Latar Belakang
Indonesia xnerupakan negara penghasi ubijalar [Ipnroea batatas &.) Lam] nomor dua di dunia walaupun produksi nasional 2.2 juta ton daIam tahun 1987 tebll
menurun tajarn menjadi 1.3 juta ton
dalam
tahun 199 1 (Biro Pusat Statistik Indonesia, 1991). Salah satu faktor penyebab penurunan tersebut karena ubijalar belum d i i t k a n untuk berbagai tujuan kegunaam Sekitar 88% produksi nasionaf diunakan sebagai pangan pokok atau selingan. Pemanfktan ubijatar sebagai bahan baku industri belum dikembmgkan di Indonesia.Peranan ubijatar untuk tujuan industri dapat ditkgkatkan apabila %or kualitas umbi rnenjadi perhatian utama dalam sasaran program pemuliaan, selain daya
hasil tinggi tahan penyakit kudis daun [ EIsinoe batatos (Viegas dan Jenkins)], dan hama penggerek umbi [ C y h fonnicarius Wab.)]. (Dimyati, Wargiono dan Mallian, 1988). Komponen utama kualitas umbi untuk bahan baku indutri arlnlah kadar &&an kering, pati, gula, protein, atau karoten (Ghosh, Ramanujam, Jos, Moorthy. dan Nair, 1988; Truong
dan
Fernentira, 1988).Dalam strategi untuk rnencapai sasaran pemuliaan tanaman terdapat dua kegiatan utama yang saling menunjang dan melengkapi, yaitu perbaikan popuksi dan
perakitan varietas (Gallais, 1979). Perbailcan pop& merupakan jangka
Kedua kegiatan utarna tersebut harus dilakukan secara terpisah. Strategi perbaikan populasi rnenjadi sumbu utama program pemuliaan d m dalam setiap daur menghasilkan cabang-cabang kegiatan untuk merakit varietas barn. Kegiatan perbaikan populasi yang dhkukan dengan terarah dan m t akan menunjang keghtan perakitan varietas baru dengan tepat guna
Populasi ubijalar Indonesia masa kini terdiri dari beberapa n i u klon dengan sifat morfologi, daya basii W t a s umbi d m &i ketahanan penyakit kudis yang sangat beragam
-warin.
Sukaya, dan Hartana, 1993). Klon-klon budidaya tersebut mexupakan hasil seleksi massa berdasarkan selera dan budaya setempat terhadap karakter agronomi dan g a s t r o n o e dan temdaptasi dengan ekosistem pertanian setempat. Keanekamgarnan klon ubijalar tersebut dapat dipandaug sebagai hasi kegiatan perbaikan populasi Dari popdasi ubijalar lokal Indonesia diharapkan dapat diseleksi Won-klon dengan perpaduan sif%i a g ~ o w m i , 6siologi biokimia atau ke- terhadap hama, penyakit, dan dapat digmakm sebagai tetus dalam program perakitan varietas baru. Atau bila terdapat klon ubijalar bkaI yangmenampilkan kombihasi karakter penting yang kbih unggul dari pa& varietas
nasional masa kki, &pat dipertimbangkan untuk dilepas sebagai varietas nasional baru.
Kegiatan perakitan varietas alutn sangat bermakna apabila menghasinEan
3
lingkungan luas (Yue, Pemg, Walter, Wassom dan
L i ,
1990). K e s t a b i h daya hasil telah mnjadi sasaran setiap program pemuliaan. Karena fkktor pembatas tertentu, pemulia tanaman jarang menguji sifkt kualitas pada berbagai h g k w g a n untuk memahami ciri k e s t a b b y a . Pemahaman kestabiLm ktditas basil suatu varietas sangat penting karena berkaitan dengan mutu suatu produk a k h (Peterson, Graybosch, Baenziger, dan Grombacher, 1992).hkblakshmi, Bidinger,
Rao,
dan Raju (1992) mengtmgkapkan alasan petani di kawasan laban kering Aftika dan Asia Selatan yang tetap membudidayakan raslokal Pennisetum glaucurn setempat karena memiliki kestabilan dan daya adaptasi baik terhadap cekaman Lingkungan, walaupun berdaya has2 lebih rendah dari pada
varietas baru. Untuk meningkatkan daya has& ras lokal terpilih digunakan sebagai tetua daIam program persilangan-puncak. Penelitian menunjukkan h i d persibngan- puncak yang dhsilkan berdaya hasil tinggi dan tetap mempertahankan kestabilan dan
daya adaptasi terhadap cekaman lingkungan
al.,
1992).yang berdaya adaptasai sempit atau luas. Varietas berdaya adaptasi luas mampu berproduksi baik pada lingkungan subur maupun kurang s u b , sedangkan yang berdaya adaptasi sempit hanya berproduksi baik pa& salah satu dari kedua tipe Lingkungan tersebut.
Pendekatan statistika telah dikembangkan untuk mem&kkan pengaruh GxL dan memperoleh gambaran tentang kestabilan genotipe. Beberap metode pendekatan parametrik telah dikemukakan oleh Plaisted dan Peterson (1959), Wricke (1962), Finlay dan Wrlkinson (1963), Eberhart dan Russell (19661, Tai (1971), Shukla (1972), dan Francis dan Kamenberg (1978). Pendekatan non- parametrik dikemukakan pula oleh Nassar dan Huhn (1987), Lm dan Birms (1988),
dan Huehn (1 990% 1990b).
Penggunaan metode regresi untuk me&irkan k e s t a b h varietas dikemukakan pertama kali oleh Mooers (1921) dan dikembmgkau oleh Yates clan Cochran (1938), kemudian oleh Finlay dan Willrinson (1%3), dan Eberhart dan Russell (1966). Regresi rataan varietas terhadap indeks lingkungan yang
dikemukakan Eberhart clan Russell (1966) lebih s e r i n g d i i untuk mengukur kestabilan varietas (Hanson, 1994). Yue g
d.
(1990) -kan bahwa hasit analisis kestabilan daya hasid jagung, gandum clan canteI di beberapa lokasi pengujietn meqgumkm metode Eberhart dan Russell menunjukkan statistik regresi lingkunganberbagai statist* kesta-bilan untuk digunakan &lam pengujian kestabilan daya hasil klon-klon ubijalar. Dikemukakan pula bahwa metode Eberhart dan Russel (1966) termasuk salah satu yang layak digunakan dalam menguji kestaKlan klon ubijalar.
MeMui penelitii
Hii
Bersaing Perguruan Tinggi, FMIPA IPB tehhmelakukan karakterisasi awal dan evaluasi pmdahuluan dalam tahun 1992 terhadap
253 klon ubijalar lokal Indonesia asal Sumatera, Jawa dan Irian Jaya. Percotman berlangsung di Bogor Jawa Barat clan Manokwari Irian Jaya Sebanyak 110 klon
ubijalar lokal yang berpotensi untuk karakter daya basil, kualitas umbi, atau toleran penyakit kudii diseleksi dan dievaluasi kembali di kedua lokasi yang sama &lam tahun 1993 (Ehtam, 1993, 1994, 1995). Dari basil evaluasi tersebut, diseleksi 28 klon ubijalar lokal untuk dievaluasi pada rentang keadaan hgkungan yang lebih luas. Evaluasi terhadap tanggapan spasial rnaupun temporal pada rentang keadaan
lingkungan yang luas dihrapkan dapat menyeleksi klon ubijalar sesuai dengan tujuan
penelitian.
Tujuan Penelitii ini bertujuan :
1. Mempertelakan keragaman sifat morfologi, daya has& dan kualitas umbi 28 klon ubijalar lokal untuk mehalt kemungkinan hubungan antar s i t 3 yang berkaitan dengan daya adaptasi klon ubijalar terhadap lingkungaa
3. Menyeleksi klon ubijalar dengan kombinasi sifat daya hasii tinggi, kualitas urnbi baik, tahan penyakit kudis, dan berperagaan stam untuk keadaan lingkungan produkt'i kurang produktii, atau keduanya.
4. Mernpelajari makna seleksi klon tetua berdaya basil stabil terhadap pemuliaan ubijalar.
Hipotesis
1 . Terdapat keragaman kestabilan daya hasil dan kualitas umbi klon ubijalar lokal Indonesia
2. Klon ubijalar lokal stabil memiliki potensi daya hasil dan kualitas umbi di bawah rata-rata umum
TINJAUAN PUSTAKA
Potensi Ubijalar
Ubijalar merupakan salah satu tamman penghasil ksrbohidrat berproduksi tinggi di dunia dan sebagai pangan penting bagi petani di sekiar seratus negara di daerah tropik dan subtropik (Horton, Prain dan Gregory, 1989). Tanaman tersebut dapat tumbuh dan berproduksi pada rentang kisaran keadaan hgkungan yang luas termasuk lahan bermasatah dan kering (Ghosh
d.,
1988).Sekitar 90% luas tanaman ubijalar di dunia terdapat di Asia, 5% di Afiilca
dan 5% lagi di negara-negara lain Cina dengan produksi per tahun sekitar 100 juta ton merupakan negara penghasil utama, diikuti Indonesia, Uganda, Vietnam, India dan Jepang yang mengbasilkan sekitar satu sampai dua juta ton (Horton @ gJ., 1989).
Pada umumnya tanaman ubijalar dipandang sebagai tanaman yang hanya
m e n g h a s h karbohidrat. Horton &
d.
