IKA
MARIANA
SEKO INSTIT
AN MAS Cy
A YERMIN
OLAH PAS TUT PERT
BOG 201
yprinus carp
NA BERUA
SCASARJA ANIAN BO GOR
12
rpio
ATJAAN
Dengan ini saya menyatakan bahwa tesis Evaluasi Penurunan Serat Kasar dan Peningkatan Nilai Kecernaan Bungkil Kelapa Dengan Penambahan Enzim Cairan Rumen Domba Sebagai Bahan Baku Pakan Ikan Mas Cyprinus carpio adalah karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka dibagian terakhir tesis ini.
Bogor, Juni 2012
MARIANA YERMINA. The Evaluation Of Sheep Rumen Liquor Enzymes On Crude Fiber Reduction End Digestibility Enhancement Of Coconut Cake Meal For Carp Cyprinus carpio. Under direction of DEDI JUSADI, NUR BAMBANG PRIYO UTOMO
The objectives of this work were to evaluate the roles of sheep rumen liquor enzymes on its capability to reduce the crude fiber contant and enhance the apperent digestibility of coconut cake meal (BK) for common carp Cyprinus carpio. The work comprised by two stages experiments. In experiment I, BK was hydrolized with 0, 25, 50, 75, 100, or 125 ml enzymes/kg BK for 0, 12, and 24 hours, respectively. Thereafter, proximate compositions of BK were analyzed. In experiment II, hydrolyzed BK (BKe) contained the least crude fiber content from the result of experiment I was tested for its digestibility. It was found that BK hydrolyzed with 125 ml enzymes/kg BK for 24 hours had the lowest value of crude fiber content, and highest value of glucose content. In this treatment, crude fiber content was decreased from 14,34% to 6,98%, while the glucose content was increased from 0,014% to 0,464%. The apparent digestibility of BKe was found to be higher than BK. The total digestibility, protein carp was 63,92% and 54,58%, 79,53% and 65,17%, 72,2% and 49,17% for BKe and BK, respectively. It was concluded that sheep rumen liquor enzymes could reduce the crude fiber content of BK, and enhance its apparent digestibility for carp Cyprinus carpio.
Kecernaan Bungkil Kelapa Dengan Penambahan Enzim Cairan Rumen Domba Sebagai Bahan Baku Pakan Ikan Mas Cyprinus carpio. Dibimbing oleh DEDI JUSADI, NUR BAMBANG PRIYO UTOMO.
Penelitian ini bertujuan mengevaluasi pengaruh penambahan enzim cairan rumen domba terhadap penurunan kandungan serat kasar bungkil kelapa sebagai bahan baku pakan ikan mas, serta mengevaluasi nilai kecernaan bahan baku pakan berbasis bungkil kelapa yang telah dihidrolisis dengan enzim cairan rumen domba pada ikan mas. Penambahan enzim cairan rumen domba dalam bungkil kelapa diharapkan menurunkan kandungan serat kasar sehingga mampu meningkatkan nilai kecernaan dalam bahan pakan dan dapat dimanfaatkan secara optimal oleh ikan. Penelitian dilakukan dalam dua tahap yaitu, tahap I menguji efektivitas enzim cairan rumen domba volume 0, 25, 50, 75, 100 atau 125 ml/kg selama 0, 12 atau 24 jam dalam menurunkan kandungan serat kasar bungkil kelapa. Rancangan percobaan yang digunakan adalah rancangan acak lengkap faktorial. Tahap II adalah menguji kecernaan bahan pakan bungkil kelapa yang dihidrolisis dengan enzim cairan rumen domba (BKe) yang merupakan hasil terbaik dari uji tahap pertama. Uji tahap II menggunakan rancangan acak lengkap dengan 3 perlakuan pakan dan 3 ulangan yang ditambahkan indikator Cr2O3. Pakan perlakuan dalam penelitian ini adalah pakan acuan (100% komersil), pakan uji A : 30% Bungkil kelapa dengan penambahan enzim cairan rumen domba (BKe) dan pakan B : 30% bungkil kelapa tanpa penambahan enzim (BK). Ikan yang digunakan adalah 25 ekor ikan mas bobot 1,0 – 1,3 g yang dipelihara dalam akuarium dengan volume 50 liter air. Pemberian pakan secara et satiation dengan frekuensi pemberian 3 kali per hari. Feses dikumpulkan selama 20 hari pemeliharaan untuk dianalisis kandungan nutrisinya. Analisis proksimat dilakukan pada pakan dan feses untuk diukur nilai kecernaannya.
Hasil penelitian menunjukan bahwa penambahan enzim cairan rumen domba volume 125 ml/kg bahan dengan lama waktu inkubasi 24 jam dapat menurunkan kandungan serat kasar bungkil kelapa terbaik yaitu dari 14,34% menjadi 6,98%. Penurunan kandungan serat kasar dapat meningkatkan nilai kecernaan total bahan pakan bungkil kelapa dari 54,58% menjadi 63,92%, kecernaan protein dari 65,17% menjadi 79,53% dan kecernaan energi dari 49,17% menjadi 72,2%.
@ Hak Cipta milik IPB, tahun 2012 Hak Cipta dilindungi Undang Undang
1. Dilarang mengutip sebahagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkn sumber.
a. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah.
b. Pengutipan tersebut tidak merugikan kepentingan yang wajar IPB.
IKAN MAS Cyprinus carpio
MARIANA YERMINA BERUATJAAN
Tesis
Sebagai salah satu syarat untuk memperoleh gelar Magister sains pada
Program Studi Ilmu Akuakultur
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
Cairan Rumen Domba Sebagai Bahan Baku Pakan Ikan Mas Cyprinus carpio
Nama : Mariana Yermina Beruatjaan
NRP : C151090101
Disetujui Komisi Pembimbing
Dr. Dedi Jusadi Dr. Nur Bambang Priyo Utomo
Ketua Anggota
Diketahui
Ketua Program Studi Dekan Sekolah Pascasarjana
Ilmu Akuakultur
Prof. Dr. Enang Harris Dr. Dahrul Syah
berkat dan anugerahNya sehingga tesis ini dapat diselesaikan. Penyelesaian Tesis ini dengan penuh perjuangan dan tidak terlepas dari bantuan berbagai pihak. Perkenankan penulis mengucapkan terima kasih yang tak tehingga kepada :
1. Komisi pembimbing, Bapak Dr. Dedi Jusadi dan Bapak Dr. Nur Bambang Priyo Utomo, yang telah banyak menuangkan pikiran, meluangkan waktu untuk membimbing, mengarahkan, dan memberikan saran hingga terselesaikannya tesis ini.
2. Dekan Pascasarjana IPB dan jajarannya yang memberikan kesempatan bagi penulis untk belajar pada Sekolah Pascasarjana IPB serta memberikan pelayanan akademis kepada penulis.
3. Direktur Jenderal Pendidikan Tinggi (Dirjen Dikti) yang telah memberikan biaya pendidikan melalui Sekolah Pascasarjana IPB kepada penulis.
4. Direktur Politeknik Perikanan Negeri Tual yang telah memberikan Izin bagi penulis untuk melanjutkan studi pada jenjang pendidikan magister (S2). 5. Ketua Mayor Ilmu Akuakultur, Staf Dosen, Kesekretariatan, Teknisi dan
Laboran FPIK atas pelayanan, pengabdian dan perhatiannya.
6. Suami tercinta Dr. Jeffry Prabowo yang setia menunggu dan menopang baik Doa, moril maupun materiil.
7. Orang Tua tercinta Bpk. Perez Beruat, Mama Mia dan Mami Ida W, atas Doa, dan motivasi setiap saat.
8. Adik-adik tersayang Yomi, Jhein, dan Asri, Alin, Ona, Tati
9. Keluarga besar Beruatjaan/Warin, Oma Yomi, Opa Andi, Kel. Besar Wiyatno, Om Nunug, Kel.Om Fredy Metengun, Kel. Om Jems Miru, Mama Yos, dan Kel. Balubun, Milka Ngurmetan, trima kasih menyumbangkan tenaga dan waktu sejak pagi sampai malam selama penelitan berlangsung.
Semua pihak yang tidak sempat disebutkan, terima kasih atas Doa dan semangat yang diberikan semoga Tuhan Yesus memberkati. Harapan penulis kiranya karya ini bermanfaat.
