MENGGUNAKAN TEKNIK REAL TIME RT-PCR
AMINAH
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERYATAAN MENGENAI TESIS DAN SUMBER INFORMASI
Dengan ini saya menyatakan bahwa tesis Kajian Persistensi dan Penularan
Virus Avian Influenza di Peternakan Itik menggunakan Teknik Real Time
RT-PCR adalah karya saya dengan arahan dari komisi pembimbing dan belum
diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber
informasi yang berasal atau dikutip dari karya yang diterbitkan mauoun tidak dari
penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di
bagian akhir tesis ini.
Bogor, Februari 2012
Aminah
AMINAH. The Study of Persistence and Transmission of Avian Influenza Virus
in Duck Farm using Real Time RT-PCR Technique. Under supervision of
SURACHMI SETIYANINGSIH and IDWAN SUDIRMAN.
Highly pathogenic avian influenza (HPAI) H5N1 virus has been a major
threat to poultry industry and human health in Indonesia over the past several
years. The existence of backyard and free-range duck raising system has been
hypothesized to play role in the disease circulation. This study investigated H5
virus infection and circulation occurring in three types of free-range duck farms in
Indramayu District, West Java, Indonesia from September 2009 to March 2010.
One hundred and eighty pairs of cloacal-oropharyngeal swab samples from
sentinel ducks placed in six farms of the three farm types were collected every
month for seven months period and screened in pools for influenza A virus. In
addition to the sentinel ducks, 30 non-sentinel ducks were included at the first and
the last sampling month. Of the total 648 pool samples collected for seven months,
91 pools (14%) were found influenza A positive at repeated events. The virus was
more commonly found on type 1 farms followed by type 2 and 3 farms. The
individual swab samples of influenza A positive pools were tested for H5 subtype.
Of the total 91 pools of influenza A positive samples, 455 individual samples
were tested for H5 subtype and 50 (11%) individual samples were found H5
positive. The H5 subtype was found at certain points of sampling time and was
more commonly found in type 2 farms followed by type 1 and was not found on
type 3 farms. The H5 virus was likely to be transmitted within duck farm but was
unlikely to be maintained for a long time.
RINGKASAN
Avian influenza (AI) telah menjadi masalah global maupun nasional yang
mengakibatkan kerugian besar bagi industri perunggasan. Virus highly pathogenic
avian influenza (HPAI) H5N1 bersifat 100% mematikan bagi ayam dan unggas
gallinaceous lainnya sedangkan unggas air seperti itik dapat mengeluarkan virus
melalui saluran pernafasan dan pencernaan dengan sedikit atau tidak ada gejala
penyakit. Unggas air merupakan reservoir utama virus influenza A dan dapat
menularkannya ke unggas domestik dan mamalia, termasuk manusia. Keberadaan
sistem pemeliharaan itik skala rumah tangga dan itik angon berperan dalam
peredaran penyakit AI. Penelitian ini mengamati infeksi dan peredaran virus AI
subtipe H5 di tiga tipe peternakan itik yang ada di kabupaten Indramayu, Jawa
Barat, Indonesia dari bulan September 2009 hingga Maret 2010. Seratus delapan
puluh pasang sampel usap kloaka dan orofaringeal itik sentinel yang ditempatkan
di enam peternakan dari ketiga tipe diambil setiap bulan selama tujuh bulan dan
diperiksa keberadaan virus influenza A dalam pool. Selain dari itik sentinel,
sampel juga diambil dari itik non-sentinel pada bulan pertama dan terakhir
pengambilan sampel. Dari total 614 pool sampel usap kloaka dan orofaringeal
yang berhasil dikoleksi selama tujuh bulan, didapati 98 (16%) pool positif VAI
secara berulang pada bulan-bulan tertentu yang menunjukkan bahwa VAI
bersirkulasi di satu peternakan untuk waktu yang lama dan mungkin melibatkan
lebih dari satu strain virus meskipun shedding virus terjadi dalam rentang waktu
tertentu pada tingkat pooldengan perbandingan sampel usap kloaka positif hampir
seimbang dengan usap orofaringeal yaitu masing-masing 52 (53,1%) dan 46
(46,9%). Virus AI lebih sering ditemukan pada peternakan tipe 1, diikuti tipe 2
dan 3. Sampel usap individual dari pool yang positif influenza A diperiksa
terhadap keberadaan subtipe H5. Dari total 98 sampel pool yang positif influenza
A, diperiksa 453 sampel individu dan didapati 49 (10,9%) sampel positif subtipe
H5 dengan perbandingan sampel usap kloaka dan orofaringeal positif H5
masing-masing 18 (36,7%) dan 31 (63,3%) serta rata-rata konsentrasi virus yang
diekskresikan melalui kloaka sedikit lebih tinggi dibandingkan orofaring, terlihat
dari nilai Ct masing-masing 27,68 (STD 5,32) dan 27,83 (STD 5,64). Subtipe H5
menghilang seiring kematian itik yang terinfeksi atau pembersihan virus
(clearance). Subtipe H5 ditemukan pada titik waktu tertentu pengambilan sampel
dan lebih sering ditemukan pada peternakan tipe 2, diikuti tipe 1 dan tidak
ditemukan pada tipe 3. Virus influenza A selain H5 lebih persisten dalam populasi
itik angon dibandingkan dengan subtipe H5 yang hanya muncul pada titik waktu
tertentu. Virus influenza A lebih persisten pada peternakan itik angon namun
penularan lebih mudah terjadi pada itik yang dikandangkan. Keberadaan VAI H5
di peternakan menunjukkan peran itik angon sebagai reservoir dan sumber
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan
atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan,
penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau
tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan
yang wajar IPB
Dilarang mengumumkan dan memperbanyak sebagian atau seluruh karya tulis
KAJIAN PERSISTENSI DAN PENULARAN
VIRUS AVIAN INFLUENZA DI PETERNAKAN ITIK
MENGGUNAKAN TEKNIK REAL TIME RT-PCR
AMINAH
Tesis
Sebagai salah satu syarat untuk memperoleh gelar
Magister Sains pada
Program Studi Mikrobiologi Medik
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Judul Penelitian : Kajian Persistensi dan Penularan Virus Avian Influenza di Peternakan Itik menggunakan Teknik Real Time RT-PCR
Nama : Aminah
Nomor Pokok : B253090011
Program Studi : Mikrobiologi Medik
Disetujui:
Komisi Pembimbing
drh. Surachmi Setiyaningsih, Ph.D Ketua
Dr. drh. Idwan Sudirman Anggota
Diketahui:
Ketua Program Studi Mikrobiologi Medik
A.n. Dekan Sekolah Pascasarjana IPB Sekretaris Program Magister
Prof. Dr. drh. Fachriyan H Pasaribu Prof. Dr. Ir. Nahrowi, M.Sc
KATA PENGANTAR
Puji syukur kepada Allah SWT atas limpahan berkahNya sehingga penulis dapat menyelesaikan penelitian dan penyusunan tesis yang berjudul “Kajian Persistensi dan Penularan Virus Avian Influenza di Peternakan Itik mengunakan Teknik Real Time RT-PCR” ini. Karya ilmiah ini disusun sebagai salah satu syarat dalam menyelesaikan studi program Magister pada Sekolah Pascasarjana Institut Pertanian Bogor.
Terima kasih penulis sampaikan kepada drh. Surachmi Setiyaningsih, PhD atas kesempatan dan segala bimbingan yang diberikan sehingga karya ilmiah ini dapat terwujud, kepada Dr. drh. Idwan Sudirman atas bimbingan dan arahan dalam diskusi-diskusi selama penyusunan tesis, dan kepada Prof. Dr. drh. Retno D. Soejoedono, MS sebagai penguji luar komisi pada ujian tesis ini. Terimakasih kepada seluruh staf pengajar serta pegawai laboratorium program studi Mikrobiologi Medik atas curahan ilmu dan tenaga yang tiada henti selama penulis menyelesaikan studi.
