IDENTIFIKASI
QUANTITATIVE TRAIT LOCI
(QTL) YANG
TERKAIT KARAKTER MORFOLOGI DAN KOMPONEN
HASIL TANAMAN PADI DENGAN MARKA SSR
LINA HERLINA
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa tesis berjudul Identifikasi Quantitative Trait Loci (QTL) yang Terkait Karakter Morfologi dan Komponen Hasil Tanaman
Padi dengan Marka SSR adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, 2014
Lina Herlina
RINGKASAN
LINA HERLINA. Identifikasi Quantitative Trait Loci (QTL) yang terkait karakter
morfologi dan komponen hasil tanaman padi dengan marka SSR Dibimbing oleh: SOBIR dan KURNIAWAN RUDI TRIJATMIKO.
Sejak dipastikan bahwa telah terjadi stagnasi pada produktivitas padi akibat sempitnya keragaman genetik di skala internasional, maka upaya untuk melakukan perbaikan secara genetik untuk menghasilkan varietas-varietas berpotensi hasil tinggi ditempuh dengan gencar untuk melakukan pencarian genotipe-genotipe baru, khususnya melalui pemanfaatan bioteknologi. Aplikasi seleksi berbasis marka molekuler pada tanaman padi juga telah mulai dikembangkan di Indonesia. Salah satu marka molekular yang telah banyak digunakan dalam pemuliaan tanaman adalah Simple Sequence Repeats (SSR).
BIO-148 (Bio-148-Mamo-Dro) adalah galur harapan yang berpotensi daya hasil tinggi. Galur Bio-148 berasal dari persilangan antara varietas Gajah Mungkur (IRAT112) dengan varietas IR64. Padi IRAT112, merupakan padi introduksi dari Kenya (Afrika) yaitu padi lokal jenis tropical japonica sedangkan
IR64 adalah padi indica. Persilangan antara spesies O. sativa ssp japonica dengan O. sativa ssp indica untuk mendapatkan variasi keragaman genetik telah banyak
dilakukan sebagai salah satu alternatif memperluas keragaman genetik padi untuk memecah stagnasi produksi padi.
Penelitian ini bertujuan untuk mendapatkan marka SSR yang berasosiasi dengan posisi QTL yang mengendalikan karakter komponen hasil dan karakter morfologi lainnya pada populasi F2 dari persilangan Bio-148 dan IR64, mengkaji korelasi antara karakter-karakter tersebut, terutama yang nyata kontribusinya terhadap hasil dan komponen hasil pada Galur Bio-148.
Penelitian dilaksanakan dalam jangka waktu 15 bulan dari Februari 2013 sampai April 2014. Persilangan tanaman, pembentukan populasi F2 dan analisis molekuler dilakukan di Rumah Kaca dan Laboratorium Molekuler Balai Besar Litbang Bioteknologi dan Sumberdaya Genetik Pertanian-BB BIOGEN- Cimanggu, Bogor. Penelitian ini meliputi: survei polimorfisme tetua dengan 553 primer SSR, melakukan seleksi marka polimorfik hingga terpilih 24 marka yang mewakili setiap kromosom padi, mengkonfirmasi heterosigositas atas individu F1 terpilih, melakukan karakterisasi fenotipik untuk sifat terkait hasil/komponen hasil terhadap populasi F2 dari turunan F1 hasil persilangan Bio-148 dan IR64, dan mengidentifikasi keberadaan QTL untuk sifat terkait hasil/komponen hasil berdasar 24 marka polimorfik terpilih.
Terdapat 4 karakter yang memiliki pengaruh sangat nyata terhadap hasil biji (Y), yaitu jumlah malai per tanaman (PN), jumlah total anakan per tanaman (TN), total bunga per malai (TSN) dan jumlah biji hampa (UG). Karakter morfologi yang sangat berpengaruh terhadap komponen hasil (TSN) adalah tinggi tanaman (PH), panjang malai (PL), panjng dan lebar daun bendera (FLL dan FLW). Distribusi fenotipik pada populasi F2 memiliki segregasi yang kontinyu, yang berarti karakter-karakter tersebut diwariskan secara kuantitatif.
0.077 – 18.1 % untuk karakter morfologi, dan 5.2-14.2% untuk karakter komponen hasil. Nilai LOD bervariasi antara 2.37 – 8.262 untuk karakter morfologi, sedangkan untuk komponen hasil nilai LOD berkisar 2.236 – 7.283. Pada metode CIM, berhasil dideteksi sebanyak 17 QTL yang tersebar di kromosom yang sama yaitu 2.4,5, 8,9 dan 12 dengan variasi fenotipik berkisar 5.4 – 17.9% dan nilai LOD antara 2.387 – 8.225.
QTL untuk TSN diidentifikasi pada kromosom 4, yaitu tsn.4.1 dengan
LOD di α0.05 = 6,607 (SMR), 7,725 (CIM) dan tsn.4.2 dengan LOD di α0.05 =
4,382 (SMR), kontribusi variasi fenotipik total 23,9 % (SMR) dan 16,4% (CIM). Bio-148 adalah tetua yang berkontribusi alel pada tsn.4.1 yang meningkatkan tanaman. QTL untuk TN diidentifikasi pada kromosom 4 yaitu tn.4 dengan LOD
di α0.05 = 2,674 (SMR) dan 3,215 (CIM) dengan variasi fenotip 6,1% (SMR) dan 7,3% (CIM). IR64 adalah tetua yang memberikan kontribusi alel pada tn.4 yang
meningkatkan jumlah anakan dari 1.587 (SMR) sampai dengan 2,244 (CIM) per tanaman.
SMR dan CIM mendeteksi QTL yang sama untuk PH pada kromosom 4, diidentifikasi sebagai ph.4 dengan LOD di α0.05 = 2,972, kontribusi variasi
fenotipik 6,8%. Bio-148 sebagai tetua yang menyumbang alel untuk tinggi tanaman mencapai 2,783 cm. QTL untuk FLL diidentifikasi pada kromosom 2 ditetapkan sebagai fll.2 dengan LOD di α0.05 = 7.13 (SMR) dan 7,285 (CIM)
dengan variasi fenotipik 15,4% (SMR) dan 15,7% (CIM). IR64 sebagai tetua yang memberikan kontribusi alel pada fll.2 yang menambah panjang daun bendera oleh
2,376 (SMR) sampai 3,94 (CIM) cm. QTL untuk FLW diidentifikasi di kromosom 4 sebagai flw.4.1 dengan LOD di α0.05 = 7,817 (SMR) atau 8,225 (CIM), dan flw.4.2 dengan LOD di α0.05 = 3,425 (SMR), yang memberikan variasi fenotipik
24,5% (SMR) dan 17,6% (CIM). Bio-148 sebagai tetua yang memberikan
1.428mm. Informasi tentang lokasi QTL akan sangat membantu untuk perbaikan genetik Bio-148 dan pada akhirnya untuk meningkatkan hasil padi di Indonesia.
SUMMARY
LINA HERLINA. Identification of Quantitative Trait Loci (QTL) associated with morphological characteristics and yield components of rice plants using SSR markers
Supervised by: SOBIR and KURNIAWAN RUDI TRIJATMIKO.
Since it is certain that there has been a stagnation in the productivity of rice due to the narrowness of the genetic diversity in the international scale, efforts to make improvements genetically to produce varieties of high yielding potential reached by aggressively searching for new genotypes, particularly through the use of biotechnology. Application of molecular marker-based selection in rice plants have also started to be developed in Indonesia. One of the molecular markers that have been widely used in plant breeding is Simple Sequence Repeats (SSR).
BIO-148 (Bio-148-Mamo-Dro) is a potentially promising lines of high yield. Bio-148 strain derived from a cross between the varieties Gajah Mungkur (IRAT112) with IR64. IRAT112 rice is the introduction of Kenya (Africa) types of tropical japonica rice locally while IR64 is indica rice. Crosses between
species O. sativa ssp japonica and O. sativa ssp indica to get a variation of
genetic diversity have been carried out as an alternative to broaden the genetic diversity of rice to break up stagnation in rice production.
This study aims to obtain SSR markers associated with QTL of yield components and morphological characters in F2 populations from crosses between Bio-148 and IR64, examines the correlation between these characters, especially which have significant correlation to yield and yield components in Bio-148 strain.
The experiment was conducted in a period of 15 months from February 2013 until April 2014. Crosses plants, the formation of the F2 population and molecular analysis performed in Greenhouse and Laboratory of Molecular Research Center for Agricultural Biotechnology and Genetic Resources
BIOGEN-BB-Cimanggu, Bogor.
This research includes: parental polymorphism survey with 553 SSR primer, the selection of polymorphic markers of 24 markers which selected to represent each rice chromosomes, confirming heterozygosity over F1 individuals selected, perform phenotypic characterization related to the nature of the results / outcome component of the F2 population derived from the F1 results crosses Bio-148 and IR64, and identify the presence of QTL for traits related to outcomes / results based on 24 components selected polymorphic markers.
There are 4 characters that have a significant effect on grain yield (Y), i.e. the number of panicles per plant (PN), the total number of tillers per plant (TN), total flowers per panicle (TSN) and the number of unfilled grain/seeds (UG). Morphological characters which were very influential on yield components (TSN) i.e. plant height (PH), panicle length (PL), flag leaf length and flag leaf width (FLL and FLW).
have been identified. Based on SMR, 16 QTLs were detected on chromosomes 2, 4, 5, 8, 9 and 12 with phenotypic variation ranging from 0.077 - 18.1% for morphological characters, and 5.2-14.2% for the yield component characters. LOD value varies between 2:37 - 8,262 for morphological characters, while the results of the component values for the LOD range 2236-7283. In the CIM method, a total of 17 QTL were detected scattered in the same chromosome are 2.4,5, 8.9 and 12 with phenotypic variation ranging from 5.4 - 17.9% and LOD score between 2387-8225.
