IMUNOMODULATOR DAN ANTITUMOR INFEKSI Marek’s
Disease Virus (MDV) SEROTIPA 1 ONKOGENIK PADA AYAM
MUHAMAD SAMSI
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI DISERTASI
Saya menyatakan dengan sebenar-benarnya bahwa disertasi saya dengan
judul : Ekstrak Benalu Teh (Scurrula oortiana) Sebagai Imunomodulator dan Antitumor pada Infeksi Marek’s disease virus (MDV) Sertipa 1 Onkogenik pada Ayam adalah benar-benar asli karya saya dengan arahan komisi pembimbing, dan bukan hasil jiplakan atau tiruan dari tulisan siapa pun serta belum diajukan dalam
bentuk apa pun kepada perguruan tinggi mana pun.
Bogor, Agustus 2007.
Muhamad Samsi NIM. B161030011
MUHAMAD SAMSI. The Tea Parasite (Scurrula oortiana) Extract as Immunomodulator and Antitumor on the Infection of Marek’s Disease Virus (MDV)
Serotype 1 Oncogenic in Chicken. Under supervision of MARTHEN B.M. MALOLE,
WASMEN MANALU, and EKOWATI HANDHARYANI.
Marek’s disease virus (MDV) is one of oncogenic herpesvirus which has a DNA as nucleic acid. It causes immunosupresion and cancer in chicken. This study was aimed to find out the mechanism of Marek’s disease in layer commercial chickens which administered orally with extract of tea parasite (Scurrula oortiana) in dose of10 mg/kg bw through drinking water, then the chickens were infected by intraperitoneal oncogenic MDV in dose 1,0 x103 TCID50. The study used 60 layer
commercial day old chicks (DOC) divided into four group of treatments. The
treatments were group A (administered S. oortiana extract and without MDV
infection), B (neither S. oortiana nor MDV infection), C(administered S. oortiana
extract and whith MDV infection), and D (none administered S. oortiana extract, but whith MDV infection). This study was conducted for 60 days.
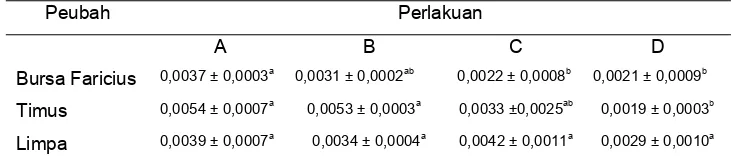
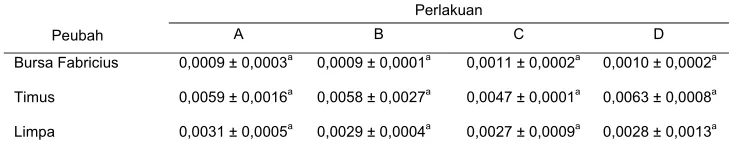
The analysis showed that MDV oncogenic caused immunosupresion at day
post infection (p.i) and recovery to be normal based on relative weight of bursa of Fabricius and thymus, and also diameter of the bursa of Fabricius follicle at 40 of post infection. Moreover, the MDV caused cancer at day 20 of post infection. and increased pathogensity based on the amount of the proventriculus limphocyte, and pathogenesis of liver cancers at day 40 of post infection. The extract of S. oortiana
had a capability as an immunomodulator as indicated by the increase of relative weight of bursa of Fabricius and thymus at day 20 of post infection. and the increase of diameter of bursa of Fabricius follicle at day 40 of post infection.
. Its effect on nonspesific immunity was indicated with the increase of inducible nitric oxyde shynthase (iNOS) enzyme at 20 of day p.i. Its effect on the humoral immunity was indicated with the increase of antibody titre against MDV at day 20 of post infection. The special property of S. oortiana extract as antivirus was indicated by the inhibition the MDV development on the bursa of Fabricius at day 20 of post infection. The extract also decrease the amount of lymphocyte of submucous proventriculus and liver pathogenesis at day 40 of post infection.
RINGKASAN
MUHAMAD SAMSI. Ekstrak Benalu Teh (Scurrula oortiana) Sebagai
Imunomodulator dan Antitumor pada Infeksi Marek’s disease virus(MDV)Onkogenik Serotipe 1 pada Ayam. Dibimbing oleh MARTHEN B.M. MALOLE, WASMEN MANALU, dan EKOWATI HANDHARYANI.
Ayam dalam kondisi normal memproduksi radikal bebas (prooksidan) sebagai proses fisiologis yang seimbang dengan antioksidan endogen yang tersedia. Infeksi Marek’s disease virus(MDV) onkogenik pada ayam diawali sitolisis pada limfosit B dan limfosit T, ayam memberikan responss imun yang didahului oleh responss imun nonspesifik, yaitu fagositosis oleh mekrofag dan neutrofil yang menghasilkan bahan penghancur mikroorganisme patogen berupa peningkatan produksi radikal bebas yang memiliki efek samping, yaitu kerusakan molekul-molekul pada sel sehingga menimbulkan sitolisis termasuk pada limfosit B dan limfosit T. Radikal bebas merupakan bahan karsinogen yang menimbulkan mutasi gen sehingga dapat menginduksi terjadinya kanker. Virus penyebab tumor disebut virus onkogen dan gen yang ada pada virus disebut viral oncogen (V-onc) yang homolog dengan sekuen DNA pada gen seluler inang, yaitu protooncogen (C-onc) yang dapat berinteraksi dengan gen virus. Terjadinya transformasi pada gen seluler inang oleh gen virus bergantung pada resistensi seluler inang, virulensi virus penyebab, dan kehadiran substansi kimia penyebab tumor, yaitu bahan karsinogen yang menginduksi terjadinya mutasi.
Tujuan umum penelitian ini adalah untuk mempelajari pengaruh ekstrak benalu teh Scurrula oortiana terhadap fenomena imunologis dan risiko kanker pada ayam yang diinfeksi virusherpes MDV onkogenik, sedangkan secara khusus tujuan penelitian ini adalah: pembuktian secara ilmiah khasiat eksstrak S. oortiana sebagai imunomodulator dan mengurangi risiko kanker, menggunakan parameter imunologi. Menjadikan benalu teh S. oortiana sebagai obat herbal berstandar.
Tahap pertama uji penentuan dosis infeksi MDV onkogenik, digunakan 20 ekor ayam dibagi ke dalam lima kelompok perlakuan masing-masing empat ekor yaitu : A. kontrol tanpa infeksi, B. diinfeksi intraperitoneal dengan dosis 0,125 x 1000 EID50, C. 0,250 x 1000 EID50, D 0,500 x 1000 EID50, dan E 1 x 1000 EID50.
Sedangkan tahap kedua digunakan 60 ekor ayam dibagi ke dalam empat kelompok perlakuan yaitu : perlakuan A. diberi ekstrak S. oortiana dengan, tanpa infeksi MDV, B tanpa pemberian ekstrak S. oortiana dan tanpa infeksi MDV, C diberi ekstrak S. oortiana dan diinfeksi MDV, dan D tanpa diberi ekstrak S.oortiana, diinfeksi MDV. Ekstrak benalu teh diberikan secara oral (dicekok) sejak ayam berumur 15 hari sampai akhir percobaan, dengan dosis 10 mg/kg bobot badan yang dilarutkan dalam air minum. Ayam diinfeksi dengan virus Marek pada umur 20 hari secara intraperitoneal dengan dosis 1.000 EID50. Parameter yang diamati : Perhitungan
dengan paningkatan rataan bobot relatif bursa Fabricius, bobot relatif timus, dan diameter folikel bursa Fabricius. Hasil tersebut tercermin dari meningkatnya rataan bobot realatif bursa Fabricius pada kelompok ayam yang diberi ekstrak S. oortiana
dibanding kelompok yang diberi ekstrak dikombinasi dengan infeksi MDV maupun keleompok ayam yang hanya diinfeksi MDV 20 hari p.i. Bobot relalatif timus pada kelompok ayam yang diberi ektrak S. oortiana dan diinfeksi MDV tidak mengalami
penurunan pada 20 hari pascainfeksi, hal ini menunjukkan bahwa ekstrak S.
oortiana mampu menghambat imunosupresi akibat infeksi MDV. Pada pengamatan ini terjadi peningkatan rataan diameter folikel bursa Fabricius 40 hari pascainfeksi pada kelompok ayam perlakuan kombinasi ekstrak S. oortiana dibanding dengan semua kelompok perlakuan.
Keberadaan iNOS berdasarkan reaksi positif dengan pewarnaan imunohistokimia pada jaringan hati diduga terkait dengan aktivitas sel-sel dalam hati yang diekspresikan oleh sel-sel Kupffer maupun sel endotel. Kelompok ayam yang
diberi ekstrak S. oortiana tanpa infeksi MDV (A) mengalami peningkatan
pembentukan iNOS yang lebih banyak jika dibandingkan dengan kelompok ayam tanpa diberi ekstrak S. oortiana tanpa infeksi MDV (B). Infeksi MDV mampu meningkatkan jumlah sel yang menghasilkan iNOS baik pada kelompok yang diberi ekstrak maupun tanpa diberi ekstrak S. oortiana dibanding dengan kelompok ayam tanpa infeksi. Hal ini menunjukkan bahwa infeksi MDV mampu meningkatkan produksi iNOS sebagai bagian dari responss imun nonspesifik.
Meningkatnya titer antibodi terhadap MDV pada 20 hari kelompok perlakuan pemberian benalu teh dan infeksi MDV (C) lebih tinggi daripada kelompok tanpa pemberian benalu teh dan tanpa infeksi MDV (B). Hal ini disebabkan adanya
kombinasi pengaruh imunomodulasi ekstrak S. oortiana dengan faktor
imunostimulasi sebagai respons imun akibat tindakan uji tantang. Pemeriksaan imunohistokimia keberadaan MDV pada bursa Fabricius menunjukkan terjadi penurunan pada kelompok ayam yang diinfeksi dengan MDV dan diberi ekstrak S. oortiana.
