RESPONS PERTUMBUHAN TANAMAN DAN KONSENTRASI
RANTAI PANJANG POLYISOPRENOID TERHADAP
VARIASI NAUNGAN DAN SALINITAS PADA
MANGROVE
Sonneratia alba
Smith.SKRIPSI
EVAN KHAROGI SINULINGGA 111201103/BUDIDAYA HUTAN
PROGRAM STUDI KEHUTANAN
FAKULTAS PERTANIAN
LEMBAR PENGESAHAN
Judul Penelitian : Respons Pertumbuhan Tanaman dan Konsentrasi Rantai Panjang Polyisoprenoid terhadap Variasi Naungan dan Salinitas pada Mangrove Sonneratia alba Smith.
Nama : Evan Kharogi Sinulingga
NIM : 111201103
Disetujui oleh, Komisi Pembimbing
Mohammad Basyuni, S.Hut, M.Si, Ph.D Dr. Ir. Yunasfi, M.Si. Ketua Anggota
Mengetahui,
ABSTRAK
EVAN KHAROGI SINULINGGA. Respons Pertumbuhan Tanaman dan Konsentrasi Rantai Panjang Polyisoprenoid terhadap Variasi Naungan dan Salinitas pada Mangrove Sonneratia alba Smith. Dibimbing oleh MOHAMMAD BASYUNI dan YUNASFI.
Tumbuhan mangrove memiliki kemampuan khusus untuk beradaptasi dengan kondisi lingkungan yang ekstrim. Kondisi ini memungkinkan mangrove untuk menghasilkan metabolit sekunder sebagai pertahanan kimia untuk kehidupan mereka. Polyisoprenoid merupakan salah satu metabolit sekunder yang belum diketahui fungsinya. Tujuan penelitian ini adalah untuk mengukur respons pertumbuhan semai S. alba pada variasi naungan dan tingkat salinitas dan pengaruhnya terhadap konsentrasi rantai panjang polyisoprenoid. Penelitian ini dilaksanakan pada Agustus sampai Desember 2014 menggunakan Rancangan Acak Lengkap Faktorial dengan perlakuan naungan 0%, 25%, 50%, dan 75% dengan tingkat salinitas 0%, 1,5% dan 3%. Analisis kandungan NSL dan polyisoprenoid dilakukan dengan membandingkan daun dan akar semai S. alba pada perlakuan variasi naungan 0% dengan salinitas 3% dengan perlakuan variasi naungan 75% dengan salinitas 3%. Hasil penelitian menunjukkan bahwa pertumbuhan semai S. alba optimum pada perlakuan variasi naungan 0% dengan salinitas 3% dengan pensentase tumbuh mencapai 100%, tinggi 2,6 cm, diameter 0,17 mm, berat basah akar 0,113 g, berat kering akar 0,08 g, berat basah tajuk 0,15 g, berat kering tajuk 0,05 g, jumlah daun 4 dan rasio tajuk dan akar 0,59. Kandungan polyisoprenoid yang diperoleh dari perlakuan variasi naungan 0% dengan salinitas 3% pada jaringan daun adalah sebesar 190,5 ml dan 133,4 ml pada jaringan akar sedangkan pada perlakuan variasi naungan 75% dengan salinitas 3% adalah sebesar 185,2 ml pada jaringan daun dan 231,9 ml pada jaringan akar. Diduga polyisoprenoid berperan dalam beradaptasi dengan ketersediaan cahaya, dimana polyisoprenoid meningkat konsentrasinya dengan berkurangnya intensitas cahaya matahari pada semai S. alba.
ABSTRACT
EVAN KHAROGI SINULINGGA. Response Plant Growth and Concentration long Chain Polyisoprenoid to Shade and Salinity Variations in Mangrove Sonneratia alba Smith under academic supervision by MOHAMMAD BASYUNI and YUNASFI.
Mangroves has a special ability to adapt with extreme environmental conditions. This condition allows mangroves to produce secondary metabolites as chemical defenses for their sustainability. Polyisoprenoid is one of the secondary metabolites which unknown function. The purpose of this study is to analyze the growth of S. alba seedling in responsse to shade variations and salinity concentration and its influence on the concentration of long-chain polyisoprenoid. The research was conducted from August to December 2014 using Factorial Completely Randomized design with shade treatment of 0%, 25%, 50%, and 75% along with salinity levels of 0%, 1.5% and 3%. Analysis of non-saponifiable lipid content and polyisoprenoid was done by comparing the leaves and roots of S. alba seedlings in variation 0% shade and 3% with salinity variations and in 75% shade with 3% salinity. The results showed that growth of S. alba seedling was optimum variation of 0% shade and 3% salinity showing 100% growth, 2.6 cm height, 0.17 mm diameter, 0.113 g fresh weight root, 0, 08 g dry weight root, 0.15 g wet weight shoot, 0.05 g dry weight shoot, 4 leaves number and ratio shoot and roots was 0.59. Polyisoprenoid content obtained at 190,5 ml in leaves an 133,4 ml in roots from variation 0% shade and with 3% salinity but in 75% shade with 3% salinity at 185,2 ml in leaves and 231,9 ml in roots. Polyisoprenoid may play an important role in adapting the availability light intensity, which polyisoprenoid concentration increased with reducing light intensity on S. alba seedling.
RIWAYAT HIDUP
Penulis lahir di Berastagi, 08 Februari 1994 dari Efendi Sinulingga dan Martini Br Ginting. Penulis merupakan anak kedua dari tiga bersaudara.
Penulis Lulusan SD Negeri 040462 Berastagi pada Tahun 2005, lulusan SMP Negeri 1 Berastagi pada tahun 2008, dan lulusan SMA Swasta Methodist Berastagi pada tahun 2011. Juni 2011 penulis diterima di Program Studi Kehutanan, Fakultas Pertanian Universitas Sumatera Utara melalui jalur SNMPTN. Selanjutnya penulis memilih peminatan Budidaya Hutan.
KATA PENGANTAR
Puji dan syukur penulis panjatkan kehadirat Tuhan Yang Maha Esa yang telah memberikan limpahan berkat dan anugerah kepada penulis, sehingga penulis dapat menyelesaikan penyusunan skripsi yang berjudul “Respons Pertumbuhan Tanaman dan Konsentrasi Rantai Panjang Polyisoprenoid terhadap Variasi Naungan dan Salinitas pada Mangrove Sonneratia alba Smith.”. Skripsi ini merupakan salah satu syarat untuk memperoleh gelar sarjana di Program Studi Kehutanan, Fakultas Pertanian, Universitas Sumatera Utara.
Pada kesempatan ini penulis mengucapkan terima kasih kepada orang tua penulis Efendi Sinulingga dan Martini Br Ginting atas doa dan dukungannya selama ini. Penulis juga menyampaikan ucapan terima kasih kepada Mohammad Basyuni, S.Hut, M.Si, Ph.D sebagai ketua komisi pembimbing skripsi dan Dr. Ir. Yunasfi, M.Si. sebagai anggota komisi pembimbing skripsi, yang telah banyak membimbing dan memberikan arahan serta masukan kepada penulis sehingga penulis dapat menyelesaikan penelitian ini.
Disamping itu penulis juga mengucapkan terima kasih kepada kepada semua staf pengajar dan pegawai di Program Studi Kehutanan, serta semua teman-teman mahasiswa yang telah membantu penulis dalam menyelesaikan skripsi ini. Semoga skripsi ini dapat bermanfaat bagi pengembangan ilmu pengetahuan, khususnya bidang kehutanan.
Medan, April 2015
DAFTAR ISI
Pelaksanaan Penelitian ... 13
Penyiapan media tanam ... 13
Pengumpulan dan penanganan buah S. alba ... 13
Penanaman dan Perlakuan ... 14
Pengamatan parameter ... 14
Persentase tumbuh semai (%) ... 14
Pertambahan tinggi semai (cm) ... 15
Pertambahan diameter semai (mm) ... 15
Berat basah akar (g) ... 15
Analisis NSL (nonsaponifieble lipids) dan Polyisoprenoid ... 17
HASIL DAN PEMBAHASAN
Korelasi perlakuan dengan parameter Pengamatan ... 18
Persentase tumbuh semai S. alba ... 20
Respons pertumbuhan tinggi tanaman S. alba ... 21
Respons pertumbuhan diameter tanaman semai S. alba ... 23
Respons pertumbuhan berat basah dan berat kering akar S. alba ... 25
Respons pertumbuhan berat basah dan berat kering tajuk S. alba ... 27
Respons pertumbuhan jumlah daunS. alba ... 29
Rasio Tajuk dan akar S. alba ... 31
Polyisoprenoid ... 36
KESIMPULAN DAN SARAN Kesimpulan ... 40
Saran ... 40
DAFTAR PUSTAKA
DAFTAR TABEL
No. Halaman
1. Tingkat salinitas optimum untuk pertumbuhan mangrove ... 7 2. Intensitas naungan optimum untuk pertumbuhan mangrove ... 9 3. Nonsaponifieble Lipids (NSL) pada semai S. alba. H0 merupakan
perlakuan naungan 0% dengan salinitas 3% dan H75 merupakan
DAFTAR GAMBAR
No. Halaman
1. Hasil pengukuran persentase tumbuh semai tanaman S. alba pada
umur 3 bulan.. ... 20 2. Hasil pengukuran respons pertumbuhan tinggi tanaman S. alba
pada umur 3 bulan (n = 0 – 13). Tanda * mengindikasikan secara
statistik berpengaruh nyata pada P < 0,05 menurut uji Dunnet.. ... 22 3. Hasil pengukuran respons pertumbuhan diameter tanaman S. alba
pada umur 3 bulan (n = 0 – 13). Tanda * mengindikasikan secara
statistik berpengaruh nyata pada P < 0,05 menurut uji Dunnet.. ... 24 4. Hasil pengukuran respons pertumbuhan berat basah akar tanaman
S. alba (A). Hasil pengukuran respons pertumbuhan berat kering akar tanaman S. alba (B) pada umur 3 bulan (n = 4 – 13). Tanda * mengindikasikan secara statistik berpengaruh nyata pada P < 0,05
menurut uji Dunnet. ... 25 5. Hasil pengukuran respons pertumbuhan berat basah tajuk tanaman
S. alba (A), Hasil pengukuran respons pertumbuhan berat basah kering tanaman S. alba (B) pada umur 3 bulan (n = 0 – 13). Tanda * mengindikasikan secara statistik berpengaruh nyata pada P <
0,05 menurut uji Dunnet ... 27 6. Hasil pengukuran respons pertumbuhan jumlah daun tanaman
semai S. alba pada umur 3 bulan (n = 0 – 13). Tanda * mengindikasikan secara statistik berpengaruh nyata pada P < 0,05
menurut uji Dunnet. ... 30 7. Hasil pengukuran rasio berat kering tajuk dan akar ( n = 0-13).