Terima kasih kepada Colorado State University (CSU) atas dukungan dana penelitian yang penulis kerjakan, khususnya kepada Dr. Kristy Pabilonia dan Christina Weller dari CSU Veterinary Diagnostic Laboratory atas pelatihan yang diberikan. Demikian juga kepada rekan-rekan dokter hewan yang tergabung
dalam Center for Indonesian Veterinary Analytical Studies (CIVAS) atas kerja
keras selama pengambilan sampel di lapangan.
Terima kasih yang tak terhingga penulis sampaikan kepada Dr. Yety Rochwulaningsih, MS atas segala dukungan agar penulis melanjutkan studi serta kepada keluarga besar Bukittinggi dan Sragen atas kebersamaan dan kehangatan keluarga yang penulis terima. Tidak lupa kepada rekan-rekan drh. Emilia, Yuliana Radja Riwu, Wury Kadarsih, Wiwin Mukti, dan Zakiyah Widowati untuk semua persahabatan. Terimakasih juga kepada semua pihak yang mendukung penelitian serta penyusunan tesis ini.
Semoga tesis ini bermanfaat dan menginspirasi banyak pihak untuk giat melakukan penelitian dan memberikan yang terbaik bagi kemajuan ilmu pengetahuan dan kesejahteraan bangsa. Segala saran dan masukan yang menunjang demi kebaikan kedepan kami terima dengan tangan dan pikiran terbuka.
Bogor, Februari 2012
Penulis lahir di Bukittinggi Sumatera Barat pada tanggal 10 Oktober 1983,
menyelesaikan pendidikan dasar hingga menengah atas di kota yang sama. Tahun
2002 penulis lulus dari SMU Negeri 3 Bukittinggi dan pada tahun yang sama
masuk sebagai mahasiswa ke Fakultas Kedokteran Hewan Institut Pertanian
Bogor. Tahun 2008 penulis menamatkan pendidikan Dokter Hewan dan mulai
aktif sebagai asisten peneliti di Laboratorium Virologi FKH IPB sekaligus asisten
DAFTAR ISTILAH
AI Avian influenza
Ct Cycle threshold
HA Hemaglutinin
HPAI Highly pathogenic avian influenza
LPAI Low pathogenic avian influenza
M1 Protein matriks 1
M2 Protein matriks 2 (ion channel)
MA Matriks
NA Neuraminidase
NEP Nuclear export protein
NLS Nuclear localization signal
NP Nukleoprotein
NS Non-struktural
ORF Open reading frame
PA Polymerase acidic
PB1 Polymerase basic 1
PB2 Polymerase basic 2
PCR Polymerase chain reaction
Rn Normalized reporter
RNA Ribonucleic acid
RNP Ribonukleoprotein
RRT-PCR Real time reverse transcriptase polymerase chain reaction
RT-PCR Reverse transcriptase polymerase chain reaction
SA Sialic acid
Sentinel Hewan yang sengaja ditempatkan untuk merekam satu atau
rangkaian kejadian penyakit dalam program sureveilans prospektif
Halaman
DAFTAR TABEL ... xvi
DAFTAR GAMBAR ... xvii
DAFTAR LAMPIRAN ... xviii
PENDAHULUAN ... 1
Latar Belakang ... 1
Tujuan ... 2
Manfaat ... 3
Hipotesis ... 3
TINJAUAN PUSTAKA ... 4
Virus Influenza A ... 4
Siklus replikasi virus ... 8
Antigenic drift ... 11
Antigenic shift ... 12
Influenza A pada Unggas Liar ... 13
Influenza A pada Unggas Domestik ... 15
Ekologi Itik dan Perannya dalam Penyebaran Influenza A ... 15
Patobiologi Avian Influenza pada Itik ... 19
AI Patogenitas Rendah (low-pathogenic avian influenza, LPAI) ... 20
AI Patogenitas Tinggi (highly pathogenic avian influenza, HPAI) ... 21
Teknik Diagnostik Avian Influenza ... 23
RRT-PCR untuk Deteksi Avian Influenza ... 23
BAHAN DAN METODE ... 31
Waktu dan Tempat ... 31
Bahan dan Alat ... 31
Sampel ... 31
Metode ... 32
Pooling ... 32
Isolasi RNA ... 33
RT-PCR Konvensional ... 33
Real Time RT-PCR ... 34
Analisis Data ... 35
HASIL DAN PEMBAHASAN ... 36
RT-PCR Konvensional dan Real Time ... 36
Influenza A ... 38
Subtipe H5 ... 41
SIMPULAN DAN SARAN ... 45
DAFTAR PUSTAKA ... 46
DAFTAR TABEL
Halaman
Tabel 1. Pasangan primer dan probe untuk deteksi gen tertentu. ... 29
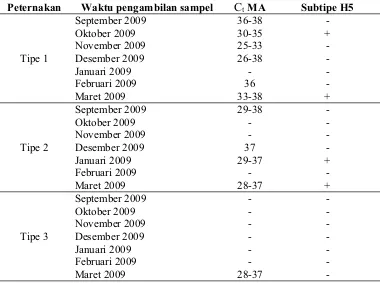
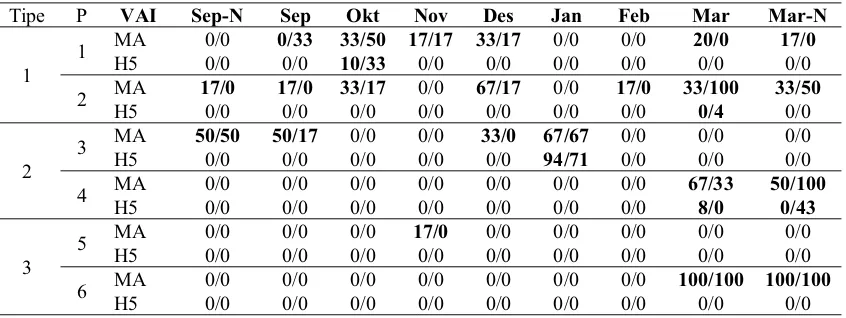
Tabel 2. Virus AI di tiga tipe peternakan itik angon. ... 39
Tabel 3. Persentase sampel usap positif di 6 peternakan itik ... 40
Gambar 1.Diagram skematis struktur virus influenza A ... 4
Gambar 2.Endositosis virus influenza ... 9
Gambar 3.Grafik tingkat kelangsungan hidup itik. ... 20
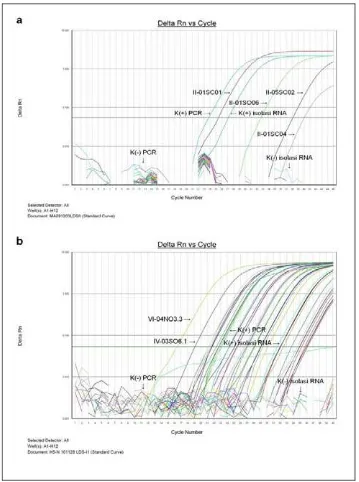
Gambar 5. Perbandingan hasil PCR konvensional dan real time ... 36
Gambar 6. Grafik amplifikasi real time RT-PCR. ... 37
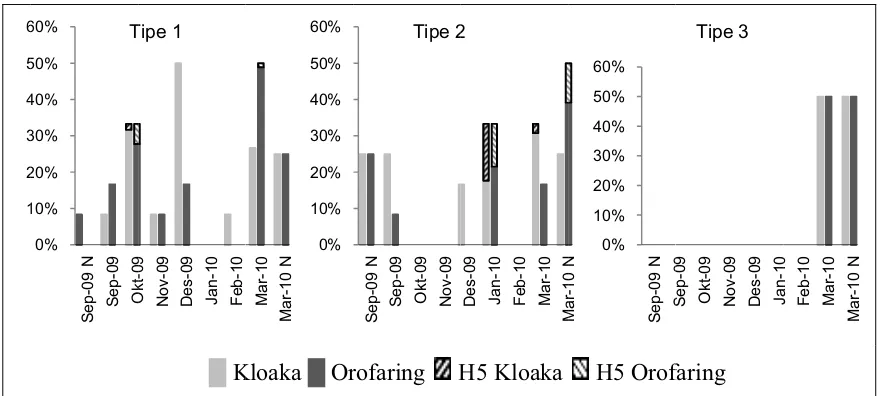
Gambar 7. Distribusi temporal VAI di tiga tipe peternakan itik angon ... 41
DAFTAR LAMPIRAN
Lampiran 1. Tabulasi nilai Ct MA dan H5 di peternakan 1 ... 59
Lampiran 2. Tabulasi nilai Ct MA dan H5 di peternakan 2 ... 60
Lampiran 3. Tabulasi nilai Ct MA dan H5 di peternakan 3 ... 61
Lampiran 4. Tabulasi nilai Ct MA dan H5 di peternakan 4 ... 62
Lampiran 5. Tabulasi nilai Ct MA dan H5 di peternakan 5 ... 63
Latar Belakang
Avian influenza (AI) telah menjadi masalah global maupun nasional yang
mengakibatkan kerugian besar bagi industri perunggasan. Sejak terjadi wabah
highly pathogenic avian influenza (HPAI) pertama pada pada unggas di akhir
tahun 2003, hingga Januari 2012 30 dari 33 Provinsi di Indonesia telah tertular
(OIE 2012). Kasus AI pada manusia di Indonesia mulai terjadi pada bulan Juni
2005, dan hingga bulan Januari 2012 WHO telah mencatat 184 kasus dengan 152
(82,6%) diantaranya mengakibatkan kematian (WHO 2012). Hingga Oktober
2010 kasus AI pada manusia di Indonesia paling tinggi terjadi di Provinsi DKI
Jakarta dan dari 46 kasus konfirmasi, 39 (84,8%) diantaranya meninggal dunia
(Dinkes-Jabar 2010). Kasus AI pada manusia tertinggi kedua terjadi di Provinsi
Jawa Barat yaitu dari 41 kasus konfirmasi, 36 (87,8%) diantaranya meninggal
dunia dan tersebar di 14 Kabupaten/Kota (Dinkes-Jabar 2010).
