DETEKSI DAN KARAKTERISASI PARSIAL CORONAVIRUS
DARI KELELAWAR ASAL PROVINSI GORONTALO DAN
SULAWESI UTARA
WENTI DWI FEBRIANI
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa tesis berjudul Deteksi dan Karakterisasi Parsial Coronavirus dari Kelelawar asal Provinsi Gorontalo dan Sulawesi Utara adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Oktober 2016
RINGKASAN
WENTI DWI FEBRIANI. Deteksi dan Karakterisasi Parsial Coronavirus dari Kelelawar Asal Provinsi Gorontalo dan Sulawesi Utara. Dibimbing oleh JOKO PAMUNGKAS dan LIGAYA ITA TUMBELAKA.
Kelelawar dikenal sebagai reservoar alami untuk virus-virus yang baru muncul, seperti Lyssaviruses, Coronavirus, Ebola virus, virus Nipah, dan banyak lainnya. Karena kelimpahan populasi, distribusi yang luas dan mobilitasnya, kelelawar memiliki risiko lebih besar sebagai sumber penularan zoonosis dibandingkan dengan hewan lain. Peran kelelawar sebagai reservoar bagi banyak pathogen semakin mendapat perhatian, setelah ditemukannya kasus epidemik Severe Acute Respiratory Coronavirus (SARS-CoV) pada tahun 2003 dan Middle East Respiratory Syndrome Coronavirus (MERS-CoV) 2012, sehingga masyarakat ilmiah mulai memberikan perhatian khusus terhadap coronavirus pada kelelawar. Virus yang mirip dengan virus penyebab SARS juga ditemukan pada kelelawar dengan tingkat prevalensi lebih tinggi.
Penelitian ini dilakukan sebagai bagian dari proyek besar PREDICT Indonesia, khususnya untuk memeriksa coronavirus pada kelelawar dari Provinsi Gorontalo dan Sulawesi Utara, Indonesia. Dari 151 sampel feses atau sampel usap rektum kelelawar besar pemakan buah (Pteropus alecto) yang dianalisis di laboratorium menggunakan teknik nested Polymerase Chain Reaction (PCR) untuk mengamplifikasi target dari gen RNA-dependent RNA polymerase (RdRp) dengan ukuran produk 434 pasang basa, ditemukan 65 sampel diduga positif (presumptive positive). Enam belas dari 65 sampel presumptive positive, diperiksa lebih lanjut dengan teknik sekuensing untuk dikonfirmasi sebagai Coronavirus. Pohon filogenetik terhadap 16 sampel yang dikonfirmasi positif coronavirus kemudian dianalisis dengan MEGA-6.0 dan hasilnya menunjukkan bahwa sekuen gen RdRp secara parsial memiliki tingkat kemiripan yang tinggi dengan coronavirus pada kelelawar yang dilaporkan sebelumnya di Indonesia dan Thailand.
SUMMARY
WENTI DWI FEBRIANI. Detection and Partial Characterization of Coronavirus from Bats Originated from Gorontalo and North Sulawesi Provinces. Supervised by JOKO PAMUNGKAS and LIGAYA ITA TUMBELAKA.
Bats are known as natural reservoirs for potential emerging infectious viruses, such as Lyssaviruses, Coronaviruses, Ebola viruses, Nipah virus, and many others. Because of their abudance in population, wide distribution and mobility, bats have a greater risk as source for zoonotic transmission than other animals. Despite the facts of their role as reservoirs for many pathogens, not until an epidemic of Severe Acute Respiratory Coronavirus (SARS-CoV) in 2003 and Middle-East Respiratory Syndrome Coronavirus (MERS-CoV) in 2012, that scientific community pay much attention about coronavirus in bats. SARS-like virus also found in bats with a higher prevalence rate.
This study was conducted as part of bigger PREDICT Indonesia project, in particular to examine coronavirus in bats from Gorontalo and North Sulawesi provinces, Indonesia. As many as 151 fecal or rectal swab samples collected from flying foxes (Pteropus alecto) were analyzed in the laboratory using Consensus Polymerase Chain Reaction (PCR) technique to amplify the target sequence from RNA-dependent RNA Polymerase (RdRP) gene with 434 basepair product, resulted 65 samples determined as presumptive positive. Sixteen out of 65 presumptive positive samples by PCR were analyzed further by nucleotide sequencing and confirmed coronavirus positive. Phylogenetic tree analyses to the 16 coronavirus confirmed-sequences were constructed with MEGA-6.0 and the results showed that the RdRP partial gene sequences have higher similarities to the previously reported bat coronavirus findings in Indonesia and Thailand.
© Hak Cipta Milik IPB, Tahun 2016
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Mikrobiologi Medik
DETEKSI DAN KARAKTERISASI PARSIAL CORONAVIRUS
DARI KELELAWAR ASAL PROVINSI GORONTALO DAN
SULAWESI UTARA
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2016
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah subhanahu wa ta’ala atas segala karunia-Nya sehingga karya ilmiah ini berhasil diselesaikan. Tema yang dipilih dalam penelitian yang dilaksanakan sejak bulan Desember 2015 ini ialah deteksi virus, dengan judul Deteksi dan karakterisasi parsial coronavirus dari kelelawar asal Provinsi Gorontalo dan Sulawesi Utara. Penelitian ini didanai oleh Kementerian Riset Teknologi dan Pendidikan Tinggi Republik Indonesia yang bekerjasama dengan PREDICT. Tesis ini merupakan salah satu syarat untuk mendapatkan gelar Magister Sains di Program Studi Mikrobiologi Medik, Sekolah Pascasarjana, Institut Pertanian Bogor.
Terima kasih penulis ucapkan kepada Bapak Dr Drh Joko Pamungkas MSc dan Ibu Dr Drh Ligaya ITA Tumbelaka MSc.,Sp.MP selaku pembimbing yang memberikan banyak, arahan, bimbingan, selama proses penelitian dan penulisan tesis, Profesor Dr Drh Fachriyan H Pasaribu selaku ketua Prodi Mikrobiologi medik, Ibu Drh. Surachmi Setyaningsih Ph.D selaku penguji luar komisi atas kesediaan waktu dan masukan yang diberikan, kepala staf laboratorium Bioteknologi Bapak Dr.Uus Saepuloh M.Biomed yang banyak memberikan arahan dan staf pusat studi satwa primata (PSSP) Institut Pertanian Bogor yang telah membantu selama pengumpulan data, Bapak dan Ibu staf pengajar yang memberi dukungan penelitian, kepada teman-teman Mikrobiologi Medik 2014, teman-teman kos Tridara yang telah memberikan dukungan, Bapak, Ibu dan kakak dirumah, serta seluruh keluarga, atas segala doa dan kasih sayangnya.
Semoga karya ilmiah ini bermanfaat.
