ANALISIS STABILITAS HASIL CABAI HIBRIDA
(
Capsicum annuum
L.)
DARMAWAN ASTA KUSUMAH
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI
Dengan ini saya menyatakan bahwa tesis yang berjudul Analisis Stabilitas
Hasil Cabai Hibrida (Capsicum annuum L.) adalah karya saya dengan arahan dari
komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di`bagian akhir tesis ini.
Bogor, Pebruari 2010
ABSTRACT
DARMAWAN ASTA KUSUMAH. The Yield Stability Analysis of Hybrid
Pepper (Capsicum annuum L.). Supervised by SRIANI SUJIPRIHATI,
MUHAMAD SYUKUR.
Multi location trials play important roles in plant breeding and agronomic research. Data from multi location trials will help farmers to estimate or predict yield potential of their varieties, provide reliable guidance for selecting the best genotypes and to determine yield stability. A number of statistical procedures have been developed over the years to analyze genotype x environment interaction and especially yield stability over environment. Joint regression analysis as the first statistical procedures to stability analysis was proposed by Yates and Cochran. The objective of this study is to identify the stability of seven hybrid peper genotypes as the breeding result of Genetic and Plant Breeding IPB, using yield stability analysis and Additive Main Effect Multiplicative Interaction (AMMI) method. The genotypes used were IPB CH1, IPB CH2, IPB CH3, IPB CH5, IPB CH25, IPB CH28, IPB CH50 and five commercial varieties i.e. Adipati, Biola, Gada, Hot Beauty and Imperial. The genotypes were planted in six different locations which included Ciherang, Leuwikopo, Tajur, Subang, Rembang and Boyolali. The design in each location was Randomized Complete Block Design (RCBD) with three replications as blocks. Based on yield stability analysis, IPB CH28 is the most stable genotypes and adaptive to the environment at 400 m above sea level. IPB CH3 has the highest fruit weight per plant (555.51 g); earlier days to flowering and days to harvesting; and bigger weight and diameter fruit compare to other genotypes. IPB CH3 also results in the highest yield and has a dynamic stability according to Eberhart and Russell’s analysis. IPB CH3 will give higher yield potential if planted in optimal environment. Based on AMMI bi-plot analysis, IPB CH3 is more suitable for Subang location. Perkins and Jinks’ stability analysis method is correlated to Finlay and Wilkinsons’ method, Francis and Kannenberg’s method, and Tai’s alpha parameter (α). Shukla’s method is
correlated to Wricke’s method, while Eberhart and Russell’s parameter S2di is
correlated to Tai’s lamdha parameter (λ), Skhula’s method and Wricke’s method.
It is possible to choose one method among others, since the correlated methods measure the same aspects. The Eberhart and Russell’s method is the most suitable method to analyze the stability of hybrid pepper.
RINGKASAN
DARMAWAN ASTA KUSUMAH. Analisis Stabilitas Hasil Cabai Hibrida (Capsicum annuum L.). Dibimbing oleh SRIANI SUJIPRIHATI dan MUHAMAD SYUKUR.
Cabai (Capsicum annuum L.) merupakan salah satu jenis sayuran buah
yang memiliki nilai ekonomi tinggi, rata-rata produktivitas cabai secara nasional pada tahun 2008 yaitu 6.37 ton per hektar. Produktivitas ini masih jauh dari potensi hasil cabai yang bisa mencapai 20 – 30 ton per hektar. Rendahnya produktivitas ini salah satunya disebabkan oleh penggunaan varietas yang tidak tepat dalam hal kesesuaian lahan. Analisis stabilitas dapat memberikan gambaran pola respon suatu genotipe terhadap perubahan lingkungan sehingga dapat digunakan petani dalam memilih varietas yang sesuai bagi lingkungan sehingga dapat memberikan produksi yang optimal.
Pemuliaan tanaman bertujuan untuk memperbaiki karakter tanaman sesuai dengan kebutuhan manusia dengan memanfaatkan potensi genetik dan interaksi genotipe dengan lingkungan. Pemulia dapat mengunakan interaksi genotipe dengan lingkungan untuk mengembangkan varietas unggul baru yang spesifik lingkungan atau varietas yang beradaptasi luas. Jika interaksi genotipe x lingkungan tinggi, maka diperlukan pengembangan suatu varietas yang spesifik lokasi, dan sebaliknya bila interaksi genotipe x lingkungannya kecil, maka dapat dikembangkan varietas beradaptasi luas. Interaksi genotipe x lingkungan adalah variasi yang disebabkan oleh pengaruh bersama dari genetik dan lingkungan. Interaksi genotipe x lingkungan merupakan hal yang menarik ketika pemulia tanaman mengevaluasi stabilitas hasil pada berbagai lingkungan. Kemampuan tanaman bertahan pada berbagai kondisi lingkungan merupakan pertimbangan penting dalam pemuliaan tanaman, dan perhatian utama dari seorang pemulian adalah menghasilkan varietas tanaman yang berdaya hasil tinggi dan stabil mengingat waktu dan biaya yang dikeluarkan.
Analisis stabilitas parametrik pertama kali diajukan oleh Yates dan Cochran, yang menyatakan bahwa derajat hubungan antara perbedaan varietas dan nilai tengah semua varietas dapat dijelaskan dengan menghitung regresi dari hasil satu varietas dengan nilai tengah hasil dari semua varietas. Yates dan Cochran menunjukkan regresi tersebut pada percobaan barley, tetapi ide mereka tidak teramati sampai Finlay dan Wilkinson menemukan ulang metode yang sama dan digunakan pada analisis adaptasi pada percobaan 277 varietas barley di tujuh lingkungan. Metode Finlay – Wilkinson mengukur stabilitas dan adaptasi tanaman berdasarkan regresi linear untuk setiap lokasi dan musim. Perhitungan regresi ini dijadikan sebagai dasar untuk menentukan tingkat derajat kelinearan yang timbul. Finlay dan Wilkinson menggunakan koefisien regresi sebagai ukuran stabilitas.
Koefisien regresi (bi) = 1.0 menyatakan rata-rata stabilitas. Penambahan nilai
koefisien terhadap 1.0 berarti meningkatkan kepekaan adaptasi terhadap perubahan lingkungan. Penurunan koefisien berarti peningkatan adaptasi terhadap perubahan lingkungan. Suatu genotipe dikatakan sangat stabil apabila nilai
hibrida harapan cabai yang memiliki daya adaptasi yang baik dan potensi hasil yang stabil, mempelajari korelasi antar metode stabilitas dan mempelajari metode analisis stabilitas yang efektif.
Percobaan dilakukan dengan menggunakan Rancangan Kelompok Lengkap Teracak dengan tiga ulangan pada 6 unit lokasi percobaan yaitu: Ciherang, Leuwikopo, Tajur, Boyolali, Rembang dan Subang. Genotipe yang diujikan adalah 12 genotipe cabai hibrida yaitu 7 hibrida harapan cabai dan 5 varietas hibrida pembanding. Setiap satuan percobaan terdiri dari 20 tanaman, dan sebelum melakukan uji gabungan dilakukan uji kehomogenan ragam untuk melakukan pendugaan komponen ragam. Untuk mengetahui pengaruh lokasi percobaan, maka dilakukan analisis. Analisis kehomogenan ragam dilakukan berdasarkan uji Barlett. Untuk mengetahui bahwa genotipe dan interaksi genotipe x lingkungan berbeda nyata, maka dilakukan uji F. Delapan metode stabilitas parametrik digunakan pada penelitian ini, yaitu metode Perkin-Jink, Finlay-Wilkinson, Eberhart-Russel, Shukla, Wricke, Francis-Kannenberg, Tai, dan Lin dan Binns serta stabilitas AMMI. Pengolahan data dan pengujian mengunakan program SAS 9.0.
Berdasarkan frekuensi kestabilan, cabai hibrida IPB CH28 memiliki frekuensi kestabilan sebanyak 10 kali sehingga dikategorikan sebagai hibrida yang paling stabil dan memiliki daya adaptasi yang lebih luas pada berbagai lingkungan di bawah 400 m di bawah permukaan laut. IPB CH28 mempunyai bobot per tanaman 418.07 g. IPB CH3 merupakan hibrida dengan produktivitas tertinggi yaitu 555.51 g/tan. IPB CH3 mempunyai umur panen dan umur berbunga lebih genjah daripada genotipe lainnya. IPB CH3 juga mempunyai bobot buah dan diameter buah yang lebih besar daripada genotipe lainnya.Berdasarkan analisis stabilitas Eberhart dan Russell, IPB CH3 merupakan hibrida yang memiliki kestabilitasan dinamis. IPB CH3 akan memberikan potensi produksi yang lebih tinggi apabila ditanam pada lingkungan yang optimal. Berdasarkan analisis biplot AMMI, IPB CH3 lebih sesuai untuk lingkungan Subang. Terdapat korelasi antara metode analisis stabilitas Perkins dan Jinks dengan stabilitas Finlay dan
Wilkinsons, Francis dan Kannenberg serta parameter alpha (α) Tai. Metode
Shukla berkorelasi dengan Wricke. Parameter S2di Eberhart dan Russell berkorelasi
dengan parameter lamdha (λ) stabilitas Tai, Shukla dan Wricke. Metode-metode
yang berkorelasi tersebut dapat dipilih salah satunya karena dalam menganalisis stabilitas mengukur aspek yang sama. Metode stabilitas Eberhart dan Russell’s merupakan metode stabilitas yang paling sesuai untuk menganalisis stabilitas cabai hibrida.
©
Hak Cipta milik IPB, tahun 2010
Hak Cipta dilindungi undang-undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau peninjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan yang wajar IPB
Judul Tesis : Analisis Stabilitas Hasil Cabai Hibrida (Capsicum annuum L.)
