ANALISIS GENETIK DAN SELEKSI GENERASI AWAL
SEGREGAN GANDUM
(
Triticum aestivum
L.)
BERDAYA HASIL TINGGI
AZIS NATAWIJAYA
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI
Dengan ini saya menyatakan bahwa tesis „Analisis Genetik dan Seleksi
Generasi Awal Segregan Gandum (Triticum aestivum L.) Berdaya Hasil Tinggi‟ adalah karya saya sendiri dengan arahan Komisi Pembimbing dan belum diajukan dalam bentuk apapun kepada Perguruan Tinggi manapun. Sumber informasi yang berasal atau kutipan dari karya yang diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Bogor, Agustus 2012
Azis Natawijaya
ABSTRACT
AZIS NATAWIJAYA. Genetic Analysis and Early Generation Selection to Identify High Yielding Segregants in Wheat (Triticum aestivum L.). Under direction of TRIKOESOEMANINGTYAS and DARDA EFENDI.
The preliminary breeding research series to develop adaptive variety in wheat to low and medium land regions had been conducted. It was started from evaluation and identification of agronomical responses of some introduced varieties, selective hybridization and development the breeding populations, until selection in early generation to identify high yielding potential segregants. First field experiment was conducted at two Indonesian agroecosystems (BIOTROP as low land representative and Cipanas as high land representative) from Mei to September 2011 and others were conducted at Research Station of BALITHI, Cipanas. The first experiment results showed that location had significant effect on wheat performance. High temperature in low land decrease the sink-source capacity, accelerate the seed filling period, and increase the empty floret. The tested genotypes were classified into three tolerance level. Oasis was classified as tolerant genotype, HP1744, LAJ, Menemen, Alibey, Selayar, and Dewata were classified as medium tolerant, and Rabe, H-21, G-21, G-18, and Basribey classified as sensitive genotypes. Analysis of agronomical traits-controlling gene action showed that all of observed traits controlled by poligenic with additive and epistatic effects. The total tillering number, productive tillering number, shoot-root dried weight, seed weight/panicle, and seed number/pland had strong phenotypic correlation with seed weight/plant. On the other hand, based on path analysis only two traits which had high direct effect to seed weight/plant. Those traits are useful for secondary trait in improving yield potential. Four selected traits (seed weight/plants, productive tillering number, seed weight/panicle, ratio of empty floret) had been used to develop an selection index. Selected segregants in Oasis x HP1744 population are O/HP-F2-87, O/HP-F2-93, O/HP-F2-100, 106, 51, 6, 16, 124, O/HP-F2-92, O/HP-F2-39, O/HP-F2-9, O/HP-F2-26, O/HP-F2-31, dan O/HP-F2-123. S/R-F2- 39, S/R-F2-13, S/R-F2-61, S/R-F2-19, S/R-F2-56, S/R-F2-7, and S/R-F2-64 are selected segregants in Selayar x Rabe population. In addition selected segregants in Dewata x Alibey population are D/A-F2-34, D/A-F2-13, D/A-F2-99, D/A-F2-22, D/A-F2-70, D/A-F2-4, D/A-F2-33, D/A-F2-80, D/A-F2-25, dan D/A-F2-65. All of the selected segregants have higher phenotypic performance in all observed traits than their parents.
RINGKASAN
AZIS NATAWIJAYA. Analisis Genetik dan Seleksi Generasi Awal Segregan Gandum (Triticum aestivum L.) Berdaya Hasil Tinggi. Dibimbing oleh TRIKOESOEMANINGTYAS dan DARDA EFENDI.
Gandum merupakan salah satu tanaman pangan penting di dunia termasuk di Indonesia. Kebutuhan gandum di Indonesia terus meningkat secara signifikan dari tahun 1984 sampai 2011. Sampai saat ini, Indonesia masih sangat tergantung terhadap produk gandum impor. Perbedaan kesesuaian kondisi agroklimat yang meliputi faktor temperatur dan curah hujan merupakan kendala utama dalam upaya produksi gandum di Indonesia. Varietas adaptif merupakan salah kunci penentu keberhasilan pengembangan gandum di Indonesia. Melalui program pemuliaan tanaman yang sistematis varietas adaptif dapat dicapai. Tujuan utama pemuliaan gandum di Indonesia yaitu meningkatkan toleransi gandum terhadap cekaman suhu tinggi dan meningkatkan potensi hasil. Rangkaian penelitian yang dimulai dengan mengevaluasi plasma nutfah introduksi, hibridisasi antar genotipe selektif, dan seleksi segregan harapan hasil rekombinasi telah dilakukan.
Secara umum penelitian ini ditujukan untuk merakit varietas gandum berdaya hasil tinggi dan toleran suhu tinggi. Secara khusus penelitian ini ditujukan untuk (1) mendapatkan informasi respon genotipe gandum terhadap cekaman suhu tinggi; (2) memperoleh informasi kendali genetik pada beberapa kelompok populasi F2 untuk karakter-karakter agronomi gandum di lingkungan berelevasi tinggi; (3) mendapatkan informasi hubungan antar karakter agronomi dan kriteria seleksi; (4) mendapatkan informasi parameter genetik pada populasi bersegregasi; (5) mengidentifikasi dan mendapatkan segregan potensial berdaya hasil tinggi.
Percobaan evaluasi respon genotipe gandum terhadap cekaman suhu tinggi dilakukan didua lokasi, Kebun Percobaan BIOTROP, Tajur (ketinggian tempat ± 250 m dpl) dan Kebun Percobaan BALITHI, Cipanas (ketinggian tempat ± 1100 m dpl) pada bulan Mei – September 2011. Studi kendali genetik, analisis hubungan antar karakter, dan seleksi generasi awal F2 dilakukan di Kebun Percobaan BALITHI, Cipanas pada bulan September 2011 – Januari 2012.
sink, serta mampu meminimalkan kehampaan malai. Sementara itu Rabe, H-21, G-21, G-18, dan Basribey merupakan genotipe sensitif suhu tinggi.
Hasil studi kendali genetik karakter agronomi diketahui bahwa hampir semua karakter agronomi yang dipelajari pada percobaan kedua dikendalikan secara poligenik dengan aksi gen aditif dengan pengaruh epistasis baik duplikat maupun komplementer. Namun demikian, terdapat beberapa karakter yang dikendalikan oleh gen mayor. Adanya epistasis yang nyata pada hampir semua karakter, menegaskan bahwa perlu lebih banyak individu-individu segregan yang ditanam di generasi berikutnya untuk meningkatkan perolehan segregan harapan. Selain itu, pengaruh epistasis komplementer dan duplikat akan menurun seiring dengan lanjutnya generasi bersegregasi.
Pada percobaan ketiga diketahui bahwa karakter jumlah anakan total, jumlah anakan produktif, bobot kering tajuk, bobot kering akar, dan jumlah biji/tanaman memiliki koefisien korelasi yang tertinggi, positif, sangat nyata dengan karakter bobot biji/tanaman. Sementara itu hasil analisis lintas menunjukkan bahwa karakter jumlah anakan produktif dan bobot biji/malai memiliki pengaruh langsung yang tertinggi dan perolehannya konsisten untuk ketiga kelompok populasi F2 yang diuji. Karakter jumlah anakan produktif dan bobot biji/malai dapat digunakan sebagai penanda untuk menyeleksi segregan-segregan berdaya hasil tinggi.
Nilai diferensial seleksi menggunakan seleksi berbasis indeks memberikan hasil yang tidak terpaut jauh dengan perolehan diferensial seleksi berbasis karakter target. Karakter – karakter yang digunakan dalam menyusun indeks seleksi didasarkan dari hasil penelitian ke 3 dan 4, berdasarkan keeratan hubungannya dengan karakter hasil dan nilai heritabilitas dengan pembobot heritabilitas dan koefisien lintas. Dengan demikian, segregan yang terpilih merupakan segregan yang memiliki karakter unggul untuk hampir semua karakter yang diamati. Diferensial seleksi berdasarkan indeks seleksi pada populasi Oasis x HP1744 yaitu 9.02 gram, populasi Selayar x Rabe yaitu 11.76 gram, dan populasi Dewata x Alibey yaitu 11.03 gram.
Segregan-segregan terseleksi di populasi Oasis x HP1744 yaitu O/HP-F2-87, O/HP-F2-93, O/HP-F2-100, O/HP-F2-106, O/HP-F2-51, O/HP-F2-6, O/HP-F2-16, O/HP-F2-124, O/HP-F2-92, O/HP-F2-39, O/HP-F2-9, O/HP-F2-26, O/HP-F2-31, dan O/HP-F2-123. S/R-F2- 39, S/R-F2-13, S/R-F2-61, S/R-F2-19, S/R-F2-56, S/R-F2-7, dan S/R-F2-64 merupakan segregan-segregan terseleksi dipopulasi Selayar x Rabe. Sedangkan segregan – segregan terseleksi di populasi Dewata x Alibey yaitu D/A-F2-34, D/A-F2-13, D/A-F2-99, D/A-F2-22, D/A-F2-70, D/A-F2-4, D/A-F2-33, D/A-F2-80, D/A-F2-25, dan D/A-F2-65. Segregan-segregan terpilih memiliki keragaan yang jauh lebih baik dibanding tetua-tetuanya untuk semua karakter.
© Hak Cipta milik IPB, tahun 2012
Hak Cipta dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan yang wajar IPB.
ANALISIS GENETIK DAN SELEKSI GENERASI AWAL
SEGREGAN GANDUM (
Triticum aestivum
L.)
BERDAYA HASIL TINGGI
AZIS NATAWIJAYA
Tesis
Sebagai salah satu syarat untuk memperoleh gelar
Magister Sains
pada Program Studi Pemuliaan dan Bioteknologi Tanaman
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
BOGOR
Judul Tesis : Analisis Genetik dan Seleksi Generasi Awal Segregan
Gandum (Triticum aestivum L.) Berdaya Hasil Tinggi
Nama : Azis Natawijaya
NRP : A253100051
Mayor : Pemuliaan dan Bioteknologi Tanaman
Disetujui
Komisi Pembimbing
Dr. Ir. Trikoesoemaningtyas, M.Sc. Dr. Ir. Darda Efendi, M.Si.
