ANALISIS JARAK GENETIK BERDASARKAN MARKA SSRs
DAN MORFOLOGI SERTA ANALISIS DAYA GABUNG
UNTUK PEMBENTUKAN HIBRIDA JAGUNG MANIS
(Zea mays L. var. saccharata)
R. NENI IRIANY M
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI DISERTASI DAN
SUMBER INFORMASI
Dengan ini saya menyatakan bahwa disertasi “Analisis Jarak Genetik Berdasarkan Marka SSRs dan Morfologi serta Analisis Daya Gabung untuk Pembentukan Hibrida Jagung Manis (Zea mays L. var. saccharata) ” adalah karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun yang tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan tercantum dalam Daftar Pustaka di bagian akhir disertasi ini.
Bogor, September 2011
ABSTRACT
R.NENI IRIANY M. Genetic Distance Analysis based on SSRs and Morphological
Marker and Combining Ability Analysis for sweet corn development (Zea mays L. var. saccharata) hybrid. Under supervised SRIANI SUJIPRIHATI (in
memoriam), MUHAMAD SYUKUR, JAJAH KOSWARA (in memoriam), and MUHAMAD YUNUS.
The objective of the research were to grouping sweet corn lines resistant to downy mildew, high specific and general combining ability and high heterosis to select parental line to breed single cross hybrid of sweet corn.
The first research was done to study genetic variability of 39 sweet corn lines based on SSR and morphological marker to select good parental lines of hybrid sweet corn. There were high genetic variability among genetic material, showed by genetic distance value about 0.18 to 0.78, indicates wide genetic variability of sweet corn. The results shows that among 20 lines having genetic similarity between 0.25 – 0.82 or genetic distance from 0.18 to 0.75; based on morphological trait 0.27 – 1.00. Correlation value between similarity matrix based on SSR and morphology was high on 0.88 of goodness of fit criteria.
The aim of second research was to study general combining ability of sweet corn lines for yield and resistant to downy mildew for hybrid varieties improvement. Six of 45 lines having high fresh ear yield per plot and best general combining ability, such as Mr4/SC/BC4-2-1B, Mr4/SC/BC4-6-1B, Mr11/SC/BC4-2-1B, Mr11/SC/BC4-4-1B, Mr12/SC/BC4-6-1B, and Mr12/SC/BC3-3-1B.
All of testing lines have resistance to downy mildew with severity about 0.0 to 7.1%. The lines having high GCA value would be good combiner to other lines to develop high yielding hybrid sweet corn and resistant to downy mildew.
The third research was done to study general and specific combining ability of sweet corn lines and higher heterosis of downy mildew resistance supporting the hybrid corn development. The combination of A x D lines (Mr12/SC/BC4-6-1B x Mr11/SC/BC4-2-1B) were medium genetic distance and having highest GCA, heterosis and heterobeltiosis for fresh ear yield per plot character. The D x C combination (Mr11/SC/BC4-2-1B x Mr4/SC/BC4-2-1B) have wide genetic distance and high fresh ear yield per plot, heterosis and heterobeltiosis, but have negative SCA value. The combination of E x C (Mr12/SC/BC3-3-1B x Mr4/SC/BC4-2-1B) have a wide genetic distance but low productivity.
RINGKASAN
R.NENI IRIANY M. Analisis Jarak Genetik Berdasarkan Marka SSRs dan Morfologi serta Analisis Daya Gabung untuk Pembentukan Hibrida Jagung Manis (Zea mays L. var. saccharata) Dibimbing oleh SRIANI SUJIPRIHATI (alm), MUHAMAD SYUKUR, JAJAH KOSWARA (alm)dan MUHAMAD YUNUS.
Penelitian ini bertujuan untuk memperoleh pengelompokan galur jagung manis yang tahan terhadap penyakit bulai, dengan nilai daya gabung umum, daya gabung khusus, serta nilai heterosis yang tinggi sehingga diperoleh jagung hibrida silang tunggal yang mempunyai potensi hasil tinggi.
Penelitian pertama untuk mengetahui keragaman genetik 39 galur jagung manis memanfaatkan marka SSRs dan karakter morfologi untuk seleksi tetua hibrida. Hasil percobaan menunjukkan keragaman genetik cukup tinggi dari materi yang diuji. Hal ini ditunjukkan dengan nilai jarak genetik berkisar dari 0,18 sampai 0,78 yang mengindikasikan variabilitas genetik galur jagung manis yang dikarakterisasi cukup luas. Hasil penelitian 20 galur berdasarkan SSRs menunjukkan koefisien kemiripan genetik pada galur yang diuji berkisar antara 0,25 – 0,82 atau pada jarak genetik 0,18 - 0,75; berdasarkan karakter morfologi menunjukkan nilai rata-rata koefisien kemiripan genetik 0,27 – 1,00. Nilai korelasi antara matrik kemiripan berdasarkan SSRs dengan matrik kemiripan berdasarkan morfologi tergolong baik berdasarkan kriteria goodness of fit (0,88).
Kegiatan pada penelitian kedua dilakukan untuk untuk mengetahui nilai daya gabung umum galur-galur jagung manis (Zea mays L. var. saccharata) untuk karakter hasil dan ketahanan terhadap penyakit bulai (downy mildew) dalam rangka perakitan varietas hibrida. Dari 45 galur yang diuji terpilih enam galur yang mempunyai hasil tongkol segar per petak dan nilai DGU tertinggi yaitu Mr4/SC/BC4-2-1B, Mr4/SC/BC4-6-1B, Mr11/SC/BC4-2-1B, Mr11/SC/BC4-4-1B, Mr12/SC/BC4-6-1B, dan Mr12/SC/BC3-3-1B.
Semua genotipe jagung manis yang diuji memiliki ketahanan terhadap penyakit bulai dengan presentase serangan 0,0 – 7,1%. Galur-galur yang bernilai DGU tinggi diharapkan mempunyai kemampuan berdaya gabung umum baik untuk menghasilkan genotip yang mempunyai potensi hasil tinggi dan tahan penyakit bulai.
Kegiatan ketiga bertujuan mengetahui nilai daya gabung umum (DGU), daya gabung khusus (DGK) galur jagung manis (sweet corn), heterosis hibrida terbaik tahan penyakit bulai (downy mildew) mendukung perakitan varietas hibrida. Persilangan galur A x D (Mr12/SC/BC4-6-1B x Mr11/SC/BC4-2-1B) mempunyai nilai DGK, heterosis, heterobeltiosis tertinggi untuk karakter hasil tongkol segar per petak, jarak genetik yang sedang dan produktivitas tinggi. Persilangan galur D x C (Mr11/SC/BC4-2-1B x Mr4/SC/BC4-2-1B) mempunyai hasil tongkol segar per petak tinggi, nilai heterosis dan heterobeltiosis tinggi serta memiliki jarak genetik yang cukup jauh, tetapi nilai DGK negatif. Persilangan galur E x C (Mr12/SC/BC3-3-1B x Mr4/SC/BC4-2-1B) mempunyai jarak genetik yang jauh, tetapi produktivitasnya rendah.
© Hak Cipta milik IPB, tahun 2011 Hak Cipta dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan yang wajar IPB
ANALISIS JARAK GENETIK BERDASARKAN MARKA SSRs
DAN MORFOLOGI SERTA ANALISIS DAYA GABUNG
UNTUK PEMBENTUKAN HIBRIDA JAGUNG MANIS
(Zea mays L. var. saccharata)
R. NENI IRIANY M
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Judul Disertasi : Analisis Jarak Genetik Berdasarkan Marka SSRs dan Morfologi serta Analisis Daya Gabung untuk Pembentukan Hibrida Jagung Manis (Zea mays L. var. saccharata)
Nama : R. Neni Iriany M
NIM : A263070011
Disetujui
Komisi Pembimbing
Prof. Dr. Ir. Sriani Sujiprihati, MSc.(alm) Dr. Muhamad Syukur, SP.,MSi
Ketua Anggota
.
Prof. Dr. Ir. Jajah Koswara (alm)
Anggota Anggota
Dr. Ir. Muhamad Yunus, MSi.
Mengetahui
Ketua Program Studi Dekan Sekolah Pascasarjana Pemuliaan dan Bioteknologi Tanaman
PRAKATA
Segala puji dan syukur bagi Allah S.W.T., atas segala limpahan berkah, rahmat, hidayah dan inayah-Nya sehingga penulis dapat menyelesaikan disertasi berjudul “Analisis Jarak Genetik Berdasarkan Marka SSRs dan Morfologi serta Analisis Daya Gabung untuk Pembentukan Hibrida Jagung Manis (Zea mays L. var. saccharata)” .
Penulis mengucapkan terima kasih dan penghargaan yang setinggi-tingginya kepada Prof. Dr. Ir. Sriani Sujiprihati MS. (alm), Dr. Muhamad Syukur,Sp., MSi., Prof. Dr. Ir. Jajah Koswara (alm), dan Dr. Ir. Muhamad Yunus, MSi, selaku pembimbing yang telah banyak memberi arahan, sumbangan pemikiran dan motivasi yang diberikan sejak penulis mengikuti pendidikan, penelitian dan penulisan hingga selesainya disertasi ini. Kepada Dr. Ir. Memen Surahman, MSc., Agr., Dr. Ir. Yudiwanti, MS., sebagai penguji luar komisi pembimbing pada ujian tertutup, serta Dr. Ir. Hajrial Aswidinnoor, MS., Dr. Ir. Firdaus Kasim MSc., sebagai penguji pada ujian terbuka. Di samping itu, penghargaan penulis sampaikan kepada Kepala Badan Litbang Pertanian dan Ketua Komisi Pembinaan Tenaga Badan Litbang Pertanian Departemen Pertanian beserta staf, Kepala Balai Penelitian Tanaman Serealia Maros beserta staf, Kepala KP. Cikeumeuh Balai Besar Penelitian Pengembangan Bioteknologi dan Sumber Daya Genetik (BB Biogen) Bogor beserta staf, KP Maros, dan Koordinator Laboratorium Molekuler Balai Penelitian Tanaman Serealia beserta staf, yang telah membantu selama penelitian dan pengumpulan data. Penulis menyadari bahwa keberhasilan penyelesaian disertasi ini tidak terlepas dari bantuan dan dukungan dari rekan-rekan teknisi dan peneliti. Untuk itu penulis mengucapkan terima kasih kepada Sampara, Sunarto, Julaiha, Haryati SP., Hasnah SP., Rismayanti SP., Hariadi, Sirajudin, Usman, Hamzah, Abd. Haris, Stepanus Misi, Muzdalifah Isnaini SP.MP., Indria Wahyu Mulsanti SP., Dr. Indrastuti A. Rumanti., Dr. Rahmi Yunianti., Dr. Muhammad Azrai, Dr. Marcia Bunga Pabendon, Ir. Sumarni Singgih MS., dan Ir. Muh. Yasin HG. MS.,
Kepada Pimpinan dan Dosen SPs IPB, penulis menyampaikan penghargaan dan ucapan terima kasih atas segala bimbingan dan pembinaannya selama penulis menyelesaikan studi, terutama kepada Ketua Program Studi Pemuliaan dan Bioteknologi Tanaman beserta staf yang telah membantu penulis selama mengikuti studi di IPB. Ucapan terima kasih juga penulis sampaikan kepada rekan-rekan mahasiswa Program Studi Pemuliaan dan Bioteknologi Tanaman angkatan 2006 dan 2007 atas segala bantuan, dukungan dan kebersamaannya selama penulis mengikuti studi di SPs IPB.
dan dukungannya, penulis juga mengucapkan terima kasih yang sebesar-besarnya. Secara khusus ucapan terima kasih penulis sampaikan kepada keluarga Dr. Marsum Dahlan (alm) dan keluarga Prof. Dr. Ir. Djafar Baco, MS atas bimbingan dan arahannya selama ini.