DAFTAR ISI
DAFTAR TABEL v
DAFTAR GAMBAR v
DAFTAR LAMPIRAN v
DAFTAR TABEL
1 Identitas persentase nukleotida Coronavirus 12
DAFTAR GAMBAR
1 Black Flying fox (Pteropus alecto) 4
2 Struktur coronavirus 5
3 Diagram skematis gen yang diamplifikasi deteksi Coronavirus 6
4 Siklus hidup coronavirus 7
5 Visualisasi hasil nested PCR 11
6 Visualisasi hasil nested PCR 11
7 Pohon filogenetik 14
DAFTAR LAMPIRAN
1 Pensejajaran sekuens Bat coronavirus INDSWBT regio RdRp isolat
1
PENDAHULUAN
Latar Belakang
Indonesia memiliki keragaman hayati melimpah, baik flora ataupun fauna dan memiliki luas wilayah hutan sekitar 162 juta hektar dengan berbagai ekosistem di dalamnya termasuk habitat kelelawar (Yudhoyono 2015). Pulau Sulawesi yang sebagian besar wilayahnya termasuk dalam zona ekologi garis Wallace menjadi rumah bagi banyak flora atau fauna di Indonesia. Sekitar 62 spesies kelelawar di dunia terdapat di Sulawesi (Heinrichs et al. 1997). Kelelawar besar pemakan buah diketahui merupakan reservoar yang berpotensi sebagai penyebab munculnya patogen. Karena kelimpahan, distribusi dan mobilitasnya kelelawar memberi resiko tinggi untuk transmisi zoonosis dibandingkan dengan hewan lain (Calisher et al. 2006). Interaksi satwa liar dengan manusia tersebut diperantarai oleh beberapa pemicu (drivers): pembukaan hutan, industri penambangan, ekowisata, transportasi, perburuan dan perdagangan satwa liar, serta konsumsi daging satwa liar. Masyarakat Sulawesi Utara (suku Minahasa) dikenal sebagai konsumen berbagai hewan domestik ataupun hewan liar. Kelelawar adalah salah satu daging yang paling disukai, bahkan menjadi extreme culinary (Whitten et al. 1987).
Pada mulanya infeksi coronavirus pada manusia kurang begitu diperhatikan hingga ditemukannya coronavirus tipe baru yang menyebabkan epidemik pada tahun 2003, yaitu SARS-CoV serta Middle-East Respiratory Syndrome Coronavirus (MERS-CoV) pada tahun 2012 yang menginfeksi lebih dari 8000 individu di dunia dan menyebabkan kematian sebanyak 774 individu (Poon et al. 2004). Gejala umum pada SARS meliputi demam tinggi dengan suhu tubuh melebihi 38°C yang diikuti dengan gejala pneumonia ([CDC] 2015; Mahon et al. 2014). Di Asia tenggara, ditemukan kelelawar yang diketahui membawa coronavirus di beberapa negara termasuk Filipina ( Watanabe et al. 2010) dan Thailand (Gouilh et al. 2011).
Implementasi konsep Satu Kesehatan atau One Health diperlukan dalam rangka pencegahan penularan agen penyakit dari satwa liar kepada manusia. Lebih jauh, implementasi konsep tersebut diharapkan akan dapat mencegah kejadian pandemik suatu penyakit yang dapat mengancam kehidupan manusia. One Health merupakan upaya kolaboratif dari berbagai disiplin yang bekerja di tingkat lokal, nasional dan global untuk mencapai kesehatan yang optimal untuk manusia, hewan, dan lingkungan (Barrett dan Osofsky 2013). Konsep One Health juga akan mendorong kemitraan yang lebih erat diberbagai sektor pemerintah dan para stakeholder terkait untuk mengembangkan dan mengevaluasi metode diagnostik baru, pengobatan dan vaksin untuk pencegahan dan pengendalian penyakit lintas spesies, secara bersamaan dengan upaya untuk menginformasikan dan mengedukasi para pemimpin politik dan publik.
2
disease) yang bersifat zoonotik. Adanya interaksi antara kelelawar dan manusia (human-animal interface) khususnya di wilayah Sulawesi Utara dapat meningkatkan resiko tertularnya manusia terhadap penyakit zoonosis. Di Indonesia masih belum banyak penelitian mengenai coronavirus pada kelelawar, oleh karena itu diperlukan adanya penelitian mengenai identifikasi dan karakterisasi parsial coronavirus dari kelelawar besar pemakan buah yang bertujuan untuk mendeteksi dan karakterisasi parsial materi genetik dari kelelawar besar pemakan buah di Provinsi Gorontalo dan Sulawesi Utara dan membandingkan tingkat kekerabatan genetik virus yang dideteksi dan diidentifikasi tersebut. Penelitian ini diharapkan dapat memberikan informasi mengenai virus yang dideteksi dan diidentifikasi pada kelelawar besar pemakan buah serta tingkat kekerabatan genetiknya untuk membantu pihak yang berwenang untuk membangun kebijakan terkait.
Perumusan Masalah
Hasil surveilans virus pada kelelawar besar pemakan buah ditemukan beberapa jenis virus yang dikategorikan sebagai penyebab penyakit yang baru muncul (emerging infectious disease) yang bersifat zoonosis. Adanya interaksi antara kelelawar dan manusia (human-animal interface) khususnya di wilayah Sulawesi Utara dan Gorontalo dapat meningkatkan risiko tertularnya manusia oleh penyakit zoonotik baru ini. Dengan melakukan penelilitan mengenai identifikasi dan karakterisasi parsial coronavirus dari kelelawar besar pemakan buah diharapkan dapat mencegah risiko terjadinya penularan coronavirus pada manusia yang dapat bersifat fatal.
Tujuan Penelitian
1. Deteksi coronavirus dari kelelawar besar pemakan buah asal Provinsi Gorontalo dan Sulawesi Utara
2. Karakterisasi pada tingkat molekular dari genom coronavirus
3. Menentukan tingkat kekerabatan (melalui pohon filogenteik) virus yang dideteksi dan diidentifikasi dari kelelawar.
Manfaat Penelitian
Manfaat yang diharapkan dari penelitian ini adalah dapat memberikan informasi mengenai identifikasi dan karakterisasi parsial virus yang dideteksi pada kelelawar besar pemakan buah dan dari pohon filogenetik terkait kekerabatannya.
Ruang Lingkup Penelitian
3
2
TINJAUAN PUSTAKA
Kelelawar
Kelelawar merupakan salah satu jenis mamalia yang dikenal oleh masyarakat Indonesia. Di Indonesia bagian timur kelelawar disebut paniki, niki atau lawa; orang Sunda menyebutnya kampret, lalai; orang Jawa menyebutnya lowo, lawa, codot, kampret; Suku Dayak di Kalimantan menyebutnya hawa, prok, cecadu, kusing dan tayo (Suyanto 2001).
Biologi Kelelawar
Kelelawar merupakan satu-satunya mamalia yang dapat terbang dan digolongkan dalam Bangsa Chiroptera yang berarti mempunyai “sayap tangan”, karena anggota tubuh bagian depannya termodifikasi menjadi sayap, meskipun berbeda dengan sayap pada burung. Sayap kelelawar merupakan perluasan tubuh yang berdaging, tidak berbulu dan terbentuk dari membran elastis berotot yang dinamakan patagium. Sayap ini berfungsi untuk terbang dan menyelimuti tubuhnya ketika bergantung terbalik (Lekagul dan McNeely, 1977). Pada kelelawar betina patagium juga berfungsi untuk memegang anaknya yang baru dilahirkan dengan posisi kepala di bawah. Berdasarkan jenis makanannya, kelelawar digolongkan menjadi dua yaitu:
1. Kelelawar pemakan buah (Megachiroptera), yang memiliki ukuran tubuh relatif besar dengan bobot berkisar antara 10-1500 g. Jenis dari Marga Balionycteris, Chironax dan Aethalops memiliki bobot sekitar 10 gram, sedangkan kalong kapuk (Pteropus vampyrus) yang berukuran paling besar berbobot lebih dari 1500 gram dengan bentangan sayap mencapai 1700 mm dan panjang lengan bawah (FA : Fore Arm) berkisar antara 36 - 228 mm.
