(S a c c h a r u m o f f i c i n a r u m L . ) D E N G A N B A N T U A N
Agrobacterium tumefaciens GV2260 DAN REGENERASINYA
ANNEKE PESIK
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
SURAT PERNYATAAN
Dengan ini saya menyatakan bahwa Tesis yang berjudul: “Penyisipan Gen Fitase (pBINPI-II EC) ke Kalus Tebu (Saccharum officinarum L.) dengan Bantuan Agrobacterium tumefaciens GV2260 dan Regenerasinya” adalah benar merupakan hasil karya saya sendiri dan belum pernah dipublikasikan.
Tesis ini belum pernah diajukan untuk memperoleh gelar pada program sejenis di perguruan tinggi lain. Semua sumber data dan informasi yang digunakan telah dinyatakan secara jelas dan dapat diperiksa kebenarannya.
Penelitian ini didanai oleh Kementerian Pendidikan dan Riset Jerman (Bundesministerium für Bildung und Forschung Germany, BMBF) melalui kerjasama bilateral Indonesia-Jerman untuk konstruksi tebu transgenik dan proyek Riset Andalan Perguruan Tinggi dan Industri (RAPID), dengan peneliti utama Dr. Ir. Dwi Andreas Santosa, MS. Sebagian pendanaan juga diperoleh dari Indonesian Center for Biodiversity and Biotechnology (ICBB) Bogor.
Bogor, September 2005
ANNEKE PESIK. Penyisipan Gen Fitase (pBINPI-II EC) ke Kalus Tebu (Saccharum officinarum L.) dengan Bantuan Agrobacterium tumefaciens GV2260 dan Regenerasinya. Dibimbing oleh DWI ANDREAS SANTOSA dan SUDARSONO.
Rekayasa genetik digunakan untuk meningkatkan karakter tebu yang pada umumnya memiliki fertilitas rendah dan polihibrid. Kami melaporkan introduksi gen fitase yang berasal dari bakteri ke dalam genom beberapa kultivar kalus tebu melalui Agrobacterium tumefaciens GV2260. Penelitian ini bertujuan untuk mendapatkan komposisi media regenerasi yang sesuai untuk kalus transforman dan nontransforman. Eksplan meristem dan kalus dari tebu kultivar PA 117, PSJT 94-33, dan PS 851 digunakan dalam uji regenerasi dan transformasi. Komposisi media regenerasi yang baik untuk menginduksi tunas, daun, dan akar dari dua kultivar PA 117 dan PSJT 94-33 berturut-turut adalah media R10 (yaitu media MS yang ditambahkan 2 mg/L + 57 mg/L dalapon) dan media R4 (yaitu media MS yang ditambahkan 1 mg/L IAA dan 57 mg/L dalapon). Regenerasi kalus transforman PSJT 94-33 mampu menghasilkan tunas dan daun. Sedangkan dua kultivar transforman lain, yaitu PA 117 dan PS 851 (dalam media MS dengan penambahan 2 mg/L IAA dan 59 mg/L dalapon yang dimodifikasi (dengan penambahan glukosa 1%, 2.5 mg/L asam sitrat dan 2 mg/L asam askosbat) hanya menghasilkan tunas. Reaksi PCR untuk amplifikasi gen fitase dari kalus transforman menunjukkan hasil positif. Dengan demikian, kalus transforman dapat terus ditumbuhkan menjadi tanaman transgenik yang lengkap.
Kata kunci: kultivar tebu, transformasi gen fitase, Agrobacterium tumefaciens,
ABSTRACT
ANNEKE PESIK. Insertion of Phytase Gene (pBINPI-II EC) into Callus of Sugarcane (Saccharum officinarum L.) by Agrobacterium tumefaciens GV2260 and Its Regeneration. Under the direction of DWI ANDREAS SANTOSA and SUDARSONO.
Genetic engineering can be used to improve the characteristic of sugarcane which much of them are infertile and polyhybrid. We report introducing of phytase gene derived from bacteria into the genome of several cultivar of sugarcane callus via Agrobacterium tumefaciens GV2260. This research were also conducted to find out the composition of regeneration media suitable for both nontransformant and transformant calli. Meristematic explants and calli of sugarcane var. PA 117, PSJT 94-33, and PS 851 were used for regeneration and transformation experiment. The best composition of regeneration media to induced buds, leaves, and roots of both var. PA 117 and PSJT 94-33 are R10 (MS media suplemented with IAA 2 mg/L + dalapon 57 mg/L) and R4 (MS media suplemented with IAA 1 mg/L + dalapon 57 mg/L), respectively. Regeneration of transformant calli of PSJT 94-33 produced buds and leaves. Another two transformant cultivar PA 117 and PS 851 (in MS media with IAA 2 mg/L + dalapon 59 mg/L modified by adding glucose 1%, citric acid 2.5 mg/L and ascorbic acid 2.5 mg/L) were only became bud. Positive result were also revealed by using polimerase chain reaction of phytase gene onto transformant calli. Therefore, transformant calli can be established into transgenic plants.
Dilarang mengutip dan memperbanyak tanpa izin tertulis dari Institut Pertanian Bogor, sebagian atau seluruhnya dalam bentuk
PENYISIPAN GEN FITASE (pBINPI-II EC) KE KALUS TEBU (S a c c h a r u m o f f i c i n a r u m L . ) D E N G A N B A N T U A N
Agrobacterium tumefaciens GV2260 DAN REGENERASINYA
ANNEKE PESIK
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
Program Studi Bioteknologi
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
tumefaciens GV2260 dan Regenerasinya Nama : Anneke Pesik
Nomor Pokok : P055010121 Program Studi : Bioteknologi
Disetujui,
Komisi Pembimbing
Dr. Ir. Dwi Andreas Santosa, MS Prof. Dr. Ir. Sudarsono, MSc
Ketua Anggota
Diketahui,
Ketua Program Studi Bioteknologi Dekan Sekolah Pascasarjana
Dr. Ir. Muhammad Jusuf, DEA Prof. Dr. Ir. Hj. Syafrida Manuwoto, MSc
PRAKATA
Segala hormat dan puji dinaikkan kepada Tuhan Yang Maha Kuasa, karena berkat kasih karuniaNya yang berlimpah dan tak berkesudahan ini, penulis telah menyelesaikan penelitian dan penulisan tesis yang berjudul “Penyisipan Gen Fitase (pBINPI-II EC) ke Kalus Tebu (Saccharum officinarum L.) dengan Bantuan Agrobacterium tumefaciens GV2260 dan Regenerasinya”.
Dengan penuh kerendahan hati, kesempatan ini penulis menyampaikan terima kasih kepada Bapak Dr. Ir. Dwi Andreas Santosa, MS, Prof. Dr. Ir. Sudarsono, MSc, dan Dr. Ir. Utut Widyastuti, serta Dr. Ir. Agus Purwito, MSc, yang dengan sabar dan tak henti-hentinya telah memberikan banyak pengarahan dan saran selama penelitian dan penulisan tesis ini.
Ucapan terima kasih yang tulus penulis sampaikan pula kepada:
1. Kepala Lab. Mikrobiologi & Bioteknologi Lingkungan PPLH, dan Lab. Bioteknologi Tanaman Fakultas Pertanian, Institut Pertanian Bogor, dan Direktur PT. Saraswanti Indo Genetech Bogor, yang telah mengizinkan penelitian ini berlangsung. Kepala Lab. Kultur Jaringan PT. Pabrik Gula Rajawali Nusantara Indonesia II di Cirebon yang telah memberikan bahan eksplan dan kalus tebu. 2. Bapak, Ibu, Suami, saudara kakak-beradik sepupu, dan keponakan atas
dukungan doa dan materil yang diberikan bagi penulis.
3. Kementerian Pendidikan dan Riset Jerman (BMBF), Proyek RAPID, ICBB Bogor, dan Yayasan Dana Beasiswa Maluku (YDBM) Jakarta atas bantuan dana sehingga penelitian dan penulisan tesis ini dapat berlangsung.
4. Rekan-rekan kerja di Lab. MBL PPLH, Lab. Bioteknologi Tanaman Fakultas Pertanian IPB, dan Lab. PT Saraswanti Indo Genetech, atas dukungan dan kebersamaan.
5. Teman-teman Bioteknologi 2001 dan kepada semua pihak yang tidak disebutkan satu-persatu atas bantuan dan dorongannya kepada penulis selama studi sampai dengan penulisan tesis ini.
Akhirnya, penulis menyadari bahwa tulisan ini masih jauh dari sempurna. Namun penulis berharap tesis ini bermanfaat bagi kepentingan semua pihak.
Penulis dilahirkan di Manado, Sulawesi Utara pada tanggal 1 Agustus 1978 sebagai anak tunggal dari ayah Herling Pesik dan ibu Hetty Waworga (†).
Pendidikan Sarjana Pertanian ditempuh di Fakultas Pertanian, Jurusan Hama dan Penyakit Tanaman, Universitas Sam Ratulangi, Manado, dan lulus pada tahun 2001.
DAFTAR ISI
Halaman
DAFTAR TABEL ... x
DAFTAR GAMBAR ... xi
DAFTAR LAMPIRAN ... xii
PENDAHULUAN ... 1
Latar Belakang ... 1
Tujuan Penelitian ... 3
TINJAUAN PUSTAKA ... 4
Perbaikan Genetik Tebu ... 4
Kultur Jaringan Tanaman Tebu ... 5
Gen Fitase ... 10
Konstruksi Vektor pBINPI-II EC ... 11
Transformasi dengan Bantuan Agrobakterium ... 14
Analisis Tanaman Transgenik ... 15
BAHAN DAN METODE ... 17
Tempat dan Waktu Penelitian ... 17
Metode Penelitian ... 17
Kultur jaringan tanaman tebu ... 17
Transformasi tebu dengan gen fitase ... 19
HASIL DAN PEMBAHASAN ... 23
Kultur Jaringan Tanaman Tebu ... 23
Transformasi Tebu dengan Gen Fitase ... 34
SIMPULAN DAN SARAN ... 39
Simpulan ... 39
Saran ... 39
DAFTAR PUSTAKA ... 40
No. Halaman
DAFTAR GAMBAR
No. Halaman
1 Konstruksi kaset gen fitase appA dalam vektor pBINPI-II EC ... 13 2 Inisiasi dan proliferasi kalus ... 23 3 Grafik rataan jumlah tunas tebu kultivar PA 117 yang terbentuk
sampai dengan minggu ke-4 ... 24 4 Grafik rataan jumlah daun tebu kultivar PA 117 yang terbentuk
sampai dengan minggu ke-4 ... 25 5 Hasil optimasi media regenerasi tebu kultivar PA 117 sampai
dengan minggu ke-4 ... 26 6 Grafik rataan jumlah akar tebu kultivar PA 117 yang terbentuk
sampai dengan minggu ke-4 pada media regenerasi R10 ... 27 7 Hasil induksi akar tebu kultivar PA 117 pada media regenerasi
R10 ... 27 8 Grafik rataan jumlah tunas tebu kultivar PSJT 94-33 yang
terbentuk sampai dengan minggu ke-4 ... 28 9 Grafik rataan jumlah daun tebu kultivar PSJT 94-33 yang terbentuk
sampai dengan minggu ke-4 ... 29 10 Hasil optimasi media regenerasi tebu kultivar PSJT 94-33 sampai
dengan minggu ke-4 ... 30 11 Grafik rataan jumlah akar tebu kultivar PSJT 94-33 yang terbentuk
sampai dengan minggu ke-4 117 pada media regenerasi R4 ... 30 12 Hasil induksi akar tebu kultivar PSJT 94-33 pada media
regenerasi R10 ... 31 13 Regenerasi tebu transforman PSJT 94-33, PA 117, dan PS 851 ... 32 14 Grafik rataan tunas transforman kultivar PA 117 pada media
regenerasi R10 ... 33 15 Grafik rataan tunas dan daun yang terbentuk dari regenerasi kalus
transforman kultivar PSJT 94-33 pada media regenerasi R4 ... 33 16 Grafik rataan tunas transforman yang terbentuk dari regenerasi kalus
transforman kultivar PS 851 pada media regenerasi Modifikasi P3GI ... 34 17 Kalus tebu transforman yang dihasilkan dari beberapa metode
transformasi ... 37 18 Hasil PCR gen fitase dari kalus transforman yang berumur 4 minggu
No. Halaman
1 Komposisi media MS untuk induksi kalus dan regenerasi
planlet yang telah dimodifikasi ... 44
2 Peta plasmid pBINPI-II EC ... 45
3 Komposisi media transformasi metode Enriquez-Obregón et al (1997) 45 4 Komposisi media transformasi metode Minarsih (2003) ... 45
5 Komposisi media transformasi metode Santosa et al (2004) ... 46
6 Komposisi media transformasi metode Modifikasi ... 46
Latar Belakang
Konsumsi gula nasional meningkat seiring dengan pertambahan penduduk Indonesia. Swasembada gula makin menurun karena industri gula dalam negeri tidak dapat memenuhi kebutuhan nasional. Tanaman tebu (Saccharum officinarum L.) sebagai bahan baku pembuatan gula adalah komoditi utama yang harus mampu berperan terhadap permasalahan kebutuhan gula.