Semoga karya ilmiah ini bermanfaat terhadap pengembangan ilmu pengetahuan.
Bogor, September 2011
DAFTAR RIWAYAT HIDUP
Penulis dilahirkan di Kaju Kabupaten Bone Sulawesi Selatan pada tanggal 1 April 1971 sebagai anak bungsu pasangan Massi Patittingi dan Hj. Siti Aminah (Almarhum). Pendidikan sarjana di Program Studi Biologi Fakultas Matematika dan Ilmu Pengetahuan Alam Universitas Hasanuddin Ujung Pandang, lulus pada tahun 1996. Pada tahun 2000, penulis diterima di Program Studi Pemuliaan Tanaman pada Program Pascasarjana UNPAD dan menamatkan pada tahun 2002. Kesempatan untuk melanjutkan ke program doktor pada Program Studi Pemuliaan dan Bioteknologi Tanaman IPB diperoleh pada tahun 2007. Beasiswa pendidikan pascasarjana diperoleh dari Badan Litbang Pertanian Departemen Pertanian Republik Indonesia.
viii
BIODATA
N a m a : R. Neni Iriany M Tempat/Tgl.Lahir : Kaju/1 April 1971
Instansi : Balai Penelitian Tanaman Serealia Maros Jabatan : Pemulia Tanaman Jagung
Alamat Kantor : Jl. Dr. Ratulangi No.274
Po.Box 1173 Makassar, Telp. 0411 371016, 371961. Fax. 0411 371961 E-mail: balitsereal@plasa.com,
balitser@yahoo.com
Maros 90514 Sulawesi Selatan.
Alamat Rumah : Jl. Kacang Hijau No.72 Komplek Balitsereal Maros Telp. 0411 371 764/081342749687 Kel: Allepolea Kec: LAU, Kab: Maros, Sulawesi Selatan 90511
Alamat E-mail : rn_iriany@yahoo.com
Riwayat Pendidikan:
1983 SD Negeri 241 Tellu Boccoe, Kab.Bone 1986 SMP Negeri 516 Mara, Kab.Bone 1989 SMA Negeri 372 Mara, Kab. Bone 1996 SSi. Universitas Hasanuddin (MIPA)
2002 MP. Universitas Padjadjaran (Pemuliaan Tanaman)
Riwayat Kerja :
DAFTAR ISI
Halaman
DAFTAR TABEL ... xii
DAFTAR GAMBAR ... xiv
DAFTAR LAMPIRAN ... xv
BAB I. PENDAHULUAN ... 1
Latar Belakang ... 1
Tujuan Penelitian ... 6
Hipotesis ... 6
Diagram Alir Penelitian ... 7
BAB II. TINJAUAN PUSTAKA ... 10
Genetika jagung manis (sweet corn)... 10
Penyakit bulai (downy mildew)... 11
Pengendalian penyakit bulai(downy mildew)... 13
Pemuliaan untuk ketahanan terhadap penyakit ... 14
Marka genetik ... 16
Keragaman genetik pada jagung ... 18
Korelasi marka DNA dengan marka fenotipik... 18
Metode dialel... 19
Daya gabung (combining ability) ... 20
Heterosis ... 22
BAB III Analisis keragaman genetik galur-galur jagung manis (Zea mays L. var. saccharata) berdasarkan marka SSRs (Simple Sequence Repeats) dan morfologi... 23
Abstrak ... 23
Pendahuluan ... 25
Bahan dan Metode ... 26
Hasil dan Pembahasan ... 33
xv
BAB IV Evaluasi daya gabung galur-galur jagung manis (Zea mays
L. var. saccharata) menggunakan silang puncak ... 43
Abstrak ... 43
Pendahuluan ... 45
Bahan dan Metode ... 46
Hasil Pembahasan ... 50
Kesimpulan ... 62
BAB V Evaluasi daya gabung galur jagung manis (Zea mays L. var. saccharata) dan heterosis hibridanya menggunakan persilangan diallel ... 63
Abstrak ... 63
Pendahuluan ... 65
Bahan dan Metode ... 67
Hasil Pembahasan ... 73
Kesimpulan ... 95
BAB VI. PEMBAHASAN UMUM ... 96
BAB VII. KESIMPULAN DAN SARAN ... 100
Kesimpulan ... 100
Saran ... 101
DAFTAR PUSTAKA ... 102
xv
DAFTAR TABEL
Halaman 1. Kandungan nilai nutrisi dalam biji jagung manis ... 10 2. Perkembangan gejala penyakit bulai (downy mildew)………... 12 3. Materi tanaman yang digunakan dalam penelitian……… 26 4. Sekuen dari 20 marka mikrosatelit yang digunakan dalam
penelitian……… 28
5. Materi tanaman yang digunakan dalam analisis berdasarkan
marka SSRs dan morfologi……… 30
6. Data pengamatan morfologi galur jagung manis...……… 31 7. Kriteria goodness of fit berdasarkan nilai korelasi ……… 32 8. Profil data marka mikrosatelit hasil karakterisasi pada galur
jagung manis menggunakan 20 marka SSRs ……… 35
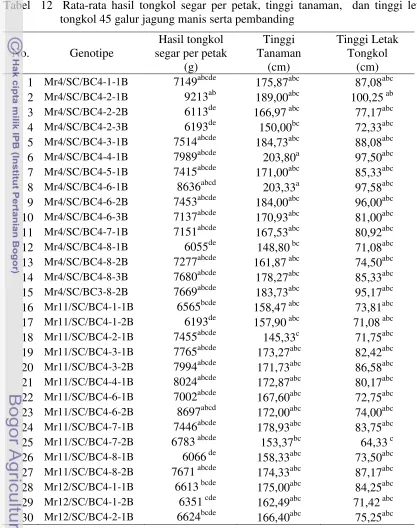
9. Deskripsi karakter morfologi 20 galur jagung manis …...……… 37 10. Materi genetik yang digunakan dalam persilangan top cross …... 47 11. Kriteria ketahanan tanaman jagung terhadap penyakit bulai... 49 12. Rata-rata hasil tongkol segar per petak, tinggi tanaman, dan
tinggi letak tongkol 45 galur jagung manis serta pembanding
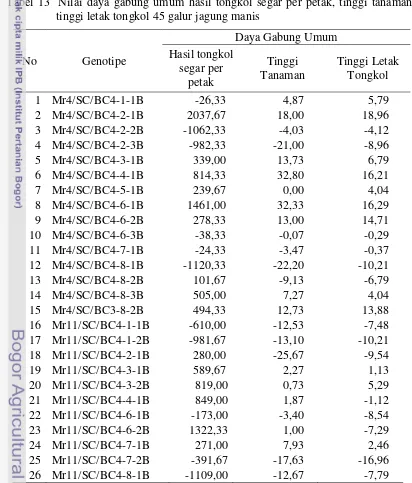
………... 51 13. Nilai daya gabung umum hasil tongkol segar per petak, tinggi
tanaman dan tinggi letak tongkol 45 galur jagung manis
……… 53
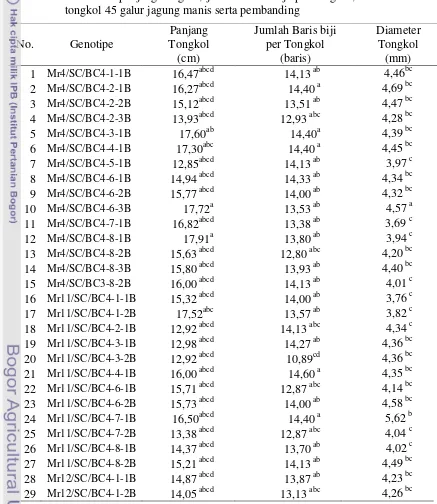
14. Rata-rata panjang tongkol, jumlah baris biji per tongkol, diameter tongkol, dan 45 galur jagung manis serta
pembanding……… 56
15. Nilai daya gabung umum panjang tongkol, jumlah baris biji per tongkol, diameter tongkol, 45 galur jagung manis
………..………. 58
16 Rata-rata presentase serangan penyakit bulai 45 galur jagung manis dan pembanding, serta nilai DGU penyakit bulai
………... 60
xv
19. Kuadrat tengah perbedaan antar genotipe karakter hasil tongkol segar per petak, panjang tongkol, diameter tongkol, dan jumlah baris biji per tongkol persilangan dialel (5 x 5) galur jagung
manis ………. 73
20. Kuadrat tengah perbedaan antar genotipe karakter tinggi tanaman, tinggi letak tongkol, persentase serangan terhadap penyakit bulai dan kandungan gula persilangan dialel (5 x 5)
galur jagung manis ……… 73
21. Kuadrat tengah genotipe, DGU, DGK dan resiprokal pada
persilangan dialel (5 x 5) galur jagung manis ………. 74
22. Kuadrat tengah genotipe, DGU, DGK dan resiprokal pada
persilangan dialel (5 x 5) genotipe jagung manis ………... 75
23. Daya gabung umum lima galur jagung manis... 76 24. Nilai Daya Gabung Khusus 20 kombinasi persilangan galur
jagung manis... 77
25. Nilai rata-rata hasil tongkol segar per petak, P1, P2, dan F1 serta
nilai heterosis dan heterobeltiosis... 84
26. Nilai rata-rata panjang tongkol P1, P2, dan F1 serta nilai
heterosis dan heterobeltiosis ... ... 85
27. Nilai rata-rata diameter tongkol P1, P2, dan F1 serta nilai
heterosis dan heterobeltiosis ... ... 86
28. Nilai rata-rata jumlah baris biji per tongkol P1, P2, dan F1 serta
nilai heterosis dan heterobeltiosis... 87
29. Nilai rata-rata tinggi tanaman P1, P2, dan F1 serta nilai
heterosis dan heterobeltiosis ... ... 88
30. Nilai rata-rata tinggi letak tongkol P1, P2, dan F1 serta nilai
heterosis dan heterobeltiosis ... ... 89
31. Nilai rata-rata kandungan gula P1, P2, dan F1 serta nilai
heterosis dan heterobeltiosis………... 90
32. Nilai rata-rata persentase serangan penyakit bulai, P1, P2, dan
xv
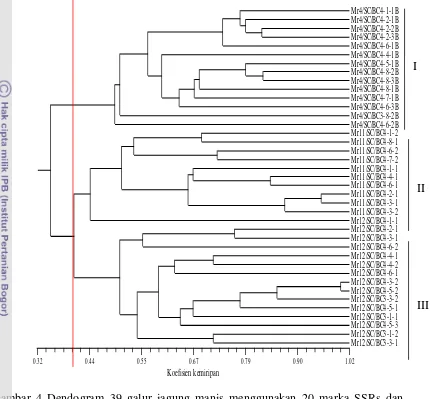
DAFTAR GAMBAR
Halaman 1. Bagan alir kegiatan penelitian ... 8
2. Daun tanaman jagung yang terserang bulai ... 13
3. Visualisasi pola pita DNA menggunakan marka SSRs phi109188 melalui elektroforesis vertikal 4,5% PAGE
(Polyacrylamide Gel Electrophoresis)……… 34
4. Dendogram 39 galur jagung manis menggunakan 20 marka SSRs dan dikonstruksi berdasarkan koefisien kemiripan