Ketua Anggota
Mengetahui
Ketua Program Studi Dekan Sekolah Pascasarjana Pemuliaan dan Bioteknologi Tanaman
Dr. Ir. Trikoesoemaningtyas, M.Sc. Dr. Ir. Dahrul Syah, M.Sc. Agr.
PRAKATA
Pengembangan genotipe-genotipe gandum yang toleran suhu tinggi dan
berproduksi tinggi di lingkungan berelevasi rendah dan medium merupakan
tujuan utama dalam kegiatan pemuliaan tanaman gandum di Indonesia. Tesis yang
berjudul “ Analisis Genetik dan Seleksi Generasi Awal Segregan Gandum
(Triticum aestivum L.) Berdaya Hasil Tinggi” merupakan salah satu upaya untuk
mengurai dan memahami kendala adaptasi gandum di lingkungan berelevasi
rendah dan upaya untuk merakit varietas gandum harapan. Di tahun mendatang
dan tidak terlalu lama diharapkan varietas – varietas gandum unggul dapat
dihasilkan.
Pada kesempatan ini penulis menyampaikan penghargaan dan ucapan
terima kasih kepada :
1. Dr. Ir. Trikoesoemaningtyas, M.Sc. dan Dr. Ir. Darda Efendi selaku komisi
pembimbing atas segala bimbingan, saran, kritikan, dan masukan selama
penyusunan proposal, pelaksanaan penelitian, dan penulisan tesis.
2. Dr. Ir. Yudiwanti Wahyu EK., M.S. selaku dosen penguji luar komisi pada
ujian akhir tesis atas saran-saran untuk perbaikan tesis.
3. Dr. Ir. Eny Widajati, M.S. selaku dosen penguji perwakilan dari Program
Studi Pemuliaan dan Bioteknologi Tanaman pada ujian akhir tesis atas
saran-saran untuk perbaikan tesis.
4. Seluruh staf pengajar di Program Studi Pemuliaan dan Bioteknologi
Tanaman IPB yang telah mendidik dan membekali penulis tentang
pengetahuan pemuliaan tanaman, bioteknologi, dan genetika.
5. Dr.Sc.Agr., Ir. Agung Karuniawan, M.Sc.Agr. dan Dr. Ir. Nani Hermiati,
M.S. staf pengajar di Program Studi Pemuliaan Tanaman UNPAD sebagai
guru penulis yang telah membina dan membekali penulis dengan
pengetahuan dasar kestatistikaan, sumber daya genetik tanaman, dan
6. Koordinator Proyek Pengembangan Gandum IPB (Dr. Ir.
Trikoesoemaningtyas, M.Sc.), Konsorsium Gandum Indonesia, Amin Nur,
S.P.M.Si., Dr. Azrai, S.P.,M.P., yang telah memberikan kesempatan kepada
penulis untuk bergabung dalam Tim Penelitian Pemuliaan Gandum Tropis.
7. Bapak Gregori G. Hambali, M.Sc. dan Ibu Ir. Indijani Kusudiarjo atas
dukungan moril maupun materil kepada penulis selama menempuh program
magister.
8. Ibu Anih, Bapak Nasirun (Alm.), Istri tercinta Zetri Ramalisa, adik-adik,
dan seluruh keluarga yang telah memberikan dorongan, motivasi, dan kasih
sayangnya.
9. Kepada seluruh teman-teman PBT-2010 dan FORSCA-AGH-IPB yang
tidak dapat penulis sebutkan satu per satu yang telah menjadi keluarga
penulis selama sama-sama menempuh pendidikan di Sekolah Pascasarjana
IPB.
10. Tim Penelitian Pemuliaan Gandum Lab. Genetika dan Pemuliaan IPB
(Amin Nur, S.P., M.Si., Rahmah, S.P., M.Si., Eka Bobby Febrianto, S.P.,
Sri Wardani, S.P., Maya, S.P.) atas kebersamaan dan bantuannya selama
melakukan penelitian.
Akhir kata, semoga karya kecil ini dapat membawa manfaat bagi penulis
dan juga bermanfaat bagi pengembangan dan kemajuan ilmu dan bidang
pertanian, khususnya di bidang genetika dan pemuliaan tanaman gandum tropis.
Bogor, Agustus 2012
RIWAYAT HIDUP
Penulis lahir di Bogor, Jawa Barat pada 27 Januari 1988 sebagai putra
pertama dari dua bersaudara dari ayah Nasirun (Alm.) dan ibu Anih. Pendidikan
sarjana ditempuh di Program Studi Pemuliaan Tanaman Jurusan Budidaya
Pertanian Fakultas Pertanian Universitas Padjadjaran pada Agustus 2006 – Maret
2010. Pada tahun 2010 penulis melanjutkan pendidikan magister di program studi
Pemuliaan dan Bioteknologi Tanaman, Departemen Agronomi dan Hortikultura,
IPB.
Penulis pernah menjadi asisten dosen mata kuliah Penerapan Metode
Pemuliaan Tanaman (PMPT) 2008/2009, Keprofesionalan Pemuliaan Tanaman II
(KPT-II) 2008/2009, Pelestarian dan Pemanfaatan Sumberdaya Hayati (PSDH)
2009/2010, dan Teknik Rancang dan Analisis Penelitian Pemuliaan Tanaman
(TRAPP) 2009/2010. Penulis juga pernah menjadi asisten peneliti di
Laboratorium Pemuliaan Tanaman UNPAD tahun 2008 – 2010, menjadi anggota
tim eksplorasi ubi-ubian lokal di Jawa Barat, koordinator tim eksplorasi
Amorphophallus di Sumatera Barat, koordinator tim eksplorasi kerabat liar ubi
jalar di Citatah Jawa Barat.
Pada tahun 2011 penulis mengikuti program summer course di IPB yang
diadakan IPB bekerja sama dengan Universitas Ibaraki Jepang, di tahun yang
sama penulis mengikuti program winter course di Universitas Ibaraki Jepang.
Menjadi asisten praktikum mata kuliah Dasar Pemuliaan Tanaman di IPB tahun
2012.