2. Kelelawar pemakan serangga (Microchiroptera) memiliki ukuran tubuh yang lebih kecil dengan kisaran bobot tubuh antara 2 - 196 g, panjang lengan bawah (FA) 22 - 115 mm (Suyanto 2001).
Kelelawar membutuhkan lebih banyak oksigen pada saat terbang, yaitu 27 ml O2/g bobot tubuh, sedangkan saat tidak terbang kelelawar hanya membutuhkan
7 ml O2/g bobot tubuh. Pada saat terbang jantung kelelawar berdenyut lebih cepat
yaitu 822 kali/menit, sedangkan pada saat tidak terbang hanya 522 kali/menit. Untuk mendukung kebutuhan oksigen yang tinggi, jantung kelelawar berukuran relatif lebih besar dibandingkan kelompok hewan lain. Jantung kelelawar berukuran 0.09% dari bobot tubuhnya, sedangkan hewan lain hanya 0.05% dari bobot tubuhnya. Kebutuhan energi yang tinggi pada saat terbang mengharuskan kelelawar makan dalam jumlah banyak (Suyanto 2001).
Family Pteropodidae
4
Notopteris. Ekor dan selaput ekor berukuran relatif kecil atau tidak terdapat sama sekali (Corbet dan Hill 1992).
Kelelawar dari famili Pteropodidae beraktivitas pada sore dan malam hari, terkadang terbang hingga puluhan kilometer untuk mencari makan. Makanan utama famili ini adalah buah, bunga, nektar dan serbuk sari. Sebagian besar kelelawar ini menggelantung di cabang pohon atau dedaunan, sendiri atau membentuk kelompok hingga ratusan individu. Kelelawar dari famili Pteropodidae tidak dapat melakukan ekolokasi namun terdapat satu jenis dari Marga Rousettus yang melakukan ekolokasi dengan menggunakan suara decak yang dihasilkan oleh lidah, dan jenis ini dapat ditemukan di gua (Corbet dan Hill 1999; Altringham 1996). Secara ekologi jenis-jenis kelelawar dari famili Pteropodidae memiliki peran sebagai penyerbuk, penyebar biji tubuhan ataupun tanaman komersial. Sehingga keberadaan mereka merupakan salah satu kunci keberhasilan regenereasi hutan maupun dan produksi buah-buahan. Famili Pteropodidae terdiri dari 42 Marga dan 169 jenis di dunia (Nowak 1999).
Gambar 1 Black flying-fox (Pteropus alecto). Sumber : Nick Edards (www.enigmatech.com.au)
Taksonomi Pteropus alecto
Kingdom : Animalia Phylum : Chordata Class : Mamalia Order : Chiroptera Family : Pteropodidae Genus : Pteropus
5
Coronavirus
Coronavirus menginfeksi berbagai mamalia serta spesies burung. Pada umumnya dapat menyebabkan gangguan pernapasan atau infeksi usus, tetapi beberapa coronavirus juga dapat menginfeksi organ lain (hati, ginjal, dan otak).
Taksonomi Coronavirus
Ordo : Nidovirales Family : Coronaviridae Subfamily : Coronavirinae
Genus Alphacoronavirus; tipe spesies – Alphacoronavirus 1 Genus Betacoronavirus; tipe spesies – Murine coronavirus Genus Deltacoronavirus; tipe spesies – Bulbul coronavirus HKU11
Genus Gammacoronavirus; tipe spesies – Avian coronavirus Subfamily: Torovirinae
Genus Bafinivirus; White Bream virus
Genus Torovirus; type species – Equine torovirus (Drexler et al. 2014)
Struktur Coronavirus
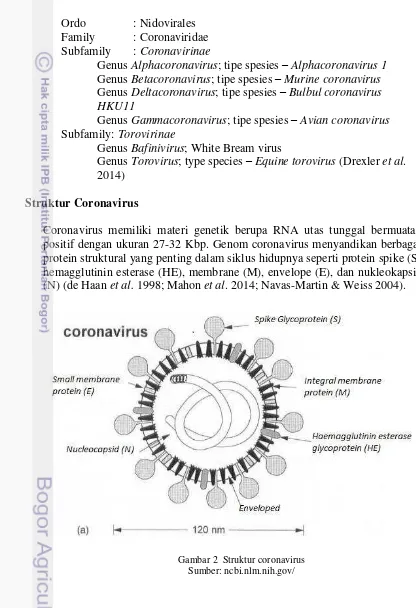
Coronavirus memiliki materi genetik berupa RNA utas tunggal bermuatan positif dengan ukuran 27-32 Kbp. Genom coronavirus menyandikan berbagai protein struktural yang penting dalam siklus hidupnya seperti protein spike (S), hemagglutinin esterase (HE), membrane (M), envelope (E), dan nukleokapsid (N) (de Haan et al. 1998; Mahon et al. 2014; Navas-Martin & Weiss 2004).
6
Genom coronavirus
Gambar 3 Diagram skematis gen yang diamplifikasi untuk deteksi coronavirus dari kelelawar berdasarkan replikasi gen coronavirus. Sumber: ncbi.nlm.nih.gov/
Protein yang terdapat dalam coronavirus berupa S (spike) protein (150k), HE protein (65kD), M (membran) protein, E (envelope) protein (9-12kD), dan N (nucleocapsid) protein (60kD).
1. S (spike) protein (150k)
S protein dapat mengikat asam salisilat (9-O-acetyl neuraminic acid) pada permukaan membrane sel inang dimana hal ini memberi kemampuan virus untuk hemagglutinasi dan merupakan antibodi yang melawan S protein dinetralisasi.
2. HE protein (65kD)
Hanya terdapat pada coronavirus yang mempunyai protein hemagglutinin-esterase. Bentuk protein ini juga seperti paku (lebih kecil dari S protein) pada permukaan virus. Protein ini juga dapat mengikat asam salisilat. Aktivitas esterase dari HE protein dapat memecah asam salisilat dari rantai gula, yang dapa membantu virus untuk masuk dalam sel inang dan bereplikasi. Antibodi yang melawan HE protein juga akan dinetralisasi oleh virus.
3. M (membran) protein
Protein ini membantu perlekatan nukleokapsid ke membran dari struktur internal seperti Badan Golgi dan tidak ditemukan pada membran plasma sel.
4. E (envelope) protein (9-12kD)
Protein kecil ini juga terdapat pada membran virus. Pada sel yang terinfeksi, protein ini ditemukan di sekitar nucleus dan permukaan sel.
5. N (nucleocapsid) protein (60kD)
Nukleokapsid protein mengikat genom RNA didahului dengan beberapa rangkaian dan menuju M protein pada permukaan dalam membrane virus. N protein merupakan protein terfosforilasi. Tidak seperti virus RNA lain, coronavirus tidak bergabung dengan RNA polymerase dalam partikel virus. Polymerase dibuat setelah infeksi dengan menggunakan genom RNA positif sebagai mRNA.
