POTENSI
Aspergillus
DAN
Penicillium
ASAL SERASAH
DIPTEROCARP SEBAGAI ENDOSIMBION AKAR
PELARUT FOSFAT
DEZI HANDAYANI
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI
Dengan ini saya menyatakan bahwa tesis Potensi Aspergillus dan Penicillium Asal Serasah Dipterocarp sebagai Endosimbion Akar Pelarut Fosfat adalah karya saya dengan arahan komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Bogor, Juli 2011
Dezi Handayani
ABSTRACT
DEZI HANDAYANI. Potency of Aspergillus and Penicillium from Dipterocarp Leaf Litter as Phosphate Solubilizer and Root Endosymbiont. Under direction of GAYUH RAHAYU and MIFTAHUDIN
The ability of soil microorganisms especially fungi to convert insoluble forms of phosphorus (P) to an accecible form is important for plant growth and development. Aspergillus sp. IPBCC.09.619 and Penicillium sp. IPBCC.09.620 showed phosphate solubilizing capacity in vitro. However, phosphate solubilizer capability and its endosymbiotic capacity with Zea mays and Shorea selanica root have not been studied. This study was aimed to evaluate the potency of Aspergillus and Penicillium from dipterocarp leaf litter as phosphate solubilizer and root endosymbiont. Microscopic observations of Z. mays and S. selanica seedlings before fungal inoculation showed that Z. mays roots were free from endosymbiotic fungi while those of S. selanica bore Dark Septate Endophyte (DSE). DSE occurred on 85% of S. selanica root seedlings and it is assumed that DSE has synergetic function with Penicillium sp. Of the two fungi studied, only Penicillium sp. lives as endosymbiont. Colonization of Penicillium sp. to Z. mays and S. selanica roots started at root hairs and then penetrated epidermal cell through epidermal junction. Some hyphae became lobed within epidermal and cortical cells and fullfiled inoculated cells with tiny hyphae. After 8 weeks inoculation, hyphae came out from the epidermal cells and developed penicillate structure on root surface. Inoculation of Penicillium sp. to Z. mays root at 100% TCP dosage (equal to 60 mg P2O5/plant) increased all of growth parameters, except S. selanica shoot length. Shoot dry weight of Z. mays and S. selanica showed higest response to fungal inoculation as indicated by 82% and 54.7% increases respectively. Colonization of Penicillium sp. to both plants increased P uptake at 60 mg P2O5/plant dosage (37% and 103% higher than plant control respectively). Phosphorus uptake at 50% P dosage was not significantly different with control. It means, inoculation of Penicillium sp. makes application of P efficient.
RINGKASAN
DEZI HANDAYANI. Potensi Aspergillus dan Penicillium Asal Serasah Dipterocarp sebagai Endosimbion Akar Pelarut Fosfat. Dibimbing oleh GAYUH RAHAYU dan MIFTAHUDIN
Fosfor merupakan hara makro esensial tanaman yang keberadaannya seringkali mengalami defisiensi dalam tanah. Pemberian pupuk fosfat ke tanah dapat mengatasi masalah ini, namun tidak efisien karena fosfat dengan cepat akan mengalami presipitasi sehingga tidak tersedia lagi bagi tanaman. Sebagian bakteri dan cendawan memiliki kemampuan untuk melarutkan fosfat, sehingga dapat digunakan sebagai penyedia fosfat bagi tanaman. Bertolak dari hal tersebut maka penelitian ini dilakukan dengan tujuan untuk menganalisis potensi Aspergillus dan Penicillium asal serasah dipterocarp sebagai pelarut fosfat simbiotik akar Z. mays bebas endosimbion dan S. selanica terkolonisasi DSE.
Kemampuan kapang asal serasah dipterocarp melarutkan fosfat diketahui dengan cara menumbuhkan masing-masing isolat pada media agar Pikovskaya. Adanya zona bening di sekitar koloni mengindikasikan bahwa kapang tersebut mampu melarutkan fosfat. Dua isolat yang memiliki indeks kelarutan fosfat cukup tinggi, yaitu Aspergillus sp. IPBCC.09.619 dan Penicillium sp. IPBCC.09.620 diinokulasikan ke akar tanaman Z. mays dan S. selanica untuk mengetahui kapasitas endosimbionnya.
Sebelum inokulasi kapang, diketahui bahwa akar Z. mays bebas cendawan endosimbion sedangkan akar S. selanica membawa DSE. Persentase jumlah kecambah S. selanica yang membawa DSE adalah 85%, namun keberadaannya tidak dominan (paling banyak 15 sel akar pada setiap potongan akar yang diamati). Umumnya DSE dalam akar berbentuk mikrosklerosia yang memenuhi sel akar. Dua jenis koloni DSE (koloni hitam dan putih) berhasil diisolasi dari akar S. selanica pada akhir perlakuan. Koloni DSE berwarna hitam memiliki hifa bersekat pendek dan bercabang dengan klamidospora interkalar sedangkan koloni berwarna putih memiliki hifa halus bercabang banyak dengan klamidospora terminal dan interkalar.
Bentuk simbiosis kapang pelarut fosfat dengan tanaman uji dianalisis melalui beberapa tahap, yaitu analisis proses kolonisasi kapang pada akar tanaman uji, respon tumbuh tanaman uji yang diinokulasi kapang pelarut fosfat dengan perlakuan berbagai taraf % TCP (dinyatakan sebagai mg P2O5
Proses kolonisasi kapang terhadap akar tanaman uji dianalisis menggunakan metode pewarnaan akar biru tripan dan ditunjang dengan pengamatan mikroskop pemindai elektron. Hasil pengamatan menunjukkan bahwa Aspergillus sp. bukanlah endosimbion akar kedua tanaman uji sedangkan Penicillium sp. dapat mengkolonisasi akar tanpa menimbulkan gejala sakit pada tanaman. Kolonisasi Penicillium sp. pada kedua akar tanaman uji terbatas pada bagian epidermis dan korteks sehingga cendawan ini dapat dikategorikan sebagai cendawan endofit akar. Kolonisasi akar Z. mays dimulai dengan masuknya hifa ke dalam epidermis akar dan dilanjutkan dengan pembentukan struktur seperti noktah. Struktur seperti noktah ini bertambah besar dan banyak, kemudian berkecambah membentuk hifa renik yang menyebar dalam sel. Hifa renik selanjutnya tumbuh kembali ke
permukaan akar membentuk hifa eksternal dengan ciri khas Penicillium. Kolonisasi pada akar S. selanica sedikit berbeda dengan Z. mays. Struktur seperti noktah tidak mengalami perkembangan lebih lanjut, namun hifa eksternal tetap terlihat pada akhir pengamatan.
Respon tumbuh dan serapan hara tanaman terhadap inokulasi kapang pelarut fosfat diuji dengan memberi perlakuan berbagai taraf % TCP dalam larutan Hoagland, yaitu 0% (setara 0 mg H2PO5/tanaman), 50% (setara 30 mg H2PO5/tanaman) dan 100% (setara 60 mg H2PO5/tanaman). Sebagai kontrol, tanaman diberi perlakuan 60 mg H2PO5/tanaman, tetapi tidak diinokulasi dengan cendawan pelarut fosfat. Hasil analisis menunjukkan bahwa inokulasi dan kolonisasi Penicillium sp. dengan pemberian 60 mg H2PO5/tanaman dapat meningkatkan semua parameter respon tumbuh kedua tanaman uji secara signifikan kecuali untuk pertambahan tinggi tanaman S. selanica bila dibandingkan tanaman kontrol. Parameter respon tumbuh Z. mays dan S. selanica yang mengalami peningkatan paling besar adalah berat kering tanaman, yaitu berturut-turut sebesar 82% dan 54.7%. Inokulasi Penicillium sp. dengan pemberian 30 mg H2PO5/tanaman pada akar Z. mays dapat meningkatkan tiga parameter respon tumbuh secara signifikan (berat basah dan berat kering tanaman serta panjang akar). Hal ini mengindikasikan bahwa Penicillium sp. dapat meningkatkan efisiensi penggunaan P. Sementara itu, kolonisasi Penicillium sp. tidak memberikan pengaruh yang nyata terhadap pertumbuhan S. selanica dengan pemberian 30 mg H2PO5
Kolonisasi Penicillium sp. meningkatkan serapan P kedua tanaman uji secara nyata pada pemberian 60 mg P
/tanaman.
2O5/tanaman yaitu berturut-turut sebesar 3.7 mg dan 1.2 mg P2O5 per tanaman (37% dan 103% lebih tinggi dari serapan P tanaman kontrol). Serapan P berkorelasi positif dengan serapan N dan K sehingga memberi keuntungan lebih pada tanaman uji.
© Hak Cipta milik IPB, tahun 2011
Hak Cipta dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan yang wajar IPB
POTENSI
Aspergillus
DAN
Penicillium
ASAL SERASAH
DIPTEROCARP SEBAGAI ENDOSIMBION AKAR
PELARUT FOSFAT
DEZI HANDAYANI
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
Mayor Mikrobiologi
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Judul Tesis : Potensi Aspergillus dan Penicillium Asal Serasah Dipterocarp sebagai Endosimbion Akar Pelarut Fosfat
Nama : Dezi Handayani NIM : G351080021
Disetujui Komisi Pembimbing
Dr. Ir. Gayuh Rahayu
Ketua Anggota
Dr. Ir. Miftahudin, M.Si
Diketahui
Koordinator Mayor Dekan Sekolah Pascasarjana
Mikrobiologi
Dr. Ir. Gayuh Rahayu Dr. Ir. Dahrul Syah, M.Sc.Agr.
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah SWT atas segala karunia-Nya sehingga karya ilmiah ini berhasil diselesaikan. Tema yang dipilih dalam penelitian yang dilaksanakan sejak bulan September 2010 ini adalah kolonisasi cendawan pelarut fosfat, dengan judul Potensi Aspergillus dan Penicillium Asal Serasah Dipterocarp sebagai Endosimbion Akar Pelarut Fosfat.