Jaccard ………. 36
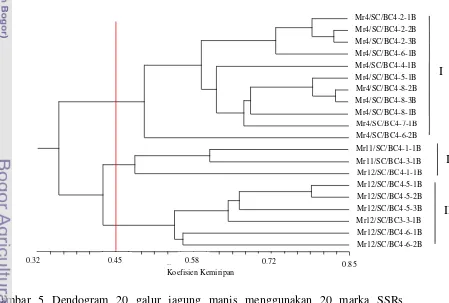
5. Dendogram 20 galur jagung manis menggunakan 20 marka
SSRs berdasarkan koefisien kemiripan Jaccard……… 38
6. Dendogram 20 galur jagung manis menggunakan data morfologi berdasarkan koefisien kemiripan Jaccard pada 11
karakter ………. 40
7. Dendogram 20 galur jagung manis menggunakan gabungan data genetik dan morfologi berdasarkan koefisien kemiripan
xv
DAFTAR LAMPIRAN
Halaman
1. Nilai matriks kemiripan 39 galur jagung manis………... 112
2. Matriks kemiripan 20 galur jagung manis berdasarkan marka
SSRs……….. 113
3. Matriks kemiripan 20 galur jagung manis berdasarkan data
morfologi……….. 114
4. Matriks kemiripan 20 galur jagung manis berdasarkan marka
SSRs dan data morfologi………. 115
BAB. I
PENDAHULUAN
Latar Belakang
Jagung (Zea mays L.) merupakan salah satu tanaman pangan terpenting yang memiliki peranan strategis dalam pembangunan pertanian dan perekonomian Indonesia, mengingat komoditas ini mempunyai fungsi multiguna, baik untuk pangan, pakan, dan bahan baku industri. Sebagai sumber karbohidrat yang menempati peringkat ketiga di Asia setelah padi dan gandum, serta menempati peringkat kedua di Indonesia setelah padi sangat berperan dalam menunjang ketahanan pangan nasional. Penduduk beberapa daerah di Indonesia, misalnya di
Madura dan Nusa Tenggara menggunakan jagung sebagai pangan pokok. Tongkol dan batangnya juga digunakan sebagai pakan ternak; biji jagung dapat diambil minyaknya dan dibuat tepung, yang dikenal dengan istilah tepung jagung atau maizena; dan bahan baku industri. Tongkol jagung kaya akan pentosa, yang dipakai sebagai bahan baku pembuatan furfural. Jagung yang telah direkayasa genetika juga sekarang ditanam sebagai penghasil bahan farmasi.
Kurun waktu lima tahun terakhir, kebutuhan jagung untuk bahan baku industri pakan, makanan, dan minuman terus meningkat berkisar antara 10% - 15% per tahun. Ditambah lagi akibat konsumsi jagung yang meningkat untuk pengembangan bioetanol terutama di Amerika Serikat (AS) yang juga produsen jagung dunia, harga jagung dunia melonjak mencapai 200 dolar AS per metrik ton atau naik sekitar 50%. Sumbangan jagung terhadap pendapatan domestikasi bruto (PDB) terus meningkat setiap tahun, sekalipun saat krisis ekonomi. Pada tahun 2000, kontribusi jagung sebesar 9,4 trilyun rupiah dan pada tahun 2003 meningkat secara tajam menjadi 18,2 trilyun rupiah. Kondisi demikian mengindikasikan besarnya peranan jagung dalam memacu pertumbuhan subsektor tanaman pangan dan pertanian serta perekonomian nasional secara umum (Departemen Pertanian, 2006).
2
jagung manis terutama untuk konsumsi di daerah perkotaan, super market, dan daerah pinggiran perkotaan yang mendukung pariwisata, sementara limbah jagung segar setelah panen sangat bermanfaat bagi petani sebagai tambahan hijauan pakan ternak. Di pasaran lokal permintaan terhadap jagung manis terus meningkat mencapai 1-1,5 ton/hari, sedangkan permintaan dari pasar di wilayah Jakarta dan Batam bahkan bisa lebih dari 1,5 ton/hari. Permintaan jagung manis terus meningkat baik permintaan dalam negeri juga permintaan untuk ekspor ke luar negeri. Peningkatan ini dikarenakan pemanfaatan jagung manis sebagai makanan selingan, jagung sayur maupun untuk bahan baku industri. Pengembangan bioetanol membawa dampak pada peningkatan kebutuhan jagung dunia. Bahan baku bioetanol
menggunakan amylase dan glukoamilase dari jagung manis. Peningkatan permintaan pasar jagung manis juga karena adanya perbaikan kadar gula dalam kultivar-kultivar jagung manis. Adanya peningkatan kadar gula karena hasil dari perbaikan gen-gen yang menyebabkan rasa manis pada jagung.
Jagung manis diperoleh dari jagung biasa yang mengalami mutasi resesif secara spontan, mutasi ini dapat mengendalikan konversi gula menjadi pati dalam endosperm. Karakter biji berkerut dan transparan, dengan kandungan gula yang tinggi dan kandungan pati yang rendah pada endosperm (Tracy, 1997). Jagung manis mengakumulasi gula sekitar dua kali (sekitar 12%) dan 8-10 kali lebih dapat larut dalam air dibanding jagung normal yang masih muda.
Jagung manis mengandung kadar gula yang relatif tinggi, rasa manis disebabkan oleh tiga gen utama yaitu gen sugary (su), sugary enhancer (se), dan shrunken (sh2). Kultivar jagung manis yang mengandung gen su, menghasilkan jumlah gula yang tinggi, tetapi terjadi perubahan gula menjadi pati secara cepat setelah panen jika tongkol tidak berada dalam temperatur yang dingin. Kultivar jagung manis yang mengandung gen se, menghasilkan jumlah gula lebih tinggi dibanding kultivar yang mengandung gen su. Kultivar yang mengandung gen se juga akan mengkonversi gula menjadi pati seperti jagung manis normal, tetapi prosesnya lebih lama setelah panen karena kandungan gulanya lebih tinggi. Kultivar jagung manis yang mengandung gen sh2, tidak langsung mengkonversi gula menjadi
pati dan oleh karena itu setelah panen rasa manisnya bertahan untuk suatu waktu
3
kombinasi-kombinasi gen mana yang memberikan kandungan gula yang lebih tinggi (Cobbledick – OMAF, 1997).
Jagung manis belum ditanam secara luas di Indonesia, padahal permintaan jagung manis semakin meningkat setiap tahun. Namun demikian, menurut Data Pusat Statistik (BPS) menunjukkan bahwa produksi jagung manis Indonesia periode 2001-2005 belum dapat mencukupi kebutuhan konsumsinya (BPS, 2006). Hal ini dikarenakan jumlah varietas jagung manis yang beredar di Indonesia masih terbatas sehingga harga benihnya mahal, dan umumnya tidak tahan penyakit bulai. Harga benih yang mahal tersebut disebabkan karena umumnya varietas yang beredar dirilis oleh perusahaan swasta, dimana materi genetiknya merupakan hasil introduksi, dan
belum ada pesaing dalam negeri. Varietas jagung manis yang beredar diantaranya adalah Manis Madu, Biji Mas, Chia Thai Seed (varietas bersari bebas), Sweet Boy, Bonansa, S & G, Thai Super Sweet (Hibrida).
Berdasarkan uraian di atas tampak bahwa permintaan jagung manis akan terus meningkat, sementara itu kemampuan produksi masih rendah dan mahalnya harga benih jagung manis. Oleh karena itu produksi jagung manis perlu ditingkatkan untuk memenuhi kebutuhan jagung manis tiap tahunnya. Namun upaya peningkatan produksi jagung selama ini menghadapi beberapa kendala, di antaranya adalah gangguan hama dan penyakit. Di negara-negara tropik dan sub tropik selain kekeringan, hama dan penyakit tanaman jagung merupakan salah satu penghambat utama yang mengganggu stabilitas produksi.
4
tahun-tahun berikutnya. Dari tahun 1973 sampai 1979 terjadi penurunan produksi berturut-turut sebesar 115, 92, 19, 44, 62, 55, dan 70 ribu ton (Sudjadi 1992).