Penyebab utama turunnya produksi tebu karena mutu bibit yang buruk. Petani tidak mengganti bibitnya dengan kultivar yang lebih baik, karena tidak memiliki daya beli. Kebanyakan petani menerapkan sistem keprasan. Sistem tersebut mempertahankan tunas baru pada pangkal tanaman setelah penebangan untuk berproduksi lagi. Cara ini beresiko terhadap penyakit busuk batang yang dapat menurunkan produksi hingga 30%. Produktivitas lahan juga berkurang akibat tidak efisiennya pemakaian pupuk fosfat (Hidayat 2002).
Pertanaman tebu yang bergeser dari lahan basah ke kering juga menyebabkan rendahnya efisiensi produksi gula. Oleh karena, lahan kering lebih banyak membutuhkan pupuk fosfat yang memerlukan biaya tinggi. Luas areal pertanaman tebu pada masa tanam 2002/2003 sebesar 351.472 ha yang terdiri dari lahan sawah sebesar 105.412 ha (30%) dan lahan kering (tegalan) mencapai 245.960 ha (70%), yang tersebar di Jawa seluas 121.738 ha dan di luar Jawa 124.222 ha (Hadi & Sutrisno 2003).
2
Pemuliaan tanaman perkebunan, termasuk tebu menggunakan metode konvensional dengan penyilangan menghadapi dua kendala utama. Kendala pertama adalah terbatasnya sumberdaya genetik yang secara seksual kompatibel dengan tanaman tebu induknya. Kendala kedua adalah siklus pemuliaan dengan metode ini dianggap terlalu lama (Minarsih 2003).
Menurut Hartatik (2000) peningkatan produksi gula secara menyeluruh dapat dilakukan dengan cara meningkatkan teknik budidaya, mengatur pola “tebang muat angkut”, meningkatkan efisiensi pabrik gula, dan menggunakan kultivar unggul baru. Usaha untuk mendapatkan kultivar tebu yang unggul terutama ditujukan untuk perbaikan kuantitas (bobot tebu per hektar) dan kualitas (rendemen gula). Usaha tersebut dilakukan dengan perbaikan genetik tanaman, salah satunya melalui rekayasa genetik yang terbukti manfaatnya dalam memperbaiki sifat tanaman. Contohnya teknik transformasi gen dengan bantuan agrobakterium yang merupakan cara alternatif untuk memperoleh tanaman dengan sifat yang diinginkan.
Keberhasilan perakitan kultivar tebu yang unggul ditentukan oleh berbagai faktor, antara lain adanya teknik transformasi dan regenerasi yang sesuai, konstruksi gen pembawa sifat unggul yang berfungsi dengan baik, dan karakter agronomis klon tanaman dan kemudahan teknis (Minarsih 1999).
Menurut Santosa (2002), salah satu upaya meningkatkan produktivitas tebu unggul dilakukan dengan transformasi gen fitase dari bakteri tanah sehingga meningkatkan ketersediaan fosfat ke dalam jaringan maupun sekitar perakaran, meningkatkan efisiensi pemupukan fosfat sehingga sedikit membutuhkan pupuk fosfat, meningkatkan jumlah klorofil dan laju fotosintesis. Ekspresi gen fitase di jaringan tebu diharapkan juga meningkatkan mutu daun tebu untuk pakan ternak.
seperti kalsium, magnesium dan kalium di dalam jaringan tanaman sehingga dapat mengurangi kebutuhan pupuk.
Vektor gen fitase untuk penyisipan ke dalam genom tanaman tebu telah dikonstruksi sebagai plasmid pBINPI-II EC. Plasmid pBINPI-II EC merupakan hasil konstruksi dan modifikasi melalui penelitian bersama antara Federal Research Centre for Nutrition, Centre for Molecular Biology Germany dan
Fakultas Pertanian IPB. Plasmid pBINPI-II EC memiliki beberapa kelebihan, di antaranya mempunyai jumlah salinan yang banyak dalam Escherichia coli, dan mempunyai ori sehingga dapat bereplikasi pada E. coli dan Agrobacterium tumefaciens, serta membawa gen penyeleksi antibiotik nptII yang menyandikan
enzim neomisin fosfotransferase yang tahan terhadap kanamisin.
Keberhasilan teknik transformasi genetik sangat dipengaruhi oleh kemampuan regenerasi yang baik dari eksplan yang digunakan untuk menjadi planlet yang utuh. Dalam hal ini teknik kultur jaringan sangat berperan penting untuk mendapatkan planlet atau tanaman transgenik yang diinginkan. Misalnya dengan memperhatikan berbagai faktor pendukung seperti media tumbuh, zat pengatur tumbuh (ZPT) yang seimbang, dan kondisi lingkungan kultur. Bahan eksplan dan genotipe tanaman juga merupakan faktor penting dalam menentukan regenerasi suatu eksplan (Trigiano & Dennis 2000).
Dengan demikian aplikasi bioteknologi untuk tanaman tebu berperan penting dalam meningkatkan produktivitas tebu unggul, mengurangi biaya untuk pemupukan dan menambah nilai tanaman tebu sebagai pakan ternak.
Tujuan Penelitian
Penelitian ini bertujuan untuk:
1. Mendapatkan media yang optimum untuk regenerasi tanaman tebu.
TINJAUAN PUSTAKA
Perbaikan Genetik Tebu
Tanaman tebu (Saccharum officinarum L.) merupakan tanaman perkebunan bernilai ekonomi tinggi karena selain menghasilkan gula juga mendukung berbagai industri lain. Tanaman tebu dapat tumbuh pada daerah tropis dan subtropis di seluruh dunia. Tebu pertama kali diketahui berasal dari daratan Indo-Gangetic India. Tebu termasuk genus Saccharum dari salah satu anggota kelompok Andropogoneae, famili Graminae. Dalam genus Saccharum ada tiga spesies yang dibudidayakan (S. officinarum L., S. barberi Jesw. dan S. sinense Roxb) dan dua spesies liar (S. robustum Brandes dan Jeswier ex Grassl, dan S. spontaneum L.) (Naik 2001).
Diketahui tanaman tebu merupakan bahan baku pembuatan gula pasir di samping berperan juga sebagai bahan baku dalam bidang industri seperti asam amino, asam organik dan bahan pangan. Beberapa industri gula mampu menghasilkan berbagai senyawa kimia tertentu seperti furfural, dextran, alkohol yang sangat tergantung pada tanaman ini (Enriquez-Obregón et al. 1997). Selain itu produk gula dapat dikembangkan menjadi bahan nutrisi pakan ternak, prosesing bahan makanan, pembuatan kertas dan sebagai sumber energi bahan bakar (Patrau 1989). Menurut Nonato et al. (2001) di Brazil tebu dikembangkan sebagai salah satu sumber energi penting (produksi etanol) serta untuk produksi bioplastik.
Perbaikan genetik pada tanaman tebu telah lama dilakukan dan belum menunjukkan hasil yang berarti, sehingga memasukkan gen tertentu dengan transformasi genetik merupakan salah satu cara penting untuk manipulasi genetik secara langsung pada tanaman. Transfer gen pada tanaman akan menghasilkan tanaman transgenik. Istilah transgenik dalam pengertian luas dipakai untuk tanaman yang memiliki gen asing yang berfungsi dan terintergrasi ke dalam genom tanaman (Uchimiya et al. 1989).
ekspresi suatu gen di dalam sel tanaman. Perkembangannya kemudian dilakukan transfer gen yang mengendalikan karakter-karakter yang bernilai ekonomis sejalan dengan ketersediaan klon-klon gen tersebut. Beberapa faktor yang berperan dalam menghasilkan tanaman transgenik antara lain metode yang efisien dalam kloning gen, ketersediaan konstruksi gen-gen baru, teknik transformasi, regenerasi tanaman, sistem vektor yang terus dikembangkan dan promotor yang spesifik untuk ekspresi gen pada organ tertentu (Aswidinoor 1995).
Teknik transformasi genetik tanaman tebu pertama kali dilakukan oleh Hauptmann et al. (1986) menggunakan penanda seleksi gen CAT (chloramphe-nicol-acetyltransferase) yang berasal dari promotor CaMV 35S untuk
mendapat-kan tanaman tebu yang tahan virus. Bower dan Birch (1992) menggunamendapat-kan partikel bombardment untuk suspensi sel dan kalus embriogenik dengan memasukkan gen neomycin phosphotransferase. Gallo dan Irvine (1993) mem-pelajari faktor fisik dan biologi yang berpengaruh terhadap ekspresi gen GUS secara transien pada tebu menggunakan teknik penembakan partikel. Hingga saat ini telah dilakukan penelitian tebu transgenik dengan sifat tertentu antara lain tahan terhadap insektisida dan beberapa herbisida yang lain, dan tahan terhadap serangan penyakit busuk batang (Arencibia et al. 1997). Wall dan Birch (1997) berhasil melakukan karakterisasi gen ketahanan albicidin dan ekspresinya pada tanaman tebu transgenik untuk ketahanan penyakit bercak daun. Pada tahun yang sama transformasi genetik tebu dengan bantuan Agrobacterium tumefaciens menggunakan antioksidan berhasil dilakukan oleh Enriquez-Obregón et al. (1997). Pada tahun 1998, Arencibia et al. melaporkan protokol yang efisien untuk transformasi tebu melalui A. tumefaciens EHA 101. Selain itu, Minarsih (2003) telah melakukan penelitian transformasi gen P5CS ke dalam sel tanaman tebu secara efektif dan efisien tanpa mengurangi kemampuan regenerasi dan ekspresi sel. Penelitian ini menghasilkan tanaman tebu dengan sifat toleransi terhadap kondisi kering.