Terima kasih penulis ucapkan kepada Ibu Dr. Ir. Gayuh Rahayu selaku pembimbing I sekaligus pemberi dana penelitian, Bapak Dr. Ir. Miftahudin selaku pembimbing II, serta Bapak Dr. Ir. Anas Dinurrohman Susila, M.Si. yang telah banyak memberi saran. Terima kasih kepada Ade, Lia, Dita, Riana, Rina dan teman-teman Laboratorium Mikologi yang tak bisa disebutkan namanya satu persatu atas kerjasama dan hari-hari yang menyenangkan. Terima kasih kepada suami saya Cipta Budiman, S.Si, MM. atas cinta, dukungan dan pengertiannya selama ini. Ungkapan terima kasih juga disampaikan kepada Apa, Ama, dan seluruh keluarga, atas segala doa dan kasih sayangnya.
Semoga karya ilmiah ini bermanfaat.
Bogor, Juli 2011
RIWAYAT HIDUP
Penulis dilahirkan di Bukittinggi pada tanggal 26 Januari 1977 dari ayah Asrul St. Marajo dan ibu Wilda. Penulis merupakan putri kedua dari tujuh bersaudara dan istri dari Cipta Budiman, S.Si., MM.
Tahun 1996 penulis lulus dari SMA Negeri 3 Bukittinggi dan pada tahun yang sama lulus seleksi masuk IPB melalui jalur Undangan Seleksi Masuk IPB pada jurusan Biologi, Fakultas Matematika dan Ilmu Pengetahuan Alam. Kesempatan untuk melanjutkan ke program Magister pada Mayor Mikrobiologi Sekolah Pascasarjana IPB diperoleh pada tahun 2008. Beasiswa Program Pascasarjana (BPPS) diperoleh dari Direktorat Jenderal Pendidikan Tinggi Republik Indonesia.
Penulis bekerja sebagai staf pengajar di Jurusan Biologi, Departemen FMIPA Universitas Negeri Padang sejak tahun 2006. Mata kuliah yang diampu adalah Mikologi, Taksonomi Tumbuhan Rendah dan Biologi Umum.
DAFTAR ISI
2.4 Analisis Proses Kolonisasi Cendawan pada Akar Tanaman ... 7
2.5 Dark Septate Endophyte (DSE) ... 8
3 BAHAN DAN METODE 3.1 Waktu dan Tempat Penelitian ... 9
3.2 Bahan ... 9
3.3 Metode Penelitian 3.3.1 Uji Aktivitas Pelarut Fosfat Aspergillus dan Penicillium Asal Serasah Dipterocarp ... 11
3.3.2 Produksi Inokulan Aspergillus dan Penicillium ... 11
3.3.3 Perkecambahan Z. mays dan S. selanica ... 11
3.3.4 Inokulasi Aspergillus dan Penicillium pada Akar Tanaman Uji, Pemberian Hara dan Pemeliharaan ... 12
3.3.5 Analisis Simbiosis Aspergillus dan Penicillium dengan Akar Z. mays dan S. selanica ... 12
3.3.6 Uji Viabilitas Endosimbion ... 16
3.3.7 Isolasi DSE ... 16
4 HASIL DAN PEMBAHASAN 4.1 Hasil Pengamatan 4.1.1 Potensi Aspergillus dan Penicillium sebagai Cendawan Pelarut Fosfat ... 18
4.1.2 Aspergillus sp. pada Akar Z. mays dan S. selanica ... 20
4.1.3 Penicillium sp. pada Akar Z. mays ... 21
4.1.4 Penicillium sp. pada Akar S. selanica ... 26
4.1.5 Respon Tumbuh Tanaman Uji yang Diinokulasi Penicillium sp. ... 31
4.1.6 Serapan Hara Tanaman Uji yang Diinokulasi Penicillium sp. ... 34
4.2 Pembahasan ... 35
5 SIMPULAN DAN SARAN 5.1 Simpulan ... 42
5.2 Saran ... 43
DAFTAR PUSTAKA ... 44
DAFTAR TABEL
Halaman 1 Indeks kelarutan fosfat Aspergillus dan Penicillium asal serasah
dipterocarp ... 18
2 Persentase kolonisasi Penicillium sp. 8 minggu setelah inokulasi
dengan perlakuan P berbeda ... 31
3 Pengaruh perlakuan P terhadap pertumbuhan tanaman uji yang
diinokulasi Penicillium sp. . ... 31
4 Pengaruh berbagai perlakuan P terhadap serapan hara tanaman
DAFTAR GAMBAR
Halaman 1 Bagan alur penelitian ... 10
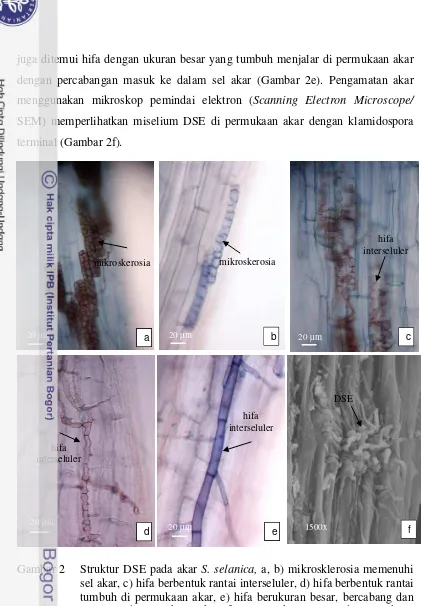
2 Struktur DSE pada akar S. selanica ... 19
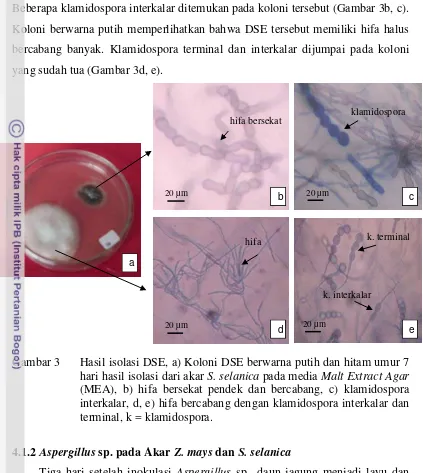
3 Hasil isolasi DSE ... 20
4 Pengaruh inokulasi Aspergillus sp. terhadap Z. mays dan
S. selanica ... 21
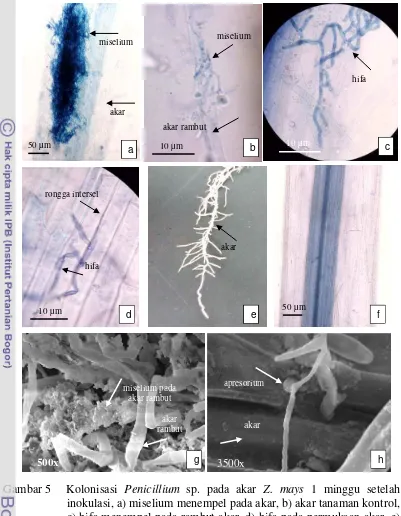
5 Kolonisasi Penicillium sp. pada akar Z. mays 1 minggu setelah
inokulasi ... 22
6 Kolonisasi Penicillium sp. pada akar Z. mays 2 minggu setelah
inokulasi ... 23
7 Kolonisasi Penicillium sp. pada akar Z. mays 3 minggu setelah
inokulasi ... 24
8 Kolonisasi Penicillium sp. pada akar Z. mays 4 minggu setelah
inokulasi ... 25
9 Kolonisasi Penicillium sp. pada akar Z. mays 8 minggu setelah
inokulasi ... 26
10 Kolonisasi Penicillium sp. pada akar S. selanica 1 minggu setelah
inokulasi ... 27
11 Kolonisasi Penicillium sp. pada akar S. selanica 2 minggu setelah
inokulasi ... 28
12 Kolonisasi Penicillium sp. pada akar S. selanica 3 minggu setelah
inokulasi ... 29
13 Kolonisasi Penicillium sp. pada akar S. selanica 4 minggu setelah
inokulasi ... 29
14 Kolonisasi Penicillium sp. pada akar S. selanica 8 minggu setelah
inokulasi ... 30
15 Kurva respon tumbuh tanaman uji terhadap dosis P ... 32
DAFTAR LAMPIRAN
1 1 PENDAHULUAN
1.1 Latar Belakang
Fosfor (P) merupakan unsur vital bagi pertumbuhan dan perkembangan tanaman dan ditemukan pada semua sel tanaman hidup. Fosfor memegang peranan penting dalam proses metabolisme energi, aktivasi metabolit intermediet, sebagai komponen aliran sinyal transduksi, elemen struktural asam nukleat dan fosfolipid (Bucher 2007). Walaupun kandungan P total dalam tanah tinggi, namun sebagian besar P ada dalam bentuk terikat dan hanya 0.1 sampai 0.5% yang dapat digunakan oleh tanaman (Pradan & Sukla 2005). Kendala ini dapat diatasi dengan penambahan P ke tanah pertanian dalam bentuk pupuk fosfat. Walaupun demikian, lebih dari 80% pupuk fosfat yang diaplikasikan dengan cepat berubah bentuk menjadi P terikat melalui reaksi presipitasi dengan ion Al3+ dan Fe3+ dalam keadaan tanah asam serta Ca2+
Alternatif lain untuk mengatasi masalah di atas adalah penggunaan
mikroorganisme pelarut fosfat. Selain mampu meningkatkan ketersediaan P dan memicu pertumbuhan tanaman, mikroorganisme pelarut fosfat juga diketahui dapat melindungi tanaman dari penyakit dan bertindak sebagai agens biokontrol (Koike et al. 2001; Shivanna et al. 1999), menghasilkan hormon tumbuh (Yadav
et al. 2011; Nenwani et al. 2010); melarutkan berbagai hara mikro (Altomare et
al. 1999); meningkatkan toleransi tanaman terhadap cekaman (Pandya & Saraf 2010) dan relatif ramah lingkungan.
pada tanah dengan pH tinggi (Hao et al. 2002; Holford 1997) sehingga penggunaan pupuk menjadi tidak efisien. Dengan demikian, pupuk fosfat harus ditambahkan secara teratur agar ketersediaan P bagi tanaman terpenuhi. Penggunaan pupuk yang terus menerus menimbulkan beberapa dampak negatif, diantaranya adalah peningkatan biaya produksi pertanian dan penurunan kualitas lahan akibat akumulasi pupuk kimiawi dalam tanah pertanian (Saraswati 1999).