DAFTAR ISI
DAFTAR TABEL ... iv
DAFTAR GAMBAR ... iv
PENDAHULUAN ... ... 1
Latar Belakang ... 1
Tujuan Penelitian ... 4
Kerangka Pemikiran ... 4
Hipotesis ... 9
TINJAUAN PUSTAKA ... 10
Asal Usul Tanaman Gandum ... 10
Aspek Morfologi dan Agronomi Gandum ... 11
Respon Tanaman Terhadap Cekaman Suhu Tinggi ... 12
Mekanisme Adaptasi Gandum Terhadap Cekaman Suhu Tinggi ... 15
Pemuliaan Gandum Terhadap Cekaman Abiotik ... 16
Studi Pewarisan Sifat dan Pendugaan Parameter Genetik ... 19
Seleksi dan Hubungan Antar Karakter Tanaman ... 20
RESPON PLASMANUTFAH GANDUM DI DUA AGROEKOSISTEM DAN ANALISIS KERAGAMAN GENETIKNYA ... 25
Abstrak ... 25
Abstract ... 25
Pendahuluan ... 26
Bahan dan Metode ... 27
Hasil dan Pembahasan ... 32
Kesimpulan ... 48
PENDUGAAN AKSI GEN KARAKTER AGRONOMI PADA TIGA KELOMPOK POPULASI F2 GANDUM DI LINGKUNGAN BERELEVASI TINGGI ... 49
Abstrak ... 49
Abstract ... 49
Pendahuluan ... 50
Bahan dan Metode ... 51
Hasil dan Pembahasan ... 54
Kesimpulan ... 95
ANALISIS LINTAS KARAKTER AGRONOMI PADA TIGA KELOMPOK POPULASI F2 GANDUM DI LINGKUNGAN BERELEVASI TINGGI ... 96
Abstrak ... 96
Abstract ... 96
Pendahuluan ... 97
Bahan dan Metode ... 99
Hasil dan Pembahasan ... 100
KERAGAMAN GENETIK DAN HERITABILITAS KARAKTER AGRONOMI PADA TIGA KELOMPOK
POPULASI F2 GANDUM ... 115
Abstrak ... 115
Abstract ... 115
Pendahuluan ... 116
Bahan dan Metode ... 117
Hasil dan Pembahasan ... 118
Kesimpulan ... 127
SELEKSI GENERASI AWAL SEGREGAN F2 GANDUM UNTUK PERBAIKAN DAYA HASIL ... 128
Abstrak ... 128
Abstract ... 128
Pendahuluan ... 129
Bahan dan Metode ... 131
Hasil dan Pembahasan ... 132
Kesimpulan ... 145
PEMBAHASAN UMUM ... 146
KESIMPULAN DAN SARAN ... 150
DAFTAR PUSTAKA ... 152
DAFTAR TABEL
No. Halaman
1. Klasifikasi Beberapa Spesies Triticum Berdasarkan Kelas
Ploidi...……... 11
2. Struktur Analisis Ragam dan Penguraian Kuadrat Tengah... 29
3. Struktur Analisis Ragam Gabungan dan Penguraian Kuadrat
Tengah...……...………... 30
4. Hasil Analisis Ragam Gabungan di Dua Lokasi …...…... 33
5. Tabel Dwi Arah Karakter Tinggi Tanaman, Jumlah Anakan
Produktif, dan Umur Berbunga..………... 36
6. Tabel Dwi Arah Karakter Umur Panen, Luas Daun Bendera, dan Kehijauan Daun Bendera...
37
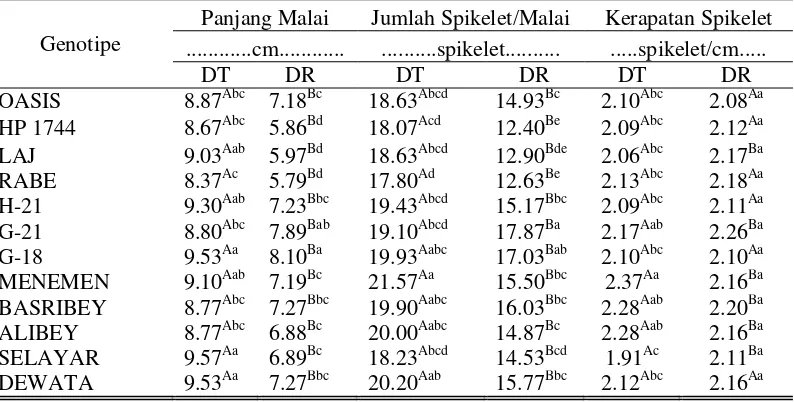
7. Tabel Dwi Arah Karakter Panjang Malai, Jumlah Spikelet/Malai, dan Kerapatan Spikelet...
39
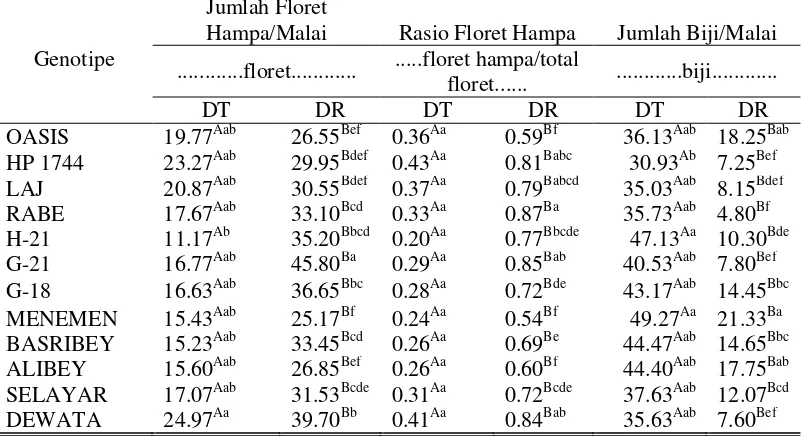
9. Tabel Dwi Arah Karakter Bobot Biji/Malai, Jumlah
Biji/Tanaman, dan Bobot Biji/Tanaman...
40
10. Indeks Sensitivitas 12 Genotipe Gandum... 41
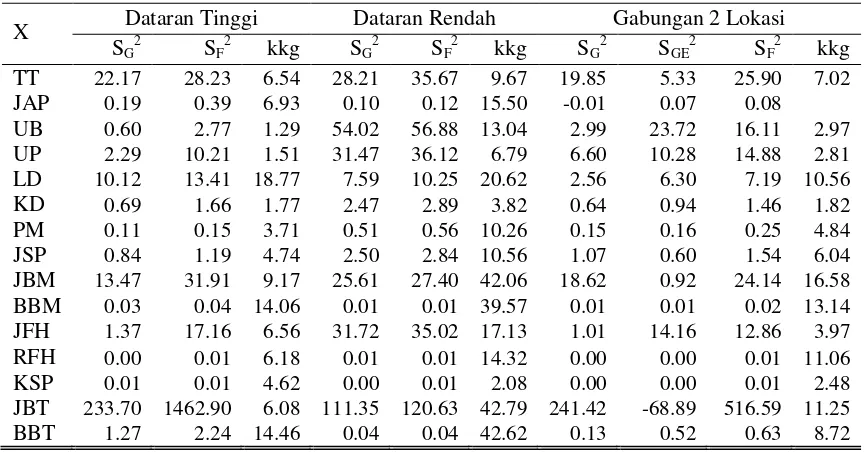
11. Komponen Ragam 15 Karakter Agronomi... 45
12. Heritabilitas dan Selang Kepercayaan Heritabilitas Karakter Agronomi...
47
13. Matriks Korelasi Karakter Agronomi Pada Populasi Oasis x HP174...
102
14. Matriks Korelasi Karakter Agronomi Pada Populasi Selayar x Rabe...
103
15. Matriks Korelasi Karakter Agronomi Pada Populasi Dewata x Alibey...
104
16. Matriks Analisis Lintas terhadap Karakter Bobot Biji/Tanaman Populasi Oasis x HP1744...
17. iks Analisis Lintas terhadap Karakter Bobot Biji/Tanaman Populasi
Selayar x Rabe... 111
18. Matriks Analisis Lintas terhadap Karakter Bobot Biji/Tanaman
Populasi Dewata x Alibey... 111
19. Hasil Analisis Komponen Ragam dan Heritabilitas pada Tiga Kelompok Populasi F2...
126
20. Diferensial Seleksi untuk Beberapa Metode Seleksi pada Tiga Kelompok Populasi F2...
139
21. Keragaan Segregan Oasis x HP1744 Hasil Seleksi Berbasis Indeks dengan Intensitas Seleksi 10%...
141
22. Keragaan Segregan Selayar x Rabe Hasil Seleksi Berbasis
Indeks dengan Intensitas Seleksi 10%...
142
23. Keragaan Segregan Dewata x Alibey Hasil Seleksi Berbasis Indeks dengan Intensitas Seleksi 10%...
DAFTAR GAMBAR
No. Halaman
1. Alur Kegiatan Penelitian... 8
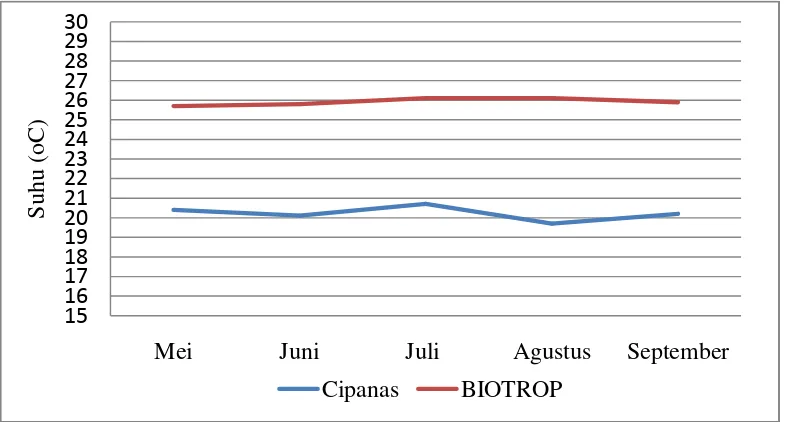
2. Rata-rata Temperatur Bulanan di Dua Agroekosistem... 32
3. Grafik BiplotGenotipe dan Karakter Berdasarkan Indeks
Sensitivitas... 42
4. Fertilitas Polen Beberapa Genotipe Gandum Berdasarkan Metode
Pewarnaan di Dua Agroekosistem... 44
5. Morfologi Polen Beberapa Genotipe Gandum... 44
6
7.1
Prosedur Persilangan Tanaman Gandum...………...
Sebaran Populasi F2 untuk Karakter Jumlah Anakan Total Hasil Persilangan Oasis x HP1744...
52
57
7.2 Sebaran Populasi F2 untuk Karakter Jumlah Anakan Total Hasil
Persilangan Selayar x Rabe... 57
7.3 Sebaran Populasi F2 untuk Karakter Jumlah Anakan Total Hasil
Persilangan Dewata x Alibey... 57
8.1 Sebaran Populasi F2 untuk Karakter Jumlah Anakan Produktif
Hasil Persilangan Oasis x HP1744... 60
8.2 Sebaran Populasi F2 untuk Karakter Jumlah Anakan Produktif
Hasil Persilangan Selayar x Rabe... 60
8.3 Sebaran Populasi F2 untuk Karakter Jumlah Anakan Produktif
Hasil Persilangan Dewata x Alibey... 60
9.1 Sebaran Populasi F2 untuk Karakter Tinggi Tanaman Hasil
Persilangan Dewata x Alibey... 62
9.2 Sebaran Populasi F2 untuk Karakter Tinggi Tanaman Hasil
Persilangan Selayar x Rabe... 62
9.3 Sebaran Populasi F2 untuk Karakter Tinggi Tanaman Hasil
10.1 Sebaran Populasi F2 untuk Karakter Kehijauan Daun Bendera
Hasil Persilangan Oasis x HP1744... 64
10.2 Sebaran Populasi F2 untuk Karakter Kehijauan Daun Bendera
Hasil Persilangan Selayar x Rabe... 64
10.3 Sebaran Populasi F2 untuk Karakter Kehijauan Daun Bendera
Hasil Persilangan Dewata x Alibey... 64
11.1 Sebaran Populasi F2 untuk Karakter Luas Daun Bendera Hasil
Persilangan Oasis x HP1744... 66
11.2 Sebaran Populasi F2 untuk Karakter Luas Daun Bendera Hasil
Persilangan Selayar x Rabe... 66
11.3 Sebaran Populasi F2 untuk Karakter Luas Daun Bendera Hasil
Persilangan Dewata x Alibey... 66
12.1 Sebaran Populasi F2 untuk Karakter Umur Berbunga Hasil
Persilangan Oasis x HP1744... 69
12.2 Sebaran Populasi F2 untuk Karakter Umur Berbunga Hasil
Persilangan Selayar x Rabe... 69
12.3 Sebaran Populasi F2 untuk Karakter Umur Berbunga Hasil
Persilangan Dewata x Alibey... 69
13.1 Sebaran Populasi F2 untuk Karakter Umur Panen Hasil
Persilangan Oasis x HP1744... 70
13.2 Sebaran Populasi F2 untuk Karakter Umur Panen Hasil
Persilangan Selayar x Rabe... 70
13.3 Sebaran Populasi F2 untuk Karakter Umur Panen Hasil
Persilangan Dewata x Alibey... 70
14.1 Sebaran Populasi F2 untuk Panjang Malai Tanaman Hasil
Persilangan Oasis x HP1744... 72
14.2 Sebaran Populasi F2 untuk Karakter Panjang Malai Tanaman
Hasil Persilangan Selayar x Rabe... 72
14.3 Sebaran Populasi F2 untuk Karakter Panjang Malai Tanaman
Hasil Persilangan Dewata x Alibey... 72
15.1 Sebaran Populasi F2 untuk Karakter Jumlah Spikelet/Malai Hasil
15.2 Sebaran Populasi F2 untuk Karakter Jumlah Spikelet/Malai Hasil
Persilangan Selayar x Rabe... 74
15.3 Sebaran Populasi F2 untuk Karakter Jumlah Spikelet/Malai Hasil
Persilangan Dewata x Alibey... 74
16.1 Sebaran Populasi F2 untuk Karakter Kerapatan Spikelet/Malai
Hasil Persilangan Oasis x HP1744... 76
16.2 Sebaran Populasi F2 untuk Karakter Kerapatan Spikelet/Malai
Hasil Persilangan Selayar x Rabe... 76