Unggas air liar merupakan reservoir utama virus influenza A dan dapat
menularkannya ke unggas domestik dan mamalia, termasuk manusia. Dari 16
subtipe HA virus influenza A, hanya subtipe H5 dan H7 yang dapat menyebabkan
highly pathogenic avian influenza (HPAI) pada inang alami. Virus HPAI H5N1
bersifat 100% mematikan bagi ayam dan unggas gallinaceous lainnya sedangkan
unggas air seperti itik dapat mengeluarkan virus melalui saluran pernafasan dan
pencernaan dengan sedikit atau tidak ada gejala penyakit (Brown et al. 2006;
Keawcharoen et al. 2008). Hal ini menunjukkan bahwa VAI yang tidak
menimbulkan gejala penyakit pada itik tetap merupakan ancaman bagi kesehatan
itik sendiri maupun inang lain. Meskipun kejadian wabah HPAI pada pada ayam
jauh lebih tinggi dibandingkan pada itik tetapi penelitian yang dilakukan di
Thailand menunjukkan adanya keterkaitan erat antara wabah HPAI dengan
keberadaan itik domestik maupun angon di negara itu (Gilbert et al. 2006) di
samping faktor-faktor lainnya seperti jumlah ayam, populasi manusia, dan
keadaan topografis (Gilbert et al. 2008). Kegiatan surveilans aktif perlu dilakukan
untuk menentukan peran itik dalam penularan dan asal usul virus. Sangat tidak
2
VAI H5 pada populasi itik karena hasilnya tidak akan akurat, demikian juga
dengan peran itik dalam wabah HPAI.
Di Indonesia beberapa jenis itik telah lama dikembangkan untuk
dimanfaatkan sebagai sumber protein hewani berupa daging maupun telur. Jenis
itik yang berkembang di beberapa daerah antara lain itik alabio di Kalimantan
Selatan, itik tegal di Jawa Tengah, dan itik pitalah di Sumatera Barat (Suswono
2011a; Suswono 2011b; Suswono 2011c). Jawa Barat memiliki populasi itik
paling tinggi di Indonesia yaitu mencapai 4,4 juta ekor pada tahun 2002 dan terus
meningkat hingga 8,2 juta ekor pada tahun 2009, 2 juta ekor diantaranya berada di
Kabupaten Indramayu (Ditjennak 2010) dengan mayoritas pemeliharaan
menerapkan sistem angon sehingga dijadikan sebagai lokasi penelitian mengenai
virus avian influenza (VAI) H5 di peternakan itik angon.
Keberadaan VAI dapat dideteksi menggunakan teknik reverse transcriptase
PCR (RT-PCR). Teknik ini memiliki beberapa keuntungan antara lain dapat
digunakan untuk berbagai jenis sampel, lebih cepat dan lebih ramah lingkungan
dibandingkan isolasi virus pada embrio ayam, dan karena virus tidak aktif sejak
awal pemrosesan, keamanan dan keselamatan biologis juga lebih mudah untuk
dipertahankan (Spackman dan Suarez 2008). Secara konvensional teknik ini
memerlukan waktu beberapa jam dan pembacaan hasilnya melibatkan bahan
kimia yang dapat merugikan pengguna maupun lingkungan. Perkembangan
teknologi telah melahirkan real-time RT-PCR (RRT-PCR) yang mulai banyak
digunakan sejak awal tahun 2000-an dalam rangka pengawasan rutin, selama
wabah, dan untuk penelitian. Beberapa keuntungan RRT-PCR dibandingkan
dengan RT-PCR konvensional antara lain dari segi sensitivitas dan spesifisitas
yang lebih tinggi, memerlukan waktu yang lebih singkat, bersifat kuantitatif,
ramah lingkungan, dan meskipun biaya yang diperlukan untuk investasi peralatan
lebih tinggi namun untuk operasional dan pengamanan lingkungan teknik ini
memerlukan biaya yang lebih sedikit.
Tujuan
Secara umum penelitian ini bertujuan untuk menjawab pertanyaan apakah
Teknik RRT-PCR digunakan untuk mendeteksi persistensi dan penularan VAI H5
dalam peternakan itik angon serta menentukan rute pengeluaran (shedding) virus.
Manfaat
Memberi informasi tentang persistensi dan penularan VAI subtipe H5 di
peternakan itik sebagai pertimbangan dalam pengendalian AI.
Hipotesis
Virus AI H5 menular dan persisten dalam peternakan itik angon dan dapat
TINJAUAN PUSTAKA
Virus Influenza A
Virus influenza merupakan virus RNA untai negatif dengan genom
tersegmentasi berisi tujuh sampai delapan segmen gen yang termasuk kedalam
famili Orthomyxoviridae. Berdasarkan perbedaan sifat antigenik protein matriks
dan nukleoprotein, virus influenza dikelompokkan kedalam tiga tipe yaitu A, B,
dan C yang masing-masing memiliki kecenderungan inang dan patogenisitas
berbeda. Virus influenza A dan B memiliki struktur yang tidak dapat dibedakan
dibawah mikroskop elektron (Bouvier dan Palese 2008) berbeda dengan virus
influenza C. Virus influenza A dan B memiliki delapan segmen gen RNA untai
tunggal, sedangkan virus influenza C memiliki tujuh segmen dan masing-masing
menyandi setidaknya satu protein (Murphy et al. 1999).
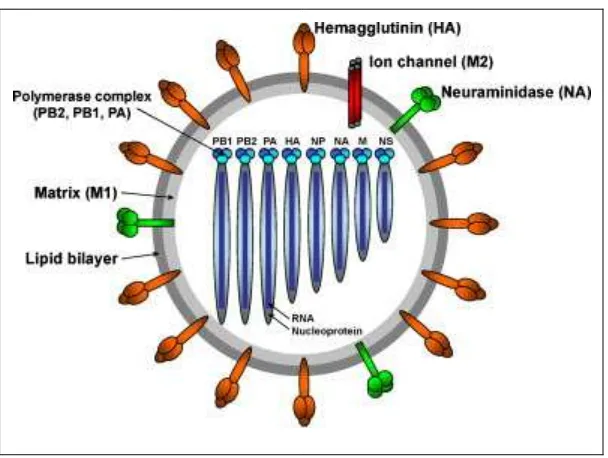
Gambar 1. Diagram skematis struktur virus influenza A (Lee dan Saif 2009)
Virus influenza A memiliki selubung yang berasal dari membran lipid sel
inang. Kedelapan segmen gen menyandi setidaknya 11 open reading frame (ORF)
(Bouvier dan Palese 2008). Permukaan virus diselubungi oleh penonjolan tiga
protein: hemaglutinin (HA), neuraminidase (NA) dan matriks 2 (M2) (Gambar 1).
