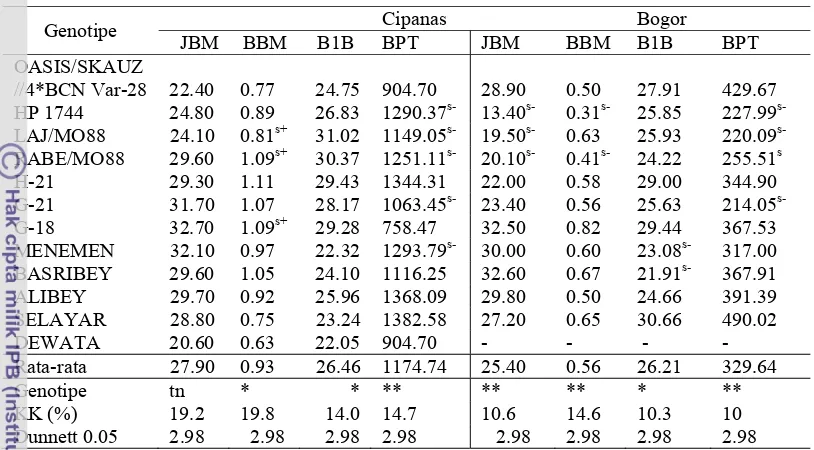
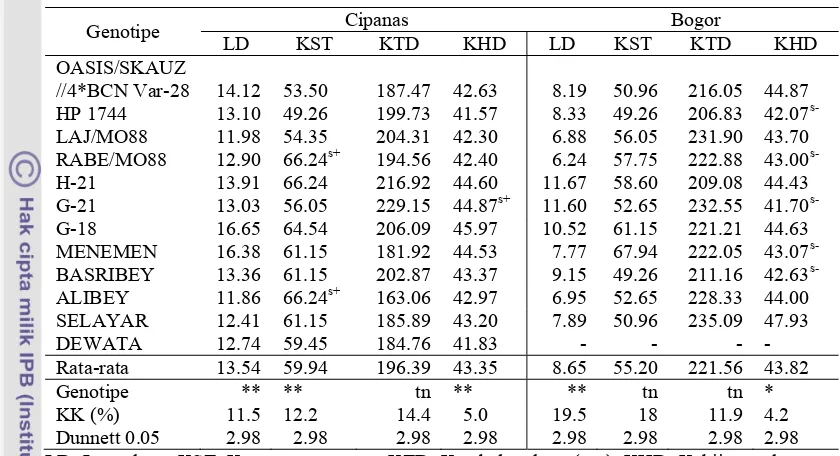


Dokumen terkait
Secara khusus penelitian ini ditujukan untuk (1) mendapatkan informasi respon genotipe gandum terhadap cekaman suhu tinggi; (2) memperoleh informasi kendali genetik
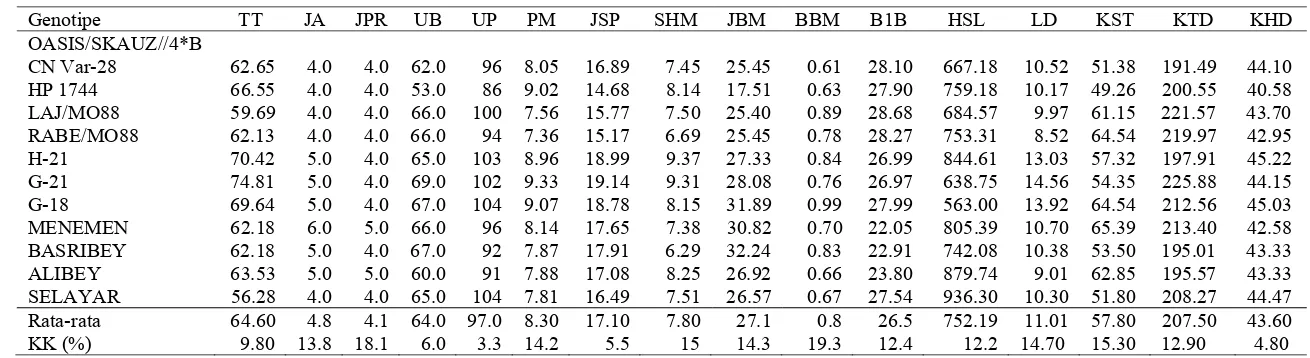
Hasil panen merupakan tujuan utama dalam budidaya tanaman sehingga seleksi galur-galur gandum introduksi dari CIMMYT ditujukan untuk mendapatkan galur gandum unggul
Dari hasil penelitian ini dapat ditarik kesimpulan sebagai berikut: genotip dan varietas gandum yang relatif mampu tumbuh lebih baik di dataran rendah Mangkang
Hasil pengamatan terhadap pertumbuhan genotipe gandum menunjukkan karakter yang memiliki ragam genetik lebih besar dari pada ragam lingkungan, maka penampilan karakter tersebut
Dari hasil penelitian dapat ditarik kesimpulan bahwa hama yang menyerang pada 13 galur dan empat varietas gandum di dataran rendah adalah jangkrik (Orthoptera:
Dalam kajian terhadap penampilan komponen pertumbuhan, terlihat bahwa beberapa hasil penelitian dari sejumlah genotipe yang diadaptasikan di wilayah tropis, mempunyai
Genotipe yang tidak berpengaruh nyata terhadap jumlah anakan produktif menunjukkan bahwa jumlah anakan produktif genotipe gandum introduksi yang terbentuk sama dengan
Hasil panen merupakan tujuan utama dalam budidaya tanaman sehingga seleksi galur-galur gandum introduksi dari CIMMYT ditujukan untuk mendapatkan galur gandum unggul