(1989) mengungkapkan beberapa keunggulan nilai nutrisi ubijalar dibandingkan dengan tanaman pangan W y a , terutama karena ubijalar menghasilkan kadar bahan kering dan energi yang dapat dicerna tertinggi Walaupun kandungan protein umbi relatif rendah, kualitas proteinnya tinggi sehingga menghasilkan jumlah protein tercerna cukup tinggi pula (Tabel 1 ).mgI100 gram bahan segar. Kandungan tersebut lebih tinggi dari yang dirniliki tanaman ubi-ubian fainnya dan beberapa tanaman sayuran utama Kultivar ubi- berdaging umbi jingga mengandllng S-karoten tinggi, sehingga dapat mencegah Illesalah kekurangan vitamin A (xeroptalxnia). Selain itu ubi rebus merupakan sumber yang cukup b d c
uIltuk
thiamin (0.09 mg/100 g). nibovlafin (0.06 @lo0 g), niacin (0.6mg/I 00 g), d a n K (243 4 100 g),
P
(47 mg/100 g), Fe (0.7 4 1OO), Ca (32 rng/100 g) dl- dengan yang terkandung di dalarn nasi berturut-turut sebanyak 0.02mg/100 g, 0.01 mg/100 g, 04 mg/IOO g, dan 28 mgf1OO g, 28 mg/100 g, 0.2 rng1100 g, 10 mg/lOO g.
Tabel 1. Nilai produksi, energi dan protein tercerna beberapa tanamao pangan (Horton,
eta,
1989)Selain sebagai pangan, ubijalar sangat berpotensi sebagai pakan maupun bahan
baku iedustri Potensi untuk bahan
baku
i n d b d i p e n g d oleh keragamankomposisi kandungan kimia umbii Komposisi kimia umbi ubijalar sangat beragam
tergantung pada kultivar, iklirn, umur panen dan lama penyimpanan setem panen (Collins, 1988). Dikemukakan bahwa kadar bahan kering umbi berkisar antara 14.7-
Tanarnan
Kent-
Ubijalar
Singkong Padl pisang --% tzmah
Energi Tercema (juta kkal/ha)
7.1 12.6
7.3
7.1
3.9
4.1
Bahan Kering
(tma)
2.3 4.0 3.4
2.6 1.5
0.9
Protein Tercerna
@a)
1% 187
9
39.9%, sedan- protein 0.5-10.4% pati 8-29%, gula total 5.6-46.8% dan minerd 0.9-1.4% bahan kering. W u m merupakan mineral utama di dalam ubijabr. Kandungan mineral K, P, Ca, Mg, Na, S, dan Fe di dalam
umbi
berturut-turut 373, 49, 30, 24, 13, 29, dan0.8 mgll00 g .Kadar gula umbi merupakan s a h h satu War penentu kuaIitas ubijalar untuk
tujuan kegunaan tertentu. Penelitian AVRDC (1983) terhadap sernbilan kultivar yang
berkadar gula 13.8-28.2% menunjukkan sukrosa merupakan gula utama (9.5-17.3%), sedangkan glukosa (0-2.0%), manosa (0-4.9%), arabiiosa ( 0 4 % ) dan rafinosa (0-
1.1 %) ditemukan dalam jurnlah sedikit.
Kadar B-karoten merupakan salah satu keunggulan ubijalar sebagai sumber
vitamin A. Dari total pigmen karotenoid di dalam ubijalar, Bkaroten mendcup 90%.
Varietas 'CentenieP dan 'Julian' yang b e r d a g i i jingga bextmut-turut berkadar karoten 30 04 1
IU
(1 8.3 @lo0 g bahan segar) dan 40 000IU
(Wang, 1982).Tepung ubijalar memiliki kegunaan yang sangat luas untuk keperluan industri kbk% h-nmi, pangan, minumao non dkohol, dan kosmetik (Collins, 1984). Setiap 100 kg tepung dapat menghasilkan 14.5 liter etanol, d i idengan 1 1.4 liter dari kentang, 11.9 liter dari gula bit, dan 17.6 liter dari gandum Produksi ubijalar tiap hektar Iebih tinggi dari pada jenis-jenis tanaman tersebut sehingga sangat berpotensi sebagai penghasil etanol (Clark dan Moyer, 1988).
gula fivktosa diperlukan 800 kg tepung ubijalar (Soenajo, 1984). Di Fipina telah diproduksi minuman ringan dari gula W o s a ubijalar dengan warna
aLuni
jmggadengan kadar karoten 2000
IU
(Truong dan Fermentira, 1988).Butiran tepung ubijalar berbentuk poligon dan berukuran 10-25 p a Kadar
amilosa berkisar antara 18-22s. Tepung ubijalar dapat membentuk gelatin stabil dengan daya rekat tinggi dan memenuhi baku mutu untuk produksi makanan, industri biskuit, atau konveksi (Ghosh
a
d.,
1988).Penyakit Kudis Ubijalar
Penyakit kudis ubijalar pertama kali dilaporkan dari Taiwan dalarn tahun 193 5 . Agen penyebab kudii daun adalah cendawan Elsinoe batatas (stadia imperfektii Sphacelorna batatus Saw.) (Jenkins dan Viegas, 1943). Persebaran penyakit kudis diketahui telah meluas di Pasifik, Asia Tenggara, Jepang, Cina, Nigeria, B r a d dan Meksiko (Wilson, Taofatofk, Pole dan Smit, 1988). Diperkidcan penyakit ini akan
lebih mengganas apabii pembudidayaan me- teknologi intensif berskala besar untuk tujuan ekommi (Clark dan Moyer, 1988).
11
Kerugian yang ditimbuIkan berupa penurunan produksi umbi 20-50% pada
klon ubijalar rentan (Goodbody, 1983; R;unsey, Vawdrey, clan Hardy, 1988; Wilson et
d.,
1988). Percotman di Papua Nugini menunjukkan bahwa penyakit kudis -rnenurunkan jumlah umbi 45 %, bobot umbi 26.5 %, produksi total 57 % dan umbi yang &pat dipasarkan 34 % (Goodbody. 1983; Kanua dan Floyd, 1988). Di Indonesia klon ubijalar rentan yang terserang kudii menurunkan produksi sebesar 30% (Soenarjo, 1984).
Pendekatan pemuliaan ke arah varietas resisten sangat diutamakan (Wilson al., 1988). Ketangkaan informasi pola pewaxisan genetik ketahanan dan keragaman -
tkiologi cendawan Elsinoe batatas merupakan faktor penghambat ubma pemuliaan ubijalar tahan kudis (Sajise and Capuno, 1990).
Seleksi ketahanan pada taraf kecambah biji hasil silangan dengan inokulasi buatan Elsinoe baratus dalam program pemuliaan ketahanan di Tonga menunjukkan pemisahan 32.5% sangat teriufkksi 27.5% terinfeksi sedang. dan 40% tidak terinfeksi (Wilson
d.,
1988). Hasil tersebut rnenunjukkan bahwa petuang untuk merakit klon ubijalar resisten penyakit kudis cukup terbukaStudi Genetik dan Pemuliaan
Ubijalar
tanaman berbunga dominan terhadap yang tidak berbunga. Pigmentasi htang, tangkai daun dan daun dominan terhadap yang tanpa pigmentasi (Pook, 1952). Daghg umbi putih dominan tidak pen& terhadap yang b e r m Pi- daging umbi dikendalhm oleh beberapa gen aditif (Hernandez, Hernandez, Constantin dan Miller. 1965).
Daya hasi dan jumlah umbi per tanaman berkorelasi positif (Warid, Dahmani
dan Kushad, 1976). Kadar bahan kering berkorelasi positif dengan kadar pati umbi (Jones, 1977). Kadar pati d i n g m u h i secara aditif oleh poligen ( S a w 1964). Jones (1986) mengungkap bahwa kadar bahan kering berkorelasi positif dengan daya hasil, warna daging umbi kadar gula, dan pati. Kadar protein berkorelasi negatif dengan daya hasii tetapi berkorefasi positif dengan kadar karoten. Dengan demikian pemuliaan ubijalar &pat diarahkan untuk merakit varietas dengan daya hasil dan
kadar pati tinggi atau h t a s dengan kadar protein dan karoten tinggi, dan keduanya memiliki kadar bahan kering tinggi.
Kadar gula umbi ubijalar dipengaruhi o k h kehadiran enzim 8-amilase. Rasa umbi manis disebabkan oleh keaktii enzim tersebut yang menghidrolisis pati menjadi maltosa selama umbi disimpan atau diolah Klon ubijalar beragam dalam sifat
rasa manis. Penelitian Kumagai, Umemura, Baba dan Iwanaga (1991) meng- unghpkan bahwa pewarkan sifat rasa manis yang diukur dengan kebadiran enzim
B-
13
ubijalar. Pernuliaan ubijalar ke arah rasa manis rnaupun tidak manis relatif mudah dilakukan.
Hammet, Heinande d a . Miller (1966) mengemukakan bahwa keragaman
kadar serat umbi d i p e n g d oleh dua gugus gen, satu mengedakan kehadiran serat
dan yang lainnya mengersdalikan ukuran serat. Ukuran serat dikendalikan oleh
beberapa gen dominan yang diwariskan secara sederhana. Kadar serat total dikendalikan oleh beberapa gen yang terpaut dengan gen-gen ukuran sera.