Pemberian ekstrak S. oortiana pada ayam yang diuji tantang dengan MDV onkogenik mampu menurunkan risiko kanker yang ditandai persentase limfosit pada leukosit 20 hari p.i dan menurunnya jumlah limfosit submukosa proventrikulus 40 hari p.i pada kelompok ayam yang diberi benalu teh dan diinfeksi. Perubahan histopatologi pada 40 hari p.i yang terjadi akibat infeksi MDV adalah infiltrasi sel-sel limfoid dan makrofag pada organ hati. Kelompok ayam yang diberi ekstrak benalu teh dan diinfeksi MDV ternyata mampu menekan pertumbuhan sel-sel limfoid atau sel tumor, yang ditunjukkan dengan jumlah sel limfoid yang lebih sedikit dibandingkan dengan hati pada kelompok ayam tanpa diberi ekstrak benalu teh dan diinfeksi MDV.
© Hak cipta milik Institut Pertanian Bogor, tahun 2007
Hak cipta dilindungi
IMUNOMODULATOR DAN ANTITUMOR INFEKSI Marek’s
disease virus (MDV) SEOTIPE 1 ONKOGENIK PADA AYAM
MUHAMAD SAMSI
Disertasi
Sebagai salah satu syarat untuk memperoleh gelar
Doktor pada
Program Studi Sains Veteriner
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Penguji Luar Komisi
Pada Ujian Tertutup : Drh. Tutik Wresdiyati, Ph.D
Pada Ujian Terbuka : Dr. drh. Retno D. Soejoedono, MS Dr. Ir. Mas Yedi Sumaryadi, MS.
Judul Disertasi
Nama NIM
: Ekstrak Benalu Teh (Scurrula oortiana) Sebagai Imunomodulator dan Antitumor pada Infeksi Marek’s Disease virus (MDV) Onkogenik Serotipe 1 pada Ayam
: Muhamad Samsi : B161030011
Disetujui
Komisi Pembimbing
Dr. Drh. Marthen B.M. Malole Ketua
Prof. Ir. Wasmen Manalu. Ph.D. Drh. Ekowati Handharyani MS. Ph.D. Anggota Anggota
Diketahui
Ketua Program Studi Sains Veteriner Dekan Sekolah Pascasarjana
Dr. drh. B. Ponco Priyosoeryanto MS. Prof. Dr.Ir. Khairil Anwar Notodiputro, MS.
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah SWT., atas segala karunia dan rahmat-Nya sehingga penulis dapat menyelesaikan penelitian sampai dengan penyusunan disertasi, dengan judul “Ekstrak Benalu Teh (Scurrula oortiana) Sebagai Imunomodulator dan Antitumor pada Infeksi Marek’s disease virus Onkogenik pada Ayam”
Ucapan terima kasih yang tulus dan penghargaan yang setinggi-tingginya penulis sampaikan kepada yang terhormat Dr.Drh. Marthen B.M. Malole, sebagai ketua komisi pembimbing, Prof.Ir. Wasmen Manalu, Ph.D. dan Drh. Ekowati Handaryani, MSi. Ph.D. masing-masing sebagai anggota komisi pembimbing yang telah menyediakan waktu, dan dengan penuh kesabaran serta keikhlasan dalam proses pembimbingan selama penulis menempuh pendidikan S3.
Ucapan terima kasih penulis juga disampaikan kepada Dekan Fakultas Peternakan Universitas Jenderal Soedirman, Rektor Universitas Jenderal Soedirman, dan pengelola beasiswa Program Pascasarjana (BPPS) Direktorat Jenderal Pendidikan Tinggi, Departemen Pendidikan Nasional yang telah memberikan kesempatan belajar dan bantuan biaya pendidikan dan penelitian kepada penulis. Penulis juga mengucapkan terimakasih kepada Direktur PPSDM-IPB dan Yayayasan Dana Sejahtera Mandiri Jakarta yang telah mamberikan bantuan biaya penulisan disertasi kepada penulis. Ucapan terimakasih kepada Dekan Sekolah Pascasarjana, Institut Pertanian Bogor beserta staf dan pegawai dan Ketua Program Studi Sains Veteriner (SVT) beserta staf dan pegawai atas kelancaran proses penyelenggaraan pendidikan, serta kepada semua pihak yang telah terlibat dalam membantu penyelesaian studi.
Penghargaan penulis disampaikan kepada Dr Retno Murwani, Dr. Drh. Marthen B.M. Malole, Dr. Drh. Sri Murtini, MSi, Dr. Drh. Fajar Satrija, MSc. atas bantuan materi penelitian awal, pak Nur, drh Farida, drh. Bongot, drh. Tanti, Kristina, Teti, pak Kasnadi, pak Endang, Anin, dan Elia atas bantuan dan kerjasamanya selama kerja dilaboratorium. Disampaing itu penulis juga menyampaikan penghargaan dan terimakasih kepada sataf dan pegawai di Laboratorium Terpadu, Laboratorium Virologi, Laboratorium Patologi, dan Laboratorium Fisiologi Fakultas Kedoteran Hewan IPB.
Rasa haru dan terima kasih yang tulus disampaikan kepada seluruh keluarga atas bantuan dan dukungan moril maupun materil kepada penulis, serta kepada istri tercinta Siti Elistjanti, S.Sos., ananda tersayang Dhaifina Asmarani (Fina), atas segala do’a, pengertian, kesabaran, dorongan semangat dan kasih sayang yang diberikan selama mendampingi penulis dalam menyelesaikan pendidikan S3.
Akhirnya, semoga karya disertasi ini dapat bermanfaat di masyarakat dan pengembangan ilmu pengetahuan, dan dapat dijadikan sebagai bahan acuan khususnya dalam bidang kesehatan dan peternakan.
.
Bogor, Agustus 2007.
Penulis dilahirkan di Banjarnegara pada tanggal 7 Oktober 1957, sebagai anak ke lima dari tujuh bersaudara dari pasangan Bapak Munidi Djogosukarto (Alm.) dan Ibu Hj. Salbingah. Pada tahun 1987 menikah dengan Siti Elistiyanti S.Sos, dan dikaruniai seorang anak, yakni Dhaifina Asmarani (Fina).
Pendidikan sarjana telah ditempuh di Fakultas Kedokteran Hewan, Universitas Gadjahmada Yogyakarta, lulus sarjana Kedokteran Hewan tahun 1982, lulus dokter hewan pada tahun 1983. Penulis menamatkan Magister Sains di Program Studi Sains Veteriner Program Pascasarjana Universitas Gadjahmada Yogyakarta tahun 1999. Kesempatan untuk melanjutkan ke program doktor pada Program Studi Sains Veteriner Sekolah Pascasarjana IPB pada tahun 2003. Beasiswa pendidikan pascasarjana diperoleh dari BPPS DIKTI Departemen Pendidikan Nasional.
Pada tahun 1985 sampai dengan tahun 1987 penulis berkesempatan bekerja di PT.Bamaindo Feeds Stuft Surabaya. Sejak tahun 1987 sampai dengan sekarang penulis adalah sebagai staf pengajar di Fakultas Peternakan Universitas Jenderal Soedirman Purwokerto dengan bidang kajian yang diminati Ilmu Kesehatan Ternak.
DAFTAR ISI
Halaman
DAFTAR TABEL . . . .. . .
DAFTAR GAMBAR . . . . .. . .
DAFTAR LAMPIRAN . . . .. . .
PENDAHULUAN . . . Latar Belakang . . . . Kerangka Pemikiran ……….. Tujuan Penelitian . . . .. . . . . . Manfaat Penelitian . . .
TINJAUAN PUSTAKA . . . Imunitas Tubuh . . . Kanker . . . . . . Penyakit Marek . . . . . . Radikal Bebas . . . Antioksidan . . . . . Benalu Teh sebagai Penurun Risiko Kanker . . .
BAHAN DAN METODE. . . . . .. . . Tempat dan Waktu Penelitian . . . Bahan dan Alat . . . . . . . . . Desain Penelitian . . . . . . Metode Penelitian . . .
HASIL DAN PEMBAHASAN . . . Penentuan Uji Dosis Infeksi MDV . . . ……….. Bobot Relatif Organ Bursa Fabricius, Timus, dan Limpa . . .
Ukuran Diameter Folikel Bursa Fabricius . . . Pemeriksaan Imunohistokimia Enzim Indusible Nitric Oxyd
Synthase (iNOS) . . . . ……… Uji Tingkat Imunitas pada MDV dengan Metode ELISA . . . Pengaruh Ekstrak S. oortiana pada Total Leukosit dan
Persentase Limfosit . . . . . . Pengaruh Ekstrak S. oortiana pada Keberadan MDV pada
Bursa Fabricius . . . Limfosit pada proventrikulus ……… Pengaruh Ekstrak S. oortiana pada Patogenesis Marek pada Hati ..
xii xiii xvi 1 1 4 6 6 7 7 9 12 17 20 25 28 28 28 39 30 37 37 39 42 44 46 49 51 52 54
PEMBAHASAN UMUM ……….
SIMPULAN . . . .. . . …...
DAFTAR PUSTAKA .. . . . . . .. . .
LAMPIRAN . . . . . . .. . . 56
63
64
DAFTAR TABEL
Halaman
1.
2.
3.
4.
5.
6.
7.
8.
9.
Rataan bobot relatif bursa Fabricius, timus, dan limpa
20 hari p.i. . . . . .
Rataan bobot relatif bursa Fabricius, timus, dan limpa
40 hari p.i. . .
Rataan diameter folikel bursa Fabricius (µm) ayam 20 dan 40 Hari pascainfeksi (p.i.) . . .
Rataan jumlah reaksi positif iNOS pada hati ayam 20 hari Pascainfeksi (p.i.) . . . .. . . . . .
Rataan nilai absorbansi uji ELISA berdasarkan perbedaan Perlakuan . . .
Rataan nilai absorbansi berdasarkan waktu pascainfeksi (p.i.) MDV . . .
Rataan jumlah leukosit per mililiter dan persentase limfosit(%) Pada ayam 20 hari pascainfeksi . . . .. . . ..
Rataan jumlah leukosit per mililiter dan proporsi limfosit (%) Pada ayam 40 pascainfeksi (p.i) . . . . . .
Rataan jumlah limfosit submukosa proventriculus 20 hari dan 40 hari pascainfeksi (p.i) . . . . . .
40
42
43
46
47
49
50
50
xiii
DAFTAR GAMBAR
Halaman
1.
2.
3.
4.
5.
6.
7.
8.
9.
10.
11.
12.
Hubungan keterkaitan MDV, ayam sebagai inang, dan
eksktrak benalu teh sebagai antioksidan eksogen . . . ….
Mekanisme secara umum sistem imun . . . . . . . ….
Kemungkinan skenario pada ketidakseimbangan ROS. . . …..