Nama : Darmawan Asta Kusumah
NIM : A151060021
Disetujui Komisi pembimbing
Prof. Dr. Ir. Sriani Sujiprihati, M.S. Dr. Muhamad Syukur, SP. M.Si.
Ketua Anggota
Diketahui
Ketua Program Studi Agronomi Dekan Sekolah Pascasarjana
Dr. Ir. Munif Ghulamahdi, M.S. Prof. Dr. Ir. Khairil A. Notodiputro, M.S.
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah SWT atas segala karunia-Nya sehingga penulis dapat menyelesaikan tesis yang berjudul “Analisis Stabilitas
Hasil Cabai Hibrida (Capsicum annuum L.)”. Tesis ini disusun sebagai salah satu
syarat untuk menyelesaikan jenjang pendidikan pendidikan S2 dan memperoleh gelas Magister Sains dari Program Studi Agronomi, Sekolah Pascasarjana, Institut Pertanian Bogor.
Terima kasih penulis ucapkan kepada Ibu Prof. Dr. Ir. Sriani Sujiprihati, M.S. selaku ketua komisi pembimbing dan Bapak Dr. Muhamad Syukur, SP. M.Si. Selaku anggota komisi pembimbing yang senantiasa memberikan bimbingan, arahan dan masukan kepada penulis selama melakukan kegiatan penelitian dan penyusunan tesis. Ucapan terima kasih juga penulis sampaikan kepada Ibu Dr. Ir. Yudiwanti W.E. Kusumo, M.S. dan Ibu Dr. Ir. Trikoesoemaningtyas, M.Sc. Selaku dosen penguji luar komisi dan perwakilan program studi atas masukan, kritik dan saran untuk kesempurnaan penyusunan tesis ini.
Terima kasih yang setulus-tulusnya penulis haturkan kepada orang tua tercinta Bapak Iking Soekara dan Emih Ratna Laelani yang telah membesarkan penulis dengan cinta yang tulus. Penulis menyampaikan rasa sayang dan terima kasih kepada istri tercinta Ismantiri Heningtyas serta anak-anakku Padmarani Syandina dan Ramadhan Adiputra yang selalu memberikan semangat untuk menyelesaikan penyusunan tesis. Kepada rekan-rekan mahasiswa satu proyek penelitian 2007-2008, Teddy, Madhumita, Habib, Shinta, Wahyu dan Dimas yang selalu memberikan dukungan selama melakukan penelitian.
Akhirnya penulis mengharapkan kritik dan saran yang konstruktif untuk perbaikan dan penyempurnaan tesis ini agar dapat bermanfaat bagi semua pihak.
Bogor, Pebruari 2010
RIWAYAT HIDUP
Penulis dilahirkan di Tangerang pada tanggal 7 Nopember 1974 sebagai anak ke enam dari tujuh bersaudara pasangan Bapak Iking Soekara dan Ibu Ratna Laelani. Pendidikan Sarjana ditempuh di Program Studi Agronomi, Fakultas Pertanian IPB, lulus pada Tahun 1998. Pada tahun 2006, penulis diterima sebagai mahasiswa pasca sarjana pada Program Studi Agronomi, Fakultas Pertanian IPB dengan pembiayaan sendiri.
Penulis berkarir dibidang perbenihan sejak tahun 1998, pada PT. Pioneer
Hibrida Indonesia (Dupont) sebagai District Agronomist untuk wilayah Jawa
Barat dengan kantor cabang di Bandung. Tahun 2000 penulis mendapatkan
tawaran bekerja di PT. Monsanto Indonesia sebagai Seed Agronomist untuk
wilayah regional Indonesia Timur dengan kantor cabang di Surabaya. Tahun 2002 penulis pindah ke wilayah regional Sumatera dengan kantor cabang di Padang. Pada tahun 2003, penulis berkarir di PT. Syngenta Seeds Indonesia sebagai
Regional Sales Manager untuk wilayah Jawa Barat dengan tugas utama pengembangan benih hortikultura untuk wilayah Jawa Barat dan Lampung. Sejak
Mei 2009 sampai sekarang, penulis bekerja sebagai Product Development
ANALISIS STABILITAS HASIL CABAI HIBRIDA
(
Capsicum annuum
L.)
DARMAWAN ASTA KUSUMAH
Tesis
Sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
Program Studi Agronomi
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
DAFTAR ISI
Halaman
DAFTAR TABEL ... xv
DAFTAR GAMBAR ... xvi
DAFTAR LAMPIRAN ... xvii
PENDAHULUAN ... 1
Latar Belakang ... 1
Tujuan ... 3
Hipotesis ... 3
Kerangka Pemikiran ... 4
TINJAUAN PUSTAKA... 6
Klasifikasi, Botani, dan Syarat Tumbuh Tanaman Cabai ... 6
Interaksi Genotipe x Lingkungan dan Stabilitas ... 8
Analisis Stabilitas Parametrik ... 12
Analisis Peubah Ganda ... 17
BAHAN DAN METODE ... 19
Waktu dan Tempat ... 19
Bahan dan Alat ... 19
Metode Percobaan ... 19
Pelaksanaan... 20
Pengamatan ... 21
Analisis Stabilitas ... 22
HASIL DAN PEMBAHASAN ... 24
Kondisi Umum ... 24
Keragaan Cabai Hibrida ... 27
Analisis Stabilitas ... 39
KESIMPULAN DAN SARAN ... 51
Kesimpulan ... 51
Saran ... 51
DAFTAR PUSTAKA ... 52
DAFTAR TABEL
Halaman
1 Pembagian grup stabilitas ... 12
2 Analisis ragam gabungan di beberapa lokasi pengujian menggunakan model tetap... 22
3 Formula statistik stabilitas ... 23
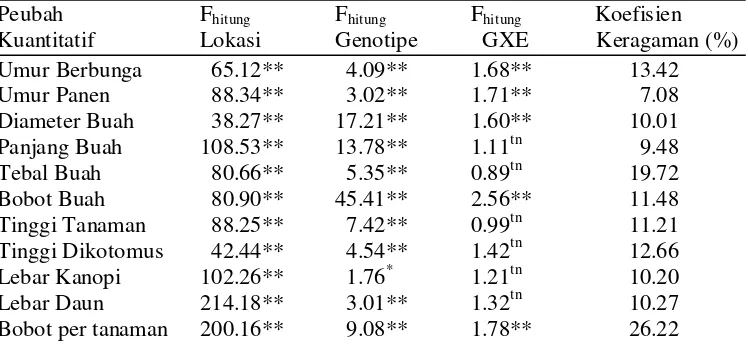
4 Rekapitulasi Fhitung, lokasi, genotipe, interaksi GXE dan koefisien keragaman ... 27
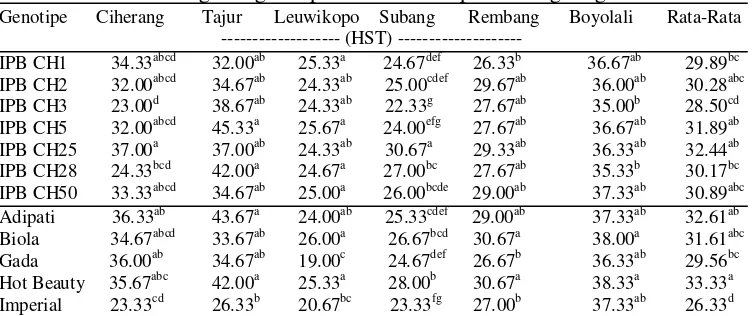
5 Umur berbunga 12 genotipe cabai hibrida pada 6 lingkungan ... 28
6 Umur panen 12 genotipe cabai hibrida pada 6 lingkungan ... 29
7 Bobot buah 12 genotipe cabai hibrida pada 6 lingkungan ... 30
8 Diameter buah 12 genotipe cabai hibrida pada 6 lingkungan... 31
9 Tebal kulit buah 12 genotipe cabai hibrida pada 6 lingkungan ... 31
10 Panjang buah 12 genotipe cabai hibrida pada 6 lingkungan ... 32
11 Tinggi tanaman 12 genotipe cabai hibrida pada 6 lingkungan ... 33
12 Tinggi dikotomus 12 genotipe cabai hibrida pada 6 lingkungan ... 34
13 Lebar kanopi 12 genotipe cabai hibrida pada 6 lingkungan ... 35
14 Lebar daun 12 genotipe cabai hibrida pada 6 lingkungan ... 36
15 Bobot buah per tanaman 12 genotipe cabai hibrida pada 6 lingkungan.. 37
16 Analisis ragam bobot per tanaman 12 genotipe cabai hibrida pada 6 lingkungan ... 39
17 Analisis stabilitas12 genotipe cabai hibrida pada 6 lingkungan ... 41
18 Korelasi Spreaman antara 10 parameter stabilitas dan bobot per tanaman 12 genotipe cabai hibrida... 46
19 Ranking analisis stabilitas 12 genotipe cabai hibrida pada 6 lingkungan ... 47
DAFTAR GAMBAR
Halaman
1 Diagram alur penelitian ... 5
2 Interpretasi umum dari pola populasi genotipe yang didapat ketika koefisien regresi genotipe diplot terhadap nilai tengah (hasil rata-rata) genotipe ... 13
3 Interpretasi parameter bi dan S2di dari pendekatan regresi ... 15
4 Gejala serangan hama tanaman ... 25
5 Gejala serangan penyakit tanaman ... 26
6 Gejala serangan hama dan penyakit sekunder ... 26
7 Bobot buah rata-rata per tanaman 12 genotipe cabai hibrida pada setiap Lingkungan ... 38
8 Pola rangking bobot buah per tanaman12 genotipe cabai hibrida pada 6 lingkungan ... 40
9 Ilustrasi stabilitas Perkin dan Jinks ... 41
10 Ilustrasi stabilitas Finlay dan Wilkinsons ... 42
11 Ilustrasi stabilitas Eberhart dan Russell... 43
12 Ilustrasi stabilitas Francis dan Kannenberg ... 44
13 Ilustrasi stabilitas Tai ... 45
14 Ilustrasi stabilitas Shukla, Wricke, Lin dan Binns ... 46
DAFTAR LAMPIRAN
Halaman
1 Listing SAS analisis stabilitas 12 genotipe cabai hibrida pada 6
lingkungan ... 58
2 Rata-rata bobot per tanaman pada 12 genotipe cabai hibrida pada 6
lingkungan ... 74
3 Rekapitulasi data karakter kuantitatif 12 genotipe cabai hibrida pada
6 lingkungan ... 75
4 Analisis ragam gabungan karakter 12 genotipe cabai hibrida yang
diuji pada enam lingkungan ... 76
PENDAHULUAN
Latar Belakang
Cabai (Capsicum annuum L.) merupakan salah satu jenis sayuran buah
yang memiliki nilai ekonomi tinggi. Tanaman cabai banyak dibudidayakan oleh
petani Indonesia karena dapat ditanam di dataran rendah maupun tinggi.