Tanda * mengindikasikan secara statistik berpengaruh nyata pada
P < 0,05 menurut uji Dunnet. ... 31 8. Thin-Layer Chromatography polyisoprenoid alkohol (dolichol) dari
semai daun dan akar S. alba dimana S merupakan standar dolichol, 1-3 merupakan daun semai S. alba perlakuan naungan 0%, 4-6 merupakan daun semai S. alba perlakuan naungan 75%, 7-9 merupakan akar semai S. alba perlakuan naungan 0% dan 10-12
DAFTAR LAMPIRAN
No. Halaman
1. Analisis sidik ragam (ANOVA) dan uji Dunnet semai S. alba ... 46 2. Korelasi antara variasi naungan dan salinitas dengan parameter
ABSTRAK
EVAN KHAROGI SINULINGGA. Respons Pertumbuhan Tanaman dan Konsentrasi Rantai Panjang Polyisoprenoid terhadap Variasi Naungan dan Salinitas pada Mangrove Sonneratia alba Smith. Dibimbing oleh MOHAMMAD BASYUNI dan YUNASFI.
Tumbuhan mangrove memiliki kemampuan khusus untuk beradaptasi dengan kondisi lingkungan yang ekstrim. Kondisi ini memungkinkan mangrove untuk menghasilkan metabolit sekunder sebagai pertahanan kimia untuk kehidupan mereka. Polyisoprenoid merupakan salah satu metabolit sekunder yang belum diketahui fungsinya. Tujuan penelitian ini adalah untuk mengukur respons pertumbuhan semai S. alba pada variasi naungan dan tingkat salinitas dan pengaruhnya terhadap konsentrasi rantai panjang polyisoprenoid. Penelitian ini dilaksanakan pada Agustus sampai Desember 2014 menggunakan Rancangan Acak Lengkap Faktorial dengan perlakuan naungan 0%, 25%, 50%, dan 75% dengan tingkat salinitas 0%, 1,5% dan 3%. Analisis kandungan NSL dan polyisoprenoid dilakukan dengan membandingkan daun dan akar semai S. alba pada perlakuan variasi naungan 0% dengan salinitas 3% dengan perlakuan variasi naungan 75% dengan salinitas 3%. Hasil penelitian menunjukkan bahwa pertumbuhan semai S. alba optimum pada perlakuan variasi naungan 0% dengan salinitas 3% dengan pensentase tumbuh mencapai 100%, tinggi 2,6 cm, diameter 0,17 mm, berat basah akar 0,113 g, berat kering akar 0,08 g, berat basah tajuk 0,15 g, berat kering tajuk 0,05 g, jumlah daun 4 dan rasio tajuk dan akar 0,59. Kandungan polyisoprenoid yang diperoleh dari perlakuan variasi naungan 0% dengan salinitas 3% pada jaringan daun adalah sebesar 190,5 ml dan 133,4 ml pada jaringan akar sedangkan pada perlakuan variasi naungan 75% dengan salinitas 3% adalah sebesar 185,2 ml pada jaringan daun dan 231,9 ml pada jaringan akar. Diduga polyisoprenoid berperan dalam beradaptasi dengan ketersediaan cahaya, dimana polyisoprenoid meningkat konsentrasinya dengan berkurangnya intensitas cahaya matahari pada semai S. alba.
ABSTRACT
EVAN KHAROGI SINULINGGA. Response Plant Growth and Concentration long Chain Polyisoprenoid to Shade and Salinity Variations in Mangrove Sonneratia alba Smith under academic supervision by MOHAMMAD BASYUNI and YUNASFI.
Mangroves has a special ability to adapt with extreme environmental conditions. This condition allows mangroves to produce secondary metabolites as chemical defenses for their sustainability. Polyisoprenoid is one of the secondary metabolites which unknown function. The purpose of this study is to analyze the growth of S. alba seedling in responsse to shade variations and salinity concentration and its influence on the concentration of long-chain polyisoprenoid. The research was conducted from August to December 2014 using Factorial Completely Randomized design with shade treatment of 0%, 25%, 50%, and 75% along with salinity levels of 0%, 1.5% and 3%. Analysis of non-saponifiable lipid content and polyisoprenoid was done by comparing the leaves and roots of S. alba seedlings in variation 0% shade and 3% with salinity variations and in 75% shade with 3% salinity. The results showed that growth of S. alba seedling was optimum variation of 0% shade and 3% salinity showing 100% growth, 2.6 cm height, 0.17 mm diameter, 0.113 g fresh weight root, 0, 08 g dry weight root, 0.15 g wet weight shoot, 0.05 g dry weight shoot, 4 leaves number and ratio shoot and roots was 0.59. Polyisoprenoid content obtained at 190,5 ml in leaves an 133,4 ml in roots from variation 0% shade and with 3% salinity but in 75% shade with 3% salinity at 185,2 ml in leaves and 231,9 ml in roots. Polyisoprenoid may play an important role in adapting the availability light intensity, which polyisoprenoid concentration increased with reducing light intensity on S. alba seedling.
PENDAHULUAN
Latar Belakang
Tumbuhan mangrove memiliki kemampuan khusus untuk beradaptasi dengan kondisi lingkungan yang ekstrim, seperti kondisi tanah yang tergenang, kadar garam yang tinggi, intensitas cahaya yang tinggi serta kondisi tanah yang kurang stabil. Dengan kondisi lingkungan seperti itu, beberapa jenis mangrove mengembangkan mekanisme yang memungkinkan secara aktif mengeluarkan garam dari jaringan tubuhnya, sementara yang lainnya mengembangkan sistem akar napas untuk membantu memperoleh oksigen bagi sistem perakarannya (Noor, 2006). Kondisi ekstrim ini memungkinkan mangrove untuk menghasilkan senyawa metabolit sekunder sebagai pertahanan kimia untuk beradaptasi. agar dapat tumbuh dan berkembang.
Hutan mangrove tersebar luas di seluruh daerah tropis dan sub tropis di dunia yang tumbuh subur di sepanjang garis pantai. Diperkirakan luas total hutan mangrove di Indonesia adalah 3,11 juta ha, yang mewakili sekitar 22,6 % dari hutan mangrove di dunia (Giri et al., 2011). Namun keberadaan hutan mangrove tersebut tidak memberikan nilai ekonomi yang signifikan terhadap pendapatan masyarakat sekitar hutan. Oleh karena itu, hutan mangrove kerap dialih fungsikan untuk pertambangan, perkebunan, pertambakan dan lahan-lahan pertanian.
Dalam 2 dekade terakhir luas hutan mangrove secara global menurun setidaknya 35% dengan laju degradasi sebesar 2,1% pertahunnya sejak tahun 1980-an dari total luas sekitar 17 juta ha (Valiela et al., 2001). Berdasarkan penelitian Giri et al. (2011) luas hutan mangrove pada 118 negara termasuk Indonesia pada tahun 2000 hanya sekitar 13,7 juta ha. Menurut Nugroho (2012) di Indonesia sendiri dari sekitar 3,11 juta ha hutan mangrove 1,8 juta ha telah mengalami kerusakan yang cukup serius, alih fungsi mangrove menjadi kawasan tambak menjadi penyebab utama kerusakan ini selain pembukaan kawasan hutan untuk perkebunan, pertanian dan pertambangan.
keberlangsungan ekosistem mangrove. Perepat (Sonneratia alba) merupakan salah satu jenis mangrove yang termasuk ke dalam jenis sekresi dan intoleran terhadap air tawar untuk periode yang lama serta merupakan salah satu jenis mangrove sejati mayor yang menyukai kondisi salinitas yang tinggi (Kusmana et al., 2003). Namun sampai saat masih sedikit studi yang membahas tentang aspek morfologi dan fisiologi tanaman ini di bawah variasi naungan dan salinitas, hal ini disebabkan karena sulitnya membudidayakan bibit dari biji S. alba yang tergolong kedalam biji normal yang berbeda dengan spesies Rhizophora, Bruguiera, maupun Avicenia yang berkembang biak menggunakan propagul dan biji yang bersifat kriptovivipari.
Tujuan Penelitian
Penelitian ini bertujuan untuk mengukur respons pertumbuhan S. alba pada variasi naungan dan tingkat salinitas serta pengaruhnya terhadap konsentrasi rantai panjang polyisoprenoid.
Manfaat Penelitian
Hasil penelitian ini dapat menjadi acuan penelitian dan sumber informasi bagi ilmu pengetahuan terkait aspek fisiologi dan morfologi semai S. alba pada variasi naungan dan salinitas serta pengaruhnya terhadap rantai panjang polyisoprenoidnya.
Hipotesis
TINJAUAN PUSTAKA
Hutan Mangrove dan Karakteristiknya
Ekosistem mangrove merupakan ekosistem yang unik yang termasuk ke dalam ekosistem hutan yang ditandai oleh vegetasi yang sangat khusus dan khas dan dibatasi oleh lingkungan (Raymold dan Queen, 1974). Tanaman mangrove tersebar meliputi bagian zona pasang surut di daerah tropis dan daerah subtropis. Mangrove adalah tanaman yang toleran terhadap garam, yang mampu tumbuh diberbagai tingkatan salinitas mulai dari air tawar sampai daerah dengan tingkat salinitas yang sangat tinggi hal ini dikarenakan mangrove memiliki mekanisme khusus untuk mengeluarkan kelebihan garam dari tubuhnya (Tomlinson, 1986). Ada sekitar 70 jenis mangrove dengan 30 genus dan 20 famili, yang setiap jenisnya memiliki peran penting di ekosistem pantai lahan basah (Duck, 1992).
sedangkan tumbuhan asosiasi adalah tumbuhan yang toleran terhadap salinitas dan dapat berasosiasi dengan mangrove mayor.
Setiap jenis mangrove yang tumbuh berkaitan erat dengan faktor lingkungannya, diantaranya media tumbuh, genangan air pasang, salinitas, erosi, penambahan lahan pesisir, fisiografi, kondisi sungai dan aktivitas manusia, sehingga secara khas membentuk pola zonasi (Watson, 1928; Chapman, 1975; Hann, 1931; Kusmana et al, 2003). Setiap spesies mangrove (terutama mangrove yang termasuk dalam genus Rhizophora, Bruguiera, Sonneratia, Heritiera dan Nypa) pada kondisi ekologi yang berkaitan dengan tingkat salinitas air dan kondisi tanah, serta rezim genangan mampu membentuk zona yang khas (Blasco et al., 1996). Bengen (1999) menyatakan bahwa zonasi mangrove Indonesia pada umumnya di daerah paling dekat dengan laut, dengan substrat agak berpasir, sering ditumbuhi Avicennia dan Sonneratia, kemudian lebih ke darat lagi dengan kadar salinitasnya agak rendah didominasi Rhizophora. Selain itu juga dijumpai Bruguiera dan Xylocarpus kemudian daerah yang memiliki substrat tanah berlumpur keras dan terendam pada saat air pasang tertinggi didominasi Bruguiera, zona yang paling belakang dan berbatasan dengan daratan biasanya ditumbuhi oleh Nypa fruticans dan pandan laut (Pandanus sp.).
Sonneratia alba Smith.