Kultur Jaringan Tanaman Tebu
menumbuh-6
kan dalam media buatan dengan kondisi aseptik dan lingkungan yang terkendali, sehingga bagian-bagian tanaman tersebut mampu memperbanyak diri dan beregenerasi menjadi tanaman baru (Gunawan 1992). Menurut Suryowinoto (1991), kultur jaringan berarti membudidayakan suatu jaringan tanaman menjadi tanaman utuh yang mempunyai sifat seperti induknya. Ada empat aplikasi metode kultur jaringan dalam bidang pertanian, di antaranya adalah: 1) untuk produksi bahan-bahan fermentasi, 2) perbaikan sifat tanaman, 3) memelihara plasma nutfah, dan 4) pelipatgandaan hasil klon dari seleksi kultivar.
Dalam usaha peningkatan produktivitas tanaman tebu secara cepat pada tingkat petani awalnya digunakan teknik kultur jaringan tapi saat ini telah berkembang sebagai sarana pendukung program perbaikan sifat tanaman, misalnya mendapatkan sifat ketahanan penyakit, stres lingkungan atau perbaikan genetik lainnya.
Penelitian kultur jaringan tanaman tebu pertama kali dilakukan oleh Nickell di Hawai tahun 1961 dan dilanjutkan pada tahun 1977 oleh Heinz et al yang menjadi perintis teknik kultur jaringan tebu pertama di dunia, mereka berhasil mendapatkan planlet utuh yang berkembang dari kultur kalus dan menunjukkan variasi yang luas pada jumlah kromosom juga beberapa karakter penting dari tanaman tebu. Sejak saat itu berkembanglah teknik kultur jaringan tebu ini ke beberapa negara di dunia (Naik 2001).
Ada beberapa hal yang perlu diperhatikan dalam teknik kultur jaringan tanaman tebu, yaitu: 1) komposisi media tumbuh, 2) bahan eksplan, 3) zat pengatur tumbuh tanaman, dan 4) kondisi lingkungan kultur.
Media tumbuh
Media yang umum dipakai untuk memelihara pertumbuhan kultur sel atau jaringan tebu adalah variasi dari media White atau media Murashige dan Skoog (MS). Menurut Heinz et al. (1977), media White lebih baik untuk mendorong per-tumbuhan kalus tapi tidak untuk totipotensinya sedangkan media MS lebih unggul dari media White dalam menginduksi diferensiasi kalus dan perkembangan selanjutnya. Murashige (1978) menekankan perlunya perimbangan tertentu dalam pemberian campuran garam-garam mineral, gula, vitamin dan ZPT. Naik (2001) menjelaskan nutrisi yang esensial terdiri dari garam anorganik dan sumber karbon (energi), vitamin, dan hormon tanaman serta bahan lain seperti nitrogen organik yang mengandung asam organik dan substansinya. Media kultur tersebut dapat ditambah dengan bahan-bahan seperti air kelapa, ekstrak khamir, sari buah tomat dan ekstrak malt. Ekstrak malt atau khamir lebih cocok untuk pertumbuhan kultur suspensi tebu dibandingkan dengan air kelapa. Sebagai sumber energi, sukrosa dan glukosa dapat dipilih di antara jenis gula yang ada karena pada tahap-tahap permulaan, eksplan baru tumbuh membentuk kalus yang belum mengadakan fotosintesis atau cara hidupnya belum autotrofik (Pierik 1987). Liu (1981) menyebutkan bahwa untuk menstimulasi pembentukan kalus lebih baik digunakan air kelapa dan mio-inositol yang ditambahkan pada media MS. Namun menurut Apriyanti (1990) bahwa pertumbuhan kalus menjadi planlet lebih baik dalam media dengan air kelapa dari pada ekstrak malt. Pierik (1987) menyatakan bahwa berhasil tidaknya suatu media untuk menumbuhkan eksplan menjadi kalus embriogenik atau planlet yang utuh tergantung pada genotip atau sifat eksplan itu sendiri dan interaksi antara faktor luar dan komponen medianya.
8
Bahan eksplan
Dalam kultur jaringan tanaman tebu untuk mempercepat penampilan sifat totipotensi sangat dianjurkan menggunakan eksplan sebagai bahan tanam yaitu organ atau jaringan muda yang masih dalam keadaan meristematis. Pierik (1987) menyatakan bahwa meristem adalah sekelompok sel-sel yang mempunyai sifat selalu membelah, sel-selnya kecil dengan dinding tipis dan penuh plasma. Dalam tanaman ada suatu zone atau daerah yang secara permanen bersifat embriogenik.
Penggunaan eksplan dari jaringan muda lebih berhasil daripada jaringan tua, hal ini disebabkan jaringan muda lebih responsif terhadap hormon tumbuh pada media. Menurut Gunawan (1992), eksplan sebaiknya diambil dari bagian-bagian tanaman yang belum banyak mengalami perubahan bentuk dan kekhususan fungsi seperti meristem batang dan akar, meristem kambium, meristem interkalar, meristem daun dan fellogen. Hal-hal penting yang harus diperhatikan dalam pemilihan eksplan yaitu sumber eksplan, ukuran dan umur fisiologi.
Bahan eksplan dari kultivar tebu yang berumur 3-4 bulan dan bebas penyakit sering dipakai dalam penelitian kultur jaringan tebu, dengan memperhatikan cara sterilisasinya karena permukaan bagian eksplan umumnya mengandung sejumlah mikroba kontaminan sehingga sterilisasi permukaan perlu dilakukan sebelum ditanam pada media (Naik, 2001).
Blanco et al. (1999) menggunakan bagian daun yang masih hijau sebagai sumber eksplan yang dipotong 3–5 mm. Minarsih (2003), untuk inisiasi kalus jaringan meristem yang belum mengalami diferensiasi menggunakan eksplan meristem daun atau jaringan muda yang masih menggulung.
Zat Pengatur Tumbuh tanaman (ZPT)
berakar baik pada media cair maupun media padat (Trigiano & Dennis 2000). Kelompok auksin menyebabkan perpanjangan sel, pembelahan sel, pembentukan akar samping, menghambat pembentukan tunas aksilar. Kelompok sitokinin dapat memacu pertumbuhan sel bila dipakai bersama auksin, dalam konsentrasi 1-10 mg/L dapat menghindari pembentukan tunas samping sehingga menurunkan dominansi pertumbuhan apikal, menghambat pertumbuhan akar (Sastrowijono 1991).
Auksin dapat menginduksi diferensiasi pembentukan kalus dan pembelahan sel secara cepat. Heinz et al. (1977) menggunakan 2,4-D 3 mg/L untuk induksi kalus dengan penambahan air kelapa dan mio-inositol. Modifikasi media MS dengan kinetin 1 mg/L, NAA (naphtahlene acetic acid) 1 mg/L dan kasein hidrolisat 400 mg/L memberikan hasil yang baik untuk diferensiasi planlet dari kalus. Ahloowalia dan Maretzki (1983) berhasil mendapatkan kalus yang bersifat embriogenik dengan menggunakan medium MS ditambah 2,4-D 3 mg/L dan air kelapa 10%. Irvine et al. (1983) telah mencoba 79 macam bahan kimia untuk menginduksi pembentukan kalus tebu, yang diketahui efektif adalah 2,4-D, picloram dan dicamba dengan konsentrasi 2–20 mg/L. Falco et al. (1996) berhasil melakukan inisiasi kultur suspensi sel dengan media MS cair ditambah 2,4-D 3 mg/L, air kelapa 5% dan kasein hidrolisat 500 mg/L. Perbandingan pengaruh beberapa auksin seperti picloram, dicamba dan 2,4-D untuk induksi kalus juga dilakukan oleh Blanco et al. (1999) dengan media dasar MS, hasilnya picloram 8,2 µM, dicamba 22,6 µM dan 2,4-D 4,5 µM yang optimum untuk pembentukan kalus embriogenik dan embriogenesis somatik pada tanaman tebu kultivar CP 5243. Minarsih (2003) berhasil mendapatkan kalus dari jaringan parenkimatis yang belum mengalami diferensiasi dengan media MS ditambah air kelapa 10%, sukrosa 30 g/L dan 2,4-D 3 mg/L.
Kondisi lingkungan kultur
10
5000–10000 lux, kelembaban relatif ruang inkubasi diatur sekitar 70–80%, bila kurang dari 50% maka media yang disimpan akan cepat mengering sedangkan kelembaban yang terlalu tinggi meningkatkan kontaminan seperti jamur dan bakteri (Sugiyarta 1991). Arencibia et al. (1992) menggunakan lingkungan kultur untuk inkubasi kalus transgenik pada suhu 25 oC di bawah cahaya selama 16 jam dengan intensitas cahaya 2000 lux. Menurut Naik (2001), regenerasi tanaman tebu dari kultur meristem apikal dapat dilakukan pada suhu 20-26 oC dengan waktu cahaya 16 jam 5000-8000 lux.
Gen Fitase
Fitase (mio-inositol heksakisfosfat fosfohidrolase) merupakan suatu fosfo-monoesterase yang mampu menghidrolisis asam fitat menjadi ortofosfat anorganik dan ester-ester fosfat dari mio-inositol yang lebih rendah. Pada kondisi tertentu bahkan menjadi fosfat dan mio-inositol bebas. Menurut IUPAC-IUB, ada dua jenis enzim fitase yaitu:
a. 3-fitase (EC 3.1.3.8) yang mengkatalisis reaksi: mio-inositol heksakisfosfat + H2O → mio-inositol-1,2,4,5,6-pentakisfosfat + Pi
b. 6-fitase (EC 3.1.3.26) yang mengkatalisis reaksi: mio-inositol heksakisfosfat + H2O → mio-inositol-1,2,3,4,5-pentakisfosfat + Pi
Jenis 3-fitase umumnya terdapat pada mikroba, sedangkan 6-fitase umumnya terdapat pada biji-bijian. Greiner et al. (1993) menjelaskan bahwa 6-fitase ditemukan pula pada Escherichia coli. Sumber 6-fitase bervariasi mulai dari berbagai jenis mikroba seperti jamur dari genus Aspergillus (A. ficuum, A. oryzae, A. carbonarius dan A. niger), ragi (Saccharomyces cerevisiae, Schwanniomyces
castellii) dan bakteri (Klebsiella aerogenes, Streptococcus bovis, Bacillus subtilis,
E. coli) hingga jaringan hewan mammalia atau usus halus (Gargova & Sariyska
2003).
tidak seimbang dengan kandungan fitatnya dan ada kemungkinan aktivitas enzim fitase dihambat oleh kandungan fitatnya yang tinggi.
Fitase juga merupakan enzim yang banyak digunakan sebagai bahan tambahan pada pakan ternak, yang dapat mengkatalisis pembebasan fosfat dari asam fitat yang menghasilkan mio-inositol, mio-inositol penta, tetra, tri, di dan monofosfat. Enzim ini sangat penting karena secara alami fosfor dalam pakan mengandung bentuk fitat yang tidak dapat dihidrolisis dalam sistem pencernaan tanpa tambahan enzim atau mikrobia mammalia yang menghasilkan fitase yang dapat digunakan sebagai probiotik (Purwadaria 2002).