Berdasarkan data luasan dari Dirjen Bina Produksi Hortikultura 2008, tanaman
cabai memiliki luasan 19.12% dari luasan sayuran di Indonesia. Cabai banyak
digunakan untuk bumbu masak sehari-hari, industri makanan dan bahan baku
obat-obatan. Berdasarkan data Badan Pusat Statistik 2009 rata-rata produktivitas
cabai secara nasional pada tahun 2008 yaitu 6.37 ton per hektar, meningkat dari
6.22 ton per hektar (tahun 2007). Menurut Pitojo (2003) produktivitas rata-rata
ini masih jauh dari potensi hasil cabai yang bisa mencapai 20 – 30 ton per hektar.
Rendahnya produktivitas ini salah satunya disebabkan oleh penggunaan varietas
yang tidak tepat dalam hal kesesuaian lahan.
Pemuliaan tanaman bertujuan untuk memperbaiki karakter tanaman sesuai
dengan kebutuhan manusia dengan memanfaatkan potensi genetik dan interaksi
genotipe dengan lingkungan. Pemulia dapat mengunakan interaksi genotipe
dengan lingkungan untuk mengembangkan varietas unggul baru yang spesifik
lingkungan atau varietas yang beradaptasi luas. Jika interaksi genotipe x
lingkungan tinggi, maka diperlukan pengembangan suatu varietas yang spesifik
lokasi, dan sebaliknya bila interaksi genotipe x lingkungannya kecil, maka dapat
dikembangkan varietas beradaptasi luas. Interaksi genotipe x lingkungan adalah
variasi yang disebabkan oleh pengaruh bersama dari genetik dan lingkungan.
Interaksi genotipe x lingkungan merupakan hal yang menarik ketika pemulia
tanaman mengevaluasi stabilitas hasil pada berbagai lingkungan. Kemampuan
tanaman bertahan pada berbagai kondisi lingkungan merupakan pertimbangan
penting dalam pemuliaan tanaman, dan perhatian utama dari seorang pemulian
adalah menghasilkan varietas tanaman yang berdaya hasil tinggi dan stabil
mengingat waktu dan biaya yang dikeluarkan. Akcura et al. (2006) menyatakan
bahwa varietas baru akan berkembang dengan baik apabila selain memiliki daya
2
secara luas pada berbagai lingkungan. Prosedur statistik analisis stabilitas telah
banyak dikembangkan oleh para ahli untuk membantu pemulia menganalisis
interaksi genotipe x lingkungan, stabilitas genotipe, dan keterkaitan antara
stabilitas genotipe dengan interaksi genotipe. Analisis stabilitas pertama kali
dibahas oleh Yates dan Cochran (1938) dengan menggunakan regresi untuk nilai
fenotipe atau interaksi dengan lingkungan. Analisis ini dimodifikasi dan
digunakan oleh Finlay dan Wilkinson (1963) serta Eberhart dan Russell (1966).
Crossa (1990) dan Flores et al. (1998) menyatakan bahwa kestabilan
genotipe tergambar oleh tiga parameter, yaitu : nilai daya hasil rata-rata, derajat
kemiringan garis regresi (bi) dan jumlah kuadrat (SS) dari regresi deviasi (S2di). Lin et al. (1986) menyampaikan dua parameter stabilitas yang diajukan oleh Eberhart dan Russell (1966) hampir sama dengan yang diajukan oleh Tai (1971).
Pada metode ini pengaruh lingkungan (i) dan perbedaan dari respon linear (i)
dapat dibuat menjadi bentuk spesial dari parameter regresi (bi) dan (S2di), dengan
indeks lingkungan diasumsikan acak. Perhitungan stabilitas dengan mengunakan
interaksi genotipe x lingkungan untuk setiap genotipe disarankan oleh Wricke
(1962) yang disebutnya sebagai ecovalance (W2i). Shukla (1972) mengembangkan
estimasi tidak bias dengan menggunakan ragam stabilitas (2i) dari genotipe, dan
menguji nyata tidaknya dari ragam stabilitas, untuk menentukan stabilitas suatu
genotipe. Francis dan Kannenberg (1978) menggunakan ragam lingkungan (S2i)
dan koefisien ragam (CVi), dan Pinthus (1973), menggunakan koefisien
determinasi (R2i) dari setiap genotipe sebagai parameter stabilitas.
Metode lain yang dapat mengurai lebih lanjut interaksi antara genotipe
dengan lingkungan salah satunya adalah AMMI (Aditif Main effect and
Multiplicative Interaction). AMMI adalah salah satu analisis dengan menggabungkan pengaruh aditif pada analisis ragam dan pengaruh multiplikasi
pada analisis komponen utama (Mattjik 2005). Analisis Biplot AMMI dapat
menjelaskan interaksi genotipe dengan lingkungan secara lebih sederhana. Biplot
AMMI meringkas pola hubungan antar genotipe, antar lingkungan, dan antara
genotipe dan lingkungan (Sumertajaya 1998). Dengan demikian analisis AMMI
dapat meningkatkan keakuratan dugaan respon interaksi genotipe dengan
3
genotipe terhadap perubahan lingkungan sehingga dapat digunakan petani dalam
memilih varietas yang sesuai bagi lingkungan sehingga dapat memberikan
produksi yang optimal.
Tujuan
Penelitian ini bertujuan untuk : (1) mengidentifikasikan hibrida harapan
cabai yang memiliki daya adaptasi yang baik dan potensi hasil yang stabil, (2)
mempelajari korelasi antar metode stabilitas, (3) mempelajari metode analisis
stabilitas yang efektif.
Hipotesis
Hipotesis yang diajukan dalam penelitian ini adalah : (1) terdapat hibrida
cabai yang memiliki daya adaptasi dan potensi hasil yang stabil di 6 unit lokasi
dataran rendah, (2) terdapat hibrida cabai yang memiliki potensi hasil yang lebih
tinggi dibandingkan dengan hibrida pembanding, (3) terdapat analisis stabilitas
4
Kerangka Pemikiran
Penelitian ini merupakan rangkaian dari penelitian pengembangan cabai
hibrida di Bagian Genetika dan Pemuliaan Tanaman IPB yang dimulai pada tahun
2006. Penelitian dimulai dengan melakukan karakterisasi daya hasil, seleksi
ketahanan terhadap penyakit antraknosa dan penyakit phytophthora. Pada tahun
2007 dilakukan persilangan, analisis silang dialel, evaluasi pendahuluan dan
evaluasi lanjutan. Dalam analisis silang dialel diperoleh tetua yang mempunyai
daya gabung umum terbaik yaitu IPB C2. Tetua ini digunakan untuk merakit
hibrida harapan. Pada tahun 2007 dilakukan uji multilokasi pada tiga Kabupaten
Bogor yang meliputi Ciherang, Leuwikopo dan Tajur. Uji multilokasi ini
merupakan awal dari rangkaian penelitian analisis stabilitas hasil cabai hibrida
yang kemudian dilanjutkan pada tahun 2008 di tiga Kabupaten yang meliputi
Kabupaten Subang (Jawa Barat), Kabupaten Boyolali (Jawa Tengah) dan
Kabupaten Rembang (Jawa Tengah).
Data yang diperoleh dari 6 lokasi percobaan diuji kehomogenan dengan uji
Barlet’s, kemudian dilakukan pengujian interaksi genotipe x lingkungan. Analisis
stabilitas hasil dilakukan apabila terjadi interaksi antara genotipe x lingkungan.
Analisis stabilitas hasil yang digunakan adalah 10 parameter. Hasil analisis
stabilitas ini digunakan untuk membandingkan antar parameter stabilitas dalam
rangka memilih parameter yang lebih efektif. Genotipe yang memiliki kategori
stabil lebih dari 50% maka dinyatakan terpilih sebagai hibrida yang paling stabil.
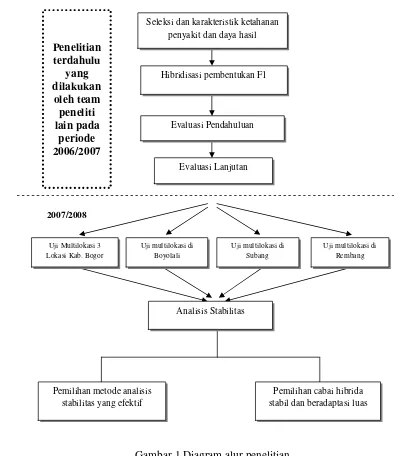
Diagram alur penelitian analisis stabilitas hasil 12 genotipe cabai hibrida disajikan
pada Gambar 1.