QTL for TSN were identified on chromosome 4 designated as tsn.4.1 with LOD at α0.05 = 6.607(SMR), 7.725(CIM) and tsn.4.2 with LOD at α0.05 = 4.382
(SMR), contributed phenotypic variation totally 23.9% (SMR) and 16.4% (CIM). Bio-148 was a parent which contributed allele at tsn.4.1 which increased spikelet
number by 24,258 (CIM) up to 28.49 (SMR) spikelet per panicle. QTL for PN were identified on chromosome 4 designated as pn.4 with LOD at α0.05 = 2.236
(SMR) and 3.96(CIM) contributed phenotypic variation 5.2% (SMR) and 9.1%(CIM). IR64 was a parent which contributed allele at pn.4 which increased
panicle number by 1.157(SMR) up to 2.16 (CIM) per plant. QTL for TN were identified on chromosome 4 designated as tn.4with LOD at α0.05 = 2.674 (SMR)
and 3.215(CIM) contributed phenotypic variation 6.1% (SMR) and 7.3%(CIM). IR64 was a parent which contributed allele at tn.4 which increased tiller number
by 1.587(SMR) up to 2.244 (CIM) per plant.
7.285(CIM) contributed phenotypic variation 15.4% (SMR) and 15.7%(CIM). IR64 was a parent which contributed allele at fll.2 which increased flag leaf length
by 2.376 (SMR) up to 3.94 (CIM) cm. QTL for FLW were identified on chromosome 4 designated as flw.4.1 with LOD at α0.05 = 7.817 (SMR) or
8.225(CIM), and flw.4.2 with LOD at α0.05 = 3.425 (SMR), which contributed
phenotypic variation totally 24.5% (SMR) and 17.6%(CIM). Bio-148 was a parent which contributed allele at flw.4 which increased flag leaf width by 0.101
mm(SMR) up to 0.137mm (CIM). QTL for PM were identified on chromosome 12 and chromosome 2 designated as pl.12. with LOD at α0.05 = 3.539 (SMR) or
3.729(CIM), and pl.2 with LOD 3.09 (CIM). Bio-148 contributed phenotypic
variation 7.9% (SMR) and totally 15.2%(CIM). Bio-148 was parent which contributed allele at pl.12 which increased panicle length 0.501 (SMR) up to
0.694 (CIM) cm. While in chromosome 2, IR64 contributed allele at pl.2 which
increased panicle length up to 1.428mm. The information on the QTL locations would be helpful for genetic improvement of Bio-148 and ultimately for increasing rice yield in Indonesia.
© Hak Cipta Milik IPB, Tahun 2014
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkansumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
IDENTIFIKASI
QUANTITATIVE TRAIT LOCI
(QTL) YANG
TERKAIT KARAKTER MORFOLOGI DAN KOMPONEN
HASIL TANAMAN PADI DENGAN MARKA SSR
LINA HERLINA
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Pemuliaan dan Bioteknologi Tanaman
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Judul Tesis: Identifikasi Quantitative Trait Loci (QTL) Yang Terkait Karakter
Morfologi dan Komponen Hasil Tanaman Padi dengan Marka SSR Nama : Lina Herlina
NIM : A253120191
Disetujui oleh
Komisi Pembimbing
Prof Dr Ir Sobir, MSi Dr Ir Kurniawan Rudi Trijatmiko, MSi Ketua Anggota
Diketahui oleh
Ketua Program Studi Dekan Sekolah Pascasarjana Pemuliaan dan Bioteknologi Tanaman
Dr Ir Yudiwanti Wahyu Endro Kusumo, MSi Dr Ir Dahrul Syah, MSAgr
PRAKATA
Alhamdulillah, segala puji bagi Allah. Tiada sekutu bagi-Nya. Dia-lah yang Maha berkehendak atas segala sesuatu. Sholawat dan salam kepada hamba dan utusan-Nya, Muhammad SAW, penutup para nabi dan rasul. Penelitian ini dilaksanakan dari bulan Agustus 2012 sampai April 2014 dengan judul Identifikasi Quantitative Trait Loci (QTL) Yang Terkait Karakter Morfologi dan
Komponen Hasil Tanaman Padi dengan Marka SSR. Penulis menyampaikan terima kasih kepada:
1. Mbah Nu, atas pengabdiannya yang senantiasa, dan tak bisa dilukiskan dalam kata-kata.
2. Prof. Dr. Ir. Sobir, MSi sebagai Ketua Komisi Pembimbing yang telah sangat berbaik hati dalam membimbing penulis melalui keluasan ilmu, wawasan maupun pengetahuan selama penulis menjalankan studi di Mayor Pemuliaan dan Bioteknologi Tanaman, serta senantiasa berkenan meluangkan waktu di sela-sela jadwal kesibukan yang sangat padat untuk selalu memberikan bimbingan, arahan, nasehat serta saran, sehingga atas support beliau jualah penulis berhasil lulus dalam studi ini.
3. Dr. Ir. Kurniawan Rudi Trijatmiko, MSi sebagai anggota Komisi Pembimbing, atas bimbingan, arahan, bantuan serta support-nya sehingga
penulis berkesempatan untuk mempelajari dan mendalami Ilmu Bioteknologi dan Genetika Molekuler secara nyata. Bahkan meski terkendala oleh jarak dan kesibukan beliau yang sangat padat, masih tetap bersemangat dan selalu memberikan banyak tambahan ilmu, wawasan maupun pengetahuan baik melalui diskusi maupun melalui paper-paper jurnal yang sangat membantu penulis dalam memahami lebih banyak tentang bioteknologi tanaman. Dan atas support beliau jualah penulis akhirnya mampu menyelesaikan studi di pascasarjana ini dengan baik. 4. Dr. Ir. Desta Wirnas, MSi selaku dosen penguji, atas ilmu dan wawasan
yang diberikan, yang sangat berarti bagi penulis, khususnya karena telah membuka pemahaman yang lebih mendalam dan komprehensif kepada penulis tentang pemuliaan untuk karakter kuantitatif.
5. Dr. Ir. Yudiwanti Wahyu EK, MSi dan Dr. Ir. Dewi Sukma, MSi, sebagai Ketua dan Wakil Program Studi Pemuliaan dan Bioteknologi Tanaman, atas bantuan dan dukungan kemudahan akses serta pelayanan akademik yang sangat baik sehingga memacu semangat penulis untuk segera menyelesaikan studi tepat waktu.
6. Seluruh jajaran staf administrasi di Mayor Pemuliaan dan Bioteknologi Tanaman (Ibu Mimin, Teh Neng, dan Pak Udin) atas pelayanan administrasi yang sangat baik selama penulis menjadi mahasiswa di almamater tersebut.
7. Badan Litbang Pertanian, Kementerian Pertanian, atas kesempatan beasiswa yang diberikan sehingga penulis dapat menempuh studi di pascasarjana IPB.
9. Teman-teman di Mayor Pemuliaan dan Bioteknologi Tanaman Pascasarjana IPB tahun angkatan 2012 atas kebersamaan yang menyenangkan
10.Putra-putraku tercinta, Asyaddu Hubbanlillah dan Adkhilna Firahmatillah, atas ‘kemewahan dunia’ yang membuat mentari di hati penulis selalu bersinar.
11.Suamiku, Bonjok Istiaji, MSi, atas cinta, kasih, sayang, pengertian, kesabaran, dukungan, doa serta perhatian yang tulus dan tanpa mengenal waktu sehingga penulis dapat memperoleh keluasan dan kedalaman ilmu, dan berhasil menyelesaikan studi ini dengan baik.
12.Keluarga tercinta yang telah memberikan doa, semangat, dan pengorbanannya dengan tulus: Ibunda Afifah Sugeng Wahyono beserta saudara-saudaraku tercinta di Semarang; dan Ibu Saban Adi Sumarto beserta saudara-saudaraku di Magelang.
Semoga karya ilmiah ini bermanfaat. Mohon maaf atas segala kekurangan.