Gen fitase dipilih untuk disisipkan ke dalam tanaman tebu karena gen ini menghasilkan enzim yang dapat mengubah senyawa fitat yaitu senyawa organik yang menyimpan unsur fosfat di dalam sel tanaman. Unsur fosfat yang tersimpan dalam fitat ini tidak dapat dimanfaatkan oleh tanaman. Namun jika senyawa fitat dihidrolisis akan menghasilkan ester yang berfosfat rendah dan melepaskan unsur fosfat anorganik. Gen fitase diharapkan dapat membuat tebu lebih efisien memanfaatkan fosfat, yang sebelumnya terikat dalam wujud senyawa organik di dalam jaringan tanaman dan tanah. Di dalam tanah senyawa fosfat organik mencapai hampir 50% yang seluruhnya tidak dapat dimanfaatkan oleh tanaman. Fosfat anorganik yang tersedia di dalam sel tanaman memberikan pengaruh positif pada proses pembentukan klorofil sehingga meningkatkan fotosintensis dan metabolisme tanaman tebu. Hasil fotosintesis ini disimpan menjadi gula tebu yang kadarnya disebut rendemen. Di samping itu, fitase juga meningkatkan ketersediaan mineral-mineral lainnya, seperti kalsium, magnesium dan kalium di dalam jaringan tanaman sehingga tanaman dapat mengurangi kebutuhan pupuk (Santosa 2002).
Konstruksi Vektor pBINPI-II EC
12
cara transformasi DNA ke dalam sel; 2) vektor harus mampu bereplikasi dalam sel inangnya (umumnya E. coli, Agrobacterium dan khamir) meskipun beberapa vektor kloning dapat terintegrasi ke dalam sel mamalia; dan 3) integrasi gen dapat dideteksi pada media seleksi (Freifelder 1987; Kleinsmith & Kish 1995).
Beberapa jenis vektor dapat digunakan sebagai vektor rekombinan di antaranya adalah plasmid, virus, dan cosmid. Pertama, plasmid yaitu molekul DNA berbentuk sirkuler yang mampu bereplikasi dalam sitoplasma bakteri secara bebas. Plasmid juga membawa gen-gen ketahanan terhadap antibiotik yang berguna sebagai penyeleksi sel-sel bakteri yang mengandung plasmid rekombinan. Kedua, virus biasanya berasal dari bacteriofage λ, DNA yang dapat disisipkan sekitar 15 kb. Vektor rekombinan bakteriofage ini dapat ditransformasi ke dalam sel bakteri. Ketiga, cosmid yaitu DNA plasmid yang juga memiliki vektor kloning λ. Cosmid memiliki satu atau lebih gen penyeleksi antibiotik dan membawa sisi cos dari λ. DNA sisipan yang dapat diklon ke dalamnya berukuran relatif besar (40-45 kb). Terakhir, shuttle vector ialah molekul DNA yang mampu bereplikasi di dalam dua jenis sel berbeda atau lebih. Beberapa jenis shuttle vector yang banyak digunakan ialah molekul DNA yang mampu bereplikasi dalam sitoplasma bakteri atau khamir (Kleinsmith & Kish 1995).
Menurut Sambrook (2001), plasmid yang dijadikan vektor harus berukuran kecil. Ukuran kecil memungkinkan untuk disisipi fragmen DNA asing yang besar, mudah dikenali dengan peta restriksi, menghasilkan jumlah salinan relatif lebih banyak dibandingkan dengan plasmid berukuran besar. Brown (1996) menyarankan agar ukuran plasmid tidak lebih dari 10 kb. Semakin besar ukuran molekul, semakin sulit dilakukan pemurnian dan manipulasi DNA.
Plasmid dapat dibedakan berdasarkan sifat utama yang dikode oleh gen-gen di dalamnya, yaitu 1) Plasmid F atau plasmid fertilitas yaitu hanya membawa gen tra yang berfungsi untuk melakukan transfer plasmid secara konjugasi; 2) Plasmid
menyebabkan patogenesis pada bakteri inang, misalnya plasmid Ti pada Agrobacterium. Plasmid Ti dapat menyebabkan penyakit crown gall pada
tanaman dikotil. Rekombinasi DNA saat ini banyak menggunakan plasmid alami yang dimodifikasi. Plasmid tersebut telah ditambah atau dikurangi gen-gen tertentunya dengan tujuan memudahkan rekombinasi. Dalam penelitian digunakan konstruksi gen kaset (Gambar 1) dalam plasmid pBINPI-II EC berukuran 11.810 bp (Lampiran 2). Plasmid pBINPI-II EC merupakan plasmid hasil konstruksi dan modifikasi yang dapat digunakan sebagai vektor bagi tanaman tebu. Plasmid ini memiliki signal peptide. Signal peptide merupakan suatu segmen yang terdiri dari 15–30 asam amino pada ujung N protein yang memungkinkan protein disekresikan melalui membran sel.
CaMV 35S SP phytase gene 900 bpappA of E. coli OCS nptII (Kan r )
EcoRI Asp718 BamHI/BglII SalI HindIII
Gambar 1. Konstruksi kaset gen fitase appA dalam vektor pBINPI-II EC. Keterangan: CaMV 35S: promotor dari virus Cauliflower mosaik 35S, SP: proteinase inhibitor II signal peptide, OCS: oktopin sintase (enhancer). Tanda panah menunjukkan situs enzim-enzim restriksi yang penting (Santosa et al. in press). Peta plasmid pBINPI-II EC disajikan dalam Lampiran 2.
Plasmid pBINPI-II EC membawa gen penyeleksi antibiotik nptII yang menyandikan enzim neomysin phosphotransferase (kanamisin). Kanamisin ber-guna sebagai penyeleksi di tingkat bakteri. Antibiotik ini tergolong amino-glikosida yang memiliki target ribosom bakteri. Golongan aminoamino-glikosida dapat mengikat sub unit kecil ribosom (30S) bakteri. Akibat pengikatan ini, penempelan sub unit besar ribosom (50S) menjadi terhalang sehingga protein tidak dapat ditranslasi. Bakteri yang membawa plasmid dengan penyeleksi kanamisin, menyebabkan enzim yang disandikannya aktif dan mampu memecah kanamisin sehingga bakteri menjadi resisten (Salyers & Whitt 1994).
Plasmid pBINPI-II EC dilengkapi dengan sekuen T-DNA, yaitu untaian DNA yang akan ditransfer ke dalam sel tanaman. Susunan gen-gen yang berada dalam T-DNA mulai dari batas kiri ke batas kanan ialah: 1) promotor Cauliflower Mosaic Virus 35S (CaMV 35S), 2) gen appA of E. coli dengan ukuran 900 bp
14
unik tersebut merupakan suatu segmen pada plasmid yang besarnya antara puluhan hingga ratusan pasang basa sebagai sekuen pengenal enzim restriksi. Sekuen pengenal ini tidak terdapat di bagian lain pada molekul plasmid. DNA sisipan biasanya diklon ke dalam sisi unik (Brown 1996).
Transformasi dengan Bantuan Agrobakterium
Agrobacterium tumefaciens merupakan bakteri aerob obligat Gram negatif
yang hidup alami di tanah dan banyak menyebabkan penyakit crown gall pada tanaman dikotil. Kemampuan untuk menyebabkan penyakit ini ada hubungannya dengan gen penginduksi tumor (tumor inducing / Ti) dalam sel bakteri (Beijersbergen dan Hooykaas 1993; Sheng & Citovsky 1996; Hiei et al. 1997).
Terdapat tiga komponen utama pada agrobakterium yang berperan dalam transfer DNA ke dalam sel tanaman (Sheng & Citovsky 1996). Komponen pertama ialah daerah DNA yaitu fragmen yang ditransfer ke sel tanaman. T-DNA terletak pada plasmid Ti (200 kb) dari agrobakterium yang diapit oleh sekuen berulang DNA (25 kb) sebagai batas T-DNA. Komponen kedua ialah daerah virulence (vir) yang berukuran 35 kb dalam plasmid Ti. Sedangkan menurut Beijersbergen dan Hooykaas (1993) gen vir berukuran sekitar 40 kb, letaknya bersebelahan dengan batas kiri T-DNA. Gen-gen vir ini terbagi atas 7 yaitu vir A, B, C, D, E, G, dan vir H. Gen vir mensintesis protein virulensi yang berperan menginduksi terjadinya transfer dan integrasi T-DNA ke dalam tanaman. Empat gen vir yang paling penting mensintesis protein virulensi ini ialah vir A, B, D dan G. Jika ada induser, vir A dan G yang terekspresi mampu mengaktifkan
chvB, pscA (atau exoC) dan att. Gen-gen tersebut terletak di kromosom
agrobakterium dan berfungsi untuk pelekatan bakteri ke dalam sel tanaman dengan membentuk senyawa protein β-1,2-glukan (Beijersbergen dan Hooykaas 1993).
Berdasarkan sifat alamiah agrobakterium tersebut, pada dasawarsa terakhir ini telah diupayakan pemindahan gen komersial untuk tanaman monokotil dengan cara menyisipkan gen tersebut ke daerah T-DNA.
Penyisipan T-DNA ke dalam plasmid Ti yang besar (200 kb) menghadapi masalah utama yaitu besarnya ukuran pTi sehingga menyulitkan dalam manipulasi dan menentukan tempat pemotongan yang khas pada plasmid Ti. Secara umum ada dua macam strategi menyisipkan DNA yaitu sistem vektor kointegrasi dan vektor ganda (Cramer & Radin 1990).
Walkerpeach dan Velten (1994) berpendapat bahwa syarat dari vektor ko-integrasi adalah 1) mempunyai tempat yang tepat untuk menyisipkan fragmen DNA, 2) memiliki gen penyeleksi antibiotik yang aktif pada E. coli maupun agrobakterium, 3) memiliki gen penanda untuk tanaman dan 4) memiliki ori yang berfungsi di E. coli tapi tidak di agrobakterium. Sistem vektor ganda (biner) membutuhkan 2 plasmid di dalam agrobakterium yaitu 1) plasmid vektor yang mengandung fragmen DNA, dan 2) plasmid penolong (helper) Ti yang menyediakan gen vir sebagai fasilitator transfer gen ke sel tanaman. Kedua plasmid dapat bereplikasi pada agrobakterium.
Perkembangan terakhir menunjukkan bahwa umumnya vektor yang digunakan untuk transformasi tanaman dikotil maupun monokotil adalah vektor ganda. Dengan menggunakan vektor penyisipan gen (kloning) menjadi lebih mudah, karena vektor yang mengandung batas T-DNA berukuran lebih kecil dari plasmid Ti yang sesungguhnya. Ukuran plasmid yang kecil memungkinkan lebih banyak penyisipan gen pada sisi unik enzim restriksi (Slamet-Loedin 1994).
Analisis Tanaman Transgenik
16
dilakukan mulai fase kalus hingga tanaman dewasa, dengan tidak merusak jaringan atau sel.