Senyawa flavonoid . . . . . . ……
Senyawa fenol . . . .. . .. . . ….
Diagram alir penelitian . . . . .. . .. . . ……….
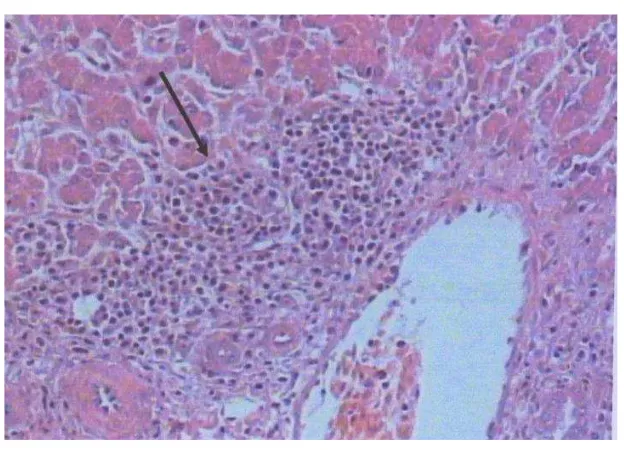
Fotomikkrograf hati ayam yang diinfeksi MDV dosis 0,125 x 103EID50 pewarnaan Hematoksilin-Eosin (HE)……….
Fotomikkrograf hati ayam yang diinfeksi MDV dosis 1 x 103EID50
pewarnaan Hematoksilin-Eosin (HE). ………
Fotomikrograf hati ayam yang diwarnai secara imunohistokimia terhadap iNOS metode sab dan counterstain hematoksilin ………
Grafik nilai absorbansi titer antibodi MDV uji ELISA
10, 20, dan 30 hari pascainfeksi . . . …….
Fotomikrograf bursa Fabricius ayam yang diwarnai secara imunohistokimia terhadap MDV, metode SAB dan counterstain Hematoksilin………
Fotomikrograf hati ayam dengan pewarnaan hematoksilin-Eosin (HE)………..
5
8
18
21
22
31
38
38
45
48
52
DAFTAR LAMPIRAN
Halaman
1.
2.
3.
4.
5.
6.
7.
8.
9.
10.
Analisis ragam bobot realif bursa Fabricius, timus, dan limpa 20 hari p.i ……….
Analisis ragam bobot realif bursa Fabricius, timus, dan limpa 40 hari p.i ... ……….
Analisis ragam diameter folikel bursa Fabricius 20 hari p.i ….
Analisis ragam diameter folikel bursa Fabricius 40 hari p.i ….
Analisis ragam reaksi positif iNOS 20 hari p.i ………
Analisis ragam titer antibodi terhadap MDV………
Analisis ragam sel darah putih dan presentase limfosit pada 20 p.i………..
Analisis ragam sel darah putih dan presentase limfosit pada 40 ………..
Analisis ragam jumlah limfosit submukosa proventrikulus pada 20 p.i. ……….
Analisis ragam jumlah limfosit submukosa proventrikulus pada 40 p.i ...
72
73
74
75
76
77
78
79
80
PENDAHULUAN
Latar Belakang
Neoplasma atau tumor adalah transformasi sejumlah gen yang menyebabkan gen tersebut mengalami mutasi. Gen yang mengalami mutasi disebut proto-onkogen dan gen supresor tumor, yang dapat menimbulkan abnormalitas pada sel somatik. Usia sel normal ada batasnya, sementara sel tumor tidak mengalami kematian sehingga multiplikasi dan pertumbuhan sel berlangsung tanpa kendali. Sel neoplasma mengalami perubahan morfologi, fungsi, dan siklus pertumbuhan, yang akhirnya menimbulkan disintegrasi dan hilangnya komunikasi antarsel. Tumor diklasifikasikan sebagai benigna, yaitu kejadian neoplasma yang bersifat jinak dan tidak menyebar ke jaringan di sekitarnya. Sebaliknya, maligna disinonimkan sebagai tumor yang melakukan metastasis, yaitu menyebar dan menyerang jaringan lain.
Kanker adalah penyakit kompleks pada sejumlah besar gen seluler yang telah mengalami perkembangan malignansi. Gen tersebut dapat dibedakan menjadi dua kelompok, yaitu sebagai onkogen dan gen supresor tumor. Onkogen dikategorikan sebagai kanker yang disebabkan oleh virus yang terbagi dalam dua grup, yaitu virus tumor DNA dan virus tumor yang mengandung RNA yang disebut juga retrovirus (Benchimol dan Minden 1998). Virusherpes sangat tumorogenik pada hewan yang pada mulanya berada pada episom sel dan tidak terintegrasi pada genom inang. Pada kejadian penyakit tumor biasanya tidak ditemukan adanya virus di dalam sel, dan DNA virus herpes hanya sedikit berada sebagai herpes yang melakukan transformasi pada sel. Hal ini dinyatakan sebagai mekanisme hit and run pada onkogenesis sehingga menyebabkan kerusakan khromosom atau kerusakan lain (Hunt 2003). Herpes virus onkogenik termasuk virus DNA yang menyebabkan penyakit Marek pada ayam, virus herpes karsinoma pada katak, virus herpes
saimiri pada primata selain manusia, virus Epstein Bar (limfosarkoma) pada manusia, virus herpes 6 pada yang berasosiasi dengan human Kaposi’s sarcoma (Cheville 1999).
kegunaan MD sebagai model pada penelitian onkologi virus herpes (Burges dan Davison 2002). Infeksi MDV pada ayam dapat dijadikan sebagai model infeksi virusherpes onkogen untuk hewan lain.
Periode infeksi MDV meliputi tiga bentuk, yaitu infeksi akut (produktif) yang menimbulkan lisis sel limfosit B dan limfosit T, infeksi laten yang bersifat nonproduktif, dan infeksi transforming, yaitu transformasi gen pada limfosit T. Pada infeksi produktif terjadi replikasi DNA virus, sintesis protein yang menghasilkan partikel virus secara lengkap. Virus menginfeksi, merusak, dan membunuh limfosit B maupun limfosit T. Selama infeksi terjadi sitolisis sehingga pada puncak replikasi virus terjadi imunosupresi dan peningkatan sensitivitas inang pada infeksi bersamaan dengan penurunan bobot relatif bursa Fabricius dan timus (Payne dan Venugopal 2000, Islam et al. 2002).
Pada infeksi laten tidak terjadi replikasi DNA, transkripsi, maupun sintesis protein. Kejadian ini dialami pada infeksi MDV serotipe 2 dan 3 nononkogen. Sel T yang terinfeksi bisa berubah menjadi infeksi laten atau bisa merespons onkogenesitas gen virus yang mengalami transformasi. Infeksi
transforming hanya terjadi pada sel yang terinfeksi oleh MDV serotipe 1. Beberapa subset limfosit T, yaitu CD4 dan CD8 merupakan target transformasi karena bagian tersebut berperan sebagai tempat perlekatan awal infeksi sitolisis (Calnek et al. 1998, Payne dan Venugopal 2000).
Virus penyebab tumor disebut virus onkogen dan gen yang ada pada virus disebut viral oncogen (V-onc) yang homolog dengan sekuen DNA pada gen seluler inang, yaitu protooncogen (C-onc) yang dapat berinteraksi dengan gen virus. Terjadinya transformasi pada gen seluler inang oleh gen virus bergantung pada resistensi seluler inang, virulensi virus penyebab, dan kehadiran substansi kimia penyebab tumor, yaitu bahan karsinogen yang menginduksi terjadinya mutasi.
3
Radikal bebas merupakan bahan karsinogen yang menimbulkan mutasi gen sehingga dapat menginduksi terjadinya kanker.
Tekanan oksidatif diinduksi secara luas oleh faktor lingkungan termasuk sinar ultraviolet, serangan patogen, reaksi hipersensitif, kerja herbisida, dan kekurangan oksigen. Spesies oksigen reaktif (ROS), hidrogen peroksida (H2O2), dan superoksida (O2-) dihasilkan oleh sejumlah reaksi seluler yang dikatalisis oleh besi (Fe-2) dan reaksi enzimatik seperti lipooksigenase, peroksidase, NADPH oksidase, dan santin oksidase. Sejumlah komponen seluler yang peka terhadap kerusakan yang disebabkan oleh radikal bebas adalah lipid, yaitu peroksidasi pada asam lemak tidak jenuh pada membran, denaturasi protein dan asam nukleat. Pembentukan ROS dapat dicegah oleh antioksidan. Pada tanaman beberapa senyawa fenolik merupakan antioksidan potensial: flavonoid, tanin, dan lignin merupakan prekursor yang bekerja pada penangkapan senyawa ROS (Blokhina et al. 2003).
Mekanisme penyerangan oleh radikal bebas termasuk ROS menginduksi peroksidasi pada asam lemak yang memiliki beberapa ikatan rangkap pada membran sel lipid bilayer yang menyebabkan reaksi berantai peroksidasi lipida sehingga terjadi kerusakan pada membran sel, oksidasi pada lipida membran dan protein, yang menyebabkan kerusakan pada bagian-bagian dari sel termasuk DNA (Miller 1996). Pada saat ini penggunaan antioksidan sintetik seperti Torlok C, Prowl galat, dan mono-tertiery-butyl-hidroquinone (TBHQ) sedang mendapat perhatian karena mempunyai efek mengurangi kerusakan oksidatif, namun mempunyai aktivitas yang dapat merugikan konsumen, antara lain gangguan fungsi hati, paru-paru, mukosa usus, dan keracunan. Untuk mengatasi hal tersebut sebaiknya dipilih memanfaatkan antoksidan alami (Manampiring et al. 2001).
oksidatif yang ditimbulkan oleh semua proses penyakit menyebabkan flavonoid layak digunakan untuk pengendalian sejumlah penyakit (Miller 1996).
Daun dan batang benalu teh mengandung senyawa alkaloid, flavonoid, terpenoid, glikosida, triterpen, saponin, dan tanin (Nugroho et al. 2000, Santoso 2001, Tambunan et al. 2003). Benalu teh secara tradisional digunakan untuk penyembuhan berbagai penyakit diare, kanker, dan amandel. Beberapa publikasi penelitian telah melaporkan bahwa benalu teh mempunyai efek sebagai antidiare (Saroni et al. 1998), antioksidan (Leswara dan Kartin 1998, Santoso 2001, Susmandiri 2002, Simanjuntak et al. 2004), perbaikan sistem imun (Winarno et al. 2000), dan hambatan pertumbuhan sel tumor (Nugroho et al. 2000, Murwani 2003, Winarno 2003).