Protein matriks 1 (M1) terdapat di bawah membran, berinteraksi dengan bagian
(RNP) virus. Protein M1 juga berikatan dengan protein pengeluaran dari inti
(nuclear export protein NEP) yang memperantarai pengeluaran M1-RNP melalui
nukleoporin ke dalam sitoplasma (Bouvier dan Palese 2008). Protein M2 yang
berukuran kecil merupakan ion channeltransmembran dan hanya ditemukan pada
virus influenza A. Protein M2 memiliki bagian luar yang berada di permukaan
selubung virus bersama dengan HA dan NA. Protein M2 merupakan target obat
anti influenza dari kelas amantadine yang memblokir aktivitas ion channel dan
mencegah pelepasan selubung virus (Pinto et al. 1992; Wharton et al. 1994; Sheu
et al. 2011). Selain itu, M2 merupakan protein permukaan sehingga dijadikan
sebagai komponen vaksin (Slepushkin et al. 1995; Neirynck et al. 1999).
Hemaglutinin merupakan protein membran integral tipe I terglikosilasi yang
berfungsi sebagai protein pengikat reseptor dan protein fusi serta merupakan
target utama netralisasi oleh antibodi inang (Cross et al. 2001; Hulse et al. 2004;
Hoffmann et al. 2005; Gambaryan et al. 2006). Protein ini dapat mengenali asam
sialat (N-acetyl neuraminic acid) yang terikat pada gula di ujung glikoprotein sel
inang. Virus influenza A memiliki berbagai HA spesifik dengan isomerisasi
ikatan glikosidik berbeda untuk disakarida yang terdiri atas sialic acid (SA) dan
galaktosa atau N-asetilgalaktosamin (GalNAc). Reseptor HA pada unggas
memiliki spesifisitas ikatan terhadap SA Į2,3 sel bersilia, sementara HA pada
manusia memiliki spesifisitas ikatan yang lebih tinggi terhadap SA Į2,6 sel tidak
bersilia (Matrosovich et al. 2004). Struktur kristal molekul HA berbentuk trimer
dengan dua regio struktural berbeda yaitu bagian batang dan kepala (Wilson et al.
1981). Bagian kepala mengandung reseptor situs pengikatan SA yang dikelilingi
oleh determinan antigenik variabel yang disebut A, B, C, dan D pada subtipe H3
(Shortridge et al. 1990) dan Sa, Sb, CA1, Ca2, dan Cb pada subtipe H1 (Palese
dan Shaw 2007). Protein HA memiliki bentuk trimer yang masing-masing
monomernya mengalami pembelahan proteolitik untuk menghasilkan rantai
polipeptida HA1 dan HA2 dengan ikatan disulfida sebelum aktivasi. Polipeptida
HA2 memperantarai fusi selubung virus dengan membran sel, sedangkan HA1
mengandung situs antigenik dan pengikatan reseptor (Steinhauer 1999).
Pembelahan HA memerlukan protease serin eksogen (enzim yang menyerupai
6
aktivasi (Chen et al. 1998). Pada manusia dan mamalia lain, enzim ini berupa
triptase Clara yang diproduksi oleh sel epitel bronkiolus (Murakami et al. 2001).
Aktivasi pembelahan HA dalam sel usus dan/atau pernafasan unggas
kemungkinan juga memerlukan protease serupa. Situs pengenalan protease dapat
berubah menjadi urutan menyerupai furin R-X-R/K-R pada subtipe H5 dan H7
bila mengalami mutasi insersional pada situs pembelahan HA. Perubahan situs
pembelahan HA menjadi polibasa ini memperluas spesifisitas protease sehingga
memungkinkan aktivasi pembelahan intraseluler dan replikasi virus secara
sistemik pada unggas yang mengakibatkan influenza unggas sangat patogen
(highly pathogenic avian influenza, HPAI) (Werner 2006). Akumulasi perubahan
yang relatif kecil pada situs antigenik HA yang dikenali oleh antibodi disebut
antigenic drift yang menghasilkan strain virus yang tidak lagi dapat dinetralisir
oleh antibodi sehingga inang menjadi rentan terhadap infeksi kembali oleh strain
yang mengalami drift.
Neuraminidase (NA) merupakan tetramer berbentuk seperti jamur yang
menancap pada selubung virus melalui domain transmembran (Colman et al.
1983; Varghese et al. 1983). Sebagai glikoprotein membran integral tipe II dengan
aktivitas enzimatik sialidase (neuraminidase), NA diperlukan untuk pembelahan
SA sel inang yang memungkinkan pelepasan virion baru dan melepaskan SA dari
glikoprotein virus untuk mencegah agregasi partikel progeni virus (Palese et al.
1974). Hemaglutinin dan NA merupakan target antigenik utama respon imun
humoral terhadap virus influenza A dengan NA menjadi target obat antivirus
oseltamivir dan zanamivir (De Clercq 2006).
Setiap segmen RNA virus influenza A diselubungi oleh nukleoprotein (NP).
Pada virion, RNA virus melilit monomer NP dan membentuk RNP bersama-sama
dengan tiga protein polimerase yaitu: polymerase acidic protein (PA), polymerase
basic protein 1 (PB1) dan polymerase basic protein 2 (PB2) (Coloma et al. 2009).
NP berperan terutama sebagai protein pengikat RNA untai tunggal dan berfungsi
sebagai protein struktural pada RNP. Selain itu, NP berperan penting dalam
transkripsi dan perpindahan RNP antara sitoplasma dan nukleus. Transkripsi RNA
virus influenza A dan replikasi terjadi di dalam inti inang karena virus ini
Sintesis RNA virus influenza A memerlukan polimerase yang terdiri atas
tiga subunit PA, PB1, dan PB2. Kompleks heterotrimer polimerase terbentuk
melalui interaksi PA dengan PB1 dan PB1 dengan PB2. Protein PA berperan
penting dalam penempelan, katalisis, dan lokalisasi inti oleh polimerase (Guu et
al. 2008). Protein PB1 berfungsi sebagai RNA polimerase sedangkan PB2
berperan dalam sintesis mRNA melalui pengikatan bagian kepala mRNA inang.
Protein non struktural kecil lainnya yaitu PB1-F2 secara bervariasi disandi oleh
gen PB1 melalui bingkai bacaan (reading frame) alternatif. Protein ini menjadikan
membran dalam mitokondria sebagai target dan mungkin berperan dalam
apoptosis selama infeksi virus influenza A selain memiliki aktivitas antagosnisme
interferon (Dudek et al. 2011). Gen PB1 juga menyandi polipeptida ketiga yang
diekspresikan melalui penggunaan kodon AUG diferensial yang disebut N40
(Wise et al. 2009).
Protein non-struktural 1 (NS1) memiliki beberapa domain fungsional antara
lain: domain N-terminal pengikat RNA (residu 1-73) yang pada in vitro mengikat
beberapa spesies RNA dengan afinitas rendah dan memiliki sinyal lokalisasi inti
(nuclear localization signal, NLS) (Hatada dan Fukuda 1992; Qian et al. 1995;
Chien et al. 2004), dan domain C-terminal 'efektor' (residu 74-230) yang
memperantarai interaksi dengan protein sel inang dan secara fungsional
menstabilkan domain pengikat RNA (Wang et al. 2002). Keseluruhan NS1
merupakan homodimer dengan domain pengikat RNA dan domain efektor
berkontribusi terhadap multimerisasi (Nemeroff et al. 1995). NS1 memiliki fungsi
pleiotropik, antara lain pengikatan dsRNA, peningkatan translasi mRNA virus,
penghambatan proses mRNA inang dan antagonisme interferon tipe I (Palese dan
Shaw 2007). Protein NS2 (disebut juga protein ekspor inti, NEP) ditemukan
dalam virion dan memfasilitasi pengeluaran kompleks RNP virus dari dalam inti
(O'Neill et al. 1998).