Integrasi gen sisipan pada kalus atau planlet hasil transformasi dapat dianalisa secara molekuler menggunakan teknik PCR (Polymerase Chain Reaction) yaitu amplifikasi fragmen spesifik dari DNA total genom. Keuntungan
teknik ini antara lain: cepat, DNA yang diperlukan sedikit, dapat dilakukan pada tahap dini dan teknik isolasi DNA sederhana. Amplifikasi DNA terjadi karena adanya enzim polimerase yang dihasilkan oleh bakteri termofilik Thermus aquaticus. Hal-hal yang menentukan keberhasilan amplifikasi antara lain adalah
desain primer dan suhu yang digunakan selama proses PCR. Sekuen primer yang tepat memungkinkan amplifikasi hanya terjadi tepat pada fragmen spesifik. Primer harus bersifat komplemen terhadap DNA target. Semakin pendek ukuran primer (8-mer) maka semakin tidak spesifik fragmen yang dihasilkan. Sebaliknya semakin panjang primer (20-mer) akan semakin spesifik fragmen yang dihasilkan. Ukuran primer yang lebih besar dari 30-mer sangat jarang digunakan. DNA target yang diamplifikasi hendaknya tidak lebih dari 3 kb, karena ukuran ideal untuk PCR ini kurang dari 1 kb (Brown 1996).
Selama amplifikasi dalam setiap siklus terjadi tiga perubahan suhu. Tahap pertama DNA didenaturasi, umumnya pada suhu 94 oC. Saat denaturasi, DNA yang semula utas ganda terurai menjadi utas tunggal karena ikatan hidrogennya lepas. Tahap kedua suhu diturunkan sehingga memungkinkan primer menempel (annealing) pada DNA cetakan. Tahap ketiga terjadi pemanjangan DNA (sintesis), biasanya pada suhu 72 oC sehingga memungkinkan enzim DNA poli-merase bekerja.
Menurut Brown (1996), optimasi suhu annealing sangat penting agar primer menempel pada DNA cetakan. Suhu annealing diduga berdasarkan suhu melting Tm antara primer dengan DNA cetakan. Suhu annealing biasanya lebih rendah 1–
Tempat dan Waktu Penelitian
Penelitian dilakukan di Laboratorium Mikrobiologi dan Bioteknologi Lingkungan Pusat Penelitian Lingkungan Hidup (PPLH) IPB dari bulan Februari 2003, Laboratorium Bioteknologi Tanaman Fakultas Pertanian IPB dari bulan Juni 2004, dan Laboratorium Molekuler PT. Saraswanti Indo Genetech di Bogor dari bulan Januari 2005 hingga bulan Mei 2005.
Metode Penelitian
Penelitian ini terdiri atas dua bagian yaitu: 1) Kultur jaringan tanaman tebu, dan 2) Transformasi tebu dengan gen fitase.
1. Kultur jaringan tanaman tebu
Sterilisasi dan penanaman eksplan
Eksplan tebu kultivar PA 117 dan PSJT 94-33 berasal dari daun muda dan sehat yang masih menggulung berumur 4–6 bulan. Pucuk tebu dipotong 20 cm di atas jaringan meristem. Secara aseptik, pucuk tebu dicelupkan ke dalam alkohol 70% lalu dibakar di atas api bunsen. Lapisan pucuk daun dibuka dan dibuang, diulang sebanyak tiga kali atau sampai kelihatan warna merah muda pada pucuk tebu. Media MS-I disiapkan lebih dahulu melalui sterilisasi dengan autoklaf 121
o
C selama 10 menit pada tekanan 1,5 atm. Selanjutnya untuk induksi kalus, bagian pucuk dipotong sepanjang 2–3 mm sebanyak 12 potong kemudian ditanam pada media MS-I (RNI 1999). Setiap petridis diisi 5-6 potong kemudian diinkubasi dalam ruang gelap bersuhu ±22 oC. Selama 1 bulan dapat diperoleh kalus dari jaringan parenkimatis yang mempunyai struktur kompak dan mampu berproli-ferasi. Untuk mempertahankan kalus dalam status embriogenik maka setiap 2-3 minggu kalus disubkulturkan pada media yang sama. Kalus embriogenik di-regenerasikan menjadi planlet pada media MS-II (P3GI 1999). Khusus untuk tebu kultivar PS 851 diberikan oleh RNI Cirebon dalam bentuk kalus sehingga siap digunakan untuk transformasi.
Regenerasi tanaman tebu
18
EC dan PA 117 pBINPI-II EC pada media optimasi regenerasi, dan regenerasi kalus transforman PS 851 pBINPI-II EC pada media Modifikasi P3GI.
Tahap pertama, sebanyak enambelas perlakuan optimasi media regenerasi kalus PA 117 dan PSJT 94-33 adalah kombinasi media yang terdiri dari media dasar MS-II (P3GI 1999) dengan penambahan zat pengatur tumbuh IAA (β -indoleacetic acid) dan dalapon (2,2-dichloropropionic acid). Secara berurutan,
perlakuan tersebut diberi prelabel R dan angka perbandingan dalam kurung menunjukan kombinasi IAA dan dalapon (mg/L). Label R1 (1:55), R2 (1:57), R3 (1:59), R4 (1:61), R5 (1,5:55), R6 (1,5:57), R7 (1,5:59), R8 (1,5:61), R9 (2:55), R10 (2:57), R11 (2:59), R12 (2:61), R13 (2,5:55), R14 (2,5:57), R15 (2,5:59), dan R16 (2,5:61). Kultur ditumbuhkan pada kondisi terang dengan suhu 22 oC. Setiap perlakuan diulang sebanyak 10 kali, pengamatan jumlah tunas dan daun dilakukan selama 4 minggu. Pada minggu berikutnya, kultur dipindahkan ke media cair yang sesuai dengan media optimum berdasarkan respon kultivar. Sebelum dipindahkan, bagian basalnya dipotong untuk induksi akar. Jumlah akar diamati selama 4 minggu.
Tahap kedua, regenerasi kalus transforman PA 117 pBINPI-II EC dan PSJT 94-33 pBINPI-II EC pada media optimasi regenerasi dilakukan berdasarkan hasil pada tahap pertama. Perbedaan respon kultivar terhadap kombinasi media mengakibatkan regenerasi kalus transforman kedua kultivar tersebut mengguna-kan media yang berbeda pula. Di bagian lain, regenerasi kalus transforman PS 851 pBINPI-II EC pada media Modifikasi (2 mg/L IAA + 59 mg/L dalapon) menggunakan media dengan penambahan glukosa 1%, asam sitrat dan asam askorbat masing-masing 2,5 mg/L (P3GI 1999).
Analisis data penelitian disusun menggunakan Rancangan Petak Terbagi (split plot experiment) dan Rancangan Acak Lengkap. Perhitungan statistik
menggunakan SAS v8.2 Statistics Software (SAS Institute, NC-USA) dengan prosedur General Linier Model. Uji lanjut signifikansi menggunakan uji beda nyata jujur (BNJ/Tukey) dengan α 5% (Steel & Torrie 1993).
faktor pengamatan berulang adalah rancangan petak terbagi. Model rancangan petak terbagi adalah:
Yijk = µ + ρk + αi + δik + βj + (αβ)ij + εijk
µ : Nilai rataan umum
ρk : Pengaruh kelompok (ulangan) ke-k (k = 1-10)
αi : Pengaruh perlakuan, yaitu kombinasi IAA dan dalapon pada media
regenerasi
δik : Galat a, interaksi kelompok dan perlakuan.
βj : Pengaruh minggu pengamatan media regenerasi ke-j (j = 1, 2, 3, 4)
(αβ)ij: Pengaruh interaksi antara minggu dan perlakuan pada media
regenerasi.
εijk : Galat percobaan (b)
Yijk : Nilai pengamatan
Analisis statistik regenerasi kalus transforman PA 117 dan PSJT 94-33 pada media hasil optimasi dilakukan menggunakan Rancangan Acak Lengkap dengan 10 ulangan dan waktu pengamatan 4 minggu. Rancangan yang sama digunakan pada hasil regenerasi kalus transforman PS 851 dalam media Modifikasi P3GI. Variabel respon yang dianalisis adalah jumlah tunas, daun dan akar yang terbentuk. Model rancangan acak lengkap adalah:
Yij = µ + αi + εij
µ : Nilai rataan umum
αi : Pengaruh minggu ke-i (i = 1, 2, 3, 4)
εij : Galat percobaan pada minggu ke-i ulangan ke-j (j = 1-10)
Yij :Nilai pengamatan
2. Transformasi tebu dengan gen fitase
Bagian ini terdiri atas tiga tahap yaitu: 1) transformasi kalus tebu dengan bantuan A. tumefaciens GV2260 pBINPI-II EC; 2) isolasi DNA genom total; dan 3) analisis gen fitase dengan PCR.
Transformasi kalus tebu dengan bantuan A. tumefaciens GV2260 pBINPI-II EC
20
Santosa et al. (2004); dan 4) metode Modifikasi yang merupakan modifikasi antara metode Minarsih (2003) dan metode Santosa et al. (2004).
Metode pertama, eksplan berasal dari jaringan meristem pucuk daun tebu. Sebelum kokultivasi eksplan diberi antioksidan (L-sistein 40 mg/L; asam askorbat 15 mg/L; perak nitrat 2 mg/L). Kultur A. tumefaciens ditumbuhkan pada media Luria Broth (tripton 10 g/L, ekstrak yeast 5 g/L, NaCl 5 g/L, sukrosa 5 g/L, dan
H2O sampai tera) yang mengandung rifampisin 100 mg/L sampai diperoleh OD620
(optical density) = 0,6 kemudian disentrifugasi selama 5 menit pada 5000 rpm. Pelet yang didapatkan dicuci dengan MS cair yang ditambah dengan antioksidan. Kokultivasi selama 10 menit dan selanjutnya eksplan dipindahkan pada media MS-I yang ditambah dengan antioksidan, dan kemudian ditutup dengan kertas saring untuk mengurangi infeksi agrobakterium. Kokultur ini dilakukan selama 3 hari kemudian dicuci dengan aquades steril dan cefotaxime 500 mg/L. Setelah pencucian, eksplan ditanam pada media MS-I padat yang mengandung kanamisin 50 mg/L dan cefotaxime 500 mg/L selama 10 hari. Pucuk daun yang tidak ditumbuhi oleh agrobakterium dipindahkan pada media seleksi MS-I yang mengandung kanamisin 100 mg/L. Komposisi media metode ini disajikan dalam Lampiran 3.
Kedua, metode Minarsih (2003) hampir sama dengan metode yang dilakukan oleh Enriquez-Obregón et al. (1997). Namun eksplan yang digunakan adalah eksplan kalus dan densitas kultur A. tumefaciens untuk kokultivasi yaitu OD620 =
0,6 dengan pengenceran sepuluh kalinya. Komposisi media metode ini disajikan dalam Lampiran 4.