Sel WEHI-164 diketahui sensitif terhadap Tumour Necrosis Factor-α
(TNF-α) dan telah digunakan sebagai model dalam penelitian. Hasil pengujian menunjukkan bahwa baik ekstrak batang maupun daun Scurrula oortiana
mampu meningkatkan sensitivitas atau suseptibilitas pada sel WEHI-164 pada TNF-α, peningkatan sensitivitas lebih dari 160 kali dibanding dengan sel kontrol tanpa perlakuan. Penelitian ini menunjukkan bahwa ekstrak air S. oortiana
secara nyata sitotoksik pada sel tumor WEHI-164 dan meningkatnya sensitivitas sel tumor pada TNF-α sehingga mengalami lisis (Murwani 2003).
Kerangka Pemikiran
Rebusan benalu teh sudah dikenal oleh masayarakat sebagai obat kanker tetapi belum diketahui bagaimana mekanismenya. Karena itu perlu dilakukan studi yang terukur yang dapat mengungkapkan mekanisme antikanker dari benalu teh, seperti yang diuraikan pada penelitian ini.
5
pada subset CD4 maupun CD8 yang menyebabkan kanker limfosit yang disebut limfoma.
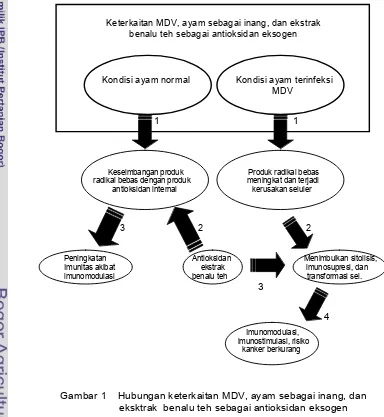
Potensi antioksidan benalu teh diharapkan mampu berperan sebagai penghambat oksidasi radikal bebas sehingga mengurangi kerusakan sel-sel pada sistem imun. Potensi antioksidan dalam mengurangi risiko kanker dengan cara menghambat induksi mutasi gen oleh kelebihan produksi radikal bebas akibat infeksi. Pensghambatan induksi mutasi oleh radikal bebas diharapkan dapat mengurangi mutasi yang disebabkan oleh MDV onkogen. Secara ringkas keseluruhan latar belakang di atas dituangkan pada Gambar 1.
Keterkaitan MDV, ayam sebagai inang, dan ekstrak benalu teh sebagai antioksidan eksogen
Kondisi ayam normal Kondisi ayam terinfeksi MDV
1 1
Keseimbangan produk Produk radikal bebas radikal bebas dengan produk meningkat dan terjadi antioksidan internal kerusakan seluler
3 2 2
Peningkatan Antioksidan Menimbulkan sitolisis, Imunitas akibat ekstrak imunosupresi, dan imunomodulasi benalu teh transformasi sel.
3
4
Imunomodulasi, imunostimulasi, risiko
kanker berkurang
Ekstrak benalu teh (Scurrula oortiana) diberikan secara oral, dan bertujuan untuk meningkatkan imunitas tubuh serta mengurangi risiko kanker pada ayam ras petelur betina yang diuji tantang dengan MDV. Ayam yang terinfeksi MDV memberikan respons imun nonspesifik berupa radikal bebas yang merusak sel-sel yang termasuk sel-sel limfosit sehingga menimbulkan sitolisis dan dapat menginduksi kejadian mutasi gen sebagai penyebab awal kejadian kanker. Peningkatan produksi radikal bebas memerlukan peningkatan antioksidan yang disuplai dari luar tubuh, yang disebut antioksidan eksogen. Benalu teh mengandung flavonoid, terpenoid, yang memiliki potensi sebagai antioksidan eksogen yang dapat dijadikan sebagai suplai antioksidan.
Tujuan Penelitian
Tujuan umum penelitian ini adalah untuk mempelajari pengaruh ekstrak benalu teh Scurrula oortiana pada fenomena imunologis dan risiko kanker pada ayam yang diinfeksi virusherpes MDV onkogen, sedangkan secara khusus tujuan penelitian ini adalah:
1. Pembuktian secara ilmiah khasiat ekstrak S. oortiana sebagai imunomodulator dan mengurangi risiko kanker.
2. Untuk memperjelas mekanisme antitumor dari benalu teh, menggunakan parameter imunologi dari ayam ras petelur yang diinfeksi MDV onkogenik.
3. Menjadikan benalu teh S. oortiana sebagai obat herbal berstandar melalui uji in vivo menggunakan ayam sebagai hewan percobaan.
Manfaat Penelitian
1. Metode dan hasil pada kajian imunitas dan onkogenisitas dapat dijadikan sebagai acuan dalam mengkaji onkogenik virus herpes pada spesies lain.
2. Membuat simulasi ayam sebagai hewan model penanggulangan kanker yang disebabkan oleh virus herpes menggunakan bahan asal tumbuhan lainnya.
3. Meningkatkan kepercayaan masyarakat tentang manfaat benalu teh yang berkhasiat mengurangi risiko kanker.
TINJAUAN PUSTAKA
Imunitas Tubuh
Resistensi dan pemulihan pada infeksi virus bergantung pada interaksi
antara virus dan inangnya. Pertahanan inang bekerja langsung pada virus atau
secara tidak langsung pada replikasi virus untuk merusak atau membunuh sel
yang terinfeksi. Fungsi pertahanan nonspesifik inang pada awal infeksi untuk
menghancurkan virus adalah mencegah atau mengendalikan infeksi, kemudian
adanya fungsi pertahanan spesifik dari inang termasuk pada infeksi virus
bervariasi bergantung pada virulensi virus, dosis infeksi, dan jalur masuknya
infeksi (Mayer 2003).
Sistem imun pada unggas bekerja secara umum seperti sistem imun
pada mamalia. Stimulasi antigenik menginduksi respons imun yang dilakukan
sistem seluler secara bersama-sama diperankan oleh makrofag, limfosit B, dan
limfosit T. Makrofag memproses antigen dan menyerahkannya kepada limfosit.
Limfosit B, yang berperan sebagai mediator imunitas humoral, yang mengalami
transformasi menjadi sel plasma dan memproduksi antibodi. Limfosit T
mengambil peran pada imunitas seluler dan mengalami diferensiasi fungsi yang
berbeda sebagai subpopulasi (Sharma 1991).
Antigen eksogen masuk ke dalam tubuh melalui endosistosis atau
fagositosis. Antigen-presenting cell (APC) yaitu makrofag, sel denrit, dan
limfosit B merombak antigen eksogen menjadi fragmen peptida melalui jalan
endositosis. Limfosit T mengeluarkan subsetnya, yaitu CD4, untuk mengenal
antigen bekerja sama dengan Mayor Hystocompatablity Complex (MHC) kelas
II dan dikatakan sebagai MHC kelas II restriksi. Antigen endogen dihasilkan
oleh tubuh inang. Sebagai contoh adalah protein yang disintesis virus dan
protein yang disintesis oleh sel kanker. Antigen endogen dirombak menjadi
fraksi peptida yang selanjutnya berikatan dengan MHC kelas I pada retikulum
endoplasma. Limfosit T mengeluarkan subsetnya, yaitu CD8, mengenali
antigen endogen untuk berikatan dengan MHC kelas I, dan ini dikatakan
sebagai MHC kelas I restriksi (Kuby 1999, Tizard 2000).
Limfosit adalah sel yang ada di dalam tubuh hewan yang mampu
mengenal dan menghancurkan bebagai determinan antigenik yang memiliki
dua sifat pada respons imun khusus, yaitu spesifitas dan memori. Limfosit
yang diproduksi, namun morfologinya sulit dibedakan (Abbas et al. 2000).
Limfosit berperan dalam respons imun spesifik karena setiap individu limfosit
dewasa memiliki sisi ikatan khusus sebagai varian dari prototipe reseptor
antigen. Reseptor antigen pada limfosit B adalah bagian membran yang
berikatan dengan antibodi yang disekresikan setelah limfosit B yang mengalami
diferensiasi menjadi sel fungsional, yaitu sel plasma yang disebut juga sebagai
membran imunoglobulin. Reseptor antigen pada limfosit T bekerja mendeteksi
bagian protein asing atau patogen asing yang masuk sel inang (Janeway et al.
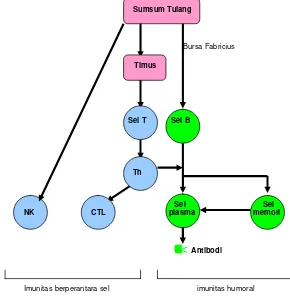
2001). Mekanisme kerja sistem imun disajikan pada Gambar 2 (Cann 1977).
Sumsum Tulang
Bursa Fabricius
Timus
Sel T Sel B
Th
Sel Sel NK CTL plasma memori
<
AntibodiImunitas berperantara sel imunitas humoral
Gambar 2 Mekanisme secara umum sistem imun (Cann 1997)
Sel limfosit B berasal dari sumsum tulang belakang dan mengalami
pendewasaan pada jaringan ekivalen bursa. Jumlah sel limfosit B dalam
keadaan normal berkisar antara 10 dan 15%. Setiap limfosit B memiliki 105 B
cell receptor (BCR), dan setiap BCR memiliki dua tempat pengikatan yang
identik. Antigen yang umum bagi sel B adalah protein yang memiliki struktur
9
membedakan antara sel B dan sel T, yang mengikat antigen yang sudah
terproses dalam sel (Kresno 2004).
Jajaran ketiga sel limfoid adalah natural killer cells (sel NK) yang tidak
memiliki reseptor antigen spesifik dan merupakan bagian dari sistem imun
nonspesifik. Sel ini beredar dalam darah sebagai limfosit besar yang khusus
memiliki granula spesifik yang memiliki kemampuan mengenal dan membunuh
sel abnormal, seperti sel tumor dan sel yang terinfeksi oleh virus. Sel NK
berperan penting dalam imunitas nonspesifik pada patogen intraseluler
(Janeway et al. 2001).