Perepat (Sonneratia alba Smith.) merupakan salah satu spesies mangrove mayor dari famili sonneratiaceae berjenis sekresi dengan klasifikasi sebagai berikut (Noor et al., 1999)
Kingdom : Plantae (Tumbuhan)
Super Divisi : Spermatophyta (Menghasilkan biji) Divisi : Magnoliophyta (Tumbuhan berbunga) Kelas : Magnoliopsida (berkeping dua / dikotil) Sub Kelas : Rosidae
Ordo : Myrtales Famili : Sonneratiaceae Genus : Sonneratia
Spesies : Sonneratia alba Smith.
S. alba, S. caseolaris, dan S. ovata memiliki pohon berukuran sedang hingga besar, tetapi memiliki viabilitas biji yang rendah (Sukristijono, 1984). S. alba umumnya tumbuh di zona intertidal rendah, S. alba intoleran terhadap air tawar untuk jangka waktu yang lama dan lebih suka salinitas tinggi. Di zona intertidal rendah, S. alba dapat menjadi spesies dominan bersama dengan A. marina, membentuk tegakan murni sepanjang margin arah laut dari jangkauan.
Salinitas dan Naungan
Salinitas memainkan peran penting dalam mengatur pertumbuhan dan distribusi mangrove, karena toleransi terhadap garam merupakan salah satu mekanisme adaptasi mangrove (Wang, 2011). Berdasarkan cara mangrove beradaptasi dari salinitas, tanaman mangrove dapat diklasifikasikan ke dalam dua kelompok hal yakni, mangrove sekresi dan nonsekresri (Scholander et al., 1962).
Tanaman mangrove berjenis sekresi yang toleran terhadap garam menyerap dan menyimpan garam di jaringannya dan kemudian kelebihan garam ini dikeluarkan melalui kelenjar-kelenjar khusus yang terdapat pada daun. Sedangkan untuk tanaman mangrove non-sekresi kelebihan garam diatasi dengan
mengakumulasikan garam pada bagian daun dan mengugurkannya (Clogh et al., 1982). Meskipun tanaman mampu hidup dan beradaptasi dengan
lingkungan salinitas yang tinggi, tanaman mangrove tetap saja membutuhkan air tawar dalam mekanisme pertumbuhannya (Naidoo, 1987).
Beberapa penelitian sebelumnya terkait dengan respons pertumbuhan tanaman mangrove terhadap salinitas menunjukkan pengaruh nyata terhadap pertumbuhan tanaman mangrove. Berdasarkan penelitian tersebut, menunjukkan bahwa setiap spesies mangrove memiliki tingkat salinitas yang optimum yang bervariasi untuk pertumbuhannya, hal ini dapat dilihat pada Tabel 1.
Tabel 1. Tingkat salinitas optimum untuk pertumbuhan mangrove
Jenis Salinitas Referensi
B.gymnorrhiza 0,00-0,70%
Hutahaen et al. (1999)
A. marina 2,25-3,00%
R. mucronata 0,75-0,15%
R. stylosa 0,50% Ester et al. (2013)
C. tagal 0,50% Ramayani et al. (2012)
R. apiculata 1,50% Prayunita et al. (2012)
Cahaya merupakan salah satu faktor penting dalam berlangsungnya kegiatan fotosintesis dalam metabolisme tumbuhan, namun setiap jenis tanaman membutuhkan intensitas cahaya yang berbeda (Farnsworth dan Ellison, 1996). Banyaknya intensitas cahaya yang masuk mempengaruhi suhu yang berperan dalam pertumbuhan tanaman di persemaian. Tingginya intensitas cahaya dapat menyebabkan tumbuhan menjadi kerdil, dan menyebabkan gugurnya daun, namun intensitas cahaya yang rendah dapat menyebabkan etiolasi pada bibit sehingga dapat menyebabkan kematian pada bibit (Marschner, 1995).
Berdasarkan hasil penelitian Anwar (1997) menunjukkan korelasi antara
tingkat naungan diterapkan pada pembibitan dan pertumbuhan bibit B. gymnorrhiza. Ada kecenderungan bahwa tinggi, jumlah pertumbuhan daun,
dan peningkatan persentase hidup mangrove ketika variasi naungan meningkat. Cahaya berpengaruh signifikan terhadap pertumbuhanan dan kelangsungan hidup anakan mangrove, di lingkungannya anakan mangrove yang berada di bawah tajuk hutan mangrove telah beradaptasi atas cahaya yang minimun sehingga pertumbuhannya tetap dapat optimum (Smith, 1987).
celah-celah tajuk yang memiliki sedimentasi yang tidak tercemar dengan tingkat salinitas rendah dan hal ini sama dengan hasil yang diperoleh oleh Smith (1987) bahwa mangrove lebih suka tempat tumbuh yang memiliki intensitas cahaya yang tinggi.
Berikut ini beberapa hasil penelitian sebelumnya terkait variasi naungan yang optimum untuk pertumbuhan beberapa jenis mangrove, yang dapat dilihat pada Tabel 2.
Tabel 2. Intensitas naungan optimum untuk pertumbuhan mangrove Jenis Intensitas naungan Referensi
B. gymnorrhiza 25 % Anwar (1997)
R. mucronato 75 % Yanti et al. (2011)
R. apiculato 50 % Simarmata et al. (2011)
A. marina 50 % Keliat et al. (2013)
Polyisoprenoid
Tumbuhan mampu menghasilkan senyawa metabolit sekunder dengan struktur molekul dan aktivitas biologik yang beraneka ragam (Radji, 2005). Polyisoprenoid alkohol (dolichols dan polyprenols) merupakan salah satu metabolit sekunder dari tumbuhan yang ditemukan di semua makhluk hidup mulai dari bakteri hingga mamalia (Surmacz dan Swiezewska, 2011). Rantai polyisoprenoid alkohol tersusun dari 5-100 lebih unit isoprenoid yang menciptakan polimer yang berbeda dalam rantai panjang konfigurasi geometrisnya (Ciepichal et al., 2011).
Danikiewicz, 2005). Perbedaaan rantai panjang polyprenol dengan dolichol terletak pada unit isoprennya dimana polyprenol terdiri dari 5-50 unit isoprena yang sering ditemui dalam fraksi lipid yang tidak tersabunkan dari daun spermatophyta sedangkan dolichols merupakan komponen struktural penting dari membran eukariotik dimana dolichols mewakili keluarga lipid polyisoprenoid memiliki 16-22 unit isoprena, terutama di cis-configuration, dengan unit-isopren jenuh, yang berfungsi membawa monosakarida dan oligosakarida (Sagami, 1992; Wieslaw et al., 1994; Haeuptle 2009). Enzim yang berperan dalam sintesis polyisoprenoid adalah cis-prenyltransferase yang berperan dalam pembangunan rantai panjang hidrokarbon.
Berdasarkan penelitian Tateyama et al. (1999) dolichol merupakan senyawa metabolit sekunder yang merupakan bagian polyisoprenoid yang dominan terdapat di jaringan akar dan daun. Perbedaan rasio kandungan dolichol dan polyprenol pada jaringan tanaman selalu berbeda-beda, rasio polyprenol dan dolichol pada daun famili Capparidaceae adalah 1:1 dan 2:1 pada daun karet, namun pada beberapa tanaman paku-pakuan hanya mengandung sanyawa dolichol (Jankowski et al., 1994; Tateyama et al., 1999; Wojtas et al., 2005).
BAHAN DAN METODE
Waktu dan Tempat
Penelitian ini dilaksanakan pada Agustus sampai dengan Desember 2014. Penelitian ini terdiri atas tiga tahapanan yang dilakukan di tempat yang berbeda, persemaian dilaksanakan di laboratorium Ekologi Hutan, Fakultas Pertanian, Universitas Sumatera Utara, Penanaman dilaksanakan di lahan pertanian, Fakultas Pertanian, Universitas Sumatera Utara dan ekstraksi polyisoprenoid dilaksanakan di Laboratorium Penelitian, Fakultas Farmasi, Universitas Sumatera Utara.
Alat dan Bahan
Alat yang digunakan dalam pelaksanaan penelitian ini adalah sprayer, ember plastik, jangka sorong, penggaris, Refractometer (Master Refractometer), timbangan (Camry; Model: EK3820), kamera digital, cutter, gunting, alat tulis, perangkat komputer yang dilengkapi paket SPSS 16,0 dan Ms. Excel 2007, oven, tube, dan vakum.
Bahan yang digunakan dalam pelaksanaan penelitian ini adalah buah Sonneratia alba yang sehat dan matang yang diambil dari Pulau Sembilan, Kecamatan Pangkalan Susu, Kabupaten Langkat, Sumatera Utara, air, bak kecambah, pasir sungai (tanpa salinitas) yang sudah disterilkan, pot plastik, paranet 25%, 50%, dan 75%, label nama, garam, klorofom, methanol, kertas filter, larutan Na, nitrogen cair, ethanol, KOH, dan larutan hexan.
Pelaksanaan Penelitian
A. Penyiapan Media Tanam
untuk membuat salinitas 0%, 1,5%, dan 3% (sama dengan level air laut, metode ini mengacu pada penelitian Basyuni et al., (2009, 2012)). Dimana garam yang dipakai adalah marine salt. Untuk membuat konsentrasi salinitas 1,5%, 3% dengan cara melarutkan 17 gram, dan 34 gram bubuk garam komersial masing-masing dalam 1 liter air.
B. Pengumpulan dan Penanganan Buah S. alba
Buah S.alba diperoleh dari pohon yang telah dewasa di Pulau Sembilan, Kabupaten Langkat, Sumatera Utara. Buah yang dikumpulkan merupakan buah yang matang secara fisiologis dan bijinya siap untuk dikecambahkan. Ciri-ciri buah S. alba yang telah matang ditandai dengan karakteristik diameter buah berukuran lebih dari 4 cm, buah berwarna hijau kekuningan dan kelopak buah mudah terlepas dari buahnya, dan bijinya sudah mengeras. Jumlah biji dalam satu buah S.alba bisa mencapai 100-200 biji.
C. Penanaman dan Perlakuan
Penanaman dilakukan ketika semai sonneratia alba telah berdaun 2 dengan tinggi ± 2 cm yang kemudian ditanam kedalam pot plastik dengan media pasir yang sudah disterilkan kemudian ditempatkan di berbagai variasi naungan yang telah ditentukan (tanpa naungan, 25%, 50%, dan 75%), untuk setiap penyiraman dengan varasi salinitas yang diberikan (0%, 1,5%, dan 3%) masing-masing 13 ulangan.