Bogor, Juni 2012
xi
DAFTAR TABEL………... xii
DAFTAR GAMBAR………... xiii
DAFTAR LAMPIRAN……….... xiv
PENDAHULUAN………... 1
Latar Belakang ……….. ... 1
Perumusan Masalah………... 3
Tujuan Penelitian ……….... 4
Manfaat Penelitian……….. 4
Hipotesis ………. 4
TINJAUAN PUSTAKA………... 5
Kebutuhan Nutrisi Ikan Mas………... 5
Bungkil Kelapa ………... 9
Cairan Rumen Sebagai Sumber Enzim ……….. 10
Serat Kasar dalam Bahan Pakan ………. 14
Kecernaan Pakan………. 15
BAHAN DAN METODE PENELITIAN………... 17
Penelitian Tahap 1. Uji Evektivitas Enzim Cairan Rumen Domba.... 17
Penelitian Tahap 2. Uji Kecernaan Bungkil Kelapa... 18
HASIL DAN PEMBAHASAN………... 22
Hasil………. 22
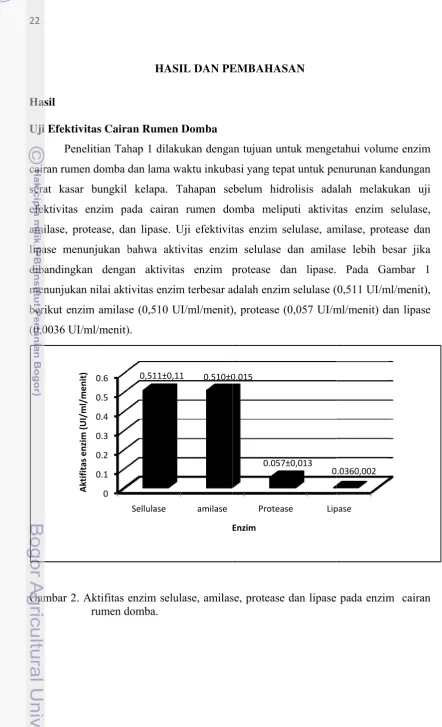
Uji Efektivitas Cairan Rumen Domba..………... 22
Analisis Kandungan Nutrisi Bungkil Kelapa……...……... 23
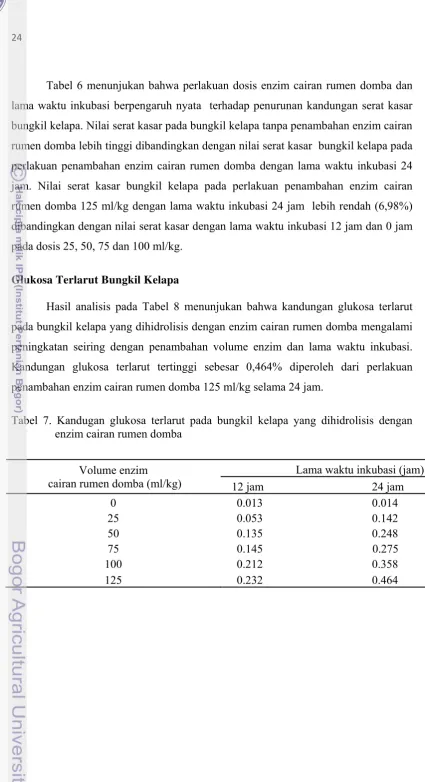
Glukosa Terlarut Bungkil Kelapa……...…………... 24
Uji Kecernaan Bungkil Kelapa yang Telah dihidrolisis dengan Enzim Cairan Rumen Domba... 25
Pembahasan………... 26
KESIMPULAN………... 31
DAFTAR PUSTAKA………... 32
xii
Halaman
1. Kebutuhan Makro Nutrisi Ikan Mas (Cyprinus carpio)... 6
2. Kandungan Nilai Nutrien Bungkil Kelapa (%)... 9
3. Komposisi Enzim Cairan Rumen Domba ………... 12
4. Komposisi Pakan Acuan, Pakan Uji A (30% BKe) dan B (30% BK)….. 19
5. Kisaran Nilai Parameter Kualitas Air Media Pemeliharaan Ikan Mas
Selama Penelitian... 19
6. Kandungan Lemak, Protein, Serat Kasar Bungkil Kelapa pada
Perlakuan Penambahan Enzim Caiaran Rumen Domba dan Lama Waktu Inkubasi ... 23
7. Kandungan Glukosa Terlarut pada Bungkil Kelapa yang Dihidrolisis
Dengan Enzim Cairan Rumen Domba... 24
8. Kecernaan Total, Kecernaan Protein, dan Kecernaan Energi... 25
xiii
Halaman
1. Bagian-Bagian Perut Hewan Ruminansia... 10
2. Aktifitas Enzim Selulase, Amilase, Protease dan Lipase pada
xiv
Halaman
1. Perosedur Analisis Kadar Protein..………... 37
2. Perosedur Analisis Kadar Lemak...………. 38
3. Perosedur Analisis Kadar Air... 39
4. Perosedur Analisis Kadar Abu... 40
5. Perosedur Analisis Kadar Serat Kasar... 41
6. Metode Uji Aktifitas Enzim... 43
7. Perosedur Analisis Kecernaan... 46
PENDAHULUAN
Latar Belakang
Sebagian besar bahan baku pakan, seperti tepung ikan, tepung kedelai,
jagung, meat bone meal, dan bahan lainnya merupakan bahan impor. Tepung ikan
yang diimpor Indonesia pada tahun 2008 sebesar 44.073,22 ton, tahun 2009
sebesar 47.518,97 ton dan pada tahun 2010 menjadi 39.261,69 ton. Sedangkan
tepung kedelai yang diimpor tahun 2008 sebesar 35.849,98 ton, tahun 2009
menjadi 53.474,80 ton, tahun 2010 meningkat menjadi 55.805,86 (Anonim 2011).
Sebagai upaya pengurangan ketergantungan terhadap bahan baku pakan yang
diimpor tersebut adalah pemanfaatan bahan baku lokal berbasis limbah yang
berpotensi dijadikan sebagai bahan baku pakan ikan, diantaranya adalah bungkil
kelapa.
Bungkil kelapa merupakan limbah agroindustri yang ketersediaannya
didukung dengan potensi kelapa di Indonesia yang mencapai luasan areal 3,9 juta
ha tahun 2009 dengan produksi 3,3 juta ton/tahun (Anonim 2010). Produksi
kelapa di Indonesia terus meningkat. Pada Tahun 2010, beberapa daerah di
Indonesia yang memiliki potensi kelapa yang cukup tinggi seperti Kepulauan
Riau memproduksi kelapa sebesar 49,4018 ton/tahun, Kalimantan Selatan (30,869
ton/tahun), Sulawesi Tenggara (41,675 ton/tahun), Maluku (75,217 ton/tahun)
(Anonim 2011). Di Bireun, Provinsi Nangroe Aceh Darussalam (NAD) memasok
buah kelapa setiap hari mencapai 35 ton, sedangkan bungkil kelapa yang
merupakan limbah dari hasil olahan tersebut mencapai ±7 ton (Anonim 2010).
Limbah bungkil kelapa yang dihasilkan dijual dengan harga Rp. 1.500/kg. Limbah
ini dijadikan sebagai pakan ternak. Harga tersebut jika dibandingkan dengan
bahan baku pakan lainnya seperti bungkil kedelai Rp. 4.400/kg, tepung jagung
Rp. 3.050/kg, pollard Rp. 2.400/kg, maka harga bungkil kelapa relatif murah.
Namun, bungkil kelapa yang digunakan sebagai alternatif bahan baku pakan
diduga sulit dicerna karena mengandung serat kasar yang tinggi. Bungkil kelapa
hasil akhir olahan minyak kelapa tersebut mengandung protein kasar 23,13%,
lemak 10,86%, serat kasar 11,64%, abu 7,21%, kadar air 2,51%, dan BETN
(NSP) terkandung dalam bungkil kelapa sehingga sulit dicerna. Salah satu upaya
yang dapat dilakukan untuk menurunkan kandungan serat kasar dalam bungkil
kelapa tersebut adalah penambahan enzim pendegradasi yang terdapat dalam
cairan rumen domba.
Rumen merupakan sumber enzim pendegradasi polisakarida. Polisakarida
dihidrolisis di rumen disebabkan pengaruh sinergis dan interaksi dari kompleks
mikroorganisme, terutama penghasil selulase dan xillanase (Trinci et al., 1994).
Mikroba-mikroba rumen mensekresikan enzim-enzim pencernaan ke dalam cairan
rumen untuk membantu mendegradasi partikel makanan. Enzim-enzim tersebut
antara lain adalah enzim selulase, hemiselulase/xylanase, amilase, pektinase,
lipase, protease dan lain-lain (Kamra, 2005). Selanjutnya dinyatakan bahwa
mikroba rumen yang mensekresikan enzim selulase diantaranya Fibrobacter
succinogenes, Ruminococcus albus, R. flavefaciens, Clostrodium lochheadii, C.
longisporum dan Eubacterium cellulosovens. Sedangkan jenis mikroba yang
banyak menghasilkan enzim amlase adalah Streptococcus bovis, Ruminococcus
amylophylus, Prevotella rumonocola, Streptococcus ruminantium dan
Lachnosphora multipharius. Hasil penelitian Pamungkas (2011) melaporkan
bahwa penambahan enzim cairan rumen domba dosis 100 ml/kg bahan dengan
lama waktu inkubasi 24 jam dapat menurunkan kandungan serat kasar bungkil
kelapa sawit dari 17,54% menjadi 6,69% dan menaikan nilai kecernaan dari
15,31% menjadi 42,26% pakan benih ikan patin siam. Penelitian Fitriliyani
(2010) menghasilkan penurunan kadar serat kasar tepung daun lamtoro tertinggi
sebesar 16,77% menjadi 7,77% dan menurunkan asam fitat 68,09% serta
meningkatkan komposisi asam amino dan nilai kecernaan pada ikan nila. Pantaya
(2003), penelitian penambahan enzim cairan rumen domba dengan dosis 1,24
IU/kg menurunkan kandungan polisakarida dari 26,32% ke 22,38%,
meningkatkan oligosakarida dari 73,68% ke 77,62% dan meningkatkan energi
wheat pollard dari 1,55 kkal/kg menjadi 1,88 kkal/kg. Penelitian Sandi (2010),
penambahan enzim cairan rumen domba dosis 1% (b/v) dengan lama waktu
inkubasi 24 jam mampu menurunkan kandungan serat kasar sebesar 17,83% dan
meningkatkan gula total terlarut sebesar 29,91% pada singkong. Terjadi
selama masa pemeliharaan dikondisikan banyak mengkonsumsi rumput. Lubis
(1992) melaporkan bahwa rumput mengandung serat kasar yang tinggi (30,86%).
Menurut Budiansyah (2010), sapi lokal yang banyak mendapatkan pakan serat
akan menghasilkan enzim selulase yang mampu mendegradasi serat. Sedangkan
pada sapi impor yang mendapatkan konsentrat banyak menghasilkan enzim
xilanase, mananase, amilase.
Bungkil kelapa sebagai alternatif bahan baku pakan ikan belum
dimanfaatkan secara optimal. Untuk itu, perlu dilakukan penelitian untuk
mengevaluasi penurunan serat kasar dan peningkatan nilai kecernaan bungkil
kelapa dengan penambahan enzim cairan rumen domba sebagai pakan benih ikan
mas Cyprinus carpio.
Perumusan Masalah
Ketersediaan bungkil kelapa sebagai alternatif bahan baku pakan impor
cukup banyak dan harganya relatif murah. Namun, tingginya kandungan serat
kasar (11,64%) menjadi faktor pembatas dalam penggunaannya sebagai pakan
ikan. Keterbatasan ikan dalam memanfaatkan serat berkaitan dengan ketersediaan
enzim sellulotik yang terbatas dalam saluran pencernaan ikan, bahkan pada level
tertentu dapat menghambat pertumbuhan ikan. Ikan tidak mampu mencerna serat
kasar, dibatasi oleh kemampuan mikroflora dalam ususnya untuk mensekresikan
sellulase (Bureau, 2000).