Antibodi diproduksi oleh sistem imun spesifik primer pada pemulihan
pada infeksi virus dan pertahanan pada serangan infeksi virus. Sel T lebih
berperan pada pemulihan infeksi virus. Sitotoksik sel T (CTLs) atau CD8
berperan pada respons imun terhadap antigen virus pada sel yang diinfeksi
dengan cara membunuh sel yang terinfeksi untuk mencegah penyebaran
infeksi virus. Sel T helper (CD4) adalah subset sel T yang berperan membantu
sel B untuk memproduksi antibodi. Limfokin disekresikan oleh sel T untuk
mempengaruhi dan mengaktivasi makrofag dan sel NK sehingga meningkat
secara nyata pada penyerangan virus (Mayer 2003).
Patogen yang mampu dijangkau oleh antibodi adalah hanya antigen
yang berada pada peredaran darah dan di luar sel, padahal beberapa bakteri
patogen, parasit, dan virus perkembangan replikasinya berada di dalam sel
sehingga tidak dapat dideteksi oleh antibodi. Penghancuran patogen ini
membutuhkan peran limfosit T sebagai imunitas yang diperantarai oleh sel.
Limfosit T mengenal sel yang terinfeksi virus, virus yang menginfeksi sel
bereplikasi di dalam sel dengan memanfaatkan sistem biosintesis sel inang.
Derivat antigen dari replikasi virus dikenal oleh limfosit T sitotoksik. Sel tersebut
mampu mengontrol sel yang terinfeksi sebelum replikasi virus dilangsungkan
secara lengkap. Sel T sitotoksik merupakan ekspresi dari molekul CD8 pada
permukaannya (Janeway et al. 2001).
Kanker
Pada keadaan normal pergantian dan peremajaan sel terjadi sesuai
kebutuhan melalui proliferasi sel dan apoptosis di bawah pengaruh
proto-onkogen dan gen supresor tumor (Silalahi 2006). Tumor adalah penyakit
memerlukan waktu yang lama. Hal inilah yang menyebabkan keterbatasan
efektivitas kemoterapi tumor. Fenomena ini akan meningkatkan jumlah
kematian (Flora dan Ferguson 2005).
Perbedaan pokok antara sel normal dan sel kanker yang teridentifikasi
bahwa sel normal usianya terbatas, sedangkan sel kanker adalah immortal. Sel
neoplastik tidak berkembang secara terintegrasi dan tidak ada ketergantungan
pada populasi. Regulasi pada kontrol mitosis, diferensiasi, dan interaksi
antarsel mengalami gangguan (Cheville 1999, Cambel dan Smith 2000).
Gen seluler inang yang homolog dengan onkogen virus disebut
protoonkogen. Gen tersebut mampu memproduksi protein yang memiliki
kemampuan menginduksi transformasi seluler setelah mengalami mutasi, yaitu
perubahan di bawah kontrol promotor yang memiliki aktivitas tinggi. Biasanya
protoonkogen berperan mengkode produksi protein pada replikasi DNA atau
mengontrol perkembangan pada beberapa stadium pertumbuhan normal.
C-onc adalah gen seluler yang diekspresikan pada beberapa stadium
perkembangan sel. Produk onkogen adalah protein inti misalnya myc, myb.
(King 2001, Hunt 2003).
Gen pengatur dapat mengalami mutasi, menjadikan gen tersebut tidak
peka terhadap sinyal regulasi normal. Gen supresor yang mengalami mutasi,
mengakibatkan gen tersebut menjadi inaktif. Untuk mengatasi penyakit
kompleks diperlukan pertahanan dengan berbagai cara yang strategis dan
pencegahan diperlukan untuk mengurangi metastasis pada kanker (Steele dan
Kellof 2005).
Gen supresor tumor yang mengalami perubahan antara lain gen p53,
adalah produk protein yang memiliki bobot molekul 53 kD. Protein tersebut
berfungsi sebagai pengatur proliferasi sel dan mediator pada apoptosis, yaitu
program kematian sel. Gen ini juga merupakan gen yang menginduksi
kerusakan DNA dengan cara menghambat mekanisme atau proses perbaikan
kembali DNA. Hilangnya fungsi gen p53 atau terjadinya mutasi gen tersebut
menjadikan sel terhindar dari kerusakan DNA, pertumbuhan dan kematian sel
tidak terkontrol, pembelahan sel terjadi secara terus menerus tanpa
mengalami apoptosis (Williamson et al. 1999, Silalahi 2006). Apoptosis
berperan penting pada fisiologi normal pada spesies hewan, termasuk program
11
jaringan, pendewasaan sel imun, dan beberapa aspek penuaan (Reed et al.
2004).
Apoptosis adalah program kematian sel yang mekanismenya
diorganisir secara fisiologis untuk merusak sel abnormal atau mengalami
kerusakan. Keadaan ini merupakan respons sel normal yang terjadi selama
pertumbuhan dan metamorfosis semua hewan multiseluler, yang merupakan
hasil kerja enzim proteolitik, yaitu caspase dimana semua enzim ini memiliki
sistin sebagai sisi aktif dan pembelahan protein target pada asam aspartat
spesifik sebagai derivat dari sistin aspartase. Sel normal dapat mengalami
transformasi oleh onkogen dan proses ini dapat dicegah oleh produk yang
dihasilkan gen lainnya yang disebut tumour suppressor genes. Satu di antara
gen ini adalah p53 yang menghasilkan 393 residu asam amino inti fosfoprotein
yang berikatan dengan DNA yang transkripsinya diaktivasi oleh beberapa
promotor. Protein p53 mampu menghambat pertumbuhan sel dan
mempengaruhi apoptosis (Cambel dan Smith 2000, Taraphdar et al. 2001).
Feng et al. (2003) pertumbuhan dan metastasis tumor bergantung pada
bertambahnya suplai darah melalui angiogenesis, ekspresi yang berlebihan
dari iNOS dan vascular endothelial growth factor (VEGF) menginduksi
angiogenesis pada tumor. P53 menekan angiogenesis dengan cara
menurunkan VEGF dan iNOS.
Transformasi sering menimbulkan hilangnya kontrol pertumbuhan,
kemampuan untuk menginvasi matriks ekstraseluler dan dediferensiasi. Pada
karsinoma, beberapa sel epitel yang mengalami transformasi adalah
mesenchimal epitelial. Pada transformasi sel sering terjadi kerusakan
kromosom. Bagian genom virus yang menyebabkan tumor disebut onkogen.
Gen asing ini dapat bergabung pada sel dan menyebabkan sel tidak
mengalami kematian sehingga menjadikan pertumbuhan tidak terkendali (Hunt
2003).
Fusi genetik dengan kromosom lain dinyatakan sebagai translokasi.
Sejumlah translokasi menimbulkan gangguan ekspresi dan fungsi gen yang
berkaitan dengan kontrol pertumbuhan sel. Translokasi terkarakterisasi pada
reseptor atau lokus sel T terlihat pada tumor sel T. Rearangement ini sering
bersamaan dengan translokasi kromosom termasuk pada lokus yang
penyebab kanker yang menyebabkan fungsi dan ekspresi terganggu sehingga
disebut onkogen (Janeway et al. 2001).
Onkogen adalah istilah untuk agen aktif oleh gen virus onkogenik,
karena pada bentuk kanker yang lain tidak jelas. Selanjutnya ekspresi yang
berlebihan pada beberapa proto-onkogen telah ditunjukkan kejadiannya pada
transformasi beberapa tipe sel dan kanker, dan level beberapa proto-onkogen
ternyata mengalami kenaikan (Cambel dan Smith 2000).
Kerusakan oksidatif pada DNA akibat radiasi, radikal bebas, dan
senyawa oksigen yang bersifat oksidatif merupakan penyebab terpenting
kanker (Silalahi 2006). Transfomasi seluler oleh virus DNA menghasilkan
protein yang berinteraksi dengan protein seluler. Terjadinya transformasi DNA
biasanya pada sel mengalami infeksi nonproduktif. Pada kejadian ini, DNA
virus berintegrasi pada DNA seluler sehingga sel mengalami perkecualian, dan
pada kasus ini adalah oleh virus papiloma dan virus herpes yang DNA virus
berada pada episom. Virus tumor berinteraksi dengan sel melalui satu dari dua
jalan, yaitu 1) infeksi produktif, yaitu virus melakukan siklus replikasi secara
lengkap dan menimbulkan lisis sel, 2) infeksi nonproduktif, yaitu transformasi
virus pada sel yang melakukan siklus replikasi secara tidak lengkap. Selama
infeksi nonproduktif, genom virus atau versi potongannya terintegrasi pada gen
seluler, v-onc, yang bertanggung jawab pada perubahan malignan (Murphy et
al. 2001, King 2001).
Penyakit Marek
Intervensi pencegahan kanker secara efektif dapat ditingkatkan dengan
cara memilih hewan model yang sesuai sehingga menghasilkan potensi baik
dari segi klinik maupun epidemiologi (Hursting et al. 2005). Virus onkogenik
penting pada peternakan maupun populasi hewan. Sumber penyakit
dikategorikan dari sejumlah familia virus, termasuk retroviridae dan
herpesviridae. Peran hewan asal penyakit dapat dijadikan sebagai hewan
model pada kejadian penyakit virus pada manusia, baik bertujuan pada
imunitas, pengobatan, maupun mekanisme patofisiologi (Lohellt 2006).
Marek’s disease (MD) disebabkan oleh virus DNA termasuk pada
kelompok virus herpes penyebab kanker pada ayam yang biasanya
menimbulkan persoalan ekonomi yang berat. Virus ini tumbuh dan berkembang
13
melalui ketombe dan debu (Simonsen 1987, Silva et al. 2004). Marek’s disease
virus (MDV) virus herpes pada unggas menimbulkan imunosupresi dan limfoma
pada ayam yang peka, biasanya menyerang ayam piaraan yang tersebar
meluas pada populasi unggas di seluruh dunia. Infeksi pada spesies lain terjadi
pada kalkun dan puyuh. Replikasi MDV sama dengan virus herpes lainnya,
yaitu sangat bergantung pada sel. Penyebaran infeksi pada sel yang lain terjadi
melalui kontak langsung dari sel yang terinfeksi, dan pemindahan virus antarsel
terjadi melalui jembatan sitoplasmik (Payne dan Venugopal 2000, Anobile et al.
2006).