Pada setiap perlakuan naungan terdapat 3 perlakuan salinitas sehingga terdapat 12 kombinasi perlakuan secara keseluruhan yaitu perlakuan kombinasi tanpa naungan dengan salinitas 0% (C0), tanpa naungan dengan salinitas 1,5% (M0), tanpa naungan dengan salinitas 3% ( H0), naungan 25% dengan salinitas 0% (C25), naungan 25% dengan salinitas 1,5% (M25), naungan 25% dengan salinitas 3% (H25), naungan 50% dengan salinitas 0% (C50), naungan 50% dengan salinitas 1,5% (M50), naungan 50% dengan salinitas 3% (H50), naungan 75% dengan salinitas 0% (C75), naungan 75% dengan salinitas 1,5% (M75), dan naungan 75% dengan salinitas 3% (H75). Konsentrasi garam pada setiap perlakuan pot diperiksa setiap selama percobaan dengan refractometer.
Pengamatan Parameter
Pengamatan dilakukan 3 bulan setelah tanam dan parameter yang diamati adalah:
1. Persentase Tumbuh Semai (%)
Perhitungan persentase tumbuh semai dilakukan sebelum dilakukan pemanenan. Persentase tumbuh semai dihitung dengan:
% �����ℎ= �����ℎ��������������ℎ
2. Pertambahan Tinggi Semai (cm)
Pengambilan data tinggi setelah 3 bulan penanaman semai dengan menggunakan penggaris, pada setiap satuan percobaan. Tinggi semai diukur mulai dari bagian plumula sampai titik tumbuh tertinggi.
3. Pertambahan Diameter Semai (mm)
Pengukuran diameter semai dilakukan pada pangkal batang dengan menggunakan jangka sorong. Pengambilan data diameter dilakukan bersamaan dengan pengambilan data tinggi semai.
4. Berat Basah Akar (g)
Untuk mendapatkan berat basah akar, bagian akar yang baru dipanen dimasukkan ke dalam amplop dan diberi label sesuai dengan perlakuan kemudian ditimbang berat akar S. alba
5. Berat Basah Tajuk (g)
Untuk mendapatkan berat basah tajuk, bagian tajuk yang baru dipanen dimasukkan ke dalam amplop dan diberi label sesuai dengan perlakuan yang kemudian ditimbang berat tajuk S. alba.
6. Berat Kering Akar (g)
Untuk mendapatkan berat kering akar, bagian akar dimasukkan ke dalam amplop dan diberi label sesuai dengan perlakuan. Kemudian akar S. alba dioven pada suhu 75ºC sampai berat kering konstan (24 jam) lalu ditimbang berat kering akar S. alba.
7. Berat Kering Tajuk (g)
pada temperatur 75ºC sampai berat kering konstan (24 jam), lalu ditimbang berat kering tajuk S. alba.
8. Rasio Tajuk dan Akar
Perhitungan rasio tajuk dan akar dilakukan pada akhir pengamatan. Perhitungan rasio tajuk dan akar dapat diperoleh dengan menggunakan rumus sebagai berikut :
����� =���������������� ���������������
Analisis Statistik
Data dianalisis menggunakan Rancangan Acak Lengkap (RAL) Faktorial dengan 4 perlakuan variasi naungan dan 3 perlakuan salinitas dengan 13 ulangan yang dilanjutkan dengan uji Dunnet untuk perbandingan seluruh perlakuan terhadap kontrol menggunakan SPSS versi 16 dengan model linear
����= �+��+��+ (��)��+∈ ��� Dimana :
���� : Hasil pengukuran respon pertumbuhan tanaman pada perlakuan variasi
naungan ke-i dan perlakuan salinitas ke-j pada ulangan ke-k � : Nilai rataan umum (mean)
�� : Pengaruh variasi naungan ke-i �� : Pengaruh tingkat salinitas ke-j
(��)�� : Interaksi perlakuan variasi naungan ke-i dan perlakuan salinitas ke-j ∈ ��� : pengaruh galat perlakuan variasi naungan ke-i dan perlakuan salinitas ke-
D. Analisis NSL (Nonsaponifiable Lipids) dan Polyisoprenoid
Daun dan akar semai S.alba yang telah berumur 3 bulan dikeringkan selama 24 jam pada suhu 60oC – 76oC. Jaringan yang telah dikeringkan kemudian dihaluskan menjadi bubuk. Bubuk kemudian direndam ke dalam 20 ml CHCl3:CH3OH (2:1) selama satu hari. Kedua jenis larutan kemudian diinkubasi
pada suhu 40oC selama 2 jam. Kemudian secara terpisah masing-masing larutan difilter dan dihasilkan filtrate. Hasil filtrate disebut juga ekstrak lipid.
Ekstrak lipid dari daun, disaponifikasi pada suhu 65oC – 70oC selama 2 jam dalam 2 ml metanol 50% yang mengandung 2 M KOH. Ekstrak lipid dari akar disaponifikasi pada suhu 55oC selama 3 jam dalam 20 ml ethanol 95% yang mengandung 15 % (w/v) KOH. Saponin yang tak tersabunkan dari lipid mentah dari masing-masing jaringan diekstraksi dengan hexane dan pelarut organik yang telah di evaporasikan. Sisa dari masing-masing sampel dilarutkan dalam methanol dan diterapkan ke dalam sebuah kolom RP-18 Sep-Pak dengan methanol dan lipid non-polar yang mengandung alkohol polyisoprenoid dengan hexane.
E. Analisis Thin-Layer Chromatography (TLC)
HASIL DAN PEMBAHASAN
Korelasi Perlakuan dengan Parameter Pengamatan
Korelasi antara variasi naungan dan salinitas dengan parameter pengamatan dapat dilihat pada Tabel 3.
Tabel 3. Korelasi antara variasi naungan dan salinitas dengan parameter pengamatan.
Respons positif yang diperlihatkan oleh semai S. alba terhadap salinitas (Tabel 3) menunjukkan bahwapertumbuhan semai S. alba sangat mencerminkan kondisi di lapangan. Menurut Tomlinson (1986) S. alba merupakan mangrove mayor berjenis sekresi yang manyukai salinitas tinggi dengan tingkat intensitas cahaya yang tinggi, oleh karena itu S. alba bersama dengan Avicenia secara zonasi menempati posisi paling dekat dengan laut pernyataan ini didukung oleh pernyataan Kusmana et al. (2003) yang menyatakan bahwa semai S. alba intoleran terhadap naungan sehingga keberadaan naungan diduga dapat mematikan bibit.
Variasi salinitas memberikan respons negatif terbesar terhadap berat basah akar, berat kering akar serta berat kering tajuk semai S. alba dengan nilai korelasi masing-masing adalah -0,451, -0,387, dan -0,276 (Tabel 3). Hal ini diduga karena keberadaan naungan mengganggu dan menghambat laju fotosintesis semai S. alba yang membutuhkan intensitas cahaya yang tinggi. Hal ini sesuai dengan studi Schmidt (2002) bahwa intensitas cahaya rendah dapat mengganggu fotosintesis hingga dapat menyebabkan kematian, hal ini didukung penelitian Simorangkir (2000) bahwa intensitas cahaya mempengaruhi tinggi dan diameter semai yang sejalan dengan proses fotosintesis sehingga berkaitan dengan pertumbuhan tajuk semai.
korelasi antara diameter dengan tinggi, sedangkan korelasi terkecil adalah 0,228 antara berat kering akar dengan rasio akar dan tajuk. Hal ini sesuai dengan studi
Husch et al. (1972) bahwa pertumbuhan merupakan pertambahan dimensi dari
suatu tumbuhan pada waktu tertentu sehingga pertambahan suatu dimensi akan
mempengaruhi pertumbuhan dimensi lainnya karena pertumbuhan merupakan
hasil yang terbentuk dari proses fotosintesis.
Persentase Tumbuh Semai S. Alba
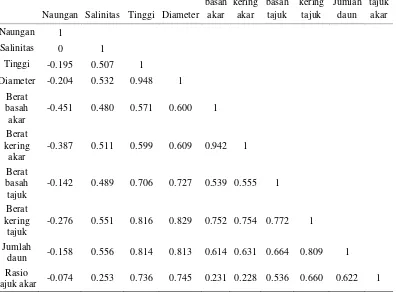
Berdasarkan hasil pengamatan yang telah dilakukan selama 3 bulan persentase tumbuh semai S. alba dapat dilihat Gambar 1.
Gambar 1. Hasil pengukuran persentase tumbuh semai tanaman S. alba pada umur 3 bulan.
Pada Gambar 1, persentse pertumbuhan tertinggi semai S. alba terdapat pada perlakuan tanpa naungan dengan tingkat salinitas 3% yakni sebesar 100%. Hachinohe et al., (1999) menyebutkan bahwa S. alba dapat tumbuh secara maksimal apabila diberi naungan dengan intensitas 20% pada umur 1-2 bulan penanaman dan pada bulan berikutnya sebaiknya dilakukan pengurangan
intensitas naungan secara berkala mengingat bahwa S. alba akan membutuhkan intensitas cahaya yang lebih tinggi di bulan-bulan berikutnya.
Pada perlakuan naungan 75% dengan tingkat salinitas 0% dan 1,5% tidak memiliki tingkat pertumbuhan (persentase tumbuh 0%). Pada fase bibit setiap tanaman memerlukan naungan karena tidak tahan terhadap intensitas cahaya penuh, namun menurut Kusmana et al.(2003) Sonneratia merupakan spesies yang intoleran terhadap naungan, pemberian naungan pada waktu yang cukup lama akan mempengaruhi pertumbuhan dan persentase tumbuh semai S. alba.
Pengaruh tingkat salinitas juga sangat berpengaruh terhadap pertumbuhan S. alba mengingat bahwa berdasarkan zonasi S. alba berada pada zona paling dekat dengan laut bersamaan dengan Avicennia. S. alba merupakan spesies yang intoleran terhadap air tawar dalam waktu yang lama dan menyukai tingkat salinitas yang tinggi. Oleh karena itu tingkat kematian yang tertinggi berada pada perlakuan naungan 75% dengan tingkat salinitas 0% dan 1,5% disebabkan kondisi S. alba yang mengalami stress akibat kekurangan intensitas cahaya matahari dan genangan air dengan salinitas yang rendah. Namun meskipun diberi perlakuan naungan dengan salinitas 3% persentase tumbuh semai S. alba lebih dari 60% diduga hal ini erat kaitannya dengan kemampuan adaptasi S. alba terhadap cekaman salinitas yang tinggi meskipun dalam kondisi semai.
Respons Pertumbuhan Tinggi Semai S. alba
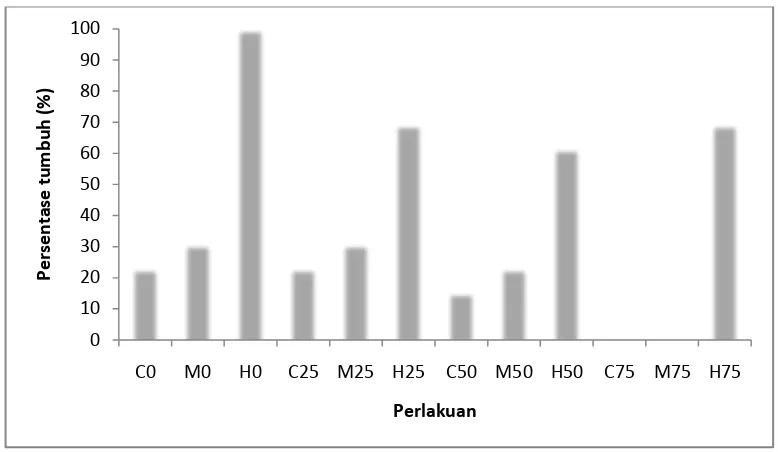
Gambar 2. Hasil pengukuran respons pertumbuhan tinggi tanaman S. alba pada umur 3 bulan (n = 0 – 13). Tanda * mengindikasikan secara statistik berbeda nyata pada P < 0,05 menurut uji Dunnet.