5
Gambar 1 Diagram alur penelitian. Seleksi dan karakteristik ketahanan
penyakit dan daya hasil
Hibridisasi pembentukan F1
Evaluasi Pendahuluan
Evaluasi Lanjutan
Analisis Stabilitas
Uji multilokasi di Rembang Uji multilokasi di
Subang Uji Multilokasi 3
Lokasi Kab. Bogor
Uji multilokasi di Boyolali
Pemilihan metode analisis stabilitas yang efektif
Pemilihan cabai hibrida stabil dan beradaptasi luas
2007/2008 Penelitian terdahulu
yang dilakukan
oleh team peneliti lain pada
TINJAUAN PUSTAKA
Klasifikasi, Botani dan Syarat Tumbuh Tanaman Cabai
Tanaman cabai tergolong divisi Magnoliophyta, kelas Magnolipsida, ordo
Solanales, famili Solanaceae, genus Capsicum dan spesies Capsicum annuum L.
(Kusandriani 1996). Tanaman cabai merupakan tanaman tropika yang memiliki
sifat menyerbuk sendiri dengan variasi penyerbukan silang yang tinggi tergantung
genotipe dan lingkungan (Daskalov 1998). Persentase penyerbukan silang pada
tanaman cabai sekitar 6 – 37 %, persentase penyerbukan silang dipengaruhi oleh
posisi dan ukuran stigma. Stigma yang lebih tinggi dibandingkan dengan kotak
sari akan menyebabkan penyerbukan silang dan sebaliknya, bunga tanaman cabai
bersifat protogeny atau kepala putik telah siap diserbuki sebelum tepung sari
masak (Permadi dan Kusandriani 1996). Tanaman cabai mempunyai jumlah
kromosom somatik diploid dengan kromosom dasar x = 12. Jumlah kromosom
normal cabai adalah 2n=2x=24 (Berke 2000). Menurut Rubatzky dan Yamaguchi
(1997), tanaman cabai merupakan tanaman herba yang berkayu pada pangkal
batangnya tetapi pada beberapa jenis menjadi semak. Batang utama tegak
ber-kayu dan bercabang banyak dengan berkisar 0.5 – 1.5 m dan memiliki perakaran
yang dangkal diawali dengan akar tunggang (akar primer) kemudian tumbuh akar
rambut ke samping (akar lateral). Panjang akar primer berkisar 35 – 50 cm dan
akar lateral berkisar 35 – 45 cm, perkembangan akar lateral yang cepat di dalam
tanah dan menyebar pada kedalaman 10 – 15 cm.
Berke (2000) menyatakan bahwa bunga tanaman cabai termasuk lengkap
berbentuk terompet terdiri dari kelopak bunga, mahkota bunga, benang sari dan
putik. Kelopak bunga berjumlah 6 helai berwarna kehijauan atau ungu, mahkota
bunga terdiri atas 5 – 7 petal berwarna putih atau ungu. Dalam satu bunga terdapat
1 putik dan 5 -7 benang sari. Menurut Greenleaf (1986), tanaman cabai mulai
berbunga pada umur 23 – 31 hari setelah tanam (HST). Buah cabai mulai masak
setelah 45 hari terjadi penyerbukan. Bunga cabai terletak pada setiap ruas, bentuk
buah cabai bervariasi dari linear, kerucut atau bulat, warna buahnya bervariasi
hijau, kuning atau ungu ketika muda, kemudian berubah menjadi merah, orange,
kuning atau ungu pada matangnya. Biji tanaman cabai terletak dalam buah dan
7
terdapat 220 biji cabai. Rubatzky dan Yamaguchi (1996) menyatakan bahwa biji
kultivar Capsicum annuum berbentuk pipih berwarna kuning pucat dan berbentuk
bulat telur dengan panjang 3 – 5 mm.
Sumarni (1996) menyatakan bahwa tanaman cabai dapat tumbuh pada
berbagai jenis tanah asalkan memiliki drainase dan aerasi yang baik. Tanaman
cabai dapat dibudidayakan pada daerah dengan ketinggian tempat hingga 2000 m
dpl. Keadaan pH tanah yang ideal untuk tanaman cabai adalah 6.0 – 6.5 dan
mengandung bahan organik sekurang-kurangnya 1.5%. Keadaan pH tanah sangat
penting karena erat kaitannya dengan ketersediaan unsur hara. Apabila ditanam
pada tanah yang mempunyai pH lebih dari tujuh maka tanaman cabai akan
menyebabkan tanaman menjadi kerdil dan gejala klorosis atau daun menguning
yang disebabkan kekurangan unsur hara besi (Fe). Pada tanah yang mempunyai
pH kurang dari lima tanaman cabai juga akan menjadi kerdil karena kekurangan
unsur hara kalsium (Ca) dan magnesium (Mg) atau keracunan aluminium (Al) dan
mangan (Mn).
Suhu udara yang baik bagi pertumbuhan tanaman cabai adalah 24 – 270 C
dengan suhu udara yang paling cocok untuk pertumbuhan cabai adalah 160C pada
malam hari dan 230C pada siang hari. Perbedaan suhu udara yang terlalu besar
akan menyebabkan kegagalan proses pembungaan dan pembuahan tanaman cabai.
Curah hujan yang sesuai bagi pertumbuhan tanaman berkisar 600 mm – 1250 mm.
Curah hujan yang terlalu rendah dapat menghambat pertumbuhan tanaman.
Me-nurut Pitojo (2003), curah hujan terlalu tinggi dapat menyebabkan kelembaban
udara meningkat dan mendorong pertumbuhan penyakit tanaman. Agar cabai
berproduksi optimal disarankan lokasi penanaman pada tempat ruang terbuka dan
tidak ternaungi. Tanaman cabai sangat cocok dibudidayakan di sawah atau tegal.
Tanaman cabai juga dapat hidup di perkarangan dan mendapat sedikit naungan
dari tanaman lain. Tanaman cabai bukan merupakan tanaman hari panjang
8
Interaksi Genotipe x Lingkungan dan Stabilitas
Pemuliaan tanaman bertujuan untuk memperbaiki karakter tanaman sesuai
dengan kebutuhan manusia. Perbaikan karakter dilakukan dengan pemanfaatan
potensi genetik dan interaksi genotipe x lingkungan. Interaksi genotipe x
ling-kungan dapat dipergunakan oleh pemulia tanaman untuk mengembangkan
va-rietas unggul baru yang spesifik lingkungan atau vava-rietas yang beradaptasi secara
luas. Pemulia tanaman memiliki tugas yang relatif mudah apabila semua
ke-ragaman fenotipe hanya dihasilkan oleh genetik saja, proses pemulian tanaman
hanya akan dibatasi pada mengidentifikasi nilai tambah genetik dari efek dominan
serta akumulasi alel-alel yang menguntungkan ke dalam populasi tanaman.
Alberts (2004) menyatakan bahwa karakter tanaman adalah hasil akhir dari
genetik yang hampir semuanya bersifat kualitatif serta kebanyakan karakter
agronomi yang penting seperti daya hasil bersifat kuantitatif dan dipengaruhi oleh
lingkungan. Sangat tidak realistis apabila suatu genotipe unggul di satu lokasi
satu musim akan menjadi unggul di semua lokasi yang lain serta unggul di semua
musim.
Indonesia merupakan negara kepulauan yang memiliki banyak perbedaan
kondisi iklim dan jenis tanah, hal ini akan lebih banyak menimbulkan kesulitan
dalam interaksi genotipe x lingkungan. Untuk mengatasi permasalahan ini,
pe-mulia tanaman harus membuat percobaan pada beberapa lingkungan (lokasi) dan
beberapa musim agar menyakinkan telah memilih genotipe yang memiliki daya
hasil yang tinggi dan penampilan yang stabil. Gauch (1992) dan De Lacy et al.
(1996) menyatakan bahwa pengaruh lingkungan terhadap genotipe dan
inte-raksinya lebih banyak berperan pada uji multi lokasi. Menurut Falconer (1952)
dan Fernandez (1991) kedua faktor tersebut tidak selalu berinteraksi positif
bahkan seringkali negatif. Untuk mendapatkan hasil interaksi genetik dan
kungan yang signifikan diperlukan pengujian multi lokasi yang beragam
ling-kungannya. Peto (1982) membagi dua bentuk dari interaksi genotipe x
lingkung-an sebagai kualitatif (perubahlingkung-an rlingkung-anking) dlingkung-an kulingkung-antitatif (perbedalingkung-an absolut lingkung-antar
genotipe). Hill (1975) dan Yau (1995) menyatakan bahwa interaksi genotipe x
lingkungan membuat tidak mudah untuk menyeleksi penampilan terbaik.
9
dalam program pemuliaan tanaman karena mengurangi kemajuan dari seleksi
pada satu lingkungan.
Lin dan Binns (1988a) menyatakan bahwa pengamatan penampilan
geno-tipe dalam percobaan genogeno-tipe x lokasi x tahun seringkali bermasalah karena
ke-hadiran interaksi lokasi x tahun atau yang disebut pengaruh lingkungan. Crossa
(1990) menyatakan bahwa data yang dikoleksi dari percobaan multilokasi akan
memiliki tiga aspek fundamental yaitu: (1) pola data yang terstruktur, dimana
jumlah genotipe yang berespon terhadap lingkungan tertentu sudah tersistematik,
signifikan dan bisa terukur; (2) data yang tidak terstruktur, dimana respon bisa
tidak terprediksi dan terukur. Fungsi dari disain percobaan dan analisis statistik
dari multilokasi adalah menghilangkan dan kemungkinan membuang data yang
tidak bisa dijelaskan; (3) hubungan antar genotipe, antar lingkungan dan interaksi
genotipe dan lingkungan.
Tanaman sangat dipengaruhi oleh lingkungan tempat tumbuhnya baik
se-cara fisik, kimia maupun biologis. Comstock dan Moll (1963) membagi
ling-kungan menjadi dua kategori, yaitu: (1) Lingling-kungan mikro, suatu lingling-kungan
di-mana satu tanaman bersaing dengan tanaman lain yang tumbuh bersamaan
wak-tunya dan tempat. Hal ini termasuk sifat fisik dan kimia seperti jenis tanah,
perbedaan cuaca, radiasi matahari, hama dan penyakit yang ada pada lingkungan
tanaman tersebut tumbuh. Menurut Roy (2000), lingkungan mikro memberikan
dampak variasi galat pada analisis statistik. Kategori (2) Lingkungan makro,
lingkungan yang berhubungan skala lokasi atau area pada satuan periode.