Usaha-usaha pengendalian penyakit pada dasarnya adalah cara-cara mengendalikan perkembangan patogen, memanfaatkan inang dan lingkungan untuk memperkecil akibat yang ditimbulkan patogen sehingga mencapai suatu titik di bawah ambang ekonomi dengan kerugian yang dapat diabaikan (Sudjadi 1992). Beberapa cara pengendalian penyakit bulai adalah penanaman kultivar tahan, pengaturan waktu tanam, sanitasi dan perlakuan benih dengan metalaksil. Perakitan kultivar unggul yang tahan terhadap penyakit dapat dilakukan dengan beberapa cara, antara lain dengan melakukan hibridisasi atau persilangan. Persilangan merupakan
salah satu upaya untuk menambah variabilitas genetik dan memperoleh genotipe baru.
Genotipe baru yang telah diperoleh diseleksi lagi untuk memperoleh genotipe yang lebih baik dan lebih unggul. Sejauh ini, para pemulia tanaman menggunakan marka morfologi dalam seleksi maupun persilangan. Namun ekspresi marka morfologi dipengaruhi oleh kondisi lingkungan dan kadang-kadang juga dipengaruhi oleh adanya interaksi epistatik dan pleiotropik. Hal ini mengakibatkan sulitnya memperoleh data yang bisa dipercaya. Oleh karena itu data karakterisasi secara morfologi harus didukung oleh karakterisasi secara molekuler.
Kemajuan yang pesat di bidang biologi molekular tanaman akhir-akhir ini telah memecahkan permasalahan di atas. Marka molekuler yang terkait erat dengan sejumlah karakter yang memiliki nilai ekonomi penting telah berhasil dikembangkan. Keadaan ini memungkinkan adanya seleksi tidak langsung dari suatu sifat yang diinginkan yaitu dengan hanya menguji pada tingkat bibit. Hal ini memberikan efisiensi dari segi waktu, sumber daya maupun energi yang sebenarnya dibutuhkan untuk memelihara suatu populasi yang besar selama beberapa generasi dan juga untuk memperkirakan parameter-parameter yang digunakan dalam proses seleksi secara langsung.
Pemanfaatan marka DNA sebagai alat bantu seleksi Marker Assisted Selection (MAS) lebih menguntungkan dibandingkan dengan seleksi secara fenotipik. Seleksi dengan bantuan marka molekuler didasarkan pada sifat genetik tanaman saja, tidak
5
menjadi lebih tepat, cepat, dan relatif lebih hemat biaya dan waktu. Seleksi berdasarkan karakter fenotipik tanaman di lapang memiliki beberapa kelemahan seperti yang disarikan oleh Lamadji et al. (1999), di antaranya memerlukan waktu yang cukup lama, kesulitan memilih dengan tepat gen-gen yang menjadi target seleksi untuk diekspresikan pada sifat-sifat morfologi atau agronomi, rendahnya frekuensi individu yang diinginkan yang berada dalam populasi seleksi yang besar, dan fenomena pautan gen antara sifat yang diinginkan dengan sifat tidak diinginkan sulit dipisahkan saat melakukan persilangan.
Dari sejumlah marka molekuler, marka mikrosatelit atau SSRs (Simple Sequence Repeats) telah dikenal secara luas banyak memberikan harapan dalam
studi keragaman genetik dan penampilannya yang kodominan sehingga dapat mengidentifikasi genotipe homozigot dan heterozigot dalam populasi.
Selama ini Balai Penelitian Tanaman Serealia (Balitsereal) belum pernah merilis varietas jagung manis, oleh karena itu untuk memperoleh varitetas jagung manis hasil rakitan Balitsereal maka dilakukan persilangan antara jagung manis
varietas ‘Thai supersweet’ dengan galur-galur koleksi Balitseral yaitu Mr4, Mr11, Mr12 dan Mr14. Keempat galur diketahui tahan terhadap penyakit bulai, mempunyai daya gabung yang baik dan berasal dari populasi yang beda. Galur-galur tersebut disilangkan dengan jagung manis melalui silang balik (back cross) sebanyak empat generasi dan silang diri (selfing) selama empat generasi. Persilangan dilakukan untuk memasukkan gen kemanisan dan mengembalikan karakter-karakter unggul yang ada pada masing-masing galur.
Galur-galur jagung manis yang sudah diperoleh dilakukan analisis kekerabatan berdasarkan karakter genotipe memanfaatkan marka SSRs, berdasarkan karakter morfologi, dan korelasinya untuk mengetahui apakah galur-galur tersebut mempunyai kemiripan genetik yang rendah atau jauh. Menurut Pabendon et al. (2005), galur Mr4 mempunyai kemiripan genetik yang rendah dengan galur Mr14 yaitu sekitar 0,27, yang berarti kedua galur memiliki hubungan genetik yang jauh sehingga jika disilangkan akan berpotensi menghasilkan turunan yang baik. Penelitian mengenai analisis korelasi kekerabatan berdasarkan data genotipe memanfaatkan marka SSRs dan morfologi pada jagung belum banyak dilakukan di
6
Galur-galur jagung manis yang diperoleh juga dilakukan analisis daya gabung sebagai penyaringan untuk mendapatkan calon tetua dalam persilangan dialel. Penelitian mengenai daya gabung pada jagung telah dilaksanakan oleh (Beck et al. 1990; Crossa et al. 1990; Vasal et al. 1992; Kang et al. 1995; Kim dan Ajala 1996; Wang et al. 1999; Mickelson et al. 2001; Betrán et al. 2002; Revilla et al. 2002; Betrán et al. 2003; Bhatnagar et al. 2004; Long et al 2004; Menkir dan Ayodele 2005) dan tanaman yang lain (Boye-Goni and Marcarian 1985; Nienhuis dan Singh 1986; Borges 1987; Tenkouano et al. 1998; Hartman and St. Clair 1999) pada berbagai karakter yang berbeda.
Berdasarkan uraian di atas dipandang perlu untuk melakukan penelitian
Analisis Jarak Genetik Berdasarkan Marka SSRs (Simple Sequence Repeats) dan Morfologi serta Analisis Daya Gabung untuk Pembentukan Hibrida Jagung Manis (Zea mays L. var. saccharata) sebagaimana bagan alir penelitian pada Gambar 1.
Tujuan Penelitian
1. Mendapatkan galur-galur jagung manis tahan terhadap penyakit bulai,
2. Memperoleh informasi kekerabatan galur jagung manis berdasarkan keragaman genetik memanfaatkan marka SSRs, dan korelasinya dengan penampilan morfologi.
3. Memperoleh informasi nilai daya gabung umum galur-galur jagung manis, yang akan dijadikan tetua dalam persilangan dialel.
4. Mendapatkan informasi nilai daya gabung umum, daya gabung khusus galur jagung manis, dan nilai heterosis hibrida jagung manis.
5. Mendapatkan satu atau lebih calon hibrida silang tunggal yang mempunyai potensi hasil tinggi dan tahan penyakit bulai
Hipotesis
1. Jarak genetik berdasarkan karakter genotipe berkorelasi dengan jarak taksonomi berdasarkan karakter morfologi jagung manis.
7
3. Terdapat galur jagung manis yang memiliki nilai daya gabung, nilai heterosis tinggi, dan tahan terhadap penyakit bulai (downy mildew) sebagai kandidat tetua pembentukan hibrida.
4. Hibrida potensial diperoleh dari pasangan tetua kelompok heterotik yang berbeda
Diagram Alir Penelitian
Kegiatan penelitian secara keseluruhan meliputi beberapa tahapan yaitu:
1. Analisis keragaman genetik galur-galur jagung manis (Zea mays L. var. saccharata) berdasarkan marka SSRs (Simple Sequence Repeats) dan morfologi.
2. Evaluasi daya gabung umum galur-galur jagung manis (Zea mays L. var. saccharata) menggunakan silang puncak.
3. Evaluasi daya gabung galur jagung manis (Zea mays L. var. saccharata) dan heterosis hibridanya menggunakan persilangan diallel.
8
Mr4 x JM Mr11 x JM Mr12 x JM Mr14 x JM
Mr4 x F1 Mr11 x F1 Mr12 x F1 Mr14 x F1
BC4F4 BC4F4 BC4F4 BC4F4
Keterangan:
- JM = varietas jagung manis. - P. Bulai = Penyakit bulai
Gambar 1 Bagan alir kegiatan penelitian POPULASI DASAR
Penelitian 1. Analisis Keragaman Genetik
Penelitian 2. Analisis Daya Gabung Umum
- Marka SSRs - Morfologi
- Gabungan SSRs dan morfologi
- Daya hasil - Ketahanan
Penyakit Bulai
Penelitian 3. Analisis Daya gabung dan Heterosis
- Daya hasil
- Ketahanan Penyakit Bulai
Informasi DG, Galur JM Tahan P. Bulai,
9
Dari peneltian 1 diperoleh pengelompokan galur jagung manis berdasarkan karakter genotipe memanfaatkan marka SSRs, dan pengelompokan galur berdasarkan kemiripan morfologi. Galur jagung manis yang terseleksi memiliki jarak genetik jauh digunakan pada percobaan 3.
Pada penelitian 2 dilakukan evaluasi daya gabung umum galur-galur jagung manis (Zea mays L. var. saccharata) menggunakan silang puncak. Informasi yang diperoleh pada percobaan ini adalah nilai efek daya gabung umum (DGU), dan galur jagung manis yang tahan penyakit bulai. Galur jagung manis yang terseleksi, dibandingkan dengan hasil percobaan 1 yang memiliki jarak genetik yang jauh. Hasil percobaan digunakan pada percobaan 3.
Pada penelitian 3 dilakukan evaluasi daya gabung galur jagung manis (Zea mays L. var. saccharata) dan heterosis hibridanya menggunakan persilangan diallel. Informasi yang diperoleh pada percobaan ini adalah nilai efek daya gabung khusus galur jagung manis sebagai tetua pembentukan varietas hibrida jagung manis dan nilai heterosisnya.
BAB. II
TINJAUAN PUSTAKA
Genetika Jagung Manis (Sweet Corn)
Jagung manis (Zea mays L. var. saccharata) merupakan salah satu sayur-mayur yang populer di negara-negara maju seperti Amerika, Brasil, Prancis dan di negara-negara berkembang. Jagung ini dikonsumsi dalam bentuk jagung muda yang direbus atau dibakar, dibuat sayur, perkedel, cream atau susu, sirup, dan bahan baku pembuatan permen karena rasanya manis dan enak. Di Europa, Cina, Korea, dan Jepan, digunakan untuk topping pizza. Rasa manis disebabkan oleh kandungan gula yang tinggi pada endosperm, mengkilap, dan tembus pandang ketika belum masak, dan bila kering berkerut. Jagung manis selain rasanya manis dan enak juga bermanfaat bagi kesehatan seperti mengurangi resiko terkena penyakit jantung dan kanker terutama jika dikonsumsi dalam bentuk jagung rebus. Kandungan nutrisi jagung manis diperlihatkan pada Tabel 1 (Larson 2003).