16.3 Sebaran Populasi F2 untuk Karakter Kerapatan Spikelet/Malai
Hasil Persilangan Dewata x Alibey... 76
17.1 Sebaran Populasi F2 untuk Karakter Jumlah Biji/Malai Hasil
Persilangan Oasis x HP1744... 78
17.2 Sebaran Populasi F2 untuk Karakter Jumlah Biji/Malai Hasil
Persilangan Selayar x Rabe... 78
17.3 Sebaran Populasi F2 untuk Karakter Jumlah Biji/Malai Hasil
Persilangan Dewata x Alibey... 78
18.1 Sebaran Populasi F2 untuk Karakter Jumlah Floret
Hampa/Malai Hasil Persilangan Oasis x HP1744... 80
18.2 Sebaran Populasi F2 untuk Karakter Jumlah Floret Hampa /Malai
Hasil Persilangan Selayar x Rabe... 80
18.3 Sebaran Populasi F2 untuk Karakter Jumlah Floret Hampa/Malai
Hasil Persilangan Dewata x Alibey... 80
19.1 Sebaran Populasi F2 untuk Karakter Rasio Floret Hampa/Malai
Hasil Persilangan Oasis x HP1744... 83
19.2 Sebaran Populasi F2 untuk Karakter Rasio Floret Hampa/ Malai
Hasil Persilangan Selayar x Rabe... 83
19.3 Sebaran Populasi F2 untuk Karakter Rasio Floret Hampa/Malai
Hasil Persilangan Dewata x Alibey... 83
20.1 Sebaran Populasi F2 untuk Karakter Jumlah Biji/Tanaman Hasil
Persilangan Oasis x HP1744... 85
20.2 Sebaran Populasi F2 untuk Karakter Jumlah Biji/Tanaman Hasil
20.3 Sebaran Populasi F2 untuk Karakter Jumlah Biji/Tanaman Hasil
Persilangan Dewata x Alibey... 85
21.1 Sebaran Populasi F2 untuk Karakter Bobot Kering Tajuk Hasil
Persilangan Oasis x HP1744... 87
21.2 Sebaran Populasi F2 untuk Karakter Bobot Kering Tajuk
Hasil Persilangan Selayar x Rabe... 87
21.3 Sebaran Populasi F2 untuk Karakter Bobot Kering Tajuk Hasil
Persilangan Dewata x Alibey... 87
22.1 Sebaran Populasi F2 untuk Karakter Bobot Kering Akar Hasil
Persilangan Oasis x HP1744... 88
22.2 Sebaran Populasi F2 untuk Karakter Bobot Kering Akar Hasil
Persilangan Selayar x Rabe... 88
22.3 Sebaran Populasi F2 untuk Karakter Bobot Kering Akar Hasil
Persilangan Dewata x Alibey... 88
23.1 Sebaran Populasi F2 untuk Karakter Indeks Panen Hasil
Persilangan Oasis x HP1744... 89
23.2 Sebaran Populasi F2 untuk Karakter Indeks Panen Hasil
Persilangan Selayar x Rabe... 89
23.3 Sebaran Populasi F2 untuk Karakter Indeks Panen Hasil
Persilangan Dewata x Alibey... 89
24.1 Sebaran Populasi F2 untuk Karakter Bobot Biji/Malai Hasil
Persilangan Oasis x HP1744... 91
24.2 Sebaran Populasi F2 untuk Karakter Bobot Biji/Malai Hasil
Persilangan Selayar x Rabe... 91
24.3 Sebaran Populasi F2 untuk Karakter Bobot Biji/Malai Hasil
Persilangan Dewata x Alibey... 91
25.1 Sebaran Populasi F2 untuk Karakter Bobot 50 Biji Hasil
Persilangan Oasis x HP1744... 92
25.2 Sebaran Populasi F2 untuk Karakter Bobot 50 Biji Hasil
Persilangan Selayar x Rabe... 92
25.3 Sebaran Populasi F2 untuk Karakter Bobot 50 Biji Hasil
26.1 Sebaran Populasi F2 untuk Karakter Bobot Biji/Tanaman Hasil
Persilangan Oasis x HP1744... 93
26.2 Sebaran Populasi F2 untuk Karakter Bobot Biji/Tanaman Hasil
Persilangan Selayar x Rabe... 93
26.3 Sebaran Populasi F2 untuk Karakter Bobot Biji/Tanaman Hasil
Persilangan Dewata x Alibey... 93
27.1 Diagram Lintas Karakter Bobot Biji/Tanaman Populasi Oasis x
HP1744... 112
27.2 Diagram Lintas Karakter Bobot Biji/Tanaman Populasi Selayar x
Rabe... 112
27.3 Diagram Lintas Karakter Bobot Biji/Tanaman Populasi Dewata x
Alibey... 112
28. Pertumbuhan individu-individu populasi F2 hasil persilangan
Oasis x HP1744... 119
29. Pertumbuhan individu-individu populasi F2 hasil persilangan
Selayar x Rabe... 119
30. Pertumbuhan individu-individu populasi F2 hasil persilangan
Dewata x Alibey... 119
31. Variasi Morfologi Beberapa Zuriat F2 Populasi Oasis x HP1744.. 121
32. Variasi Morfologi Malai Populasi Oasis x HP1744... 122 33. Variasi Morfologi Malai Populasi Selayar x Rabe... 122
34. Variasi Karakteristik Akar dan Tajuk Pada 3 Kelompok
Populasi F2... 123
35.1 Sebaran Segregan-segregan Oasis x HP1744 untuk Karakter
Bobot Biji/Tanaman dan Jumlah Anakan Produktif... 133
35.2 Sebaran Segregan-segregan Selayar x Rabe untuk Karakter
Bobot Biji/Tanaman dan Jumlah Anakan Produktif... 133
35.3 Sebaran Segregan-segregan Dewata x Alibey untuk Karakter
Bobot Biji/Tanaman dan Jumlah Anakan Produktif... 133
36.1 Sebaran Segregan-segregan Oasis x HP1744 untuk Karakter
36.2 Sebaran Segregan-segregan Selayar x Rabe untuk Karakter
Bobot Biji/Tanaman dan Bobot Biji/Malai... 134
36.3 Sebaran Segregan-segregan Dewata x Alibey untuk Karakter
Bobot Biji/Tanaman dan Bobot Biji/Malai... 134
37.1 Sebaran Segregan-segregan Oasis x HP1744 untuk Karakter
Bobot Biji/Tanaman dan Rasio Floret Hampa... 136
37.2 Sebaran Segregan-segregan Selayar x Rabe untuk Karakter
Bobot Biji/Tanaman dan Rasio Floret Hampa... 136
37.3 Sebaran Segregan-segregan Selayar x Rabe untuk Karakter
Bobot Biji/Tanaman dan Rasio Floret Hampa... 136
38.1 Sebaran Segregan-segregan Oasis x HP1744 untuk Karakter
Bobot Biji/Tanaman dan Umur Panen... 138
38.2 Sebaran Segregan-segregan Selayar x Rabe untuk Karakter
Bobot Biji/Tanaman dan Umur Panen... 138
38.8 Sebaran Segregan-segregan Dewata x Alibey untuk Karakter
PENDAHULUAN Latar Belakang
Pertumbuhan populasi penduduk yang terus meningkat harus diimbangi
dengan pemenuhan sumber pangan yang berkualitas. Menurut Amien (2004)
jumlah penduduk dunia diperkirakan akan meningkat dari enam milyar pada tahun
2004 menjadi 8 milyar pada tahun 2020. Di Indonesia laju pertambahan penduduk
terus meningkat. Saat ini populasi penduduk Indonesia sekitar 237 juta jiwa (BPS
2010). Upaya pemenuhan sumber pangan yang berkualitas merupakan
permasalahan dan tantangan saat ini terkait dengan permasalahan lain yaitu
penurunan luasan areal pertanaman dan perubahan kondisi agroekosistem akibat
perubahan iklim global. Serealia merupakan kelompok tanaman yang memiliki
peran penting sebagai sumber pangan. Gandum sebagai tanaman dari kelompok
serealia merupakan sumber pangan penting di dunia.
Gandum (Triticum aestivum L.) merupakan tanaman alloheksaploid yang
berasal dari daerah subtropis. Gandum memiliki peranan sebagai pendukung
ketahanan pangan dunia karena secara global tanaman ini merupakan komoditas
serealia yang paling banyak diusahakan di dunia (Sleper dan Poehlman 2006).
Sebagai sumber pangan, gandum dikonsumsi oleh sekitar dua milyar penduduk di
dunia yaitu sekitar 36% dari total penduduk dunia (Wittenberg 2004).
Di Indonesia, kebutuhan terhadap gandum cenderung meningkat setiap
tahun seiring meningkatnya populasi penduduk dan meningkatnya produk olahan
berbasis tepung terigu. Gandum digunakan sebagai bahan baku untuk produk
makanan seperti roti, mie instan, biskuit, dan juga dijual dalam bentuk tepung
terigu. Pada tahun 1984 konsumsi tepung terigu mencapai 6.18 kg/kapita/tahun,
tahun 1988 meningkat menjadi 6.59 kg/kapita/tahun, tahun 1990 9.17 kg, tahun
1999 sebesar 14.29 kg/kapita/tahun. Pada tahun 2003 konsumsi terigu 15.00
kg/kapita/tahun, tahun 2007 17.10 kg/kapita/tahun, dan diproyeksikan terus
meningkat setiap tahun (APTINDO 2012).