Ketiga, metode Santosa et al. (2004), kalus tebu dimasukkan ke dalam 2 mL eppendorf dan diinokulasi dengan 0,5 mL kultur suspensi A. tumefaciens (OD578 =
kertas saring steril, kemudian kalus dipindahkan pada ke media MS cair sebanyak 25 mL yang mengandung 0,5 g/L kasein hidrolisat, cefotaxime 1000 mg/L dan inkubasi pada kondisi gelap 28 oC sambil dishaking pada 60 rpm selama 2 jam. Perlakuan diulang kembali. Kalus ditransfer ke dalam 30 mL media MS yang mengandung 0,5 mg/L kasein hidrolisat, 500 mg/L cefotaxime, lalu diinkubasi dengan kondisi gelap pada 28 oC sambil dishaking 60 rpm selama 2 hari. Jika ada pertumbuhan agrobakterium media diganti dengan media baru. Kemudian dipindahkan pada media MS yang mengandung 0,5 mg/L kasein hidrolisat, 500 mg/L cefotaxime, 100 mg/L kanamisin dan diinkubasi selama seminggu untuk memastikan tidak ada pertumbuhan bakteri. Setelah seminggu kalus dikeringkan lagi dengan kertas saring steril dan dipindahkan ke media MS padat yang mengan-dung 500 mg/L cefotaxime, 100 mg/L kanamisin, dengan kondisi gelap 28 oC selama 2 minggu atau lebih untuk memastikan tidak ada pertumbuhan agrobakterium. Selanjutnya, kalus ditanam pada media MS-I dengan kanamisin 100 mg/L untuk mendapatkan struktur yang kompak dan mampu berproliferasi. Komposisi media metode ini disajikan dalam Lampiran 5.
Keempat, metode Modifikasi (Minarsih 2003 dan Santosa et al. 2004) yang dilakukan adalah memodifikasi perlakuan pasca pencucian A. tumefaciens selama dua jam yang dilakukan oleh Santosa et al. (2004) kemudian tidak dipindahkan pada perlakuan inkubasi media cair selama satu minggu melainkan langsung dipindahkan pada media padat yang mengandung cefotaxime 500 mg/L (Minarsih 2003). Komposisi media metode ini disajikan dalam Lampiran 6.
Isolasi DNA genom total
DNA total diisolasi dari kalus yang berumur 4 minggu setelah transformasi pada kultivar PA 117, PSJT 94-33, dan PS 851 dengan metode DNeasy Plant Mini Kit (Qiagen). Setelah preparasi kit isolasi, sebanyak 100 mg sampel digerus menggunakan nitrogen cair dalam mortar untuk menghancurkan dinding sel menjadi bubuk. Segera setelah sampel berbentuk bubuk dilanjutkan menurut petunjuk kit isolasi.
Analisis gen fitase dengan PCR
22
gen fitase EC1: 5’–CAGGCTCTATCCGCTAATCG–3’ dan EC3: 5’– GGCGCGGTGGGGCAATAATC–3’ yang setiap reaksi diatur sebagai berikut; pre-start 95 oC selama 10 menit, denaturasi pada 95 oC selama 30 detik, annealing pada 50 oC selama 30 detik dan sintesis pada 72 oC selama 1 menit, dan stop 72
o
C selama 5 menit. Jumlah setiap campuran reaksi sebanyak 20 µL yang terdiri dari 10 µL Master Mix, 1 µL [0,5 µM/µL] masing-masing primer spesifik untuk gen fitase, 1 µL DNA [0,5 µg/mL] dari tanaman transforman dan kontrol dan 7 µL ddH2O. Reaksi dijalankan sebanyak 40 siklus setelah pre-start menggunakan
mesin PCR Eppendorf Mastercycler Personal. Kontrol negatif (tanpa DNA cetakan) selalu digunakan tiap kali mengerjakan PCR. Selanjutnya DNA hasil amplifikasi dimasukkan dalam sumur pada gel agarose 2% (b/v). Elektroforesis dijalankan selama 30 menit pada 100 volt dalam bufer 1x TAE (4.84 g Tris-base; 0.186 g Na2EDTA; 1,142 ml asam asetat glasial; 1 L dH2O) pada perangkat
elektroforesis mini submarine. Kemudian gel direndam dalam ethidium bromida 10 µg/mL selama 10 menit lalu dibilas dengan dH2O. Gel dilihat pada GelDoc
Kultur Jaringan Tanaman Tebu
Sterilisasi dan penanaman eksplan
Sterilisasi bahan tanaman (eksplan) tebu dalam kultur jaringan ditujukan untuk memperoleh kalus embriogenik steril. Kalus ini akan digunakan untuk mendapatkan media yang optimum dalam regenerasi tanaman tebu, dan untuk transformasi gen fitase ke dalam genom tebu melalui A. tumefaciens GV2260. Cara sterilisasi yang dipakai dalam percobaan ini berturut-turut adalah dengan pencelupan alkohol 70%, pembakaran di atas nyala api spiritus, dan pengambilan eksplan yang agak jauh dari pucuk apikal sehingga eksplan menjadi steril, segar dan dapat tumbuh dengan baik dalam media MS-I (RNI 1999).
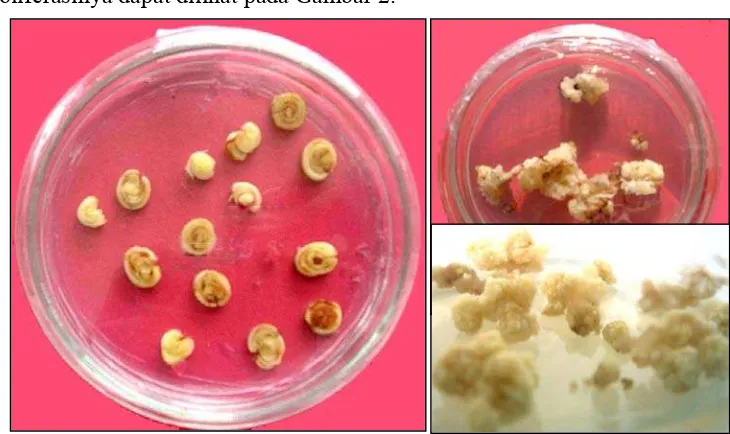
Kalus yang diinduksi dari eksplan daun menggulung (jaringan meristem) mulai terbentuk setelah 4 minggu pada media kultur MS-I. Berdasarkan teksturnya kalus yang dihasilkan bersifat kompak, viabel atau kombinasi antara keduanya. Umumnya tingkat viabilitas mengikuti tingkat konsentrasi auksin yang digunakan dan dalam percobaan ini auksin yang dipakai yaitu 2,4-D dengan konsentrasi 3 mg/L mampu membentuk kalus yang kompak. Warna kalus beragam mulai dari putih dan putih kekuningan. Inisisasi kalus dan hasil proliferasinya dapat dilihat pada Gambar 2.
Gambar 2. Inisiasi dan proliferasi kalus. a) Eksplan jaringan meristem b) Inisiasi kalus c) Hasil proliferasi setelah 4 minggu.
a
b
24
Regenerasi tanaman tebu kultivar PA 117 dan PSJT 94-33
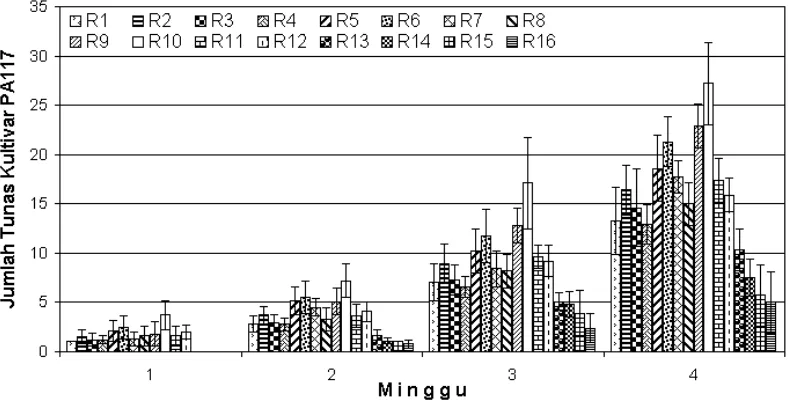
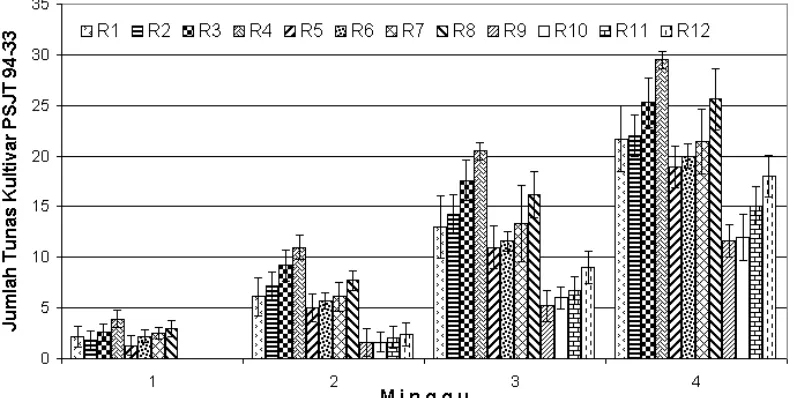
Regenerasi tanaman tebu nontransforman kultivar PA 117 dan PSJT 94-33 bertujuan untuk mendapatkan media yang optimum untuk induksi tunas, daun, dan akar. Media yang digunakan adalah MS yang ditambahkan dengan zat pengatur tumbuh IAA dan dalapon dalam beberapa kombinasi konsentrasi. Induksi tunas dan daun dilakukan pada media padat. Jumlah tunas dan daun yang tumbuh sampai dengan minggu ke-4 dari kultivar PA 117 dapat dilihat berturut-turut pada Gambar 3 dan 4. Hasil optimasi media regenerasi untuk induksi tunas dan daun dapat dilihat pada Gambar 5. Sedangkan induksi akar dilakukan pada media cair dengan waktu pengamatan sampai dengan minggu ke-4. Jumlah akar yang terbentuk ditampilkan dalam Gambar 6.
Gambar 3. Grafik rataan jumlah tunas tebu kultivar PA 117 yang terbentuk sampai dengan minggu ke-4.
Gambar 4. Grafik rataan jumlah daun tebu kultivar PA 117 yang terbentuk sampai dengan minggu ke-4.
26
R1 R2 R3 R4
R5 R6 R7 R8
R9 R10 R11 R12
R13 R14 R15 R16
Gambar 6. Grafik rataan jumlah akar tebu kultivar PA 117 yang terbentuk sampai dengan minggu ke-4 pada media regenerasi R10.
Rata-rata jumlah akar yang terbentuk pada kultivar PA 117 yang diamati sampai dengan minggu ke-4 dapat dilihat pada Gambar 6. Pada minggu pertama induksi akar langsung terbentuk walaupun jumlahnya masih sedikit. Jumlah akar terbanyak berada pada minggu ke-4 ini berarti bahwa media induksi akar yang digunakan yaitu media cair dengan komposisi IAA 2 mg/L, dalapon 57 mg/L mampu menginduksi akar kultivar PA 117. Media cair yang digunakan sangat efektif untuk induksi akar sehingga akar yang terbentuk sangat kuat dan kokoh seperti yang ditunjukkan pada Gambar 7.
28
Kedua jenis zat pengatur tumbuh yaitu IAA dan dalapon berperan sangat penting dalam regenerasi kalus menjadi planlet. IAA termasuk dalam kelompok auksin alamiah yang banyak terdapat pada tanaman. Sedangkan dalapon secara komersial termasuk dalam kelompok herbisida. Penambahan IAA dan dalapon dalam regenerasi kalus tanaman tebu kultivar PA 117 terbukti mampu meng-induksi tunas, daun, dan akar. Konsentrasi kedua zat pengatur tumbuh adalah sebanyak 2 mg/L IAA, dan 57 mg/L dalapon.