Tabel 1 Kandungan nilai nutrisi dalam biji jagung manis
Nilai nutrisi dalam biji jagung manis per 100 g (3,5 oz)
Energy 90 kkal 360 kJ
Karbohidrat 19 g
- Gula 3.2 g
- Dietary fiber 2.7 g
Lemak 1.2 g
Protein 3.2 g
Vitamin A equiv. 10 μg 1% Asam Folat (Vit. B9) 46 μg 12%
Vitamin C 7 mg 12%
Besi 0.5 mg 4%
Magnesium 37 mg 10%
Kalium 270 mg 6%
Sumber: database nutrisi USDA
11
kemanisan jagung ada tiga yaitu : (1) gen sugary (su); (2) gen sugary enhancer (se); dan (3) gen shrunken (sh2). Ketiga gen tersebut merupakan gen resesif sehingga harus ditanam terpisah dari varietas jagung biasa.
Penyakit Bulai (Downy Mildew)
Penyebab penyakit bulai pada tanaman jagung Peronosclerospora dilaporkan ada lima spesies (Renfro 1980 dikutip Wakman 2001). Namun di Indonesia hanya ada dua spesies yang dilaporkan yaitu P. maydis dan P. philippinensis, tetapi umumnya P. maydis terdapat dimana-mana, sedangkan P. philippinensis khusus terdapat di daerah Sulawesi Utara.
Penyakit bulai atau embun berbulu (downy mildew) memiliki banyak nama setempat, antara lain di Jawa Tengah dikenal dengan nama omo putih, omo londo, dan omo bule, di Jawa Timur, omo putih, dan potehan, sedangkan di Jawa Barat dikenal sebagai omo bodas dan hama lieur (Semangun 1996).
Serangan penyakit bulai dapat dilihat pada permukaan daun terdapat garis-garis sejajar tulang daun berwarna putih sampai kuning diikuti dengan garis-garis klorotik sampai coklat. Bila infeksi makin lanjut, tanaman terlihat kerdil dan tidak berproduksi, tetapi jika infeksinya terlambat tanaman masih ada kemungkinan untuk
berproduksi, hanya saja biji yang dihasilkan sudah terinfeksi patogen. Jamur berkembang secara sistemik, sehingga bila patogen mencapai titik tumbuh maka seluruh daun muda yang muncul mengalami klorotik, sedangkan daun pertama
sampai ke empat masih terlihat sebagian hijau. Ini merupakan ciri-ciri dari infeksi patogen melalui udara. Bila biji jagung sudah terinfeksi, maka bibit muda yang tumbuh memperlihatkan gejala klorotik pada seluruh daun dan tanaman cepat mati. Di permukaan bawah daun yang terinfeksi dapat dilihat banyak terbentuk tepung putih yang merupakan spora patogen tersebut (Agrios 1988).
12
dengan kelobot yang tidak menutup pada ujungnya, dan hanya membentuk sedikit biji.
Kajiwara (1974) dari hasil penelitiannya dengan menggunakan daun jagung kultivar Harapan menggambarkan proses perkembangan gejala penyakit bulai yang tercantum dalam Tabel 3.
Tabel 2 Perkembangan gejala penyakit bulai (downy mildew) Hari setelah
inokulasi Gejala
2 5 6 7 9
14-25
Muncul bintik-bintik hijau pucat (1-2 mm). Muncul garis-garis berwarna hijau kekuningan.
Garis-garis tadi semakin jelas. Beberapa daun berubah warna menjadi kuning.
Seluruh daun berwarna kuning dan beberapa diantaranya mulai layu.
Daun pertama sampai daun ke tiga mati. Garis-garis kuning yang jelas muncul pada daun ke empat.
Gejala serangan sistemik mulai terlihat pada daun-daun yang baru terbentuk.
Jamur dapat bertahan hidup sebagai mycelium dalam embrio biji yang terinfeksi jika kondisi biji lembab. Bila biji ditanam, jamurnya ikut berkembang dan menginfeksi bibit, selanjutnya dapat menjadi sumber inokulum. Infeksi terjadi melalui stomata daun jagung muda umur di bawah satu bulan jika pada permukaan daun terdapat air gutasi atau tetesan air serta jamur berkembang secara sistemik. Sporangia (konidia) dan sporangiofora dihasilkan pada permukaan daun yang basah dalam gelap. Sporangia berperan sebagai inokulum sekunder. Pembentukan spora patogen membutuhkan udara yang lembab (lebih dari 90%) dan hangat pada suhu sekitar 220 C serta gelap. Produksi sporangia (sporulasi) sangat banyak terjadi pada dini hari antara pukul 03.00 sampai 05.00 (Semangun 1996). selanjutnya oleh tiupan angin di pagi hari, spora tersebut tersebar sampai jarak jauh dan bila spora menempel pada daun jagung muda yang basah, maka dalam waktu satu jam spora tersebut sudah mulai berkecambah dan menginfeksi daun melalui stomata jika ada air gutasi atau embun (Subandi 1996).
13
yang dilakukan di Bogor menunjukkan bahwa konidium berkecambah paling baik
pada suhu 20C. Varietas-varietas jagung berbeda ketahanannya terhadap penyakit
bulai, ada yang tahan dan rentan. Hal ini ditentukan oleh gen-gen penyusun yang terdapat dalam tanaman jagung. Gambar 1 menunjukkan penampilan daun tanaman jagung yang terinfeksi penyakit bulai.
Gambar 2 Daun tanaman jagung yang terserang bulai
Pengendalian Penyakit Bulai (Downy Mildew)
Pemberantasan penyakit bulai harus dilakukan secara terpadu diantaranya penggunaan varietas resisten, penanaman serentak, tanaman yang sakit dicabut dan dikubur dalam tanah atau dibakar agar tidak menjadi sumber penyakit, pengaturan
pola tanam, dan penggunaan fungisida (Fagi et al. 1997). Pergiliran tanaman merupakan salah satu cara menekan tingkat serangan penyakit bulai. Karena tidak terbentuknya oospora memungkinkan patogen dapat bertahan di dalam daun, batang, kelobot, bunga jantan, bunga betina ataupun tongkol yang masih muda sehingga dapat bertahan dalam waktu yang cukup lama. Dengan demikian pertanaman jagung terus menerus sepanjang tahun akan menyebabkan patogen dapat bertahan terus.
14
fungisida yang dapat digunakan untuk menekan serangan bulai adalah fungisida sistemik. Namun cara yang paling baik untuk memberantas serangan penyakit dan hama adalah dengan merakit kultivar resisten (Singh 1986; Mochizuki 1974).
Penanganan pemberantasan penyakit bulai selain penggunaan fungisida sistemik juga diusahakan mendapatkan varietas baru yang resisten. Hal ini telah dilakukan karena dengan penggunaan fungisida akan membutuhkan biaya tambahan kepada petani.
Pemuliaan untuk Ketahanan terhadap Penyakit
Program pemulian untuk menghasilkan varietas resisten terhadap penyakit harus melalui beberapa tahapan yakni mendapatkan sumber ketahanan dan menentukan pola pewarisan karakter ketahanan tanaman inang serta karakter genetik dari interaksi antara inang dengan patogen (Allard 1960). Hal tersebut dapat terlaksana dengan baik apabila pengkajian dilakukan pada lingkungan epidemik bagi patogen, baik di laboratorium, rumah kaca, maupun di lapangan. Beberapa kendala yang sering ditemukan adalah penentuan dan penilaian ketahanan, identifikasi genetik dari karakter ketahanan yang melibatkan interaksi gen yang tidak sealel, dan kaitan gen.
Untuk mengantisipasi hal tersebut di atas maka pada setiap pelaksanaan pengujian dan seleksi ketahanan tanaman perlu diusahakan terciptanya lingkungan epidemik yang mampu memberikan kondisi epifitotik patogen. Langkah yang harus
dilaksanakan untuk menciptakan kondisi seluruh tanaman yang diuji terinfeksi patogen penyakit adalah dengan melakukan inokulasi buatan. Beberapa hal penting untuk keberhasilan inokulasi buatan tersebut adalah inokulum harus tetap bermutu tinggi, penerapan inokulasi diusahakan homogen untuk setiap tanaman, dan kondisi lingkungan pada saat inokulasi sesuai bagi pertumbuhan patogen, serta tanaman inang yang akan diuji harus bebas dari penyakit lain dan harus dalam keadaan fisiologik yang cocok untuk terjadinya serangan patogen.
15
resisten dan tanaman rentan dalam populasi yang bersegregasi. Varians ketahanan tersebut bersifat kontinyu dengan perubahan perbedaan ketahanan yang kecil (Herison 2002).
Pegetahuan tentang adanya gen-gen pengendalian karakter ketahanan terhadap penyakit bulai akan memudahkan pemulia tanaman untuk merakit varietas resisten. Pamin (1980) menyimpulkan bahwa terdapat keragaman genetik yang cukup luas terbukti dari pembentukan varietas sintetik yang tahan penyakit bulai, dengan pembentukan intercross S1 dan S2 telah berhasil diturunkan tingkat serangan penyakit bulai dari 60.8 persen pada bahan asal menjadi 22.4 persen pada intercross S1 dan 16.8 persen pada intercross S2.
Aday (1974) meringkas bahwa program pemuliaan yang dilakukan di Philipina guna menekan serangan bulai berlangsung dalam empat tahap yaitu:
a. isolasi dan seleksi galur-galur murni yang resisten terhadap bulai (DMR) (1953 – 1963).
b. skrening varietas lokal untuk ketahanan penyakit bulai pada areal yang luas dan introduksi plasmanutfah yang memiliki hasil tinggi (1964 – 1968).
c. pembentukan varietas hibrida persilangan antara varietas lokal tahan bulai dengan varietas introduksi plasmanutfah produksi tinggi (1967 – 1973).
d. perbaikan dalam populasi untuk ketahanan terhadap bulai dan seleksi karakter argonomi yang baik.