MDV menginduksi paralisis dan limfoma secara cepat dan pada fase
transformasi tidak ditemukan virus secara utuh. Penyakit Marek adalah
penyakit limfoproliferatif dan neurotropik pada ayam piaraan, yang disebabkan
oleh virus herpes alfa. Jozsef Marek menemukan penyakit tersebut pada tahun
1907. Simtom penyakit tersebut ditandai dengan paralisis pada leher dan
sayap, yang bersamaan dengan inflamasi pada syaraf perifer yang dikenal
sebagi polyneuritis. Duapuluh tahun kemudian penyakit tersebut ditemukan di
Amerika dan Belanda, dan namanya berubah menjadi fowl paralysis (Parcells
et al. 1999, Payne dan Venagupol 2000).
Segera setelah infeksi melalui alat pernafasan virus menyebar ke organ
limfoid primer bursa Fabricius dan timus. Target pertama diantaranya adalah
derivat bursa Fabricius (limfosit B), namun sejumlah derivat timus (limfosit T)
juga mengalami infeksi. Selama 3 sampai 6 atau 7 hari pascainfeksi (p.i.)
terjadi infeksi sitolisis, dan sering juga terjadi limforetikulitis, dan pembesaran
limpa yang disertai nekrosis dan atrofi bursa Fabricius dan timus (Calnek et
al.1998). MDV isolat Austalia MPV 57 menimbulkan imunosupresi pada ayam
pedaging bersamaan dengan penurunan bobot relatif bursa Fabricius dan
timus, penurunan jumlah limfosit B dan limfosit T, dan penurunan titer antibodi
pada infeksius bronchitis (IB), dan peningkatan kepekaan pada infeksi
Escherichia colli pada hari ketiga sampai dengan hari ke-35 setelah dilakukan
uji tantang (Islam et al. 2002).
Dalam perjalanan waktu, virulensi MDV mengalami peningkatan. Infeksi
virus secara umum menjadi lebih akut dan lebih bervariasi dalam kejadian
secara alami dan spektrum penyebarannya pada inang. Sejak dekade tahun
1990, virus MDV menjadi lebih patogen dengan munculnya strain vvMDV dan
tetapi juga gejala klinis yang baru (bervariasi). Keragaman gejala klinis terlihat
dari neoplastik MD sebelumnya yang terjadi hanya pada ayam bibit umur di
bawah 16 minggu. Dengan isolat baru, kejadian neoplastik MD tersebut tetap
terjadi pada umur di atas 20 minggu walaupun telah dilakukan vaksinasi
(Zavala 1997, Witter 1998).
Peningkatan virulensi virus lebih cepat daripada strategi intervensi
pengembangan teknologi vaksin (Witter 1998). Terdapat tiga bentuk infeksi
MDV. Infeksi awal adalah infeksi akut yang terjadi pada epitelium folikel bulu,
yang menghasilkan virion beramplop yang sangat infektif. Infeksi
restriktif-produktif terjadi juga pada organ limfoid. Limfosit adalah sel target khusus
siklus kehidupan virus. Genom MDV dapat masuk ke dalam limfosit muda
pada saat pembelahan sel. Virion MDV juga dapat pindah antarsel melalui
jembatan sitoplasma. Infeksi MDV pada limfosit menimbulkan imunosupresi
sehingga ayam menjadi lebih rentan terhadap patogen dan pengembangan
tumor. Pada infeksi transforming, tumor hanya terjadi pada limfosit T dan
penyebabnya adalah MDV serotipe 1 (Davison 1997, Fadly 1997).
Periode infeksi akut (produktif) ditandai dengan terjadinya lisis sel,
dilanjutkan dengan infeksi laten yang bersifat nonproduktif, dan infeksi
transforming. Pada infeksi produktif terjadi replikasi DNA virus, sintesis protein,
dan menghasilkan partikel virus. Virus menginfeksi dan merusak limfosit B
maupun limfosit T. Selama infeksi produktif terjadi lisis pada puncak replikasi
virus sehingga menyebabkan imunosupresi, dan meningkatnya kepekaan
terhadap infeksi, bersamaan dengan turunnya bobot relatif bursa Fabricius dan
timus. Penyakit imunosupresi pada unggas berpengaruh nyata secara
ekonomis pada faktor produksi (Calnek et al. 1998, Payne dan Venugopal
2000, Islam et al. 2002).
Replikasi produktif virusherpes pada bursa fabricius dan timus
menimbulkan imunosupresi transien (sementara) dan perubahan akut sitolitik
pada organ ini yang ditandai dengan atropi. Infeksi eksperimental
menyebabkan lesi pada bursa fabricius dalam bentuk lesi folikuler, nekrosis
limfoid sehingga mengalami penipisan, dan pembentukan kista. Timus
mengalami atropi dan limfosit hilang baik pada korteks maupun medula. Badan
inklusi intrafolikuler dapat muncul pada sel yang mengalami lesi degeneratif
15
Pada sel yang menerima masuknya virus secara keseluruhan yang
seluruh bagian gen virus terekspresi, menjadikan virus mampu melakukan
replikasi, sel inang lisis dan mengalami kematian. Bila sel tidak bisa menerima
virus secara keseluruhan untuk replikasi, sebagian DNA virus berintegrasi pada
bagian kromosom sel inang secara acak. Pada keadaan ini hanya sebagian
genom virus yang terekspresi, protein virus tidak terbentuk dan tidak dihasilkan
virus anakan (Hunt 2003). Pada infeksi laten tidak terjadi replikasi DNA,
transkripsi, maupun sintesis protein. Kejadian ini dialami pada infeksi MDV
serotipe 2 dan 3 nononkogen. Sel limfosit T yang terinfeksi bisa berubah
menjadi infeksi laten atau sel T bisa merespons onkogenesitas gen virus dan
selanjutnya mengalami transformasi. Infeksi transforming hanya terjadi pada
sel T yang terinfeksi oleh MDV serotipe 1. Beberapa subset sel T, yaitu CD4
dan CD8 merupakan target transformasi karena bagian tersebut berperan
sebagai tempat perlekatan pada awal infeksi sitolisis (Calnek et al. 1998, Payne
dan Venugopal 2000).
Infeksi transforming ditandai oleh marker permukaan spesifik yang
hanya ditemukan pada sel yang mengalami tumor. Marker tersebut ditandai
dengan 2 antigen potensial, yaitu Marek’s Disease Tumour-assciated Surface
Antigen (MATSA) yang dideteksi pada sel limfomas dan limfoblastoid. Antigen
kedua yang dideteksi dengan menggunakan antibodi monoklonal AV37,
ditemukan pada CD4+ yang mengalami transformasi. Limfokin yang
disekresikan oleh Antigen Presenting Cell (APC) merangsang aktivitas
fungsional CD4 sel T. CD8 sel T yang terinfeksi MDV selanjutnya akan
mengenal antigen yang berikatan dengan molekul MHC-1 pada respons imun
normal, MHC-1 yang sudah berikatan tersebut menghasilkan nukleated. Jika
MHC-1 berikatan dengan fragmen antigen MDV, atau jika MHC-1 mengalami
transformasi neoplastik, CD-8 (cytotoxic) sel T akan mengalami hambatan
imunitas. Hal ini diketahui bahwa limfosit T adalah sel target untuk transformasi
pada neoplastik (tumoral) sel pada infeksi MDV (Zavalo 1997, Payne dan
Venugopal 2000).
MDV memiliki fosfoprotein unik, yaitu pp38, yang merupakan penanda
yang berperan penting dalam patogenesis penyakit Marek. pp38 diperlukan
pada infeksi sitolisis pada limfosit B, namun bukan pada feather folicle
epithelium (FFE), untuk mencukupi pada saat infeksi laten pada sel T dan
Disimpulkan bahwa pp38 potensial digunakan pada tingkat kejadian tumor dan
respons imun terhadap MDV (Barrow et al. 2003, Gimeno et al. 2005). Deteksi
fosfoprotein spesifik MDV pp38, identifikasi pp38 pada limfosit menggunakan
antibodi monoklonal tarhadap marker permukaan limfosit. Ekspresi pp38 pada
limpa ditunjukkan pada saat fase infeksi sitolitik. Proporsi pp38 pada limfosit
saat 4 hari p.i adalah sebesar 0.43%. Dari jumlah tersebut 95% berada pada
limfosit B dan hanya 4% berada pada limfosit T baik pada CD4 maupun CD8.
(Baigent et al. 1996). Potensi yang nyata menunjukkan tidak lengkapnya blok
gen pada internal repeat HVT seperti yang ada pada MDV-1. Disini termasuk
pp38 dan meq gene. Dengan implikasi bahwa MDV-1 manginduksi limfoma.
Ada kemungkinan munculnya gen lain pada sisi ini, tetapi tidak muncul pada
HVT, mungkin memiliki peran panting yang mempengaruhi potensi biologis dari
virus (Kingham et al. 2001).
Kejadian pada kelompok ayam resisten dan peka, terjadi pertambahan
virus sampai hari ke-10 pascainfeksi (pi), setelah itu terjadi penambahan virus
pada kelompok ayam peka dan terjadi penurunan pada ayam resisten. Untuk
dua sitokin ada perbedaan antar galur, yaitu interleukin-6 (IL-6) dan IL-18.
Splenosit ayam yang peka mengekspresikan tingginya level transkipsi kedua
interleukin yang mengakibatkan sitolitik. Sebaliknya pada splenosit ayam
resisten hal itu tidak terjadi. Keadaan ini menunjukkan adanya indikasi bahwa
dua sitokin dapat berperan dalam perbedaan respons imun, yaitu galur resisten
menunjukkan adanya infeksi laten dan pada galur peka menghasilkan limfoma
(Kaiser et al. 2003). Beberapa subset sel T, yaitu CD4 dan CD8, berperan
sebagai target transformasi karena bagian tersebut merupakan target infeksi
sitolisis. Sel-sel T yang terinfeksi bisa berubah menjadi infeksi laten atau sel T
merespons onkogenesitas gen virus dan selanjutnya terjadi transformasi
(Calnek et al. 1998).
Pada sistem imun, tumor diklasifikasikan pada limfoma dan leukemia.