Kebutuhan gandum yang terus meningkat belum mampu diimbangi dengan
gandum terus meningkat. Pada tahun 2007, Indonesia mengimpor 4.770.000 ton
atau setaran dengan US$697.524.000 (APTINDO 2012). Upaya untuk pemenuhan
kebutuhan gandum dalam negeri dapat dilakukan dengan pengusahaan gandum di
Indonesia. Upaya ini dapat mengurangi dan menekan ketergantungan impor
gandum. Menurut Sastrosoemarjo et al. (2004), pengembangan areal pertanaman
gandum di Indonesia diharapkan tidak menggunakan daerah – daerah berelevasi
tinggi, karena akan bersaing dengan produksi komoditas hortikultura.
Pengembangan gandum harus diarahkan pada daerah berelevasi rendah sampai
sedang yang di Indonesia masih tersedia cukup luas, yaitu 313 502 ha
(Sastrosoemarjo et al. 2004).
Permasalahan yang dihadapi dalam upaya pengembangan gandum di
elevasi rendah dan medium yaitu perbedaan kesesuaian kondisi agroklimat dan
belum tersedianya varietas yang mampu beradaptasi baik pada daerah dengan
elevasi yang rendah dan sedang. Perbedaan kesesuaian kondisi agroklimat yang
dominan yaitu perbedaan faktor temperatur dan ketersediaan air.
Cekaman suhu tinggi menjadi salah satu faktor pembatas dalam upaya
pengusahaan gandum di daerah berelevasi rendah dan medium, karena pada
dasarnya gandum merupakan tanaman subtropis yang menghendaki suhu 10-21oC sebagai suhu optimalnya untuk proses pertumbuhan dan perkembangan (Ginkel
dan Villareal 1996). Menurut Peet dan Willits (1998) cekaman suhu tinggi sering
didefinisikan sebagai kenaikan suhu yang melebihi ambang kerusakan untuk
periode waktu yang cukup menyebabkan kerusakan yang tidak dapat balik
(irreversible) pada pertumbuhan dan perkembangan tanaman, sehingga batasan
suhu tinggi untuk tiap tanaman akan tergantung habitat asal tanaman.
Setiap tanaman memiliki respon yang berbeda dan spesifik untuk tiap tipe
cekaman. Genotipe selektif terhadap cekaman merupakan genotipe yang memiliki
mekanisme adaptasi baik fisiologi, anatomi, maupun morfologi. Suhu tinggi yang
bersifat sementara maupun konstan menyebabkan perubahan morfo-anatomis,
fisiologis, dan biokomiawi pada tumbuhan. Hal tersebut dapat mempengaruhi
pertumbuhan dan perkembangan tumbuhan serta menyebabkan berkurangnya
hasil yang bernilai ekonomi (Wahid et al. 2007). Secara umum cekaman suhu
signifikan. Barnabas et al. (2008) mengemukakan terjadinya penurunan hasil dan
kualitas hasil gandum terjadi karena ketidakseimbangan antara sink-source,
kerusakan protein yang menyebabkan gangguan selama proses pembelahan sel
mitosis, menginduksi biosintesis asam absisat yang menyebabkan penuaan dini,
terhambatnya proses fotosintesis, penutupan stomata, sterilitas polen, penurunan
viabilitas polen dan stigma yang menyebabkan kegagalan proses fertilisasi.
Sifat toleransi gandum terhadap cekaman suhu tinggi dapat diperbaiki dan
ditingkatkan melalui perakitan varietas baru dalam program pemuliaan tanaman.
Tujuan program pemuliaan tanaman gandum di Indonesia juga diarahkan untuk
merakit varietas gandum berdaya hasil tinggi selain toleran pada lingkungan
bersuhu tinggi. Pendekatan pemuliaan yang dipilih adalah pendekatan pemuliaan
yang mampu mengintegrasikan sifat toleransi terhadap lingkungan bersuhu tinggi
dan sifat daya hasil tinggi.
Pendekatan seleksi berulang di dua lingkungan berbeda (shuttle breeding)
merupakan pendekatan yang cocok untuk mencapai tujuan tersebut. Shuttle
breeding merupakan pendekatan pemuliaan yang melibatkan dua lingkungan
(optimum dan bercekaman) sebagai lingkungan seleksi. Pada lingkungan
optimum gen-gen produktivitas akan terekspresi secara optimal, sehingga
eksplorasi dan identifikasi genotipe untuk daya hasil tinggi dilakukan di
lingkungan ini. Sementara eksplorasi untuk sifat toleransi suhu tinggi dilakukan di
lingkungan bercekaman karena gen-gen pengendali sifat toleransi akan terekspresi
secara penuh pada lingkungan ini. Efisiensi dan efektifitas pendekatan shuttle
breeding dalam merakit varietas gandum berdaya hasil tinggi dan mampu
beradaptasi pada beberapa tipe lingkungan telah dilaporkan oleh Ortiz et al.
(2011).
Tim Penelitian Pemuliaan Gandum Laboratorium Genetika dan Pemuliaan
Tanaman IPB bekerja sama dengan Konsorsium Gandum Indonesia telah
mengintroduksi sejumlah varietas gandum dari beberapa negara dan
mengevaluasinya di lingkungan tropis Indonesia baik lingkungan yang berelevasi
tinggi maupun di lingkungan berelevasi rendah dan medium. Varietas-varietas
terseleksi hasil uji adaptasi akan digunakan dalam membentuk varietas baru
Tujuan Penelitian
Penelitian ini merupakan bagian dari penelitian yang dikoordinasikan oleh
Tim Konsorsium Gandum Indonesia yang bertujuan untuk memperoleh varietas
gandum yang berdaya hasil tinggi dan mampu beradaptasi baik di lingkungan
tropis Indonesia secara umum dan adaptif di lingkungan agroekosistem dataran
rendah dan medium secara khusus dalam mendukung kemandirian produksi
gandum.
Secara khusus penelitian ini bertujuan untuk : (1) mendapatkan informasi
respon genotipe gandum terhadap cekaman suhu tinggi; (2) memperoleh
informasi kendali genetik pada beberapa kelompok populasi F2 untuk karakter-karakter agronomi gandum di lingkungan berelevasi tinggi; (3) mendapatkan
informasi hubungan antar karakter agronomi dan kriteria seleksi; (4) mendapatkan
informasi parameter genetik pada populasi bersegregasi; (5) mengidentifikasi dan
mendapatkan segregan potensial berdaya hasil tinggi dan putatif toleran suhu
tinggi.
Kerangka Pemikiran
Sebagai salah satu tanaman menyerbuk sendiri, gandum memiliki
konstitusi genetik homozigot dan homogenous dalam satu populasi. Keragaman
genetik pada tanaman ini terlihat antar populasi maupun antar famili pada
populasi bersegregasi. Perbedaan latar belakang genetik pada tanaman ini akan
menyebabkan perbedaan respon terhadap cekaman suhu tinggi.
Gandum (Triticum aestivum L.) merupakan spesies alloheksaploid dengan
jumlah kromosom 2n=6x=42. Satu set kromosom gandum berjumlah 7. Tanaman
gandum (Triticum aestivum L.) tergolong tanaman menyerbuk sendiri karena
bersifat Cleistogami (pollen dan stigma terdapat dalam satu bunga dan matang
secara bersamaan sebelum bunga mekar) (Acquaah 2007). Sebagai tanaman
menyerbuk sendiri, secara alami tanaman gandum bersifat homozigot pada setiap
lokus gen, homogen dalam satu populasi, dan heterogen antar populasi. Anggapan
ini didasarkan kepada mekanisme pembentukan homozigositas pada tanaman
tanaman gandum bersifat disomik karena secara alami orientasi perpasangan
disomik dan homolog dikendalikan oleh gen Ph1 di kromosom 5 genome B
(Acquaah 2007). Adanya orientasi perpasangan disomik pada gandum,
memungkinkan pendugaan jumlah gen dan pewarisan sifatnya dapat dilakukan
dan dianalisis seperti halnya tanaman diploid.
Penelitian tentang pengaruh suhu tinggi terhadap pertumbuhan dan
perkembangan tanaman pada tanaman serealia maupun komoditas tanaman lain
telah banyak dilaporkan oleh para peneliti (Stone 2001; Maestri et al. 2002;
Jagadish et al. 2007; Wahid et al. 2007; Barnabas et al. 2008; Bukovnik et al.
2009; Hurkman et al. 2009; Thuzar et al. 2010; Xu et al. 2010; Yildirim & Bahar
2010; Sakata et al. 2010). Secara umum cekaman suhu tinggi berpengaruh
terhadap penurunan hasil dan kualitas hasil tanaman secara signifikan. Barnabas
et al. (2008) mengemukakan terjadinya penurunan hasil dan kualitas hasil gandum
terjadi karena ketidakseimbangan antara sink-source, kerusakan protein yang
menyebabkan gangguan selama proses pembelahan sel mitosis, menginduksi
biosintesis asam absisat yang menyebabkan penuaan dini, terhambatnya proses
fotosintesis, penutupan stomata, sterilitas polen, penurunan viabilitas polen dan
stigma yang menyebabkan kegagalan proses fertilisasi. Sakata et al. (2010) telah
berhasil mengidentifikasi dan melaporkan bahwa fase awal pada saat
perkembangan anther adalah fase yang paling peka terhadap suhu tinggi pada
tanaman gandum dan barley. Hasil ini selaras dengan penelitian Farrell et al.
(2006) dalam Jagadish et al. (2007) pada tanaman padi.
Beberapa mekanisme toleransi dan adaptasi gandum terhadap cekaman
suhu tinggi dikemukakan dan diusulkan oleh para peneliti (Mohammadi et al.
2007; Wahid et al. 2007; Sakata et al. 2010) . Pada dasarnya mekanisme toleransi
gandum terhadap cekaman suhu tinggi meliputi toleransi sifat morfologi, anatomi,
dan fisiologi. Belum banyak laporan mengenai bagaimana pola pewarisan dan
aksi gen karakter-karakter toleransi.