Constantin, Jones, dan Hernandez (1 977) rnengernukakan bahwa pemberian kalium 0-140 k g h a di empat lokasi percobaan lebih berpengaruh terhadap komponen kualitas umbi d l k d i n g k a n dengan pemberian fosfat 0-73.90 kgha. Peningkatan pemberian kalium menuruDkan kadar bahan kering dan sedikit * &&m kadar
serat kasar. Pemberian pupuk K dan
P
bersarna-sama tidak mernpengaruhi kadar karoten dan kadar serat kasar umbi. Dikemukakan pula bahwa walaupun terdapat keragarnan, secara u m u m lokasi dan tahun tidak mempengaruhi kualitas umbiKeterwarisan merupakan ukura. hubungan antara nilai enotipe dengan nilai pemukan. Nilai keterwarisan
(h2)
beberapa sifat tanaman ubijalar tercantum dalam Tabel 2. Keterwarisan diukur sebagai nisbah ragam &hat per- genotipew>
terhadap total ragam fenotipe(Vd.
Jones (1 986) memegasken bahwa nilai keterwarisan tidak mengukur siht yang
14
buruk bahan pemuliaan, kecuali bahwa tetua superior cenderung rnenghasilkim zuriat superior.
Tabel 2. Nilai keterwarisan (b2) beberapr sifnt ubijalar (Jones, 1986)
Analisis Kestabilan Genetik Sifat
Daya Hasil Bobot Umbi Warna Kulit Umbi
Warna Daging Umbi
W a r Bahan Kering Kadar Pratein Kadar Serat Panjang Batang Resistensi
Cylas puncricollis
Percobaan pengujian kuhivar di beberapa hghmgan yang berbeda mem-
bangkitkan data fenotipe daya hasi dan siht-siht lain yang diamati Data tersebut muncuI sebagai & i t pengaruh bersama faktor genetik dan Lingkungan selama perhunbuhan dan perkembangan tanaman. Saling pengaruh antara kedua faktor tersebut dikenal sebagai interaksi genotipe x hgkungan (GxL) <Moll dan Stuber,
1974).
h2
( O h )
41 4 1-+4 44 81 66 53k14 65+12 57 81 60 84 Teknik Statistika Var-Cov Regresi Var-Cov Var-Cov Var-Cov Regresi Regresi Var-Cov Var-Cov Var-Cov Var-Cov Pustaka
Pennasabhan interaksi GxL telah lama menjadi perhatian pemulia tanaman
da- lam mengernbangkan varietas baru. Interaksi yang nyata
untuk
sifat kuantitatifmempengaruhi ketidakstabilan peragaan kultivar di berbagai lingkungan dan mengurangi peran nilai tengah dalarn program seleksi. Interaksi GxL mengurangi korehsi antara genotipe dan fenotipe, dan menurunkan kemanjuran seleksi kdiivar unggul (Cornstock dan Moll, 1963; Kang dart Martin, 1987).
Batasan kestabilan kultivar dapat ditinjau dari dua ski, biologi atau agronomi (Becker, 1981). Dalarn konsep biologi, suatu kultivar dinyatakan stabil bih selalu
meragakan daya hasil yang tetap di berbagai keadaan lingkungan tumbuh. Konsep ini sejalan dengan konsep homeostatis. Kultivar demikian mermlila
.
.
ragam antar- lingkungan kecil (Lin, Binns dan Lefkovitch, 1986). Kestabilan berdasarkan konsep biologi kurang mendapat perhatian agronomiwan. Kultivar yang berperagaan tetap di semua lingkungan tidak tanggap terhadap tindakan perbaikan hgkungan tumbuh yang bertujuan meningkatkan daya hasil. Dalarn konsep agronomi suatu kultivar dinyatakan stabil apabila daya basil optimal diragakan sejalan dengantaraf
kesubxan tiap lingkungan dan interaksi genotipe x hgkungan tidak nyata. Tanggap tiap kultivar di setiap lingkungan sejajar dengan tanggap rata-rata semua WiC
r
n
&aJ., 1986).
Berbagai metode telatt dikembangkan untuk menguraikan i n t e n h i GxL agar
(Jowet, 1972; Tai, Rice, Chew dan Miller, 1982; Singh d m Bains, 1984; Kang dan Martin, 1987; Mahahkshmi
g
&l.. 1992; Ablen, Buzzell, Benersdorf dan Allen,1994, Stoffela &
&.,
1995). dan beberapa parameter kestabilan telah diandingkan satu sama lain secara statistika(Li
a
a.,
1986; Guilan ga,
1990; Helms, 1993; Ngeve dan Bouwkamp, 1993).Beberapa statistik yang dikemukakan untuk menduga kestabilan adalah : 1 . Rataan Komponen Ragam ( 8,).
Plaisted clan Peterson (1959) rnembuat analisis ragam dari setiap pasang genotipe untuk menduga ragam interaksi masing-masing k o m b i i i pasangan genotipe. Rata-rata ragam interaksi dugaan dari semua pasangan genotipe yang mendcup genotipe i merupakan indiitor sumbangan genotipe terhadap total interaksi GxL dan menjadi ukuran k e s t a b i untuk genotipe i.
2
2. Ekovalensi, Wi
.
Wricke (1962) mengajukan analisis yang mirip dengan Plaisted dan Peterson (1959). Dikemukakan bahwa sumbangan suatu genotipe terhadap jumlah kuadrat interaksi dalam analisis ragam dua-arah &pat dijadikan ukuran k e s t a b i i Pengaruh interaksi GxL untuk genotipe ke-i di setiap Kn&mgan yang
dikuadratkan dan dijurnlahkan mempakan ukuran kestabilan untuk genotipe
17
3. Statistik Kestabilan a, dan rj.
Tai (1 97 1) menggunakan teknik regresi antara genotipe dengan indeks bgkungan
yang mirip dengan yang dikemukakan oleh Eberhart dan Russell (1966).
Perbedaan terletak pada metode penyuaian. Tai me- pendugaan kesesuaian optimum suatu hubungan struktural.
4. Ragam Kestabilan ot2.
Shukla (1972) menunjukkan bahwa ragam genotipe antar-hgkungan merupakan ukuran kestabilan suatu gewtipe.
5. Koefisien Keragaman (KKS.
Francis dan Kannenberg (1978) menggunakan koefisien keragaman
(YO)
setiap genotipe sebagai s u a t u ukuran kestabilan gewtipe.Metode regresi telah digunakan untuk menganalisis k e s t a b i i genotipe (Mooers, 1921; Yates dan Cochran, 1938; Finlay dan W-n, 1963). Eberhart dan
Russel (1966) mengemukakan regresi tiap varietas terhadap indeks hngkungan dan
suatu h g s i kuadrat sjmpangan dari regresi tersebut dapat dipakai sebagai parameter untuk menduga k e s t a b i varietas yang diuji pada beberapa Parameter tersebut dibataskan dengan model,
Y,, = rata-rata varietas ke-i pada bgkungan ke-j
(k1.2 ,..., g; j=lJ
,...,
n)p, = rata-rata varietas ke-i di sernua h g k u n g a n
S,, = regresi varietas ke-i pada lingkungan ke-j.
I,
-
indeks hgkmgan yang sebagai -an rata-rata semua va-rietas di suatu lokasi terhadap rata-rata umua
Dua parameter kestabilan diajukan. Pertama, koefisien regresi lingkungan yang &pat diduga dengan,
ZY,,I, = jumlah hasid kali rata-rata varietas ke-i pada lingkungan ke-j dengan indeks -an, dan ZIZj = jumlah kuadrat indeks hgkuugan ke-j.
Ni
j& hasilksli
dapat diperaleh dengan pendekatan :[x3F,I = [~YijI,l
[XI
= matriks ratarm tiap varietas di tiap lokasiiF,]
= vehor indeks hgkungan ke-j,EZYi,IjI = vektor j& W d kali rata-rata vdetas ke-i pada lingkungan ke-j
dengan indeks lingkungsn.
Kedua, sinpangan kuadrat tengah ( s ~ , ~ ) dari regresi linear,
2 2 2
s
,,
= XS d ( n - 2 )-
s,
/r,...
-
...
..
....
(2)XS
',
= [ZY*,-
y a + / n )-
(ZY~I,)~~,,
clans l ~ r = penduga galat gabungan.
19
tidak bebas (ragam akibat regresi) dan yang menerangkan ragarn akibat s a a n
terhadap kelinearan. Atau dapat dinyatakan sebagai : a2u = a2 regresi +cr2 s w a n
2
BAHAN
DAN METODE
Percobaan 1
Seleksi Kloo Ubijalar untuk Produksi Tinggi, Kualitas Umbi Baik, Berperagaan Stabil Antar Lingkungao dan Resisten Kudis Daun Ubijalar
BaLan Tanaman
Bahan tanaman terdiri dari 28 klon ubijalar Indonesia dan dua varietas pembanding : varietas nasional Prambanan dan varietras introduksi SQ-27. Ke-28 klon lokal terdiri dari 1 1 klon asal Sumatera, I3 klon asal Jaw* dan 4 klon asal lrian Jaya (Tabel Lampiran 1). Klon-klon tersebut dipilih dari 110 kIon bahan penelitian Hibah Bersaing LE'B yang berturut-turut telah dievaluasi di lokas'i Bogor clan Manokwari dalam tahun 1992 dan 1993 (Hartam, 1993,1994 dan 1995).