7 respiratory syndrome/coronavirus (MERS-CoV) merupakan sindrom coronavirus yang ditemukan di Saudi Arabia pada tahun 2012 (WHO). MERS-CoV menimbulkan gejala berupa demam, batuk, dan sesak napas. Virus tersebut ditularkan dari hewan (unta) kepada manusia melalui kontak langsung dengan hewan maupun produknya (daging, susu). Populasi berisiko tinggi antara lain lanjut usia, penyandang diabetes melitus. Daging atau susu unta harus dipasteurisasi sebelum dikonsumsi untuk mencegah MERS-CoV.
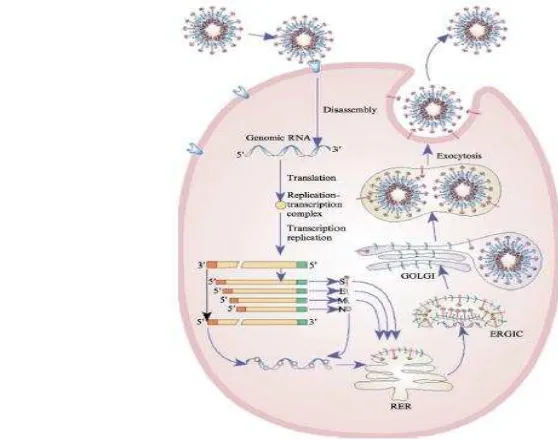
Gambar 4 Siklus hidup coronavirus (Sumber: ncbi.nlm.nih.gov) Replikasi coronavirus diawali dengan proses pelekatan protein S ke reseptor sel inang yang akan menyebabkan fusi dari sel inang dan membran virion. Setelah virion masuk ke dalam sitoplasma, genom dari virus akan ditranslasikan menjadi poliprotein replikase. Replikase ini kemudian akan menggunakan genom cetakan untuk sintesis RNA genom baru dan RNA subgenom melalui bentuk perantara berupa RNA utas negatif. RNA subgenom ditranslasikan menjadi protein struktural sedangkan RNA genom akan melalui proses replikasi yang menghasilkan RNA genom positif. Setelah komponen-komponen virus dibuat, komponen-komponen tersebut disusun menjadi partikel virus baru dan virus akan keluar dari sel inang. Secara singkat, replikasi coronavirus dapat dilihat pada gambar 2 (Masters 2006). Sebagai hasil dari mekanisme yang unik dari replikasi virus, coronavirus memiliki frekuensi rekombinasi yang tinggi. Coronavirus memiliki tingkat rekombinasi dan mutasi yang tinggi memungkinkan untuk beradaptasi dengan host baru (Lai dan Cavanagh 1997).
Kelelawar sebagai reservoar agen penyakit
8
spesies, dan subordo microchiroptera yaitu pemakan serangga, terdiri atas 16 famili, 145 genus dan 788 spesies. Ada sekitar 62 spesies kelelawar, baik jenis dalam ukuran yang relatif besar, maupun sampai pada jenis yang terkecil di dunia, terdapat di Sulawesi (Whitten et al. 1987; Heinrichs et al. 1997). Salah satu spesies kelelawar yang memiliki ukuran tubuh lebih besar, yang hidup tersebar di beberapa habitat hutan hujan tropis di Sulawesi bagian Utara adalah rubah hitam terbang (Black flying fox) Pteropus alecto. Spesies ini pertama kali dijelaskan oleh Temminck (1837) dalam Wilson dan Reeder (2005) berdasarkan spesimen yang diperoleh dari Manado, Sulawesi Utara. Spesies Pteropus alecto (Black flying fox, termasuk dalam famili Pteropodidae (Ten Pas 2004; Myers et al. 2014). Di Sulawesi, suborodo megachiroptera terdapat 11 genus dan 22 spesies (Flanery 1995; Suyanto 2001; Maryanto dan Yani 2003)
Kelelawar merupakan reservoir aneka macam jenis virus termasuk virus yang memiliki potensi zoonosis seperti Nipah, Hendra, Marburg, dan Rabies (Calisher et al. 2006; Sasaki et al. 2012; Schountz 2014 ). Selain virus-virus yang telah disebutkan sebelumnya, diketahui bahwa kelelawar juga merupakan reservoir alami dari coronavirus (Lau et al. 2005; Memish et al. 2013). Kelelawar membawa virus yang menyebabkan kejadian SARS dan MERS pada manusia (Schountz 2014). Kelelawar dapat bertahan terhadap infeksi virus dan menjaga perkembangan virus dalam tubuhnya. Hal ini didukung oleh kemampuannya untuk berhibernasi. Suhu tubuh kelelawar saat hibernasi adalah 8°C sampai dengan 24°C. Suhu dingin dapat mengendalikan viremia yang terjadi dalam tubuh kelelawar sehingga kelelawar tetap bertahan hidup. Akibatnya, kelelawar dapat menularkan virus melalui sekretnya dan individu yang rentan terhadap virus terinfeksi (Calisher et al. 2006).
9
3
METODE
Waktu dan Tempat Penelitian
Penelitian dilaksanakan pada bulan Desember 2015 sampai Mei 2016. Analisis sampel dilakukan di Laboratorium Bioteknologi Pusat Studi Satwa Primata, Lembaga Penelitian dan Pengabdian kepada Masyarakat, Institut Pertanian Bogor (PSSP LPPM-IPB), Bogor.
Bahan
Bahan-bahan yang digunakan antara lain feses kelelawar, buffer AVL,etanol absolut, buffer AW1 AW2, buffer AVE QIAamp® Viral RNA Mini kit (QIAGEN). Transkripsi balik dilakukan dengan SuperScript™ III First-Strand Synthesis System for RT-PCR (Invitrogen), random hexamer, dNTP mix, RT buffer, MgCl2,
DTT, RNAse OUT, dan SuperScript III Reverse-Transcriptase (SS III RT), Kappa HotStart ready mix (Kappa HS), nuclease-free water, gel agarosa.
Alat
Peralatan yang akan digunakan dalam penelitian adalah mesin PCR, mikropipet, mikrotips, freezer, refrigerator, sentrifus, vortex, inkubator, dan BSC (Biosafety cabinet), timbangan digital, spatula steril, GelDoc (Bio-Rad).
Analisis Laboratorium
Ekstraksi Sampel RNA
Sebanyak 151 sampel feses (fecal swab) dari kelelawar diekstraksi menggunakan QIAamp® Viral RNA Mini kit (QIAGEN). Sampel didapat dari arsip milik PREDICT Indonesia-PSSP-IPB. Proses ekstraksi dimulai dengan tahap lisis menggunakan buffer AVL. Tahapan dilanjutkan dengan mengendapkan RNA dengan etanol absolut, RNA dipisahkan dari pengotornya menggunakan buffer AW1 dan AW2 yang disertai dengan proses sentrifugasi menggunakan kolom yang memiliki membran yang dapat mengikat RNA. Tahapan akhir adalah, RNA dielusi dengan menggunakan buffer AVE.
Transkripsi Balik
10
SuperScript III Reverse-Transcriptase (SS III RT). Campuran yang telah dibuat kemudian diinkubasi pada kondisi 25 °C selama 10 menit, 50 °C selama 50 menit, 85 °C selama 5 menit, dan didinginkan pada suhu 4 °C.