Bogor, 2014
DAFTAR ISI
DAFTAR TABEL ii
DAFTAR GAMBAR ii
DAFTAR LAMPIRAN iii
1 PENDAHULUAN 1
Latar belakang 2
Perumusan Masalah 2
Tujuan Penelitian 3
Manfaat Penelitian 3
Ruang Lingkup Penelitian 3
2 KARAKTERISASI MORFOLOGI DAN AGRONOMI YANG TERKAIT HASIL PADA TANAMAN PADI POPULASI SEGREGASI PERSILANGAN BIO-148 DAN IR64
Pendahuluan 4
Tujuan Penelitian 6
Bahan dan Metode 6
Hasil 9
Pembahasan 17
Simpulan 23
3 ANALISIS MOLEKULER DAN DETEKSI QTL YANG TERKAIT HASIL PADA TANAMAN PADI POPULASI F2 SEGREGASI PERSILANGAN BIO-148 DAN IR64
Pendahuluan 23
Tujuan Penelitian 25
Bahan dan Metode 25
Hasil 28
Pembahasan 41
Simpulan 45
4 PEMBAHASAN UMUM 45
5 SIMPULAN DAN SARAN 57
DAFTAR PUSTAKA 58
LAMPIRAN 64
DAFTAR TABEL
1 Karakterisasi fenotipik populasi F2 11
2 Korelasi antara karakter morfologi terhadap hasil dan komponen hasil 14
3 Kelayakan model persamaan regresi 16
4 Pengaruh parsial tiap variabel bebas terhadap hasil 16 5 Daftar primer SSR polimorfik antara IR64 dan Bio-148 31
6 Hasil survei polimorfisme tetua dengan 553 primer 32
7 Segregasi 24 marka SSR terpilih pada populasi F2 35
8 QTL untuk Jumlah Biji Isi (FG) 36
9 QTL untuk Jumlah Spikelet per Malai (TSN) 37
10 QTL untuk Total Anakan (TN) 37
11 QTL untuk Jumlah Malai per Rumpun (PN) 38
12 QTL untuk Tinggi Tanaman (PH) 38
13 QTL untuk Panjang Daun Bendera (FLL) 38
14 QTL untuk Panjang Malai (PL) 39
15 QTL untuk Eksersi Malai Tak Sempurna (IPE) 39
16 QTL untuk Lebar Daun Bendera (FLW) 40
17 QTL untuk Sudut Daun Bendera (FLA) 40
18 QTL untuk Bobot 100 biji (100GW) 40
19 Distribusi alel asal Bio-148 dan IR64 di F2 44
20 Pola kecenderungan frekuensi fenotipik segregan di F2 terhadap tetua 45
DAFTAR GAMBAR
1 Bagan alir penelitian 4
2 Populasi F2 dari hasil pindah tanam benih umur 3 minggu ke dalam ember-ember percobaan di rumah kaca
7
3 Variasi panjang sungut (awn) pada individu di populasi F2 8
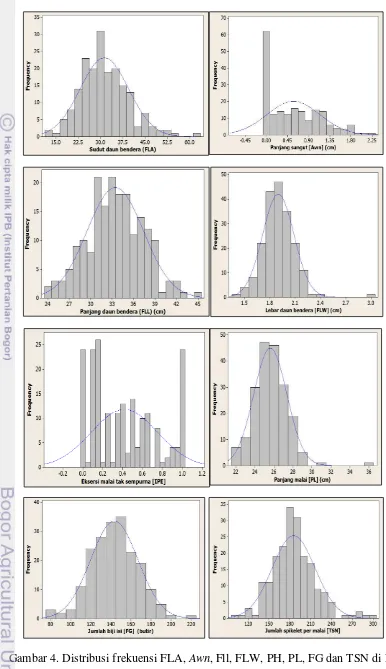
4 Distribusi frekuensi FLA, Awn, Fll, FLW, PH, PL, FG dan TSN di F2 12
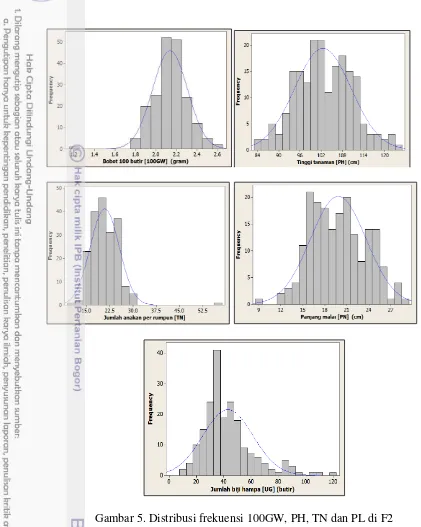
5 Distribusi frekuensi 100GW, PH, TN dan PL di F2 13
6 Infeksi patogen pada eksersi malai yang tidak sempurna 18 7 Contoh survei polimorfisme tetua pada seleksi tahap 1 27 8 Contoh survei polimorfisme tetua pada seleksi tahap 2. Lingkaran
ungu pada posisi marka, menandai ukuran terpilih untuk marka yang mewakili kromosom lengan pendek; lingkaran hijau pada posisi marka menandai ukuran terpilih untuk marka yang mewakili kromosom lengan panjang
29
9 Konfirmasi heterosigositas F1 terpilih dengan marka polimorfik 33 10 Contoh distorsi segregasi alel-alel individu F2 pada pengujian dengan
marka RM402
DAFTAR GAMBAR (lanjutan)
11 Contoh hasil Analisis QTL beberapa karakter morfologi dan
komponen hasil padi dengan metode SMR. 42
12 Contoh hasil Analisis QTL beberapa karakter morfologi dan komponen hasil padi dengan metode CIM
43
DAFTAR LAMPIRAN
1 Daftar marka terpilih yang digunakan untuk analisis QTL pada populasi F2
65
2 QTL yang dideteksi pada populasi F2 berdasar SMR 66
3 QTL yang dideteksi pada populasi F2 persilangan Bio-148/IR64 berdasar CIM
67
4 Permutasi untuk jumlah biji hampa berdasar SMR dan CIM 68
5 Deskripsi karakter varietas IR64 (BB PADI) 69
1
PENDAHULUAN
Latar Belakang
Pemuliaan padi untuk mendapatkan varietas unggul berdaya hasil tinggi telah cukup lama menjadi prioritas di Indonesia (Susanto et.al., 2003). Sebagai
negara dengan penduduk terbesar ke-4 di dunia, dengan total jumlah penduduk lebih dari 251.160.254 jiwa, maka pemenuhan terhadap kebutuhan pangan yang selalu meningkat setiap tahun menjadi agenda wajib yang harus dihadapi pemerintah. Untuk itu, kebijakan di sektor pertanian menempatkan swasembada pangan melalui peningkatan produksi padi sebagai salah satu target utama dalam pembangunan pertanian 2010-2014 (DKP 2010).
Sejak dipastikan bahwa telah terjadi stagnasi pada produktivitas padi akibat sempitnya keragaman genetik (Yamamoto et al. 2010, Peng et al. 2009),
maka mulai tahun 1980 pemuliaan padi di Indonesia mencoba melakukan terobosan melalui pengembangan padi tipe baru yang didasari pemikiran peningkatan hasil melalui rekayasa arsitektur tanaman (Susanto et al 2003).
Pemanfaatan marka molekular sebagai alat bantu seleksi lebih menguntungkan daripada seleksi secara fenotipik konvensional saja (Azrai 2005). Seleksi berdasarkan fenotipik tanaman di lapang mempunyai beberapa kelemahan, antara lain perlu waktu cukup lama, kesulitan memilih dengan tepat gen-gen yang menjadi target seleksi untuk diekspresikan pada sifat-sifat morfologi atau agronomi, frekuensi individu yang diinginkan rendah, dan fenomena pautan gen dengan sifat yang tidak diinginkan sulit dipisahkan saat melakukan persilangan (Lamadji et al. 1999)
Aplikasi seleksi berbasis marka molekuler pada tanaman padi juga telah mulai dikembangkan di Indonesia (Miftahuddin et al. 2010). Strategi yang kini
banyak dilakukan adalah mengidentifikasi marka yang berasosiasi dengan QTL (Quantitave Traits Loci) dari karakter yang ingin diperbaiki, yaitu dengan
melakukan analisis pautan dan pemetaan QTL dengan menggunakan data kuantitatif maupun data molekuler.
BIO-148 (Bio-148-Mamo-Dro) adalah galur harapan yang berpotensi daya hasil tinggi- dengan potensi gabah kering giling yang melebihi varietas Ciherang (Suratmini et al. 2010). Varietas ini memiliki bobot 1000 butir yang relatif tinggi
serta tanaman relatif tahan terhadap penyakit blast (Trijatmiko et al. 2001). Selain
spikelet per malai yang lebih padat, Galur Bio-148 juga memiliki jumlah anakan yang tinggi, daun bendera lebih luas, tanaman lebih tinggi, malai lebih panjang, umur berbunga dan umur panen yang lebih cepat (Suratmini et al. 2010).
Sebagai bahan genetik yang memiliki perbedaan latar belakang genetik padi indica dan japonica, Galur Bio-148 berpotensi sebagai sumber genetik dalam
QTL untuk karakter morfo-agronomi dan komponen hasil pada galur Bio-148 tersebut. Oleh karenanya penelitian ini menginisiasi kegiatan tersebut.
Perumusan Masalah
Produktivitas tanaman merupakan puncak dari berbagai proses yang terjadi dalam siklus hidup tanaman yang sangat dipengaruhi oleh setiap fase pertumbuhan dan perkembangan (Khanna-Chopra 2000). Untuk mengetahui secara tepat karakter tanaman yang berperan nyata dalam menyumbang kenaikan hasil pada tanaman padi, maka perlu dilakukan analisis untuk mendapatkan informasi karakter-karakter apa saja yang memiliki hubungan erat terhadap hasil dengan melakukan karakterisasi atau pengamatan terhadap fenotipik tanaman, yang selanjutnya diuji dalam analisis korelasi dan regresi.
Hasil adalah visualisasi akhir dari proses yang kompleks, yang melibatkan kerja dari sejumlah gen yang mengendalikan sifat-sifat tertentu, yang berpengaruh terhadap besar-kecilnya ekspresi pada tataran fenotipik, baik untuk karakter morfologi maupun agronomi. Gen-gen yang secara spesifik mengendalikan sifat yang terkait dengan hasil dikenal dalam kelompok gen-gen yang terdapat dalam lokus sifat kuantitatif atau quantitative trait loci (QTL). Lokus sifat kuantitatif
yang secara khusus mengendalikan karakter terkait komponen hasil pada Galur Bio-148 masih belum diketahui.
Dalam rangka memberdayakan potensi genetik khususnya yang terkait dengan hasil tinggi pada galur tersebut, maka Galur Bio-148 dapat dijadikan bahan pemetaan genetik untuk alasan tersebut. Letak lokus kuantitatif dapat dideteksi dengan marka SSR yang telah terbukti mudah aplikasinya, tidak memerlukan template DNA dalam jumlah besar dan tingkat kemurnian tinggi,
dapat mendeteksi alel, kodominan, polimorfisme tinggi, melimpah dalam genom, relatif murah dan terjangkau biayanya namun dengan kualitas output yang akurat
dan dapat diandalkan.
Penelitian ini oleh karenanya akan mencakup kegiatan karakterisasi fenotipik terhadap morfo-agronomi tanaman padi dan analisis molekuler dengan marka SSR terhadap individu di populasi F2, yang selanjutnya data analisis yang diperoleh dari analisis fenotipik dan analisis molekuler dapat dikombinasikan untuk melakukan analisis QTL. Dengan demikian diharapkan dapat diidentifikasi marka-marka SSR apa saja yang terkait dengan karakter hasil atau komponen hasil pada populasi tersebut.
Tujuan Penelitian
Penelitian ini bertujuan untuk
1) Mengkaji korelasi antara karakter-karakter morfologi dan agronomi yang memiliki kontribusi nyata terhadap hasil dan komponen hasil pada populasi F2 persilangan Galur Bio-148 dengan IR64.