Lingkungan makro merupakan kumpulan dari lingkungan mikro, dimana setiap
lingkungan mikro memberikan dampak yang berbeda pada lingkungan makronya.
Dengan kata lain lingkungan makro merujuk pada kondisi iklim, tanah, serta
manajemen penanamannya (pemupukan, pengairan, kerapatan tanaman, tanggal
tanam, curah hujan dan lain-lain).
Roy (2000) membagi empat klasifikasi genotipe berkaitan dengan
kemampuan genotipe beradaptasi dengan lingkungannya, yaitu: (1) Genotipe tidak
responsif, dimana tidak menunjukkan dalam perbedaan penampilan dibandingkan
dengan genotipe lain walaupun lingkungan sudah diperbaiki seperti penambahan
10
atau tidak ada perubahan dibandingkan dengan genotipe lain apabila lingkungan
dibuat menjadi lebih jelek, seperti kekurangan nutrisi, air dan lain-lain; (3)
Genotipe stabil, dimana menunjukkan sedikit atau tidak ada perubahan
diban-dingkan dengan genotipe lain walaupun lingkungan berubah dratis dan tidak bisa
dikontrol seperti perbedaan antar musim pada wilayah agroklimat yang sama.
Secara singkat stabilitas dikategorikan sebagai mengurangi variasi antar musim;
(4) Genotipe adaptasi luas atau fleksibel, genotipe yang tidak atau sedikit
menun-jukkan perbedaan dibandingkan dengan genotipe lain ketika ditanam pada wilayah
agroklimat yang berbeda. Kemampuan adaptasi dapat didefinisikan berkurangnya
variasi dalam semua lingkungan.
Berdasarkan respon terhadap perubahan Roy (2000) juga membagi dua
kategori genotipe, yaitu: (1) Homeostatis: kemampuan genotipe apabila ditanam
pada lingkungan berbeda, tanaman secara menyeluruh (ukuran, bentuk waktu
berbunga atau waktu panen) seragam dan stabil seperti yang dideskripsikan; (2)
Stabilitas berkembang (developmental stability): kemampuan genotipe
memper-cepat tahap pertumbuhan baik secara fisiologi maupun morfologi dalam
meng-hadapi perubahan lingkungan dibandingkan dengan genotipe lain.
Menurut Alberts (2004), pemulia tanaman setuju akan pentingnya
sta-bilitas hasil, tetapi sedikit sekali yang menjelaskan definisi stasta-bilitas serta sedikit
metode untuk mengukur dan meningkatkan stabilitas hasil. Stabilitas suatu
ge-notipe adalah kemampuan gege-notipe untuk hidup pada berbagai lingkungan yang
beragam, sehingga fenotipenya tidak banyak mengalami perubahan pada
ling-kungan lain. Penyebab stabilitas adalah adanya mekanisme penyangga individu
dan penyangga populasi, genotipe dengan hasil tinggi dan stabil akan
berpe-nampilan baik pada semua lingkungan. Stabilitas fenotipe disebabkan oleh
ke-mampuan tanaman untuk dapat menyesuaikan dirinya terhadap lingkungan yang
beragam sehingga tanaman tidak banyak mengalami perubahan sifat fenotipenya.
Lin et al. (1986) mengajukan tiga tipe konsep stabilitas, yaitu : Konsep stabilitas tipe 1, suatu genotipe cenderung stabil apabila ragam antar
lingkung-annya kecil. Becker dan Leon (1988) menyebutnya stabilitas statik atau konsep
stabilitas biologis. Genotipe ini sangat stabil walaupun berada pada berbagai
11
dan penyakit atau stres lingkungan. Parameter stabilitas yang bisa mengambarkan
ini adalah koefisien ragam (CVi) (Francis dan Kannenburg 1978) pada setiap
genotipe dan ragam genotipe pada seluruh lingkungan (S2i).
Konsep stabilitas Tipe 2, suatu genotipe cenderung stabil apabila respon
terhadap lingkungannya adalah sejajar dengan respon daya hasil untuk semua
genotipe. Becker dan Leon (1988) menyatakan sebagai stabilitas dinamis atau
stabilitas agronomis. Suatu genotipe stabil apabila tidak memiliki perbedaan
secara umum respon terhadap lingkungannya dan bisa diprediksikan responnya
terhadap lingkungan yang lain. Koefisien regresi (bi) (Finlay dan Wilkinson
1963), komponen ragam nilai tengah terhadap interaksi genotipe x lingkungan (i)
(Plasteid dan Peterson 1959), komponen ragam dari interaksi genotipe x
lingkungan ( (i)) (Plaisteid (960), ecovalen (W 2
i) (Wricke 1962) dan ragam
stabilitas (2i) (Shukla 1972) dapat digunakan untuk mengukur stabilitas tipe ini.
Konsep stabilitas Tipe 3, suatu genotipe cenderung stabil apabila residu
kuadrat tengah (MS) dari model regresi terhadap indeks lingkungannya kecil.
Indeks lingkungan digambarkan dari nilai tengah semua genotipe dari setiap
lokasi dikurangi total nilai tengah semua genotipe pada semua lokasi. Tipe 3 ini
juga bagian dari stabilitas dinamis atau agronomis menurut Becker dan Leon
(1988). Metode yang menjelaskan stabilitas tipe 3 adalah metode Eberthart dan
Russell (1966), Perkins dan Jinks (1968) dan Tai (1971). Becker dan Leon (1988)
menyatakan bahwa semua prosedur stabilitas yang berdasarkan kuantitatif
pengaruh interaksi genotipe x lingkungan termasuk kedalam konsep stabilitas
dinamis. Lin et al. (1986) mendefinisikan empat grup stabilitas hasil yang
didasarkan dari deviasi dari pengaruh rata-rata genotipe (DG) dan pola interaksi
genotipe x lingkungan (GE).
Lin dan Binns (1988a) mengajukan konsep stabilitas tipe 4 yang
berdasarkan variasi bukan genetik yang bisa diprediksi dan tidak diprediksi.
Komponen yang bisa diprediksi berkaitan dengan lokasi sedangkan komponen
yang tidak bisa diprediksi berkaitan dengan tahun. Lin dan Binns (1988a)
menyarankan untuk menggunakan pendekatan regresi pada bagian yang bisa
diprediksi dan kuadrat tengah (MS) dari tahun x lokasi untuk setiap genotipe
12
(2002) konsep stabilitas tipe 4 memiliki pengertian yang sama dengan konsep
stabilitas statis. Simmonds (1991) menyatakan bahwa stabilitas statis akan lebih
banyak berguna dibandingkan dengan stabilitas dinamis pada semua kondisi,
terutama untuk negara berkembang.
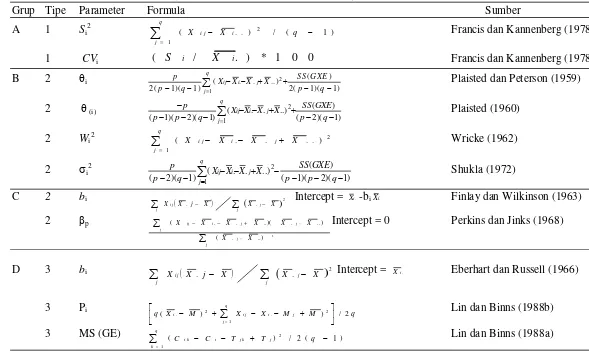
Tabel 1 Pembagian grup stabilitas (Lin et al. 1986)
Grup Dasar Perhitungan Sumber Ragam
Grup A DG (pengaruh rata-rata genotipe) jumlah kuadrat (SS)
Grup B GE (pola interaksi Genetipe X Lingkungan) jumlah kuadrat (SS)
Grup C DG atau GE koefisien regresi
Grup D DG atau GE deviasi regresi
Analisis Stabilitas Parametrik
Beragam metode telah banyak diajukan untuk menganalisis interaksi
genotipe x lingkungan dari pengujian multi lokasi. Apabila terdapat interaksi dan
terbukti nyata maka dilanjutkan dengan menganalisis stabilitas dari genotipe
untuk mendapatkan genotipe yang berdaya hasil tinggi dan stabil. Alberts (2004)
menyatakan bahwa secara umum metode yang tersedia untuk menganalisis
interaksi genotipe x lingkungan dapat dibagi menjadi empat grup, yaitu; (1)
analisis komponen ragam atau anova; (2) analisis stabilitas parametrik; (3) metode
peubah ganda (multivariate); (4) metode kualitatif atau stabilitas non parametrik.