Limfoma adalah proliferasi tumor pada jaringan limfoid, yaitu sumsum tulang,
limfonodus, atau timus. Leukemia cenderung menyebabkan proliferasi pada sel
tunggal dan terdeteksi oleh adanya peningkatan jumlah aliran darah atau cairan
limfe. Leukemia dapat berkembang pada jalur limfoid maupun mieloid (Kuby
1999). Limfoma dapat terjadi pada satu atau lebih sejumlah organ atau
jaringan. Lesi limfomatus dapat terjadi pada gonad khususnya ovarium,
17
genetik strain ayam maupun virus dapat mempengaruhi distribusi lesi yang
terjadi. Pada limfoma pada MD, kebanyakan organ viseral mengalami
pembesaran yang difus, beberapa yang lain berukuran normal dan perubahan
warna menjadi pucat atau kecokelatan (Fadly 2000). Dalam hal ini kasus
penyakit Marek, ditandai dengan lesi limfomatus dan infiltrasi limfosit pada
proventrikulus, sel limfoblas dan sel retikuler pada sel-sel kelenjar (Larbier dan
Leclerco 1992).
Antibodi terhadap MDV dan HVT dapat dideteksi pada serum dengan
berbagai uji serologis termasuk imunofluresensi, agar gel presipitasi, netralisasi
virus, atau enzyme-linked immuno sorbant assay (ELISA). Polymerase chain
reaction (PCR) juga dapat digunakan untuk mendeteksi adanya 132 pasang
basa yang terulang dari MDV pada ekstraksi DNA dari limfoma MD, tetapi tidak
terdapat pada induksi limfoma oleh avian leucosis virus (ALV) atau reticulo
endothelialis virus (REV). Limfoma dari MD biasanya mengandung lebih sering
sel terinfeksi, dan lebih banyak kopi DNA virus per sel yang terinfeksi pada
jaringan yang tidak mengalami tumor pada ayam (Fadly 1997).
Peran Radikal Bebas
Oksigen merupakan unsur penting bagi kehidupan organisme. Sebagai
kekuatan oksidan, oksigen molekuler di satu pihak bermanfaat sebagai
kemampuan dasar degradasi oksidatif, yaitu sebagai substrat pada respirasi..
Di pihak lain, oksigen dapat menimbulkan kerusakan karena berperan sebagai
prekursor pada spesies oksigen reaktif (reactive oxygen spescies, ROS) yang
menimbulkan kerusakan komponen intraseluler termasuk DNA. Untuk
mengurangi pengaruh kerusakan yang ditimbulkan oleh ROS, organisme hidup
mampu menjalankan mekanisme multisistem antiROS, namun pada saat
tertentu ROS diperlukan untuk kepentingan biologis. ROS berperan sebagai
pertahanan biologis, yaitu fagositosis dan pesan jelek apoptosis, yaitu
prgogram kematian sel, dan mungkin sebagai komponen yang diduga berperan
pada sistem mutator dengan meningkatnya penyimpangan genetik pada
populasi (Skulachev 2000).
Walaupun oksigen (O2) esensial untuk kebanyakan proses kehidupan,
molekul tersebut dapat berubah menjadi molekul yang memiliki toksisitas tinggi.
Satu dari kebanyakan senyawa reaktif adalah superoksida anion (O2- ) yang
mengandung elektron yang tidak berpasangan pada orbit luarnya. Molekul
terdiri atas atom dengan elektron yang berpasangan pada kulit terluarnya,
namun pada suatu kondisi, molekul atau atom yang memiliki elektron yang
tidak berpasangan biasanya mengambil elektron lain dari sekitarnya untuk
dijadikan sebagai pasangannya. Radikal bebas umumnya merusak molekul
lain, misalnya molekul pada sel (Noguchi dan Niki 1999, Cambel dan Smith
2000).
Perubahan pada kondisi lingkungan
Memperburuk kondisi intraseluler
Ketidakseimbangan antara terbentuknya ROS dan proses penangkapan ROS
[ROS]
Peningkatan pada mutasi dan proliferasi
Gambar 3 Kemungkinan skenario pada ketidakseimbangan ROS (Skulachev 2000).
Spesies oksigen reaktif selalu dihasilkan secara normal dalam proses
produksi energi, sintesis senyawa biologis, dan fagositosis pada sistem imun.
Di lain pihak peningkatan aktivitas spesies oksigen reaktif bisa menyebabkan
sejumlah penyakit termasuk penyakit jantung, kanker, dan penuaan (Noguchi
dan Niki 1999). Asam lemak tidak jenuh mengakibatkan lemak peka terhadap
serangan oksigen sehingga menimbulkan perubahan struktur kimia. Dalam
sistem seluler peroksidasi terjadi pada biomembran di mana kandungan asam
lemak tidak jenuh yang ada menjadi sangat reaktif. Peroksidasi lemak adalah
proses reaksi kimia yang sangat kompleks termasuk melibatkan radikal
bebas, ion logam, dan sistem biologik (Jadhav et al. 1996). Ada beberapa
hubungan saling mempengaruhi antara kesehatan diet antioksidan dan ROS,
mungkin bergantung pada status kesehatan, secara individual dan mungkin
juga kepekaan secara genetik. Pada penelitian secara klinik pada
19
penyakit atau kejadian penyakit yang telah mempengaruhi kesehatan individu,
risiko sejumlah penyakit pada populasi atau pasien yang sedang menjalani
pengobatan (Seifried et al. 2003).
Pada saat fagositosis, makrofag dan neutrofil sebagai sel efektor juga
memproduksi oksigen toksik gabungan fagosom dan lisosom menjadi
fagololisosom yang bertugas membantu membunuh dan menelan
mikroorganisme. Kebanyakan kejadian yang penting di antaranya adalah kerja
hidrogen peroksida (H2O2), superoksida anion (O2-), dan nitrogen oksida (NO),
secara langsung toksik pada bakteri. Semuanya ini dihasilkan melalui oksidasi
oleh NADPH dan enzim yang lain dalam proses yang dinamakan respiratory
burst, sebagai akibat dari naiknya jumlah konsumsi oksigen sementara.
Aktivitas makrofag sangat efisien dalam menghancurkan patogen, aktivitas ini
secara in vivo biasanya bersamaan dengan kerusakan jaringan secara lokal
yang disebabkan oleh keluarnya mediator antimikrobial sebagai radikal bebas,
NO dan protease, yang juga toksik terhadap sel inang. Kemampuan aktivitas
makrofag untuk mengeluarkan mediator toksik adalah pada pertahanan inang
karena kemampuannya melawan patogen ekstraseluler yang tidak tertelan
(Abbas et al. 2000, Janeway et al. 2001).
Nitrogen oksida adalah molekul yang penting yang mempengaruhi
sistem kardiovaskuler, NO merupakan senyawa yang bersifat toksik dan
berumur pendek, berupa molekul gas yang diproduksi oleh enzim NO synthase,
dengan cara mengubah asam amino arginin menjadi NO dan sitrulin (Becker et
al. 2000). Molekul NO berperan penting sebagai regulator kardiovaskuler
bertindak untuk mengatur tekanan darah. Molekul ini diproduksi oleh neuron
dan makrofag, memiliki jumlah elektron ganjil dan sebagai radikal bebas..
Molekul ini relatif stabil namun bereaksi cepat bila bertemu dengan senyawa
yang mengandung elektron yang tidak berpasangan, misalnya molekul oksigen
misalnya anion superoksida dan ion logam (Cambel dan Smith 2001).
Penelitian terahir menggambarkan bahwa inducible nitric oxyde synthase
(iNOS) terlibat dalam kelainan metabolik yang dihubungkan dengan inflamasi
kronis tingkat ringan, aterosklerosis, dan peningkatan tumour necrosis factor
(TNF) (Muntalib 2003).
Peran sitokin pada patogenesis dan imunitas terhadap MD, yang
diinduksi oleh virus herpes menyebabkan limfoma pada sel T. Pada ayam
sampai akhir percobaan, yaitu 15 hari p.i, dimana iNOS dan IL-1ß mengalami
peningkatan antara 6 sampai 15 hari p.i. Pada ayam umur 1 hari p.i mRNA
untuk untuk mengekspresikan IF-Y dan iNOS, antara 16 sampai dengan 64
kali pada 9 hari p.i. Kesimpulan dapat diambil dimana iNOS berperan pada
patogenesis MD (Xing dan Schat 2000).
Radikal bebas diproduksi secara normal pada fungsi imunitas,
diperlukan oleh sel imun untuk membunuh patogen dan mengeluarkannya,
dalam keadaan overproduksi pada kondisi patogenik menyebabkan kerusakan
sel imun dan menimbulkan imunosupresi. Eritrofagositosis juga terjadi pada
penyakit Marek oleh makrofag. Dibutuhkan keseimbangan oksidan-antioksidan
untuk mengatur fungsi sistem imun dalam menjaga integritas dan fungsi lipida
membran, protein seluler, asam nukleat serta mengatur ekspresi gen (Wu dan
Meydani 1999, Gilka dan Spencer 1995).
Antioksidan
Antioksidan yang berasal dari tanaman telah lama dikenal potensinya
dan telah lama diketahui untuk menstabilkan senyawa radikal yang dapat
diukur aktivitas antioksidan tersebut (Kim et al. 2002). Berbeda kondisinya
dengan hewan, tumbuh-tumbuhan tingkat tinggi sebagai mahluk hidup yang
tidak bergerak, mampu menghindar dari serangan predator maupun patogen
dan juga efek tekanan kuat dari kondisi lingkungan. Tanaman menghasilkan
sejumlah senyawa kimia kompleks yang biasanya merupakan bagian dari sel
yang disebut “metabolit sekunder” yang kandungannya bukan bahan dasar
biokimia untuk hidup, tetapi sebagai bagian yang berinteraksi dengan
lingkungan. Manusia memilih makanan yang dihasilkan tanaman dan bahan
kimia pertahanan tanaman tersebut bisa dimanfaatkan untuk diet dan
kesehatan (Houghton dan Raman 1998, Williamson et al. 1999). Fito-kimia
(bahan kimia dari tanaman) mempunyai efek biologi yang efektif menghambat
pertumbuhan kanker, sebagai antioksidan, menghambat pertumbuhan mikroba,
menurunkan kolesterol darah, dan menurunkan glukosa darah (Amelia 2006).
Flavonoid merupakan suatu metabolit sekunder pada tanaman yang
terdapat pada semua bagian tanaman tersebut dan struktur kimianya secara
umum adalah kerangka C6C3C6. Penamaan sub-grup dan klasifikasi berdasar
pada subsitusi pada bagaian cincin C dan posisi pada cincin B. Sebagian besar
21
antosianin (Larbier dan Leclerco 1992, Rajalkshmi dan Narasimhan 1996).