16 Marka Genetik
Marka genetik merupakan bagian kecil dari materi genetik yang mudah diidentifikasi. Menurut Liu (1998) penanda genetik ada tiga tipe (a) marka morfologi, protein (biokimia) dan DNA (molekuler). Marka morfologi mudah dilakukan tetapi sulit mengidentifikasi gen dalam jumlah besar jika populasi spesiesnya sedikit. Marka protein yang biasa digunakan adalah isozim, marka DNA dengan mengisolasi sebagian kecil daerah DNA. Menurut Tanksley dan McCouch (1997), marka morfologi merupakan secara visual dikarakterisasi secara fenotipik seperti warna bunga, bentuk biji, tipe tumbuh atau pigmentasi. Marka isozim adalah marka yang dapat membedakan enzim yang dideteksi melalui elektroforesis dan merupakan penanda spesifik. Keterbatasan dari marka-marka biokimia dan morfologi terbatas dalam jumlah dan dipengaruhi oleh faktor-faktor lingkungan atau fase perkembangan dari tanaman.
Marka molekuler pada awal perkembangannya diperkenalkan untuk mengatasi kesulitan seleksi secara konvensional. Apabila marka molekuler yang terpaut dengan gen-gen yang dimaksud sudah diidentifikasi, maka marka tersebut dapat membantu mengurangi ukuran populasi dan waktu yang dibutuhkan dalam program pemuliaan
per siklus seleksi. Beberapa kelebihan marka molekuler, khususnya memiliki kemampuan menyeleksi tanaman pada tahap pembibitan untuk sifat yang baru bisa diamati setelah tanaman tumbuh menjadi besar dan kemampuan menyeleksi sifat
yang sangat sulit bila menggunakan seleksi fenotipe saja yang membutuhkan waktu relatif panjang (Couch et al. 1991).
17
Primer SSR dibentuk berdasarkan pada daerah pengapit konservatif (conserved flanking region). Variasi dalam jumlah pengulangan untuk suatu batasan lokus diantara genotip-genotip yang berbeda dengan mudah dapat dideteksi dengan teknik PCR (Polymerase Chain Reaction) (Hamada et al. 1982; Powell et al. 1996). Kemudahan SSR dalam mengamplifikasi dan mendeteksi fragmen-fragmen DNA (Deoxyribo Nucleic Acid), serta tingginya tingkat polimorfisme yang dihasilkannya menyebabkan metode ini ideal untuk dipakai dalam studi genetik, terutama pada studi dengan jumlah sampel yang banyak. Selain itu, teknik PCR pada SSR hanya menggunakan DNA dalam jumlah kecil dengan daerah amplifikasi yang kecil, sekitar 100 - 300 bp (base-pair) dari genom. Selain itu, SSR dapat diaplikasikan
tanpa merusak bahan tanaman karena hanya sedikit saja yang digunakan dalam ekstraksi DNA atau dapat menggunakan bagian lain, seperti biji atau polen (Senior et al. 1996).
Marka SSRs juga bersifat multialelik dan mudah diulangi sehingga penggunaan marka SSRs lebih menarik dalam mempelajari keragaman genetik di antara genotip-genotip yang berbeda (Senior et al. 1998). Keunggulan lain adalah selain produk PCR dari SSR dapat dielektroforesis dengan gel agarose, juga dapat dielektroforesis dengan menggunakan gel akrilamid terutama pada alel suatu karakter memiliki tingkat polimorfis yang rendah, dimana gel agarose tidak mampu digunakan (Macaulay et al. 2001).
18 Keragaman Genetik pada Jagung
Dengan munculnya metode molekuler untuk menduga variasi genetik, berbagai studi mengenai keragaman genetik dan hubungannya dengan galur-galur inbrida jagung, hibrida dan bersari bebas. Keragaman molekuler pada jagung telah diteliti untuk berbagai kepentingan. Mum dan Dudley (1994) telah berhasil mengidentifikasi kelompok heterotik utama dan subgrup dalam set yang terdiri atas 148 galur inbrida Amerika Serikat (A.S.), menggunakan 46 marka RFLP.
Berdasarkan analisis molekuler dengan menggunakan 83 marka SSR, Lu dan Bernardo (2001) menyimpulkan bahwa keragaman genetik sejumlah galur inbrida A.S. terbaru telah menurun pada level genetik tetapi tidak pada level populasi jika
dibandingkan dengan galur-galur penting yang telah lama digunakan. George et al. (2004) menduga keragaman genetik untuk penyakit bulai terhadap 102 galur inbrida Asia menggunakan 76 marka SSR dan menyimpulkan bahwa aktivitas pemuliaan jagung di Asia tidak menyebabkan penurunan keragaman genetik dalam skala besar pada daerah tertentu dimana aktivitas dilakukan.
Korelasi Marka DNA dengan Marka Fenotipik
Penelitian mengenai hubungan antara marka-marka dan karakter-karakter yang
bernilai ekonomi, penting dalam kaitannya dengan pemanfaatan marka molekuler sebagai alat bantu dalam pemuliaan. Perhatian utama dalam pogram pemuliaan jagung hibrida adalah mengidentifikasi galur-galur murni dimana dari hasil
persilangannya akan diperoleh tingkat heterosis yang optimal (Lee et al. 1989). Estimasi kedekatan genetik antara tanaman bermanfaat di dalam studi evolusi populasi atau spesies dan dalam perencanaan persilangan untuk hibrida atau dalam pengembangan kultivar homozigous (Cox et al. 1985).
19
Estimasi jarak genetik sejumlah galur murni jagung yang dilakukan oleh Lee et al. (1989) berdasarkan MRD (Modified Rogers’ Distance) menunjukkan bahwa hasil (ton/ha) dan kemampuan daya gabung khusus (DGK) mempunyai korelasi yang nyata dengan MRD pada enam dari 10 peta kromosom jagung. Oleh sebab itu, gerombolisasi galur-galur jagung ke dalam kelompok heterosis sebelum pengujian di lapangan akan memungkinkan bagi peneliti pemulia untuk mengurangi biaya karena dapat menghindari terjadinya persilangan di dalam kelompok heterosis.
Riday et al. (2003) membandingkan antara nilai jarak genetik dengan morfologi dan heterosis pada Medicago sativa sub sp. sativa dan subsp. Falcata, menemukan bahwa jarak genetik berdasarkan AFLP tidak berkorelasi dengan hasil
daya gabung khusus (DGK) namun sebaliknya matriks jarak berdasarkan morfologi terhadap 17 karakter agronomik dan karakter kualitatif mempunyai korelasi yang nyata dengan heterosis.
Metode Dialel
Metode dialel merupakan cara analisis keturunan untuk daya gabung, baik daya gabung umum maupun daya gabung khusus (Hallauer and Miranda 1981). Analisis dialel membantu para pemulia menentukan pola heterosis antar populasinya serta
memilih bahan dan metode yang akan digunakan dalam program pemuliaannya. Persilangan dialel yakni persilangan yang melibatkan sejumlah genotip (varietas, galur atau klon) dalam semua kombinasi. Masing-masing genotip
mempunyai kesempatan untuk disilangkan dengan genotip lain, bahkan dapat ditambah dengan persilangan sendiri genotip itu. Melalui persilangan dialel dapat diketahui kombinasi mana yang cocok dipasangkan sehingga dapat menghasilkan suatu varietas yang lebih baik.
Menurut Griffing (1956), terdapat empat macam metode percobaan yang digunakan untuk analisis dialel yakni:
1. Metode 1: kombinasi lengkap p2 , terdiri dari tetuanya, F1, dan persilangan resiprokalnya
20
4. Metode 4: ½ p (p-1) kombinasi, terdiri dari F1 saja tanpa tetua dan resiprokalnya.
Penggunaan salah satu metode dialel tergantung dari tujuan analisisnya atau dihubungkan dengan penyederhanaan analisisnya. Berdasarkan cara penentuan tetua-tetua yang dipakai dalam persilangan, interpretasi hasil analisis dialel dibedakan ke dalam dua model, yaitu :
1. Model I atau model tetap (fixed model), dengan menggunakan tetua-tetua tertentu yang merupakan genotip yang dimaksud (reference genotype). Estimasi yang diperoleh hanya berlaku untuk genotip yang dimasukkan dalam pengujian dan tidak berlaku untuk populasi lain.
2. Model II atau model acak, dengan menggunakan tetua-tetua yang merupakan contoh acak dari populasi tetua yang dimaksud (reference population). Estimasi yang diperoleh diinterpretasikan berkaitan dengan populasi tetua, dari mana genotip yang dievaluasi diambil secara acak (Griffing 1956).
Dalam model I, tetua-tetua yang dipelajari atau diteliti adalah populasi, maka
cukup tepat untuk menganalisis galur-galur elit yang terpilih. Analisis model I digunakan untuk mengestimasi efek DGU dan DGK yang hasilnya berlaku hanya untuk tetua-tetua yang dievaluasi saja. Hasil akan berubah bila tetua-tetua tersebut
dievaluasi bersama-sama dengan tetua lain yang berbeda. Sedangkan tujuan utama analisis model II adalah estimasi komponen varians populasi yang dimaksud (reference population) dan berkaitan dengan tipe aksi gen dalam populasi (Moentono 1985).
Daya Gabung (Combining Ability)
21
Daya gabung ada dua macam yakni daya gabung umum (general combining ability) dan daya gabung khusus (specific combining ability). Daya gabung umum (DGU) adalah nilai rata-rata dari galur-galur dalam seluruh kombinasi persilangan bila disilangkan dengan galur-galur lain. Daya gabung umum yang baik adalah nilai rata-rata kombinasi persilangan mendekati nilai rata-rata keseluruhan persilangan. Daya gabung khusus (DGK) adalah penampilan kombinasi pasangan persilangan tertentu. Bila nilai pasangan persilangan tertentu lebih baik daripada nilai rata-rata keseluruhan persilangan yang terlibat, dikatakan daya gabung khususnya baik (Poehlman dan Sleeper 1990).
Daya gabung umum merupakan simpangan dari nilai rata-rata seluruh
persilangan, sehingga nilai daya gabung umum dapat positif atau negatif. Dengan demikian jumlahnya sama dengan nol. Jadi nilai daya gabung umum merupakan angka yang relatif terhadap nilai daya gabung umum yang lain. DGU yang besar menunjukkan tetua/galur yang bersangkutan mempunyai kemampuan bergabung dengan baik, sedangkan nilai DGU yang rendah menunjukkan bahwa tetua tersebut mempunyai kemampuan bergabung yang lebih jelek daripada tetua yang lain. Nilai positif atau negatif dari DGU tergantung pada karakter yang diamati dan bagaimana cara menilainya.