2
dalam penyediaan P bagi tanaman sehingga memungkinkan untuk pemberian pupuk P secara berkelanjutan dan efisien. Mikroorganisme ini termasuk dalam kelompok bakteri, cendawan dan aktinomiset. Perubahan fosfat menjadi bentuk terlarut umumnya dilakukan melalui asidifikasi, pengkelatan logam dan reaksi pertukaran ion (Pradhan & Sukla 2005).
Cendawan diketahui memiliki kemampuan yang lebih tinggi daripada bakteri dalam proses pelarutan fosfat (Nahas 1996). Aspergillus sp. dan
Penicillium sp. secara berurutan dapat melarutkan 480 µg/ml dan 275 µg/ml fosfat
dari 0.5% trikalsium fosfat (TCP) setelah 4 hari (Pradhan & Sukla 2005). Richa et al. (2007) melaporkan bahwa A. tubingensis dan A. niger merupakan cendawan pelarut fosfat yang baik diaplikasikan pada tanah alkalin dengan sumber P berupa
rock phosphate. Hal senada juga dilaporkan oleh Barrow & Osuna (2002) yang
menyatakan bahwa cendawan dark septate endophyte (DSE) dapat meningkatkan efisiensi penyerapan fosfat pada tanaman Atriplex canescens.
Penapisan cendawan pelarut fosfat sampai sekarang terus dilakukan untuk mendapatkan isolat potensial. Cendawan ini dapat digunakan sebagai pupuk hayati untuk meningkatkan efisiensi penyerapan P dalam tanah (Kucey 1987; Barrow & Osuna 2002; Richa et al. 2007). Cendawan pelarut fosfat dapat dikategorikan sebagai cendawan mutualistik akar yang terdiri dari cendawan mikoriza, misalnya Glomus fasciculatum dan Acaulospora laevis (Sabannavar & Lakshman 2009) dan cendawan non mikoriza seperti Penicillium rugulosum
(Reyes et al. 2002), P. citrinum, Trichoderma harzianum dan Aspergillus niger
(Yadav et al. 2011). Penelitian terhadap cendawan-cendawan pelarut fosfat telah banyak dilakukan, namun lebih terkonsentrasi pada jenis cendawan mikoriza. Sementara itu, penelitian terhadap cendawan mutualistik akar non mikoriza serta respon tanaman inang dan proses kolonisasi yang terjadi belum banyak diketahui (Varma et al. 1999).
Imaningsih (2010) berhasil mengisolasi beberapa isolat Aspergillus dan
Penicillium dari serasah hutan dipterocarp asal Kalimantan Tengah dan
3 1.2 Tujuan Penelitian
Penelitian ini bertujuan untuk menganalisis potensi Aspergillus dan
Penicillium asal serasah dipterocarp sebagai pelarut fosfat simbiotik akar Z. mays
bebas endosimbion dan S. selanica terkolonisasi DSE.
1.3 Hipotesis Penelitian
Hipotesis dalam penelitian ini adalah:
1. Aspergillus dan Penicillium asal serasah dipterocarp adalah cendawan
pelarut fosfat
2. Aspergillus dan Penicillium asal serasah dipterocarp dapat hidup sebagai
cendawan endosimbion akar Z. mays bebas endosimbion dan akar S.
5 2 TINJAUAN PUSTAKA
2.1 Peranan dan Ketersediaan Fosfor
Fosfor merupakan unsur hara makro esensial bagi tanaman dan menempati kurang lebih 0.2% bobot kering tanaman. Fosfor adalah komponen penyusun molekul asam nukleat, fosfolipid dan ATP yang penting. Oleh karena itu tanpa adanya asupan P yang cukup, tanaman tidak dapat tumbuh dengan baik. Fosfor organik (Pi) juga terlibat dalam pengontrolan serangkaian reaksi enzimatik dan regulasi lintasan metabolik (Theodorou & Plaxton 1993).
Meskipun jumlah total P dalam tanah tinggi, namun P tersedia dalam bentuk terikat atau jauh dari daerah rizosfer sehingga tidak dapat dijangkau oleh sistem perakaran tanaman. P dalam tanah ditemukan dalam beberapa bentuk berbeda, seperti asam organik dan mineral (Schachtman et al. 1998). Pada banyak sistem produksi pertanian, fosfor merupakan unsur hara esensial yang paling sering dijumpai dalam keadaan kahat setelah N (Mosali et al. 2005).
Sebagian mikroorganisme tanah memiliki kemampuan untuk melarutkan fosfat. Mekanisme pelarutan fosfat dapat dilakukan dengan beberapa cara yaitu asidifikasi, pengkelatan logam dan reaksi pertukaran ion (Pradhan & Sukla 2005). Umumnya proses solubilisasi fosfat oleh mikroorganisme terjadi melalui mekanisme pengkelatan logam (Whitelaw 2000). Mikroorganisme menghasilkan berbagai asam organik dengan bobot molekul rendah seperti asam glukonat, asam
sitrat dan asam laktat untuk mengkelat kation dari senyawa fosfat (Chen et al. 2006). Gugus hidroksil dan karboksil dari asam yang dihasilkan akan mengkelat kation (Al, Fe, Ca) dari senyawa fosfat sehingga Pi dapat dilepaskan. Biasanya, reaksi ini diikuti oleh penurunan pH tanah (Sagoe et al. 1998). Solubilisasi kalsium fosfat umumnya terjadi melalui proses asidifikasi. Proses asidifikasi lingkungan sekitar sel mikroba dilakukan dengan substitusi proton/ekskresi H+
(Ca
diikuti absorbsi kation dalam jumlah yang besar. Goldstein (1986) secara umum menggambarkan proses asidifikasi kalsium fosfat dalam persamaan berikut:
2+
)m(PO43-)n + (HA) = (H+) (PO43-) + Ca2+ (A
-Solubilisasi fosfat organik (Po) di dalam tanah terjadi melalui proses mineralisasi melalui aksi fosfatase. Fosfatase (alkaline fosfatase dan acid fosfatase)
6
menggunakan fosfat organik sebagai substrat dan mengubahnya menjadi bentuk fosfat inorganik (Nenwani et al. 2010).
Fosfor diserap akar tanaman dalam dua bentuk anion, masing-masing dihidrogen fosfat (H2PO4-) dan monohidrogen fosfat (HPO42-) (Jones 1998). Bentuk Pi dalam larutan tergantung pada pH larutan. Nilai pK disosiasi H3PO4 menjadi H2PO4- dan HPO42- secara berurutan adalah 2.1 dan 7.2. Oleh karena itu, di bawah pH 6 Pi umumnya ada dalam bentuk monovalen H2PO4- . Sedangkan HPO42- ada dalam proporsi minor (Schachtman et al. 1998).
2.2 Cendawan Mutualistik Akar
Cendawan mutualistik akar dapat dikategorikan dalam kelompok cendawan mikoriza dan non mikoriza. Cendawan mikoriza merupakan cendawan non patogen paling umum yang bersimbiosis dengan sekitar 80% akar tanaman berpembuluh (Smith & Read 1997). Cendawan mikoriza memberi kemudahan bagi tanaman untuk mendapatkan nutrisi. Kemampuan hifa eksternal cendawan mikoriza mengeksploitasi P yang berlokasi di sekitar daerah deplesi P dapat mengatasi keterbatasan difusi fosfat anorganik yang lambat dalam tanah. Ukuran hifa yang jauh lebih kecil (1/10) dibandingkan dengan akar tanaman memberi kemudahan bagi hifa masuk jauh sampai ke pori-pori tanah untuk menjangkau P dan air (Smith et al. 2003).
Dalam habitat alami, asosiasi antara cendawan mikoriza dan tanaman inang
saling menguntungkan. Namun dalam sistem yang melibatkan campur tangan manusia, asosiasi tersebut bisa berkembang ke arah parasitisme. Hal ini dapat terjadi pada media tanam dengan konsentrasi P terlarut tinggi (Bucher 2007).
Berbagai jenis cendawan lain termasuk cendawan endofit dapat berasosiasi dengan sistem perakaran tanaman membentuk simbiosis mutualisme. Cendawan endofit dapat membantu tanaman inang dalam berbagai hal, diantaranya yaitu adaptasi di habitat yang kurang menguntungkan, perlindungan terhadap stress lingkungan baik biotik maupun abiotik, peningkatan pertumbuhan dan penyerapan nutrisi (Maciá-Vicente et al. 2009).
7 panen tanaman pertanian, mengontrol penyakit dan hama tanaman, adaptasi tanaman terhadap lingkungan yang kurang menguntungkan serta untuk kegiatan penghutanan lahan kembali. Ekploitasi cendawan endofit untuk tujuan-tujuan di atas dapat dilakukan dengan dua cara yaitu menggunakan cendawan endofit yang secara alami sudah terbukti memberikan keuntungan bagi tanaman inang dan melalui kolonisasi jaringan tanaman oleh cendawan yang diduga memiliki ciri-ciri yang diinginkan (Maciá-Vicente et al. 2009; Bhagobaty et al. 2010).
2.3 Kolonisasi Cendawan pada Akar
Kolonisasi cendawan dapat didefinisikan sebagai suatu keadaan dimana cendawan berada dalam tubuh inang untuk jangka waktu tertentu (Casadeval & Pirovski 2000). Barker et al. (1998) menyatakan bahwa inisiasi awal simbiosis cendawan mikoriza dengan inangnya ditandai dengan pelekatan cendawan pada jaringan inang. Umumnya cendawan mikoriza melekat pada jaringan inang melalui apresoria (cendawan vesikula arbuskula/VAM) atau dengan hifa (ektomikoriza). Tahapan ini dilanjutkan dengan kolonisasi bagian internal sel dan perkembangan hifa intraselular (hanya cendawan VAM).
Prekolonisasi cendawan VAM dimulai dengan perkecambahan spora dalam air menghasilkan hifa aseptat. Pertumbuhan hifa tidak berlanjut tanpa keberadaan akar atau eksudat akar. Perkecambahan spora dipengaruhi oleh senyawa kimia yang dihasilkan akar. Senyawa kimia yang bertindak sebagai sinyal pemicu
respon hifa atau spora cendawan adalah berbagai senyawa fenolik dan iso/flavonoid yang umum terbentuk akibat interaksi tanaman-mikroba (Barker et al. 1998).