Analisis stabilitas parametrik pertama kali diajukan oleh Yates dan
Cochran (1938), yang menyatakan bahwa derajat hubungan antara perbedaan
varietas dan nilai tengah semua varietas dapat dijelaskan dengan menghitung
regresi dari hasil satu varietas dengan nilai tengah hasil dari semua varietas. Yates
dan Cochran (1938) menunjukkan regersi tersebut pada percobaan barley, tetapi
ide mereka tidak teramati sampai Finlay dan Wilkinson (1963) menemukan ulang
metode yang sama dan digunakan pada analisis adaptasi pada percobaan 277
varietas barley di tujuh lingkungan. Metode Finlay – Wilkinson mengukur
stabilitas dan adaptasi tanaman berdasarkan regresi linear untuk setiap lokasi dan
musim. Perhitungan regresi ini dijadikan sebagai dasar untuk menentukan tingkat
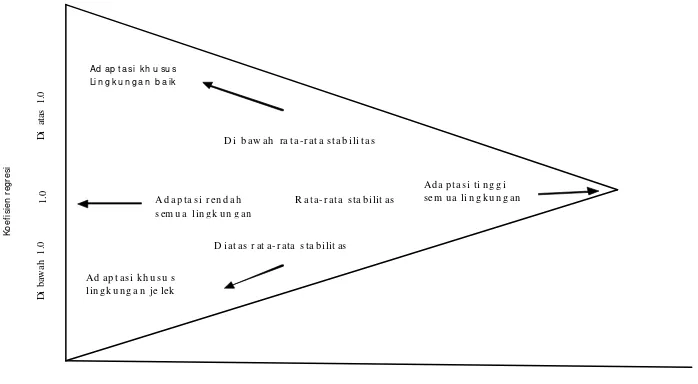
derajat kelinearan yang timbul. Finlay dan Wilkinson (1963) menggunakan
13
rata-rata stabilitas. Penambahan nilai koefisien terhadap 1.0 berarti meningkatkan
kepekaan adaptasi terhadap perubahan lingkungan. Penurunan koefisien berarti
peningkatan adaptasi terhadap perubahan lingkungan (Gambar 2). Suatu genotipe
dikatakan sangat stabil apabila nilai koefisien regresinya (bi) = 0.
Gambar 2 Interpretasi umum dari pola populasi genotipe yang didapat ketika koefisien regresi genotipe diplot terhadap nilai tengah (rata-rata hasil) genotipe (Finlay dan Wilkinson 1963).
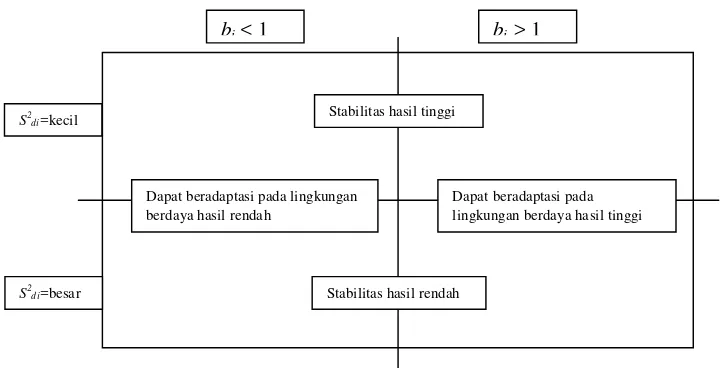
Eberhart dan Russell (1966) mengajukan pengabungan jumlah kuadrat dari
lingkungan (E) dan interaksi genotipe x lingkungan (GE) serta membaginya ke
dalam pengaruh linear antar lingkungan (derajat bebas = 1) dan pengaruh linear
dari genotipe x lingkungan (derajat bebas E = 2). Pengaruh dari residual kuadrat
tengah dari model regresi antar lingkungan digunakan sebagai indeks stabilitas.
Suatu genotipe dikatakan stabil apabila memiliki nilai deviasi regresi kuadrat
tengah (S2di) = 0 dan memiliki nilai koefisien regresi (bi) = 1.
Perkins dan Jinks (1968) mengajukan model koefisien regresi yang mirip
dengan Finlay dan Wilkinson (1963) tetapi data yang diamati disesuaikan terlebih
dahulu dengan pengaruh lingkungan sebelum dibuat regresi. Menurut Roy (2000),
model Perkins dan Jinks menjelaskan bahwa genotipe yang sangat stabil apabila
memiliki nilai βi = 0 dan genotipe ini tidak memiliki interaksi genotipe x
lingkungan. Genotipe dengan nilai βi > 0.0 tidak terlalu sensitif dengan
lingkungan, tetapi genotipe ini direkomendasikan khusus untuk lingkungan yang
14
optimal. Genotipe dengan nilai βi < 0.0 atau negatif akan lebih sedikit
perbedaannya antar lingkungan dan sangat cocok untuk ditanam pada semua
lingkungan.
Pendekatan regresi telah menjadi alat yang berguna untuk pemulia
ta-naman, tetapi ada beberapa penulis yang menunjukkan keterbatasan regresi baik
secara statistik maupun biologis. Keterbatasan pertama seperti dikemukakan oleh
Freeman dan Perkins (1971) serta Freeman (1973) adalah secara statistik nilai
tengah genotipe tidaklah bebas dari nilai marginal lingkungan. Membuat regresi
satu set peubah terhadap peubah yang lain tidak akan bebas dari saling
mem-pengaruhi satu dengan yang lain. Masalah ini akan menjadi lebih besar apabila
menggunakan jumlah genotipe yang banyak. Keterbatasan secara statistik yang
kedua seperti yang dikemukan oleh Crossa (1990) adalah galat berhubungan
dengan slope dari genotipe dan tidak secara statistik bebas, karena deviasi jumlah
kuadrat dengan derajat bebas (G-1)(E-2) dapat tidak terbagi secara orthogonal
antara G genotipe. Permasalahan yang ketiga seperti yang dikemukakan oleh
Mungomery et al. (1974) dan Wescott (1986) adalah hubungan linear antara
in-teraksi dan nilai lingkungan. Ketika asumsi ini tidak dipenuhi maka keefektifan
dari analisis akan berkurang dan menyebabkan salah pengertian dari hasil analisis.
Permasalahan secara biologi seperti dikemukan oleh Westcott (1986) dan
Crossa (1990) adalah jika lingkungan yang diikutkan hanya memiliki sedikit
lokasi yang optimal atau marjinal. Genotipe terpilih akan dijelaskan secara umum
hanya cocok pada lingkungan ekstrem. Hal ini dapat menyebabkan salah
pe-ngertian oleh karena itu analisis regresi dipergunakan dengan hati-hati apabila set
data mengikutsertakan hasil dari sedikit lokasi yang rendah atau tinggi. Becker
dan Leon (1988) mencatat ketika mempelajari metode biometrical utama, bahwa
pendekatan regresi jarang digunakan jika koefisien regresi (bi) dimasukkan ke
dalam definisi stabilitas. Dengan alasan ini, koefisien regresi (bi) oleh keba-nyakan penulis dipandang bukan sebagai ukuran kestabilan tetapi hanya sebagai
tambahan informasi dari respon rata-rata genotipe terhadap perubahan kondisi
15
Gambar 3 Interpretasi parameter bi dan S2di dari pendekatan regresi.
Wricke (1962) menggunakan interaksi genotipe x lingkungan pada setiap
genotipe sebagai ukuran kestabilan. Ukuran kestabilan Wricke (1962) disebut
ecovalance (W2i), yang merupakan jumlah kuadrat yang disumbangkan oleh satu genotipe kepada interaksi genotipe x lingkungannya. Ukuran perbedaan kestabilan
merupakan nilai konsistensi dari suatu genotipe pada semua lingkungan.
Geno-tipe yang memiliki nilai ecovalance (W2i) terkecil merupakan genotipe yang
pa-ling stabil.
Tai mengajukan dua parameter stabilitas, yaitu alpha () dan lamdha (),
yang menyerupai koefisien regresi dan ragam regresi, tapi didapat dengan cara
melanjutkan analisis ragam dan mengunakan prinsip hubungan struktural
(principle of structural relationships) (Kendall dan Stuart 1979). Mempartisikan interaksi genotipe x lingkungan kedalam regresi jumlah kuadrat dan deviasi
jumlah kuadrat dari regresi bisa dilakukan apabila pengaruh lingkungan dapat
diukur tanpa melakukan kesalahan. Tai (1971) menggunakan metode alternatif ini
dikarenakan pengaruh lingkungan tidak dapat diukur tanpa melakukan kesalahan.
Berdasarkan asumsi bahwa pengaruh lingkungan dan pengaruh interaksi genotipe
x lingkungan berdistribusi normal, Tai (1971) melakukan hubungan struktural
menurut metode Kendall dan Stuart (1979) untuk menemukan estimasi
mak-simum dari dan dari komponen yang didapat langsung dari anova. Genotipe
yang paling stabil akan tidak berubah penampilannya dari lingkungan satu ke
bi < 1 bi > 1
Stabilitas hasil tinggi
Stabilitas hasil rendah Dapat beradaptasi pada lingkungan
berdaya hasil rendah
Dapat beradaptasi pada lingkungan berdaya hasil tinggi
S2di=besar
16
lingkungan lainnya. Parameter = -1 dan = 1 merupakan parameter yang
me-nunjukkan genotipe yang paling stabil sedangkan parameter = 0 dan = 1
me-nunjukkan genotipe yang memiliki stabilitas rata-rata.
Shukla (1972) mendefinisikan ragam stabilitas genotipe sebagai ragam
seluruh lingkungan setelah pengaruh utama dari nilai lingkungan dihilangkan.
Karena pengaruh utama genotipe telah stabil, ragam stabilitas didasarkan pada
residual matrik interaksi genotipe x lingkungan dan galat sebagai klasifikasi dua
arah. Stabilitas Shukla dinamakan sebagai ragam stabilitas (σ2i). Suatu genotipe
dikatakan stabil apabila ragam stabilitas (σ2i) adalah sama dengan ragam
ling-kungan (σ2e) dimana nilai (σ2i) = 0. Nilai (σ2i) relatif besar menunjukkan
keti-dakstabilan dari genotipe. Karena ragam stabilitas adalah perbedaan antara dua
jumlah kuadrat, maka bisa bernilai negatif. Tetapi estimasi (σ2i) negatif tidak
akan menjadi masalah pada komponen ragam karena estimasi negatif dari ragam
stabilitas (σ2i) dapat dianggap sebagai nol. Pinthus (1973) mengajukan pengunaan
koefisien determinasi (ri2) dari setiap genotipe sebagai parameter stabilitas. Suatu
genotipe dikatakan stabil apabila memiliki nilai koefisien determinasi (ri2) = 1.