Berdasarkan karakterisasi antigen glikoprotein permukaan HA dan NA virus
influenza A dikelompokkan kedalam 16 subtipe HA dan 9 NA (Fouchier et al.
2005). Secara teoritis kombinasi HA-NA dapat membentuk 144 subtipe, dan
setidaknya 116 kombinasi subtipe ini telah diisolasi dari unggas (Krauss et al.
8
nomenklatur virus influenza adalah sebagai berikut: pertama, tipe virus (A, B,
atau C), kemudian inang (jika bukan manusia), tempat isolasi, nomor isolasi dan
tahun isolasi (dipisahkan dengan garis miring). Untuk virus influenza A, subtipe
HA (H1-H16) dan NA (N1-9) ditulis dalam tanda kurung. Sebagai contoh, strain
yang termasuk dalam vaksin trivalen influenza manusia untuk musim 2010-2011
di Amerika Serikat adalah: A/California/7/2009 (H1N1), A/Perth/16/2009 (H3N2)
dan B/Brisbane/60/2008.
Virus influenza tipe B dan C menginfeksi dan hampir selalu diisolasi dari
manusia meskipun virus influenza B pernah diisolasi dari anjing laut dan virus
influenza C pernah diisolasi dari babi dan anjing (Wright et al. 2007). Sebaliknya,
virus influenza A dapat menginfeksi berbagai hewan berdarah panas seperti
unggas, babi, kuda dan manusia. Virus AI yang menjadi penyebab flu burung/AI
termasuk kedalam virus influenza A dengan unggas air sebagai reservoir alami
untuk semua subtipenya (Webster et al. 1992). Tiga sifat penting yang membuat
virus influenza mudah beradaptasi, mampu menghindari respon kekebalan inang,
dan mampu menginfeksi spesies inang baru (Webster et al. 1992; Bahl et al.
2009) yaitu: pertama, enzim polimerase yang mengkatalisis replikasi RNA dari
cetakan RNA mudah melakukan kesalahan; kedua, kurangnya koreksi kesalahan
selama replikasi; dan ketiga, struktur genom virus influenza memungkinkan untuk
pertukaran segmen antar virus-virus yang menginfeksi sel di waktu bersamaan
melalui proses yang disebut reassortment.
Siklus replikasi virus
Virus influenza mengenali SA (N-asetilneuraminik) pada permukaan sel
inang. Monosakarida asam sembilan karbon yang dapat ditemukan pada ujung
berbagai glikokonjugat ini terdapat di banyak tempat pada berbagai tipe sel dan
spesies hewan. Karbon 2 SA dapat mengikat karbon 3 atau 6 galaktosa
membentuk ikatan Į2,3 atau Į2,6. Perbedaan ikatan ini menghasilkan konfigurasi
sterik yang unik pada SA. Bagian SA dapat dikenali oleh dan berikatan dengan
HA pada permukaan virus influenza yang memiliki spesifisitas ikatan Į2,3 atau
Į2,6. Pada sel epitel trakea manusia lebih dominan reseptor Į2,6 sedangkan
juga terdapat pada epitel saluran pernafasan manusia meskipun jumlahnya lebih
sedikit dibandingkan Į2,6 (Couceiro et al. 1993; Matrosovich et al. 2004)
sehingga manusia dan primata lain juga dapat terinfeksi oleh VAI meskipun
dengan efisiensi yang lebih rendah dibandingkan infeksi oleh strain manusia (Tian
et al. 1985; Beare dan Webster 1991). Perbedaan ekspresi SA pada saluran
pernafasan mamalia membantu menjelaskan infektivitas rendah tetapi
patogenisitas tinggi pada beberapa strain VAI. Pada manusia protein SA dengan
ikatan Į2 ,3 dalam jumlah sedikit terdapat di saluran pernafasan bawah seperti
bronkiolus dan alveoli. Akses partikel virus dari udara ke paru-paru tidak
semudah virus mencapai saluran pernafasan bagian atas seperti nasofaring, sinus
paranasal, trakea, dan bronkus, sehingga infeksi VAI relatif jarang terjadi pada
manusia. Namun ketika strain VAI menginfeksi paru-paru manusia, pneumonia
berat dan progresif dapat terjadi dengan angka kematian melebihi 60% (Gambotto
et al. 2008).
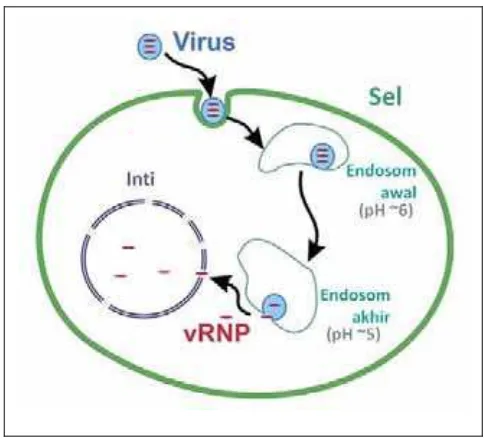
Gambar 2. Endositosis virus influenza diadaptasi dari Lakadamyali et al. (2004)
Setelah protein HA virus influenza (atau protein HEF virus influenza C)
menempel pada SA, virus mengalami endositosis. Keasaman kompartemen
endosomal sangat penting untuk pelepasan selubung virus influenza (Gambar 2).
Rendahnya pH memicu perubahan konformasi HA, memaparkan peptida fusi
yang menjadi mediator penggabungan selubung virus dengan membran
10
inang (Stegmann 2000; Sieczkarski dan Whittaker 2005). Ion hidrogen dari
endosom dipompa ke dalam partikel virus melalui ion channel M2. Pengasaman
internal virion influenza melalui channel M2 mengganggu interaksi
protein-protein internal sehingga RNP dapat dilepaskan keluar dari matriks virus ke dalam
sitoplasma sel (Martin dan Helenius 1991).
Setelah keluar dari virion, RNP masuk kedalam inti sel inang dengan
memanfaatkan sinyal lokalisasi inti (NLS) oleh protein virus (NS1) yang
memerintahkan protein sel untuk memasukkan RNP dan protein virus lainnya ke
dalam inti sel inang (Cros dan Palese 2003). Inti merupakan tempat dimana semua
sintesis RNA virus terjadi, tempat RNA poliadenilasi (mRNA) yang bertindak
sebagai cetakan bagi sel inang untuk translasi, dan tempat segmen RNA virus
yang membentuk genom progeni virus. Polimerase RNA yang merupakan
komponen RNP juga masuk kedalam inti dan menggunakan RNA virus untai
negatif sebagai cetakan untuk mensintesis dua RNA untai positif, yaitu cetakan
mRNA untuk sintesis protein virus, dan RNA komplementer (cRNA) untuk
membentuk lebih banyak RNA virus untai negatif penyusun genom (Bouvier dan
Palese 2008).
Berbeda dengan mRNA sel inang yang terpoliadenilasi oleh poli (A)
polimerase spesifik, ujung penutup poli (A) mRNA virus influenza disandi dalam
bentuk RNA virus untai negatif dengan lima sampai tujuh residu urasil yang
ditranskripsikan oleh polimerase virus menjadi untai positif dengan adenosin
membentuk ekor poli (A) (Robertson et al. 1981; Li dan Palese 1994).
Pembentukan ujung penutup RNA messenger juga terjadi dengan cara unik yang
sama, di mana protein PB1 dan PB2 "mencuri" primer berujung penutup 5' dari
transkrip pre-mRNA inang untuk memulai sintesis mRNA virus, proses ini
disebut "cap snatching" (Krug 1981). Setelah terpoliadenilasi dan ujungnya
ditutup, mRNA asal virus dapat keluar dan diterjemahkan seperti mRNA inang.