Manfaat Penelitian
Manfaat penelitian adalah :
1. Marka yang diperoleh dapat digunakan untuk mencari informasi gen-gen QTL yang terkait karakter hasil dan komponen hasil berdasar marka-marka terpaut. Gen-gen tersebut selanjutnya dapat diklon dan digunakan untuk pengujian lebih lanjut khususnya melihat ekspresinya di tingkat fenotipik.
2. Marka juga dapat digunakan dalam program pemuliaan melalui pendekatan marker assisted selection (MAS).
Ruang Lingkup Penelitian
Penelitian ini meliputi: pembentukan populasi F2, seleksi marka polimorfik terhadap tetua dengan 553 primer SSR, seleksi lanjut marka hingga terpilih 24 marka SSR yang mewakili setiap kromosom padi, mengkonfirmasi heterosigositas atas individu F1 terpilih, melakukan karakterisasi fenotipik untuk sifat terkait hasil/komponen hasil terhadap populasi F2 dari turunan F1 hasil persilangan Bio-148 dan IR64, dan mengidentifikasi keberadaan QTL untuk sifat terkait hasil/komponen hasil berdasar 24 marka polimorfik terpilih.
Percobaan 1 :
Karakterisasi Morfologi dan Agronomi yang Terkait Hasil pada Tanaman Padi Populasi Segregasi Persilangan Bio148 dan IR64
1. Pembentukan populasi F2
2. Melakukan karakterisasi fenotipik untuk karakter morfologi dan agronomi pada populasi F2 hasil persilangan Bio-148 x IR64.
3. Melakukan analisis korelasi dan regresi terhadap data fenotipik yang diperoleh
Percobaan 2 :
Analisis Molekuler dan Deteksi QTL yang Terkait Hasil pada Tanaman Padi Populasi F2 Segregasi Persilangan Bio148 dan IR64
1. Melakukan seleksi marka dua tahap terhadap kedua tetua materi persilangan
2. Melakukan konfirmasi heterogenisitas terhadap genotipe F1 terpilih 3. Melakukan analisis PCR terhadap populasi F2 dengan 24 marka terseleksi 4. Melakukan analisis linkage dan QTL terhadap 24 marka SSR untuk
mendeteksi QTL yang terkait hasil/komponen hasil
Bagan Alir Penelitian
Gambar 1. Bagan alir penelitian
2 KARAKTERISASI MORFOLOGI DAN AGRONOMI YANG
TERKAIT HASIL PADA TANAMAN PADI POPULASI
SEGREGASI PERSILANGAN BIO-148 DAN IR64
Pendahuluan
terbaik yang tersedia dan hasil teoritis maksimal. Hal ini memunculkan pertanyaan apakah kesenjangan ini bisa dipersempit dengan perbaikan genetik tanaman.
Saat ini, akibat lazimnya inkompatibilitas pada subpopulasi, padi dunia umumnya diproduksi dari inbred varitas yang telah dikembangkan secara ekslusif melalui persilangan antar aksesi dari sub populasi yang sama atau antar sub populasi yang sekerabat (tropical japonica x temperate japonica; indica x australiens). Inkompatibilitas yang terdapat pada subpopulasi merupakan
rintangan utama yang menyebabkan sangat sulit memperoleh rekombinan yang fertil secara acak dari hasil persilangan lintas-subpopulasi (misalnya O.sativa
ssp.indica x O.sativa ssp.japonica) (Harushima et al. 2002). Namun demikian
perkembangan penelitian molekuler kini memungkinkan para pemulia untuk mengidentifikasi area pada genom yang berasosiasi dengan komponen khusus dari fenotipe dan menentukan tetua yang berkontribusi atas alel yang diinginkan pada lokus tertentu (Cheema et al. 2008).
Salah satu galur yang dikembangkan oleh Balai Besar Bioteknologi dan Sumberdaya Genetik Pertanian (BB BIOGEN) adalah BIO-148 (Bio-148-Mamo-Dro). Galur ini dikembangkan dari hasil persilangan antara varietas IR64 dengan Gajah Mungkur (IRAT112) (Putra 2013) dalam suatu program pemuliaan padi varietas unggul yang tahan cekaman kekeringan. Padi IRAT112, merupakan padi introduksi dari Kenya (Afrika) yaitu padi lokal jenis tropical japonica yang di
Indonesia dilepas tahun 1994 sebagai varietas Gajah Mungkur (BBPTP 2008). Padi IRAT sering dijadikan sebagai tetua donor dalam kegiatan pemuliaan untuk toleransi terhadap cekaman kekeringan, misalnya IRAT109 (Liu et al. 2010).
Varietas Gajah Mungkur pun di Indonesia sering digunakan sebagai tetua donor (donor parent) untuk toleransi terhadap kekeringan (Effendi 2008, Trijatmiko et al. 2001).
Sebagai bahan genetik yang memiliki perbedaan latar belakang genetik padi indica dan tropical japonica, Galur Bio-148 berpotensi sebagai sumber
genetik dalam kegiatan perakitan variatas unggul melalui pemuliaan, khususnya sebagai bahan pemetaan genetik untuk potensi hasil. Karakter-karakter unggul pada Bio-148 merupakan kontribusi yang menguntungkan dari kedua tetuanya, yaitu IR64 dan Gajah Mungkur. Persilangan intraspesies padi untuk mendapatkan variasi keragaman genetik telah banyak dilakukan sebagai salah satu alternatif memperluas keragaman genetik padi untuk memecah stagnasi produksi padi (Mei
et al. 2005, Jin et al. 2013). Padi tropical japonica belum lama ini dilaporkan
memiliki gen SPIKE (SPIKELET NUMBER) yang berpotensi untuk meningkatkan hasil dan introgresi gen tersebut kepada padi indica dilaporkan
mampu meningkatkan hasil mencapai 30% (Fujita et al. 2013).
terkait proses identitifikasi spesies untuk keperluan koleksi. Selain itu, karakterisasi biasanya juga menjadi prosedur rutin yang dilakukan dalam rangka mempertahankan kemurnian koleksi plasma nutfah yang kita miliki.
Dalam penelitian ini, untuk mengetahui karakter-karakter yang sangat erat hubungannya maupun kontribusinya terhadap hasil dan menentukan karakter komponen hasil, maka sebagai langkah awal dipandang perlu melakukan karakterisasi fenotipik terhadap individu di F2. Selanjutnya informasi yang diperoleh akan dapat dimanfaatkan untuk tahapan pemuliaan selanjutnya.
Tujuan Penelitian
Penelitian ini bertujuan untuk mengetahui karakter morfologi dan agronomi yang terkait erat dengan hasil pada tanaman padi di populasi F2 hasil persilangan antara galur Bio-148 dan IR64.
Bahan dan Metode
Penelitian dilaksanakan di Laboratorium Biologi Molekuler, Balai Besar Bioteknologi dan Sumberdaya Genetika Pertanian, Bogor dari bulan Februari 2013 sampai Mei 2014. Bahan tanaman terdiri dari tetua 1 (P1) Galur Bio-148, tetua 2 (P2) varietas IR64, F1 (Galur Bio-148 x IR64) masing-masing sebanyak 10 tanaman dan tanaman F2 sebanyak 200 tanaman.
Pembentukan Populasi F2
Materi persilangan dipersiapkan dengan mensinkronkan waktu berbunga antara tetua jantan (IR64) dan tetua betina (Bio-148). Setelah induk persilangan memasuki fase pembungaan (ditandai dengan munculnya malai padi sekitar sepertiga bagian), dilakukan pembuangan anter IR64 (emaskulasi) menggunakan bantuan alat penyedot anter (vacuum pump). Proses emaskulasi ini dilakukan pada
sore hari sekitar jam 17.00-19.00 dan bulir padi yang berada pada bagian paling atas dan bagian paling bawah malai dibuang terlebih dahulu untuk menghindarkan kemungkinan terjadinya penyerbukan sendiri (selfing). Bulir padi pada malai yang
sudah diemaskulasi tersebut ditutup dengan kertas minyak dan dibiarkan selama semalam.
Esoknya, pot tetua betina yang sudah diemaskulasi diletakkan berdekatan dengan tetua jantan sebagai sumber polen untuk memudahkan proses penyerbukan berlangsung. Penyerbukan dilakukan dengan cara menggoyangkan malai dari tetua jantan sebagai sumber polen tepat pada malai tetua betina yang sudah dipersiapkan untuk menerima polen. Malai-malai tetua betina yang sudah diserbuki tersebut ditutup kembali menggunakan kertas minyak sampai terbentuk dan matangnya biji-biji F1.
sekitar 3 minggu dipindahkan ke ember percobaan yang berisi tanah dan pupuk yang sesuai dengan kebutuhan hara untuk pertumbuhan tanaman padi (Gambar 2). Setiap satu ember ditanami satu rumpun yang berasal dari satu benih, sampai diperoleh sebanyak 200 ember. Selanjutnya tanaman dipelihara sebagai individu F2 yang akan diamati karakter fenotipiknya. Pemeliharaan tanaman padi meliputi penyiraman, pengendalian hama dan penyakit, serta pemupukan NPK 1:1:1 dengan dosis 0.8 g/tanaman diberikan pada saat 30 dan 60 hari setelah tanam.
Gambar 2. Populasi F2 dari hasil pindah tanam benih umur 3 minggu ke dalam ember-ember percobaan di rumah kaca
Pengamatan Karakter Fenotipik
Pengamatan fenotipik dilakukan terhadap kedua tetua (Bio-148 dan IR64) dan 200 individu dalam populasi F2 (selfing F1). Pengamatan fenotipik bertujuan
untuk mendapatkan informasi normalitas atau tipe sebaran frekuensi individu pada populasi F2 untuk setiap karakter yang diamati. Pengamatan fenotipik tersebut terdiri dari : panjang dan lebar daun bendera, tinggi tanaman, panjang eksersi malai, panjang malai, panjang sungut (awn), jumlah anakan, jumlah malai
per tanaman, jumlah bunga per malai, jumlah biji hampa, jumlah biji isi, bobot 100 biji, bobot biji per rumpun.
