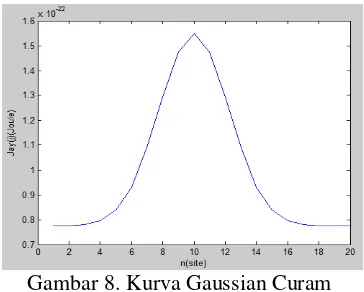
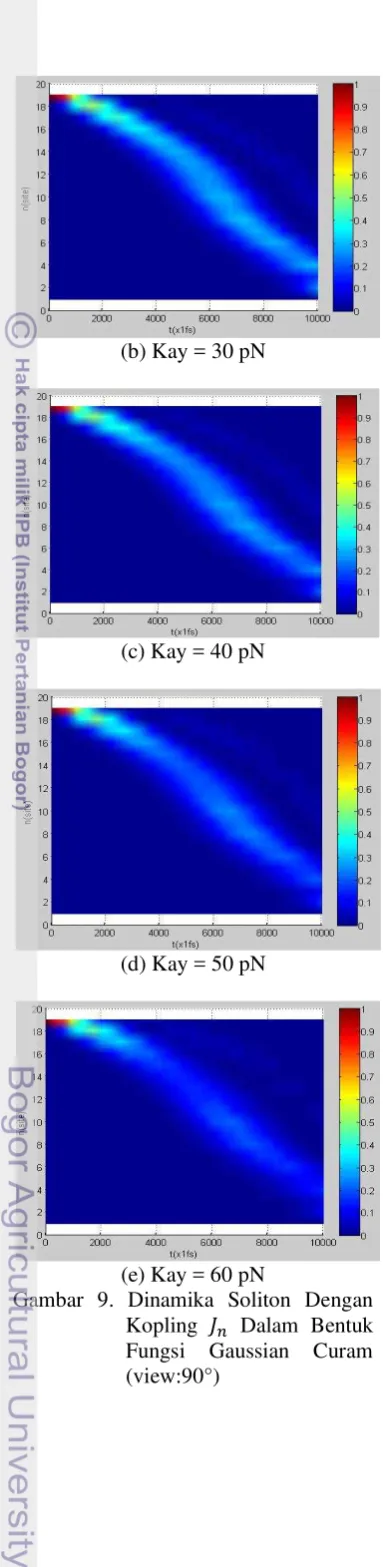
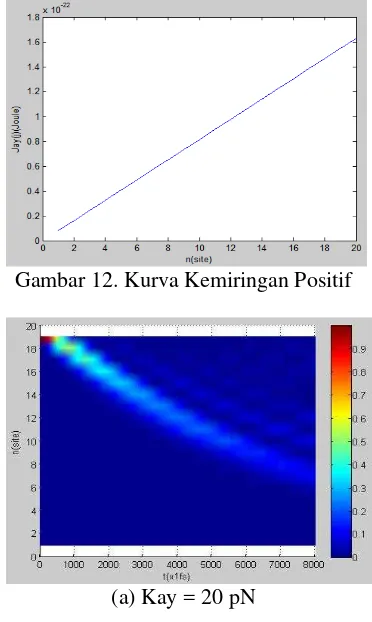
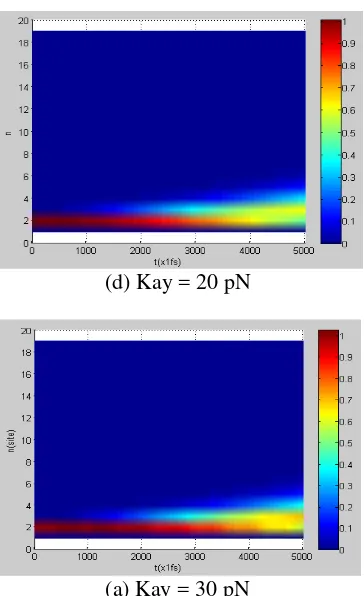
Dinamika Transfer Energi Pada Protein α-Heliks Berdasarkan Ansatz I Model Davydov Dengan Variasi Kopling Antar Peptida
Bebas
45
0
0
Teks penuh
Gambar
+7
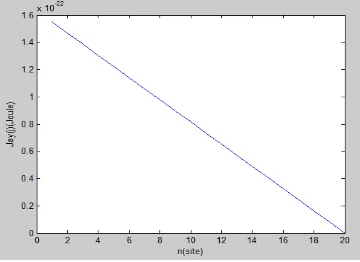
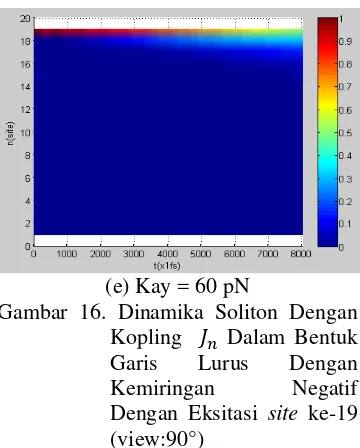
Dokumen terkait