Upaya untuk meminimalkan kandungan serat kasar bungkil kelapa perlu
dilakukan, yakni dengan menghidrolisis kandungan serat kasarnya. Untuk
menghidrolisis serat kasar bisa dilakukan dengan kapang maupun menggunakan
enzim yang dihasilkan mikroba. Penelitian Indariyanti (2011), penggunaan
kapang Trichoderma harzianum Rifai untuk menurunkan serat kasar bungkil inti
sawit dari 16,78% ke 9,35% membutuhkan waktu 8 hari. Sedangkan dengan
cairan enzim rumen domba hanya membutuhkan 24 jam untuk menurunkan
kandungan serat kasar daun lamtoro, bungkil kelapa sawit dan kulit kakao,
masing-masing dari 16,77% ke 7,74%; 17,54% ke 6,69% dan 27,97% ke 21,67%
(Fitriliyani 2010, Pamungkas 2011, Kurniansyah, 2012). Selain efisiensi waktu,
menurunkan kandungan serat dalam jumlah yang lebih banyak dibanding kapang.
Penggunaan rumen domba sebagai enzim pendegradasi serat kasar pada bungkil
kelapa dibanding ternak lainnya karena keberadaan ternak domba ditemukan
hampir diseluruh daerah di Indonesia sehingga diperoleh dengan mudah.
Berdasarkan hal-hal tersebut, penelitian ini dirancang untuk menurunkan
serat kasar bungkil kelapa dengan enzim cairan rumen domba. Penambahan enzim
cairan rumen domba dalam bungkil kelapa tersebut diharapkan dapat
mengevaluasi efektivitas enzim cairan rumen domba yang ditambahkan dalam
bahan pakan serta mampu meningkatkan nilai kecernaan dalam pakan sehingga
dapat dimanfanfaatkan secara optimal oleh ikan mas.
Tujuan Penelitian
1. Mengevaluasi pengaruh penambahan enzim cairan rumen domba terhadap
penurunan kandungan serat kasar bungkil kelapa.
2. Mengevaluasi nilai kecernaan bungkil kelapa sebagai bahan baku pakan ikan
mas.
Manfaat Penelitian
Manfaat dari penelitian ini diharapkan memberikan informasi tentang
ketersediaan pakan bungkil kelapa sebagai bahan baku alternatif dari bahan baku
lokal yang berkulitas dalam pakan ikan mas.
Hipotesis
1. Penambahan enzim cairan rumen domba pada bungkil kelapa dapat
mengurangi kandungan serat kasar bahan pakan. Semakin banyak enzim yang
digunakan dalam waktu tertentu, akan semakin banyak serat kasar yang
dihidrolisis
2. Bungkil kelapa yang telah dihidrolisis dengan enzim cairan rumen domba
TINJAUAN PUSTAKA
Kebutuhan Nutrisi Ikan Mas
Fungsi utama makanan adalah sebagai penyedia energi bagi aktivitas
sel-sel tubuh. Karbohidrat, lemak dan protein merupakan zat gizi dalam makanan yang
berfungsi sebagai energi tubuh. Protein bersama dengan mineral dan air merupakan
bahan baku utama dalam pembentukan sel-sel dan jaringan tubuh, sedangkan protein
bersama-sama dengan mineral dan vitamin berfungsi dalam pengaturan
keseimbangan asam basa, pengaturan tekanan osmotik cairan tubuh, serta pengaturan
proses metabolisme dalam tubuh. Adapun lemak dalam bentuk fosfolipid dan
kolesterol juga sedikit berperan dalam pembentukan dinding sel (NRC, 1993).
Ikan, seperti juga hewan lainnya tidak mempunyai kebutuhan nutrisi yang
pasti, namun ikan membutuhkan nutrisi yang seimbang untuk keberlangsungan
hidupnya. Afrianto dan Liviawati (2005) mengemukakan bahwa kebutuhan nutrisi
untuk tiap species ikan berbeda-beda dan sering berubah-ubah dipengaruhi oleh
beberapa faktor seperti jenis ikan, ukuran, lingkungan dan musim. Protein merupakan
komponen utama jaringan dan juga senyawa nitrogen lainnya seperti asam nukleat,
enzim, hormon, dan vitamin, sehingga keberadaannya harus secara terus-menerus
disuplai dari makanan untuk pertumbuhan dan perbaikan jaringan tubuh (Furuichi,
1988). Protein mempunyai peran penting untuk fungsi jaringan yang normal,
pertahanan dan petumbuhan (Watanabe and Cho,1988). Protein dalam fungsinya
tidak hanya sebagai penyusun utama tubuh ikan tetapi juga berperan penting sebagai
enzim dan hormon-hormon yang menunjang metabolisme.
Kebutuhan protein ikan menurut (Hepher, 1990) pada umumnya berkisar
35-50%, kebutuhan protein ikan karnivora 40-50% dan omnivora 25-35% (Craig and
Helfrich, 2002). Pemanfaatan protein sangat beragam diantara spesies ikan,
bergantung pada sumber energi non-protein pakan karena kemampuan ikan dalam
memanfaatkan lemak atau karbohidrat pakan juga berbeda untuk setiap speses ikan.
Protein pakan yang tidak mencukupi akan menghambat pertumbuhan, sedangkan
energi sehingga protein yang digunakan untuk membangun jaringan tubuh hanya
sedikit (NRC, 1983). Dalam Standar Nasional Indonesia dinyatakan bahwa
kebutuhan protein minimal untuk benih ikan mas adalah 30% (SNI 2000).
Takeuchi et al. 2002, mengemukakan bahwa ikan mas mampu mencerna lemak dengan baik. Oleh karena itu, jumlah energi yang dapat tercerna (digestible energy) lebih penting dari pada jumlah lemak dalam pakan. Lemak merupakan sumber energi yang tinggi dalam pakan. Watanabe (1988) mengemukakan satu gram
energi memiliki energi dalam pakan (gross energy) sebesar 9,4 Kkal, sedangkan dalam protein dan karbohidrat sebesar 5,6 dan 4,1 Kkal. Takeuchi (1988), kebutuhan
karbohidrat ikan mas tergolong tinggi dibandingkan dengan ikan yang lain karena
ikan tersebut tergolong omnivora. Jobling (1993) dalam Midlen & Redding (1998), mengemukakakn bahwa ikan mas dapat mencerna sebagian besar karbohidrat dalam
pakan, sementara golongan karnivora seperti salmon dan yellowtail hanya mampu mencerna sekitar 25% saja. Secara umum kebutuhan makro nutrisi ikan mas disajikan
pada Tabel 1. (Takeuchi 1988).
Tabel 1. Kebutuhan makro nutrisi ikan mas (Cyprinus carpio) Makro Nutrisi Kebutuhan
Protein 30 - 35 g.100 g-1
Lemak 5 - 15 g 100 g-1
Energi 13 - 15 MJ kg-1 (310-360 Kcal)
Karbohidrat 30 - 40 g. 100 g-1
Energi pakan diperoleh dari karbohidrat, lemak dan protein. Proporsi
energi yang dikonsumsi meningkat, dengan meningkatnya ukuran ikan, namun
efisiensi pencernaan dan absorpsi menurun yang akhirnya memperlambat
pertumbuhan akibat energi yang hilang melalui feses meningkat (De Silva dan
Anderson, 1995). Imbangan protein dan energi sangat penting dalam menunjang
pertumbuhan ikan. Pakan yang mempunyai kadar protein tinggi belum tentu dapat
terlebih dahulu dipakai untuk kegiatan metabolisme standar, seperti respirasi,
transport ion, dan pengeluaran cairan tubuh serta aktifitas lainnya. Energi untuk
seluruh aktifitas tersebut diharapkan sebagian besar berasal dari bahan nutrien
non-protein, dalam hal ini karbohidrat dan lemak. Apabila sumbangan energi dari bahan
non-protein ini rendah maka protein akan digunakan sebagai sumber energi untuk
berbagai aktifitas tersebut sehingga pertumbuhan akan berkurang. Dengan kata lain,
penambahan energi non-protein dapat meningkatkan fungsi protein dalam menunjang
pertumbuhan ikan (Furuichi, 1938). Rasio energi protein optimum telah ditemukan
pada berbagai spesies ikan, dan rasio tersebut berkisar antara 8 sampai 10 kkal DE
per gram protein pakan (Halver, 2002). Sedangkan pada catfish rasio ini berkisar antara 7,4-l2 kkal/g. Peningkatan rasio DE/P pakan catfish diatas kisaran ini akan meningkatkan deposit lemak dan jika energi terlalu rendah, pertumbuhan ikan akan
melambat (Robinson et a1, 2007).
Peran utama karbohidrat dalam nutrisi hewan adalah sebagai sumber energi
(Takeuchi, l988). Menurut Furuichi (1988) karbohidrat adalah sumber energi yang
paling murah dan mudah didapatkan untuk komposisi pakan ikan dan juga bertindak
sebagai protein sparing efect. Sumber karbohidrat yang berkualitas baik menjadi sangat penting karena akan berfungsi sebagai energi nonprotein, sehingga akan
sedikit protein yang digunakan sebagai sumber energi tetapi lebih banyak digunakan
untuk pertumbuhan. Kemampuan menggunakan karbohidrat sebagai sumber energi
berbeda diantara spesies ikan. Yamamoto et al., (200l), menyatakan bahwa ikan umumnya lebih efisien dalam mencerna dan memanfaatkan protein dan lemak, tetapi
dalam memanfaatkan karbohidrat sangat bervariasi bergantung pada kompleksitas
karbohidrat. Menurut Mokoginta et al. (1999), hal tersebut disebabkan oleh aktivitas enzim amylase yang berbeda untuk spesies ikan, dan biasanya ikan karnivor lebih
terbatas dalam memanfaatkan karbohidrat dibandingkan ikan omnivor dan herbivor.
Kebanyakan ikan perairan tropis, termasuk catfish dapat memanfaatkan lebih banyak karbohidrat dibandingkan ikan perairan dingin dan ikan laut. Ikan omnivora
umumnya mampu memanfaatkan karbohidrat lebih tinggi (kadar optimum 30-40%)
(Furuichi, 1999). Ikan yang diberi pakan tanpa karbohidrat memiliki laju
pertumbuhan yang relatif lebih rendah jika dibandingkan dengan pakan yang diberi
karbohidrat (Wilson, 1994). Namun pemberian karbohidrat yang terlalu tinggi akan
mengakibatkan pertumbuhan ikan menurun dan tidak efektifnya pakan yang
diberikan (Zonneveld et al., 199l). Pertumbuhan fingerling catfish lebih tinggi ketika pakannya mengandung karbohidrat dibandingkan hanya mengandung lemak sebagai
sumber energi nonprotein (NRC, 1993).