Struktur khas cendawan VAM yang dibentuk secara interselular adalah arbuskula dan vesikula. Arbuskula adalah percabangan dikotomus yang intensif dari hifa intraselular dan berperan dalam transfer nutrisi antara cendawan dan tumbuhan. Vesikula dibentuk secara intra dan interseluler. Struktur ini berfungsi sebagai cadangan makanan bagi cendawan. Varma et al. (1999) menyatakan bahwa cendawan mutualistik akar non mikoriza Pirimospora indica
8
dalam korteks melalui pembentukan struktur percabangan dan koil atau struktur menyerupai klamidospora. Namun pada cendawan mutualistik akar non mikoriza tidak terjadi pembentukan arbuskula.
2.4 Analisis Proses Kolonisasi Cendawan pada Akar Tanaman
Proses kolonisasi cendawan pada akar tanaman dapat dipelajari dengan menggunakan beberapa metode. Beberapa metode yang sering digunakan diantaranya adalah metode pewarnaan akar, metode biokimiawi dan metode molekular. Dua metode terakhir membutuhkan biaya yang cukup besar dan waktu preparasi yang relatif lama sehingga tidak digunakan secara rutin. Mewarnai akar dan pengamatan mikroskopis merupakan metode standar yang banyak digunakan sampai sekarang (Vierheilig et al. 2005).
Brundrett (2004) menyatakan bahwa teknik pewarnaan akar dan pengamatan mikroskop tidak saja dapat memberikan data tingkat kolonisasi cendawan pada akar secara akurat tetapi juga mampu memperlihatkan keberadaan struktur kunci seperti arbuskula yang penting dalam penentuan ciri morfologi cendawan yang berasosiasi dengan tanaman inang. Untuk tujuan pengamatan ciri morfologi, material akar perlu diproses sedemikian rupa agar ciri morfologi yang ingin diamati dapat dengan mudah terdeteksi.
Pengamatan simbiosis cendawan dengan akar tanaman dapat dilakukan menggunakan metode non destruktif dan destruktif. Metode non destruktif
mungkin dilakukan dalam keadaan tertentu. Metode ini dapat dilakukan dengan beberapa cara seperti pengamatan langsung dibawah mikroskop cahaya, mikroskop epiflouresens dan mikroskop pemindai laser konfokal (confocal
laser-scanning microscopy) (Vierheilig et al. 2005; Maciá-Vicente et al. 2009).
Pengamatan menggunakan mikroskop pemindai elektron (SEM) dilakukan untuk tujuan mendapatkan hasil observasi dengan detail yang lebih baik daripada mikroskop cahaya.
9 Teknik pewarnaan non vital merupakan teknik yang paling sering digunakan untuk visualisasi cendawan pada akar tanaman. Struktur cendawan biasanya tidak nampak karena terhalang oleh pigmen alami tanaman dan isi sel. Oleh karena itu, diperlukan prosedur untuk menghilangkan pigmen yang ada pada dinding sel tanaman dan mengeluarkan isi sel (Gardner 1975). Struktur cendawan yang terdapat dalam akar setelah proses penjernihan kemudian diikat oleh zat warna. Zat warna ini biasanya diaplikasikan dalam larutan asam yang mengandung asam laktat, gliserin dan air (Brundrett et al. 1984).
2.5 Dark Septate Endophyte (DSE)
Jumpponen & Trappe (1998) mendefinisikan DSE sebagai cendawan Askomiset dengan ciri hifa bersekat, memiliki konidia atau steril, membentuk struktur termelanisasi (hifa interseluler, hifa intraseluler dan mikroskleria) di dalam akar tanaman. Kolonisasi DSE dilaporkan terjadi pada sekitar 600 spesies tanaman meliputi 320 genus dan 114 famili dan tersebar luas mulai dari daerah tropis sampai kutub dan pegunungan. DSE merupakan kelompok cendawan heterogen yang secara ekologi dan fungsinya tumpang tindih dengan cendawan tanah, cendawan saprob akar, cendawan patogen dan mikoriza.
Keragaman DSE secara taksonomi tidak begitu banyak diketahui walaupun keberadaaannya melimpah. Umumnya DSE tidak bersporulasi, atau bila bersporulasi, konidia yang dihasilkan sangat sedikit (Jumpponen & Trappe 1998).
Beberapa strain hanya dapat bersporulasi bila diberi stimulus temperatur rendah (Fernando & Currah 1995).
11 3 BAHAN DAN METODE
3.1 Waktu dan Tempat Penelitian
Penelitian dilaksanakan pada bulan September 2010 sampai Maret 2011 di Laboratorium Mikologi, Departemen Biologi FMIPA IPB dan Laboratorium Zoologi LIPI, Cibinong.
3.2 Bahan
Aspergillus sp. IPBCC.09.619, Aspergillus sp. IPBCC.07.503, Aspergillus
sp. IPBCC.10.643, Penicillium sp. IPBCC. 09.620 dan Penicillium sp.
IPBCC.09.621 merupakan kapang koleksi IPBCC yang berasal dari serasah hutan dipterocarp Kalimantan Tengah dan Kalimantan Timur. Biji Z. mays varietas Bismo diperoleh dari Balai Penelitian Bioteknologi dan Sumberdaya Genetik Pertanian Cimanggu, Bogor. Biji S. selanica diperoleh dari Balai Penelitian dan Pengembangan Kehutanan, Bogor. Media agar Pikovskaya digunakan untuk uji aktivitas pelarut fosfat. Larutan Hoagland dengan TCP sebagai sumber P digunakan untuk perlakuan berbagai taraf % P. Pewarna biru tripan 0.05% digunakan untuk analisis tahapan kolonisasi kapang terhadap tanaman uji. Komposisi media dan larutan hara yang digunakan dapat dilihat pada Lampiran 1 dan 2.
3.3 Metode Penelitian
Secara garis besar penelitian dibagi menjadi 3 tahapan (Gambar 1), yaitu 1. Uji aktivitas pelarut fosfat Aspergillus dan Penicillium asal serasah
dipterocarp dan produksi inokulan isolat
2. Perkecambahan tanaman inang, inokulasi isolat dan pemeliharaan
3. Analisis simbiosis Aspergillus dan Penicillium dengan akar Z. mays dan
S. selanica meliputi, analisis kolonisasi, respon tumbuh dan serapan hara
12
jika ada
Gambar 1 Bagan alur penelitian Potensi Aspergillus dan Penicillium
Asal Serasah Dipterocarp sebagai EndosimbionAkar Pelarut Fosfat
Z. mays dan
S. selanica
Uji aktivitas pelarut fosfat
Pemilihan biji
Aspergillus dan
Penicillium
Produksi inokulan
Inokulasi Aspergillus Penicillium dan pada akar
Pemberian larutan hara dan pemeliharaan tanaman
Aspergillus dan Penicillium
pelarut fosfat potensial Perkecambahan biji
Kecambah
Inokulan Pengamatan status endosimbion akar
Reisolasi endosimbion
13 3.3.1 Uji Aktivitas Pelarut Fosfat Aspergillus dan Penicillium Asal Serasah
Dipterocarp
Uji aktivitas pelarut fosfat Aspergillus dan Penicillium dilakukan terhadap kapang koleksi IPBCC yang berasal dari serasah dipterocarp. Aspergillus dan
Penicillium diremajakan pada media agar kentang dekstrosa (PDA) selama 7 hari.
Bagian tepi koloni yang tumbuh aktif diambil dengan menggunakan cork borer
steril (diameter 5 mm) dan kemudian diinokulasikan ke media Pikovskaya dengan TCP sebagai sumber P, lalu diinkubasi pada suhu ruang. Adanya zona bening di sekitar koloni mengindikasikan bahwa kapang memiliki kemampuan untuk melarutkan TCP menjadi fosfat anorganik (Pi). Indeks kelarutan fosfat dihitung berdasarkan rumus berikut:
Dua isolat Aspergillus dan Penicillium yang memiliki aktivitas cukup tinggi digunakan untuk analisis potensi dan proses kolonisasi terhadap tanaman uji.
3.3.2 Produksi Inokulan Aspergillus dan Penicillium
Produksi inokulan dua isolat terpilih dilakukan dengan menumbuhkan masing-masing tiga potong koloni kapang (diameter 5 mm) dalam 100 ml media dekstrosa kentang cair. Inkubasi dilakukan pada suhu ruang selama 1 minggu dengan agitasi 100 rpm. Miselium dipanen dan kemudian disaring menggunakan kertas saring Whatman steril. Sebanyak kurang lebih 3 gram miselium diinokulasikan pada perakaran tanaman uji.
3.3.3 Perkecambahan Z. mays dan S.selanica
Biji yang digunakan memiliki ukuran yang relatif seragam, tingkat
kematangan yang baik dan tidak cacat. Perkecambahan dilakukan pada media zeolit steril yang telah dicuci dalam wadah plastik. Sebelum ditanam sayap biji S.
selanica dibuang. Sterilisasi permukaan biji dilakukan untuk menghilangkan
14
dan dicuci kembali dengan air steril sebanyak 3-5 kali. Biji dikeringkan dengan tissue steril dan ditanam dalam zeolit. Untuk merangsang perkecambahan biji, permukaan wadah ditutup dengan aluminium foil selama 1 malam.
3.3.4 Inokulasi Aspergillus dan Penicillium pada Akar Tanaman Uji, Pemberian Hara dan Pemeliharaan
Sebelum kecambah diinokulasi, kecambah diamati status endosimbionnya. Bila ditemukan, maka endosimbion diisolasi dengan media yang sesuai dan dimurnikan pada akhir perlakuan. Kecambah Z. mays yang telah berumur 1 minggu dan 3 minggu untuk S. selanica diinokulasi dengan kurang lebih 3 gram inokulan kapang. Zeolit steril lembab (200g) dimasukkan ke dalam gelas plastik sebanyak kurang lebih sepertiga wadah. Ke dalam wadah kemudian diletakkan inokulan dan diaduk supaya inokulan tidak menumpuk pada satu daerah saja. Kecambah dimasukkan ke dalam wadah, lalu ditutup dengan sisa media. Larutan Hoagland digunakan sebagai sumber hara tanaman dengan TCP sebagai sumber P. Tanaman dipelihara selama delapan minggu dan disiram dengan air steril sekali dua hari atau sesuai kebutuhan.