Potensi flavonoid sebagai antioksidan dan kemampuannya mengurangi
aktivitas radikal hidroksi, anion superoksida, dan radikal peroksida lemak
menjadikan flavonoid bereperan penting. Kerusakan sangat erat kaitannya
dengan proses dan epidemiologi penyakit. Uji klinik dan laboratorium pada
flavonoid dan antioksidan yang lain menjadikan penggunaan senyawa ini
penting pada pencegahan dan pengobatan sejumlah kasus penyakit. Flavonoid
telah diketahui sebagai antibakteri, antiviral, antiinflamasi, antialergi,
antimutagenik, antitrombotik, dan aktivitas vasodilatasi (Larbier dan Leclerco
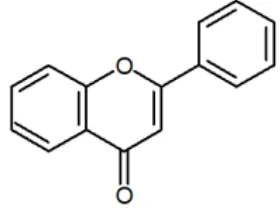
[image:36.612.269.409.263.368.2]1992, Miller 1996).
Gambar 4 Senyawa flavonoid (Miller 2004)
Sejumlah senyawa fenolat berperan sebagai bahan baku pangan yang
berasal dari tanaman, di antaranya asam fenolat, flavonoid tanin, dan lignin.
Perbedaan kultivar beberapa tanaman menunjukkan variasi yang luas baik
pada kandungan fenolat maupun kapasitas antioksidan secara in vitro (Larbier
dan Leclerco 1992, Imeh dan Khokhar 2002).
Fenolat penting diketahui sebagai substansi yang terbaik yang berperan
sebagai antioksidan sebagai kelompok donor elektron dari fenol, meningkatkan
aktivitas antioksidan melalui efek induksi. Namun aktivitas kimia sejumlah
antioksidan bergantung pada sejumlah faktor, yaitu stabilitas dan reaktivitas.
Antioksidan primer membentuk ikatan dengan radikal setelah pemindahan
hidrogen lebih penting daripada faktor lainnya (Jadav et al. 1996). Senyawa
fenol mencakup sejumlah senyawa yang umumnya mempunyai sebuah cincin
aromatik dengan satu atau lebih gugus hidroksil. Senyawa fenol cenderung
untuk larut dalam air karena paling sering terdapat bergabung dengan gula
glukosida dan biasanya terdapat dalam rongga sel. Di antara senyawa fenol
alami yang telah diketahui, lebih dari seribu struktur, flavonoid merupakan
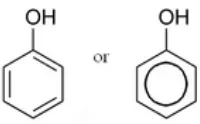
Gambar 5 Senyawa fenol (Cann 1997)
Penelitian terakhir telah menunjukkan adanya kemampuan ekstrak apel
menghambat proliferasi sel tumor secara in vitro karena diduga mengandung
senyawa fenolat atau flavonoid sebagai antioksidan. Hasil penelitian
menunjukkan hambatan secara tidak langsung, yaitu pada H2O2 setelah
berinteraksi dengan senyawa fenolat pada kultur sel, flavonoid berguna untuk
perbaikan kondisi kesehatan. Senyawa polifenolat yang diisolasi dari teh hijau
6 sendok teh per-hari, mampu menghambat perkembangan dan mestastasis
kanker prostat pada manusia (Murphy 2003, Adhami et al. 2003).
Antioksidan adalah senyawa kimia yang memilki kemampuan untuk
memberikan hidrogen radikal. Sebagai akibatnya, senyawa tersebut mampu
mengubah sifat radikal menjadi nonradikal dan terjadi perubahan oksidasi
radikal oleh antioksidan. Struktur molekul antioksidan bukan hanya memiliki
kemampuan melepas atom hidrogen tetapi juga mengubah radikal menjadi
reaktivitas rendah sehingga tidak terjadi reaksi dengan lemak. Antioksidan
terdiri atas antioksidan endogen yang dihasilkan oleh tubuh sendiri dan
antioksidan eksogen yang berasal dari makanan (Jadav et al. 1996,
Manampiring et al. 2000).
Antioksidan menjadi bentuk aktif pada oksigen reaktif termasuk pada
step inisiasi oksidasi, atau dapat memecah rantai reaksi oksidatif dengan cara
bereaksi dengan radikal peroksida membentuk ikatan antioksidan-radikal yang
stabil sehingga tidak terjadi reaksi selanjutnya atau bentuk nonradikal (Howell
dan Saeed 1999). Pertahanan antioksidan pada sel mampu mencegah
terjadinya peroksidasi lipida dan beberapa molekul biologi yang mengalami
kerusakan. Dalam hal ini ada tiga level pertahanan sebagai dasar pada sistem
eliminasi kerusakan dengan cara menghambat inisiasi atau propagasi dan
perbaikan kembali. Level pertahanan antioksidan pada enzim termasuk lipolitik
(fosfolipase), proteolitik (peptidase atau protease), dan enzim yang lain, yaitu
23
dan Niki 1996). Hambatan terhadap enzim bergantung pada reaktivitas
senyawa fenol terhadap sisi rantai asam amino enzim (Rohn et al. 2002).
Diet dan antioksidan eksogen mencegah kerusakan seluler melalui
reaksi yang dilakukan oleh radikal bebas. Ayam yang diberi pakan diet
semisintetik rendah antioksidan menunjukkan penurunan yang nyata stabilitas
eritrosit terhadap H2O2 atau 2,2’-azobis (2-amidinopropan) dihidrokhlorid
(AAPH), tetapi peningkatan pada aktivitas katalase pada hepar, karbonil pada
protein otot tak larut, dan peningkatan oksidasi lemak pada perlakuan
pemanasan pada hepar dibandingkan dengan ayam yang diberi pakan
konvensional. Pada percobaan ini, ayam model menjadi lebih peka terhadap
perubahan oksidatif daripada ayam yang diberi pakan konvensional, yang
ditunjukkan oleh rendahnya pertahanan antioksidan (Young et al. 2002).
Aktivitas antioksidan dapat diukur dengan metode tiosianat dengan cara
melihat jumlah peroksida yang terbentuk pada emulsi selama inkubasi sampel
yang diukur secara spektrofotometri, yaitu mengukur absorbansi pada panjang
gelombang 500 nm. Tingginya nilai absorbansi mengindikasikan tingginya
konsentrasi peroksida (Yildirim et al. 2001). Pengukuran aktivitas antioksidan
dilakukan dengan menggunakan metode tiosianat berdasarkan kemampuan
terbentuknya senyawa-senyawa radikal yang bersifat reaktif. Proses terjadinya
senyawa radikal bebas ini disebabkan oleh oksidasi senyawa asam linoleat
dalam buffer yang diinkubasi pada suhu 40oC selama beberapa jam. Asam
linoleat pada uji ini berperan sebagi substrat yang dioksidasi. Setiap periode
tertentu aborbans hasil oksidasi diukur dengan menggunakan FeCl2 dan
amonium thiosinat (NH4CN). Besi (Fe2+) berperan sebagai mediator
mengkatalisisi peroksidasi lipida telah banyak diketahui, juga berperan
meningkatkan absorbsi dan transport lipida intraseluler (Osborn dan Casimir
2003).
Degradasi isoflavon dan flavonoid dalam saluran pencernaan menjadi
senyawa monofenolat memiliki daya tarik karena beberapa monofenolat
memiliki sifat berefek sebagai antiproliferatif, misalnya senyawa metil
p-hidroksifenolat dapat menghambat sel MCF-7 secara in-vitro (Hendrich et al.
1999). Setelah diabsorbsi pada saluran pencernaan, antioksidan masuk ke
dalam peredaran darah. Ikatan dengan protein menghasilkan pelapisan
substansi yang merupakan kapasitas antioksidan flavonoid. Laporan terakhir
misalnya kemampuan melawan peroksidasi lemak. Ikatannya juga menurun
dengan tidak adanya antioksidan. Pada kejadian ini penambahan aktivitas
intrinsik dari senyawa, metabolisme, ikatan terhadap protein juga menentukan
untuk mempengaruhi efek pemberian flavonoid secara invivo (Arts et al. 2002).
Aktivitas antioksidan ekstrak Pomegranate (Punica granatum)
memproteksi hati terhadap efek tosik CCl4 secara histologi menjadi normal dan
mampu memperbaiki fungsi enzimatik akibat serangan ROS (Murthy et al.
2002). Ekspresi berlebihan antioksidan tidak selalu menghasilkan pertahanan
antioksidan dan bila ditingkatkan kapasitas antioksidan tidak selalu berkorelasi
positif dengan tingkat ketahanan. Dalam hal ini perlu diperhatikan faktor-faktor
yang dapat mempengaruhi kemanjuran dan proteksi antioksidan terhadap
penghapusan oksigen dan juga tekanan lingkungan (Blokhina et al 2003).
Tampaknya sangat beralasan untuk mempertimbangkan antara potensi, risiko,
dan manfaat antioksidan dosis tinggi secara kasus per kasus dan konsumsi
antioksidan supleman tunggal dosis tinggi harus dihindari (Silalahi 2006).
Aktivitas benalu teh sebagai antioksidan yang terkandung dalam ekstrak
ditandai dengan daya mereduksi kaliumferisianida [K3Fe (CN)6], menghambat
oksidasi asam linoleat, kemampuan eliminasi terhadap H2O2 (Leswara dan
Kartin 1998, Santoso 2001, Susmandari 2002). Seduhan daun S. artopurpurea
mengandung antioksidan yang tinggi dan ternyata mampu menurunkan
konsentrasi H2O2. Seduhan tersebut mengandung senyawa penurun risiko
kanker, karena radikal bebas dalam tubuh dapat menimbulkan reaksi berantai
yang menyebabkan kerusakan membran sel, asam nukleat, protein, dan lipid
(Sudihartini 2003). Uji antioksidan benalu teh (Scurrula oortiana) dengan
oksidator 1,1-diphenyl 2-pyrohidrxyl (DPPH), menunjukkan bahwa aktivitas
antioksidan tertinggi ditunjukkan oleh ekstrak metanol dengan daya hambat
sebesar 93.59 ppm (Simanjuntak et al. 2004).
Benalu Teh sebagai Penurun Risiko Kanker
Kanker merupakan hasil proses jangka panjang yang mengakibatkan
efek penyimpangan genetik dan perubahan molekuler yang proses
perubahannya berjalan secara berangsur-angsur. Biasanya diperlukan waktu
lama untuk perubahan dari normal ke peningkatan level puncaknya, displasia,
yaitu invasi dan metastasis secara fenotip. Akumulasi perubahan secara
25
intervensi bidang klinik untuk pence