Adapun kriteria pengamatan antara lain :
1. Tinggi tanaman, diukur dalam cm dari pangkal batang sampai ujung malai tertinggi
2. Panjang daun bendera, diukur dalam cm.
3. Lebar daun bendera, diukur dalam cm pada bagian daun yang terlebar. 4. Jumlah anakan per rumpun, dihitung setelah fase pembungaan penuh. 5. Jumlah malai per rumpun, dihitung setelah fase pembungaan penuh.
6. Jumlah biji isi, dihitung setelah panen dan biji dikeringkan, dengan cara menekan biji.
7. Jumlah biji hampa, dihitung setelah panen dan biji dikeringkan, dengan cara menekan biji.
9. Sudut daun bendera diukur ketika tanaman sudah membentuk malai secara sempurna. Sudut yang terbentuk antara daun bendera dengan poros malai utama diukur besarnya menggunakan busur derajat.
10. Eksersi malai tak sempurna, ditentukan berdasar rasio dengan membandingkan jumlah malai yang eksersinya tidak sempurna (sebagian malai masih terbungkus/tertutup oleh pelepah daun bendera) dengan malai yang eksersinya penuh (malai yang keluar sempurna sampai terlihat ruas internode di dekat leher malai) untuk setiap individu tanaman di populasi F2.
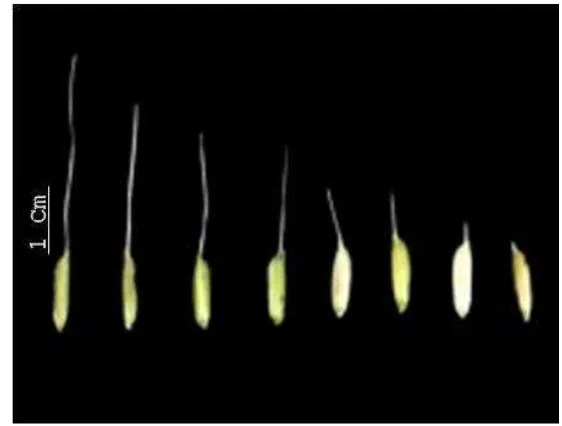
11. Panjang sungut/bulu pada gabah, diukur pada biji yang terletak di apikal/ujung malai, dipilih yang terpanjang dari setiap malai (Gambar 3).
Gambar 3. Variasi panjang sungut (awn) pada individu di populasi F2
12. Panjang malai, diukur dalam cm, mulai leher sampai ujung malai.
13. Bobot 100 butir, ditimbang dalam gram dari 100 butir secara random setelah biji dikeringkan
14. Hasil biji, ditentukan berdasar bobot total biji yang isi yang dihasilkan per satu rumpun (satu individu).
Data yang diperoleh merupakan data fenotipik yang akan dianalisis lebih lanjut.
Analisis Normalitas, Korelasi dan Regresi
Data fenotipik yang diperoleh diuji normalitas sebarannya. Pengujian normalitas didasarkan pada Uji Kolmogorov-Smirnov dengan software yang terintegrasi pada program Qgene 4.3.8. Konsep dasar dari normalitas Kolmogorov Smirnov adalah membandingkan distribusi data (yang akan diuji normalitasnya) dengan distribusi normal baku. Distribusi normal baku adalah data yang telah ditransformasikan ke dalam bentuk Z-Score dan diasumsikan normal. Jadi uji normalitas Kolmogorov Smirnov adalah uji beda antara data yang diuji normalitasnya dengan data normal baku.
dengan data normal baku, berarti data tersebut tidak normal. Lebih lanjut, jika signifikansi di atas 0,05 maka berarti tidak terdapat perbedaan yang signifikan antara data yang akan diuji dengan data normal baku, artinya data yang kita uji normal.
Analisis korelasi yang diterapkan terhadap karakter-karakter fenotipik utamanya bertujuan untuk mendapatkan informasi ada/tidaknya hubungan dan arah hubungan (positif/negatif) dari karakter morfo-agronomi terhadap hasil dan atau komponen hasil. Koefisien korelasi dihitung dengan bantuan program dalam MSExcel, sedangkan untuk menguji nyata-tidaknya kontribusi karakter terhadap hasil maupun komponen hasil, dihitung nilai t-hitung yang dibandingkan dengan t-tabel pada taraf α = 0.05, dimana t hitung diperoleh berdasar formula :
dimana t = nilai t hitung, . r =koefisien korelasi Pearson, . n=jumlah sampel.
Berdasarkan hasil analisis korelasi, kemudian persamaan regresi disusun untuk mengetahui karakter yang memiliki kontribusi nyata langsung terhadap hasil. Pemilihan variabel bebas didasarkan pada analisis korelasi yang dilakukan sebelumnya. Faktor-faktor yang berhubungan dengan hasil padi dapat diduga menggunakan analisis regresi linier berganda, dengan persamaan sebagai berikut:
Y = α + β1 X1+ β2 X2 + β3 X3 + ………. + βm Xn
Variabel hasil yang merupakan variable tak bebas (Y) pada penelitian ini adalah hasil per rumpun. Variabel bebas (X) yang digunakan untuk menduga faktor-faktor yang berhubungan dengan hasil per rumpun pada percobaan ini terdiri dari : bobot 100 biji/100GW (X1), jumlah biji isi/FG (X2), sudut daun
Analisis fenotipik karakter morfologi dan komponen hasil pada populasi F2
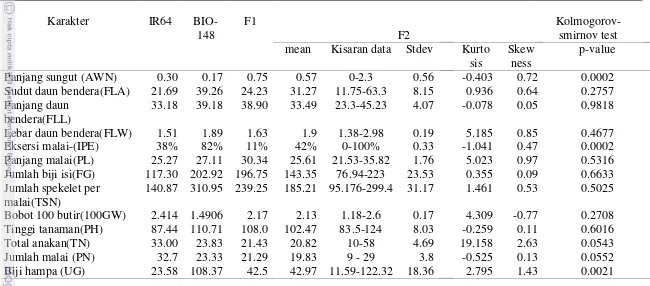
Variasi fenotipik yang diperlihatkan pada generasi F2 secara umum berada di dalam kisaran nilai kedua tetuanya, kecuali untuk karakter panjang sungut (AWN), lebar daun bendera (FLW), eksersi malai (IPE), panjang malai (PL),
jumlah biji isi (FG),tinggi tanaman (PH) dan jumlah anakan (TN) (Tabel 1). Bahkan nilai rataan untuk AWN pada generasi F2 mencatat selisih nilai cukup jauh
terhadap rataan tetua yang tertinggi, yaitu mencapai 0.27 mm. Variasi AWN pada
Variasi karakter pada populasi F2 untuk FLA, FLL, FLW, IPE, PL, FG, TSN, dan PH memperlihatkan skewness atau ke arah tetua Bio-148, seperti yang
diperlihatkan dalam grafik untuk karakter-karakter tersebut dimana posisi grafik lebih condong atau menjulur ke tetua Bio-148 (Gambar 4 dan 5). Karakter TN dan AWN memiliki kemenjuluran sebaliknya, yaitu mengarah kepada tetua IR64.
Berdasar Tabel 1 juga dapat diketahui bahwa TSN dan FG pada tetua Bio-148 terbukti lebih tinggi dari IR64. Hal tersebut mendukung pernyataan mengenai Bio-148 yang berpotensi sebagai sumber genetik untuk potensi hasil. Demikian pula PL galur tersebut yang lebih panjang daripada malai IR64 juga memperkuat pernyataan tersebut.
Namun demikian, beberapa karakter lainnya ternyata juga mencatat nilai yang kurang mendukung pernyataan bahwa Galur Bio-148 berpotensi sebagai bahan genetik untuk pemetaan populasi berdaya hasil tinggi, diantaranya adalah biji hampa (UG) pada Bio-148 juga lebih tinggi, dan persentase IPE yang jauh lebih besar (82%), dan bobot 100 biji (100GW) yang jauh lebih rendah (Tabel 1). Sebaran data pada karakter fenotipik yang diamati pada populasi F2 secara umum adalah normal, kecuali untuk karakter panjang sungut (AWN), eksersi malai yang tak sempurna (IPE) dan jumlah biji yang hampa (UG). Nilai probabilitas atau p-value untuk ketiga karakter tersebut lebih kecil dari α 0.05 yang berarti kurva
normalitas data ternyata signifikan berbeda terhadap kurva normal standar/baku (Tabel 1) atau ketiga karakter tersebut menyebar tidak normal.
Grafik panjang sungut (AWN) pada Gambar 4 memperlihatkan bahwa AWN meskipun memiliki sebaran yang tidak normal, namun kurva yang terbentuk
adalah kontinyu. Adanya puncak yang sangat berbeda nyata di kurva bagian kiri pada grafik awn tersebut menandakan bahwa terdapat satu gen yang kemungkinan merupakan gen mayor yang memiliki efek yang besar dalam mengendalikan karakter panjang sungut. Grafik IPE pada Gambar 4 meski memiliki sebaran data yang tidak normal, namun bersifat kontinyu dengan beberapa puncak, yang berarti bahwa karakter IPE kemungkinan dikendalikan oleh beberapa gen mayor. Adapun sepuluh karakter fenotipik lainnya, antara lain FLA, FLL, FLW, PL, FG, TSN, 100GW, PH, TN, dan PN memiliki nilai probabilitas (p-value) lebih besar dari α
0.05 yaitu antara 0.0543 - 0.9818. Artinya, kesepuluh karakter fenotipik tersebut memiliki kurva normalitas yang tidak signifikan bedanya terhadap kurva normal standar, atau artinya data memiliki sebaran yang normal dan kontinyu (Gambar 4 dan 5). Dengan demikian dapat diinterpretasikan bahwa karakter-karakter tersebut dikendalikan oleh banyak gen (poligenik) atau merupakan karakter kuantitatif. Adapun dua karakter lainnya yaitu IPE dan AWN apabila ingin dianalisis lebih
lanjut karakter kuantitatif yang mengendalikan sifat tersebut, pada dasarnya juga dapat dilakukan, dengan cara melakukan transformasi data untuk mendapatkan sebaran data yang normal. Namun dalam penelitian ini hal tersebut tidak dilakukan. AWN telah banyak diteliti dengan pendekatan analisis QTL, dimana
banyak QTL untuk sifat ini yang telah dipetakan. Beberapa laporan juga telah berhasil mendapatkan gen untuk QTL tersebut. Tiga gen, yaitu An-1, An-2 dan An-3 yang masing-masing dipetakan ke kromosom 3, kromosom 4 dan kromosom
5, dilaporkan mengendalikan panjang sungut melalui efek aditif (Hu et al. 2010).