3.3.5 Analisis Simbiosis Aspergillus dan Penicilliumdengan Akar Z. mays dan
S. selanica
Bentuk simbiosis Aspergillus dan Penicillium pelarut fosfat dengan tanaman uji dianalisis melalui beberapa tahap, yaitu analisis kolonisasi Aspergillus dan
Penicillium terpilih pada akar tanaman uji, respon tumbuh tanaman uji yang
15 A Analisis kolonisasi dengan biru tripan 0.05% dan mikroskop pemindai
elektron
Analisis kolonisasi dilakukan dengan cara mewarnai akar tanaman yang telah diinokulasi dengan Aspergillus dan Penicillium menggunakan biru tripan 0.05%. Analisis dilakukan terhadap lima contoh dari masing-masing tanaman setiap minggu dengan pemberian dosis P 100% (setara 60 mg P2O5/tanaman). Metode pewarnaan akar mengikuti prosedur Kormanick & McGraw (1982) dan diperkuat dengan pengamatan mikroskop pemindai elektron. Akar tanaman dipisahkan dari media dengan hati-hati agar sistem perakaran tidak rusak. Akar dicuci dengan air mengalir selama 10 menit untuk menghilangkan sisa media tanam dan setelah itu dipotong dengan ukuran 1 cm. Akar direndam dalam larutan KOH 10% pada suhu 90o
Pengamatan kolonisasi Aspergillus dan Penicillium pada akar tanaman menggunakan mikroskop pemindai elektron sesuai dengan metode Lewinson
(Lewinson et al. 1994). Preparasi diawali dengan merendam sampel akar di dalam
caccodylate buffer selama 2 jam, kemudian diagitasi dalam ultrasonic cleaner
selama 5 menit. Akar direndam dalam larutan glutaraldehyde 2.3% selama 2 hari dan difiksasi dengan tannic acid 2% selama 6 jam. Akar dicuci 4 kali dengan
caccodylate buffer selama 5 menit, selanjutnya didehidrasi secara bertahap dalam
alkohol 50% (5 menit), alkohol 70% (20 menit), alkohol 85% (20 menit), alkohol 95% (20 menit) dan 2 kali dalam alkohol absolut (10 menit). Akar selanjutnya direndam dua kali dalam tert butanol selama 10 menit, dibekukan dalam freezer
dan terakhir dimasukan ke dalam freezed drier sampai kering. Akar yang sudah kering diletakkan di atas batang besi dan dilapisi dengan 30 nm emas menggunakan mesin ion coater IB2. Struktur cendawan yang terdapat pada
16
permukaan akar diamati menggunakan mikroskop pemindai elektron model JSM 5000 LV yang dioperasikan pada tegangan 20 kv. Pengamatan proses kolonisasi menggunakan mikroskop pemindai elektron dilakukan di Laboratorium Zoologi, LIPI Cibinong.
Akar dinyatakan terkolonisasi bila minimal setengah dari panjang potongan akar membawa Aspergillus dan Penicillium. Kolonisasi dinyatakan dalam persen jumlah akar terkolonisasi dari seluruh akar yang diamati tiap sampel. Struktur-struktur khusus cendawan yang terbentuk semuanya diamati dan didokumentasikan menggunakan kamera digital.
B Analisis respon tumbuh dan serapan hara tanaman inang yang diinokulasi kapang pelarut fosfat
Respon tumbuh dan serapan hara tanaman terhadap inokulasi Aspergillus
dan Penicillium pelarut fosfat diuji dengan memberi perlakuan berbagai dosis %
TCP (dinyatakan sebagai mg P2O5) dalam larutan Hoagland pada tanaman Z.
mays dan S. selanica. Dosis TCP yang digunakan yaitu, 0% (0 mg P2O5/tanaman), 50% (setara 30 mg P2O5/tanaman) dan 100% (setara 60 mg P2O5/tanaman). Tanaman kontrol diberi perlakuan 60 mg P2O5
Setelah pengukuran parameter respon tumbuh tanaman selesai, maka dilakukan analisis serapan hara (NPK) terhadap tanaman tersebut. Analisis NPK dilakukan di Balai Penelitian Tanah, Bogor secara komposit. Serapan NPK (mg hara/tanaman) dihitung dengan mengalikan persentase kadar hara (%) dan bobot kering masing-masing tanaman.
/tanaman tetapi tidak diinokulasi dengan Aspergillus dan Penicillium pelarut fosfat. Parameter respon tumbuh yang diamati yaitu pertambahan tinggi tajuk tanaman, bobot basah dan kering tanaman, panjang akar serta bobot basah akar.
17 Analisis kadar N dilakukan menggunakan metode Kjehdahl. Sebanyak 0.25 g contoh tanaman dimasukkan ke dalam tabung digestion dan ditambah dengan 1 g campuran selen (siap pakai) dan 2.5 ml H2SO4. Campuran diratakan dan dibiarkan selama satu malam. Blanko dibuat dengan cara yang sama tetapi tidak mengandung sampel. Sampel selanjutnya dipanaskan hingga suhu 350oC dan dihentikan setelah terlihat ada uap putih yang keluar sehingga didapat ekstrak jernih. Tabung diangkat dan didinginkan, kemudian ekstrak diencerkan dengan air bebas ion hingga tepat 50 ml. Ekstrak dikocok sampai homogen dan dibiarkan semalam agar partikel mengendap. Satu ml ekstrak contoh dipipet ke dalam tabung reaksi, ditambah dengan 9 ml air bebas ion dan divorteks. Dua ml ekstrak encer dimasukkan dalam tabung reaksi baru dan ditambah larutan sangga Tartrat dan Na-fenat masing-masing sebanyak 4 ml, lalu divorteks dan dibiarkan 10 menit. Masing-masing tabung ditambah 4 ml NaOCl 5%, divorteks dan diukur dengan spektrofotometer pada panjang gelombang 636 nm. Larutan (NH4)2SO4
Penetapan unsur P dan K dilakukan dengan metode pengabuan basah menggunakan campuran asam nitrat (HNO
dengan konsentrasi 0-20 ppm digunakan sebagai standar.
3) dan asam perklorat (HClO4). Sebanyak 0.5 g sampel tanaman dimasukkan ke dalam tabung digestion, lalu ditambah dengan 5 ml HNO3 dan HClO4 dan dibiarkan selama satu malam. Sampel dipanaskan dalam digestion blok pada suhu 100oC selama satu jam, kemudian suhu ditingkatkan menjadi 150oC. Setelah uap kuning habis, suhu
digestion blok ditingkatkan menjadi 200o
Pengukuran kadar P dilakukan dengan mengencerkan masing-masing ekstrak contoh sampai 10x. Sebanyak 2 ml ekstrak encer contoh ditambah dengan 10 ml pereaksi pewarna P dan divorteks sampai homogen lalu dibiarkan selama 30 menit. Kadar P diukur pada panjang gelombang 693 nm dengan spektrofotometer UV-VIS. Sebagai standar digunakan larutan PO
C. Proses destruksi dihentikan setelah terlihat ada asap putih yang keluar dan sisa ekstrak kurang lebih 0,5 ml. Tabung diangkat dan dibiarkan dingin. Ekstrak diencerkan dengan air bebas ion hingga tepat 50 ml dan dikocok sampai homogen dan siap digunakan untuk penentuan kadar P dan K.
18
Pengukuran kadar K dilakukan dengan cara memipet masing-masing 1 ml ekstrak ke dalam tabung reaksi dan ditambah dengan 9 ml larutan La 0,25 %. Larutan divorteks sampai homogen dan kadar K diukur menggunakan
flamefotometer. Larutan standar K yang digunakan adalah K2SO4
Percobaan dilakukan menggunakan rancangan acak lengkap (RAL) dengan 5 ulangan. Data selanjutnya diolah menggunakan program SPSS 16.0 dan diuji lanjut menggunakan uji Jarak Berganda Duncan.
dengan konsentrasi 0-250 ppm. Larutan pereaksi yang digunakan dalam penentuan kadar NPK dan rumus yang dipakai dapat dilihat pada Lampiran 3.
3.3.6 Uji Viabilitas Endosimbion
Untuk mengetahui viabilitas endosimbion, dilakukan reisolasi kapang dari akar tanaman 8 minggu setelah inokulasi menggunakan media PDA. Akar tanaman yang telah diinokulasi dengan kapang pelarut fosfat dipisahkan dari media secara hati-hati, dicuci dengan air mengalir selama kurang lebih 10 menit lalu dipotong dengan ukuran kurang lebih 1 cm. Akar direndam dalam alkohol 70% selama 30 detik lalu NaOCl 0.05% selama 5 menit. Setiap selesai satu tahap, akar dibilas dengan air steril sebanyak 3-5 kali. Akar dikeringkan dengan tissue steril dan ditanam pada media PDA lalu diinkubasi pada suhu ruang. Viabilitas endosimbion dapat diketahui apabila reisolasi berhasil dilakukan. Selanjutnya endosimbion yang berhasil direisolasi dimurnikan dan diremajakan menggunakan
media yang sama.