Lemak pada pakan mempunyai peranan penting bagi ikan, karena berfungsi
sebagai sumber energi dan asam lemak esensial, memelihara bentuk dan fungsi
membran atau jaringan sel yang penting bagi organ tubuh tertentu, membantu dalam
penyerapan vitamin yang larut dalam lemak dan untuk mempertahankan daya apung
tubuh. Menurut Craig dan Helfrich (2002), lemak adalah salah satu makronutrien
dengan kandungan energi yang tinggi yang dapat dimanfaatkan sebagai protein sparing effect dalam pakan budidaya. Satu unit lemak yang sama mengandung energi dua kali lipat dibandingkan dengan protein dan karbohidrat. Jika lemak dapat
menyediakan energi untuk pemeliharaan metabolisme maka sebagian besar protein
yang dikonsumsi dapat digunakan tubuh untuk pertumbuhan dan bukan digunakan
sebagai sumber energi (NRC, 1993).
Ikan menggunakan lemak untuk energi, komponen struktur sel dan
pemeliharaan integritas biomembran (Takeuchi, l988). Furuichi (1998) juga
menyatakan bahwa lemak juga dapat dimanfaatkan untuk membangun struktur sel
dan mempertahankan integritas membran melalui penggunaan fosfolipid. Lemak
adalah sumber energi dan mengandung 2,25 kali energi karbohidrat, dan memegang
peranan penting dalam metabolisme hewan seperti mensuplai asam lemak esensial,
Bungkil Kelapa
Bungkil kelapa adalah hasil ikutan yang diperoleh dari ekstraksi daging
buah kelapa segar/kering. Didaerah tropik, bungkil kelapa adalah salah satu bahan
makanan ternak sumber protein nabati yang sering digunakan sebagai bahan
penyusun pakan (Wahju, 1988). Parakkasi (1990) mengemukakan bahwa bahan
pakan bungkil mengandung bahan protein nabati dan sangat potensial untuk
meningkatkan karkas. Kandungan nilai nutrien bungkil kelapa disajikan pada Tabel 2.
Tabel 2. Kandungan nilai nutrien bungkil kelapa (%)
Kandungan Zat Jumlah (%)
Kadar Air
Sumber : a. Hoffman, A (1981) c. Mepba dan Achinewhu (2003) b. Creswell dan Brooks (1971) d. Moorthy,M dan Viswanathan (2009)
e. Hasil analisis proksimat bungkil kelapa dari Aceh
Hasil penelitian Ng dan Chen (2004) menunjukkan bahwa bungkil kelapa
sawit (BKS) yang difermentasi oleh Trichoderma koningii menghasilkan peningkatan kandungan protein kasar, yaitu dari 17% menjadi 32%. Penggunaan BKS sebagai
pakan ikan lele telah dilakukan oleh Ng dan Chen (2002) membuktikan bahwa
pernberian bungkil sawit sebanyak 20% dalam pakan tidak berpengaruh negatif
terhadap pertumbuhan ikan lele. Penelitian yang dilakukan oleh Lim et al., (2001) pada ikan nila tilapia menunjukkan bahwa penggunaan BKS 30% dalam pakan
memberikan pertumbuhan yang tidak berbeda nyata dengan ikan yang diberi pakan
sebagai sumber protein walaupun tingkat kecernaan proteinnya lebih rendah dari
pakan kontrol. Ng dan Chong (2002), dalam hasil penelitiannya melaporkan bahwa
penggunaan BKS 20% bobot kering dalam pakan ikan nila Tilapia (Oreochromis sp) tidak menunjukkan pengaruh negatif terhadap laju pertumbuhan dan efisiensi pakan
bila dibandingkan dengan pakan kontrol yang menggunakan tepung ikan 2l,19% dan
tepung bungkil kedele 30,73% sebagai sumber protein. Selain itu juga dilaporkan
bahwa penambahan enzim pada BKS mampu meningkatkan nilai nutrisi BKS dan
dapat meningkatkan penggunaan BKS dalam pakan ikan nilia sebesar 4,0% dapat
memberikan pertumbuhan yang lebih baik dari penggunaan BKS tanpa enzim.
Cairan Rumen sebagai Sumber Enzim
Ternak ruminansia mempunyai organ pencernaan yang berbeda dengan
organ pencernaan monogastrik. Ternak ruminansia terdapat empat lambung yang
terdiri atas retikulum, rumen, omasum dan abomasum (Gambar 1). Proses pencernaan
bahan makanan yang terjadi dalam rumen adalah proses fermentasi oleh mikroba
rumen. Proses fermentasi ini yang menjadikan perbedaan antara ruminansia dan
monogastrik.
Gambar 1. Bagian-bagian perut hewan ruminansia.
Organ pencernaan ternak monogastrik yang berfungsi untuk mencerna
tidak dapat mencerna serat yang terlalu banyak karena tidak terdapat mikroba dalam
organ pencernaannya yang menghasilkan enzim pendegradasi selulosa. Pada dasarnya
hewan ruminansia juga tidak mampu memecah ikatan β1-4 glikosida, akan tetapi karena adanya mikroba di dalam rumen, maka ruminansia dapat memecah ikatan β 1-4 glikosidik (Arora, 1989).
Wizna et al., (2008) menyatakan bahwa xanthophyll yang terdapat dalam isi rumen, sebagian besar terdiri dari hijauan, diduga dibutuhkan oleh pigmen kuning
telur dan kulit unggas. Manfaat lain dari penggunaan isi rumen sebagai bahan pakan
adalah adanya vitamin Bl2 sebagai faktor protein hewan yang menyebabkan isi
rumen mempunyai nilai secara biologi yang sama dengan tepung ikan dan ekstrak
hati. Kandungan nutrisi dari isi rumen sapi adalah 9,29% air, 8,45% protein kasar,
1,23% lemak kasar, 33,53% serat kasar. 0,20% Ca, 0,45% P, l6,l9% abu, dan,
31,60% NFE. Menurut Hardiyanto, (2001) isi rumen berpotensi sebagai feed additive. Cairan rumen telah digunakan sebagai sumber inokulan dalam pengolahan silase
jerami padi. Lebih lanjut dinyatakan bahwa cairan rumen pada onggok sebagai bahan
baku penyusun ransum komplit dapat meningkatkan kandungan VFA (Volatile Fatty Acid).
Rumen diakui sebagai sumber enzim pendegradasi polisakarida.
Polisakarida dihidrolisis di rumen disebabkan pengaruh sinergis dan interaksi dari
kompleks mikroorganisme, terutama selulase dan xillanase (Trinci et al., 1994). Ada dua grup jenis mikroorganisme yang diyakini pada cairan rumen (Iiquid phase) dan yang menempel pada digesta rumen. Enzim yang aktif mendegradasi struktural
polisakarida hijauan kebanyakan aktif pada mikroorganisme yang menempel pada
partikel pakan.
Mikroba-mikroba rumen mensekresikan enzim-enzim pencernaan ke dalam
cairan rumen untuk mernbantu mendegradasi partikel makanan. Enzim-enzim
tersebut antara lain adalah enzim yang mendegradasi substrat selulosa yaitu selulosa,
hemiselulosa/xylosa adalah hemiselulase/xylanase, pati adalah amilase, pektin adalah
pektinase, lipid/lemak adalah lipase, protein adalah protease dan lain-lain (Kamra,
mensekresikan enzim selulase diantaranya Fibrobacter succinogenes, Ruminococcus albus, R. flavefaciens, Clostrodium lochheadii, C. longisporum dan Eubacterium cellulosovens. Sedangkan jenis mikroba yang banyak menghasilkan enzim amilase adalah Streptococcus bovis, Ruminococcus amylophylus, Prevotella rumonocola, Streptococcus ruminantium dan Lachnosphora multipharius.
Aktivitas enzim dalam cairan rumen juga tergantung dari komposisi atau
perlakuan makanan (Moharrery dan Das, 2002). Agarwal et al., (2002) melaporkan bahwa anak domba dengan berat badan 23,5 kg yang diberi makan minum susu
sampai 8 minggu dan diteruskan dengan 50% konsentrat dan 50% rumput sampai
umur 24 minggu mendapatkan bahwa enzim-enzim yang ada dalam cairan rumen
antara lain carboxymethyl cellulase dengan aktivitas enzim 3,60 mol glukosa per jam
per ml, alpha amylase 0,33 umol glukosa per menit per ml, xylanase 0,29 umol
xylosa per menit per ml, beta-glukosidase 0,20 umol p-nitrophenol per menit per ml,
alpha-glukosidase 0,008 U/mol p-nitrophenol per menit per ml, urease 0,05 U/mol
NHs-N per menit per ml dan protease 452,7 U/ g hidrolisis protein per jam per ml.
Moharrery dan Das (2001) mengukur aktivitas enzim protease, selulase, amylase,
lipase dan urease pada cairan rumen domba dan mendapatkan bahwa cairan rumen
yang berisi enzim-enzim dari sel-sel bakteri, aktivitas enzimnya lebih tinggi dari
cairan rumen tanpa protozoa dan tanpa sel-sel bakteri. Komposisi enzim cairan rumen
domba ditunjukan pada Tabel 3.
Tabel 3. Komposisi enzim cairan rumen domba
Enzim1) Cairan rumen
738,5±3,45 162,2±33,70 405,30±44,19
Protease (unit/ml) 0,201±0,078 0,090±0,027 0,220±0,046 Amilase (ug glukosa/menit/ml 172,2±45,9 60,05±10,96 208,7±97,0 Lipase (unit/ml) 1,076±0,309 0,339±0,080 1,225±0,803
1)
Cairan rumen telah digunakan sebagai sumber inokulan dalam pengolahan
silase jerami padi dan menghasilkan penurunan bahan kering 10,6%, kadar serat
15,98% serta meningkatkan kandungan protein 4,5% (Purnomohadi, 2006). Hasil
penelitian Hardiyanto (2001), menyatakan bahwa cairan rumen yang ditambahkan
pada onggok sebagai bahan pakan penyusun ransum komplit dapat meningkatkan
kandungan VFA (volatile fatty acid). Penambahan cairan rumen sebesar 62 dan 1,240 U/g pada wheat pollard menghasilkan penurunan kandungan polisakarida
berturut-turut sebesar 4,0% dan 3,9% dan kandungan polisakarida wheat pollard tanpa enzim lebih tinggi dibandingkan dengan yang ditarnbahkan enzim (Pantaya, 2003).