Mohammadi et al. (2007) dalam penelitiannya menyatakan bahwa
karakter bobot biji di bawah kondisi cekaman suhu tinggi merupakan karakter
yang lebih baik untuk menyaring genotipe toleran dibanding karakter – karakter
senyawa fenolik. Pola pewarisan karakter komponen hasil dan hasil gandum pada
kondisi optimal telah dilaporkan oleh Erkul et al. (2010). Terdapat pengaruh gen
aditif dan dominan serta interaksinya pada karakter panjang malai, jumlah
spikelet/malai, bobot 1000 biji, dan daya hasil. Novoselovic et al. (2004)
melaporkan terdapat pengaruh gen aditif yang nyata pada beberapa sifat
kuantitatif. Dalam penelitiannya tinggi tanaman merupakan sifat kuantitatif yang
memiliki nilai heritabilitas arti sempit yang tinggi (81%).
Genotipe – genotipe gandum yang digunakan dalam penelitian ini yaitu 2
genotipe gandum yang sudah dikembangkan di Indonesia dan 4 genotipe
introduksi yang berasal dari wilayah yang berbeda. Perbedaan wilayah asal
mengindikasikan latar belakang genetik yang berbeda. Genotipe – genotipe yang
berasal dari wilayah yang berbeda akan memiliki kekerabatan yang lebih jauh
dibanding genotipe – genotipe yang berasal dari satu wilayah. Analisis pola
pewarisan sifat dan pendugaan model genetik dapat dilakukan menggunakan dua
pendekatan yaitu analisis dialel dan analisis populasi bersegregasi (populasi F2). Pendugaan pewarisan pada populasi F2 memberikan gambaran sebaran populasi berbasis fenotipe sebagai refleksi variasi genotipe maksimum.
Rekombinasi di antara tetua selektif pada tanaman gandum merupakan
upaya memfiksasi alel-alel dominan ke dalam segregan-segregannya, sehingga
upaya ini sering menghasilkan segregan-segregan yang memiliki keragaan lebih
baik dibanding tetua-tetuanya. Menurut Sleper dan Poehlman (2006), segregasi
gen-gen yang menyebabkan mengumpulnya sejumlah alel dominan pada beberapa
segregan disebut segregasi transgresif. Individu – individu hasil segregasi
transgresif yang memiliki keragaan di luar rentang keragaan tetuanya disebut
segregan transgresif. Segregan transgresif secara teori dapat diperoleh pada
generasi selfing ke-1 (F2) dengan asumsi populasi minimumnya terpenuhi.
Identifikasi dan seleksi segregan-segregan yang memiliki potensi hasil
tinggi dapat dilakukan pada generasi awal. Perbaikan pada karakter hasil tidak
dapat dilakukan hanya pada karakter target karena karakter hasil umumnya tidak
dapat berdiri sendiri melainkan dipengaruhi oleh karakter-karakter yang lain.
Kajian dan pendugaan hubungan antar karakter hasil dengan karakter agronomi
yang akan digunakan. Karakter umur panen, jumlah anakan per tanaman, jumlah
malai per tanaman, bobot malai kering, dan bobot malai panen dilaporkan
memiliki korelasi positif yang sangat nyata dengan hasil per tanaman. Karakter
jumlah anakan per tanaman, jumlah malai per tanaman, jumlah butir per malai,
dan bobot 100 butir dilaporkan memiliki pengaruh langsung tertinggi terhadap
hasil per tanaman. Sementara itu karakter-karakter yang diduga memiliki
pengaruh tidak langsung tertinggi terhadap hasil per tanaman yaitu karakter umur
panen, bobot malai kering, dan bobot malai panen melalui karakter jumlah anakan
dan jumlah malai per tanaman (Budiarti et al. 2004). Banyaknya karakter
komponen hasil yang berkorelasi dengan hasil mengindikasikan bahwa metode
seleksi berbasis indeks merupakan salah satu metode seleksi yang cocok dan
efektif digunakan untuk menyeleksi segregan-segregan berpotensi hasil tinggi.
Acquaah (2007) menjelaskan bahwa penerapan metode seleksi berbasis
indeks menghasilkan segregan atau genotipe-genotipe berkarakter ideal untuk
karakter-karakter yang dilibatkan dalam indeks seleksi. Pembobot yang dapat
digunakan untuk indeks seleksi yaitu nilai parameter genetiknya dan nilai
ekonomis karakter. Pembobot berbasis parameter genetik digunakan dalam
kegiatan seleksi segregan harapan di generasi awal, sementara itu pembobot
berbasis nilai ekonomis karakter digunakan dalam kegiatan seleksi di generasi
lanjut atau pada saat uji adaptasi dan stabilitas.
Perakitan varietas gandum berdaya hasil tinggi dan adaptif di lingkungan
tropis terutama dataran rendah merupakan upaya yang saat ini sedang dilakukan
untuk mengurangi ketergantungan terhadap gandum impor. Dalam penelitian ini
dilakukan serangkaian percobaan untuk memperbaiki potensi genetik plasma
nutfah gandum. Percobaan pertama yaitu mengevaluasi, karakterisasi, dan
observasi respon genotipe terhadap perbedaan kondisi agroekosistem terutama
difokuskan kepada perbedaan faktor temperatur. Percobaan kedua dan selanjutnya
diarahkan untuk menghasilkan segregan-segregan terbaik melalui kegiatan
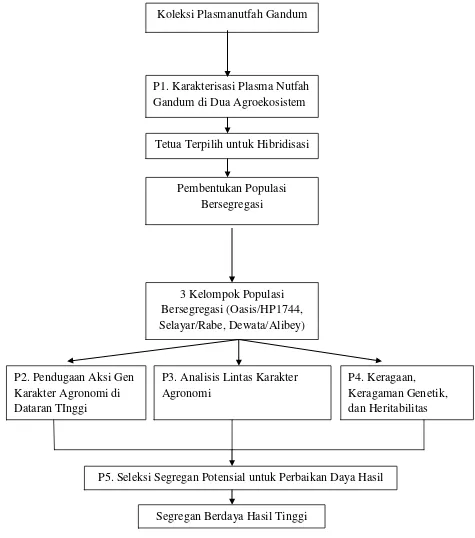
hibridisasi antar varietas selektif. Alur kegiatan penelitian yang dilakukan
Gambar 1. Alur Kegiatan Penelitian
P1. Karakterisasi Plasma Nutfah Gandum di Dua Agroekosistem
Tetua Terpilih untuk Hibridisasi
Pembentukan Populasi Bersegregasi
3 Kelompok Populasi Bersegregasi (Oasis/HP1744, Selayar/Rabe, Dewata/Alibey)
P2. Pendugaan Aksi Gen Karakter Agronomi di Dataran TInggi
P3. Analisis Lintas Karakter Agronomi
P4. Keragaan, Keragaman Genetik, dan Heritabilitas Karakter Agronomi
P5. Seleksi Segregan Potensial untuk Perbaikan Daya Hasil
Hipotesis
Hipotesis yang dikemukakan pada penelitian ini adalah : (1) terdapat
perbedaan respon diantara plasma nutfah gandum di dua agroekosistem ; (2)
Karakter-karakter agronomi pada tiga kelompok populasi F2 dikendalikan secara poligenik dengan aksi gen aditif; (3) Terdapat keragaman genetik yang luas di
populasi bersegregasi dengan nilai heritabilitas dari sedang sampai tinggi; (4) Ada
beberapa karakter agronomi yang memiliki pengaruh langsung yang tinggi
terhadap karakter hasil; (5) Ada beberapa genotipe terseleksi yang memiliki
TINJAUAN PUSTAKA Asal Usul Tanaman gandum
Gandum pertama kali ditanam di Asia Tengah di kawasan yang kini
dikenal sebagai lembah subur (Fertile-crescent). Bukti tertua bagi penanaman
gandum berasal dari Syria, Jordan, Turki, Armenia dan Irak (Wikipedia 2011).
Sleper dan Poehlman (2006) menjelaskan bahwa asal usul genetik gandum
merupakan contoh klasik bagaimana spesies – spesies liar dapat saling
berekombinasi di alam untuk menghasilkan suatu bentuk poliploidi alami.
Gandum (Triticum aestivum) merupakan spesies yang berasal dari genus Triticum,
Tribe Triticeae, dan Famili Poaceae. Triticeae merupakan Tribe dari famili
Poaceae yang terdiri lebih dari 15 genus dan 300 spesies yang termasuk gandum
dan barley. Genus Triticum berkerabat dengan Hordeum, Avena, Secale, Zea, dan
Oryza (Wittenberg 2004).
Secara taksonomi gandum pertama kali diklasifikasikan pada tahun 1753
oleh Linnaeus. Spesies – spesies yang termasuk di dalam genus Triticum
dikelompokkan ke dalam tiga kelas ploidi yaitu diploid, tetraploid dan heksaploid
(Sakamura 1918 dalam Wittenberg 2004; Fehr 1987; Sleper dan Poehlman 2006).
Beberapa spesies yang termasuk ke dalam genus Triticum diploid, tetraploid, dan
heksaploid disajikan pada Tabel 1. Sleper dan Poehlman (2006) melaporkan
bahwa saat ini terdapat 11 spesies diploid, 11 spesies tetraploid, dan 6 spesies
heksaploid yang sudah diidentifikasi dan dideskripsikan. Lebih lanjut Sleper dan
Poehlman mengemukakan hanya dua spesies dari genus Triticum yang memiliki
nilai ekonomis penting yaitu Triticum aestivum dan Triticum turgidum. Triticum
aestivum merupakan gandum yang umum dikenal yang dimanfaatkan untuk bahan
baku roti. Triticum turgidum yang dikenal dengan gandum durum digunakan
untuk membuat pasta. Wilson (1955) mengklasifikasikan gandum berdasarkan
kegunaannya yang meliputi gandum keras, gandum lunak, dan gandum durum.
Gandum keras (hard wheat) adalah gandum yang memiliki kandungan gluten dan
protein tinggi. Tipe ini cocok untuk pembuatan roti. Gandum lunak (soft wheat)
adalah gandum yang memiliki kandungan gluten dan protein yang lebih rendah.