3.3.7 Isolasi DSE
Dark Septate Endophyte yang ditemukan pada akar S. selanica diisolasi
21 4 HASIL DAN PEMBAHASAN
4.1 Hasil Pengamatan
4.1.1 Potensi Aspergillus dan Penicillium sebagai Cendawan Pelarut Fosfat
Aspergillus dan Penicillium koleksi IPBCC yang berasal dari serasah
dipterocarp dapat membentuk zona bening pada media Pikovskaya dengan indeks kelarutan bervariasi (Tabel 1). Dua isolat, yaitu Aspergillus sp. IPBCC.09.619 dan
Penicillium sp. IPBCC.09.620 memiliki indeks kelarutan fosfat yang cukup tinggi dan
selanjutnya potensi pembentukan endosimbion dengan akar tanaman Z. mays dan
S. selanica dianalisis.
Tabel 1 Indeks kelarutan fosfat Aspergillus dan Penicillium asal serasah dipterocarp
Nama Isolat Indeks kelarutan fosfat pada hari ke-
1 2 3 4 5
Aspergillus sp. IPBCC.09.619 0.35 0.19 0.13 0.14
-Aspergillus sp. IPBCC.07.503 0.16 0.04 0.03 0.26 0.01
Aspergillus sp. IPBCC.10.643 0.11 0.04 0.03 0.25 0.01
Penicillium sp. IPBCC.09.620 0.33 0.27 0.31 0.69 0.21
Penicillium sp. IPBCC.09.620 0.2 0.12 0.41 0.38 0.09
Sebelum potensi endosimbiosis Aspergillus dan Penicillium diuji pada Z.
mays dan S. selanica, akar-akar dari kedua tanaman tersebut diperiksa status
endofitnya. Akar Z. mays ditemukan bebas endofit sedangkan akar S. selanica
membawa DSE. Jumlah sel akar S. selanica yang mengandung DSE dalam setiap potongnya paling banyak 15 sel dan penyebarannya tidak merata. Persentase DSE
dalam kecambah adalah sebesar 85% (35 dari 40 kecambah membawa DSE). Kolonisasi DSE pada setiap kecambah bervariasi mulai dari 7.5-83.3% dengan
rata-rata 28.2% (data tidak disajikan).
22
juga ditemui hifa dengan ukuran besar yang tumbuh menjalar di permukaan akar dengan percabangan masuk ke dalam sel akar (Gambar 2e). Pengamatan akar menggunakan mikroskop pemindai elektron (Scanning Electron Microscope/
SEM) memperlihatkan miselium DSE di permukaan akar dengan klamidospora terminal (Gambar 2f).
Gambar 2 Struktur DSE pada akar S. selanica, a, b) mikrosklerosia memenuhi sel akar, c) hifa berbentuk rantai interseluler, d) hifa berbentuk rantai tumbuh di permukaan akar, e) hifa berukuran besar, bercabang dan menjalar di permukaan akar, f) penampakan DSE pada permukaan akar.
Dua jenis koloni DSE (putih dan hitam) berhasil diisolasi dari akar S.
selanica (Gambar 3a). Pengamatan mikroskopik terhadap koloni DSE berwarna
hitam memperlihatkan DSE memiliki hifa bersekat pendek dan bercabang.
20 µm a
f e
d
c b 20 µm
20 µm
20 µm
20 µm 1500x
mikroskerosia mikroskerosia
hifa
interseluler
hifa
interseluler hifa
interseluler
23 Beberapa klamidospora interkalar ditemukan pada koloni tersebut (Gambar 3b, c). Koloni berwarna putih memperlihatkan bahwa DSE tersebut memiliki hifa halus bercabang banyak. Klamidospora terminal dan interkalar dijumpai pada koloni yang sudah tua (Gambar 3d, e).
Gambar 3 Hasil isolasi DSE, a) Koloni DSE berwarna putih dan hitam umur 7 hari hasil isolasi dari akar S. selanica pada media Malt Extract Agar
(MEA), b) hifa bersekat pendek dan bercabang, c) klamidospora interkalar, d, e) hifa bercabang dengan klamidospora interkalar dan terminal, k = klamidospora.
4.1.2 Aspergillus sp. pada Akar Z. mays dan S. selanica
Tiga hari setelah inokulasi Aspergillus sp., daun jagung menjadi layu dan
pada hari ketujuh terlihat bahwa akar membusuk serta mudah putus. Hasil pengamatan mikroskop stereo memperlihatkan akar yang terinfeksi Aspergillus
sp. menjadi berair dan transparan (Gambar 4a). Inokulasi Aspergillus sp. pada perakaran S. selanica menyebabkan akar menghitam dan mati setelah dua minggu. Daun S. selanica bebercak kuning dan agak keriput (Gambar 4b, c).
24
Gambar 4 Pengaruh inokulasi Aspergillus sp. terhadap Z. mays dan S. selanica,
a) akar Z. mays berair dan lapuk, b) akar S. selanica menghitam dan mati, c) bercak kuning pada daun S. selanica, d) daun sehat. Tanda panah menunjukkan bagian yang terkena pengaruh Aspergillus sp.
4.1.3 Penicillium sp. pada Akar Z. mays
Penicillium sp. dapat mengkolonisasi akar Z. mays dan tidak menyebabkan
gejala sakit pada tanaman. Satu minggu setelah inokulasi, miselium cendawan pada umumnya terlihat menempel pada permukaan akar termasuk akar rambut (Gambar 5a, b dan g). Penetrasi hifa pada sebagian besar akar belum terjadi (Gambar 5c). Walaupun demikian, pada sebagian kecil potongan akar teramati adanya penetrasi hifa pada rongga intersel epidermis akar dan tidak terlihat adanya struktur khusus untuk penetrasi (Gambar 5d). Pertumbuhan hifa di dalam akar pada awal kolonisasi terjadi secara interseluler. Satu minggu setelah inokulasi Penicillium sp., tidak terlihat adanya perubahan morfologi pada akar Z.
mays (Gambar 5e). Akar-akar dari tanaman kontrol tidak memperlihatkan adanya
kolonisasi endofit (Gambar 5f). Namun analisis dengan SEM mendeteksi adanya struktur menyerupai apresorium berbentuk noktah (Gambar 5h). Struktur menyerupai apresorium ini menempel pada dinding sel akar dan menyebabkan bagian akar berlubang sehingga hifa dapat masuk ke dalam sel akar.
a b c d
busuk
mati
bercak kuning
25 inokulasi, a) miselium menempel pada akar, b) akar tanaman kontrol, c) hifa menempel pada rambut akar, d) hifa pada permukaan akar, e) hifa masuk ke rongga intersel, f) akar setelah 1 minggu inokulasi, g) gumpalan miselium menyelubungi rambut-rambut akar, h) struktur apresorium.
Dua minggu setelah inokulasi terlihat miselium Penicillium sp. mulai menyebar di permukaan akar (Gambar 6a, b). Hifa yang masuk ke dalam celah intersel menjalar di sepanjang ruang antar sel epidermis dan terbentuk kompartementasi hifa (Gambar 6c). Hifa membentuk semacam tonjolan sampai
26
akhirnya berbentuk bulat. Struktur ini semakin lama semakin membesar dan bertambah banyak (Gambar 6d-g). Pada saat ini tidak terdeteksi adanya perubahan morfologi akar. Perubahan hanya terjadi di tempat lebih banyak miselium menempel. Akar-akar ini berwarna kecoklatan (Gambar 6h).
Gambar 6 Kolonisasi Penicillium sp. pada akar Z. mays 2 minggu setelah inokulasi, a, b) miselium mulai tumbuh menjalar di permukaan akar, c-g) proses pembentukan struktur seperti noktah, h) bentuk akar 2 minggu setelah inokulasi.
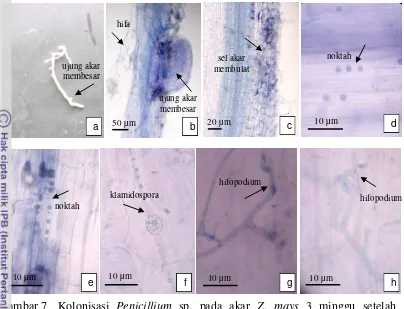
Sebagian ujung akar terlihat mulai membesar pada minggu ketiga setelah inokulasi (Gambar 7a, b). Miselium semakin menyebar di permukaan akar dan terjadi perubahan morfologi sel akar pada bagian yang memiliki tingkat kolonisasi lebih tinggi. Sel-sel ini membulat dan ukurannya lebih besar dari sel-sel normal
(Gambar 7c). Struktur seperti noktah terlihat semakin banyak memenuhi sel akar pada bagian akar yang memiliki tingkat kolonisasi tinggi sehingga sel menjadi
berwarna gelap (Gambar 7d, e). Pada saat yang sama juga ditemukan struktur seperti klamidospora dan sel-sel seperti hifopodium (Gambar 7f-h).
27
Gambar 7 Kolonisasi Penicillium sp. pada akar Z. mays 3 minggu setelah inokulasi, a) bentuk akar setelah 3 minggu inokulasi, b) miselium mulai memenuhi permukaan akar dan ujung akar membesar, c) sel-sel dengan tingkat kolonisasi tinggi membesar dan membulat, d-e) struktur bulat seperti noktah semakin banyak, f) struktur seperti klamidospora, g-h) struktur seperti hifopodium pada permukaan akar.
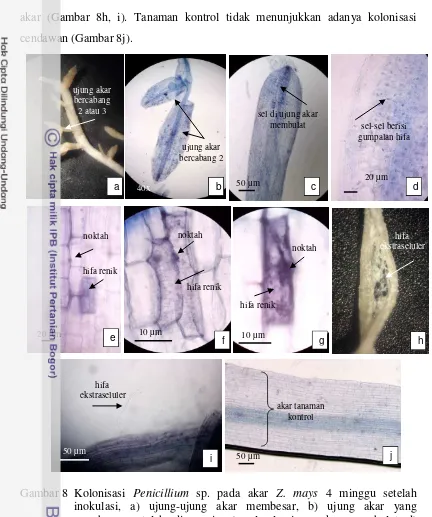
Kolonisasi Penicillium sp. pada empat minggu setelah inokulasi menyebabkan sebagian besar ujung-ujung akar Z. mays bertambah besar, bercabang 2 atau 3 dan tidak memanjang lagi (Gambar 8a, b). Pengamatan mikroskopik terhadap ujung akar yang membesar menunjukkan bahwa morfologi
sel-sel akar yang semula panjang dan ramping berubah bentuk menjadi membulat dan bertambah besar. Bagian ujung akar yang membesar terlihat berisi gumpalan
hifa (Gambar 8c-d). Hifa tumbuh menjalar di ruang antar sel dan membentuk noktah pada beberapa bagian. Selain itu juga terlihat hifa masuk ke dalam sel membentuk jalinan hifa renik yang memenuhi sel akar (Gambar 8e). Struktur seperti noktah di dalam sel akar mulai berkecambah membentuk jalinan hifa renik yang memenuhi sel-sel akar, menyebabkan sel terlihat berwarna ungu kebiruan (Gambar 8f-g). Pada minggu ini juga terlihat miselium tumbuh di luar permukaan
28
akar (Gambar 8h, i). Tanaman kontrol tidak menunjukkan adanya kolonisasi cendawan (Gambar 8j).