Selulosa menurut Hardjo et al. (1989) adalah polimer tak bercabang dari glukosa yang dihubungkan melalui ikatan 1,4 β glikosida. Enzim yang mendegradasi selulosa yaitu endoglukonase atau karboksil metal selulase (endo-1,4- β-glukonase). Kompleks enzim selulase mempunyai tiga komponen utama yang bekerja
bersama-sama atau bertahap dalam menguraikan selulosa menjadi unit glukosa, yaitu :
1. Endo-selulase yang memotong bagian dalam struktur kristal dari selulosa dan
mengeluarkan unit selulosa dari rantai polisakarida.
2. Ekso-selulase yang memotong 2-4 unit selulosa dari rantai akhir hasil produksi
endo-selulase dan menghasilkan tetrasakarida atau disakarida seperti selubiosa.
3. Selubiosa atau β-glukosidase yang menghidrolisis produk dari ekso-selulase menjadi monosakarida.
Tiga reaksi tersebut yang dikatalis oleh selulase memotong interaksi
nonkovalen dalam bentuk ikatan hidrogen yang ada dalam struktur kristal selulosa
oleh enzim endo-selulase, menghidrolisis serat selulosa menjadi sakarida yang lebih
sederhana oleh ekso-selulase, serta menghidrolisis disakarida dan tetrasakarida
menjadi glukosa oleh enzim β-glukosidase. Penelitian Malathi dan Devegowda (2002), menambahkan multienzim ke dalam pakan broiler memperoleh hasil terjadi
peningkatan nilai total gula sunflower meal, soybean meal, deoiled rice bran yang lebih besar dibandingkan dengan penggunaan enzim tunggal. Dinyatakan bahwa
kandungan multienzim ini juga menjadi kelebihan yang dimiliki oleh ekstrak enzim
Serat Kasar Dalam Bahan Pakan
Sundu dan Dingle (2003) mengemukakan bahwa penggunaan bungkil
kelapa sawit dalam ransum unggas dibatasi oleh tiga faktor; pertama secara fisik
bungkil kelapa bersifat “gritty (berbatu / mengandung grit) dan tidak palatable; kedua
secara nutrisional mengandung bahan atau zat seperti manan atau galaktomanan dan
xilan atau arabinoxilan yang dapat menurunkan penyerapan nutrient. Lebih lanjut
dikatakan bahwa dari total karbohidrat bungkil kelapa, 26 persen adalah manan, 61
persen galaktomanan, dan 13 persen selulosa.
Kadar serat kasar yang berbeda pada bahan penyusun pakan dapat
mempengaruhi nilai energi yang tersedia dalam pakan karena terdapat korelasi
negatif antara kadar serat kasar dalam pakan dengan energi yang tersedia dalam
pakan. Semakin tinggi serat kasar pakan maka semakin rendah jumlah energi yang
tersedia. Hal tersebut disebabkan serat kasar tidak mampu menyediakan energi yang
dapat dimanfaatkan oleh ikan. Anggorodi (1990) mengemukakan bahwa, serat kasar
yang berisi selulosa, hemiselulosa, dan lignin relatif sulit dicerna dan merupakan
sumber energi yang rendah.
Tillman (1999) menyatakan bahwa serat kasar adalah penyusun utama
dinding sel tumbuhan dan merupakan fraksi karbohidrat yang telah dipisahkan
dengan bahan ekstrak tanpa nitrogen (BETN) yang tidak larut dalam basa dan asam
encer setelah pendidihan selama 30 menit. Serat kasar terdiri dari sellulosa,
hemisellulosa dan lignin yang sulit dicerna (Anggorodi l994; Tillman l999). Serat
kasar dibutuhkan dalam pakan untuk membantu proses pencernaan makanan.
Menurut Piliang (2006), serat kasar mernbantu mempercepat ekskresi sisa-sisa
makanan rnelalui saluran pencernaan. Dalam keadaan tanpa serat, feses dan
kandungan air rendah akan lebih lama tinggal dalam saluran usus yang dapat
rnenyebabkan gangguan pada gerakan peristaltik pada usus besar sehingga eksresi
feses menjadi lebih lamban. Sebaliknya, (Slae and Hinz 1969 dalam Fitriliyani, 2010) bahwa pakan dengan kandungan serat kasar yang tinggi dapat menyebabkan absorbs
Keterbatasan ikan dalam memanfaatkan serat berkaitan dengan
ketersediaan enzim sellulotik yang terbatas dalam saluran pencernaan ikan, bahkan
pada level tertentu dapat menghambat pertumbuhan ikan (Bureau, 1999). Labih lanjut
dinyatakan bahwa, kemampuan ikan dalam mencerna serat kasar dibatasi oleh
kemampuan mikroflora dalam ususnya untuk mensekresikan sellulase.
Kecernaan Pakan
Nilai kecernaan suatu makanan atau disebut juga dengan koefisien
penecernaan (digestibility) disamping menggambarkan kemampuan ikan dalam memanfaatkan makanan juga dapat menggambarkan kualitas pakan yang dikonsumsi
oleh ikan. Lovell (1989) mendefenisikan kecernaan sebagai bagian dari pakan yang
diserap oleh hewan. Pakan yang masuk ke dalam saluran pencernaan akan dicerna
menjadi senyawa sederhana berukuran mikro, dimana protein dihidrolisis menjadi
asam-asam amino atau peptida sederhana, lemak menjadi gliserol dan asam lemak
menjadi gula sederhana (Halver 2002).
Proses kecernaan pakan baik fisik maupun kimia memegang peranan
penting. Hidrolisis nutrient makro dimungkinkan dengan adanya beberapa enzim
pencernaan seperti protease, karboksilase, dan lipase (Zonneveld et al. 1991). Robinson, (2001) mengemukakan bahwa rendahnya serat kasar dalam pakan
menyebabkan tingginya daya cerna dan penyerapan zat-zat makanan didalam alat
pencernaan ikan. Selama pakan berada dalam usus ikan, nutrient dicerna oleh
berbagai enzim menjadi bentuk yang dapat diserap oleh dinding usus dan masuk
dalam sistim peredaran darah. Sebaliknya pakan yang mengandung serat kasar tinggi
akan menghasilkan feses yang lebih banyak sehingga serat kasar yang tidak tercerna
tersebut dapat membawa zat-zat makanan yang seharusnya dicerna.
Metode pengukuran kecernaan menurut Takeuchi (1988) ada dua cara yaitu
metode langsung dan tidak langsung. Metode langsung yang diukur yaitu jumlah
pakan yang dikonsumsi dan jumlah feses yang dikeluarkan. Sedangkan metode tidak
langsung yaitu dengan menambahkan indikator dalam pakan dimana indikator
dapat dianalisa dalam jumlah yang sedikit dan indikator yang mempunyai sifat
tersebut adalah Cromium oxide. Jumlah Cromium oxide yang digunakan dalam penentuan kecernaan adalah 0,5-1,0%. Selanjutnya dikatakan bahwa keuntungan
menggunakan metode tidak langsung ini adalah feses yang telah dikumpulkan dapat
dianalisa kandungan nutriennya sehingga dapat diketahui koefisien daya cerna suatu
nutrien dalam pakan tersebut.
Amonia yang diekskresikan ikan merupakan indikator yang baik dalam
menentukan kadar optimum protein dalam pakan terutama jika dihubungkan dengan
pertumbuhan (Wermerskirchen et al. 1996). Menurut NRC (1993), nitrogen yang diekskresikan berkorelasi dengan nitrogen yang dikonsumsi. Eksresi amonia
menunjukan jumlah relatif protein pakan yang dicerna untuk sintesis protein atau
METODE PENELITIAN
Penelitian dilakukan dalam dua tahapan. Tahap 1 adalah uji efektivitas enzim
cairan rumen domba terhadap penurunan kandungan serat kasar bungkil kelapa. Uji
Tahap 2 adalah mengevaluasi kecernaan bungkil kelapa sebagai pakan benih ikan
mas (Cypinus carpio).
Penelitian Tahap 1: Uji Efektivitas Enzim Cairan Rumen Domba Terhadap Penurunan Kandungan Serat Kasar Bungkil Kelapa
Penelitian Tahap 1 dilakukan dengan tujuan untuk mengetahui efektivitas
penambahan enzim cairan rumen domba dalam bungkil kelapa terhadap penurunan
kandungan serat kasar pada bungkil kelapa. Pada tahap pertama dilakukan
pengambilan cairan rumen domba. Cairan rumen domba yang diambil adalah cairan
rumen dari domba yang selama pemeliharaan diberikan pakan rumput. Cairan rumen
domba yang dihasilkan diusahakan selalu dalam kondisi dingin. Cairan rumen yang
diambil kemudian disentrifugasi dengan kecepatan 10.000 rpm selama 20 menit pada
suhu 40C. Supernatan yang terbentuk direaksikan dengan amonium sulfat 60% menggunakanmagnetic stirer selama 1 jam dan didiamkan selama 24 jam pada suhu
40C. Selanjutnya cairan rumen yang telah diinkubasi selama 24 jam disentrifugasi pada kecepatan 10.000 rpm selama 20 menit kondisi 40C. Endapan yang terbentuk digunakan sebagai sumber enzim. Enzim kemudian dilarutkan dalam buffer fosfat
dengan perbandingan 1:1, yakni endapan dari 100 ml supernatan cairan rumen
dilarutkan dalam 100 ml buffer fosfat pH 7,0 dan disimpan pada suhu 40C (Budiansyah, 2010; Fitriliyani 2010). Enzim yang telah diperoleh dari hasil isolasi,
selanjutnya diuji aktivitas enzimnya yang meliputi aktivitas enzim selulase (FP-ase
Ghosse 1987), amilase dan protease, serta lipase (Tiests and Friedreck dalam
Barlongan 1990). Enzim yang diperoleh dari hasil isolasi kemudian ditambahkan ke
dalam bungkil kelapa dengan volume yang berbeda, yaitu: 0, 25, 50, 75, 100 atau
125 ml/kg bungkil kelapa. Lama waktu inkubasi di masing-masing dosis tersebut
Parameter yang diamati adalah kandungan serat kasar BK sebelum dan
sesudah inkubasi. Untuk mengetahui pengaruh penambahan enzim cairan rumen
terhadap parameter yang diukur dilakukan analisis statistik menggunakan analisis
ragam (uji F). Jika terdapat perbedaan antar perlakuan dilanjutkan dengan Uji Duncan
(Steel dan Torrie, 1993). Rancangan percobaan pada penelitian Tahap 1
menggunakan Rancanagan Acak Lengkap Faktorial dengan 2 faktor peubah dan 3
ulangan. Faktor peubah tersebut adalah dosis enzim cairan rumen domba dan lama
waktu inkubasi. Analisis kimia yang dilakukan pada tahapan ini meliputi analisis
serat kasar, kadar protein, lemak, abu, dan air bahan pakan hidrolisis (Takeuchi,
1988).