Amplifikasi coronavirus
Amplifikasi coronavirus dilakukan dengan metode nested PCR menggunakan empat primer berbeda. Proses amplifikasi dibagi menjadi dua protokol. Protokol pertama menggunakan primer CoV-FWD1 dan CoV-RVS1 yang dilanjutkan dengan proses nested PCR menggunakan primer CoV-FWD2 dan RVS2. Protokol kedua menggunakan primer FWD3 dan CoV-RVS3 yang dilanjutkan dengan proses nested PCR menggunakan primer
CoV-FWD4/Bats dan CoV-RVS3 dengan forward 5’
-GAYTSYCCHAARTGTGAYAGAGC-3’ dan reverse 5’
-CCATCATCASWYRAATCATCATA-3’ yang sejajar dengan kontrol positif yang digunakan (Watanabe et al. 2010).
Visualisasi Hasil Amplifikasi
Visualisasi dilakukan pada gel agarosa 1,8% yang mengandung etidium bromida 10mg/ml sebanyak 10 µl. Elektroforesis dilakukan pada tegangan 100 Volt selama 45 menit di dalam buffer TAE 1×. Setelah proses elektroforesis selesai, hasil yang didapat divisualisasi dengan GelDoc (Bio-Rad).
Analisis Hasil sekuensing
Hasil sekuensing yang didapat akan dianalisis lebih lanjut dengan teknik alignment menggunakan program Basic Local Alignment Search Tool (BLAST), NCBI.
Analisis Data
Data penelitian dianalisis secara deskriptif menggunakan MEGA 6.0. Pohon filogenetik juga dibuat untuk melihat kekerabatan virus dengan menggunakan program BioEdit dan Molecular Evolutionary Genetics Analysis 6.0 (MEGA 6.0) dengan sekuen pembanding berupa RNA-dependent RNA polymerase
11
4
HASIL DAN PEMBAHASAN
Hasil
Sebanyak 151 sampel feses atau usap rektum kelelawar besar pemakan buah (Pteropus alecto) yang dideteksi, ditemukan 65 sampel presumptive positive yang ditunjukkan dengan keberadaan pita sebesar 434 pasang basa (Gb. 1 dan Gb. 2) parsial gen RNA-dependent RNA polymerase (RdRp). Enambelas dari 65 sampel presumptive positive diperiksa lebih lanjut dengan dilakukan sekuensing.
M 11 11 22 22 33 33 97 97
Gambar 5 Visualisasi hasil nested PCR dari 4 sampel fecal swab kelelawar asal Tomohon dan Bersehati menggunakan primer CoV F4/Bat dan CoV RVS3. M: marker Vc 100 bp DNA ladder, lane 3-10: Hasil positif nested PCR
M 37 49 52 55 1 2 3 10 80 84 25 16
Gambar 6 Visualisasi hasil nested PCR dari 12 sampel fecal swab kelelawar asal Tomohon, Bersehati dan hutan bakau Olibuu menggunakan primer CoV F4/Bat dan CoV RVS3. M: marker Vc 100 bp, DNA ladder, lane 3-14: Hasil positif nested PCR
Enambelas sampel positif yang dikonfirmasi sebagai coronavirus kemudian dianalisis lebih lanjut kekerabatannya dengan menggunakan perangkat lunak bioinformatika. Pem-buatan pohon filogenetik menggunakan program Molecular Evolutionary Genetics Analysis 6 (MEGA 6). Program ini dapat digunakan untuk mengestimasi jarak evolusi, menyusun pohon filogenetik, dan menghitung kuantitas statistika dari data molekuler (Kumar et al. 1994).
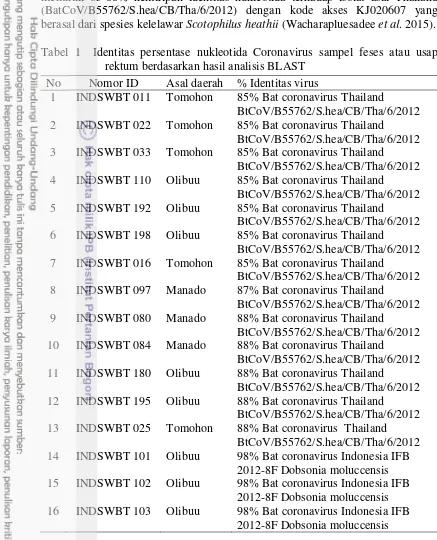
Hasil analisis BLAST menunjukkan bahwa sampel INDSWBT 101, INDSWBT 102, dan INDSWBT 103 memiliki 98% identitas nukleotida terhadap isolat asal Indonesia (BatCoV IFB2012-8F) dengan kode akses AB18719 yang berasal dari spesies kelelawar Dobsonia moluccensis (Anindita et al. 2015). Sedangkan sampel lainnya INDSWBT033, INDSWBT110, INDSWBT192, INDSWBT011, INDSWBT198, INDSWBT016, INDSWBT097, INDSWBT025,
434 bp
12
INDSWBT180, INDSWBT195, INDSWBT080, INDSWBT084, INDSWBT022 memiliki 85-88% kemiripan identitas nukleotida terhadap isolat asal Thailand (BatCoV/B55762/S.hea/CB/Tha/6/2012) dengan kode akses KJ020607 yang berasal dari spesies kelelawar Scotophilus heathii (Wacharapluesadee et al. 2015). Tabel 1 Identitas persentase nukleotida Coronavirus sampel feses atau usap
rektum berdasarkan hasil analisis BLAST No Nomor ID Asal daerah % Identitas virus
1 INDSWBT 011 Tomohon 85% Bat coronavirus Thailand
BtCoV/B55762/S.hea/CB/Tha/6/2012 2 INDSWBT 022 Tomohon 85% Bat coronavirus Thailand
BtCoV/B55762/S.hea/CB/Tha/6/2012 3 INDSWBT 033 Tomohon 85% Bat coronavirus Thailand
BtCoV/B55762/S.hea/CB/Tha/6/2012 4 INDSWBT 110 Olibuu 85% Bat coronavirus Thailand
BtCoV/B55762/S.hea/CB/Tha/6/2012 5 INDSWBT 192 Olibuu 85% Bat coronavirus Thailand
BtCoV/B55762/S.hea/CB/Tha/6/2012 6 INDSWBT 198 Olibuu 85% Bat coronavirus Thailand
BtCoV/B55762/S.hea/CB/Tha/6/2012 7 INDSWBT 016 Tomohon 85% Bat coronavirus Thailand
BtCoV/B55762/S.hea/CB/Tha/6/2012 8 INDSWBT 097 Manado 87% Bat coronavirus Thailand
BtCoV/B55762/S.hea/CB/Tha/6/2012 9 INDSWBT 080 Manado 88% Bat coronavirus Thailand
BtCoV/B55762/S.hea/CB/Tha/6/2012 10 INDSWBT 084 Manado 88% Bat coronavirus Thailand
BtCoV/B55762/S.hea/CB/Tha/6/2012 11 INDSWBT 180 Olibuu 88% Bat coronavirus Thailand
BtCoV/B55762/S.hea/CB/Tha/6/2012 12 INDSWBT 195 Olibuu 88% Bat coronavirus Thailand
BtCoV/B55762/S.hea/CB/Tha/6/2012 13 INDSWBT 025 Tomohon 88% Bat coronavirus Thailand
BtCoV/B55762/S.hea/CB/Tha/6/2012 14 INDSWBT 101 Olibuu 98% Bat coronavirus Indonesia IFB
2012-8F Dobsonia moluccensis 15 INDSWBT 102 Olibuu 98% Bat coronavirus Indonesia IFB
2012-8F Dobsonia moluccensis 16 INDSWBT 103 Olibuu 98% Bat coronavirus Indonesia IFB
2012-8F Dobsonia moluccensis
14
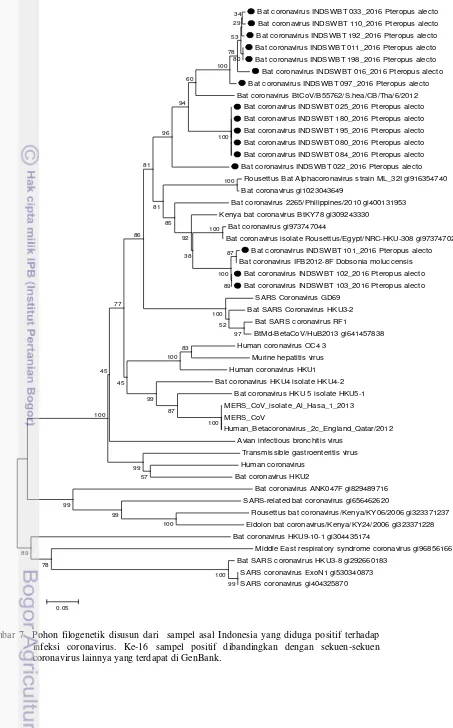
Gambar 7 Pohon filogenetik disusun dari sampel asal Indonesia yang diduga positif terhadap infeksi coronavirus. Ke-16 sampel positif dibandingkan dengan sekuen-sekuen coronavirus lainnya yang terdapat di GenBank.