Pada gambar 2 terlihat bahwa respons pertumbuhan tinggi tanaman semai S. alba tertinggi terdapat pada perlakuan naungan 25% dengan tingkat salinitas 0%, dengan rata-rata tinggi 2,83 cm, dan yang terendah pada perlakuan naungan 75% dengan tingkat salinitas 0% dan 1,5% dengan tinggi rata-rata 0 cm. Hal ini diduga disebabkan oleh ketersediaan cahaya yang cukup pada naungan 25 % maka intensitas cahaya yang masuk mencapai 75%, hal ini sesuai dengan studi Hachinohe et al., (1999) yang menyarankan persemaian S. alba menggunakan naungan dengan intensitas 20 % hal ini berkaitan dengan spesies S. alba yang merupakan spesies intoleran terhadap naungan.
Berdasarkan uji Dunnet perlakuan naungan 75% memiliki respons berbeda nyata terhadap pertumbuhan semai S. alba tanpa naungan dengan P < 0,05, akan tetapi naungan 75% yang dibandingkan dengan tanpa naungan memberikan respons negatif terhadap pertumbuhan semai. Selain itu salinitas 3% juga sangat berbeda nyata terhadap terhadap respons pertumbuhaan tinggi tanaman semai S.
alba dengan P < 0,05, hal ini dapat dilihat dari tingkat keseragaman tinggi tanaman meskipun berada pada tingkat naungan yang berbeda yakni secara berurutan mulai dari 0%, 25%, 50% dan 75% yakni dengan rata-rata sebesar 2,6 cm, 2,66 cm, 2,78 cm, dan 2,7 (Gambar 2).
Pada perlakuan dengan variasi naungan 75% untuk tingkat salinitas 0% dan 1,5% tidak memiliki tinggi rata-rata (Gambar 2) hal ini dikarenakan tingkat kematian yang mencapai 100% pada perlakuan ini. Pada bulan-bulan pertama kondisi tanaman mengalami pertumbuhan yang sama dengan perlakuan yang lainnya, namun memasuki bulan kedua satu persatu tanaman mengalami pengguguran daun dan kemudian mati hal ini disebabkan kurangnya intensitas cahaya yang masuk. Menurut Schmidt (2002) intensitas cahaya yang rendah akan menggangu proses fotosintesis sehingga menggangu pertumbuhan tanaman dan akan menyebabkan tanaman mengalami etiolasi yang pada akhirnya akan mati.
Respons Pertumbuhan Diameter Semai S. alba
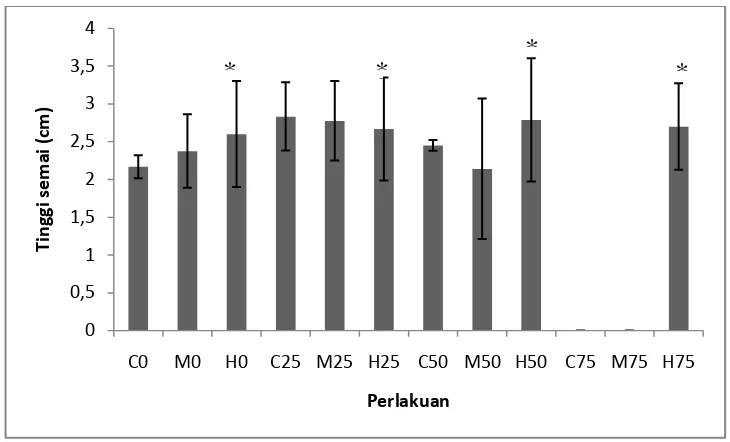
Gambar 3. Hasil pengukuran respons pertumbuhan diameter tanaman S. alba pada umur 3 bulan (n = 0 – 13). Tanda * mengindikasikan secara statistik berbeda nyata pada P < 0,05 menurut uji Dunnet.
Berdasarkan Gambar 3, bahwa respons pertumbuhan diameter tertinggi berada pada perlakuan variasi naungan 50% dengan tingkat salinitas 3% dengan diameter rata-rata mencapai 0,18 mm. sedangkan diameter terendah berada pada perlakuan dengan variasi naungan 75% dengan tingkat salinitas 0% dan 1,5% dengan diameter rata-rata 0 mm. Tinggi rata-rata dari perlakuan denga variasi naungan 50% dengan tingkat salinitas 3% mencapai 2,78 mm. Hal ini sesuai dengan penelitian Simorangkir (2000) bahwa laju pertumbuhan diameter batang pada tanaman akan sejalan dengan laju efektifitas dari fotosintesis yang erat kaitannya dengan banyaknya intensitas cahaya yang masuk karena akan mempengaruhi diameter atau tinggi serta morfologi tanaman secara keseluruhan.
Berdasarkan uji Dunnet yang dilakukan menunjukkan bahwa perbedaan variasi naungan dan tingkat salinitas berbeda nyata terhadap respons pertumbuhan diameter semai tanaman S. alba. Dimana untuk variasi naungan 75%, P < 0,05 yang dibandingkan dengan tanpa naungan. Salinitas 3% juga berbeda nyata
terhadap pertumbuhan diameter dengan nilai P < 0,05 yang dibandingkan dengan salinitas 0%. Secara keseluruhan perbedaan tinggi setiap perlakuan variasi naungan didominasi oleh tingkat salinitas 3%, menunjukkan bahwa semai S. alba lebih menyukai kondisi salinitas tinggi dibandingkan dengan kondisi air tawar.
Respons Pertumbuhan Berat Basah dan Berat Kering Akar Semai S. alba
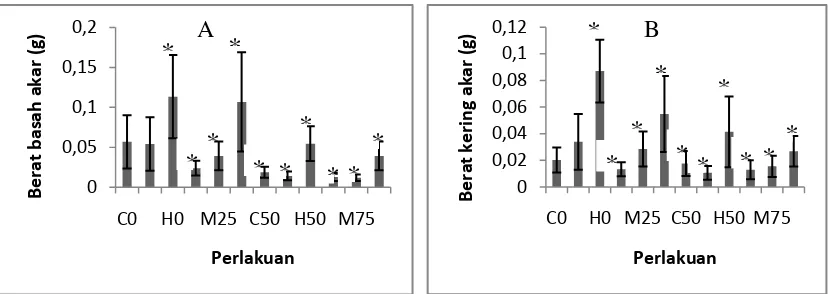
Pengukuran berat basah akar tanaman semai S. alba dilakukan dengan memotong bagian akar tanaman pada sisi tempat daun tembaga tumbuh yang kemudian ditimbang. Untuk berat kering tanaman semai S. alba diperoleh dari hasil pengovenan akar tanaman semai S. alba. Pengukuran berat basah dan berat kering akar semai S. alba dapat dilihat pada Gambar 4
Gambar 4. Hasil pengukuran respons pertumbuhan berat basah akar tanaman S. alba (A). Hasil pengukuran respons pertumbuhan berat kering akar
tanaman S. alba (B) pada umur 3 bulan (n = 4 – 13). Tanda * mengindikasikan secara statistik berbeda nyata pada P < 0,05 menurut uji Dunnet.
Berdasarkan grafik pertumbuhan berat basah akar tanaman semai S. alba menunjukkan berat basah akar tanaman semai S. alba tertinggi pada perlakuan dengan variasi tanpa naungan dengan tingkat salinitas 3% dengan berat basah mencapai 0,113 gr sedangkan berat basah tanaman semai S. alba terkecil berada pada perlakuan dengan variasi naungan 75% dengan tingkat salinitas 1,5% dengan
berat hanya 0,011 g. Untuk berat kering akar tanaman semai S. alba berat tertinggi berada pada perlakuan dengan variasi tanpa naungan dengan tingkat salinitas 3%
dengan berat mencapai 0,087 g sedangkan untuk berat kering tanaman semai S. alba terkecil berada pada perlakuan variasi naungan 75% dengan tingkat
salinitas 0% yakni sebesar 0,012 g.
Berdasarkan uji Dunnet respons pertumbuhan berat basah dan berat kering tanaman menunjukkan pengaruh nyata dari perlakuan variasi naungan dan tingkat salinitas dengan P < 0,05. Untuk variasi naungan, perlakuan naungan 25%, 50% dan 75% berbeda nyata terhadap pertumbuhan semai dibandingkan dengan tanpa naungan selain itu salinitas 3% juga berbeda nyata terhadap pertumbuhan semai dibandingkan dengan salinitas 0% Perlakuan naungan dan salinitas memberikan pengaruh terhadap respons pertumbuhan tinggi dan diameter sehingga erat kaitannya dengan biomassa akar karena merupakan hasil akumulasi dari fotosintesis.
dan Purwoko (2003) keberadaan naungan akan mengakibatkan pertambahan luas dan bentuk daun untuk mengefisienkan tangkapan cahaya yang masuk sehingga pertumbuhan tajuk lebih cepat dibandingkan pertumbuhan akar.
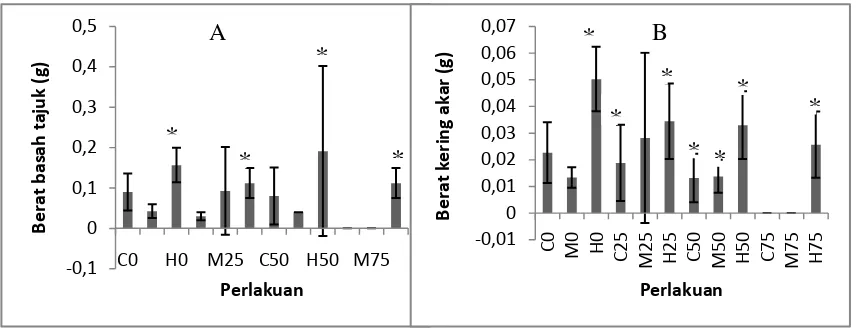
Respons Pertumbuhan Berat Basah dan Berat Kering Tajuk Semai S. alba Pengukuran berat basah akar tanaman semai S. alba dilakukan dengan memotong bagian tajuk tanaman pada sisi tempat daun tembaga tumbuh yang kemudian ditimbang. Untuk berat kering tanaman semai S. alba diperoleh dari hasil pengovenan tajuk semai S. alba. Pengukuran berat basah dan berat kering tajuk semai S. alba dapat dilihat pada Gambar 5.