Penelitian Tahap 2: Uji Kecernaan Bungkil Kelapa yang telah Dihidrolisis dengan Enzim Cairan Rumen Domba Sebagai Pakan Benih Ikan Mas
Pakan Uji
Tahapan uji kecernaan bahan pakan dilakukan berdasarkan metode kecernaan
bahan yang dikemukakan oleh Watanabe (1988), yaitu pakan acuan (reference diet)
yang terdiri dari 100% pakan komersil, dan 2 jenis pakan uji (test diet) yang terdiri
dari pakan A (65% pakan komersil dan 30% bungkil kelapa + enzim (BKe), pakan B
(65% pakan komersil dan 30% bungkil kelapa (BK). Uji kecernaan dilakukan di
Laboratorium Nutrisi Ikan, Departemen Budidaya Perairan, Fakultas Perikanan dan
Ilmu Kelautan, Institut Pertanian Bogor pada bulan Juni Juli 2011. Pakan yang
dibuat, kemudian dianalisis proksimat untuk perhitungan nilai kecernaan di akhir
penelitian. Penelitian tahap dua dilakukan bertujuan untuk mengetahui nilai
kecernaan bahan pakan bungkil kelapa yang dihidrolisis dengan enzim cairan rumen
domba sebagai pakan benih ikan mas. Komposisi pakan acuan dan pakan uji dengan
Tabel 4. Komposisi pakan acuan (100% komersil), pakan uji (30% BKe) dan (30% BK) dalam persen
Komposisi (100% komersil) A (30% BKe) B (30% BK)
Pakan komersil 95 65 65
Bke 0 30 0
BK 0 0 30
Tepung tapioka 3 3 3
Kadar air 1,5 1,5 1,5
Cr2O3 0,5 0,5 0.5
Total 100 100 100
Kualitas Air
Pengukuran kualitas air dilakukan selama penelitian berlangsung meliputi
parameter suhu, oksigen terlarut (DO), pH dan amoniak. Pengukuran dilakukan pada
awal, pertengahan dan pada akhir penelitian. Kisaran nilai parameter kualitas air
disajikan pada Tabel 5.
Tabel 5. Kisaran nilai parameter kualitas air media pemeliharaan ikan mas selama penelitian
Uji Kecernaan Ikan Mas dan Pengumpulan Data
Ikan uji yang digunakan telah diadaptasikan dalam wadah pemeliharaan selama
7 hari. Ikan uji pada awal penelitian berjumlah 225 ekor benih ikan mas dengan bobot
rata-rata 1,0 - 1,5 g/ekor yang terdistribusi pada 9 akuarium berukuran 60x40x50 cm
volume air 50 liter, masing-masing akuarium berisi 25 ekor ikan. Setelah dipuasakan
selama 24 jam, ikan ditimbang dalam bobot basah tubuhnya. Pemeliharaan ikan
dilakukan selama 20 hari dan diberikan pakan uji yang mengandung indikator Cr2O3
dengan frekuensi pemberian pakan sebanyak tiga kali sehari, yaitu pukul 07.00, 12.00
Parameter Kualitas Air Kualitas air selama penelitian
Suhu (0C) 29 30
pH 6,5 7,5
DO (mg/l) 4,6 6,7
dan 17.00 WIB. Pemberian pakan dilakukan secara at satiation (sampai kenyang).
Pergantian air dilakukan satu kali per tiga hari dengan volume 50% dari total air
dalam wadah. Feses yang telah terkumpul dikeringkan di dalam oven dengan suhu
110°C selama 4-6 jam. Jumlah ikan uji setelah penelitian 222 ekor dengan bobot
akhir 2,0 3,0 g. Selanjutnya dilakukan analisis kandungan protein dan kadar air
Cr2O3 terhadap feses yang sudah dikeringkan dengan bantuan alat spektrofotometer
yang memiliki panjang gelombang 350 nm (Watanabe, 1988). Parameter yang
diamati pada penelitian tahap 2 berdasarkan prosedur Takeuchi (1988) meliputi:.
Parameter yang Diukur
a. Kecernaan Total (Takeuchi 1988)
Kecernaan Total (%) = 1 − 100
Keterangan :
ADC = Koefisien daya cerna pakan
IF =Cr2O3 dalam feses (%)
IP = Cr2O3 dalam pakan (%)
b. Daya Cerna Nutrien (Takeuchi 1988)
ADC(%) = 1 − 100
Keterangan :
ADC = Koefisien daya cerna pakan
IF =Cr2O3 dalam feses (%)
IP = Cr2O3 dalam pakan (%)
c. Kecernaan Energi (Takeuchi 1988)
Keterangan :
DE = Digestible Energi (energi tercerna) (kcal/100gr)
EP = Energi dalam pakan (kcal/100gr)
d. Kecernaan Bahan Watanabe (1988) :
Kecernaan bahan = (ADT - 0,7 AD) / 0,3
Keterangan :
ADT = nilai kecernaan pakan uji (kcal/100gr)
AD = nilai kecernaan pakan acuan (kcal/100gr)
e. Analisis Kimia
Analisis meliputi analisis proksimat pakan uji, analisis kandungan Cr2O3
pakan uji dan feses ikan. Analisis proksimat pakan uji meliputi pengukuran kadar
protein, lemak, serat kasar, abu dan air. Seluruh analisis proksimat dilakukan
berdasarkan Takeuchi (1988).
f. Analisis Statistik
Untuk mengetahui pengaruh pakan uji terhadap kecernaan yang dukur
tersebut maka digunakan analisis ragam (Uji F). Jika terdapat perbedaan antar
perlakuan dilanjutkan dengan uji Duncan dengan selang kepercayaan 95% (Steel and
Analisis Kandungan Nutrisi Bungkil Kelapa
Hasil uji efekvitas enzim diketahui maka selanjutnya dilakukan proses
hidrolisis bungkil kelapa. Analisis proksimat bungkil kelapa dengan perlakuan
penambahan volume enzim cairan rumen domba dan lama waktu inkubasi yang
berbeda disajikan pada Tabel 6.
Tabel 6. Kandungan lemak, protein, serat kasar bungkil kelapa pada perlakuan penambahan enzim cairan rumen domba dan lama waktu inkubasi
Volume
Tabel 6 menunjukan bahwa perlakuan dosis enzim cairan rumen domba dan
lama waktu inkubasi berpengaruh nyata terhadap penurunan kandungan serat kasar
bungkil kelapa. Nilai serat kasar pada bungkil kelapa tanpa penambahan enzim cairan
rumen domba lebih tinggi dibandingkan dengan nilai serat kasar bungkil kelapa pada
perlakuan penambahan enzim cairan rumen domba dengan lama waktu inkubasi 24
jam. Nilai serat kasar bungkil kelapa pada perlakuan penambahan enzim cairan
rumen domba 125 ml/kg dengan lama waktu inkubasi 24 jam lebih rendah (6,98%)
dibandingkan dengan nilai serat kasar dengan lama waktu inkubasi 12 jam dan 0 jam
pada dosis 25, 50, 75 dan 100 ml/kg.
Glukosa Terlarut Bungkil Kelapa
Hasil analisis pada Tabel 8 menunjukan bahwa kandungan glukosa terlarut
pada bungkil kelapa yang dihidrolisis dengan enzim cairan rumen domba mengalami
peningkatan seiring dengan penambahan volume enzim dan lama waktu inkubasi.
Kandungan glukosa terlarut tertinggi sebesar 0,464% diperoleh dari perlakuan
penambahan enzim cairan rumen domba 125 ml/kg selama 24 jam.
Tabel 7. Kandugan glukosa terlarut pada bungkil kelapa yang dihidrolisis dengan enzim cairan rumen domba
Volume enzim cairan rumen domba (ml/kg)
Uji Kecernaan Bungkil Kelapa yang telah Dihidrolisis dengan Enzim Cairan Rumen Domba Sebagai Pakan Benih Ikan Mas
Tahapan uji kecernaan bertujuan untuk mengetahui tingkat kecernaan ikan
terhadap bungkil kelapa yang digunakan sebagai bahan baku pakan. Hasil uji
kecernaan bungkil kelapa yang dihidrolisis dengan enzim cairan rumen domba
volume 125 ml/kg dengan lama waktu inkubasi 24 jam dan bungkil kelapa tanpa
hidrolisis meliputi nilai kecernaan total, kecernaan protein dan kecernaan energi
disajikan pada Tabel 8. Hasil analisis data menunjukan bahwa pakan komersil dan
pakan 30% BKe memiliki kecernaan yang lebih tinggi dari pakan 30% BK.
Tabel 8. Kecernaan total, kecernaan protein dan kecernaan energi
Parameter Uji Pakan
(100% Pakan komersil) A (30% BKe) B(30% BK)
Kecernaan (%)
Total
Protein
Energi
67,35±2,01a
76,93±1,42ab
71,50±0,81b
66,32±2,17a 63,52±0,39b
77,71±1,80a 75,51±0,65a
69,60±0,81b 64,80±0,77a
Notasi yang sama pada baris yang sama menunjukan pengaruh perlakuan yang tidak berbeda nyata (P<0,05).
Hasil perhitungan uji kecernaan bahan baku menunjukan bahwa baik
kecernaan total, protein dan energi mengalami peningkatan nilainya. Nilai kecernaan
BKe lebih tinggi dari BK (Tabel 9).