Bahkan An-1 dilaporkan merupakan gen utama untuk panjang sungut yang
Tabel 1. Karakteristik fenotipik populasi F2
Karakter IR64
BIO-148 F1 F2 Kolmogorov-smirnov test
mean Kisaran data Stdev Kurto sis
Skew ness
p-value
Panjang sungut (AWN) 0.30 0.17 0.75 0.57 0-2.3 0.56 -0.403 0.72 0.0002
Sudut daun bendera(FLA) 21.69 39.26 24.23 31.27 11.75-63.3 8.15 0.936 0.64 0.2757 Panjang daun
bendera(FLL)
33.18 39.18 38.90 33.49 23.3-45.23 4.07 -0.078 0.05 0.9818
Lebar daun bendera(FLW) 1.51 1.89 1.63 1.9 1.38-2.98 0.19 5.185 0.85 0.4677
Eksersi malai-(IPE) 38% 82% 11% 42% 0-100% 0.33 -1.041 0.47 0.0002
Panjang malai(PL) 25.27 27.11 30.34 25.61 21.53-35.82 1.76 5.023 0.97 0.5316
Jumlah biji isi(FG) 117.30 202.92 196.75 143.35 76.94-223 23.53 0.355 0.09 0.6633 Jumlah spekelet per
malai(TSN) 140.87 310.95 239.25 185.21 95.176-299.4 31.17 1.461 0.53 0.5025
Bobot 100 butir(100GW) 2.414 1.4906 2.17 2.13 1.18-2.6 0.17 4.309 -0.77 0.2708
Tinggi tanaman(PH) 87.44 110.71 108.0 102.47 83.5-124 8.03 -0.259 0.11 0.6016
Total anakan(TN) 33.00 23.83 21.43 20.82 10-58 4.69 19.158 2.63 0.0543
Jumlah malai (PN) 32.7 23.33 21.29 19.83 9 - 29 3.8 -0.525 0.13 0.0552
Gambar 5. Distribusi frekuensi 100GW, PH, TN dan PL di F2
Pendugaan karakter yang berhubungan dengan Hasil dan Komponen Hasil
Analisis korelasi bermanfaat untuk mengetahui hubungan antara berbagai karakter, dalam hal ini adalah untuk mengetahui karakter yang berpengaruh kuat/sangat nyata terhadap hasil dan komponen hasil. Hasil dan komponen hasil merupakan karakter penting dalam pemuliaan tanaman, terutama yang bertujuan mencapai hasil yang tinggi dan stabil (Lanceras et al. 2004). Ketika komponen
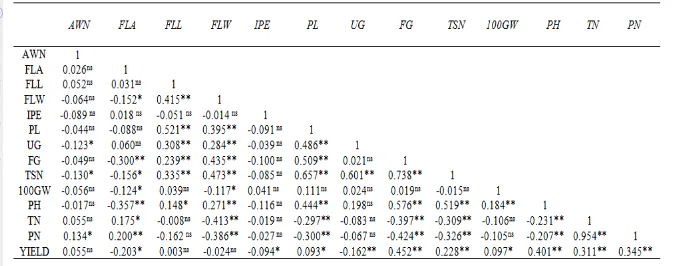
Tabel 2. Korelasi antara karakter morfologi terhadap hasil dan komponen hasil
Keterangan :* berbeda nyata pada taraf 5%, **sangat berbeda nyata pada taraf 1%, nstidak berbeda nyata
AWN = Panjang sungut (Awn) ; PN = Jumlah Malai (Panicle Number); FLA = Sudut Daun Bendera (Flag Leaf Angle) ;YIELD = Hasil
; FLL = Panjang Daun Bendera (Flag Leaf Length); TSN = Jumlah Spikelet per Malai (Total Spikelet Number); FLW = Lebar Daun
Bendera (Flag Leaf Width) ; FG = Jumlah Biji Isi (Filled Grain) ; IPE = Eksersi Malai tak sempurna (IncompletePanicle Exertion)
;100GW = Bobot 100 butir (100 Grain Weight) ; PL = Panjang Malai (Panicle Length) ; PH = Tinggi Tanaman (Plant Height); UG =
Analisis regresi dalam kaitannya dengan penelitian ini bermanfaat untuk menduga secara lebih akurat pengaruh karakter-karakter apa saja yang sangat nyata terhadap peningkatan hasil atau komponen hasil. Disinilah perlunya dilakukan kombinasi analisis korelasi dan regresi untuk memilih karakter-karakter tertentu sebagai informasi dasar yang akan digunakan dalam analisis molekuler pada deteksi QTL terkait karakter tersebut.
Korelasi antara hasil maupun komponen hasil terhadap karakter morfo-agronomi yang diamati, dievaluasi pada taraf nilai Pvalue 0.05 dan 0.01 (Tabel 2).
Korelasi sangat nyata-positif (Pvalue 0.01) antara hasil dengan karakter lainnya
memiliki koefisien korelasi antara 0.228 – 0.452, yang terdiri dari karakter TSN, TN, PN, PH, dan FG. Ini berarti, perbaikan genetik terhadap karakter-karakter tersebut akan dapat meningkatkan hasil. Adapun FLA dan UG memiliki korelasi nyata negatif terhadap hasil. Artinya, kedua karakter tersebut sebaiknya diminimalisir pengaruhnya, misalnya melalui tindakan seleksi.
Nilai koefisien korelasi yang sangat nyata pada FG, PH, TN, PN, dan TSN terhadap Yield merupakan bukti keeratan hubungan antara jumlah biji isi, tinggi tanaman, jumlah malai per rumpun, dan jumlah spikelet per malai terhadap hasil. Namun demikian, eratnya hubungan tersebut tidak menjadi jaminan bahwa hanya karakter-karakter tersebut saja yang memberi kontribusi tinggi terhadap kuantitas maupun kualitas hasil.
Analisis regresi berganda selanjutnya dilakukan terhadap seluruh fenotipik yang diamati. Model regresi yang terbaik dicari dengan membandingkan nilai Radjusted yang diperoleh dari setiap kemungkina model yang didapat berdasar
jumlah dan jenis variabl yang dimasukkan dalam pengujian regresi.
Menurut persamaan tersebut, perbaikan genetik atau peningkatan terhadap bobot 100 biji, jumlah biji isi, panjang daun bendera, tinggi tanaman, jumlah malai per tanaman, jumlah anakan per rumpun, serta penurunan terhadap sudut daun bendera, lebar daun bendera, panjang malai, jumlah biji hampa dan jumlah spikelet per malai secara simultan atau bersamaan maka akan meningkatkan hasil secara sangat nyata (P =0.000) (Tabel 3).
Nilai koefisien determinasi yang terkoreksi (Radjusted) dari persamaan
regresi tersebut adalah 65.6. Hal itu berarti bahwa 65.6% keragaman pada hasil yang diperoleh terjadi dapat dijelaskan oleh 100GW, FG, FLA, FLL, FLW, TSN, PH, PN, TN UG, IPE, dan PL dalam model regresi linear berganda. Sedangkan sisanya (34.4%) diterangkan oleh faktor lain yang tidak terdapat dalam model. Koefisien determinasi (Rseq) sedikit lebih tinggi dari Radjusted yaitu sebesar
67.8. Baik Rseq maupun Radjusted menjadi indikator kebaikan model yang
dibentuk, namun untuk model regresi berganda nilai Radjusted yang dijadikan
acuan.
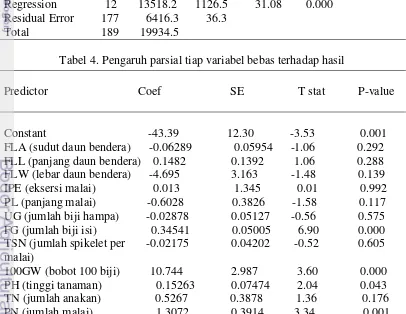
Tabel 3. Kelayakan model persamaan regresi S = 6.02080 R-Sq = 67.8% R-Sq(adj) = 65.6%
Analysis of Variance (ANOVA)
Source DF SS MS F P
Regression 12 13518.2 1126.5 31.08 0.000 Residual Error 177 6416.3 36.3
Total 189 19934.5
Tabel 4. Pengaruh parsial tiap variabel bebas terhadap hasil
Predictor Coef SE T stat P-value
Constant -43.39 12.30 -3.53 0.001 FLA (sudut daun bendera) -0.06289 0.05954 -1.06 0.292 FLL (panjang daun bendera) 0.1482 0.1392 1.06 0.288 FLW (lebar daun bendera) -4.695 3.163 -1.48 0.139 IPE (eksersi malai) 0.013 1.345 0.01 0.992 PL (panjang malai) -0.6028 0.3826 -1.58 0.117 UG (jumlah biji hampa) -0.02878 0.05127 -0.56 0.575 FG (jumlah biji isi) 0.34541 0.05005 6.90 0.000 TSN (jumlah spikelet per -0.02175 0.04202 -0.52 0.605 malai)
Jika diperhatikan, nilai konstanta atau intersep dari persamaan model regresi yang diperoleh adalah negatif 43,4. Hal ini menunjukkan bahwa untuk mendapatkan peningkatan hasil yang signifikan, maka jumlah keseluruhan kontribusi dari variabel dalam persamaan regresi tersebut minimal harus lebih besar dari 43,4.