Gambar 8 Kolonisasi Penicillium sp. pada akar Z. mays 4 minggu setelah inokulasi, a) ujung-ujung akar membesar, b) ujung akar yang membesar setelah diwarnai, c) sel-sel ujung akar membulat d) gumpalan hifa mengisi sel-sel akar yang membesar, e-g) sel-sel akar berisi noktah dan hifa renik, h, i) hifa ektraselular mulai muncul ke permukaan akar, j) akar tanaman kontrol.
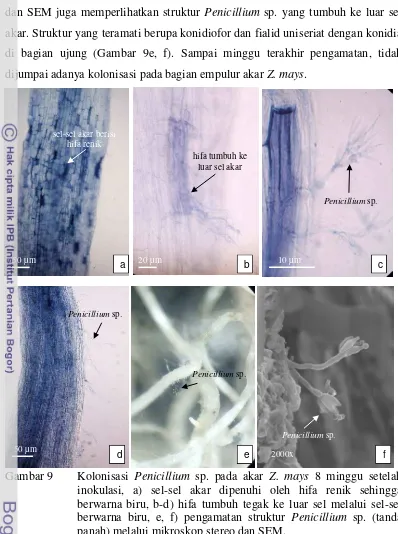
Delapan minggu setelah inokulasi, hifa Penicillium sp. telah memenuhi sebagian besar sel-sel epidermis dan korteks akar. Sel-sel akar terlihat berwarna biru muda sampai biru gelap (Gambar 9a). Hifa tumbuh ke luar sel melalui sel-sel berwarna biru membentuk hifa bercabang, konidiofor dan fialid, tetapi belum menghasilkan konidia (Gambar 9b-d). Hasil pengamatan dengan mikroskop stereo
a b c d
29 dan SEM juga memperlihatkan struktur Penicillium sp. yang tumbuh ke luar sel akar. Struktur yang teramati berupa konidiofor dan fialid uniseriat dengan konidia di bagian ujung (Gambar 9e, f). Sampai minggu terakhir pengamatan, tidak dijumpai adanya kolonisasi pada bagian empulur akar Z. mays.
Gambar 9 Kolonisasi Penicillium sp. pada akar Z. mays 8 minggu setelah inokulasi, a) sel-sel akar dipenuhi oleh hifa renik sehingga berwarna biru, b-d) hifa tumbuh tegak ke luar sel melalui sel-sel berwarna biru, e, f) pengamatan struktur Penicillium sp. (tanda panah) melalui mikroskop stereo dan SEM.
4.1.4 Penicillium sp. pada Akar S. selanica
Penicillium sp. juga dapat mengkolonisasi akar S. selanica tanpa
menimbulkan gejala sakit (Gambar 10a). Minggu pertama setelah inokulasi
Penicillium sp., terlihat bahwa miselium umumnya menempel pada permukaan
akar dan mulai tumbuh menjalar disepanjang permukaan akar (Gambar 10b, c). Pada bagian tertentu, hifa membentuk tonjolan untuk menempel dan penetrasi
a
d e f
b c
50 µm
50 µm 20 µm 10 µm
2000x sel-sel akar berisi
hifa renik
hifa tumbuh ke luar sel akar
Penicillium sp.
Penicillium sp. Penicillium sp.
30
pada permukaan sel kemudian membentuk struktur seperti apresorium (Gambar 10d-f).
Gambar 10 Kolonisasi Penicillium sp. pada akar S. selanica 1 minggu setelah inokulasi, a) penampakan akar, b) miselium menempel pada permukaan akar, c) hifa menjalar di permukaan akar, d) tonjolan hifa untuk menempel dan penetrasi, e, f) struktur seperti apresorium.
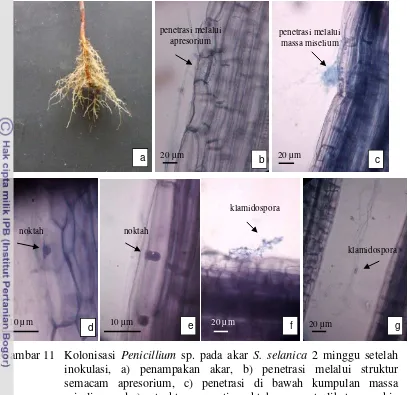
Dua minggu setelah inokulasi terlihat adanya penetrasi pada sel epidermis. Penetrasi terjadi langsung pada permukaan sel melalui struktur mirip apresorium. Di dalam sel, hifa membentuk semacam tabung yang kemudian menembus sel dibawahnya membentuk struktur seperti noktah (Gambar 11b). Penetrasi juga terlihat di bawah kumpulan massa miselium yang menempel pada permukaan akar dan menembus dinding sel lalu membentuk struktur seperti noktah di bagian dalam sel (Gambar 11c). Noktah membesar dan hifa eksternal sebagian besar sudah tidak terlihat lagi menempel pada permukaan akar (Gambar 11d, e).
Penicillium sp. yang berada di luar sel akar terlihat juga membentuk klamidospora
bersel dua (Gambar 11f, g).
31
Gambar 11 Kolonisasi Penicillium sp. pada akar S. selanica 2 minggu setelah inokulasi, a) penampakan akar, b) penetrasi melalui struktur semacam apresorium, c) penetrasi di bawah kumpulan massa miselium, d-e) struktur seperti noktah yang terlihat semakin membesar, f-g) struktur seperti klamidospora di luar sel akar
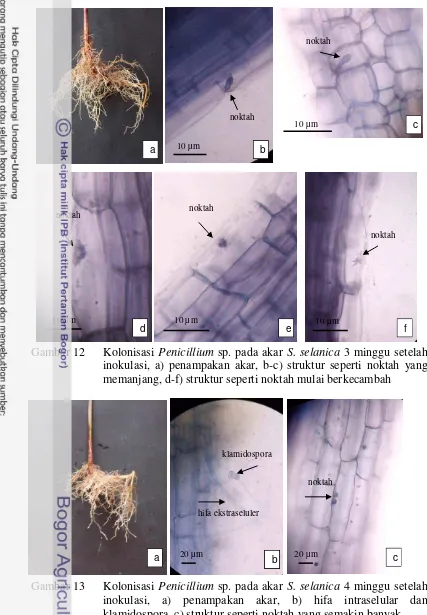
Kolonisasi Penicillium sp. setelah tiga minggu tidak menunjukkan perkembangan yang signifikan. Hal ini terlihat dengan tidak ditemukannya penetrasi hifa yang lebih jauh ke dalam sel. Umumnya struktur seperti noktah hanya ditemukan pada sel epidermis. Bedanya, pada minggu ini terlihat struktur seperti noktah mulai tumbuh memanjang dan berkecambah (Gambar 12b-f).
Minggu keempat setelah inokulasi terlihat bahwa sebagian hifa tumbuh di permukaan akar dan bagian lain berada di ruang interseluler dan intraseluler. Di luar akar, hifa membentuk klamidospora (Gambar 13b). Pada minggu ini terlihat
bahwa miselium sudah mulai menjalar di dalam sel akar membentuk struktur seperti noktah yang lebih banyak (Gambar 13c). Penetrasi hifa ke dalam sel tidak meluas. Hifa umumnya hanya berada pada bagian epidermis akar.
g
apresorium penetrasi melalui massa miselium
noktah
klamidospora noktah
32
Gambar 12 Kolonisasi Penicillium sp. pada akar S. selanica 3 minggu setelah inokulasi, a) penampakan akar, b-c) struktur seperti noktah yang memanjang, d-f) struktur seperti noktah mulai berkecambah
Gambar 13 Kolonisasi Penicillium sp. pada akar S. selanica 4 minggu setelah inokulasi, a) penampakan akar, b) hifa intraselular dan klamidospora, c) struktur seperti noktah yang semakin banyak.
f e
d
c
b a
c b
a
noktah
hifa ekstraseluler klamidospora 10 µm
10 µm
10 µm 10 µm 10 µm
20 µm 20 µm
noktah
noktah noktah
noktah
33
Gambar 14 Kolonisasi Penicillium sp. pada akar S. selanica 8 minggu setelah inokulasi, a) miselium pada permukaan akar, b, c) struktur seperti noktah pada epidermis sel, d-f) hifa ekstraseluler yang tumbuh ke luar akar setelah 8 minggu inokulasi.
Delapan minggu setelah inokulasi Penicillium sp., miselium sudah tumbuh menyebar memenuhi beberapa bagian sel akar (Gambar 14a). Pengamatan
mikroskopik menunjukkan bahwa hifa yang ada dalam akar kebanyakan hanya berupa noktah yang semakin banyak. Noktah tidak menunjukkan perkembangan lebih lanjut seperti halnya noktah pada tanaman Z. mays (Gambar 14b, c). Hifa yang terlihat pada permukaan akar merupakan hifa ekstraseluler, tetapi belum terlihat adanya struktur khas Penicillium (Gambar 14d-f).
Perlakuan berbagai taraf % TCP tidak mempengaruhi tingkat kolonisasi
Penicillium sp. pada tanaman Z. mays. Sementara itu, kolonisasi cendawan pada
perakaran S. selanica dipengaruhi oleh dosis % TCP yang diberikan. Persentase kolonisasi tertinggi diperoleh dengan pemberian TCP sebesar 60 mg
34
P2O5/tanaman. Pada perlakuan kontrol tidak dijumpai kolonisasi cendawan baik pada Z. mays maupun S. selanica (Tabel 2).
Tabel 2 Persentase kolonisasi Penicillium sp. 8 minggu setelah inokulasi dengan perlakuan P berbeda
Jenis Tanaman Perlakuan % Kolonisasi % Kolonisasi (mg P2O5/tanaman) Penicillium sp. DSE
Ket: Angka dalam kolom yang sama pada masing-masing kelompok dan diikuti huruf yang sama tidak berbeda nyata berdasarkan uji jarak berganda Duncan, P<0.05
4.1.5 Respon Tumbuh Tanaman Uji yang Diinokulasi Penicillium sp.
Tabel 3 Pengaruh perlakuan P terhadap pertumbuhan tanaman uji yang diinokulasi Penicillium sp.