Analisa sidik ragam nilai kecernaan bahan uji menunjukan bahwa nilai
kecernaan (total, protein dan energi) pada pakan BKe ternyata memiliki nilai
kecernaan yang lebih tinggi dibandingkan nilai kecernaan pada pakan BK. Nilai
kecernaan total BKe sebesar (63,92%), kecernaan protein (79,53%), dan kecernaan
energi (72,2% ), sedangkan nilai kecernaan total BK (54,58%), kecernaan protein
Tabel 9. Kecernaan bahan uji
Bahan Uji Kecernaan Bahan (%)
Total Protein Energi
Bugkil kelapa + enzim
Bungkil kelapa
63,92±0,35
54,58±0,70
79,53±0,65
65,17±0,43
72,2±0,28
49,17±0,53
Pembahasan
Tingginya aktivitas enzim selulase dan amilase pada cairan rumen domba
diduga akibat jenis pakan yang dikonsumsi selama masa pemeliharaan. Domba yang
diambil cairan rumennya ini mengkonsumsi rumput, dimana rumput mengandung
serat yang cukup tinggi sehingga dikonversi pada saluran pencernaan menjadi enzim
pendegradasi serat yang dibutuhkan di dalam rumen domba untuk mencerna rumput.
Menurut Moharery dan Das, (2002), aktifitas enzim dalam cairan rumen tergantung
dari komposisi atau perlakuan makanan. Cairan rumen domba yang digunakan untuk
penelitian tersebut mengkonsumsi rumput selama pemeliharaan, sehingga aktifitas
enzim selulase lebih tinggi. Lubis (1992) mengemukakan bahwa kandungan serat
kasar rumput sangat tinggi yaitu 30,86%, sehingga enzim selulase yang dihasilkan di
rumen juga lebih banyak. Pada penelitian ini diperoleh hasil aktifitas enzim tertinggi
pada perlakuan volume 125 ml/kg bahan dengan lama waktu inkubasi 24 jam yaitu
pada enzim selulase sebesar 0,511 IU/ml/menit diikuti enzim amilase sebesar 0,510
μg glukosa/ml/menit; enzim protease 0,057 μg protein/ml/menit dan lipase 0,036 μg lemak/ml/menit. Penelitian Pamungkas (2011) mendapatkan efektivitas enzim
tertinggi adalah enzim selulase 0,31 IU/ml/menit sedangkan Fitriliyani (2010)
menghasilkan aktivitas enzim tertinggi adalah enzim selulase 1,66 IU/ml/menit.
Perbedaan respon aktivitas enzim tersebut diduga akibat perbedaan domba
yang digunakan enzim cairan rumennya. Budiansyah (2010) menemukan aktifitas
enzim tertinggi dihasilkan oleh rumen sapi asal lokal dibandingkan rumen sapi impor.
Hal ini disebabkan karena sapi lokal lebih banyak mengkonsumsi hijauan sehingga
banyak mengkonsumsi konsentrat, sehingga lebih sedikit menghasilkan mikroba
pencerna serat. Akibatnya, aktivitas enzim selulase pada cairan rumen sapi lokal lebih
tinggi dibandingkan dengan aktivitas enzim selulase pada sapi impor. Perbedaan nilai
tersebut juga diduga akibat dari umur domba yang diambil cairan rumennya berbeda.
Agarwal et al., (2002) melaporkan bahwa anak domba dengan berat badan 23,5 kg yang diberi makan minum susu sampai 8 minggu dan diteruskan dengan 50%
konsentrat dan 50% rumput sampai umur 24 minggu mendapatkan bahwa
enzim-enzim yang ada dalam cairan rumen antara lain carboxymethyl cellulase dengan
aktivitas enzim 3,60 mol glukosa per jam per ml, alpha amylase 0,33 umol glukosa
per menit per ml, xylanase 0,29 umol xylosa per menit per ml. Perbedaan respon
aktivitas enzim amilase juga terjadi.
Penelitian ini menghasilkan aktifitas enzim amylase 0,510 IU/ml/menit.
Pamungkas (2011) mendapatkan aktivitas enzim amilase 0,14 IU/ml/menit,
sedangkan Fitriliyani memperoleh aktivitas enzim amilase 1,32 IU/ml/menit.
Perbedaan nilai tersebut diduga adanya perbedaan domba dan umur domba serta
makanan yang dimakan selama masa pemeliharaan. Aktivitas enzim amylase yang
dilaporkan Fitriliyani (2010) lebih tinggi dibanding yang dilaporkan penelitian
lainnya disebabkan domba yang digunakan mengkonmsi rumput dan ditambahkan
daun lamtoro yang mengandung serat kasar yang tinggi sehinggi enzim amylase yang
dihasilkan di rumen juga cukup tinggi. Moharery dan Das (2002) melaporkan bahwa
cairan rumen domba yang berisi sel-sel bakteri mempunyai aktivitas enzim selulase,
amylase, protease dan lipase yang lebih tinggi dari cairan rumen tanpa sel-sel
mikroba.
Di dalam penelitian ini, hasil hidrolisis terbaik mampu menurunkan serat
kasar bungkil kelapa dari 14,34% menjadi 6,98%, sehingga kadar serat kasar yang
tertinggal dari semula menurun lebih dari setengahnya. Penelitian Kurniansyah
(2012) juga menggunakan enzim cairan rumen domba volume 150 ml/kg untuk
mendegradasi kandungan serat kasar kakao dengan lama waktu inkubasi 24 jam.
Hasil yang diperoleh terjadi penurunan kandungan serat kasar kakao dari 27,97%
serat pada tepung daun lamtoro dengan menggunakan enzim cairan rumen domba
volume 100 ml/kg, lama waktu inkubasi 24 jam dapat mendegradasi serat kasar dari
16,77% menjadi 7,774%, atau turun sebanyak 53,7% (Fitriliyani, 2010). Penelitian
dengan menggunakan enzim cairan rumen domba volume 100 ml/kg untuk
menghidrolisis bungkil kelapa sawit selama 24 jam juga dilakukan Pamungkas
(2011). Hasil penelitian tersebut berhasil menurunkan kandungan serat kasar BKS
dari 17,54% menjadi 10,63% (turun 40%). Prosen penurunan serat kasar bervariasi
diduga karena bahan yang dirumen berbeda dan substrat yang digunakan juga
berbeda. Aktifitas enzim dalam rumen yang semakin tinggi akan efektif menurunkan
serat kasar dibandingkan penambahan rumen yang memiliki aktivitas enzim yang
lebih rendah.
Meningkatnya kandungan glukosa terlarut dalam bungkil kelapa seiring
penambahan volume enzim cairan rumen domba. Kandungan glukosa terlarut pada
BKe meningkat dari 0,013% menjadi 0,464%. Pamungkas (2011) menggunakan
enzim cairan rumen domba dosis 100 ml/kg bahan bungkil kelapa sawit dengan lama
waktu inkubasi 24 jam meningkatkan kandungan glukosa terlarut sebesar 0,469%.
Perbedaan nilai tersebut diduga erat hubungannya dengan kerja enzim selulase.
Enzim selulase terbukti efektif mendegradasi serat kesar menjadi gula-gula sederhana
yang dapat diserap oleh ikan. Hardjo et al.(1989) mengemukakan bahwa selulosa adalah polimer tak bercabang dari glukosa yang dihubungkan melalui ikatan 1,4 β glukosida. Enzim yang mendegradasi selulosa yaitu endoglukanase atau karboksil
metal selulase (endo-1,4- β-glukanase). Lebih lanjut dinyatakan bahwa kompleks enzim selulase mempunyai tiga komponen utama yang bekerja bersama-sama atau
bertahap dalam menguraikan selulosa menjadi unit glukosa, yaitu :
1. Endo-selulase yang memotong bagian dalam struktur Kristal dari selulosa
dan mengeluarkan unit selulosa dari rantai polisakarida.
2. Ekso-selulase yang memotong 2-4 unit selulosa dari rantai akhir hasil
produksi endo-selulase dan menghasilkan tetrasakarida atau disakarida
3. Selubiosa atau β-glukosidase yang menghidrolisis produk dari ekso-selulase menjadi monosakarida.
Tiga reaksi tersebut yang dikatalis oleh selulase memotong interaksi
nonkovalen dalam bentuk ikatan hidrogen yang ada dalam struktur kristal selulosa
oleh enzim endo-selulase, menghidrolisis serat selolusa menjadi sakarida yang lebih
sederhana oleh ekso-selulase, serta menghidrolisis disakarida dan tetrasakarida
menjadi glukosa oleh enzim β-glukosidase.
Kandungan serat kasar bungkil kelapa yang rendah mampu meningkatkan
kecernaan benih ikan mas. Hasil penelitian menunjukan bahwa kecernaan total
(63,92%), protein (79,53%), maupun energi (72,2%) pada pakan BKe memiliki nilai
kecernaan yang lebih tinggi dibandingkan nilai kecernaan total (54,58%), protein
(65,17%) dan energy (49,17%) pada pakan BK. Rendahnya serat kasar dalam pakan
BKe menyebabkan peningkatan penyerapan zat-zat makanan dalam saluran
pencernaan sehingga mempercepat pertumbuhan dan proses fisiologis lainnya.
Proses tersebut terjadi akibat mekanisme kerja enzim yaitu memperbaiki kecernaan,
menurunkan kekentalan (viskositas) digesta, memodifikasi morfologi dan histologi
saluran pencernaan dan memodifikasi komunitas mikroba saluran pencernaan (Khan
et al. 2006). Hidrolisis nutrient makro dimungkinkan dengan adanya beberapa enzim pencernaan seperti protease, karboksilase, dan lipase (Zonneveld et al. 1991). Robinson (2001) mengemukakan bahwa rendahnya serat kasar dalam pakan
menyebabkan tingginya daya cerna dan penyerapan zat-zat makanan didalam alat
pencernaan ikan. Selama pakan berada dalam usus ikan, nutrient dicerna oleh
berbagai enzim menjadi bentuk yang dapat diserap oleh dinding usus dan masuk
dalam sistim peredaran darah. Sebaliknya pakan yang mengandung serat kasar tinggi
akan menghasilkan feses yang lebih banyak sehingga serat kasar yang tidak tercerna
tersebut dapat membawa zat-zat makanan yang seharusnya dicerna. Kebutuhan serat
didalam pakan dalam jumlah maksimal berkisar 7%. Serat yang melebihi batas
maksimal akan menurunkan kandungan gizi dalam pakan.