Gandum durum yaitu gandum yang memiliki kandungan gluten dan protein sangat
Tabel 1. Klasifikasi Beberapa Spesies Triticum Berdasarkan Kelas Ploidi
Species Genome Status
Diploid Species (2n = 14)
T. Monoccocum var. monoccocum AA Budidaya
T. Monoccocum var. boeoticum AA spesies liar
T. Dichasians CC spesies liar
T. Tauschii DD spesies liar
T. Comosum MM spesies liar
T. Speltoides SS spesies liar
T. Umbellatum UU spesies liar
Spesies Tetraploid (2n = 4x = 28)
T. turgidum L. var. dococcon AABB Budidaya
T. turgidum L. var. durum AABB Budidaya
T. turgidum L. var. turgidum AABB Budidaya
T. turgidum L. var. polonicum AABB Budidaya
T. turgidum L. var. carthlicum AABB Budidaya T. turgidum L. var. dicoccoides AABB spesies liar T. timopheevii var. araraticum AAGG spesies liar
T. cylindricum DDCC spesies liar
T. ventricosum DDMM spesies liar
T. triunciale UUCC spesies liar
T. ovatum UUMM spesies liar
T. kotschyi UUSS spesies liar
Spesies Heksaploid (2n = 6x = 42)
T. aestivum L. var. aestivum AABBDD Budidaya
T. aestivum L. var. spelta AABBDD Budidaya
T. aestivum L. var. compactum AABBDD Budidaya T. aestivum L. var. sphaerococcum AABBDD Budidaya
T. syriacum DDMMSS spesies liar
T. juvenale DDMMUU spesies liar
T. triaristatum UUMMMM spesies liar
Sumber : Fehr (1987)
Aspek Morfologi dan Agronomi Gandum
Tanaman gandum yang normal memiliki dua macam akar, yaitu akar
kecambah dan akar adventif. Akar kecambah merupakan akar pertama yang
tumbuh dari embrio, sedangkan akar adventif adalah akar yang berkembang dari
buku dasar tumbuh setelah akar embrio. Sistem perakaran tanaman gandum
dibentuk oleh akar adventif. Sistem perakaran dengan perakaran serabut dan
kedalaman perakaran gandum sekitar 10-30 cm di bawah permukaan tanah.
Batang gandum tegak, berbentuk silinder, dan membentuk tunas. Ruas-ruasnya
pendek dan buku-bukunya umumnya berongga. Rata-rata tanaman dewasa
memiliki enam ruas buku. Anakan primer dari buku batang utama terus
rumpun. Tinggi tanaman gandum bervariasi tergantung genotipe dan lingkungan
tumbuh. Daun pertama yang tumbuh disebut koleoptil berongga dan berbentuk
silinder, diselaputi plumula yang terdiri dari dua sampai tiga helai daun. Setiap
daun gandum terdiri dari tangkai pelepah, helai daun, dan ligula dengan dua
pasang telinga pada dasar helai daun. Tulang daun sejajar dan memanjang
(Nurmala 1980).
Bunga gandum tersusun dalam rangkaian bunga berbentuk malai (spike)
terdiri dari beberapa bunga (spikelet). Umumnya 1 spikelet terdiri dari 3 floret (2
floret primer dibagian kanan dan kiri dan 1 floret sekunder dibagian tengah
spikelet). Sebuah floret disusun oleh lemma dan palea yang menutupi biji. Secara
botani biji gandum disebut caryopsis. Floret gandum ada yang berbulu (lemma
dan paleanya memanjang dan bentuknya meruncing) dan tidak. Biji gandum
bervariasi warnanya yaitu merah, ungu, coklat, dan putih (Acquaah 2007).
Secara alami tanaman gandum menyerbuk sendiri karena berbunga
sempurna. Waktu anthesis dan reseptis terjadi secara bersamaan, namun stigma
dapat reseptif lebih awal. Umumnya bunga-bunga yang berada di bagian tengah
rangkaian bunga yang anthesis dan reseptis terlebih dahulu kemudian bunga
bagian atas dan bawah. Malai gandum umumnya keluar sempurna (heading stage)
pada temperatur 13-250C. Pertumbuhan tabung pollen sekitar 15-60 menit setelah penyerbukan terjadi atau pollen menempel di stigma. Periode pengisian biji
umumnya sekitar 14-21 hari setelah terjadi fertilisasi (Acquaah 2007).
Faktor temperatur dan curah hujan merupakan faktor dominan yang
menentukan pertumbuhan dan perkembangan tanaman gandum. Tanaman gandum
beradaptasi sangat baik pada lingkungan bertemperatur rendah dengan temperatur
optimalnya sekitar 10-210C dengan curah hujan tidak lebih dari 40-60 cm/tahun (Acquaah 2007). Di Indonesia kondisi lingkungan tersebut berada di wilayah
agroekosistem berelevasi tinggi.
Respon Tanaman Terhadap Cekaman Suhu Tinggi
Pertumbuhan dan perkembangan tanaman sebagai visualisasi ekspresi gen
Secara umum faktor lingkungan yang mempengaruhi pertumbuhan tanaman yaitu
faktor biotik dan faktor abiotik. Faktor temperatur merupakan faktor lingkungan
abiotik. Cekaman suhu tinggi merupakan salah satu faktor pembatas dalam usaha
produksi tanaman.
Cekaman suhu tinggi sering diartikan sebagai peningkatan suhu yang
melebihi level ambang batas selama periode waktu tertentu sehingga
menyebabkan kerusakan bagi pertumbuhan dan perkembangan tumbuhan. Secara
umum, peningkatan suhu sementara, biasanya 10–15 °C di atas suhu udara di
sekitar, dapat dianggap sebagai cekaman panas (heat shock). Cekaman panas
merupakan sebuah fungsi yang kompleks dari intensitas (derajat suhu), durasi, dan
kecepatan peningkatan suhu. Tingkat kemunculan (cekaman) pada zona iklim
tertentu bergantung pada probabilitas dan periode dari suhu tinggi yang terjadi
selama siang hari dan atau malam hari. Toleransi terhadap panas secara umum
diartikan sebagai kemampuan tumbuhan untuk tumbuh dan memproduksi hasil
(yang bersifat) ekonomis pada kondisi suhu yang tinggi. Beberapa peneliti
meyakini bahwa suhu di malam hari merupakan faktor pembatas utama,
sementara yang lain berpendapat bahwa suhu siang dan malam hari tidak
mempengaruhi tumbuhan secara mandiri, dan suhu rata – rata diurnal merupakan
prediktor yang lebih baik untuk respons tumbuhan terhadap suhu tinggi di mana
suhu siang hari berperan sebagai faktor sekunder (Peet dan willits 1998).
Cekaman panas akibat suhu udara yang tinggi merupakan ancaman yang
serius bagi produksi tanaman di seluruh dunia. Emisi gas yang diakibatkan
aktifitas manusia secara substansial menambah konsentrasi gas – gas rumah kaca,
terutama CO2, metana, klorofluorokarbon, dan oksida – oksida nitrus. Sirkulasi global yang berbeda memprediksi bahwa gas – gas rumah kaca secara bertahap
akan meningkatkan rata – rata suhu udara dunia. Menurut laporan International
Panel on Climatic Change (IPCC), rata – rata suhu global akan meningkat 0,3 °C
setiap dekadenya (Jones et al. 1999), secara berturut – turut akan mencapai kurang
lebih 1 dan 3 °C di atas nilai suhu sekarang pada tahun 2025 dan 2100, dan
mengarah pada pemanasan global. Peningkatan suhu dapat mengakibatkan
perubahan persebaran geografis dan musim tanam komoditas pertanian dengan
kemasakan tanaman yang lebih awal (Porter 2005). Secara umum perubahan yang
terjadi pada tanaman yang tercekam suhu tinggi dikelompokkan menjadi beberapa
tipe yaitu perubahan morfologis, anatomis, fenologis, dan fisiologis (Wahid et al.
2007).
Suhu tinggi dapat menyebabkan perubahan morfologis pada tanaman yang
berupa kerusakan pra dan pasca panen, termasuk luka bakar pada daun dan
ranting, hangus (terbakar sinar matahari) pada daun, cabang dan batang, absisi dan
penuaan daun, terhambatnya pertumbuhan akar dan pucuk, kerusakan dan
kehilangan warna pada buah, serta berkurangnya hasil. Selain itu, suhu tinggi
menyebabkan penurunan yang signifikan untuk bobot tajuk, kecepatan relatif
pertumbuhan, dan kecepatan asimilasi pada jagung, pearl millet, dan tebu (Wahid
et al. 2007).
Cekaman panas, baik itu secara mandiri maupun berkombinasi dengan
kekeringan, merupakan hambatan utama selama proses antesis dan pengisian biji
pada berbagai tanaman serealia pada wilayah bersuhu sedang. Sebagai contoh,
cekaman panas mempercepat durasi pengisian biji dengan adanya reduksi pada
pertumbuhan biji, yang mengarah pada hilangnya kepadatan dan bobot biji
hinggamencapai 7% pada gandum musim semi (Guilioni et al. 2003). Penurunan
sejenis juga terjadi untuk kandungan pati, protein, dan minyak pada biji jagung
dan penurunan kualitas pada tanaman serealia lainnya dalam kondisi tercekam
panas (Maestri et al. 2002). Pada gandum, bobot dan jumlah biji nampak sangat
sensitif terhadap cekaman panas, di mana jumlah biji per bulir pada saat masak
berkurang seiring dengan meningkatnya suhu (Ferris et al. 1998). Pada tomat,
proses – proses reproduktif sangat terpengaruh secara negatif oleh suhu tinggi,
dimana ternasuk di dalamnya adalah pembelahan meiosis pada organ jantan dan
betina, perkecambahan polen dan pertumbuhan tabung polen, jumlah butir polen
yang disimpan dalam stigma, proses fertilisasi dan paska fertilisasi, pertumbuhan
endosperma, pra embrio dan embrio yang sudah terfertilisasi. Pengaruh negatif
yang paling jelas akibat suhu tinggi tehadap proses reproduktif pada tomat adalah
dihasilkannya stilus yang terdesak sehingga dapat menghambat penyerbukan
dengan rendahnya kadar karbohidrat dan dihasilkannya pengatur pertumbuhan
pada jaringan sink tumbuhan (Kinet dan Peet 1997 dalam Wahid et al 2007).