Francis dan Kannenberg (1978) menggunakan ragam lingkungan (S2i) dan
koefisien ragam (CVi) untuk menentukan kestabilan suatu genotipe. Suatu
geno-tipe dikatakan stabil apabila memiliki nilai ragam lingkungan (S2i) dan koefisien
ragam (CV i) kecil serta memiliki hasil yang optimal.
Lin dan Binns (1988b) mengajukan stabilitas berdasarkan ukuran
su-perioritas genotipe (Pi) atau yang disebut sebagai stabilitas tipe 4. Superioritas
genotipe (Pi), merupakan parameter stabilitas yang sangat spesifik. Suatu
geno-tipe dikatakan stabil apabila memiliki nilai (Pi) terkecil. Menurut Lin dan Binns
(1988), stabilitas tipe 4 lebih konsisten dibandingkan dengan stabilitas tipe 3
ka-rena mengunakan ekspresi nyata dari prilaku genetik suatu genotipe. Oleh kaka-rena
itu suatu genotipe yang memiliki nilai (Pi) terkecil akan memiliki sedikit
17
Analisis Peubah Ganda
Menurut Crossa (1990), analisis peubah ganda memiliki tiga tujuan utama
yaitu: (1) menghilangkan gangguan pola data, seperti membedakan ragam
sis-tematik dengan non sissis-tematik; (2) menyimpulkan data; (3) mengungkap struktur
data. Berbeda dengan metode statistik biasa, fungsi dari analisis peubah ganda
adalah mengurai struktur internal data dari hipotesis yang dapat dijabarkan dan
kemudian diujikan dengan metode statistik. Alberts (2004) menyimpulkan bahwa
analisis peubah ganda mengutamakan analisis matrik dua arah dari genotipe dan
lingkungan. Respon dari setiap genotipe pada lingkungan tertentu dapat dianggap
sebagai pola ruang dimensi lingkungan, dengan koordinat masing-masing hasil
dari genotipe dalam satu lingkungan. Menurut Gauch (1982) dan Crossa (1990),
terdapat dua grup teknik peubah ganda yang bisa digunakan untuk mengurai
struktur internal interaksi genotipe x lingkungan, yaitu :
1. Teknik ordinat, seperti principal component analysis (PCA), principal
coordinate’s analysis dan analisis faktor. Asumsi yang diperlukan adalah data merupakan satu kesatuan. Tehnik ini menempatkan genotipe yang
terpilih dan kaitannya dengan lingkungan adalah sangat mungkin berada
pada ruang dimensi yang dekat.
2. Teknik klasifikasi, seperti analisis gerombol dan analisis diskriminasi.
Metode ini melibatkan kelompok yang mirip pada satu gerombol dan
efektif untuk menyimpulkan kelebihan pada data.
Menurut Crossa (1990), principal component analysis sering digunakan
pada metode peubah ganda (multivariate). Tujuannya adalah untuk
mentrans-formasikan satu set koordinat axis ke yang lain dengan segala kemungkinannya.
Konfigurasi asli dari set utama dari struktur data ditempatkan pada komponen axis
utama. Principal coordinates analysis adalah generalisasi dari analisis PCA yang
mana setiap ukuran kemiripan antara individu dapat digunakan. Analisis faktor
masih terkait dengan PCA, faktor yang terbentuk menjadi mirip seperti principal
component. Jumlah besar peubah yang berkolerasi akan berkurang menjadi jum-lah kecil faktor utama. Keragaman dijelaskan sebagai pola faktor umum untuk
18
Menurut De Lacy et al. (1996), analisis gerombol adalah teknik klasifikasi
numerik yang mendefinisikan grup dari gerombol individu-individu. Tujuan
utama dari analisis gerombol adalah untuk mengklasifikasikan obyek baik
geno-tipe maupun lingkungan ke dalam grup-grup dimana telah dibuat minimal
kera-gaman dalam grup, sementara kerakera-gaman antar grup dimaksimalkan. Banyak cara
untuk membuat skala dan standarisasi data melalui pemusatan pada lingkungan,
standarisasi lingkungan, pembobotan heritabilitas lingkungan, atau membuat
ranking pada lingkungan. Beragam metode gerombol kadang kala membuat
per-bedaan hasil, walaupun data yang digunakan sama. Dasar dari semua metode
ge-rombol adalah mengunakan ukuran kemiripan (proximity) atau ketidakmiripan
(distance) untuk mengklasifikasikan obyek ke dalam grup.
Analisis biplot AMMI dapat menjelaskan interaksi genotipe x lingkungan,
menampilkan pola sebaran posisi relatif genotipe pada lingkungan. Hasil
pengu-raian nilai singular diplotkan antara satu komponen genotipe dengan komponen
lingkungan secara simultan. Biplot AMMI dapat meringkas pola hubungan antar
genotipe, antar lingkungan, dan interaksi genotipe x lingkungan. Biplot
menam-pilkan nilai komponen utama pertama dan nilai tengah. Interpretasi biplot nilai
komponen pertama dan nilai tengah respon, dibuat jarak titik amatan yang
ber-dasarkan sumbu datar dimana titik amatan menunjukkan perbedaan pengaruh
utama amatan-amatan tersebut (Mattjik 2005).
Sumertajaya (1998) menyatakan bahwa jarak titik-titik amatan
berdasar-kan sumbu tegak menunjukberdasar-kan perbedaan pengaruh interaksinya atau perbedaan
tingkat sensitifitas terhadap lokasi. Sedangkan interpretasi untuk titik sejenis
yang diperoleh dari biplot nilai komponen utama kedua dan nilai komponen utama
pertama merupakan jarak titik-titik amatan yang menunjukkan perbedaan
in-teraksi. Interpretasi titik-titik amatan yang berlainan jenis biplot nilai komponen
utama kedua dan nilai komponen utama pertama menunjukkan jenis interaksi
antar titik-titik amatan. Titik-titik amatan yang mempunyai arah yang sama
me-nunjukkan berinteraksi positif (saling menguatkan) dan titik-titik yang berbeda
BAHAN DAN METODE
Waktu dan Tempat
Penelitian dilakukan pada dua periode percobaan dengan pemilihan lokasi
percobaan di bawah ketinggian 400 m di atas permukaan laut. Periode pertama
percobaan dilakukan di tiga unit lokasi percobaan di Kabupaten Bogor (Jawa
Barat), yaitu Ciherang, Leuwikopo dan Tajur yang dilaksanakan pada bulan
September 2006 – Mei 2007. Periode kedua dilakukan pada tiga kabupaten yaitu
Boyolali (Jawa Tengah), Rembang (Jawa Tengah) dan Subang (Jawa Barat) yang
dilaksanakan pada bulan Desember 2007 – Agustus 2008. Bogor terletak pada
ketinggian 190 m di atas permukaan laut (m dpl), suhu rata-rata 22.6-31.80C dan
curah hujan 382 mm/bulan. Subang terletak pada ketinggian 47 m dpl, rata-rata
curah hujan 104 mm/bulan. Temperatur minimum 22.60C dan maksimum 31.70C
dengan suhu rata-rata 270C. Boyolali mempunyai ketinggian tempat 104 m dpl
dengan suhu rata-rata 260C dan curah hujan 233.5 mm/bulan.
Bahan dan Alat
Bahan penelitian yang digunakan adalah tujuh hibrida cabai harapan hasil
perakitan Bagian Genetika dan Pemuliaan Tanaman IPB, yaitu : IPB CH1, IPB
CH2 IPB CH3, IPB CH5, IPB CH25, IPB CH28 dan IPB CH 50 dengan lima
hibrida komersial cabai sebagai pembanding yaitu Adipati, Gada, Biola, Hot
Beauty dan Imperial. Hibrida pembanding ini digunakan karena sudah menjadi
hibrida rekomendasi dari PT Heinz ABC dalam kemitraan dengan petani. Bahan
lain yang digunakan adalah media tanam, pupuk NPK, pupuk kandang, pupuk
urea, KCl dan SP-36, pestisida sebagai bahan perawatan tanaman. Alat yang
digunakan adalah tray, gembor, ajir, cangkul, kored, meteran, timbangan analitik,
timbangan kasar, spayer, tali rafia, mulsa plastik, label dan jangka sorong.
Metode Percobaan
Percobaan dilakukan dengan menggunakan Rancangan Kelompok
Lengkap Teracak dengan tiga ulangan pada 6 unit lokasi percobaan yaitu
Ciherang, Leuwikopo, Tajur, Boyolali, Rembang dan Subang. Setiap satuan
20
hibrida yaitu: 7 hibrida harapan cabai dan 5 varietas hibrida pembanding. Sebelum
melakukan uji gabungan dilakukan uji kehomogenan ragam untuk melakukan
pendugaan komponen ragam. Untuk mengetahui pengaruh lokasi percobaan,
maka dilakukan analisis gabungan dari tiap lokasi percobaan. Model linear
Rancangan Acak Kelompok dengan pola gabungan adalah sebagai berikut
(Gomez dan Gomez 1985):
Yijk= µ + Lk + βi/k + Gj + (LG)kj + εijk
Keterangan :
Yijk = nilai pengamatan dari ulangan ke-i, genotipe ke-j dan lokasi ke-k
µ = nilai rataan umum Lk = pengaruh lokasi ke-k
Βi/k = pengaruh ulangan ke-i dalam lokasi ke-k
Gj = pengaruh genotipe ke-j
(LG)kj = pengaruh interaksi lokasi ke-k dengan genotipe ke-j
εijk = pengaruh galat percobaan
i = 1, 2, 3 j = 1, 2, 3,.... 16 k = 1, 2, 3,...6
Pelaksanaan
Pelaksanaan percobaan dilakukan dengan tahapan sebagai berikut :
1. Persemaian. Persemaian dilakukan dengan mengunakan tray semai.
Media yang digunakan yaitu tanah dan pupuk kandang yang telah
diayak dengan perbandingan 1:1.