Keberadaan serat kasar yang tinggi dalam pakan tidak diharapkan karena
ikan terhambat. Keterbatasan ikan dalam memanfaatkan serat berkaitan dengan
ketersediaan enzim sellulotik yang terbatas dalam saluran pencernaan ikan, bahkan
pada level tertentu dapat menghambat pertumbuhan ikan. Pakan dengan kandungan
serat kasar yang tinggi dapat menyebabkan absorbs zat makanan berkurang dan
koefisien cerna semua zat makanan menurun. Namun, tak dapat dihindari bahwa serat
kasar juga dibutuhkan dalam pakan walaupun dalam jumlah yang sangat sedikit.
Serat kasar dibutuhkan dalam pakan untuk membantu proses pencernaan makanan.
Menurut Piliang (2006), serat kasar mernbantu mempercepat ekskresi sisa-sisa
makanan rnelalui saluran pencernaan. Dalam keadaan tanpa serat, feses dan
kandungan air rendah akan lebih lama tinggal dalam saluran usus yang dapat
rnenyebabkan gangguan pada gerakan peristaltik pada usus besar sehingga eksresi
Afrianto E, dan Liviawati E. 2005. Pakan ikan: pembuatan, penyimpanan, pengujian, pengembangan. Penerbit Kanisius.
Agarwal N, Kamra DN, Chaunhary LC, Agarwal I, Sahoo A and Pathak NN. 2002. Microbial status and rumen enzyme profile of crossbred calves fed on different microbial feed additives. Letter in Applied Microbiology, 34:329-336.
Anggorodi R. 1990. Kemajuan mutakhir dalam ilmu makanan ternak unggas.Cetakan Pertama. Universitas Press. Jakarta.
Anonim. 2010. PT. Bireuen Coconut Oil. Bireuen. Nangroe Aceh Darusallam.
. 2011. Kebutuhan bahan baku lokal untuk pengembangan industri pakan nasional. Forum Inovasi Teknologi Akuakultur Kementrian Kelautan dan Perikanan. Bali.
. 2011. Statistik komoditi hasil perkebunan di Indonesia. Kementerian Pertanian Direktorat Jenderal Perkebunan. Jakarta.
Arora SP. 1989. Pencernaan mikroba pada ruminansia. Penerbit Gajah Mada Press, Yogyakarta.
Barlongan TG. 1990. Studies on Lipases of milkfish (Chanos chanos). Aquakulture,89:315-325.
Budiansyah A. 2010. Aplikasi cairan rumen sapi sebagai sumber enzim, asam amino, mineral dan vitamin pada ransum broiler berbasis pakan lokal. Disertasi. Sekolah Pascasarjana Institut Pertanian Bogor.
Bureau DP. 2000. Feather meals and meat bone meal from different origin as protein sources in rainbow trout (Onchorynchus mykis) diets. Aquaculture: 181.281-291.
Choet M. 2001. Feed Non-starch polyssacharides: chemical structure and nutritional significance. Feed milling International, June Issuem pp. 13-26.
Fitriliyani I. 2010. Peningkatan kualitas nutrisi tepung daun lamtoro dengan penambahan ekstrak enzim cairan rumen domba untuk pakan ikan nila (Oreochromis sp). Disertasi. Sekolah Pascasarjana Institut Pertanian Bogor. Furuichi M. 1988. Fish nutrition, p.1-78. In : Watanabe T (ed). Fish nutrition and
mariculture. Tokyo. Departemen of Aquatic Biosciences Tokyo University Of Fisheries.
Hardiyanto S. 2001. Kecernaan (in vitro) dan kelarutan ransum komplit domba berbahan baku jerami teramoniasi dan onggok yang mendapat perlakuan cairan rumen. Skripsi. Institut Pertanian Bogor.
Hardjo S.S, Indrasti N.S, Tajuddin B. 1989. Biokonversi: pemanfaatan limbah industri pertanian. Pusat Antar Universitas Pangan dan Gizi. IPB.
Harver, JE dan Hardy, RW, editor. 2002. Fish Nutrition. Third Edition. California USA:Academic Press.822 pp.
Hepher B. 1990. Nutrition and pond fishes. Cambridge University Press. Cambridge, New York.
Indariyanti N. 2011. Evaluasi kecernaan campuran bungkil inti sawit dan onggok yang difermentasi oleh Trichoderma harzianum Rifai untuk pakan ikan nila Oreochromis sp [Tesis]. Sekolah Pascasarjana Institut Pertanian Bogor. Kamra DN. 2005. Special Section microbial diversity: Rumen microbial ecosystem.
Current Science, 89(10):124-135.
Khan SH, Sardar R, Siddique B. 2006. Influence of enzymes on performance of broilers fed sunflower-corn based diets. Pakistan Vet J26(3): 109-114.
Kung LJR, Treacher RJ, Nauman GA, Smagala AM, Endres KM and Cohen MA. 2000. The effect of treating forages with fibrolytic enzyms on its nutritive value lactation performance of dairy cows. J. Dairy Sci.83:115-122.
Kurniansyah A. 2012. Uji efektivitas penambahan enzim cairan rumen domba terhadap penurunan serat kasar dan nilai kecernaan kulit kakao sebagai pakan ikan nila.
Lovell T. ,1988. Nutrition and feeding of fish. Auburn Univercity. Published by : Van Nostrand Academy of Sciences Washington DC. 260 pp.
Lubis D. A. 1992. Ilmu makanan ternak. PT. Pembangunan, Jakarta.
Malathi V and Devegowda G. 2002. In Vitro evaluation of nonstarch polysaccharide digestibility of feed ingredients by enzymes. Department of Poultry Science, University of Agricultural Sciences, Hebbal, Bangalore-India.
Midlen A and Redding T. 1998. Environmental Management for Aquaculture. Chapman Hall, London.
Moharrery A and Das Tirta K, 2002. Corealtion between microbial enzyme activities in the rumen fluid of sheep under different treatments. Reprod.Nutr.Dev, 41:513-529.
Mokoginta I, Takeuchi T, I Suprayudi AM , Wiramihardja Y, dan Setiawati M. 1999. Pengaruh sumber karbohidrat yang berbeda terhadap kecernaan pakan, Efesiensi pakan dan pertumbuhan benih gurame (Osphronemus gouramy Lac. Jurnal Ilmu Ilmu Perairan dan Perikanan Indonesia. IV (2): 13-19.
Ng.W.K and Chen, M.I. 2002. Replacement of soybean meal with palm kernel meal in practical diets for hybrids Asian-African Catfish. Aquaculture 12:67-76. Ng.W.K and Chen, M.I. 2004. Researching the use of Palm Kernel Cake in
Aquaculture feeds. Fish Nutrition Laboratory, Universitas Sains Malaysia,Penang.
Ng.W.K and Chong K. 2002. The Nutritive value of palm meal end the Effact of enzyme suplementation in practical diets for Red Hybrid Tilapia (Oreocromis sp.). Asian Fisheries Science 15 (2002):167-176. Asian Fisheries Society, Manila, Philipines.
National Research Council (NRC). 1993. Nutrient requirement of fish. National Academy of Science. Washington DC. 86pp.
Pamungkas WS. 2011. Uji Efektivitas penambahan enzim cairan rumen domba terhadap penurunan serat kasar dan nilai kecernaan bungkil kelapa sawit sebagai pakan benih ikan Patin Siam Pangasius pangasius. Thesis. Pascasarjana IPB. Bogor.
Parakkasi A. 1990. Ilmu Nutrisi dan makanan ternak monogastrik. UI Press. Jakarta Piliang WG. Sastradipraja D. 2006. Studi Analisa metabolisme kalsium dan
kolesterol serta kebutuhan kalsium pada ayam petelur yang mendapat ransum dengan serat kasar tinggi asal dedak padi (Laporan Penelitian DP3M Ditjen Dikti). IPB. Bogor.
Purnomohadi M. 2006. Peranan bakteri selulotik cairan rumen pada fermentasi jerami padi terhadap mutu pakan. Jurnal Protein, Vol 13, No 2.
Robinson EH. Menghe HLi, Bruce BM. 2001, A Practical guide to nutrition feeds and feeding of catfish. Bull. 1113. Missisippi Agricultural & Forestry Experiment Station. Missisipi State University.
Sandi S. 2010. Peningkatan kualitas nutrisi silase berbahan baku singkong varietas pahit dengan enzim cairan rumen dan bakteri Leuconostoc mesenteroides sebagai pakan ternak unggas. Tesis. Institut Pertanian Bogor.
Steel RGD and JH. Torrie, 1993. Principles and procedure of statistic. McGraw Hill. London.
Sundu BA. Kumar and JG. Dingle. 2003. Perbandingan dua produk enzim komersial pencerna beta manan pada ayam pedaging yang mengkonsumsi bungkil kelapa sawit dengan level yang berbeda. Prosiding Seminar Nasional Pemanfaatan Sumberdaya Hayati Berkelanjutan.pp : 19-25
Takeuchi T. 1988. Laboratory work chemical evaluation of dietary nutrition. p. 179 – 229. In Watanabe T. Fish Nutrition and Mariculture JICA Textbook the General Aquaculture Course. Tokyo: Kanagawa International Fisheries Training Center.
Wahju J. 1988. Ilmu nutrisi unggas. Yogyakarta: Gadjahmada University Press.
Watanabe T. 1988. Fish nutrition and mariculture. Department of aquatic bioscience. Tokyo university of fisheries.JICA. 223 pp.
Watanabe T and Cho CY. 1988. Nutritional energetic. P. 79-94. In : Fish nutrition and mariculture. Kanagawa Fisheries Training Center. Japan International Cooperation Agency.
Wilson R.P. 1994. Utilization of dietary carobohidrate by fish. Aquaculture, 124: 67-80.
Wizna H. Abbas, Y, Rizal, I.P. Kompiang and A. Dharma. 2008. Potensi bakteri Bacillus amylseraquefaciens serasah hutan sebagai inokulum fermentasi pakan berserat tinggi. Jurnal Ilmiah Ilmu-Ilmu Peternakan. Vol VIII. No 3.p. 212-220.
Yamamoto T. Konishi K. Shima T. Furuita H. Suzuki N. and Tabata M. 2001. influence of dietery fat and carbohydrate levels on growth and body composition of ranbow trout Onchorhyncus mykiss under self feeding conditions. Fisheries Science. 67:221-227.
Zonneveld N, Huisman EA, Boon JH. 1991. Prinsip-prinsip budidaya ikan. PT. Gramedia Pustaka Utama. Jakarta. 318 p.