Lokasi Percobaan
Percobaan dilakukan di 6 lokasi, 3 di Jawa Barat dan 3 di lrian Jaya Lokasi
di Jawa Barat terdiri dari Kebun Percobaan IPB Darmaga (240 m dpl) dan Kebun Percobaan Balittan Muara (260 m dpl.) di Bogor, serta Kebun Percobaan Balittan Pacet (1 130
rn
dpL) di Cianjur. Lokasi di Irian Jaya terdm dari Kebun PercobaanDinas Pertanian di Prafi (45 m dpL) clan Kebun Percobaan Faperta Uncen Menggoapi
(1 10 m dpL) di M a n o w serta Kebun Percobaan DinasPertanian di Wamena
Metode Percobaan
Percobaan di 6 lokasi dilakukan selama 2 musim tanam (MT) atau pada 12 Lingkungan tumbuh MT-1 berlangsung pada bulan Juli-November 1994 dan MT-2
pada bulan Januari-Mei 1995. Pelaksanaan percobaan diperlakukan sama di semua
lingkungan tumbuh. Percobaan menggunakan rancangan acak kelompok lengkap dengan tiga ulangan. Setek pucuk ubijalar panjang 30 cm ditanam dalam bedeng berukuran 6x1 meter persegi dengan jumlah 18 tanamanlbedeng. Pernupukan dengan pupuk urea, fosfat (P2051, c l a n kaliurn (KCl) menggunakan dosis 90-45-90 k g h Pengendalian penyakit tidak
d
i
selama pe&sanaan percobaan untuk memberikan peluang tanaman terserang kudii dam ubijalar secara alamiUntuk tujuan pengamatan, dua tanaman terluar di setiap bedeng tidak diikutsertakan Sebanyak 5 tanaman contoh dipilih secara acak dari I6 tanaman percobaan Pengarnatan dilakukan terhadap peubah tanaman maupun lingkungan tumbuh
Peubah Tanaman
1. Karakter morofologi.
Sebanyak 25 karakter morfologi (Tabel Lampiran 2) diamati dengan meng-
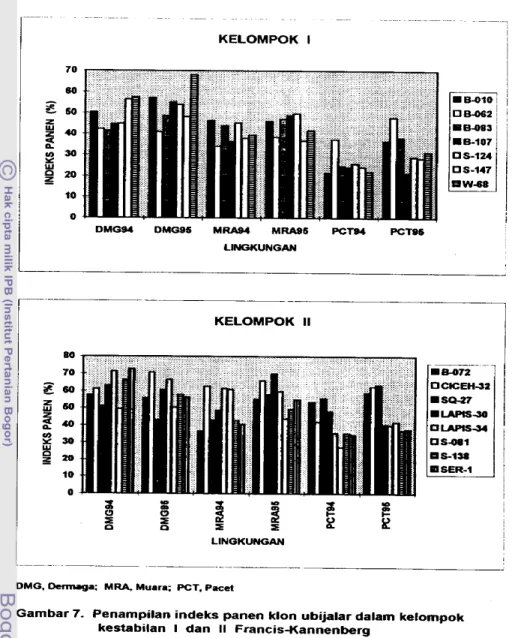
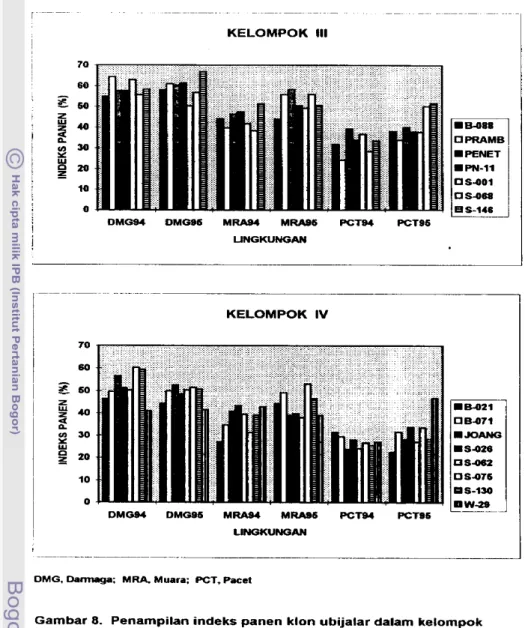
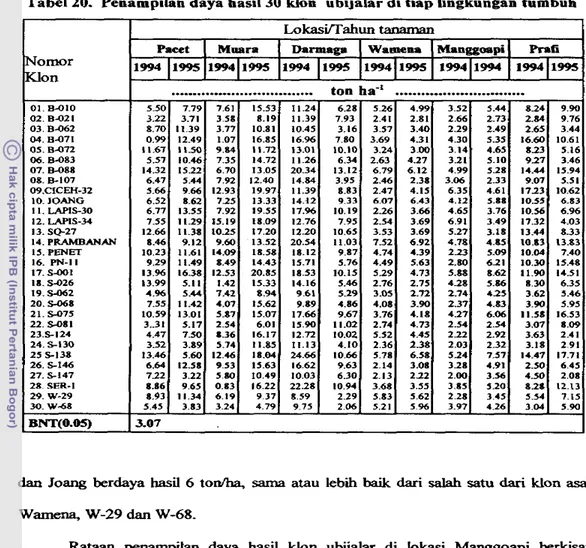
gunakan petunjuk deskriptor ubijalar (CIP, AVRDC, IBPGR, 199 1). 2 . Indeks panen (% bobot basah).
lndeks panen (IP) adalah nisbah hasid urnbi terhadap hasil biornassa (Donald
IP = (bobot u r n b i i b o t umbi
+
batang + bun) x 1W/o 3. Daya hasil (tonlha).Daya hasil dihitung dari bobot umbi &pat d i k a n per bedeng dan dinyatakan daIam ukuran tonha. Umbi yang dapat dipasarkan adalah yang berbobt di atas
1 SO gram clan berpenampilan baik (Wargiono, 1980).
4. Skor m a npenyakit
Ketahanan alami penyakit kudis diukur dengan intensitas serangan be- 5 (Tabel Lampiran 3)-
5 . Kadar bahan bering (%)
Umbi tanarnan contoh masing-masing klon dalam tiap ulangan digabung. Ujung- ujung umbi sebesar 20% dibuang. Dari bagian umbi terpilih dibuat sayatan tipis melintang dengan aIat pembuat kripik. Sayatan umbi basah diimbang 500 gram sebagai bobot basah awd (Bb) dan dijemur selama dua hari di bawah cahaya matahari. Pengeringan dilanjutkan rnenggtwbn mesin pengering daIam s d u 60% selama 48 jam. Bobot kenhg akhir (Bk) ditimbang. Kadar baban kering (BK) bahan segar d i n y a w sebagai :
BK = B k (g)/Bb (g) x 100%.
dan Biokimia, PAU Pangan dan Gizi IPB. Analisis dilakukan dengan menggunakan metode
baku.
6. Kadar 0-karoten (mg/lOO g bahan segar)
Pengamatan kadar D-karoten dan protein berdasarkan bahan =gar hanya
d iuntuk klon ubijalar berdaging umbi jingga 7. Kadar protein (% bahan kering)
8. Kadar gula total (% bahan kering)
Peubab Lingkangan T u m b u h :
Peubah h g k u q a n yang diamati meliputi komponen iklim rata-rata bulanan
curah hujrtn (mm), suhu maksirnum dan rninbum
( c ) ,
kelem- nisbi (%), lamapenyinaran (% hari). Peubah kesuburan tanah yang &adisis meliputi pH ( H f l ) , kadm C-organik
(%I,
P-tersedia @pm), basa K, Ca, M g dapat ditukar (me/100 g), dan kapasitas tukar kation (me/lOO g).24
1. Analisis Komponen Utama
Analisis korespondensi diiakukan terhadap karakter kualitatif morfologi dan komponen utama untuk karakte?. kuantitatif (Iezzoni and Pritts, 1991). Wis
menghasii rnatriks korelasi yang terdiri atas koreiasi fenotipe antar semua
kemungkinan pasangan hgkungan. Nib& keragaman total dihitung dari setiap komponen utama, yaitu nisbah akar cirinya terhadap junk& akar cfi digunakan untuk memntukan jumlah kornponen yang dilibatkaa Hanya komponen yang lebih besar dari 15% dari keragarnan total digunakan untuk analisis lanjutan.
Lkgkungan diplot terbadap korelasi balik setiap komponen utama Hasil yang
diperoleh merupakm korelasi antam rata-rata klon dalarn satu Lingkungan dan skor klon pada komponen utama Dalam plot ini, lingkwgan yang memberikan tanggap klon yang mirip akan berdekatan. Kemiripan yang tergambar dalarn hasil analisis komponen utama d i idengan gambanrn yang d i h a s i i dari analisis gugus.
2. Analisis Ragam dam Komponen Ragam
Won ubijalar yang diuji terpilih secara acak dari populasi koleksi k e j a Penelitian f i i Bersaing IPB dan d i i k a n sebagai contoh acak. Lokasi dan tahun tanam diasumsikan pula sebagai contoh acak dari semua kemungkioan iokasi clan tahun Oleh karena itu semua analisis ragam d h k d c m berdasarkan asumsi model a d .
dan bobot umbi per tanarnau dapat dipasarkan (BUT), analisis ragam dilakukan terhadap hasi pengamatan dari percobaan tahun pertama, 199411995, di lokasi Muara. Setiap pengamatan dilakukan terhadap lima tanaman w n t o h Model linear
yang digunakan :
yUk = m
+
g,+ r,+ (gr)q+
( ~ / g r ) # ~...