Bat coronavirus INDSWBT 033_2016 Pteropus alecto
Bat coronavirus INDSWBT 110_2016 Pteropus alecto
Bat coronavirus INDSWBT 192_2016 Pteropus alecto
Bat coronavirus INDSWBT 011_2016 Pteropus alecto
Bat coronavirus INDSWBT 198_2016 Pteropus alecto
Bat coronavirus INDSWBT 016_2016 Pteropus alecto
Bat coronavirus INDSWBT 097_2016 Pteropus alecto
Bat coronavirus BtCoV/B55762/S.hea/CB/Tha/6/2012
Bat coronavirus INDSWBT 025_2016 Pteropus alecto
Bat coronavirus INDSWBT 180_2016 Pteropus alecto
Bat coronavirus INDSWBT 195_2016 Pteropus alecto
Bat coronavirus INDSWBT 080_2016 Pteropus alecto
Bat coronavirus INDSWBT 084_2016 Pteropus alecto
Bat coronavirus INDSWBT 022_2016 Pteropus alecto
Rousettus Bat Alphacoronavirus strain ML_32I gi916354740
Bat coronavirus gi1023043649
Bat coronavirus 2265/Philippines/2010 gi400131953
Kenya bat coronavirus BtKY78 gi309243330
Bat coronavirus gi973747044
Bat coronavirus isolate Rousettus/Egypt/NRC-HKU-308 gi973747026
Bat coronavirus INDSWBT 101_2016 Pteropus alecto
Bat coronavirus IFB2012-8F Dobsonia moluccensis
Bat coronavirus INDSWBT 102_2016 Pteropus alecto
Bat coronavirus INDSWBT 103_2016 Pteropus alecto
SARS Coronavirus GD69
Bat coronavirus HKU4 isolate HKU4-2
Bat coronavirus HKU 5 isolate HKU5-1
MERS_CoV_isolate_Al_Hasa_1_2013
Middle East respiratory syndrome coronavirus gi968561661
Bat SARS coronavirus HKU3-8 gi292660183
15
Pembahasan
Penelitian ini merupakan bagian dari proyek PREDICT Indonesia, dimana pengambilan sampel feses atau usap rektum kelelawar besar pemakan buah dilakukan di tiga daerah di Provinsi Gorontalo (hutan bakau Olibuu) dan Sulawesi Utara: Manado (pasar Bersehati), Tomohon (pasar Beriman). Pengambilan sampel di lapang dilakukan oleh tim PREDICT Indonesia yang diambil secara oportunistik dari pengumpul kelelawar besar pemakan buah di daerah setempat sebelum kelelawar besar pemakan buah didistribusikan untuk memenuhi kebutuhan pasar. Penjualan daging kelelawar besar pemakan buah di pasar tradisional di daerah tersebut tanpa adanya regulasi layaknya pemotongan hewan ternak, sehingga potensi penularan agen penyakit yang dibawa oleh kelelawar yang di transmisikan ke manusia sangat dimungkinkan terinfeksi virus. Namun demikian tidak hanya konsumen yang memiliki resiko terpapar agen petogen tapi juga para penangkap atau pemburu (hunters) kelelawar, pengumpul, keluarga dari pengumpul, orang yang mendistribusikan, serta penjual memiliki resiko atau potensi tertular infeksi coronavirus yang dibawa oleh kelelawar. Cara pemasakan ataupun pengolahan menjadi faktor resiko terinfeksi virus. Dalam hal ini, Malole (1987) mengatakan bahwa sebagian besar virus dapat diinaktifkan dengan suhu 56°C selama 30 menit atau 100°C selama beberapa detik, tetapi sifat virus berbeda tergantung jenisnya sehingga sifat ketahanan terhadap pemanasan, dan pengaruh pH yang berbeda. Semakin tinggi suhu pemanasan, maka peluang untuk rusaknya partikel virus akibat terjadinya denaturasi protein semakin besar. Pemanasan suhu tertentu mempunyai pengaruh terhadap struktur protein virus sehingga terjadi denaturasi protein yang menyebabkan rusaknya partikel virus dan akan menghilang inefektifitas virus atau menyebabkan virus tersebut inaktif.
Hasil penelitian menunjukkan 65 sampel dari 151 sampel (43,04%) Pteropus alecto spesimen feses atau usap rektum adalah presumptive positive coronavirus. Sementara itu, 16 dari 65 sampel presumptive positive coronavirus dikonfirmasi postif coronavirus (24,6%), selebihnya masih belum dilakukan sekuensing karena keterbatasan dana penelitian. Penelitian menggunakan hewan coba dapat dilakukan untuk mengidentifikasi apakah virus-virus tersebut berbahaya atau tidak (Clay et al. 2014). Data ini konsisten dengan hasil kelelawar CoV deteksi dari sampel feses atau jaringan usus yang dilaporkan sebelumnya (Anindita et al. 2015; Wacharapluesadee et al. 2015 Watanabe at al 2010) yang menyatakan bahwa coronavirus dapat ditemukan di berbagai spesies kelelawar. Spesies kelelawar sebagai sumber sampel yang diambil dari Provinsi Gorontalo (hutan bakau Olibuu) dan Sulawesi Utara: Manado (pasar Bersehati), Tomohon (pasar Beriman) telah diidentifikasi adalah merupakan spesies kelelawar besar pemakan buah Pteropus alecto (black flying fox). Sementara itu, isolat virus yang terkait erat dengan CoV yang ditemukan di Indonesia adalah spesies kelelawar Dobsonia moluccensis dan spesies kelelawar Scotophilus heathii dari Thailand. Data menunjukkan bahwa CoV yang identik secara genetic ditemukan di spesies kelelawar yang berbeda. Data ini juga didukung penelitian sebelumnya tentang keragaman CoV yang dideteksi pada spesies kelelawar yang berbeda (Wacharapluesadee et al. 2015).