Pengeluaran segmen RNA virus dari inti diperantarai oleh protein M1 dan
NEP/NS2 virus (Cros dan Palese 2003).
Protein selubung HA, NA, dan M2 disintesis dari mRNA asal virus di
kedalam aparatus Golgi untuk modifikasi pasca-translasi. Ketiga protein tersebut
memiliki sinyal penyusun apikal yang kemudian mengarahkan mereka ke
membran sel untuk perakitan virion. Meskipun relatif sedikit yang diketahui
tentang translasi dan penyortiran protein yang bukan bagian dari selubung, M1
diperkirakan berperan dalam membawa kompleks RNP-NEP berkontak dengan
protein selubung HA, NA, dan M2 untuk dikemas di membran sel inang (Palese
dan Shaw 2007).
Virus influenza tidak sepenuhnya menular kecuali virion yang lengkap
berisi genom delapan segmen, atau tujuh segmen untuk virus influenza C.
Sebelumnya pengemasan RNA virus dianggap sebagai sebuah proses yang
sepenuhnya acak, di mana segmen RNA virus secara tidak beraturan dimasukkan
ke dalam tunas partikel virus dan hanya yang memiliki genom lengkap yang dapat
menular. Namun bukti baru menunjukkan bahwa pengemasan merupakan proses
selektif di mana sinyal pengemasan pada semua segmen RNA virus memastikan
bahwa genom lengkap dimasukkan ke dalam setiap partikel virus (Bancroft dan
Parslow 2002; Fujii et al. 2003).
Pertunasan (budding) virus influenza terjadi di membran sel yang dimulai
dengan akumulasi protein matriks M1 di sisi sitoplasma dari lipid bilayer. Ketika
budding selesai, tonjolan HA tetap menempelkan virion pada SA di permukaan
sel hingga partikel virus secara aktif dilepaskan oleh aktivitas sialidase protein NA
(Colman et al. 1983; Varghese et al. 1983).
Antigenic drift
Virus influenza A terus berevolusi dengan tingkat mutasi tinggi yang
berkisar antara 1×10-3 sampai 8×10-3 substitusi/situs/tahun (Chen dan Holmes 2006). Mutasi selektif pada domain antigenik yang terjadi secara bertahap dalam
satu strain dan menghindarkan virus dari sistem kekebalan disebut antigenic drift
(Rambaut et al. 2008). Bagian HA1 dari gen HA mengalami evolusi dengan
tingkat mutasi 5,7 substitusi nukleotida/tahun atau 5,7×10í3 substitusi/situs/tahun (Fitch et al. 1997). Dengan antibodi terhadap protein HA mencegah pengikatan
reseptor, menetralisir, dan mencegah infeksi ulang oleh subtipe yang sama
12
glikoprotein permukaan seperti HA dapat menguntungkan virus karena
memungkinkan virus menghindar dari sistem kekebalan. Proses replikasi virus
sangat rawan mutasi karena enzim polimerase yang mengkatalisis replikasi RNA
dari cetakan RNA mudah melakukan kesalahan disertai kurangnya koreksi
kesalahan selama replikasi. Hal ini menjadi penyebab terjadinya antigenic drift.
Antigenic drift merupakan salah satu strategi virus influenza untuk
menghindar dari sistem kekebalan inang yang meningkat karena vaksinasi.
Perubahan antigenik yang terjadi di daerah epitop merupakan hambatan untuk
pengembangan vaksin karena vaksinasi yang efektif hanya dapat terjadi bila strain
epidemik sesuai dengan strain vaksin (Stohr 2002). Gen HA sebagai target
netralisasi antibodi menjadi contoh klasik protein antigen yang mengalami mutasi
titik yang menumpuk pada epitop atau daerah yang dikenali antibodi (Webster et
al. 1982; Wilson dan Cox 1990). Antigenic drift pada gen HA dapat dipercepat
oleh vaksinasi (Lee et al. 2004) yang suboptimal karena tekanan oleh kekebalan
hasil imunisasi terhadap virus yang sebelumnya bereplikasi dan beredar antar dan
intra spesies (Abdelwhab dan Hafez 2011) memaksa virus untuk beradaptasi.
Virus H5N1 dapat bermutasi secara intensif pada unggas yang divaksinasi
sehingga berpotensi menimbulkan pandemi. Gen HA dari 4 strain H5N1 yang
beredar di Mesir mengalami perubahan asam amino pada epitop HA sehingga
berbeda dengan VAI H5N1 awal yang ditemukan sejak program vaksinasi dimulai
pada tahun 2006 yang berdampak pada virulensi H5N1 pada mamalia
(Abdel-Moneim et al. 2011). Contoh lain virus influenza A yang mengalami antigenic
drift adalah virus pandemi H1N1 2009 (pH1N1 2009) yang berasal dari babi.
Residu 227 HA pada H1N1 babi yang berupa asam amino alanin mengalami
perubahan menjadi asam glutamat sehingga mampu menular dan menimbulkan
pandemi pada manusia (van Doremalen et al. 2011).
Antigenic shift
Genom virus influenza A terdiri dari 8 segmen RNA sehingga ko-infeksi
satu sel inang dengan dua virus influenza A berbeda dapat menghasilkan progeni
virus yang berisi segmen gen dari kedua virus. Proses penyusunan (reassortment)
penting dalam evolusi virus influenza A (Holmes et al. 2005; Dugan et al. 2008)
dan adaptasi inang (Garten et al. 2009, Scholtissek et al. 1978). Secara teori dapat
terjadi 256 (28) kombinasi 8 segmen gen hasil reassortment antara dua virus
dalam satu inang. Rekombinasi homolog jarang terjadi pada virus RNA negatif
seperti virus influenza A (Boni et al. 2008) tetapi rekombinasi dengan pertukaran
segmen gen diketahui berperan dalam perubahan virulensi dan adaptasi inang
(Wright et al. 2007).
Influenza A pada Unggas Liar
Virus influenza A memiliki keragaman genetik dan antigenik yang tinggi
dan tersebar pada berbagai spesies unggas liar di seluruh dunia. Penularan virus
influenza A pada unggas air liar terjadi melalui rute fekal-oral dan menginfeksi
sel-sel epitel saluran pencernaan dengan sedikit atau tanpa gejala penyakit. Virus
bertahan melalui infeksi asimtomatik (low pathogenic, LPAI) pada unggas air dari
ordoAnseriformes seperti itik dan angsa, ordo Charadriiformes seperti camar dan
burung laut, serta ordo Passeriformes dan setidaknya 105 spesies unggas liar telah
teridentifikasi membawa virus influenza A (Munster et al. 2007). Distribusi
subtipe HA dan NA virus pada isolat unggas liar tidak merata. Sebagian besar
subtipe HA dapat ditemukan pada Anseriformes sedangkan subtipe H13 dan H16
ditemukan pada Charadriiformes (Munster et al. 2007).
Pola umum keragaman VAI pada unggas liar dapat dijelaskan dengan dua
model evolusi yaitu spesiasi alopatrik (cekaman geografis) dan simpatrik
(cekaman selektif) (Dugan et al. 2008). Analisis filogenetik menunjukkan bahwa
semua subtipe HA VAI memiliki nenek moyang yang sama namun subtipe HA
tidak berasal dari radiasi tunggal. Hal ini dapat dilihat dari tingginya keragaman
genetik antar subtipe HA sedangkan dalam subtipe HA yang sama keragaman
genetik cukup rendah. Pola ini juga terjadi pada evolusi kesembilan subtipe NA.
Analisis menunjukkan keragaman yang mencerminkan bahwa nenek moyang
bersama terdekat (the most recent common ancestors TMRCA) subtipe HA yang
berbeda pernah ada dalam rentang waktu beberapa ratus tahun yang lalu (Chen
dan Holmes 2010). Segmen gen NS VAI pada unggas memiliki perbedaan jelas
14
keseimbangan (Dugan et al. 2008). Keragaman genetik yang lebih rendah dimiliki
oleh lima segmen gen VAI lainnya (PB2, PB1, PA, NP dan M). Analisis
filogenetik juga menunjukkan perbedaan urutan asam inti yang jelas antara VAI
yang berasal dari unggas di belahan dunia timur dan barat, sesuai dengan evolusi
cekaman alopatrik (Dugan et al. 2008; Munster dan Fouchier 2009).