Gambar 8. Grafik rataan jumlah tunas tebu kultivar PSJT 94-33 yang terbentuk sampai dengan minggu ke-4.
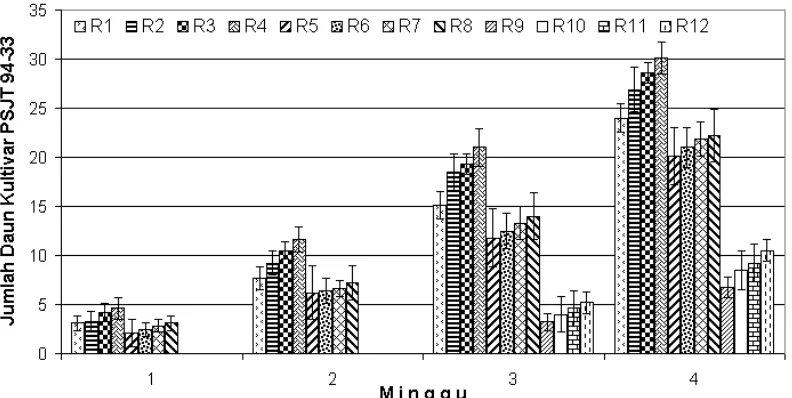
Gambar 9. Grafik rataan jumlah daun tebu kultivar PSJT 94-33 yang terbentuk sampai dengan minggu ke-4.
30
Gambar 10. Hasil optimasi media regenerasi tanaman tebu kultivar PSJT 94-33 sampai dengan minggu ke-4. Keterangan nomor sama seperti yang dijelaskan dalam Gambar 5. Untuk nomor R13-R16 tidak disajikan karena kalus tidak mampu tumbuh. Optimasi media terbaik tampak pada gambar bernomor R4.
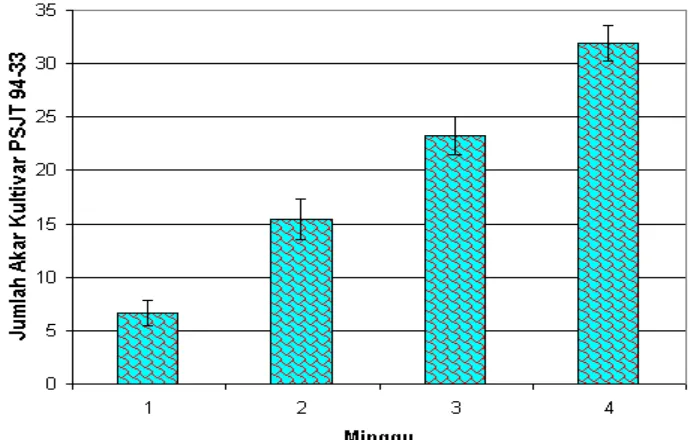
Gambar 11. Grafik rataan jumlah akar tebu kultivar PSJT 94-33 yang terbentuk sampai dengan minggu ke-4 pada media regenerasi R4.
R1 R2 R3 R4
R5 R6 R7 R8
Jumlah akar kultivar PSJT 94-33 yang terbentuk pada media cair dengan kombinasi IAA 1 mg/L, dalapon 61 mg/L sangat nyata pada minggu pengamatan yang ke-4. Pada minggu pertama pembentukan akar sangat sedikit dan masih sangat lemah dibandingkan dengan minggu keempat yang struktur akarnya lebih kokoh, seperti ditunjukan pada Gambar 12.
Gambar 12. Hasil induksi akar tebu kultivar PSJT 94-33 pada media regenerasi R4.
Secara menyeluruh, regenerasi tanaman tebu kultivar PA 117 dan PSJT 94-33 dengan kombinasi IAA dan dalapon berhasil dilakukan dengan baik. Respon regenerasi yang optimum antar kedua kultivar berbeda pada kombinasi kedua zat pengatur tumbuh yang digunakan. Dari laporan penelitian yang ada ternyata untuk pertumbuhan dan perkembangan kultur jaringan diperlukan komposisi atau konsentrasi zat pengatur tumbuh yang berbeda untuk satu kultivar dengan kultivar lain dari suatu jenis tanaman (Wattimena 1992).
Regenerasi kalus transforman
32
perlakuan pada saat transformasi. Selain itu, respon masing-masing kultivar terhadap zat pengatur tumbuh pada media regenerasi juga berbeda karena genotip sumber jaringan atau organ yang digunakan (Wattimena et al. 1992).
Gambar 13. Regenerasi tebu transforman PA 117, PSJT 94-33, dan PS 851.
Rendahnya hasil regenerasi kalus transforman menjadi tanaman hijau sangat berbeda dengan kalus nontransforman. Chung (1992) dan Chen (1983) menyatakan proporsi tanaman hijau dan albino tersebut selain tergantung pada genotipa tanaman juga pada kondisi kultur, misalnya suhu inkubasi. Dari studi molekuler, gen inti maupun sitoplasmik dan interaksinya mungkin mempengaruhi produksi tanaman albino.
Rataan tunas transforman kultivar PA 117 yang terbentuk selama pengamatan sampai dengan minggu keempat disajikan dalam Gambar 14. Sejak minggu pertama, telah terbentuk tunas meskipun dalam jumlah sangat sedikit dengan struktur yang lemah. Struktur tunas yang kokoh mulai terbentuk pada minggu keempat. Hal ini diduga karena kemampuan regenerasi kalus transforman yang berkurang akibat sub kultur yang dilakukan.
Gambar 14. Grafik rataan jumlah tunas transforman kultivar PA 117 pada media regenerasi R10.
34
Gambar 16. Grafik rataan tunas transforman yang terbentuk dari regenerasi kalus transforman kultivar PS 851 pada media regenerasi Modifikasi P3GI.
Transformasi Tebu dengan Gen Fitase
Transformasi tanaman tebu dengan gen fitase dilakukan menggunakan empat metode transformasi, yaitu: Metode Enriquez-Obregón et al. (1997), metode Minarsih (2003), metode Santosa et al. (2004) dan metode Modifikasi dari metode Minarsih (2003) dan Santosa et al. (2004). Transformasi gen fitase ke dalam genom tebu menggunakan eksplan jaringan meristem dan kalus. Keempat metode transformasi dan hasil transformasinya disajikan pada Tabel 1.
Tabel 1. Empat metode transformasi gen fitase pada eksplan tebu
Pada metode Enriquez-Obregón (1997), eksplan yang digunakan adalah jaringan meristem dari tiga kultivar tanaman tebu (PSJT 94-33, CB 6979 dan BR 194). Pada tahapan setelah kokultivasi eksplan dengan A. tumefaciens terjadi kontaminasi agrobakterium pada semua eksplan. Usaha untuk menghilangkan kontaminan dilakukan pencucian dengan aquades dan media MS-I yang mengandung cefotaxime 500–750 mg/L secara berulang-ulang, tetapi kontaminan tidak dapat dihilangkan. Pengaruh selanjutnya karena pencucian berulang-ulang tersebut, maka eksplan jaringan meristem mengalami kerusakan yang mengakibatkan daya tumbuhnya berkurang.
Metode kedua yang dilakukan yaitu metode Minarsih (2003). Tahapan kerja metode ini hampir sama dengan metode Enriquez-Obregón (1997) perbedaannya pada sumber eksplan yang digunakan yaitu kalus dan adanya pengenceran kultur A. tumefaciens sebelum kokultivasi. Hasil transformasi pada media seleksi dengan
metode ini disajikan pada Gambar 17a dan b. Pada tahapan kerja yang dilakukan, kontaminan agrobakterium dapat dihilangkan dengan pencucian menggunakan media MS-I yang mengandung cefotaxime 500 mg/L dan kalus sebagai eksplannya tidak mudah rusak karena struktur kalusnya lebih kompak dibandingkan dengan eksplan pada metode yang pertama. Pengenceran kultur A. tumefaciens yang dilakukan sebelum kokultivasi menyebabkan makin kecilnya
kemungkinan gen fitase masuk ke dalam genom tanaman, hal ini nampak pada kalus transforman yang dapat tumbuh di media seleksi sekitar 50%.
36
transforman yang bebas dari kontaminan bakteri sebesar 100%. Akan tetapi metode ini memiliki kendala yaitu pada saat pencucian dengan cara dishaking bila tidak dilakukan secara hati–hati dapat menyebabkan over growth population yang penanganannya akan lebih sulit. Pada penelitian yang dilakukan, kontaminan A. tumefaciens masih tinggi sekitar 50% hal ini disebabkan oleh kondisi lingkungan
kultur yang mempengaruhi pertumbuhan A. tumefaciens. Optical Density (OD) A. tumefaciens yang diinkubasi selama tujuh jam OD578 = 1,9. Sedangkan Santosa et
al (2004) mendapatkan OD578 = 0,5 untuk inkubasi selama tujuh jam. Hal ini
menunjukkan bahwa kecepatan pertumbuhan, jumlah sel yang ditumbuhkan dan kecepatan agitasi pada masing–masing kondisi kultur berbeda. Dengan demikian metode ini dapat dipakai dengan mempertimbangkan OD dari kultur A. tumefaciens yang akan digunakan pada saat kokultivasi.
Gambar 17. Kalus tebu transforman yang dihasilkan dari beberapa metode transformasi. Keterangan: a & b = Minarsih pada kultivar BR 194 & PA 183; c & d = Santosa pada kultivar PSJT 94-33 & PA 183; e, f & g = Modifikasi pada kultivar PSJT 94-33, PA 117 & PS 851.
a b
c d
e f
38
Analisis Gen Fitase dengan PCR
Verifikasi introduksi gen fitase ke dalam genom tanaman tebu hanya dapat dilakukan pada kalus. Hal ini disebabkan karena kesulitan untuk meregenerasikan kalus transforman menjadi planlet utuh. Setelah isolasi DNA dilakukan, analisis gen fitase dilakukan amplifikasi gen fitase dengan teknik PCR menggunakan primer EC1 dan EC3 berhasil dilakukan. Pada Gambar 18 ditampilkan hasil elektroforesis produk PCR gen fitase.
Gambar 18. Hasil PCR gen fitase dari kalus transforman yang berumur 4 minggu setelah transformasi. Keterangan: M = Marker 100 bp InvitrogenTM, K+ = Kontrol positif pBINPI-II EC, 1 = Kultivar PSJT 94-33, 2 = Kultivar PA117, 3 = Kultivar PS 851, 4 = Kultivar PS 851 (negatif), 5 = Kultivar PA117 (negatif), K─ = Kontrol negatif, tanpa DNA cetakan.
Pada Gambar 18 di atas menunjukkan bahwa ketiga kalus transforman terbukti memiliki gen fitase dengan ukuran fragmen hasil amplifikasi kurang lebih 900 bp (sumur gel 1-3). Beberapa kalus lain yang diuji hasilnya negatif (sumur gel 4 dan 5) mungkin disebabkan oleh kualitas hasil isolasi DNA, antara lain karena jumlah kalus sedikit sehingga DNA yang dihasilkan terlalu sedikit.