16
conserved pada semua CoV (Xu et al. 2003). Hasil BLAST dan hasil pohon filogenetik dari 16 sampel positif CoV terdapat dua kelompok CoV yang terdeteksi berasal dari daerah Olibuu (Gorontalo) dengan spesies kelelawae besar pemakan buah adalah Pteropus alecto. Tiga sampel (INDSWBT101, INDSWBT 102, dan INDSWBT103) berkerabat dengan CoV yang dilaporkan sebelumnya dari Paguyaman, Indonesia dengan identitas nukleotida 98%. Desa Olibuu pada kecamatan Paguyaman pantai sebagai sumber sampel pada penelitian ini dan kecamatan Paguyaman sebagai sumber sampel yang digunakan pada penelitian Anindita et al. 2015 sebagai salah satu acuan referensi merupakan wilayah di dalam kabupaten yang sama di Provinsi Gorontalo Indonesia. Tigabelas spesimen INDSWBT033, INDSWBT110, INDSWBT192, INDSWBT011, INDSWBT198, INDSWBT016, INDSWBT097, INDSWBT025, INDSWBT180, INDSWBT195, INDSWBT080, INDSWBT084, INDSWBT022 memisah dalam kelompok berbeda (cluster) baru karena memiliki persentase kesamaan yang rendah (85%) dengan data yang ada di GenBank, meskipun masih berkelompok dengan CoV dari Thailand. Karakterisasi lebih lanjut untuk gen RdRp keseluruhan atau wilayah gen lain dari CoV ini diperlukan untuk membuktikan asumsi ini. Secara umum, CoV pada kelelawar yang diambil dari tiga daerah di Provinsi Gorontalo (hutan bakau Olibuu) dan Sulawesi Utara: Manado (pasar Bersehati), Tomohon (pasar Beriman) secara genetik masih memiliki kekerabatan dengan CoV yang ditemukan di beberapa daerah di dunia (Kenya, China, Thailand, Filipina, Timur Tengah, dan Madagaskar).
Studi ini menunjukkan bahwa kelelawar besar pemakan buah dari Indonesia merupakan reservoar untuk virus yang memiliki potensi zoonosis, karena kelelawar besar pemakan buah juga dikonsumsi oleh masyarakat di beberapa daerah di Indonesia, sehingga temuan ini penting sebagai informasi untuk kebijakan terkait. Meskipun CoV tidak menyebabkan penyakit pada kelelawar dan kekerabatan dari temuan filogenetik dari virus ini dalam penelitian ini cukup jarak jauh dengan SARS dan MERS-CoV, namun perlu kewaspadaan tentang dampak dari CoV untuk kesehatan manusia dan bisa menjadi peringatan dini. Data ini dapat disajikan sebagai data tambahan dan informasi monitoring global dari munculnya kelelawar CoV yang berpotensi menjadi patogen berbahaya bagi kesehatan manusia. Ancaman penyakit pandemik muncul diperantarai oleh interaksi dari satwa liar dan manusia.
Coronavirus diketahui menyebabkan infeksi pada manusia dan hewan, dengan berbagai macam gejala mulai dari gangguan saluran pernafasan hingga terjadinya kematian (Christian et al. 2010; Anindita et al. 2015). Selain saluran pernafasan, coronavirus juga menyebabkan infeksi pada organ tubuh termasuk saluran pencernaan, liver, ginjal (Lelli et al. 2013). Pengambilan jumlah sampel di provinsi Gorontalo dan Sulawesi Utara akan lebih representatif apabila jumlah sampel ditambah untuk penelitian sejenis untuk mewakili data persebaran virus yang dibawa oleh kelelawar. Dengan adanya mutasi dan rekombinasi pada Coronavirus juga menjadi suatu kewaspadaan sebagai early warning untuk masyarakat luas sehingga surveilans perlu terus dilakukan untuk meningkatkan kesehatan hewan, manusia, dan lingkungan sebagai upaya kolaborasi dari One Health.
17 pathogen. Gangguan habitat, seperti pembukaan hutan untuk tujuan pertanian atau pembangunan, transportasi, penjualan dan konsumsi satwa liar, memberikan peluang kemungkinan berpindahnya patogen dari satwa liar ke manusia. Pengembangan sistem peringatan dini (early warning), termasuk upaya pengawasan dan kapasitas diagnostik untuk potensi ancaman penyakit ke manusia, sangat dibutuhkan oleh instansi pemerintah, kegiatan penelitian, dan lembaga akademis dalam rangka untuk lebih melayani dan melindungi masyarakat. Data ini dapat berfungsi sebagai data tambahan dan informasi pengawasan global dari munculnya coronavirus yang berpotensi sebagai patogen berbahaya untuk kesehatan manusia.
5
SIMPULAN DAN SARAN
Simpulan
Enampuluh lima dari 151 sampel diketahui presumptive positive coronavirus. Enambelas dari 65 sampel ditemukan positif terhadap infeksi dari coronavirus. Tiga dari 16 sampel positif coronavirus memiliki identitas persentase tertinggi 98% terhadap BatCoV isolat asal Indonesia IFB2012_8F dengan spesies kelelawar Dobsonia moluccensis dan 85-88% terhadap Bat coronavirus asal Thailand isolat BtCoV/B55762/S.hea/CB/Tha/6/2012 dengan spesies kelelawar Scotophilus heathii.
Saran
18
DAFTAR PUSTAKA
Anindita PD, Sasaki M, Setiyono Agus, Handharyani E, Orba Yasuko, Kobayashi S, Rahmadani Ibnu, Taha S, Adiani S, Subangkit M, et al. 2015. Detection of coronavirus genomes in Moluccan naked-Backed fruit bats in Indonesia. Arc Virol. doi: 10.1007/s00705-015-2342-1.
Altringham JD. 1996. BATS. Biologi and Behaviour. Oxford University Press. New York
Barrett MA, Osofsky SA. 2013. One Health: Interdependence of People, Other Species, and the Planet. In Katz DL, Elmore JG, Wild DMG, Lucan SC,editors. Jekel’s Epidemiology, Biostatistics. Preventive Medicine, and Public Health (4th ed.). Philadelphia: Elsevier/ Saunders; 2013. pp. 364-377 and online supplement pp. 407-416.
Breed AC, Meng Y, Barr JA, Crameri G, Thalmann CM, Wang LF. 2010. Prevalence of henipavirus and rubulavirus antibodies in pteropid bats, Papua New Guinea. Emerg Infect Dis.;16:1997–9.doi: 10.3201/eid1612.100879 Calisher CH, Childs JE, Field HE, Holmes KV, Schountz T. 2006. Bats important
reservoirs hosts of emerging viruses. Clin microbiol. Rev 19:531-545. [CDC] Centers for Disease Control and Prevention. 2015. Severe Acute
Respiratory Syndrome. [Internet]. [diunduh 2015 Desember 23]. Tersedia pada: http://www.cdc.gov/coronavirus/mers/downloads/factsheet-mers_en.pdf.