Banyaknya kombinasi HA-NA yang ditemukan pada unggas liar
menunjukkan bahwa infeksi campuran dan reassortment VAI sering terjadi pada
unggas liar (Wang et al. 2008) dan bahwa subtipe HA-NA memiliki kombinasi
spesifik yang rendah. Keragaman genetik yang tinggi pada HA, NA dan NS
bertolak belakang dengan 5 segmen gen penyandi protein internal yang memiliki
stabilitas tinggi di tingkat asam amino. Hal ini menandakan bahwa kelima segmen
gen tersebut telah melalui alur seleksi pemurnian. Kecocokan kelima gen tersebut
untuk saling terkait dalam genom ditentukan oleh viabilitas fungsional, dengan
sedikit cekaman selektif untuk mempertahankan mutasi yang menguntungkan.
Urutan asam amino yang sangat stabil menunjukkan bahwa reassortment terjadi
antara segmen-segmen yang secara fungsional setara. Dugan et al. berhipotesis
bahwa VAI pada unggas liar berperan sebagai kolam (pool) besar yang berisi
segmen-segmen gen yang memiliki kesetaraan fungsional sehingga dapat saling
tukar membentuk konstelasi genom sementara tanpa ada cekaman selektif yang
kuat agar tetap bertahan sebagai genom (Dugan et al. 2008).
Virus influenza A pada unggas liar dapat berpindah ke inang yang baru
seperti ayam, kuda, babi, bahkan manusia dan tetap stabil sehingga dapat menjadi
virus menular di kelompok inang yang baru. Virus influenza A sering beradaptasi
terhadap inang yang berasal dari spesies unggas domestik (Wright et al. 2007).
Kemampuan virus untuk tetap stabil setelah berganti inang memerlukan akuisisi
sejumlah mutasi, tergantung pada virus dan spesies inang yang memisahkan
individu virus dari pool gen virus influenza A di unggas liar. Adaptasi terhadap
inang baru ini dapat mengurangi kemampuan virus untuk kembali ke pool gen
virus influenza A pada unggas liar (Swayne 2007) sehingga ia harus membangun
konstelasi genom delapan segmen yang berbeda dari klonnya di unggas liar
Influenza A pada Unggas Domestik
Unggas domestik dari ordo Galliformes seperti kalkun, ayam, dan burung
puyuh bukan merupakan reservoir virus influenza A unggas namun rentan
terhadap infeksi oleh virus influenza A dari unggas liar yang telah beradaptasi.
Virus influenza A yang telah beradaptasi pada Galliformes jarang kembali dan
beredar di unggas liar (Swayne 2007) kecuali virus HPAI H5N1 Eurasia yang
baru-baru ini diisolasi dari populasi unggas liar di Eropa dan Asia. Virus
panzootik HPAI H5N1 galur Asia memiliki keunikan (Webster et al. 2007) yang
dapat mengakibatkan kematian jutaan unggas di 64 negara di tiga benua. Adaptasi
virus influenza A pada inang Galliformes secara molekuler belum sepenuhnya
dapat dijelaskan namun diketahui melibatkan seleksi positif mutasi HA, NA
(Perez et al. 2003; Campitelli et al. 2004), dan protein RNP (Wasilenko et al.
2008).
Virus influenza A yang diisolasi dari unggas domestik umumnya
mempertahankan spesifisitas pengikatan reseptor HA Į2,3-SA (Wright et al.
2007). Ciri lainnya yaitu penghapusan in-frame sekitar 20 asam amino di daerah
batang NA yang mengurangi aktivitas enzimatik NA (Baigent dan McCauley
2001) sebagai kompensasi terhadap penurunan aktivitas pengikatan reseptor HA
virus influenza A dari unggas liar yang beradaptasi untuk bereplikasi di saluran
pernafasan unggas domestik (Matrosovich et al. 1999). Strain virus influenza A
H5 atau H7 yang beradaptasi pada unggas domestik berkembang menjadi HPAI
melalui akuisisi mutasi insersi yang mengakibatkan situs pembelahan asam amino
polibasa pada HA (Wright et al. 2007).
Ekologi Itik dan Perannya dalam Penyebaran Influenza A
Itik adalah anggota subfamili Anatinae yang menaungi spesies unggas air
Anseriformes. Subfamili ini tersebar di seluruh dunia dan menempati hampir
semua habitat perairan. Ekologi unggas ini memungkinkan pemeliharaan dan
penyebaran VAI.
Replikasi VAI terjadi di saluran pernafasan (Webster et al. 1978) tetapi
situs utama infeksi VAI pada itik adalah usus (Webster et al. 1978) meskipun
16
sering menginfeksi saluran pernafasan bagian atas. Virus LPAI dalam populasi
itik ditularkan melalui rute fekal-oral (Webster et al. 1992) yang dicirikan oleh
tingginya jumlah usap kloaka positif dibandingkan trakea dan titer virus yang
tinggi pada kotoran serta didukung oleh stabilitas virion dalam air meskipun
penularan melalui aerosol tidak dapat diabaikan. Itik yang diinfeksi secara
eksperimental mengeluarkan virion H4N7, H7N3, dan H11N9 dalam waktu lebih
lama dan titer lebih tinggi melalui feses dibandingkan melalui trakea (Webster et
al. 1978). Virus AI memasuki lingkungan ketika inang defekasi atau
mengeluarkan leleran kemudian menginfeksi inang yang rentan melaui proses
makan dan minum. Ketika segerombolan itik berenang di kolam kecil,
diperkirakan sebanyak 1010 EID50/g/hari virion ditularkan ke lingkungan melalui
kotoran masing-masing itik yang terinfeksi (Webster et al. 1978) dan VAI relatif
stabil dalam air (Stallknecht et al. 1990; Webster et al. 1992). Keadaan ini
menjelaskan mengapa prevalensi infeksi pada itik yang makan di permukaan lebih
tinggi dibandingkan itik yang mencari makan di air yang lebih dalam (Olsen et al.
2006).
Data surveilans menunjukkan bahwa penularan VAI dalam populasi itik
terjadi sepanjang tahun. Prevalensi infeksi menunjukkan pola siklus tahunan pada
populasi itik di Amerika Utara (Olsen et al. 2006) (Krauss et al. 2004) dan
Eurasia (Munster et al. 2007) yang memuncak sebelum dan selama migrasi
musim gugur sebagai akibat dari masuknya itik remaja yang secara imunologis
naif kedalam populasi (Hinshaw et al. 1985; Webster et al. 1992; Olsen et al.
2006). Itik Pekin putih yang diinfeksi secara ekperimental mengeluarkan virus
selama lebih dari tiga minggu setelah inokulasi (Kida et al. 1980). Itik yang
terinfeksi mengeluarkan virus selama beberapa minggu pertama migrasi musim
gugur, menebarkan virus di sepanjang koridor migrasi dengan morbiditas dan
respon antibodi serum yang rendah (Kida et al. 1980). Meskipun demikian,
prevalensi infeksi jauh lebih rendah di sepanjang rute migrasi dan di lokasi
migrasi musim dingin dibandingkan di tempat itik istirahat dan mencari makan
(Okazaki et al. 2000; Munster et al. 2007; Wallensten et al. 2007). Perbedaan ini
beredar dalam populasi itik atau penurunan transmisi karena penyebaran populasi
(Hinshaw et al. 1985).
Secara umum prevalensi infeksi di tempat migrasi musim dingin dan di
tempat bersarang musim semi lebih tinggi pada populasi itik Eropa dibandingkan
populasi itik Amerika Utara. Penjelasan yang paling mungkin untuk perbedaan ini
adalah variasi acak, karena penelitian surveilans pada populasi itik di beberapa
daerah di Amerika Utara dan Eropa sering memperoleh nilai prevalensi yang
sedikit berbeda. Banyak faktor dapat mempengaruhi prevalensi termasuk ukuran
populasi itik, lokasi pengambilan sampel, waktu pengambilan sampel, dan
lain-lain.