Daya gabung yang diperoleh dari suatu persilangan antar kedua tetua, dapat memberikan informasi tentang kombinasi-kombinasi yang dapat memberikan turunan yang berpotensi hasil tinggi. Hasil tinggi dapat diperoleh apabila kombinasi tersebut memiliki nilai heterosis dan daya gabung khusus yang besar. Galur yang mempunyai efek daya gabung umum yang tinggi tidak selalu memberikan efek daya gabung khusus yang tinggi pula (Silitonga et al. 1993).
22 Heterosis
Pada umumnya apabila dua tanaman yang berlainan (unrelated or distantly related individuals) disilangkan, maka turunannya sering memperlihatkan gejala heterosis atau umumnya disebut vigor hibrida (Hybrid Vigour) (North 1979 dikutip Baihaki 1989). Tanaman turunan pertamanya memperlihatkan pertumbuhan yang lebih subur dan terkadang lebih genjah daripada kedua tetuanya. Untuk persilangan yang melebar, misalnya persilangan antar species tanaman atau antar genera, dapat menghasilkan tanaman turunan pertamanya menunjukkan pertumbuhan yang lemah. Dengan demikian terlihat bahwa terdapat variasi penampilan gejala heterosis dan tergantung pada materi yang digunakan dalam persilangan.
Fenomena heterosis telah banyak dimanfaatkan secara intensif pada pemuliaan jagung dalam membentuk kultivar hibrida. Keberhasilan jagung dalam memanfaatkan heterosis mendorong pemulia menggunakannya pada jenis tanaman lain seperti pada tanaman terung, tomat, mentimum, sorgum, dan lain-lain. (Dahlan et al. 1998).
Terdapat tiga konsep heterosis yang dapat menjelaskan gejala heterosis. Konsep pertama adalah konsep yang berdasarkan hipotesis bahwa vigor hibrida merupakan hasil terkumpulnya gen-gen dominan yang baik (Favourable dominant genes) dalam satu genotip tanaman dan dikenal sebagai hipotesis dominan.
Menurut Shull dan East (1908) dikutip Sriani et al. 2007, Davenport merupakan orang pertama yang mengajukan hipotesis dominan dalam upaya menjelaskan secara genetik mengenai gejala heterosis. Konsep kedua didasarkan kepada hipotesis bahwa vigor hibrida merupakan hasil penampilan superioritas heterozigositas terhadap homozigositas, artinya bahwa individu yang penampilan superior adalah individu yang memiliki jumlah alil dalam keadaan heterosigos yang terbanyak, konsep ini dikenal sebagai hipotesis overdominan.
23 BAB III
Analisis Keragaman Genetik Galur-galur Jagung Manis
(Zea mays L. var. saccharata) Berdasarkan Marka SSRs
(Simple Sequence Repeats) dan Morfologi.
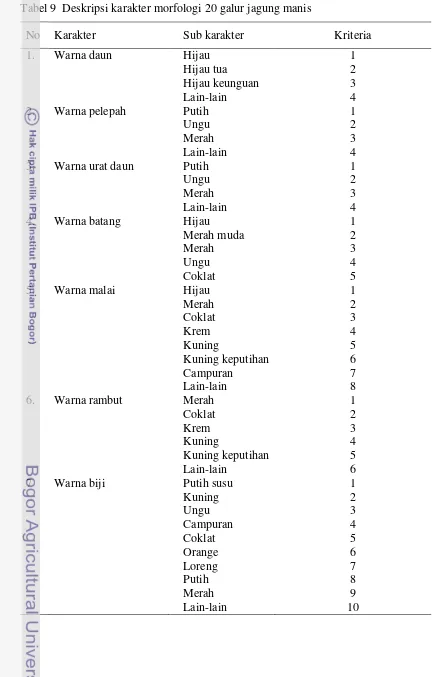
ABSTRAK
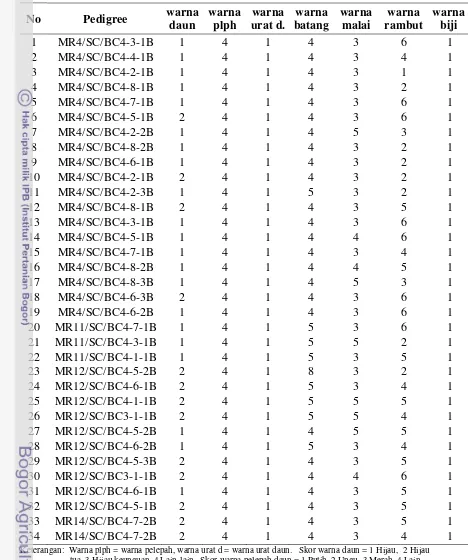
Penelitian ini bertujuan untuk (a) mengetahui hubungan genetik dan tingkat keragaman genetik 45 galur jagung manis memanfaatkan marka SSRs dan karakter morfologi, (b) mendapatkan informasi kelompok heterotik dan nilai jarak genetik yang akan digunakan untuk seleksi tetua hibrida. Data yang dianalisis adalah data genotype dan karakter morfologi. Data genotipe meliputi tingkat polimorfisme, keragaman genetik, dan jarak genetik menggunakan koefisien Jaccard. Karakter morfologi meliputi warna daun (skor 1-4), warna urat daun (skor 1-4), warna pelepah (skor 1-4), warna batang (skor 1-5), warna malai (skor 1-8), warna rambut (skor 1-6), warna biji (skor 1-10). Hasil penelitian menunjukkan bahwa 20 primer yang digunakan memiliki rata-rata tingkat polimorfisme 0,69 dan total alel 69 dengan rata-rata 3,45 alel/lokus. Nilai jarak genetik 39 galur jagung manis berdasarkan marka SSRs berkisar 0,18 sampai 0,78. Korelasi antara matrik kemiripan berdasarkan marka SSRs dengan matrik kemiripan berdasarkan morfologi memiliki korelasi yang kuat berdasarkan kriteria goodness of fit. Galur yang dapat direkomendasikan untuk dipasangkan dengan galur lainnya adalah Mr4/SC/BC4-2-1B, Mr4/SC/BC4-6-Mr4/SC/BC4-2-1B, Mr11/SC/BC4-2-Mr4/SC/BC4-2-1B, Mr11/SC/BC4-4-Mr4/SC/BC4-2-1B, Mr12/SC/BC4-6-1B, dan Mr12/SC/BC3-3-1B.
24
Genetik Diversity Analysis of Sweet Corn (Zea mays L. var. saccharata) Lines based on Simple Sequence Repeats (SSRs) Markers and Morphological
ABSTRACT
The present study is undertaken to evaluate genetic divergence among inbreds and to assess the relationship between heterosis and parental genetic distance. The objective of this study are (a) to estimate the level of genetic diversity among 34 inbred lines from different population sources based on microsatellite or single sequence repeats (SSR) markers and pedigree data, and (b) to obtain information of heterotic groups and genetic distance of the inbreds as used in parental hybrid selection. The data are analyzed based on genotype and morphological trait. The data genotype by polymorphic information content (PIC), genetic diversity, and genetic distance
(Jaccard’s coefficient). Morphological trait are leaf color (score 1-4), vein color (score 1-4), sheath color (score 1-4), coloration of steam (score 1-5), anther color (score 1-8), silk color (score 1-6), seed color (score 1-10). The results showed that the used 0f 20 primers had 0.69 of polymorphism level and total 0f 69 alleles with mean of 3.45 alleles per locus. Genetic distance value ranging from 0.18 to 0.78. correlation between similarity matrices SSR-based and similarity matrices morphological-based using goodness of fit criteria is strong. Lines Mr4/SC/BC4-2-1B, Mr4/SC/BC4-6-Mr4/SC/BC4-2-1B, Mr11/SC/BC4-2-Mr4/SC/BC4-2-1B, Mr11/SC/BC4-4-Mr4/SC/BC4-2-1B, Mr12/SC/BC4-6-1B, and Mr12/SC/BC3-3-1B were recommended for diallel cross.
25
PENDAHULUAN
Program utama pemuliaan hibrida membutuhkan prediksi bagaimana penampilan hibrida yang dihasilkan. Pemulia selalu tertarik dalam memilih galur tetua, yang akan memberikan kombinasi heterotik tanpa perlu membuat semua persilangan yang mungkin di antara semua tetua potensial. Pemilihan tetua dengan keragaman genetik genetik yang luas dalam populasi genetik memegang peranan penting terhadap kemajuan program pemuliaan.
Selama ini karakter morfologi dan informasi pedigree telah banyak digunakan dalam menentukan jarak genetik dan hubungan kekerabatan dalam sejumlah plasma nutfah, sehingga membentuk sejumlah kultivar. Walaupun hasilnya kadang bias karena karakter-karakter morfologi sering tidak tepat menjelaskan hubungan genetik antar individu oleh adanya interaksi lingkungan, dan kadang-kadang data pedigree tidak tersedia untuk semua materi. Oleh karena itu informasi keragaman genetik secara morfologi perlu didukung oleh data molekuler, untuk memperjelas perbedaan dan hubungan kekerabatan antara galur-galur yang dimiliki.
Menurut Weising et al. (1995) karakterisasi genetik yang didasarkan pada penanda fenotip biasanya dipengaruhi oleh lingkungan makro dan mikro, serta umur
suatu individu. Kesulitan lain akan terjadi apabila karakter kuantitatif yang diatur oleh banyak gen tersebut terekspresi pada akhir pertumbuhan, seperti karakter hasil. Oleh karena itu karakterisasi fenotip perlu didukung oleh karakterisasi yang
dilakukan melalui penanda molekuler. Lefebvre et al. (2001) menjelaskan bahwa penanda molekuler dapat memberi gambaran hubungan kekerabatan yang akurat antarspesies maupun kerabat jauhnya, karena analisis DNA sebagai material genetik tidak dipengaruhi oleh kondisi lingkungan.
Pengembangan marka molekular sebagai alat bantu untuk memprediksi keragaman genetik pada level DNA suatu tanaman (Melchinger dan Gumber, 1998). Khususnya, marka SSR menunjukkan potensi untuk sidik jari (finger printing) DNA dari genotipe jagung dalam kaitan dengan deteksi tingkat polymorphism (Smith et al. 1997), system analisisnya otomatis, dan mempunyai ketelitian tinggi dan repeatability(=daya pengulangan tinggi) (Heckenberger et al. 2002).
26
Warburton et al. (2002); Pabendon et al. (2007); Yu et al. (2007); tetapi analisis korelasi keragaman genetik dan morfologi masih sedikit dipelajari terutama pada jagung manis.