Jenis Ket: Angka dalam kolom yang sama pada masing-masing kelompok dan diikuti
35 Gambar 15 Kurva respon tumbuh tanaman uji terhadap dosis P. ( = Z.
36
Inokulasi Penicillium sp. terhadap sistem perakaran tanaman uji dapat meningkatkan pertumbuhan kedua tanaman uji secara signifikan dengan
pemberian P dosis normal (60 mg P2O5/tanaman) kecuali untuk pertambahan tinggi tanaman S. selanica bila dibandingkan dengan perlakuan tanpa inokulasi
(Tabel 3). Respon tumbuh tanaman uji dinyatakan dengan pertambahan tinggi tanaman, bobot basah dan bobot kering tanaman, panjang akar serta bobot basah akar. Pemberian P sebesar 30 mg P2O5/tanaman juga dapat meningkatkan tiga parameter respon tumbuh Z. mays (bobot basah dan bobot kering tanaman serta panjang akar) walaupun peningkatannya tidak sebesar dosis 60 mg P2O5/tanaman. Sementara itu, kolonisasi Penicillium sp. tidak memberikan pengaruh yang nyata terhadap pertumbuhan S. selanica pada pemberian P 30 mg P2O5/tanaman. Perlakuan P sebesar 0 mg P2O5
Hasil analisis kurva respon tumbuh tanaman uji terhadap pemberian berbagai taraf P (0 mg, 30 mg dan 60 mg P
/tanaman memberikan respon tumbuh paling rendah bila dibandingkan dengan perlakuan lainnya.
2O5/tanaman) menunjukkan adanya hubungan antara dosis P dengan respon pertumbuhan tanaman. Semua parameter respon tumbuh yang diamati membentuk kurva linear kecuali parameter bobot basah tanaman Z. mays. Bobot basah tanaman ini membentuk kurva kuadratik (Gambar 15).
4.1.6 Serapan Hara Tanaman Uji yang Diinokulasi Penicillium sp.
Kolonisasi Penicillium sp. mempengaruhi serapan hara kedua tanaman uji
37 Tabel 4 Pengaruh berbagai perlakuan P terhadap serapan hara tanaman uji
Tanaman Perlakuan Ket: Angka dalam kolom yang sama pada masing-masing kelompok dan diikuti
huruf yang sama tidak berbeda nyata berdasarkan uji jarak berganda Duncan, P<0.05. BK= bobot kering; Jar = jaringan; tan = tanaman.
4.1.7 Viabilitas Endosimbion
Delapan minggu setelah inokulasi Penicillium sp., endosimbion berhasil diisolasi kembali dari akar tanaman Z. mays dan S. selanica dan dapat tumbuh dengan baik pada media PDA. Penicillium sp. hasil reisolasi dari akar S. selanica
bercampur dengan DSE dan sulit untuk dipisahkan. Pemisahan koloni dilakukan dengan cara pengenceran bertingkat. Hasil reisolasi dapat dilihat pada Gambar 16.
Gambar 16 Reisolasi Penicillium sp. dari akar tanaman uji 8 minggu setelah inokulasi, (a) dari akar Z. mays dan (b) S. selanica.
a a
Penicillium sp.
DSE
38
4.2 Pembahasan
Aspergillus dan Penicillium koleksi IPBCC yang berasal dari serasah hutan
dipterocarp dapat membentuk zona bening di sekitar koloni pada media agar Pikovskaya. Hal ini mengindikasikan bahwa isolat-isolat tersebut memiliki
kemampuan untuk melarutkan TCP secara in vitro. Aspergillus sp. IPBCC.09.619
dan Penicillium sp. IPBCC.09.620 merupakan dua isolat yang memiliki indeks
kelarutan fosfat tertinggi, yaitu berturut-turut sebesar 0.35 (1 hsi) dan 0.69 (4 hsi) sehingga berpotensi untuk digunakan sebagai cendawan pelarut fosfat. Kedua isolat ini diinokulasikan pada akar kecambah Z. mays dan S. selanica untuk dianalisis kemampuan pembentukan endosimbionnya. Aspergillus dan Penicillium
telah dilaporkan mampu melarutkan fosfat dan berpengaruh terhadap pertumbuhan tanaman (Pradan & Sukla 2005; Richa et al. 2007; Yadaf et al. 2011), namun kemampuan simbiosisnya belum pernah dilaporkan.
Dark Septate Endophyte ditemukan pada sebagian besar kecambah S.
selanica yang digunakan sebagai tanaman uji (85% dari total kecambah membawa
DSE), namun hanya sedikit sel akar yang terkolonisasi DSE pada setiap potongan akar yang diamati. Keberadaan DSE tersebut diasumsikan tidak akan mengganggu kolonisasi cendawan pelarut fosfat dan pertumbuhan tanaman uji. Selain itu, adanya DSE dalam akar tanaman tidak menyebabkan gejala sakit pada tanaman uji. Jumpponen et al. (1998) menyatakan bahwa DSE diduga berfungsi sebagai cendawan mutualistik yang berperan dalam pengambilan nutrisi dan air terutama dalam kondisi lingkungan tidak menguntungkan. Kolonisasi DSE bersamaan
dengan cendawan mikoriza pada suatu tanaman inang diduga membuat DSE bertindak sebagai suatu sistem back up pada saat pertumbuhan cendawan
mikoriza terhambat oleh kondisi lingkungan yang tidak menguntungkan. Kemungkinan besar DSE yang ditemukan berasal dari cendawan tular benih (seed
borne fungi) atau cendawan kontaminan yang masuk ke dalam biji pada saat biji
jatuh ke tanah.
39 percabangan dan panjang hifa cendawan mikoriza Gigaspora rosea. Jumpponen
et al. (1998) menumbuhkan benih Pinus contorta di tanah glasier dengan kondisi
kandungan nitrogen (N) rendah. Mereka menyatakan bahwa inokulasi DSE
Phialocephala fortinii dapat meningkatkan konsentrasi P pada daun dan apabila
inokulasi DSE dikombinasikan dengan penambahan N, maka biomassa P.
contorta meningkat sebesar 50%. Haselwandter & Read (1982) juga menyatakan
bahwa inokulasi dua jenis DSE dapat meningkatkan kandungan P pada daun benih Carex firma dan C. sempervirens. Inokulasi kedua jenis DSE tersebut hanya dapat merangsang pertumbuhan C. firma. Usuki & Narisawa (2007) menyatakan bahwa DSE dapat bertindak sebagai cendawan mikoriza melalui transfer nutrisi dua arah secara in vitro.
Akar tanaman merupakan salah satu habitat alami berbagai mikroorganisme termasuk cendawan. Hubungan yang terbentuk antara tanaman dan cendawan dapat berupa interaksi positif, negatif atau netral (Atlas & Barta 1998). Bentuk interaksi cendawan dengan tanaman inang dapat diketahui melalui pengamatan respon tanaman uji terhadap inokulasi cendawan. Inokulasi Aspergillus sp. pada akar kedua tanaman uji menyebabkan kerusakan akar dan daun tanaman. Gejala yang terlihat akibat inokulasi Aspergillus sp. pada Z. mays adalah layu daun dan busuk akar sedangkan daun S. selanica bebercak kuning dan agak keriput, akar menjadi hitam dan mati.
Adanya kerusakan jaringan tanaman inang yang disebabkan oleh inokulasi
Aspergillus sp. mengindikasikan bahwa cendawan ini bukan endosimbion akar
kedua tanaman uji sehingga dapat dikatakan bahwa Aspergillus sp. adalah cendawan patogen Z. mays dan S. selanica. Hal ini sesuai dengan definisi patogen
yang dinyatakan oleh Cassadeval & Pirofski (2000). Menurut definisi tersebut, patogen adalah mikroba yang mampu menyebabkan kerusakan tubuh inang.
Penicillium sp. dapat mengkolonisasi epidermis dan korteks akar Z. mays
dan S. selanica tanpa menyebabkan gejala sakit pada tanaman. Kolonisasi pada
40
sesuai dengan pendapat Lagopodi et al. (2002); Olivain & Alabouvette (1999) bahwa Fusarium oxysporum pertama kontak dengan akar tomat juga pada bagian
akar rambut.
Kolonisasi Penicillium sp. pada akar Z. mays dimulai dengan masuknya hifa
ke dalam akar melalui rongga intersel epidermis. Struktur khusus penetrasi tidak terlihat melalui teknik pewarnaan biru tripan. Namun melalui pengamatan SEM terlihat bahwa hifa menembus epidermis akar melalui struktur mirip apresorium yang menyebabkan sel akar berlubang sehingga hifa dapat melakukan penetrasi. Kolonisasi pada akar S. selanica dimulai dengan terbentuknya tonjolan hifa pada permukaan akar dan kemudian berkembang menjadi struktur seperti apresorium. Massa miselium yang menempel pada permukaan akar juga dapat mempenetrasi dinding sel. Cara penetrasi Penicillium sp. pada akar Z. mays dan S. selanica
berbeda, namun dapat dikategorikan ke dalam mekanisme penetrasi secara aktif, artinya cendawan dapat secara langsung menembus sel akar tanpa pelukaan terlebih dahulu (Lagopodi et al. 2002).
Setelah berada di dalam akar, hifa Penicillium sp. tumbuh secara interseluler dan mengalami kompartementasi. Hifa menonjol dan diikuti dengan terbentuknya struktur berbentuk bulat yang semakin lama semakin membesar dan bertambah banyak. Proses ini mirip dengan kolonisasi cendawan endofit Heteroconium
chaetospira pada akar tanaman kubis Cina. Tiga minggu setelah inokulasi, hifa H.
chaetospira membentuk cuping bulat tidak teratur di dalam sel epidermis tanaman
sehat dan selanjutnya terbentuk mikrosklerosia (Ohki et al. 2002).
Struktur bulat seperti noktah pada akar Z. mays selanjutnya membentuk jalinan hifa renik yang memenuhi sel akar sehingga sel terlihat berwarna biru