Ill
ygk
adalah
nilai pengamatan yang diukur dari tanaman contoh ke-k dari klon ke-i dalam -an ke-j. rn = rataan mum, g, = pengaruh klon ke-i, rj = pengaruh ulangan ke-j, (grIu = pengaruh interaksi antar klon ke-i clan ulangan ke-j, d m (plgr) d a b h#k
pengaruh tanaman contoh ke-k pada klon ke-i dalam ulangan ke-j. Komponen ragam diduga dari analisis ragam (Wricke and Weber, 1986).
Analisis ragam dan komponen ragam gabungan dilakukan terhadap indeks panen, days has& kadas bahan kering, kadar protein, kadar guia total, dan kadar karoten umbi klon ubijalar yang diuji. Indeks panen terdiri dari 5 data tamman contoh untuk t - p Won ubijalar dan ulangau, kadar protein dan gula total umbi d i j i k a n dalarn dua data pengukuran ulang untuk setiap klon dan ulangau, sedangkan untuk daya hasid (to-), kadar bahan kering, dan kadar karoten terdiri dari satu data
tiap klon ubijalar untuk setiap ulangan Model linear yang d iadalab :
Yy,
adalah nilai pengamatan tanaman contoh ke-m dari klon ke-i pada lokasi koj, tahun tanam ke-k clan ulang-an ke-L rn = rata-rata umum. G, = pengaruh klon ke-iL,
= pengaruh lokasi ke-j. (GL)g, = pengaruh interaksi antar klon ke-i clan lokasi ke-j. T, = pengaruh tahun tanam ke-k. (GT), = pengaruh intecaksi antara klon ke-i clan tahun tanam ke-k. Rw = pengaruh ulangan ke-1 pada lokasi ke-j clan tahun tanam ke-e
w-
adalah pengaruh galat m b a a n akibat tanaman contoh ke-m dari klon ke-ipada lokasi ke-j, tahun tanam ke-k, dan ulangan ke-I.
Analisis ragam dan komponen ragam untuk setiap lbgkurgan tumbuh dilakukan pula terhadap daya has& kadar bahan kering, kadar gula total, kadar
protein, dan kadar karoten ubi Analisis menggunakan model linear :
yw = nilai pengamatan tanaman contoh ke-k dari klon ke-i pada ulangan ke-j,
,"
= nilai tengah urnum, r, = pengaruh ulangan ke-j, e,, = pengaruh galat percobaan akibat tanaman contoh ke-k dari klon ke-i pada ulangan ke-j.Untuk tujuan seleksi klon ubijalar, pembandingan daya dan kualitas umbi antar klon me- uji beda nyata terkecil ( B N T O . 0 5 ) (Steel and Torie, 1980).
Nilai ketenvarisan (h2) rat- antar-lokasi dan di setiap Iingkungan diduga dengan menggunakan komponen ragam galat dan ragam genet* dugaan. Keterwarisan (hZ) dalarn arti luas dihitung me- rumus sebagai berikut (Abd- El Moneim, Cocks, dan Mawlawy, 1990; Caradus dan Woodiield, 1990; Rowe dan Brink. 1993):
h2, pada pe- [4] men- komponen rag- dari model [I], h2, pada
prsamaan
151
kompown ragam dari model [2], dan h2, pada3. Analisis Kestabilan
Analisis kestabilan menggunakan parameter koefkien regresi hgkungan bi
dan simpaagan rejgesi d2ii (Eberhart dan Russell, 1966) diilakukan b i interaksi lingkungan dan klon ubijalar berbeda nyata Parameter kestab'llan ketiga adalah koel%ien determinasi R ~ . Klon dengan R' lebih besar dari 50% dinyatakan stab3 (Stoffella gt
&,
1995). Parameter kestabilan keempat adalah koefkien keragaman tiap klon ubijalar antar Iingkungan-3
(Francis dan Kannenberg, 1978) yangdiitung sebagai KK, = (S(x> x 100%. Si = simpangan baku klon ubijalar ke-i dan xi = rataan
M
i
pengarnatan klon ke-i antar hgkunganSuatu Won ubijalar diiyatakan stabil untuk sifat tertentu apabii rataan klon ubijalar lebih besar dari rata-rata umum, koefisien regresi lingkungan bi=l, simpangan
regresi kecil (s2, = O), R' > 50 %, dan K, < KK rata-rata umum 4. Analisis Gugus
Analisis gugus merupakan salah satu alternatif penunjang analisiis k e s t a b i berdasarkan teknik regresi, terutama apabii banyak klon uji menunjukkan interaksi genotipe x lingkungan sangat nyata (Ghaderi, Everson, dan Cress, 1980; Carver, Smith dan England, 1987; Hanson, 1994). Hasii anal%= gugus memberikan gambaran u m u m tentang pengebmpokan klon b e r k k a n kemiripan tanggapan dan
Strategi penggugusan berhirarki dengan jarak Euclides digunakan untuk mengelornpokkan lokasi berdasarkan daya adaptasi klon dan pengelompokan klon ubijalar ke dalam gugus dengan kemiripan daya adaptasi terhadap hgkungan Analisis gugus k d a s a r k a n interaksi GxL dilakukan untuk mengelornpokkan Lokasi (Ghaderi &
d.,
1980; Carver &., 1987). Setiap lokasi dinyatakan sebagai s a t uvektor yang mu-unsurnya k h u b u n g a n dengan pengaruh interaksi GxL. Koefisien
jarak Euclides diiuga antar semua kemungkinan pasangan lokasi dalam satu ruang
berdimensi 30. Penggugusan dimulai dengan membentuk satu gugus dari dua lokasi terdekat berdasarkan interaksi GxL dan d i i j u t k a n secara berumtan sarnpai men- cakup semua bkasi clan rnernbentuk satu gugus tuggal. Untuk mengelompokka. klon berdasarkan kemiripan daya adaptasi, setiap klon dinyatakan sebagai satu vektor yang unsur-unsurnya berhubungan dengan daya tanggap fknotipe dalam setiap 12 lingkungan tumbuh Koefisin jarak Euclides antar klon ubijalar diduga berdasarkan mode1 geometri berdimensi 12.
P
ercobaan 2Evaluasi Ketahanan Alarni Klon Ubijalar Terhadap Penyakit Kudis
Baban Taoaman
Metode Percobsan
Percobaan berlangsung pada bulan November 1995-Februa~i 1996. Sebulan sebelum pot berisi tunas sebat d i p i i ke lapangan, tempat percobaan diianami klon ubijalar sangat peka, yaitu klon G-09, G-022, S-150, dan &etas Kabsan untuk memperoleh gambaran tentang inokuhun alami dan sebagai sumber inokulutn Pot
den- tanaman ubijalar berumur satu eengah butan (tinggi tunas sew 25 cm)
ditempatkan di sela bedeng sumber inoLculum Penataan mengjkuti mncangan acak kelompok dengan tiga ulangen Pengamatan dilakukan terhadap perkembangan j& bercak yang terarnati di batang setiap hari selarna 100
hari
Data &ringkas rnenjadi pengarnatan berkala lima hari. Serangan kudis dinyatakan dalam % mengikuti metode Sajise dan Capuno (1990) yang dimow. Lima skala dipakai, 0-10 bercak/30 cm, 11-20 b e r c a 3 0 cm, 21-30 bercak/30 cm, 31-40 bercaW3O cm, dan jumlah di atas 41 bercaW30 c m batang berturut-twut menggambsrkan tingkat serangan 0-20%, 21 4%, 4 1 -60%, 6 1 -80% dan diatas 800/0-Analisis Data
Dua peubah digunakan untuk me& taraf ketahanan kton, yaitu laju
perkem- bangan penyakit kudis (Nilai-r) dan daemh di bawah kurva laju perkemberngan
penyakit
( N i - A ) (Singh dan Rao, 1989). Nilai-r dan nilai-A dihitung sebagai berikut (Broscious, Pataky, dan Kirby, 1987):ti adakh waktu pengamatan ke-i setelah tanggal pengamatan pertama (hari)
dan Xi adalah persen serangan kudii pada pengamatan ke ti.