Christian KA, Ijaz K, Dowell SF, Chow CC, Chitale RA, Bresee JS, Mintz E, Pallansch MA, Wassilak S, McCray E et al. 2013.What we are watching five top global infectious disease threats, 2012: a perspective from CDC’s Global Disease Detection Operation Center. J Emerg Health Threats. 6(20632):1-8.
Clay CC, Donart N, Fomukong N, Knight JB, Overheim K, Tipper J, Westrienen JV, Hahn F, Harrod KS. 2014. Severe acute respiratory syndrome-coronavirus infection in aged nonhuman primates is associated with modulated pulmonary and systemic immune responses. Immunity & Ageing. 11:4. doi: 10.1186/1742-4933-11-4.
Corbet GB, Hill JE. 1992. The mammals of the Indomalayan Region : A systematic review. Natural history museum publication & Oxford University Press.
De Haan CAM, Kuo L, Masters PS, Vennema H, Rottier PJM. 1998. Coronavirus particle assembly: primary structure requirements of the membrane protein. J Virol 8(72): 6838-6850.
Flannery T. 1995. Mammals of the Soth-Wes Pacific & Moluccan Island. Sydney. Australian Museum/ReedBook.
Drexler JF, Corman VM, Drosten C. 2014. Ecology, evolution and classification of bat coronaviruses in the aftermath of SARS. Antiviral Res. 101:45-56. Gouilh MA, Puechmaille SJ, Gonzales JP, Teehng E, Kittayapong P, Manuguerra
19 Heinrichs S, Zahnke K. 1997. The fruit bats of Sulawesi. BATS Magazine Media and Education. 15(3). [Internet]. [diunduh 2015 Desember 23]. Tersedia pada: http://www.batcon.org/resources/media-education/bats-magazine/bat . Kumar S, Tamura K, Nei M. 1994. MEGA: Molecular Evolutioary Genetics Analysis software for microcomputers. Comput Appl Biosci 10(2):189-191. Lai MM, Cavanagh D. 1997. The molecular biology of coronaviruses Res 48:1-10 Lau SK, Woo PC, Li KS, Huang Y, Tsoi HW, Wong BH, Wong SS, Leung SY, Chan KH, Yuen KY. 2005. Severe acute respiratory syndrome coronavirus-like virus in Chinese horseshoe bats. Proc Natl Acad Sci U S A 102(39): 14040-5.
Lekagul B, McNeely JA. 1977. Mammals of Thailand. Sahakarnbhat. Bangkok. Lelli D, Papetti A, Sabelli C, Rosti E, Moreno A, Boniotti MB. 2013. Detection of
coronavirus in bats of various species in Italy. Viruses. 5:2679-2689.
Mahon CR, Lehman DC, Manuselis Jr. G. 2014. Textbook of Diagnostic Microbiology. Ed ke-5. Maryland Heights (US): Elsevier.
Malole B dan Martin. 1987. Virologi Pusat Antar Universitas. Institut Pertanian Bogor. Lembaga Sumberdaya Informasi IPB.
Maryanto I, Yani M. 2003. A new species of Rousettus (Chiroptera; Pteropodidae) from Lore Lindu, Central Sulawesi. Mammal Study 28;111-120
Masters PS. 2006. The Molecular Biology of Coronaviruses. Di dalam: Maramorosch K, Shatkin AJ, editor. Advances in Virus Research. San Diego (US): Academic Pr.
Memish ZA et al. 2013. Middle east respiratory syndrome coronavirus in bats, 8Saudi Arabia. Emerg Infect Dis. 11(19):1819-1823.
Myers P, R Espinosa, CS Parr, T Jones, GS Hammond, and TA. Dewey. 2014. The Animal Diversity. [internet] [diunduh pada 19 April 2016] Tersedia pada: http://amimaldiversity.org.
Navas-Martin S, Weiss SR. 2004. Coronavirus replication and pathogenesis: Implications for the recent outbreak of severe acute respiratory syndrome (SARS), and the challenge for vaccine development. J Neurovirol 10:75-85. DOI: 10.1080/13550280490280292.
Nowak RM. 1999. Walker’s bats of the word. The Johns Hopkins University Press. Australia. Ed.5( 2): 1629.
Poon L, Guan Y, Nicholls J, Yuen K, Peiris J, 2004. The aetiology, origins, and diagnosis of severe acute respiratory syndrome. Lancet Infect Dis. 4: 663– 671.
Sasaki M, Setiyono A, Handharyani E, Rahmadani I, Taha S, Adiani S, Subangkit M, Sawa H, Nakamura I, Kimura T. 2012. Molecular detection of a novel paramyxovirus in fruit bats from Indonesia. J Virol 9: 240.
Schountz T. 2014. Immunology of bats and their viruses: Challenges and opportunities. Viruses 6: 4880-4901.
Sompayrac L. 2012. How Pathogenic Virus Think. Ed ke-2. Burlington (US): Jones & Bartlett.
Suyanto A. 2001. Kelelawar di Indonesia. Seri Panduan Lapangan. Bogor. Pusat Penelitian dan Pengembangan Biologi LIPI.
20
Smith EC, Denison MR. 2013. Coronavirus as DNA wannabes: a new model for the regulation of RNA virus replication fidelity. Plos Pathogens. 9(12). doi:10.1371/journal.ppat.1003760.
Wacharapluesadee S, Duengkae P, Rodpan A, Kaewpom T, Maneeom P, Kanchanasaka B, Yingsakmongkon S, Sittidetboripat N, Chareesaen C, Khlangsap N. 2015. Diversity of coronavirus in bats from eastern Thailand. J Virol, doi: 10.1186/s12985-015-0289-1.
Watanabe S, Masangkay JS, Nagata N, Morikawa S, Mizutani T, Fukushi S, Alviola P, Omatsu T, Ueda N, Iha K et al. 2010. Bat Coronaviruses and Experimental Infection of Bats, the Philippines. Emerg Infect Dis. 16(8):1217-1223.
Whitten AJ, Henderson GS, Mustafa M. 1987. Ekologi Sulawesi. Yogyakarta: Gadjah Mada University Press.
Wilson DE and DeeAnn M Reeder (Editors). 2005. Mammal Species of the World. A Taxonomic and Geographic Reference 3rd ed, Johns Hopkins University Press. [internet] [diunduh pada 23 Desember 2015] Tersedia pada:
http://www.departments.bucknell.ed.u/biology/resources/msw3/.
Xu X, Liu Y, Weiss S, Arnold E, Sarafianos SG, Ding J. 2003. Molecular model of SARS coronavirus polymerase: implications for biochemical function and drug design. Nucleic acids res 31:7117-7130.
21
23 Lampiran 1 Pensejajaran sekuens Bat coronavirus INDSWBT regio RdRp isolat
26
>CoV4Bats#033
27
RIWAYAT HIDUP
Penulis dilahirkan di Magetan, pada tanggal 14 Februari 1992, sebagai anak bungsu dari pasangan Tawar Hadi Wiyono dan Nasiyah. Pendidikan sarjana ditempuh di Program Studi Pendidikan Biologi, Fakultas Keguruan dan Ilmu Pendidikan Universitas Jember, lulus pada tahun 2014. Pada tahun 2014 penulis diterima di Program Studi Mikrobiologi Medik pada Program Pascasarjana Institut Pertanian Bogor dan menamatkannya pada tahun 2016.