2. Pengolahan lahan. Pengolahan lahan dilakukan satu bulan sebelum
penanaman. yaitu dengan membajak tanah dan mengaplikasikan pupuk
kandang dengan dosis 20 ton/ha. Lahan dibuat bedengan-bedengan
dengan ukuran bedengan 1 X 5 m. Jarak antar petak percobaan 0.5 m.
Setelah itu diberi pupuk urea, SP-36, dan KCl, kemudian ditutup
dengan mulsa plastik hitam perak.
3. Transplanting. Pemindahan bibit ke lapang dilakukan setelah bibit
berumur 4 minggu setelah semai dengan jumlah daun mencapai 4-5
21
tanaman. Penyulaman dilakukan satu minggu setelah tanam pada bibit
yang tidak sehat pertumbuhannya dengan bibit baru yang umurnya
sama.
4. Pemeliharaan. Pemeliharaan meliputi pengajiran yang dilakukan pada
2 MST. Pengendalian hama dan penyakit dilakukan apabila terlihat
adanya gejala serangan hama dan penyakit pada tanaman.
5. Panen. Panen dilakukan setelah populasi mencapai 75% buah matang.
Pemanenan dilakukan bertahap sampai delapan minggu dan setiap
minggu dilakukan satu kali panen.
Pengamatan
Pengamatan dilakukan pada 10 tanaman contoh setiap satu satuan
percobaan. Karakter kuantitatif yang diamati mengacu pada IPGRI Descriptor
(1995), yaitu:
1. Umur berbunga (HST). Umur berbunga adalah jumlah hari setelah pindah
tanam sampai 50% populasi tanaman dalam petakan telah mempunyai
bunga mekar pada percabangan tanaman.
2. Umur panen (HST). Umur panen adalah jumlah hari setelah transplanting
sampai 50% tanaman dalam petakan mempunyai buah masak pada
percabangan pertama.
3. Tinggi tanaman (cm). Tinggi tanaman diukur dari permukaan tanah
sampai pucuk, pengukuran dilakukan setelah panen pertama.
4. Tinggi dikotomus (cm). Tinggi dikotomus diukur dari permukan tanah
sampai percabangan utama, pengukuran dilakukan setelah panen pertama.
5. Lebar kanopi (cm). Lebar kanopi diukur pada kanopi tanaman terlebar
pada fase generatif (20 MST).
6. Lebar daun (cm). Lebar daun diukur dari 20 daun dewasa setelah 50%
populasi tanaman berbuah masak.
7. Bobot per buah (g). Bobot per buah dihitung berdasarkan rata-rata bobot
buah dari 10 buah segar dari panen kedua.
8. Panjang buah (cm). Panjang buah diukur dari 10 buah segar dari panen
22
9. Diameter buah (cm). Diameter pangkal-tengah-ujung diukur dari 10 buah
segar dari panen kedua.
10. Tebal kulit buah (cm). Tebal kulit buah dihitung berdasarkan rata-rata
tebal kulit buah dari 10 buah segar dari panen kedua.
11. Bobot per tanaman (g/tan). Bobot per tanaman adalah jumlah keseluruhan
bobot buah yang dipanen dari 10 tanaman contoh pada panen ke-1 sampai
panen ke-8.
Analisis Stabilitas
Analisis ragam gabungan untuk beberapa lokasi menurut Annicchiarico
(2002) disajikan pada Tabel 2. Untuk mengetahui bahwa genotipe dan interaksi
genotipe x lingkungan berbeda nyata, maka dapat dilihat nilai F hitungnya. Jika
nilai F hitung > nilai F tabel pada taraf α0.05 maka perlakuan tersebut dinyatakan
berbeda nyata. Untuk mengetahui stabilitas hasil pada populasi cabai hibrida
dilakukan analisis stabilitas. Analisis stabilitas dilakukan dengan menggunakan
delapan metode analisis yaitu 1) Perkins dan Jinks, 2) Finlay dan Wilkinsons, 3)
Eberhart dan Russell, 4) Francis dan Kannenberg, 5) Tai, 6) Shukla, 7) Wricke
dan 8) Lin dan Binns. Formula statistik stabilitas disajikan pada Tabel 3 dan
listing SAS untuk menganalisis stabilitas hasil dapat dilihat pada Lampiran 1.
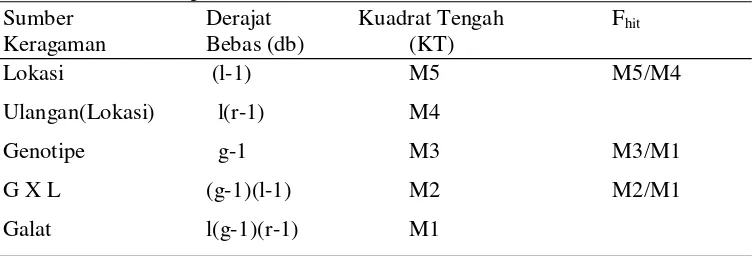
Tabel 2 Analisis ragam gabungan di beberapa lokasi pengujian menggunakan model tetap (Annicchiarico 2002)
Sumber Derajat Kuadrat Tengah Fhit
Keragaman Bebas (db) (KT)
Lokasi (l-1) M5 M5/M4
Ulangan(Lokasi) l(r-1) M4
Genotipe g-1 M3 M3/M1
G X L (g-1)(l-1) M2 M2/M1
Galat l(g-1)(r-1) M1
Tabel 3 Formula statistik stabilitas (Lin et al. 1986 dan Hussein et al. 2000)
Grup Tipe Parameter Formula Sumber
HASIL DAN PEMBAHASAN
Kondisi Umum
Penelitian ini dilakukan pada enam lokasi dengan ketinggian di bawah 400
meter di atas permukaan laut. Dua lokasi dilakukan pada kebun percobaan IPB,
yaitu kebun percobaan IPB Tajur II dan Leuwikopo, Kabupaten Bogor, Jawa
Barat (± 190 m dpl). Empat lokasi percobaan lainnya dipilih lahan petani agar
terjadi penyebaran lokasi dan mendekati kondisi lingkungan yang sebenarnya,
yaitu satu lokasi di Ciherang Kabupaten Bogor, Jawa Barat (± 190 m dpl),
Kabupaten Subang, Jawa Barat (± 47 m dpl), Kabupaten Boyolali, Jawa Tengah
(± 104 m dpl), dan Kabupaten Rembang, Jawa Tengah (± 47 m dpl). Penelitian
ini dilakukan dua periode, yaitu periode pertama dilakukan pada tiga lokasi
percobaan di Kabupaten Bogor. Penanaman dimulai pada bulan September 2006
dan panen terakhir pada bulan Mei 2007, periode kedua dilakukan di dua Propinsi
dengan periode tanam mulai Desember 2007 dan panen terakhir pada bulan
Agustus 2008.
Selama masa pembibitan, penyakit layu bakteri dan etiolasi menghambat
pertumbuhan bibit tanaman. Bibit tanaman yang normal dan sehat mulai dipindah
ke lokasi percobaan setelah berumur 4 minggu atau bibit tanaman sudah memiliki
4-5 daun. Penanaman dilakukan pada sore hari untuk menghindari panas matahari
dan menghindari stres yang berlebihan pada bibit tanaman. Setelah bibit tanaman
dipindah ke lapangan kadangkala mengalami gangguan oleh hembusan angin
yang kencang sehingga menyebabkan rebah. Penyulaman dilakukan pada bibit
tanaman yang mati dan rusak agar jumlah tanaman tiap petak tetap.
Pada awal penanaman, tanaman diserang hama siput dan belalang. Hama
tersebut menyerang tanaman yang masih muda dan memakan pangkal batang
sampai pucuk tanaman. Pengendalian dilakukan dengan menyemprotkan pestisida
curacon dan segera dilakukan penyulaman. Hama kutu, tungau dan thrips mulai
menyerang pada satu bulan pertama tanaman (Gambar 4A dan 4B). Hama-hama
tersebut menyerang daun tanaman sehingga menyebabkan daun menjadi kering
dan keriting. Hibrida IPB CH3 terlihat lebih toleran terhadap serangan hama
thrips dan tungau. Pengendalian hama kutu, thrips dan tungau dilakukan dengan
25
sampai tanaman siap dipanen. Hama lalat buah menyerang pada fase generatif
yang menyebabkan buah cabai menjadi busuk dan rontok (Gambar 4C) untuk
mengendalikan lalat buah digunakan pestisida petrogenol sebagai perangkap hama
lalat bibit. Hibrida yang lebih banyak terserang oleh lalat buah adalah IPB CH2
dan IPB CH3.
Gambar 4 Gejala serangan hama tanaman. Kutu (A), thrips (B) dan lalat buah (C).
Serangan penyakit rebah pangkal batang banyak terjadi pada percobaan
ini (Gambar 5A). Penyakit yang banyak merusak tanaman percobaan dan sangat
besar pengaruhnya terhadap penurunan produksi adalah layu fusarium dan
antraknosa. Layu fusarium dapat menyebabkan tanaman layu dan mengering
(Gambar 5B), penyebaran penyakit layu fusarium sangat cepat dan dapat
menghabiskan populasi tanaman. Pengendalian penyakit ini dilakukan dengan
segera mencabut tanaman yang terserang dan membuangnya agar tanaman lain
tidak ikut terserang. Hibrida yang paling rentan terserang penyakit layu fusarium
adalah Biola, Gada dan IPB CH5.
Penyakit antraknosa merupakan penyakit yang sangat sulit dikendalikan,
apabila buah cabai sudah terkena penyakit ini dapat menyebabkan buah menjadi
busuk dengan warna coklat kehitaman, pengendalian dilakukan dengan
meng-gunakan pestisida antracol mulai berbunga sampai panen minggu kesepuluh.
Hibrida yang lebih rentan terserang penyakit ini adalah IPB CH28. Penyakit
antraknosa sangat mudah menular apabila satu hibrida sudah terkena maka lambat
laun semua hibrida akan terkena pula (Gambar 5C).