Seieksi Klon Ubijalar Loknl
Seleksi d i i s e h h untuk rnemperoleh klon ubijaiar stabd, juga untuk
penampilan karakter daya Mil dan kualitas umbi di atas rata-rata unmm serta tahan penyakit Mi.Sekksi simultan d i iterhadap peubah kestabih d m rataan
HASIL
D A N
PEMBAHASAN
Lokasi Percobaan
Seleksi genotipe berdaya adaptasi luas telah menjadi sasaran umum pemuliaan tanaman Kestabilan genotipe berdasarkan konsep agronomi untuk karakter tertentu dijadikan ukuran untuk menilai daya adaptasi d a . menyeleksi bahan genetik. Percobaan ini bertujuan menilai kestabilan beberapa karakter ekonomi klon ubijak. Seleksi terhadap k e s t a b i i klon &&an pemuliaan akan lebih berdayaguna apabii dilakukan pada rentang keadaan lingkungan yang beragam Pertimbangan untuk
memilih jumlah lokasi atau ternpat pengujian serta musim tanam dipengaruhi o b h tujuan dan strategi pendekatan pemuliaan yang akan diterapkan dan berkaitan dengan bentuk tanggap lingkungan yang diharapkan. Keputusan yang diambil mem-
pertimbangan faktor keragaman lingkungan dan harapan bagaimana klon akan
b e ~ t e r a k s i dengan -or lingkungan. PenampiIan klon di suatu lingkungan dan tanggap yang dl 'berikan terhadap perbedaan satu atau beberapa aspek lingkungan diperlukan untuk mengidentifikasi klon yang bernilai komersial atau digunakan sebagai balzan genetik
daIam
program pemuliaan. Karena itu pengungkapan keragaman hgkungan tempat pelaksanaan percobaan sangat penting.Keragamaa Lingkungaa Percobaan
Perbedaan jarak letak petak percobaan antara musim tanam pertama clan kedua di lokasi Pacet sekitar 20 meter sedangkan di lokasi lain kurang dari lima meter. M u s h tanam pertama berIangsung dari pertengaban bulan Agustus sampai dengan Desember
1994 dart diharapkan mewakili periode musirn kering. Musirn tanam kedua dari bulan Januari sampai dengan April 1995, d i p k a n sebagai periode b a . Keadaan urnurn lokasi percobaan meneakup kedudukan geografi, elevasi dan jenis
tanah
di lokasi percobaan tercantum dalam Tabel 3.Tabel 3. Kedudukan geografi, elevasi dan jenis tanah enam lokasi percobaan
BT = Bujur ~ i m u r ,
LS
= Lintang Selatan dpl = di atas permukaan lautLokasi KP. Pacet KP. Muam KP. Darmaga KP. Wameoa KP. Manggoapi KP. Prafi
Dua lokasi percobaan betternpat di dataran tin& yaitu Wamena di Irian Jaya clan Pacet di Jawa Barat, berturut-turut berelevasi 1550 dan 1150 meter di atas permukaan laut. Lokasi h y a bertempat di dataran rendah dengan kisaran elevasi 45-260 meter di atas permukaan laut. Data hasid analisis kesuburan
tanah
di lokasi percobaan dan data iklirn selama percobaan berlangsung disajikan berturut-turut dalam Tabel 4 dan Tabel 5.Sumber : Stasiun iklim di tiap lokasi ~ercobaan Sandi
PCT MRA DMG
W M N
MGP
PRF
Kedudukan Elevasi
(m dpl) 1150 260 250 1550 110 45 BT 107'.001 106*.45' 106'.45' 13S0.59' 134'.05' 133*.10' Jenis Tanah Andosol Latosol Latosol Inseptisol A l f i i l Entisol LS
Tabel 4. Data kesuburan taaab di tiap lokasi percobaan
L I I I I I I I I
Catatan : Dianalisis di Labomtorium Jurusan Tanah, Faperta IPB. KP. Pacet
KP. Muara KP. Darmaga KP. Wamena KP. Manggoapi KP. Pmfi
Analisis komponen utama terhadap data peubah h g k m g a n fisik kesuburan tanah dan iklim dilakukan untuk melacak keragaman lingkungan percobaan dan
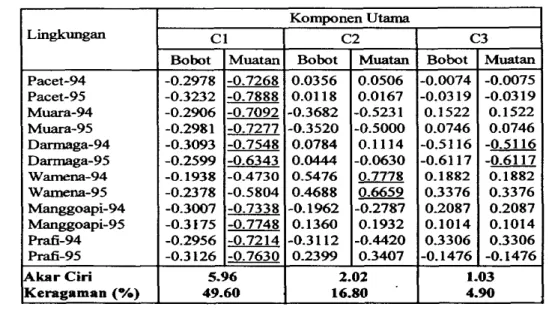
mengelompokkannya berdasarkan kemiripan aspek hgkungan. Hasil analisis komponen utama terhadap rnatriks ragam--am data hgkxmgan menunjukkau ragam pada komponen utama pertama sangat tinggii Sumbangan kedua komponen
m a m a pertama terhadap keragaman totd mencapai 99.9%.
Korelasi antara peubah awal dan komponen utruna terkait
digunakan
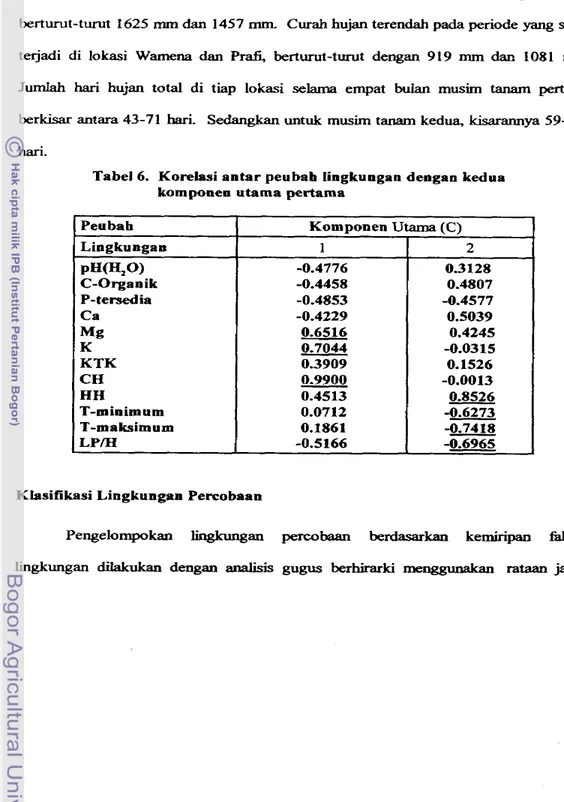
untukrnengungkap peubah yang memberikan sumbangan berarti bagi keragaman b g k u g a n Korelasi antar peubah dengan komponen utama tercantum dalam Tabel 6. U r n hara, M g , K, dan iklim, jumlah curah hujan (CH), hari hujan (HH), suhu minimum (T-minimum) clan maksimum (T-maksimum) di tiap lokasi merupakan komponen utama yang rnembedakan lingkungan percobaan. Ju& curah hujan dan hari hujan
r
Lo kasi
C-organik (Walker & Black) p H (H,O) 4.73 5.45 5.50 7.45 6.55 5.55 P-tersedia
(Bray Oben)
(%I
0.62 0.52 1.55 4.26 2.62 0.66Basa dapat ditukar
@Pm> 2.5 2.4 0.7 3.2 20.6 15.7 I
Ca
1
M g I K~ K T K
...
(me1100 g)....,
mernberikan sumbangan terbesar pada komponen utama pertama dan kedua. Kedua unsur tersebut lebih diutamakan dalam menguraikan lingkungan percobaan.
Tabel 5. Total c u r a b hujan dan jumlah hari hujan empat bnlan, rataan subu minimum dam maksimum, dan lama penyinarctn h a n a n d i tiap Liugkungan
1. KP. Pacet '94
2. KP. Pacet '95 3. KP. Muara194 4. KP. M u a r a '95 5. KP. Darmaga '94
6. KP. Darmaga '95
7. KP. Wamena '94 8. KP. Wamena '95 9. KP. Manggoapi '94
10. KP. Manggoapi '95
11. KP. Prafi '94
12. KP. Prafi '95
1
1391(
59 Sumber : Stasiun iklim di tiap lokasi percobaan.LPM
(Ye)
Lingkungan
CH = c u r a h huja;, ~ F I = j " m l a h hari hujan, T-min = suhu minimum, T-maks=suhu maksimum, LPM = lama p e n y i n a r ~ n l h a r i
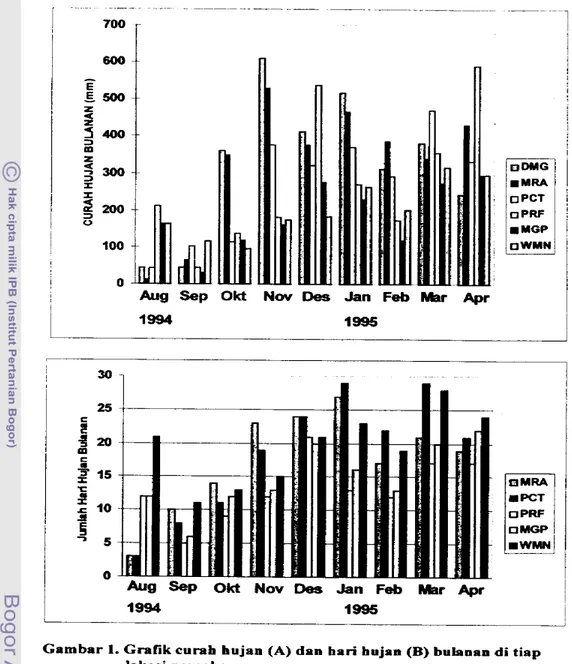
G r a pola persebaran curah hujan clan jumlah hari hujan b u h a a selama percobaan tercantum dalam Gambar 1. Gainbar 1A memperlihatkan bahwa di awal periode musim tanam pertarna, 1994/1995, percoban berlangsung pada bulan yang relatifkering. J u m h h curah hujan selama kurun waMu empat
bulan
percobam dilokasi Pacet, Wamena, Manggoapi, d m Prafi berkisac antara 526-775 mm, lebih rendah dibandingkan dengan di lokasi Muara dan Dannaga yang mencapai jumlah
1135 mm dan 1706 mm Di m u s h tanam kedua, lokasi Muara dan Darmaga tetap CH
fmm)
T-maks ("C)
HH T-min
rnenunjukkan total curah hujantertinggi selama empat bulan pelaksanaan percobaan, berturut-turut 1625 mm dan 1457 mm Curah hujan terendah pada periode yang sama terjadi di lokasi Wamena dan Praf3, berturut-turut dengan 919 mm dan 1081 mm.