Gambar 5. Hasil pengukuran respons pertumbuhan berat basah tajuk tanaman S. alba (A), Hasil pengukuran respons pertumbuhan berat basah kering tanaman S. alba (B) pada umur 3 bulan (n = 0 – 13). Tanda * mengindikasikan secara statistik berbeda nyata pada P < 0,05 menurut uji Dunnet.
Berdasarkan Gambar 5 pertumbuhan berat basah tajuk tanaman semai S. alba menunjukkan berat basah tajuk tanaman semai S. alba tertinggi pada perlakuan dengan variasi naungan 50% dengan tingkat salinitas 3% dengan berat basah mencapai 0,191 g sedangkan berat basah tajuk tanaman semai S. alba terkecil berada pada perlakuan variasi naungan 75% dengan tingkat salinitas 0%
S. alba berada pada perlakuan dengan variasi tanpa naungan dengan tingkat salinitas 3% dengan berat basah mencapai 0,050 g sedangkan untuk berat kering tajuk tanaman semai S. alba terkecil berada pada perlakuan variasi naungan 75% dengan tingkat salinitas 0% dan 1,5% yakni sebesar 0 g.
Berdasarkan uji Dunnet respons pertumbuhan berat basah dan berat kering tanaman menunjukkan pengaruh nyata dari perlakuan variasi naungan dan tingkat salinitas dengan P < 0,05. Untuk berat basah tajuk naungan 75% dan salinitas 3% berbeda nyata terhadap respons pertumbuhan berat basah semai. Untuk berat kering tajuk naungan 25%, 50%, dan 75% serta salinitas 3% berbeda nyata terhadap pertumbuhan berat kering semai. Perlakuan naungan dan salinitas memberikan pengaruh terhadap respons pertumbuhan tinggi dan diameter sehingga erat kaitannya dengan biomassa akar karena merupakan hasil akumulasi dari fotosintesis.
Tingginya angka standar deviasiasi pada perlakuan variasi naungan 25% dengan tingkat salinitas 1,5% dan variasi naungan 50% dengan tingkat salinitas 3% (Gambar 6A) diduga karena tingginya tingkat kematian pada semai tersebut selain itu kondisi semai pada saat dilakukan pengukuran tidak berada dalam kondisi sehat dan sebagian semai telah mengalami kerusakan mulai dari pembusukan sebagian tajuk sampai kondisi tanaman yang layu. Oleh karena itu kandungan kadar air pada tajuk tanaman yang mengalami pembusukan sangat tinggi apabila dibandingkan dengan kondisi tanaman segar dan setelah pengovenan kehilangan kadar air tersebut mengakibatkan penurunan berat yang sangat signifikan, akibatnya perbandingan berat kering tajuk pada tanaman segar dan tanaman yang rusak sangat tinggi.
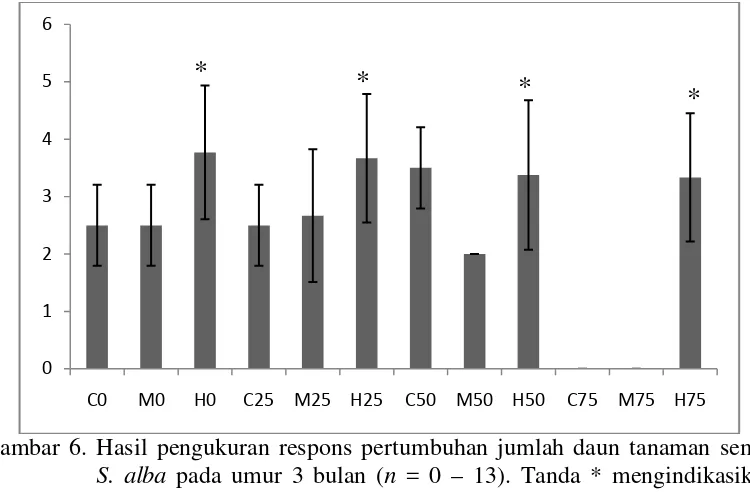
Respons Pertumbuhan Jumlah Daun Semai S. alba
Pengukuran respons pertumbuhan jumlah daun semai S. alba dilakukan pada saat pemanenan yakni pada umur 3 bulan. Daun semai S. alba yang dihitung dalam kegiatan ini adalah daun muda dan daun tua yang telah memiliki bentuk sempurna yakni telah memiliki tangkai daun, tulang daun dan bentuk daun yang jelas. Respons pertumbuhan jumlah daun semai S. alba dapat diligat pada Gambar 6.
Gambar 6. Hasil pengukuran respons pertumbuhan jumlah daun tanaman semai S. alba pada umur 3 bulan (n = 0 – 13). Tanda * mengindikasikan secara statistik berbeda nyata pada P < 0,05 menurut uji Dunnet. Berdasarkan uji Dunnet respons pertumbuhan jumlah daun semai S. alba menunjukkan respons berbeda nyata terhadap perlakuan variasi naungan 75% yang dibandingakan dengan perlakuan tanpa naungan dengan P < 0,05. Salinitas 3% juga memberikan respons berbeda nyata dengan perlakuan tanpa salinitas terhadap pertumbuhan semai dengan P < 0,05. Perlakuan naungan dan salinitas memberikan pengaruh terhadap respons pertumbuhan jumlah daun tanaman semai S. alba karena diduga kedua perlakuan yang diberikan erat kaitannya dengan proses fotosintesis.
Di setiap perlakuan naungan tingkat salinitas 3% merupakan perlakuan dengan rata-rata jumlah daun terbanyak dengan rata-rata setiap naungannya mencapai 3 daun. Hal ini menunjukkan bahwa pemberian salinitas 3% merupakan tingkat salinitas yang optimum untuk pertumbuhan tanaman semai S. alba selain itu tingkat intensitas cahaya yang tinggi juga merupakan kebutuhan bagi pertumbuhan tanaman semai S. alba. Hal ini sesuai dengan pernyataan Dwijoseputro (1980) yang menyatakan bahwa perbedaan jumlah daun yang
tumbuh dibawah naungan dipengaruhi oleh adanya perbedaan intensitas cahaya yang diperoleh dengan adanya hambatan cahaya masuk mengakibatkan energi foton yang dibutuhkan untuk proses fotosintesis menjadi berkurang sehingga fotosintat yang dihasilkan berkurang oleh karena itu pertumbuhan vegetatif terutama pertumbuhan daun terhambat dan dapat bisa mengakibatkan kematian jaringan tanaman dan diikuti kematian tanaman itu sendiri.
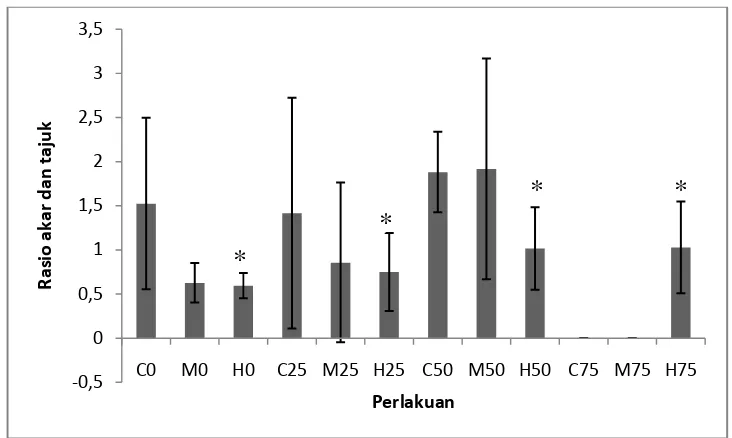
Rasio Tajuk dan Akar Semai S. alba
Pengukuran rasio tajuk dan akar dilakukan pada saat tajuk dan akar telah dipisahkan dan pengukuran berat basah telah dilakukan, sampel kemudian dioven di selama 24 jam dengan suhu 75 oC yang kemudian ditimbang beratnya. Grafik rasio berat tajuk dan akar dapat dilihat pada gambar 7.
Gambar 7. Hasil pengukuran rasio berat kering tajuk dan akar (n = 0 – 13). Tanda * mengindikasikan secara statistik berbeda nyata pada P < 0,05 menurut uji Dunnet.
Pengukuran rasio berat kering tajuk dan akar pada Gambar 7 menunjukkan bahwa rasio akar dan tajuk tertinggi terdapat pada perlakuan dengan variasi naungan 50% dengan tingkat salinitas 1,5% dengan rasio sebesar 1,916 dan rasio
tajuk dan akar terkecil berada pada perlakuan tanpa naungan dengan tingkat salinitas 3% dengan rasio sebesar 0,594. Sedangkan rasio tajuk dan akar yang mendekati 1 adalah perlakuan dengan variasi naungan 50% dengan tingkat salinitas 3%.
Berdasarkan uji Dunnet respons rasio tajuk dan akar tanaman semai S. alba menunjukkan pengaruh nyata dari salinitas 3% yang dibandingkan dengan
salinitas 0% dengan P < 0,05. Rasio tajuk dan akar terkecil pada perlakuan variasi tanpa naungan dengan tingkat salinitas 3% dengan nilai 0,594 (Gambar 7) menunjukkan bahwa berat kering tajuk lebih kecil dari berat kering akar. Hal ini diduga karena ketersediaan energi cahaya matahari yang cukup untuk melakukan fotosistesis namun karena kurangnya ketersediaan unsur hara pada media pasir yang telah disterilkan mengakibatkan pertumbuhan akar lebih cepat. Kekurangan unsur hara tersebut membuat tanaman memiliki pertumbuhan akar yang lebih cepat untuk mencari dan menyerap unsur hara yang lebih banyak, hal ini sesuai dengan pernyataan Lakitan (1994) yang menyatakan bahwa berat kering merupakan akumulasi senyawa organik melalui proses sintesis yang tergantung pada laju fotosintesis tanaman tersebut, sedangkan fotosintesis dipengaruhui oleh kecepatan penyerapan unsur hara di dalam tanaman melalui akar sehingga pertumbuhan akar tanaman akan lebih cepat dalam kondisi kering dan kekurangan unsur hara.
fotosintesis serta karena sifat S. alba yang intoleran terhadap air tawar. Pada perlakuan dengan variasi naungan 50% dengan tingkat salinitas 3% (Gambar 7) menunjukkan bahwa pertumbuhan tajuk dan akar semai S. alba berkembang secara normal dan seimbang dengan nilai rasio akar dan tajuk 1,015, diduga dengan ketersediaan unsur hara yang sedikit pada media pasir yang telah disterilkan dan cadangan makanan yang sedikit pada kotiledon semai S. alba maka energi cahaya matahari yang sesuai adalah sebesar 50%. Hal ini sesuai dengan pernyataan Mulyani et al., (1999) yang menyatakan bahwa pertumbuhan semai suatu tanaman sangat dipengaruhi oleh cadangan makanan yang ada dalam jaringan sel tanaman tersebut dan ketersediaan unsur hara pada media tumbuhnya. Hal ini didukung oleh pernyataan Jumin (2002) menyatakan bahwa pertumbuhan tanaman tidak terlepas dari ketersediaan unsur hara di dalam tanah yang akan menentukan produksi berat kering tanaman. Menurut Nyakpa et al. (1986) pertumbuhan tanaman ditandai dengan pertambahan berat kering tanaman, ketersediaan hara yang optimal bagi tanaman akan diikuti peningkatan aktifitas fotosintesis yang menghasilkan asimilat yang mendukung berat kering tanaman.
mempunyai pengertian bahwa pertumbuhan satu bagian tanaman diikuti pertumbuhan bagian tanaman lainnya.