Perubahan anatomis pada tanaman terhadap cekaman suhu tinggi meliputi
berkurangnya ukuran sel, penutupan stomata dan terbatasnya kehilangan air,
meningkatnya kepadatan stomata dan trikorma, pembesaran pembuluh xilem pada
akar dan tajuk (Anon et al. 2004). Pada tanaman anggur (Vitis vinifera), cekaman
panas dapat merusak sel – sel mesofil dan meningkatkan permeabilitas membran
plasma (Zhang et al. 2005). Pada kondisi suhu tinggi, tanaman Zygophillum
qatarense akan menghasilkan daun – daun polimorfik dan cenderung mereduksi
kehilangan air transpirasi dengan menunjukkan perilaku stomata bimodal. Pada
level subseluler, modifikasi mayor terjadi pada kloroplas, dan menyebabkan
perubahan dalam fotosintesis. Sebagai contoh, suhu tinggi mereduksi fotosintesis
dengan cara mengubah organisasi struktural tilakoid (Karim et al. 1997).
Perubahan fisiologis tanaman terhadap cekaman suhu tinggi meliputi
berkurangnya ketersediaan air, akumulasi senyawa – senyawa organik tertentu
yang secara umum sering disebut sebagai osmolit – osmolit kompatibel, degradasi
klorofil a dan b pada daun yang sedang berkembang (Karim et al., 1997).
Mekanisme Adaptasi dan Toleransi Gandum Terhadap Cekaman Suhu Tinggi
Secara alami, tumbuhan memiliki mekanisme adaptasi terhadap setiap
perubahan lingkungan baik biotik maupun abiotik. Dalam kaitannya dengan
adaptasi, terdapat dua kriteria adaptasi yaitu adaptasi biologis dan adaptasi
agronomis. Adaptasi biologis yaitu kemampuan tanaman untuk tetap
mempertahankan hidupnya dan menghasilkan keturunannya dengan memodifikasi
seluruh karakter baik morfologi, anatomi, fenologi, maupun anatomi. Adaptasi ini
umumnya menghasilkan tanaman yang dapat tetap tumbuh pada lingkungan
bercekaman namun berproduksi rendah. Adaptasi agronomis merupakan
mekanisme adaptasi tanaman yang berkaitan dengan hasil tanaman. Tanaman
yang adaptif agronomis merupakan tanaman yang dapat menjaga stabilitas
Terkait dengan mekanisme adaptasi tanaman terhadap cekaman suhu
tinggi, Wahid et. al. (2007) mengungkapkan bahwa terdapat dua mekanisme
adaptasi (1) mekanisme penghindaran (avoidance); (2) mekanisme toleransi.
Berumur genjah merupakan salah satu bentuk mekanisme penghindaran terhadap
cekaman suhu tinggi. Mekanisme ini akan efektif jika cekaman suhu tinggi hanya
terjadi beberapa saat atau pada fase tertentu saja. Mekanisme adaptasi ini dapat
memberikan konsekuensi negatif bagi produksi tanaman, karena menurut Wahid
et. al. (2007) karakter umur genjah umumnya berkorelasi dengan penurunan daya
hasil. Genotipe berumur genjah umumnya berdaya hasil rendah. Selain umur
berbunga, perilaku pembungaan tanaman juga menjadi salah satu mekanisme
penghindaran. Genotipe-genotipe tanaman yang menunjukkan mekanisme ini
mampu berbunga di luar waktu pembungaan yang normal. Seperti padi yang
memiliki mekanisme ini mampu berbunga dini hari untuk mencegah gangguan
mikrogametogenesis dan megagametogenesis serta fertilisasi pada kondisi
cekaman suhu tinggi.
Karena tanaman bersifat immobil secara fisik, Wahid et. al. (2007)
menjelaskan bahwa umumnya mekanisme toleransi tanaman terhadap cekaman
suhu tinggi berupa mekanisme fisiologi yang berupa (1) peningkatan kandungan
antioksidan tanaman untuk mencegah terjadinya penuaan dini (senescence) karena
pengaruh oksigen reaktif; (2) mekanisme stabilitas suhu membran untuk
mencegah kerusakan fungsi membran; (3) peningkatan akumulasi protein-protein
yang teraktivasi di suhu tinggi (heat shock protein); (4) mekanisme renaturasi
protein; (5) mekanisme osmoprotektan.
Pemuliaan Gandum Terhadap Cekaman Abiotik
Tujuan utama pemuliaan gandum yaitu perbaikan karakter daya hasil,
stabilitas hasil, dan peningkatan kualitas hasil (Fehr 1987; Sleper dan Poehlman
2006). Karakter hasil merupakan karakter penting karena kunci dalam
peningkatan produksi untuk pemenuhan sumber pangan bagi manusia. Karakter
stabilitas hasil mencakup karakter – karakter toleransi terhadap cekaman abiotik
dan biotik. Upaya merakit kultivar tahan cekaman abiotik dan biotik diperlukan
lingkungan. Karakter kualitas hasil merupakan karakter lain yang penting karena
wujud dari optimalisasi produksi yang dicapai.
Cekaman abiotik merupakan salah satu faktor pembatas baik dalam
kegiatan ekstensifikasi maupun intensifikasi tanaman. Secara mendasar
lingkungan bercekaman didefinisikan sebagai lingkungan suboptimum untuk
pertumbuhan dan produksi tanaman (Wirnas 2007). Upaya perbaikan daya hasil
dan adaptasi tanaman terhadap cekaman abiotik dapat dilakukan melalui
serangkaian program pemuliaan tanaman.
Keberhasilan perakitan kultivar baru toleran cekaman abiotik ditentukan
oleh beberapa faktor yang meliputi ketersediaan sumber genetik dan variabilitas
genetiknya yang luas, kemampuan dalam mengidentifikasi genotipe toleran dan
pemilihan metode seleksi yang cepat, tepat dan efisien, serta pembentukan
populasi bersegregasi dengan rancangan persilangan yang tepat. Baihaki (2000)
memaparkan faktor – faktor penentu keberhasilan suatu program pemuliaan yang
mencakup pemilihan pasangan – pasangan tetua, metode persilangan, jumlah
kombinasi persilangan pada generasi F1, dan jumlah tanaman yang ditumbuhkan
pada generasi F2.
Pengembangan dan peningkatan keragaman genetik sebagai langkah awal
untuk merakit kultivar toleran cekaman abiotik dapat dilakukan melalui
introduksi, domestikasi, hibridisasi, induksi mutasi, dan rekayasa genetik.
Introduksi dan domestikasi merupakan upaya untuk meningkatkan keragaman
genetik dengan memanfaatkan variasi yang telah tersedia di alam. Hibridisasi
merupakan upaya penggabungan dua sifat baik yang terdapat pada dua tanaman
menjadi satu tanaman. Mutasi merupakan upaya peningkatan keragaman genetik
dengan memanfaatkan mutagen fisik dan kimia sebagai agen penginduksi mutasi.
Induksi mutasi dapat menghasilkan alel baru sehingga karakter fenotipe dapat
dihasilkan. Rekayasa genetik merupakan pendekatan untuk peningkatan
keragaman genetik berbasis gen pada level seluler.
Identifikasi dan penyaringan awal genotipe – genotipe toleran akan
menentukan keberhasilan program pemuliaan selanjutnya. Berdasarkan
lingkungan seleksi, seleksi dan identifikasi genotipe toleran dapat dilakukan
breeding) dan seleksi tidak langsung dilingkungan optimal (indirect breeding).
Menurut Ceccareli et al. (2007) dalam Wirnas (2007) seleksi untuk perbaikan
toleransi cekaman harus dilakukan di lingkungan target sehingga dapat
memaksimalkan ekspresi gen – gen yang mengendalikan daya adaptasi tanaman.
Keberhasilan seleksi dilingkungan target ditentukan oleh pemilihan
karakter seleksi. Seleksi pada lingkungan bercekaman umumnya tidak
menggunakan seleksi langsung atau seleksi berdasarkan hasil, karena pada
lingkungan bercekaman sangat sulit memisahkan variabel – variabel lingkungan
yang dapat menurunkan hasil. Seleksi tidak langsung menjadi pilihan untuk
melakukan seleksi pada lingkungan bercekaman. Seleksi tidak langsung
didasarkan kepada karakter toleransi yang berkontribusi atau memiliki pengaruh
langsung yang tinggi untuk daya hasil dan memiliki heritabilitas yang tinggi untuk
mendukung kemajuan genetik yang akan dicapai. Karakter – karakter toleransi
pada tanaman dapat meliputi karakter morfologi, karakter anatomi, karakter
fisiologi, karakter molekuler, dan karakter komponen hasil.
Penggabungan karakter - karakter toleransi ke dalam satu genotipe
tanaman dapat dilakukan melalui persilangan. Fehr (1987) mengemukakan bahwa
pada umumnya kultivar baru gandum dihasilkan melalui hibridisasi buatan.
Hibridisasi gandum secara buatan pertama kali dilakukan sekitar tahun 1890.
Lebih lanjut Fehr (1987) menyebutkan di Amerika, A.E. Blount, W.J. Spillman,
dan L.F. Waldron adalah orang – orang yang pertama melakukan hibridisasi
buatan dalam merakit kultivar baru gandum. Kultivar – kultivar yang telah mereka
hasilkan yaitu „Gypsum‟, „Hybrid 128‟, dan „Ceres‟.
Terdapat beberapa metode persilangan yang telah dikembangkan dan
digunakan oleh para pemulia tanaman yaitu persilangan top cross, line x tester,
single cross, double cross, suksesif, dan dialel (Baihaki 2000; Singh dan
Chaudhary 1979). Persilangan dialel merupakan metode persilangan yang banyak
digunakan dan memiliki beberapa kelebihan (Yunianti 2007). Persilangan dialel
merupakan metode persilangan yang memungkinkan seluruh kombinasi
persilangan diantara sekelompok genotipe tetua yang dilibatkan. Persilangan