Selain mengetahui pengaruh variabel bebas terhadap hasil secara simultan, pengaruh variabel bebas secara parsial atau individual juga dapat diketahui, yaitu dengan melihat nilai p-value setiap variabel bebas yang diuji (Tabel 4). Menurut
tabel tersebut, pada pengujian model regresi yang disusun dengan tingkat kepercayaan 95% (α =0.05), FG, 100GW, PH dan PN memberi kontribusi yang nyata terhadap peningkatan hasil, yang ditunjukkan oleh nilai p-value
masing-masing karakter tersebut lebih kecil dari 0.05. Karakter lainnya, yaitu FLA, FLL, FLW, IPE, PL, UG, TSN dan TN ternyata tidak nyata pengaruhnya terhadap hasil (pada α =0.05).
Pembahasan
Variasi karakter fenotipik
Karakter fenotipik pada populasi F2 ternyata cukup bervariasi, dimana secara umum memiliki sebaran frekuensi yang normal dan kontinyu sehingga relevan untuk digunakan dalam analisis kuantitatif (QTL). Selain itu, keragaan tanaman yang diperlihatkan oleh tetua bio-148, seperti telah disampaikan sebelumnya, beberapa karakter relevan dengan pernyataan mengenai potensi Bio-148 sebagai bahan pemetaan genetik khususnya untuk potensi hasil tinggi. Karakter tersebut terutama adalah TSN yang sangat tinggi, dan FLL/FLW yang lebih besar (Tabel 1).
Terdapat hal yang menarik berdasar informasi karakteristik fenotipik yaitu 100GW pada tetua P1 (Bio-148) ternyata jauh lebih rendah dari tetua IR64 maupun pada populasi F1 dan F2. Selain itu, UG pada Bio-148 juga lebih tinggi, dan persentase IPE yang jauh lebih besar (82%). Tingginya UG dan IPE pada turunan Bio-148 sebelumnya juga dilaporkan oleh Trijatmiko (komunikasi pribadi). Kedua karakter tersebut sangat mungkin memiliki hubungan yang erat. Dalam hal ini, malai yang tidak sempurna eksersinya berakibat pada terganggunya proses pengisian malai, sehingga hasil fotosintesa tidak terdistribusi secara sempurna.
Eksersi malai yang tak sempurna juga berimplikasi pada kelembaban di dalam bagian yang tertutup pelepah menjadi lebih tinggi. Kelembaban yang tinggi merupakan wahana yang sangat baik bagi perkembangan patogen tanaman. Pada kondisi yang buruk, bulir biji yang terkurung dalam lilitan pelepah daun bendera pada malai yang eksersinya tak sempurna tersebut akan sangat rawan terserang penyakit. Biji yang terserang cendawan akan mudah busuk, kemudian berpotensi menular ke biji-biji yang lain (Gambar 6). Pada kondisi akumulatif hal ini berakibat penurunan secara signifikan terhadap bobot biji maupun jumlah biji isi yang dihasilkan.
untuk galur Bio-148 cukup tinggi, yaitu rata-rata sebesar 2,43 gram. Bobot biji yang lebih rendah dalam penelitian ini sangat mungkin disebabkan oleh eksersi malai yang tidak sempurna. Seperti yang dikemukakan sebelumnya, biji yang terjebak dalam pelepah daun bendera menjadi terhambat perkembangannya. Penetrasi cahaya matahari, sirkulasi udara maupun respirasi menjadi tidak maksimal pada bagian yang tertutupi oleh pelepah daun bendera. Kondisi ini secara langsung maupun tak langsung dapat berpengaruh terhadap keberlangsungan proses fisiologis lainnya yang terkait dengan perkembangan biji. Perkembangan biji yang tidak sempurna selanjutnya berdampak pada tingginya jumlah biji yang hampa maupun rendahnya bobot biji.
Gambar 6. Infeksi patogen pada eksersi malai yang tidak sempurna
Galur Bio-148 menurut keragaan fenotipiknya merupakan representasi yang mendekati desain dari tipe Padi Tipe Baru, dengan karakter utama jumlah spikelet per malai yang tinggi, namun kapasitas jumlah anakan dan jumlah malai yang lebih kecil (Tabel 1). Bobot 100 biji yang rendah kemungkinan juga terkait erat dengan kapasitas anakan yang lebih kecil pada galur Bio-148 ini. Produksi biomassa juga ditentukan oleh kapasitas anakan, dimana pada kapasitas anakan yang kecil berdampak pada tingkat pertumbuhan fase vegetatif tanaman yang lebih rendah. Dalam hal ini, biomassa yang rendah adalah indikator rendahnya potensi hasil.
Menurut Peng et al.(2008), beberapa faktor yang menyebabkan rendahnya
bobot hasil biji pada Padi Tipe Baru diantara adalah kapasitas anakan yang kecil dan pengisian biji yang buruk pada varietas tersebut. Penurunan kapasitas atau jumlah anakan berkontribusi terhadap produksi biomassa yang rendah karena tingkat pertumbuhan tanaman selama fase vegetatif pada PTB lebih rendah dibandingkan varietas indica. Produksi biomassa yang lebih rendah juga terkait
Suhu rumah kaca yang relatif tinggi diduga menjadi faktor lainnya yang menyebabkan rendahnya bobot biji yang dihasilkan oleh Galur Bio-148. Berdasarkan informasi yang diperoleh dari penanggung-jawab rumah kaca BB BIOGEN, rata-rata suhu terpanas pada siang hari dalam rumah kaca mencapai kisaran 49o - 55o C (Iman Ridwan, komunikasi pribadi). Pencatatan suhu sayangnya tidak dilakukan secara khusus (rutin) selama percobaan ini di rumah kaca. Respon tanaman padi terhadap suhu memang berbeda-beda, beberapa diantaranya adalah akibat pengaruh faktor genetik. Varietas yang memiliki toleransi yang tinggi terhadap suhu tinggi, ketika ditanam pada rumah kaca, maka panasnya suhu di dalam rumah kaca tidak akan terlalu besar pengaruhnya dibandingkan varietas yang rentan.
Suhu yang terlalu panas akan menyebabkan peningkatan laju transpirasi pada permukaan tubuh tanaman, dengan demikian tanaman akan lebih cepat mengalami defisit air. Apabila suplai terhadap kebutuhan air tidak memadai, maka akan menyebabkan tanaman kekeringan, dan akibatnya tanaman akan menderita layu atau bagian tanaman terutama daun akan lebih cepat menguning/mengering dan mati. Selain itu, pada kondisi suhu yang lebih tinggi, metabolisme maupun proses-proses fisiologis yang berlangsung dalam tanaman menjadi terpacu untuk berlangsung lebih cepat. Akibatnya, beberapa proses fisiologi yang terjadi menjadi berlangsung secara tidak sempurna. Misalnya, periode pengisian dan pematangan biji yang menjadi semakin pendek, berdampak pada penurunan kualitas biji yang dihasilkan, antara lain berupa makin rendahnya bobot biji.
Galur Bio-148 mungkin memiliki toleransi yang lebih rendah terhadap suhu tinggi daripada varietas IR64. Bobot 100 biji pada galur yang dijadikan tetua betina tersebut adalah yang paling rendah dibandingkan IR64, F1 maupun F2 (Tabel 1). Meskipun hal yang berbeda dilaporkan oleh Suratmini (2010), yang menanam pada kondisi di lapangan. Namun pada rumah kaca di penelitian ini, jika asumsinya suhu di dalam rumah kaca adalah seragam, maka dugaan Galur Bio-148 memiliki toleransi rendah terhadap suhu tinggi cukup logis. Alel IR64 menyumbang ketahanan terhadap suhu tinggi pada individu F1 maupun F2, yang secara fenotipik terlihat baik F1 maupun F2 berupa rata-rata bobot 100 biji yang lebih tinggi daripada Bio-148. Meski demikian, untuk mendapatkan kesimpulan yang tepat, memerlukan investigasi lebih lanjut terhadap hal ini.
Argumen lain yang juga relevan terhadap rendahnya bobot 100 biji pada tetua Bio-148 ini adalah kemungkinan adanya alel pada Galur Bio-148 yang mengendalikan karakter jumlah spikelet per malai namun juga menyebabkan penurunan bobot biji. Sebagaimana telah diuraikan di awal, Bio-148 adalah ‘progenitor jauh’ dari padi tropical japonica yaitu Padi IRAT112 atau varietas
Gajah Mungkur. Padi IRAT112 memiliki karakter jumlah spikelet per malai yang lebat, anakan sedikit, dan tanaman lebih tinggi. Galur Bio-148 memiliki riwayat sebagai keturunan persilangan IR64 dengan IRAT112 yang telah mengalami beberapa kali backcrossing dengan IR64 sebagai tetua recurrent (KR Trijatmiko,
komunikasi pribadi). Tingginya TSN dan TN yang lebih sedikit pada Bio-148 mencerminkan adanya konservasi terhadap alel-alel atau gen-gen yang mengendalikan TSN dan TN pada galur tersebut.
belum diketahui secara pasti, apakah gen yang mempengaruhi jumlah spikelet per malai tersebut adalah gen SPIKE – gen yang berasal dari variasi alami yang unik pada padi tropical japonica dan telah berhasil diklon dan dimanfaatkan dalam
pemuliaan untuk meningkatkan jumlah spikelet secara signifikan- akan tetapi fenomena yang ditunjukkan oleh Bio-148 maupun turunannya (individu pada populasi F2) sangat mirip dengan pengaruh gen SPIKE yang dilaporkan oleh Fujita et al (2013).