S. alba merupakan spesies yang berkembang biak dengan biji (generatif) sehingga meskipun diperoleh dari pohon yang sama setiap biji memiliki sifat genetik yang berbeda oleh karena itu kemampuan untuk hidup dan beradaptasi dari setiap semai tanaman S. alba berbeda-beda. Hal ini sesuai dengan pernyataan Mohr dan Schopfer (1995) yang menyatakan bahwa kemampuan suatu tanaman dalam beradaptasi untuk tumbuh dan berkembang sangat dipengaruhi oleh faktor genetik. Dimana pertumbuhan dan perkembangan tanaman dipengaruhi oleh faktor lingkungan dan faktor genetik. Di lingkungannya S. alba tumbuh dengan tingkat salinitas dan intensitas cahaya yang tinggi dengan media tumbuh substrat berpasir. Oleh karena itu perlakuan perbedaan variasi naungan dan tingkat salinitas diduga menjadi penyebab randahnya persentase tumbuh bibit semai S. alba karena kurang dapat beradaptasi.
Rendahnya persentase hidup semai S. alba dibandingkan dengan penelitian jenis-jenis mangrove lainnya diduga karena S. alba berkembangbiak secara generatif dimana cadangan makanan yang tersimpan di kotiledonnya relatif sedikit dibandingkan dengan jenis mangrove yang berkembang biak dengan
vivipari dan kriptovivipari. Hal ini sesuai dengan pernyataan Hachinohe et al. (1999) yang menyatakan S. alba berkembangbiak secara
Media tanam berupa pasir juga sangat mempengaruhi pertumbuhan tanaman semai S. alba mengingat bahwa Sonneratia tumbuh paling dekat dengan laut dengan substrat agak berpasir (Bengen, 1999). Meskipun pasir sungai yang digunakan telah diseterilkan sehingga bakteri dan jamur potogen yang ada pada pasir tersebut telah berkurang (Fahmi, 2011), namun karena kurangnya keberadaan unsur hara pada media tersebut diduga mengakibatkan pertumbuhan tanaman semai S. alba tidak dapat tumbuh dengan maksimal dan rentan terkena hama dan penyakit. Selain itu S. alba juga memiliki biji dengan kotiledon yang kecil hal ini diduga menyebabkan pertumbuhan semai kurang maksimal pada media pasir yang miskin unsur hara dan rentan mati.
Selain itu tingkat salinitas terhadap pertumbuhan semai S. alba juga sangat berpengaruh dimana hasil pengukuran respons pertumbuhan semai S. alba tertinggi umumnya dengan tingkat salinitas 3%. Hal ini didukung oleh pernyataan Bengen (1999) yang menyatakan bahwa zonasi mangrove di Indonesia di daerah paling dekat dengan laut, dengan substrat agak berpasir, umumnya sering ditumbuhi Avicennia dan Sonneratia. Selain itu tingginya tingkat kematian pada perlakuan tingkat salinitas 0% dan 1,5% diduga karena tanaman semai S. alba kurang dapat beradaptasi dengan tingkat salinitas yang rendah mengingat bahwa tanaman semai S. alba merupakan spesies halopita obligat yang mampu tumbuh di lingkungan dengan salinitas yang tinggi namun kurang dapat beradaptasi dengan lingkungan dengan salinitas rendah (Ewusie, 1990). Hal ini didukung oleh pernyataan Tomlinson (1986) yang menyatakan bahwa S. alba merupakan spesies yang intoleran terhadap air tawar dalam kurun waktu yang lama.
Polyisoprenoid
Dari hasil ekstraksi daun dan akar S. alba diperoleh nilai NSL (Nonsaponifiable Lipids) dan polyisoprenoid. Adapaun hasil ekstraksi yang diperoleh dapat dilihat pada Tabel 3.
Tabel 3. Nonsaponifieble Lipids (NSL) pada semai S. alba.
Jenis Perlakuan Jaringan Berat
Sampel (g)
NSL (mg) Polyisoprenoid
(ml)
perlakuan tanpa naungan dengan salinitas 3% pada jaringan daun adalah sebesar 190,5 ml dan sebesar 133,4 ml pada jaringan akar sedangkan pada perlakuan dengan variasi naungan 75% dengan salinitas 3% adalah sebesar 185,2 ml pada jaringan daun dan sebesar 231,9 ml pada jaringan akar.
Metabolit sekunder merupakan salah satu senyawa dari lipid yang berperan dalam adaptasi mangrove terhadap cekaman lingkungan. Secara umum polyisoprenoid merupakan fraksi lipid. Kandungan polyisoprenoid pada perlakuan tanpa naungan dengan salinitas 3% pada yang terakumulasi pada jaringan daun lebih banyak dibandingkan dengan kandungan polyisoprenoid pada jaringan akar hal ini diduga karena pada perlakuan ini semai S. alba mendapatkan intensitas cahaya yang tinggi untuk fotosintesis namun karena penggunaan media pasir yang miskin hara menyebabkan hasil fotosintesis difokuskan untuk pertumbuhan akar dibandingkan pertumbuhan tajuk. Sehingga lipid yang merupakan salah satu hasil fotosintesis digunakan sebagai energi untuk metabolisme pertumbuhan akar. Hal ini sesuai dengan studi Estiti (1995) bahwa lipid berfungsi sebagai pembentuk struktur membran sel, sebagai bahan cadangan makanan dan sebagai sumber energi dalam metabolisme pertumbuhan.
pada semai berjalan lambat dan metabolisme sel tanaman yang berlangsung juga berjalan lambat sehingga penggunaan energi untuk kegiatan metabolisme yang diperlukan semakin sedikit.
Tingginya kandungan polyisoprenoid pada perlakuan perlakuan variasi naungan 75% dengan salinitas 3% dibandingkan tanpa naungan dengan salinitas 3% (Tabel 3) diduga disebabkan oleh rendahnya intensitas cahaya yang diterima oleh S. alba yang merupakan tanaman yang lebih menyukai intensitas cahaya yang tinggi. Hal ini sesuai dengan studi Sudha dan Ravishankar (2002) yang menyatakan bahwa faktor biotik dan abiotik yang bersifat mencekam pertumbuhan dapat meningkatkan senyawa lipid sebagai metabolit sekunder.
Polyisoprenoid yang dihasilkan dari S. alba pada standar dolichol dapat dilihat pada Gambar 8.
Gambar 8. Thin-Layer Chromatography polyisoprenoid alkohol (dolichol) dari semai
S. alba dimana S merupakan standar dolichol, 1-3 merupakan daun semai S.
Berdasarkan Gambar 8 dapat dilihat bahwa konsentrasi dolichol pada perlakuan variasi tanpa naungan lebih pekat baik pada jaringan akar maupun pada jaringan daun semai S. alba jika dibandingkan dengan perlakuan variasi naungan 75%. Hal ini menunjukkan bahwa dengan adanya penambahan naungan maka akan terjadi penurunan kadar dolichol pada jaringan semai S.alba. Hal ini diduga karena dolichol berperan dalam adaptasi semai S. alba terhadap cekaman lingkungan dalam hal ini cekaman intensitas cahaya yang tinggi. Hal ini sesuai dengan pernyataan Swiezewska dan Danikiewicz (2005) yang menyatakan bahwa konsentrasi dolichol dan polyprenol akan meningkat di setiap jaringan tanaman dengan pertambahan umur dan dengan meningkatnya cekaman lingkungan.
Pada jaringan akar dan daun semai S. alba pada perlakuan yang sama terdapat perbedaan konsentrasi dolichol hal ini diduga karena perbedaan umur jaringan dan perbedaan gen dari S. alba. Hal ini sesuai dengan pernyataan Tateyama et al. (1999) yang menyatakan distribusi rantai panjang polyprenols belum tentu sama dengan rantai panjang dolichol di jaringan yang sama, hal ini didukung pernyataan Suga et al.(1989) yang menyatakan konsentrasi polyisoprenoid pada tanaman mengalami perubahan yang disebabkan oleh perbedaan umur dan musim.
KESIMPULAN DAN SARAN
Kesimpulan
Perlakuan variasi naungan dan salinitas yang diberikan ke semai S. alba memberikan pengaruh nyata terhadap respons pertumbuhan yang diamati. Perlakuan variasi tanpa naungan dengan salinitas 3% memberikan respons pertumbuhan yang optimum dibandingkan dengan perlakuan yang lainnya dan diduga polyisoprenoid berperan dalam adaptasi semai S. alba terhadap ketersediaan intensitas cahaya.
Saran
DAFTAR PUSTAKA
Anwar C. 1997. Pengaruh pemberian naungan di persemaian terhadap pertumbuhan semai mangrove. Buletin Pengelolaan DAS 4 (1) : 97-106 Ball MC., Pidsley SM. 1995. Growth responsse to salinity in relation to
distribution of two mangrove species, Sonneratia alba and Sonneratia lanceolata. Funct Ecol 9 : 77–85
Basyuni, M., Baba, S., Inafuku, M., Iwasaki, H., Kinjo, K., Oku, H. 2009. Expression of terpenoid synthase mRNA and terpenoid content in salt stressed mangrove. Journal of Plant Physiology 166 : 1786-1800
Basyuni, M., Baba, S., Kinjo, Y., Lollie, A. P. P., Hakim, L., Oku, H. 2012. Salt-dependent increase in triterpenoids is reversible upon transfer to fresh water in mangrove plants Kandelia candel And Bruguiera gymnorrhiza. Journal of Plant Physiology 169 : 1903-1908.
Basyuni, M., Lollie, A. P. P., Julayha, Nurainum, H., Oku, H. 2012. Non-saponifiable lipid composition of four salt-secretor and non-secretor mangrove species from North Sumatra, Indonesia. Makara Journal of Science 16(2) : 89-94.
Basyuni, M., Baba, S., Kinjo, Y., Oku, H. 2012. Salinity increases the triterpenoid content of a salt secretor and a non-salt secretor mangrove. Aquatic Botany 97: 17-23.
Bengen, D. 1999. Pedoman Teknis Pengenalan dan Pengelolaan Ekosiistem Mangrove. PKSPL. IPB. Bogor.
Chapman, V. J. 1975. Mangrove vegetation. Strauss and Cramer GmbH, German.
Clarke, P.J. & Allaway, W.G. .1993. The regeneration niche of the grey mangrove (Avicennia marina): effects of salinity, light and sediment factors on establishment, growth and survival in the field. Oecologia 93 : 548–556. Djukri, B.S. Purwoko. 2003. Pengaruh naungan paranet terhadap sifat toleransi
tanaman talas (Colocasia esculenta (L.) Schott). Ilmu Pertanian 10 : 17–25. Dwidjoseputro, D. 1978. Pengantar Fisiologi Tumbuhan. PT Gramedia : Jakarta Ewusie, J. Y. 1990. Ekologi Tropika. Bandung: Penerbit ITB.