JumIah hari hujan total di tiap lokasi selama ernpat buian m u s h tanam pert- berkisar antara 43-71 hari+ Sedangkan untuk mush tanam kedua, kisarannya 59-101 hari.
Tabel 6. Korelasi antar peubab lingkungan dengan kedua komponen utama pertama
Klasifikasi Lingkungan Percobaan Peubah
Lingkungan PH(H,O) C-Organik P-tersed ia Ca
M g
K KTK CH HH
T-minimum T-maksimum LPM
Komponen Utarna ( C ) 1
-0.4776 -0.4458 -0.4853 -0.4229 0.6516 0.7044 0.3909 0.9900 0.4513 0.0712 0.1861 -0.5166
2 03128
600 - -
1
fE 500 - -
Z
I
loo
Aug Sep OM Nov Des Jan Feb Mar Apr
I
I
25
i
3
29 mC
I
15
I
c 10 1I
s
OMGP5
0
I
terbobot Euclides. Dendrogram kelompok lingkungan percobaan berdasarkan jarak EucIides dan basii pemetaan skor komponen pertama (Cl) clan kedua (C2) ditampilkan dalarn Gambar 2. Kedua pendekatan memberikan has2 penggugusan yang serupa. Pemotongan hirarki pada titik h i 18 (Gambar 2A) mengelompokkan 12 Iingkungan percobaan ke dalam enam kelompok yang relatif berkemiripan Keragaman antar- kelompok 63%. Hasil pengelompokan tercantum dalam Tabel 7.
Dengan ti& memasukkan fdctor tinggi tempat dalam analisis gugus, hgkungan Wamena atau Pacet di dataran tinggi tergugus dalam kelompok yang sama dengan lingkungan percobaan di dataran rendah L i a n Wamena-94 ter- kelompok dengan Lingkungan Manggoapi-94, Wamena-95 dengan Muara-94, Pacet-94 dengan Pr&-94, dan Pacet-95 dengan Darmaga-95 dan Pr&-95. Sedangkan lingkungan Manggoapi-95 terkelompok tersendiri.
Tabel 7. Hasil peogelompokan lingkungangan percobaan berdasarkan data iklim dao kesubumn tanah
Tabel 7 rnemperlihatkan bahwa antar musirn tanam di satu lokasi tidak ter- gabung dalam satu kelompok. Hal ini menandakan m u s h tanam pertama relatif
No.
1 2 3 4 5
6
Kelompok
14 15 16 17 18 19
Lokasi Prcobaan
Pacet-94, Prafi-94
Wamena-94, Manggoapi-94 Warnena 95, Muara-94
Pacet-95, Darmaga-95, Pmfi-95 Muara-95, Darmaga-94
berbeda dengan rnusirn tanam kedua di masing-masing lokasi percobaan. Kelompok
18 berdasarkan skor komponen utama (Gambar 2B) terdiri dari lingkungan Muara-95 dan Darmaga-94 k c i r i jumlah curah hujan relatif tinggi s e w percobaan
berlangsung. Sedangkan kelompok 15 yang terdiri dari lingkungan Wamena-94 dan Manggoapi-94, bercm jumlah curah hujan relatif reladah
Dari hasil pengelompokan dengan jar& Euclides dan slcor komponen utama
disimpulkan bahwa lingkungan tempat percobaan cukup beragam. Lingkungan Pacet-95 dan Darmaga-95 di Jawa Barat yang tergabung &lam satu kelornpok (kelompok 17) dan berkemiripan dalarn hgkungan fisik kesuburan tanah, jumlah curah hujan dan hari hujan, tetapi sangat berbeda &lam elevasi, &pat digunakan
untuk memperoleh garnbaran tentang tanggap klon terhadap tinggi ternpat untuk
karakter yang diamati di lokasi iawa Barat.
Keragrmman Morfologi Klon Ubijalar
Pemuliaan tanaman bertujuan pada upaya memperbaiki kuantitas dan kualitas tanaman budidaya. Keragaman plasma nut% merupaican syarat utama untuk rnenyeleksi b h a n genetik. Klon-klon ubijalar lokal yang digunakan sebagai bahan genetik dalam percobaan
ini
berasal dari beberapa dnerah di Indonesia yang perlu diungkap keragamannya Informasi tentang keragannrn diperIukan pula untuk m e n g k l a s i i i klon ubijalar untuk tujuan pemuliaan. Hasil m i akan19 16 15 18 17
KELOMPOK LINGKUNGAM
Gambar 2. Dendrogram Jarak Euclidea (A) dan Kefompok Lingkungan Percobaan
heterosigositas tinggi. Pengungkapan derajat hubungan antar klon ubijalar di dalam populasi bahan percobaan memiliki makna penting dalam merancang strategi pemuliaan, terutama untuk pengbibridan. Klon-klon berkerabat dekat cenderung me- miliki kerniripan genetik. Upaya untuk mernasukkan satu atau dua karakter penting tertentu dan rnempertahankan karakter penting lainnya yang telah dim= suatu klon ubijaIar akan lebih tepat guna apabiia penghibridan dilakukan antar klon berkerabat dekat.
Keragaman Karakter Kualitatif Analisis Korespondensi
Deskripsi tujuh belas karakter kualitatif morfologi batang, daun dan umbi tiap klon ubijalar hasit pengamatan dari percobaan di Kebun Percobam Muara tercanlum dalam Tabel Lampiran 4. Data morfologi bunga tidak termasuk karena tidak semua klon berbunga selama masa percobaan.
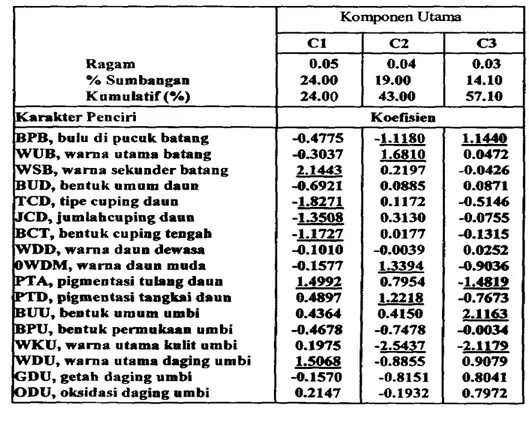
Analisis korespondensi terhadap data kualitat~f morfologi dilakukan untuk memperoleh gambaran tentang keragaman klon. Hasil analisis tercantum dalarn Tabel 8. Ketiga komponen utama pertama memberikan sumbangan 57.1% dari total keragaman. Karakter warna sekunder batang (WSB), tipe cuping d a m (TCD), jumIah
Tabel 8. Hasil analisis korespondensi uotuk deskriptor karakter kualitatif morfologi 30 k l o m ubijalar
O/o Sumbangam
ma daun dew-
,
warna daun muda A, pigmentasi tulang daunm,
pigmentasi tanghi daun 0.4897 1.2218 -0.7673BUU,
bentuk umum umbi 0.4364 0.4150 2.1163BPU, beotuk pewukaan umbi -0.4678 -0.7478 -0.0034 WKU, warna utama kulit umbi 0.1975 -2.5437 -2.1179 WDU, warna utama daging umbi 1.5068 -0.8855 0.9079 GDU, getab daging umbi -0.1570 -0.8151 0.8041 ODU, oksidasi daging umbi 0.2147 -0.1932 0.7972
-
(WCTB), wama daun muda (WDM), pigrnentasi tangkai daun (PTD), dan warm kulit umbi (WKU) menonjol pada komponen utama kedua (C2). Pada komponen utama
tiga (C3), karakter yang menonjol adalah bent.& umbi (F3UU). Ekspresi
karakter-karakter kualitatif utama tersebut dipakai untuk melakukan k l a s i i i klon ubijalar. Warna kulit dan d a g i i urnbi merupakan karakter kualitatif yang n&ndapat
karakter ini dijadikan pernbobot dalam klasi6kasi. Sebaran keragarnan beberapa karakter utama rnorfologi bat- c l a n daun klon ubijak tercantum dalam Gamba~ 3. Batang
Deskriptor untuk warna utama batang terdiri dari delapan k e h . Hasil karakterisasi menunjukkan klon ubijalar tersebar dalam enam kelas (Gambar 3A). Tidak ada klon ubijalar di dalam kelas 5 dan 9 yang berciri warna batang hijau dengan banyak bintik
ungu
tua sepanjang permukaan atau keseluruhan batangberwarna ungu sangat t u a Sebanyak 8 klon me- warna batang hijau pobs, 14
klon berwarna batang hijau dengan sedikit bintik ungu sepanjang batang, 1 klon
berwarna batang hijau dengan banyak bin& ungu muda, 2 klon dengan hampir
kesehrruh-an batang berwarna ungy satu kIon @m)e' dengan hsmpir keseluruhan
batang berwarna ungu tua, dan 4 klon (Joang. S-062, S-068 clan SER-1) dengan
keseluruhan batang berwarna ungu.
A
WARNA UTAMA BATANG
--
SKOR
-
-C PlGMENTASl TANGKAl DAUN
10 8 8
g
7 W 6 3 65
4E X
10
1 2 3 4 6 6 7 8 8
SKOR
WARNA DAUN MUDA
1
B WARNA SEKUNDER BATANG
8
T
Z 6
W
3 x 4
W
L
*
0
0 1 2 3 4 6 6 7 8
SKOR
D
PlGMENTASl TULANG DAUN AEAKSIAL
10 fl 1
SKOR
. - - - - --.. -
JUMLAH CUPING DAUN
1
SKORI SKOR
-~ .