Gen SPIKE adalah gen mayor yang memiliki efek pleiotrofik, antara lain meningkatkan jumlah spikelet per malai, lebar daun bendera, bobot kering akar, diameter leher malai, jumlah pembuluh ikat, namun menurunkan bobot 1000 butir dan jumlah malai (Fujita et al. 2013). Introgresi gen SPIKE ini kepada padi-padi indica ternyata menyebabkan meningkatnya jumlah spikelet per malai yang nyata
pada galur-galur introgresi yang dihasilkan, namun di sisi lain jumlah biji hampa tinggi dan bobot 1000 biji rendah. Diduga rendahnya bobot 100 biji pada tetua Bio-148 pada penelitian ini adalah salah satu bentuk dari ekpresi sejenis gen SPIKE asal padi IRAT112 (tropical japonica) yang terkonservasi secara baik
dalam kromosom 4 Bio-148. Jumlah spikelet yang meningkat signifikan pada cabang-cabang sekunder malai berdampak pada ketatnya kompetisi antar spikelet untuk memperoleh asimilat, sehingga banyak biji yang hampa atau mengalami aborsi.
Fakta bahwa populasi F2 memiliki nilai rataan 100GW yang lebih tinggi dari Bio-148, dan TSN yang tinggi namun tidak setinggi Bio-148 membuktikan bahwa: pada generasi keturunan Bio-148 gen SPIKE masih terkonservasi, namun dalam ‘konsentrasi’ yang semakin rendah, dan individu turunan juga telah mampu mengembangkan mekanisme ‘adaptasi’ terhadap efek pleiotrofik gen tersebut, dengan cara menekan efek buruk yang dihasilkan. Hal tersebut dapat dijelaskan berdasarkan efek gen terhadap peningkatan pembuluh ikat. Dengan semakin meningkatnya jumlah pembuluh ikat, maka distribusi asmilat ke cabang-cabang sekunder pada malai dapat berlangsung secara lebih baik. Pada akhirnya, selain bobot biji dapat lebih ditingkatkan, spikelet yang aborsi (jumlah biji hampa) pun semakin berkurang jumlahnya. Oleh karena itu, populasi F2 memiliki 100GW yang lebih tinggi.
Dalam penelitian ini, meski secara tidak langsung informasi heritabilitas dalam arti luas (HBS) sebenarnya dapat diduga melalui pendekatan analisis ragam
pada populasi yang ada, namun dalam hal ini tidak dilakukan. Populasi F2 dapat digunakan untuk menghitung ragam fenotip (Vf), sedangkan rataan ragam pada populasi P1, P2 dan F1 dapat digunakan untuk menghitung ragam lingkungan (Ve). Selanjutnya dari selisih antara ragam fenotip dan ragam lingkungan maka dapat diketahui nilai ragam genetik (Vg). Dari rasio antara ragam genetik dengan ragam fenotip maka nilai heritabilitas arti luas dapat diketahui.
Selisih jumlah sampel pada populasi F2 terhadap sampel pada ketiga populasi lainnya (P1, P2 dan F1) sangat besar. Implikasinya, nilai dugaan heritabilitas yang disebabkan oleh faktor genetik (ragam genetik) yang didapatkan pun cenderung akan besar. Perbedaan jumlah sampel sangat berimplikasi pada perbedaan ragam masing-masing populasi, karena besarnya nilai ragam ditentukan oleh jumlah sampel yang diamati. Pada populasi P1, P2 dan F1 yang memiliki sampel lebih kecil (n=10) dengan demikian akan memiliki ragam yang lebih kecil bila dibandingkan ragam di populasi F2 (yang memiliki 200 sampel). Nilai dugaan heritabilitas yang cenderung besar tersebut adalah karena kecilnya ragam gabungan (P1, P2 dan F1) akibat kecilnya nilai masing-masing sampel populasi pembentuknya tersebut. Hal ini jika diadopsi, dikawatirkan justru akan menyebabkan kesalahan interpretasi maupun pengambilan kesimpulan yang fatal terhadap studi pewarisan sifat untuk karakter kuantitatif yang diamati.
Untuk studi heritabilitas yang penduganya adalah ragam, sebaiknya menggunakan populasi yang lebih besar, supaya ragam yang diperoleh lebih mewakili kondisi yang sebenarnya. Dalam studi-studi heritabilitas yang biasa dilakukan, dugaan heritabilitas akan lebih valid jika jumlah sampel P1 dan P2 masing-masing minimal 50 sampel, dan F1 minimal 100 sampel, dengan jumlah minimal sampel F2 sebesar 200 sampel.
Hubungan karakter fenotipik terhadap hasil
Hasil biji (Y) pada tanaman padi merupakan karakter kompleks yang sangat tergantung pada berbagai karakter yang terkait dengan perkembangan tanaman dan komponen hasil. Beberapa karakter yang berkontribusi terhadap hasil dan berpengaruh langsung terhadap hasil diantaranya adalah jumlah spikelet per malai, jumlah malai per tanaman, dan bobot 1000 biji (Hittalmani et al. 2003).
Dalam penelitian ini, bobot 100 biji yang rendah tentu juga secara langsung berakibat pada hasil yang akan rendah pula.
Hasil pada tanaman padi juga ditentukan oleh komponen hasil yang terdiri dari jumlah malai efektif per tanaman, jumlah spikelet per malai, persentase biji isi, bobot 1000 biji atau bobot biji, fertilitas bunga maupun persentase bunga steril (Sui et al. 2013). Jumlah malai per tanaman dan jumlah spikelet per malai
merupakan komponen hasil yang sering dijadikan parameter tak langsung untuk mengukur hasil pada tanaman padi (Fujita et al 2013).
Diantara semua karakter yang sangat nyata kontribusinya terhadap hasil, karakter FG, PH dan TSN memiliki korelasi positif satu-sama lain yang sangat nyata. Artinya, perbaikan genetik yang diterapkan terhadap ketiga karakter tersebut akan positif pula pengaruhnya secara bersama-sama. Korelasi yang kuat yang didukung dengan regresi yang nyata antara hasil dengan komponen hasil pada penelitian ini relevan dengan yang dilaporkan Li et al. (2009) yang
menyebutkan bahwa jumlah spikelet per m2 (sink-size) berkorelasi tinggi dengan
hasil (r=0.96**).
bahwa semakin banyak jumlah spikelet per malai berkorelasi negatif dengan jumlah malai efektif yang dihasilkan per tanaman (Xu et al. 2005).
Hasil biji dilaporkan sangat dipengaruhi oleh berbagai proses perkembangan reproduktif maupun karakter-karakter sekunder yang berpengaruh langsung maupun tak-langsung terhadap formasi hasil, misalnya karakter yang meregulasi fertilitas bunga, jumlah malai dan panjang malai (Sellamuthu et al.
2011). Dalam penelitian ini panjang malai (PL) juga nyata korelasinya terhadap hasil.
Jumlah biji isi (FG) adalah komponen hasil dengan kontribusi sangat signifikan terhadap hasil yang mencatat nilai koefisien tertinggi yaitu 0.452. Karakter tersebut ternyata sangat dipengaruhi oleh beberapa karakter morfologi, antara lain FLW, PL, PH, dan FLL. Demikian pula komponen hasil lainnya yang sangat nyata pengaruhnya terhadap hasil, yaitu TSN, ternyata juga sangat nyata dipengaruhi oleh FLL, FLW, PL dan PH.
Tinggi tanaman (PH) merupakan karakter morfologi yang sangat nyata kontribusinya terhadap hasil. Tinggi tanaman juga dilaporkan memiliki korelasi yang nyata-positif pada hasil padi dari populasi DH, namun dalam cekaman defisit air (Lanceras et al. 2004). Dalam penelitian ini tinggi tanaman mayoritas
memiliki hubungan positif terhadap karakter lainnya kecuali terhadap FLA dan IPE. Perbaikan genetik terhadap karakter ini secara umum akan berpengaruh positif bagi hasil.
Adanya kemiripan faktor morfologi yang mempengaruhi FG dan TSN antara lain FLL, FLW dan PL memperlihatkan bahwa ketiga morfologi tersebut merupakan atribut penting dalam formasi hasil. Menariknya, baik FLL maupun FLW ternyata tidak nyata pengaruhnya terhadap hasil (Y). Meski demikian tidak berarti bahwa kedua karakter tersebut boleh diabaikan dalam proses pemuliaan. Demikian pula terhadap morfologi lainnya.
Hittalmani et al (2003) dan Sellamuthu et al.(2011) melaporkan bahwa,
hasil merupakan karakter yang kompleks, perbaikan hasil biji harus melibatkan berbagai proses baik reproduktif maupun karakter-karakter sekunder, baik yang berpengaruh langsung maupun tak langsung terhadap formasi hasil. Selain itu Sui
et al. (2013) juga menyatakan bahwa peningkatan potensi hasil relatif lebih sulit
dilakukan jika hanya memanipulasi karakter tertentu saja, perbaikan terhadap hasil perlu dilakukan melalui perluasan kapasitas sink dan penyesuaian terhadap
karakteristik formasi hasil.
Luas daun padi, yang ditentukan oleh panjang dan lebar daun, memberikan kontribusi terhadap hasil gabah, karena merupakan organ utama untuk produksi karbohidrat melalui fotosintesis (Samonte et al. 2013). Panjang daun, lebar daun
dan sudut daun merupakan karakter utama tanaman padi berdasar konsep Padi Tipe Baru, dan perbedaan pada karakter tersebut berasosiasi dengan hasil tinggi pada tanaman padi (Chen et al. 2005).
Laju fotosintesa pada daun ditentukan oleh nitrogen spesifik pada daun, dan konduktansi stomata (Li et al. 2009). Semakin pendek, lebar dan tebal daun,
cenderung pada struktur daun menjadi lebih tegak (tegak), sudut yang terbentuk lebih sempit, merupakan ilustrasi varietas berpotensi hasil tinggi . Penetrasi cahaya lebih baik dan asimilasi karbon per unit area pun lebih tinggi pada struktur kanopi dengan daun yang tegak (Long et al. 2006). Selain itu, panjang dan lebar