Farnsworth, E. J., Ellison, A. M. .1996. Sun-shade adaptability of the red mangrove, rhizophora mangle (rhizophoraceae): changes trough ontogeny at several levels of biological organization. America Journal of Botany 83 : 1131-1143.
Field, C. 1996. Restoration of Mangrove Ecosystems. ISME.
Giri C., Ochieng E., Tieszen L. L., Zhu Z., Singh A., Loveland T., Masek J., Duke N. 2011.Status and distribution of mangrove forests of the world using earth observation satellite data. Global Ecology and Biogeography 20 : 154-159 Haan, J. H. De. 1931. Het een en ander over de Tjilatjapꞌ sche vloedbosschen.
Tectona 24 : 39 – 76. (In Dutch with English summary).
Hachinohe, H.,Suko, O., dan Ida, A.1999. NURSERY MANUAL FOR MANGROVE SPECIES-at Benoa Port in Bali.The Development Of Sustainable Mangrove Management Project. Ministry of Forest and Estate Crops, Indonesia and Japan Internatio.
Hoffman, L.L., Anten, N. P. R., Ramos, M. M. Ackerly, D. D. 2007. Salinity and light interactively affect neotropical mangrove seedlings at the leaf and whole plant levels.Oecologia 150 : 545-556.
Husch, B., Miller, C.I. and Beers, T.W. 1972. Forest Mensuration. The Ronald Press Company. New York.
Keliat, S. R., Yunasfi, Utomo, B. 2013. Pertumbuhan bibit avicenia marina pada berbagai intensitas naungan. Universitas Sumatera Utara.
Kusmana, C., Wilarso, S., Hilwan, I., Pamoengkas, P., Wibowo, C., Tiryana, T., Triswanto, A., dan Yunasfi, H. 2003. Teknik rehabilitasi mangrove. Fakultas Kehutanan IPB, Bogor.
Lakitan, B. 1994. Dasar-Dasar Klimatologi. Penerbit PT. Raja Grafindo Persada. Maghfoer, M.D. dan Koesriharti, 1998. Rekayasa 12 teknologi penaungan dalam
sistem budidaya tanaman paprika (Capsicum annum L.). Jurnal Penelitian Ilmu-ilmu Teknik (Enginering) 10 : 89-95.
Marjenah. 2001. Pengaruh perbedaan naungan di persemaian terhadap pertumbuhan dan respons morfologi dua jenis semai meranti. Jurnal Ilmiah Kehutanan Rimba Kalimantan 6 : 16–23.
Marschner.1995. Mineral nutrition of higher plants. Academic press. United States of America.
Prayunita, Basyuni, M., Putri, L. A. P. 2012. Respons pertumbuhan dan biomassa semai bakau minyak (Rhizophora apiculata BI) terhadap salinitas dan kandungan lipidnya pada tingkat pohon. Universitas Sumatera Utara. Medan.
Naidoo G. 1987. Effects of salinity and nitrogen on growth and water relations in the mangrove, Avicennia marina (Forsk.) Vierh. New Phytologist 107 : 317–325.
Noor, Y., M. Khazali, dan I N.N. Suryadiputra. 1999. Panduan Pengenalan Mangrove di Indonesia. PHKA/WI-IP, Bogor.
Nugroho, H. 2012. 1,8 juta hektare hutan mangrove di Indonesia rusak. Kompas, 5 November 2012.
Radji, M. 2005. Peranan bioteknologi dan mikroba endofit dalam pengembangan obat herbal.laboratorium mikrobiologi dan bioteknologi. Departemen Farmasi, FMIPA-UI. Majalah Ilmu Kefarmasian 3: 113–126.
Ramayani, Basyuni, M., Putri L. A. P. 2012. Pengaruh salinitas terhadap pertumbuhan dan biomassa semai non-sekresi Ceriop tagal dan kandungan lipid pada tingkat pohon. Universitas Sumatera Utara. Medan.
Reimold R. J., Queen W. H. 1974. Ecology of Halophytes. Academic Press, New York.
Sagami, H., Kurisaki, A., Ogura, K., and Chojnacki T. 1992. Separation of dolichol from dehydrodolichol by a simple two-plate thin-layer chromatography. Journal of Lipid research 33 : 1857–1861.
Schmidt. L. 2002. Pedoman penanganan benih tanaman hutan tropis dan subtropis. Ditjen Rehabilitasi Lahan dan Perhutanan Sosial. Jakarta.
Scholander, P.F., Hammel, H.T., Hemmingsen, E., Garey, W., 1962. Salt balance in mangroves.Plant Physiol 37: 722–729.
Sialoho, P. E., Basyuni, M., Putri, L. A. P. 2013. Pengaruh variasi konsentrasi garam dan adaptasi air tawar terhadap respons pertumbuhan dan komposisi triterpenoid dan fitosterol semai bakau (Rhizophora stylosa). Universitas Sumatera Utara. Medan.
Simarmata, E., Yunasfi, Utomo, B. 2011. Pertumbuhan bibit Rhizophora apiculata pada berbagai intensitas naungan. Universitas Sumatera Utara. Medan.
Smith, T.J. III (1987b) Effects of seed predators and light level on the distribution of Avicennia marina (Forsk.) Vierh. in tropical, tidal forests. Estuarine, Coastal and Shelf Science, 25 : 43–51.
Suga, T., Ohta, S., Nakai, A., Munesada, K. 1989. Glycinoprenols: novel polyprenols possessing a phytyl residue from the leaves of soybean. The Journal of Organic Chemistry 54: 3390-3393.
Sukristijono Sukardjo. 1984. EKOSISTEM MANGROVE .Oseana 4 : 102-115. Surmacz, L., Swiezewska E. 2011. Polyisoprenoids – Secondary metabolites or
physiologically important superlipids?. Biochemical and Biophysical Research Communications 407 : 627 – 632.
Swiezewska E., Danikiewicz, W. Polyisoprenoids: structure, biosynthesis and function, Prog. Lipid Res 44 : 235–258.
Tateyama, S., R. Wititsuwannakul, D. Wititsuwannakul, H. Sagami, K. Ogura. 1999. Dolichols of rubber plant, ginkgo and pine. Phytochemistry 51: 11-16. Thomas J. Smith III.1987.Effects of seed predators and light level on the
distribution of Avicennia marina (Forsk.) Vierh. in tropical, tidal forests. Estuaria, Coastal and Shelf Science 25: 43–51.
Tomlinson PB. 1986.. The Botany of Mangroves. London: Cambridge University Press.
Valiela, I., Bowen, J. L., York, J. K. 2001. Mangrove forest: one of the world threatened major tropical environments. BioScience. 10 : 807-815
Watson, J. G. 1928. Mangrove Forest of The Malay Paninsula. Malay Forest Wieslaw, J. J., Swiezewska, E., Sasak, W., Chojnacki, T. 1994. Occurrence of
Polyprenols and Dolichols in Plants. Journal of Plant Physiology 143 : 448–452
Lampiran 1. Analisis Sidik Ragam (ANOVA) dan Uji Dunnet Semai S. alba
1. Hasil sidik ragam tinggi semai S. alba
Sumber Keragaman Jumlah Kuadrat Db Kuadrat Tengah F Sig.
Model Koreksi 103.658a 11 9.423 7.924 .000
Total Koreksi 274.907 155
2. Uji Dunnet tinggi semai S. alba pada variasi naungan
(I)
Indeks Terkecil Indeks Terbesar
25 0 -.1897 .24695 .783 -.7760 .3965
50 0 -.4538 .24695 .166 -1.0401 .1324
75 0 -.6846* .24695 .017 -1.2709 -.0983
3. Uji Dunnet tinggi semai S. alba pada variasi salinitas
(I)
Indeks Terkecil Indeks Terbesar
1.5 0 .1308 .21387 .766 -.3470 .6085
4. Hasil sidik ragam diameter semai S. alba
Sumber Keragaman Jumlah Kuadrat Db Kuadrat Tengah F Sig.
Model Koreksi .400a 11 .036 9.469 .000
5. Uji Dunnet diameter semai S. alba pada variasi naungan
(I)
Indeks Terkecil Indeks Terbesar
25 0 -.0209 .01403 .315 -.0542 .0124
50 0 -.0239 .01403 .216 -.0572 .0094
75 0 -.0465* .01403 .003 -.0798 -.0131
6. Uji Dunnet diameter semai S. alba pada variasi salinitas
(I)
Indeks Terkecil Indeks Terbesar
1.5 0 .0050 .01215 .885 -.0222 .0322
7. Hasil sidik ragam berat basah akar semai S. alba
Sumber Keragaman Jumlah Kuadrat Db Kuadrat Tengah F Sig.
Model Koreksi .302a 11 .027 25.015 .000
8. Uji Dunnet berat basah akar semai S. alba pada variasi naungan
(I)
Indeks Terkecil Indeks Terbesar
25 0 -.0392* .00750 .000 -.0570 -.0214
50 0 -.0610* .00750 .000 -.0788 -.0432
75 0 -.0656* .00750 .000 -.0834 -.0478
9. Uji Dunnet berat basah akar semai S. alba pada variasi salinitas
(I)
Indeks Terkecil Indeks Terbesar
1.5 0 .0069 .00649 .461 -.0076 .0214
10.Hasil sidik ragam berat kering akar semai S. alba
Sumber Keragaman Jumlah Kuadrat Db Kuadrat Tengah F Sig.
Model Koreksi .077a 11 .007 19.777 .000
11. Uji Dunnet berat kering akar semai S. alba pada variasi naungan
(I)
Indeks Terkecil Indeks Terbesar
25 0 -.018128* .0042601 .000 -.028242 -.008014
50 0 -.026715* .0042601 .000 -.036829 -.016602
75 0 -.030213* .0042601 .000 -.040327 -.020099
12.Uji Dunnet berat kering akar semai S. alba pada variasi salinitas
(I)
Indeks Terkecil Indeks Terbesar
1.5 0 .007017 .0036894 .106 -.001225 .015259
13.Hasil sidik ragam berat basah tajuk semai S. alba
Sumber Keragaman Jumlah Kuadrat Db Kuadrat Tengah F Sig.
Model Koreksi .382a 11 .035 7.751 .000
14.Uji Dunnet berat basah tajuk semai S. alba pada variasi naungan
(I)
Indeks Terkecil Indeks Terbesar
25 0 -.0254 .01515 .227 -.0613 .0106
50 0 -.0169 .01515 .544 -.0529 .0190
75 0 -.0372* .01515 .041 -.0731 -.0012
15.Uji Dunnet berat basah tajuk semai S. alba pada variasi salinitas
(I)
Indeks Terkecil Indeks Terbesar
1.5 0 .0025 .01312 .974 -.0268 .0318