Tujuan penelitian ini adalah untuk (a) mengetahui hubungan genetik dan tingkat keragaman genetik galur-galur jagung manis memanfaatkan marka SSRs dan karakter morfologi, (b) mendapatkan informasi kelompok heterotik dan nilai jarak genetik yang akan digunakan untuk seleksi tetua hibrida.
BAHAN DAN METODE
Penelitian dilaksanakan di Kebun Percobaan Balai Penelitian Tanaman Serealia Maros dan Laboratorium Molekuler, Balai Penelitian Tanaman Serealia (Balitsereal) Maros dari Pebruari sampai Mei 2009.
Percobaan 1 : Analisis keragaman genetik berdasarkan marka SSRs.
Bahan Genetik
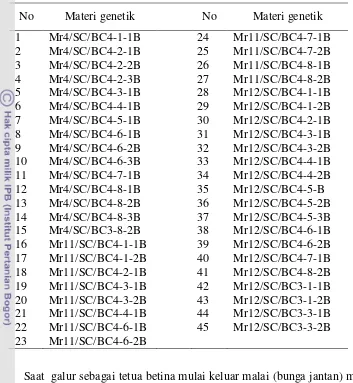
Materi tanaman yang digunakan adalah 45 galur jagung manis (Tabel 3), merupakan galur hasil persilangan backcross ke empat (BC4F4) antara jagung manis dengan galur Mr4, Mr11, Mr12, dan Mr14.
Tabel 3 Materi tanaman yang digunakan dalam penelitian
No Materi genetik No No Materi genetik
27 Percobaan di Laboratorium
Pelaksanaan
Biji ditanam sebanyak 10 individu untuk masing-masing galur pada baki yang menggunakan media tanah. Proses isolasi, amplifikasi, dan visualisasi pola pita
DNA mengikuti prosedur George et al. (2004) dengan beberapa modifikasi. Materi tanaman yang digunakan untuk ekstraksi DNA adalah tanaman yang berumur 10
sampai 15 hari setelah tanam.
Isolasi DNA
Bagian tanaman yang diambil adalah daun muda yang telah membuka sempurna sebanyak 5 - 8 individu tanaman dipotong-potong kecil, dicampur kemudian ditimbang 0,4 g per sampel. Sampel daun tersebut diisolasi DNAnya
dengan bantuan buffer (0,7 ml per sampel) di laboratorium, masing-masing sampel digerus menggunakan mortal sampai halus dan dimasukkan ke dalam tabung ependorf berukuran 2,0 ml, ditambahkan 0,5 ml buffer ektraksi CTAB panas yang
mengandung -mercaptoethanol konsentrasi 2 l/ml larutan kemudian diinkubasi
pada suhu 65o C dalam waterbath dan didinginkan ±30 menit.
Amplifikasi dan Visualisasi Pola Pita DNA
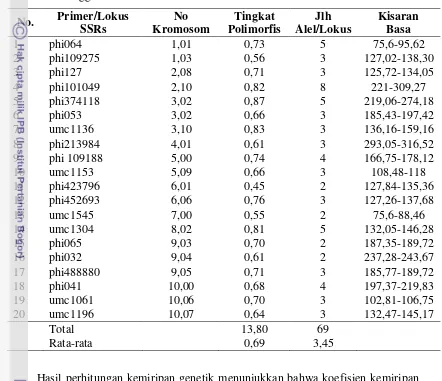
Primer mikrosatelit yang digunakan sebanyak 20 primer (forward dan reverse),
diperoleh dari Research Genetik, Inc, dan dari Invitrogen. Primer-primer tersebut diseleksi berdasarkan tingkat polimorfisme (referensi dari CIMMYT) (Tabel 4).
28 Pengamatan Data Molekuler
Analisis data molekuler dilakukan berdasarkan hasil skoring pita DNA yang muncul pada plate. Pita DNA diberi skor berdasarkan penampilan pita DNA ditransformasi ke dalam kode data biner dengan cara: jika ada pita diberi skor satu (1) dan jika tidak ada pita diberi skoring nol (0). Pita yang tidak sempurna dan tidak jelas diberi skor 9 (missing data). Jika ada galur yang menghasilkan banyak pita maka pita yang paling jelas diberi skor ‘1’ sedangkan yang lainnya diberi skor ‘9’. Data pada kolom menunjukkan inbrida sedangkan baris menunjukkan lokus SSRs.
Tabel 4 Sekuen dari 20 marka mikrosatelit yang digunakan dalam penelitian
No. Primer Bin No. Repeat
type Primer Sequence
1 phi109275 1.03 AGCT CGGTTCATGCTAGCTCTGC // GTTGTGGCTGTGGTGGTG 2 phi96100 2.01 ACCT AGGAGGACCCCAACTCCTG // TTGCACGAGCCATCGTAT 3 phi374118 3.02 ACC TACCCGGACATGGTTGAGC // TGAAGGGTGTCCTTCCGAT
4 phi079 4.05 AGATG TGGTGCTCGTTGCCAAATCTACGA // GCAGTGGTGGTTTCGAACAGACAA 5 phi109188 5.03 AAAG AAGCTCAGAAGCCGGAGC // GGTCATCAAGCTCTCTGATCG
6 phi299852 6.07 AGC GATGTGGGTGCTACGAGCC // AGATCTCGGAGCTCGGCTA 7 phi328175 7.04 AGG GGGAAGTGCTCCTTGCAG // CGGTAGGTGAACGCGGTA 8 phi233376 8.09 CCG CCGGCAGTCGATTACTCC // CGAGACCAAGAGAACCCTCA
9 phi065 9.03 CACTT AGGGACAAATACGTGGAGACACAG // CGATCTGCACAAAGTGGAGTAGTC 10 umc1196 10.07 CACACG CGTGCTACTACTGCTACAAAGCGA // AGTCGTTCGTGTCTTCCGAAACT 11 phi109642 2.03-2.04 ACGG CTCTCTTTCCTTCCGACTTTCC // GAGCGAGCGAGAGAGATCG 12 phi101049 2.10 AGAT CCGGGAACTTGTTCATCG // CCACGTCCATGATCACACC 13 phi102228 3.06 AAGC ATTCCGACGCAATCAACA // TTCATCTCCTCCAGGAGCCTT
14 phi093 4.08 AGCT AGTGCGTCAGCTTCATCGCCTACAAG // AGGCCATGCATGCTTGCAACAATGGATACA 15 umc1153 5.09 (TCA)4 CAGCATCTATAGCTTGCTTGCATT // TGGGTTTTGTTTGTTTGTTTGTTG
16 phi423796 6.01 AGATG CACTACTCGATCTGAACCACCA // CGCTCTGTGAATTTGCTAGCTC 17 phi114 7.03 GCCT CCGAGACCGTCAAGACCATCAA // AGCTCCAAACGATTCTGAACTCGC 18 phi420701 8.00 CCG GATGTTTCAAAACCACCCAGA // ATGGCACGAATAGCAACAGG 19 phi448880 9.06-9.07 AAG CGATCCGGAGGAGTTCCTTA // CCATGAACATGCCAATGC 20 phi96342 10.02 ATCC GTAATCCCACGTCCTATCAGCC // TCCAACTTGAACGAACTCCTC * Sumber: Applied Biotechnology Center at CIMMYT, Mexico
Analisis Data Genotipik
Analisis data molekuler dilakukan berdasarkan hasil skoring pita DNA yang muncul pada plate, hasil skoring dalam bentuk data biner. Tingkat polimorfisme (PIC = Polimorphism Information Content) dari primer yang digunakan dihitung untuk masing-masing marka SSRs (Smith et al. 1997), dengan formula:
n fi PIC1 2
1 i = 1, 2, 3,………n , dimana
f
i2 adalah frekuensi alel ke-i.29 Analisis Tingkat Heterosigositas
Salah satu kelebihan dari metode SSRs adalah deteksinya kodominan sehingga lokus yang heterozigot akan dapat dibedakan dari lokus homozigot. Lokus yang homozigot akan mucul hanya satu pita/alel per primer per genotipe. Jika lebih dari satu alel berarti lokus tersebut heterozigot. Analisis ini penting untuk menghindari terseleksinya genotipe-genotipe dengan tingkat heterosigositas yang tinggi dimana pada pengamatan secara fenotipik tidak terdeteksi karena pengaruh faktor lingkungan. Tingkat heterosigositas untuk setiap genotipe dapat diketahui dengan formula:
x 100%
digunakan yang
SSRs lokus Total
t heterozigo lokus
Jumlah sitas
heterozigo
Persentase
Untuk memperoleh hasil analisis data yang lebih akurat, maka dalam penelitian ini hanya genotipe yang mempunyai tingkat heterozigositas <20% yang dianalisis lebih
lanjut.
Estimasi Kekerabatan Genetik dan Analisis Klaster
Tingkat kemiripan genetik (GS=genetik similarity) diestimasi dengan
menggunakan koefisien Jaccard (Rohlf, 2000) dengan formula:
n u
m S
,
dimana m = jumlah pita (alel) DNA yang sama posisinya, n = total pita (alel) DNA, dan u = jumlah pita (alel) DNA yang tidak sama posisinya. Kemiripan genetik dianalisis berdasarkan Unweighted Pair Group Method using Arithmetic Averages
30
Percobaan 2. Korelasi antara tingkat kemiripan berdasarkan marka SSRs dengan kemiripan berdasarkan morfologi
Bahan Genetik
Sebanyak 20 materi genetik yang DNA-nya dapat diamplifikasi dan terkarakterisasi di lapangan (Tabel 5).
Tabel 5 Materi tanaman yang digunakan dalam analisis berdasarkan marka SSRs dan morfologi
No Materi genetik No Materi genetik
1 Mr4/SC/BC4-2-1B 11 Mr4/SC/BC4-8-3B 2 Mr4/SC/BC4-2-2B 12 Mr11/SC/BC4-1-1B 3 Mr4/SC/BC4-2-3B 13 Mr11/SC/BC4-3-1B 4 Mr4/SC/BC4-4-1B 14 Mr12/SC/BC4-1-1B 5 Mr4/SC/BC4-5-1B 15 Mr12/SC/BC4-5-1B 6 Mr4/SC/BC4-6-1B 16 Mr12/SC/BC4-5-2B 7 Mr4/SC/BC4-6-2B 17 Mr12/SC/BC4-5-3B 8 Mr4/SC/BC4-7-1B 18 Mr12/SC/BC4-6-1B 9 Mr4/SC/BC4-8-1B 19 Mr12/SC/BC4-6-2B 10 Mr4/SC/BC4-8-2B 20 Mr12/SC/BC3-3-1B
Pelaksanaan
Materi genetik galur jagung