2072 bp 1500
900 600
100
K-Simpulan
Simpulan penelitian ini adalah:
1. Optimasi media regenerasi untuk kultivar PA 117 dan PSJT 94-33 berbeda. Media regenerasi untuk induksi tunas, daun, dan akar dari kalus kultivar PA 117, dan untuk induksi tunas dan akar kalus kultivar PSJT 94-33 berturut-turut adalah R10 (IAA 2 mg/L, dalapon 57 mg/L) dan R4 (IAA 1 mg/L, dalapon 57 mg/L).
2. Regenerasi kalus transforman tiga kultivar PA 117, PSJT 94-33, dan PS 851 berbeda. Kalus kultivar PSJT 94-33 pada media R4 mampu membentuk tunas dan daun, sedangkan kultivar PA 117 (pada media R10) dan PS 851 (pada media Modifikasi P3GI) hanya membentuk tunas.
3. Metode transformasi terbaik yang digunakan adalah metode Santosa dan Modifikasi.
4. Uji keberadaan gen fitase dalam kalus transforman berhasil diamplifikasi menggunakan primer spesifik dengan ukuran fragmen hasil PCR sebesar ± 900 bp.
Saran
DAFTAR PUSTAKA
Ahloowalia BS, Maretzki A. 1983. Plant regeneration via somatic embryogenesis in sugarcane. Plant Cell Reports 2:19-20.
Apriyanti A. 1990. Percobaan sterilisasi jaringan tebu (Saccharum officinarum L.) var. F 154. [skripsi]. Bogor: Institut Pertanian Bogor.
Arencibia A, et al. 1997. Transgenic sugarcane plants resistant to stem-borer attack. Mol Breed 3:247-255.
Arencibia A, et al. 1998. An efficient protocol for sugarcane (Saccharum spp.) transformation mediated by Agrobacterium tumefaciens. Transgenic Res 7:213-222.
Aswidinoor H. 1995. Transformasi gen: sumber baru keragamaan genetik dalam pemuliaan tanaman. Zuriat 6:56-67.
Beijersbergen A, Hooykaas PJL. 1993. Transkingdom promiscuty similarities between T-DNA transfer by A. tumefaciens and bacteria conjugation. [disertation]. pp 9-26.
Blanco MA, Magdalena SN, Ramiro C, Nadina N. 1999. Storage protein in sugarcane: an interesting exception in monocots. Plant Cell Tissue Organ Cult 59:217-218.
Bower R, Birch RG. 1992. Transgenic sugarcane plants via microprojectile bombardment. Plant J 2:409-416.
Brown TA. 1996. Gene cloning: an introduction. Chapman & Hall.
Chen Y. 1983. Cell And Tissue Culture Techniques for Cereal Crop Improvement. Proceedings of a Workshop Cosponsored by the Institute of Genetics, Academia Sinica and the International Rice Institute. China: Sci. Pr. pp 11-26.
Chung GS. 1992. Anther culture for rice improvement in Korea. In: Zheng K, Murashige T (Eds.). Anther culture for rice breeders. Seminar and training for rice anther culture at Hangzhou, China. pp 8-37.
Cramer CL, Radin DN. 1990. Molecular biology of plants. In: Nakas JP, Hagedorn C (Eds.). Biotechnology of Plant Microbes Interaction. New York: McGraw-Hill. pp 1-49.
Eechout W, de Paepe M. 1994. Total phosphorus, phytate-phosphorus and phytase activity in plant foodstuff. Feed Sci Tech 47:19-29.
Erwidodo. 2002. Getting sugar policy right. Jakarta: Dewan Ketahanan Pangan. Brief Policy 1-7.
Falco MC, Neto AT, Mendes BMJ, Arias FJZ. 1996. Histological characterization of in vitro regeneration of Saccharum sp. R Bras Fisiol Veg 8:93-97.
Freifelder D. 1987. Molecular Biology. Boston: Jones and Bartlett Pubs. Inc. Gallo MM, Irvine JE. 1993. Physical and biological factors affecting transient
GUS expression in sugarcane following particle bombardment. Plant Physiol 102 Suppl 165.
Gargova S, Sariyska M. 2003. Effect of culture conditions on the biosynthesis of Aspergillus niger phytase and acid phosphatase. Enzyme Microbial Tech 32:231-235
Greiner R, Konietzny U, Jany KD. 1993. Purification and characterization of two phytases from Escherichia coli. Arch Biochem Biophys 303:107-113.
Gunawan LW. 1992. Teknik kultur jaringan tumbuhan. Bogor: Institut Pertanian Bogor, Pusat Antar Universitas Bioteknologi.
Hadi S, Sutrisno. 2003. Ikhtisar angka perusahaan pada masa giling tahun 2003. Pasuruan: P3GI.
Hartatik S. 2000. Studi genetik plasma nutfah tebu (Saccharum spp) berdasarkan penanda morfologi, agronomi dan isozim [disertasi]. Bogor: Institut Pertanian Bogor.
Hauptmann et al. 1986. Expression of electroporated DNA in gramineous monocots and dicotyledonous species. Int Congr Plant Tissue Cell Cult. 16 Meet 18.
Heinz DJ, Krishnamurthi M, Nickell LG, Maretzki A. 1977. In: Reinert J, Bajaj YPS (Eds.). Applied and fundamental aspects of plant cell, tissue and organ culture. Berlin: Springer.
Hidayat D. 29 Sep 2002. Tawaran manis tebu transgenik. Tempo:11 (kolom 1-5). Hiei Y, Otha S, Komari T, Kumashiro T. 1994. Efficient transformation of rice
(Oryza sativa L.) mediated by Agrobacterium and sequence analysis of the boundaries of the T-DNA. Plant J 6:271–282.
Hiei Y, Komari T, Kubo T. 1997. Transformation of rice mediated by Agro-bacterium tumefaciens. Plant Mol Biol 35:205–218.
[HKTI] Himpunan Kerukunan Tani Indonesia. 1999. Permasalahan produksi dan tataniaga gula di akhir abad XX. Gula Indonesia XXIV:7-11.
42
Kleinsmith LJ, Kish VM. 1995. Principles of cell and molecular biology. New York: Harper Coliins College Pubs.
[KCM] Kompas Cyber Media. 2005. Menperindag siap dipanggil polisi. http://www.kompas.co.id. [15 Agustus 2005].
Liu MC. 1981. In vitro methods applied to sugarcane improvement. In: Thorpe TA (Eds.). Plant Tissue Culture Methods and Application in Agriculture. New York: Academic Pr. p 23.
Lolas MG, Markakis G. 1977. The phytase of nary beans (Phaseolus vulgaris). J Food Sci 42:1094–1101.
Minarsih H. 1999. Transformasi melalui Agrobacterium pada tanaman monokotil dan prospeknya pada tanaman tebu. Bul P3GI 151:13-23.
Minarsih H. 2003. Rekayasa genetik tebu (Saccharum officinarum L.) untuk toleransi kekeringan. [laporan riset]. Riset Unggulan Terpadu VIII Bidang Bioteknologi. Kementerian Riset dan Teknologi RI Lembaga Ilmu Pengetahuan Indonesia.
Murashige T. 1978. In: Frontiers of plant tissue culture. Thorpe TA, (Eds.). Canada: University of Calgary, Calgary.
Naik GR. 2001. Sugarcane biotechnology. Enfield NH: Science Pub.
Nonato RV, Mantelatto PE, Rossel CEV. 2001. Integrated production of bio-degradable plastic, sugar and ethanol. App Microbiol Biotechnol 57:1-5. [P3GI] Pusat Penelitian Perkebunan Gula Indonesia. 1997. Data produksi gula
tahun giling 1997. Pasuruan.
[P3GI] Pusat Penelitian Perkebunan Gula Indonesia. 1999. Petunjuk pembuatan media untuk kultur jaringan tebu. Pasuruan.
Patrau JM. 1989. By-products of the cane sugar industry: an introduction to their industrial utilization. Sugar Series 11. Amsterdam: Elsevier Science.
Pierik RLM. 1987. In vitro culture of higher plants. Dordrecht: Martinus Nijhoff Pubs.
Purwadaria T. 2002. Application of hydrolityc enzymes to improve the quality of feed. Third International Training Course on Advances in Molecular Biology Techniques to Assess Microbial Diversity, Bogor 17-27 Sep 2002.
[RNI] Rajawali Nusantara Indonesia. 1999. Metode kultur jaringan. Puslitagro, PT Pabrik Gula RNI II. Cirebon.
Sambrook J, Russell DW. 2001. Molecular cloning. A laboratory manual. Ed ke-3. New York: Cold Spring Harbor Laboratory Pr.
Santosa DA. 24 Sep 2002. IPB meneliti tebu transgenik. Kompas:11 (kolom 1-5). Santosa DA, Hendroko R, Farouk A, Greiner R. 2004. A rapid and high efficient
method for transformation of sugarcane callus. Mol Biotech 28:113-119. Santosa DA, Hendroko R, Farouk A, Greiner R. Agrobacterium-mediated
transformation of sugarcane (Saccharum officinarum L.) with bacterial phytase gene. Sugar Cane International. In press.
Sastrowijono S. 1991. Aspek fisiologis dalam mikropropagasi tebu. Di dalam: Diktat pelatihan penanganan mikropropagasi tanaman tebu, Pasuruan 4-9 Feb 1991. Pasuruan: P3GI.
Sheng J, Citovsky V. 1996. Agrobacterium-plant cell DNA transport: have virulence proteins will travel. Plant Cell 8:1699-1710.
Slamet-Loedin IH. 1994. Transformasi genetik beberapa tanaman: beberapa aspek penting. Hayati 1:66-67.
Soeponohardjo. 1997. Produktivitas gula Indonesia vs dunia. Majalah Gula Indonesia XXII:51-53.
Steel RGD, Torrie JH. 1993. Prinsip dan pendekatan statistika: suatu pendekatan biometrika. Jakarta: PT. Gramedia.
Sugiyarta E. 1991. Persyaratan laboratorium dan penetapan media pada teknik kultur jaringan tebu. Di dalam: Diktat pelatihan penanganan mikropropagasi tanaman tebu, Pasuruan 4-9 Feb 1991. Pasuruan: P3GI.
Suryowinoto M. 1991. Budidaya jaringan terobosan bermanfaat dalam biotekno-logi. Yogyakarta: Universitas Gadjah Mada, Fakultas Biobiotekno-logi.
Trigiano RN, Dennis JG. 2000. Plant tissue culture concepts and laboratory exercises. Ed ke-2. Florida: CRC Pr., Boca Raton.
Uchimiya H, Handa T, Brar DS. 1989. Transgenic plant. J Biotechnol 12:1-20. Walkerpeach CR, Velten J. 1994. Agrobacterium mediated gene transfer to plant
cells: cointegrated and binary vector system. Plant Mol Biol 12:1-19.
Wall MK, Birch RG. 1997. Genes for albicidin biosynthesis and at least 69 kb in the genome of Xanthomonas albilineans. Lett Appl Microbial 24:256-260. Wattimena et al. 1992. Bioteknologi tanaman. Bogor: Pusat Antar Universitas
44
Lampiran 1. Komposisi media MS untuk induksi kalus dan regenerasi planlet yang telah dimodifikasi.