I
Daua
Tangkai daun ubijalar bisa tidak, sebagian, atau kesehrruhan permukaan terpigmentasi antosianin Deskriptor untuk karakter pigmentasi tangkai daun ter- bagi dahn sembilan kelas. Pigmentasi tangkai daun 30 klon ubijalar tersebar dalam tujuh kelas (Gambar 3C). Ekspresi pigmmtasi dapat b e r k m d c bintik berwama ungu di pangkal dan ujung tangkaitimtik-bintik ungu sepanjang tangkai atau keseluruhan tangkai daun berwama ungu. Sebanyak 6 kIon dari populasi klon percobaan berwama
tangkai daun hijau polos. 6 klon berhngkai hijau dengan t i t i ungu pada ujung tangkai dekat pangkal daun, 9 klon mengekspresikan warna tangkai hijau dengan wama ungu pada daerah ujung dan paagkal tangkai 5 klon menampilkan warna hijau dengan
titik-tit& ungu sepanjang t a m dan 4 klon, yaitu S-062, S-068, PN-11 dan Joang,
me& keseluruhan atau hampir keseluruhan tangkai daun berwarna ungu (&or 7, 8
dan 9).
Seperti pada tangkai daun, tulang daun abaksii ubiialar bisa tidak atau terpigmentasi antosianin. BiIa tidak terpigmentasi, tulang dam berwama kuning atau hijau. Beragam ekspresi pigmentasi dapat terlihat pada tulang daun. Pigmentasi dapat berbentuk bintik wama ungu pada pangkal, s e b a g i i atau keseluruhan tulang daun utama terpigmentasi warm ungu. Selain itu dapat pula s e b a g i i atau kesehuuhan
tulang daun berwama ungu, atau keseiuruhan sisi daun bawah clan tuhng daunya
berwarna ungu. Ekspresi pigmentasi tulang daun abaksial klon uji didominasi oleh keIas skor 3 dan 8, berturut-turut wama ungu pada pangkal tulang daun utama yang
(Garnbar 3D). Tuiang daun berwarna kuning tanpa terpigmentasi diekspresikan oleh 5 klon, sedangkan sebagian tulang daun utama terpigmentasi diekspresikan oleh lima Mon lain. Dua kbn, Joang dan PN-11, menampilkan daun dewasa dan tulang daun abaksial berwarna ungu.
Warna daun muda ubijalar dikategorikan dalam sembilan kelas. Klon uji tersebar beragarn dalam tujuh kelas. Klon ubijalar tersebar beragarn dalarn tujuh kelas. Gambar 3E menunjukkan 7 klon merniliki dam muda berwarna hijau-kekuningan, 6 klon berdaun Gjau dengan pinggiran daun bewarna ungu, 1 klon dengan warna hijau keabu- abuan karena pucuk bat- berbulu lebat, 5 klon berdaun hijau dengan tulang dam -a1 ungu, 6 klon dengan daun mu& berwarna hijau-keungwn dan 3
klon ungu-kehijauan, serta 3 klon yaitu Joang, PN-11, dan W-68 dengan kedua sisi daun muda berwarna ungu tua Di dalam populasi kJon uji tidak terdapat Won dengan ciri sisi atas daun muda berwarna hijau dan sisi bawahnya ungu.
bercuping agak dalam, dan dua klon bercuping dalam. Tiga klon bercuping tujuh, dua klon bertipe sedang dan satu klon bertipe dalam.
Sebaran keragaman karakter morfologi bentuk, warna kulit dan daging umbi tercantum dalam Gambar 4. Bentuk urnbi terbagi dalam sembillan kelas deskriptor, yaitu membundar, menjorong bundar, menjorong, membundar tehrr, memburKlar telur sungsang, melonjong, melonjong panjang, menjorong panjang, atau memanjang takteratur. Hasil karakterisasi (Gambar 4A) mengidentitlkasi 12 klon ubijalar memiliki bentuk umbi menjorong panjang, 8 klon dengan bentuk umbi menjorong bundar, 4 klon menjorong, 3 klon membundar telur sungsang, dan berturut-turut satu klon me^ bentuk umbi mernbundar telur, melonjong, atau memaajang takteratur.
UbGalar rnemiliki ernpat warna utama kulif dan daging urnbi yaitu put* kuning, jingga, atau ungu. Populasi klon ubijalar terdiri dari 14 klon berkulit umbi put& 7 klon berkulit jingga, dan 9 klon berwarna kulit ungu (Gambar 4B). Sedangkan untuk warm daging umbi, 9 klon berdaging putih atau gad@ (skor 1, 2, 3). 4 klon berdaging kuning pucat, 13 klon jingga (skor 6, 7, 8), d m 4 klon ungu (Gambar 4C). Semua kion ubijalar berdaging ungu berkuiit umbi putih. Klon ubi- yang berdagimg umbi putih atau gading terdiri dari enam klon berkulit umbi putih clan tiga ungu, sedangkan yang berdaging umbi jmgga terdiri dari yang berkulit putih, jingga dan
* 2 3 4 5 6 7 8 8
SKOR
' 8
WARNA KULrr U111BI
1 2 3 4 5 6 7 8 8
SKOR
. - - -
WARNA DAGlNG UlllBl
7
5 "
2 5
J
Y 3
W
E t
0
1 2 3 4 5 6 7 8 9
SKOR
KIasifikasi Klon Ubijalar
Untuk mengungkap derajat hubungan antar klon ubijalar, k l a s i a s i berhirarki menanjak berdasarkan jarak Euclides d idengan menggunakan rata-rata jarak terbobot sebagai kriteria penggugusan. Dendrogram klasifikasi jarak Euclides dan analisis korespondensi tercantum d a b Gambar 5.
Pemotongan gugus hirarki klon ubijalar pada titik h s i 55 dengan taraf ketakrniripan 85 % (Gambar 5A) menghasilkan 6 keiompok klon ubijalar. Hasil pengelompokkan 30 klon ubijalar yang ditetiti tercantum dalam Tabel 9.
Tabel 9. Kelompok klon ubijalar berdasarkan deskriptor karakter morfologi kualitatif
Kelompok I
Klon-klon ubijalar dalarn kelornpok-I berkerniripan dalam karakter warm utama batang, tipe cuping dan jumlah cuping daun. Won-klon menampilkan warna
Kelompok I
I1
I11
Jumlah 13
5
9
Sandi Klon Ubijalar B-010, B-071, B-083, B-088, B- 107, Ciceh-32, Prambanan, S-001, S-026, S-07.5, S-138,s-146, W-29
8-02 I , Lapis-30. Lapis-34, Penet, S- 124 B-062, B-072, SQ-27, S-062, S-068 S-130, S-147, SER-I, W-68
Joang PN-11 S-08 l IV
V
VI
KELOMPOK KLON
I
I
C
batang hijau polos atau hijau dengan sediit bmtik-bmtik pigmentasi ungu. Jurnlah cuping d a m satu atau tiga, dengan tipe cuping dangkal atau sangat dangkal. Tiga belas klon ubijalar tergabung dalarn kelompok ini. Berdasarkan warna daging umbi, empat klon uji berdaging ungu yaitu B-010, B-083, W-29, dan B-107; delapan dari 13 klon berdaging jingga, B-088, S-075, S-001, 5-026, S-138, Prambanan, B-071, Ciceh-32; dan satu klon, S-147, berda-ging umbi gading menjadi anggota kelompok I yang terbagi dalam dua anak gugus.
Keempat kIon b e r d a g i i ungu dan klon B-088 berda- ging umbi j i g g a menjadi anggota anak gugus pertama. Semua klon ubijalar b e r d a g i i ungu selain berkulit umbi putih, bercuping daun tiga, dan tulang daun abaksii terpigmentasi bmtik ungu di pangkal tulang utama. Klon B-010, B-083 clan, W-29 me* bentuk umbi rnenjorong panjang, sedangkan untuk 8- 107 menjorong bulat. KIon B-010
asal
Bogor tanpa catatan nama daerah dan klon B-083asal
Margasari Jawa Barat dengan nama daerah 'mantang-bm' yang paling berkerniripan d a b anak gugus ini. Kedua kIon ubijalar tersebut tergabung pada titik h i 33 (Gambar 5A) dengan derajat kemiripan 60%. Klon W-29 asal lrian Jaya dengan nama daerah 'tinta' berbeda dengan kedua klon berdaging ungu tersebut untuk karakter warna sekunder batang dan warna daun mu&. Klon W-29 rnengekspresikan pucuk batang dan dam muda berwarna ungudaerah 'dayak'. Kedua klon bergabung pada titik h i 36 dengan derajat kemiripan klon 56%.
Anak gugus kedua terdiri dari tujuh klon ubijaIar berdagiig jingga clan satu
klon ubijalar (5-147) dengan wama daging gading. Varietas nasional Prambanan berkulit dan berdaging umbi jingga tergabung dalam anak gugus ini. Klon-klon berwarna daging umbi jingga menunjukkan keragaman untuk karakter wama daun muda, bentuk umbi dan wama kulit umbi. Klon-klon ubijalar asal Sumatera (bersandi S) dalarn anak gugus ini menunjukkan hubungan kekerabatan yang lebih dekat. Kion S- 147 asal Nias clan S-075 asal Gido, keduanya tanpa catatan nama daerah bergabung pada titik fisi 39 dengan derajat kemiripan