Prevalensi infeksi paling rendah terjadi selama migrasi musim semi namun
kembali meningkat setelah musim kawin ketika itik pindah ke tempat bersarang
musim panas (Hinshaw et al. 1985; Krauss et al. 2004; Wallensten et al. 2007).
Tidak jelas bagaimana populasi itik memperoleh virus AI selama musim semi
setiap tahun. Ada dua kemungkinan bahwa itik mungkin membawa virus terus
menerus selama migrasi yang ditunjukkan oleh prevalensi pada itik yang terus ada
sepanjang tahun, meskipun daya tahan virus di habitat beku juga dapat berperan
dalam kelangsungan hidup virus (Olsen et al. 2006) karena virion infektif
mungkin dapat bertahan di dalam air beku melewati musin dingin di tempat itik
berkembang biak dan menginfeksi saat itik tersebut kembali pada musim semi
(Webster et al. 1978; Webster et al. 1992).
Beberapa subtipe virus lebih sering ditemukan daripada yang lain (Krauss et
al. 2004; Olsen et al. 2006). Tiga subtipe HA yaitu H3, H4, dan H6 paling sering
ditemukan pada itik di Amerika Utara dan Eropa (Krauss et al. 2004; Munster et
al. 2007) dengan kombinasi subtipe yang paling umum yaitu H4N6 dan H6N2
(Wallensten et al. 2007). Banyak penjelasan mengapa subtipe HA dan NA tertentu
dan kombinasi keduanya sering atau jarang ditemukan pada unggas liar. Hipotesis
umum adalah bahwa subtipe tertentu memiliki kecocokan tertinggi, dengan
tingkat replikasi dan virulensi seimbang yang cukup untuk meningkatkan
kemungkinan keberhasilan transmisi. Hal ini diyakini sangat dipengaruhi oleh
18
(Wagner et al. 2002). Meskipun gen H6 berasal dari Eurasia dan secara luas
tersebar pada itik di Amerika Utara, analisis genom virus menunjukkan bahwa
pertukaran gen antar benua antara Eurasia dan Amerika sangat terbatas (Krauss et
al. 2007). Oleh karena itu kemunculan genotipe virus baru harus melalui mutasi
dan reassortment genom-genom yang bersirkulasi dalam wilayah geografis
tertentu. Kesempatan untuk mutasi dan reassortment ini terbuka lebar di daerah
tempat itik istirahat dan mencari makan karena populasi itik dari berbagai tempat
dan koridor migrasi berbeda datang dengan membawa kombinasi subtipe
masing-masing (Wallensten et al. 2007). Koinfeksi itik dengan dua atau lebih subtipe
virus sering terjadi (Sharp et al. 1997) sama seperti reassortment memunculkan
virus yang sangat virulen pada unggas Galliformes namun memiliki patogenisitas
rendah pada inang itik (Sturm-Ramirez et al. 2005).
Peran itik dalam pemeliharaan dan penyebaran virus influenza, dan terutama
dalam pemunculan genotipe baru tergantung pada perilaku migrasi. Itik yang
bermigrasi setiap tahun cenderung menyebarkan virus influenza di sepanjang rute
migrasi terutama pada populasi itik domestik dan peliharaan di berbagai lokasi
persinggahan (Olsen et al. 2006; Wallensten et al. 2007). Selanjutnya itik
domestik membawa virus berdekatan dengan spesies lain dan berperan dalam
penyebaran LPAI dan HPAI pada unggas domestik dan unggas darat lainnya
(Hulse-Post et al. 2005; Sturm-Ramirez et al. 2005; Gilbert et al. 2006).
Bebek domestik dan itik angon telah dikaitkan dengan penyebaran virus
HPAI H5N1 di Asia Tenggara (Gilbert et al. 2006). Itik sebagai salah satu unggas
air domestik dianggap sebagai sumber penularan virus H5N1 pada wabah di Cina
tahun 1999-2002 (Chen et al. 2004; Li et al. 2004) dan Hongkong tahun 2001
(Sturm-Ramirez et al. 2005). Penelitian seroprevalensi AI pada unggas air (itik,
entog, dan angsa) secara signifikan lebih tinggi dibandingkan seroprevalensi pada
ayam kampung. Hal ini terlihat pada pemeriksaan serologis itik di daerah Jawa
Barat (BALITVET 2006). Pemeriksaan serologis yang dilakukan oleh Balitvet
pada bulan Oktober 2006 menunjukkan sejumlah unggas yang seropositif
terhadap H5 VAI dengan prevalensi pada ayam 22,96% (n=591), itik 41,74%
(n=43), entog 27,04% (n=43), dan angsa 75,0% (n=12). Hasil pemeriksaan
seropositif terhadap H5 VAI masing-masing pada ayam 33,37% (n=1.038), itik
43,44% (n=63), entog 28,21% (n=68), dan angsa 42,3% (n=11) (BALITVET
2006).
Patobiologi Avian Influenza pada Itik
Wabah AI pertama kali dilaporkan tahun 1878 terjadi pada ayam dan
burung di Italia yang saat itu disebut penyakit Lombardia kemudian pada tahun
1901 Centanini dan Savonucci dapat mengidentifikasi organisme berukuran mikro
yang menyebabkan wabah tersebut namun baru pada tahun 1955 Schafer dapat
menunjukkan ciri-ciri organisme itu sebagai virus influenza A (Werner 2006).
Virus influenza A biasanya tidak patogenik terhadap reservoir alaminya yaitu itik
dan unggas air lain. Namun virus HPAI telah berevolusi dari yang tidak
mengakibatkan atau sedikit menimbulkan gejala infeksi pada saluran pernafasan
itik menjadi virus yang menyebabkan penyakit sistemik parah dan kematian
(Pantin-Jackwood dan Swayne 2007). Wabah virus highly pathogenic avian
influenza (HPAI) H5N1 pertama kali dilaporkan di Cina Selatan pada tahun
1996-1997, kemudian menyebar dan menyebabkan kematian unggas di Vietnam,
Thailand, Indonesia dan Negara Asia Timur sejak awal tahun 2004 (Smith et al.
2006).
Gejala HPAI H5N1 sebelum tahun 2002 tidak terlihat pada itik. Penelitian
eksperimental yang dilakukan oleh (Perkins dan Swayne 2002) yang
menginfeksikan A/chicken/HK/220/97 pada itik tidak menemukan gejala klinis.
Sedangkan infeksi virus HPAI H5N1 pada itik menggunakan isolat yang
diperoleh pada tahun 2002 sampai 2004 menimbulkan gejala klinis seperti
penurunan berat, lesu, diare, mata berkabut, dan ataksia kemudian mati meskipun
beberapa strain yang diisolasi selama tahun 2002-2004 juga menunjukkan gejala
yang ringan atau tidak ada sama sekali (Gambar 3) (Sturm-Ramirez et al. 2004).
Itik yang diinfeksi virus A/duck/Thailand/71.1/2004 menunjukkan gejala panas
tinggi, kesulitan bernafas, depresi, diare, gejala syaraf (ataksia, konvulsi, dan
inkoordinasi), dan konjungtivitis dengan mortalitas 20-100% (Songserm et al.
2006). Saat nekropsi ditemukan pendarahan titik dan terlokalisir pada kaki dan
20
paling menonjol ditemukan pada paru-paru seperti pneumonia, edema, kongesti,
dan peradangan perivaskuler (Songserm et al. 2006). Pada itik yang menunjukkan
gejala syaraf ditemukan kumpulan sel radang di sekitar pembuluh darah dan
peradangan neuroglia. Pada itik ras pedaging ditemukan degenerasi otot jantung
dengan perubahan patologis lain yang dapat terlihat antara lain hepatitis,
tubulonefritis, pengecilan kelenjar limfoid dan enteritis (Songserm et al. 2006).
Gambar 3. Grafik tingkat kelangsungan hidup itik yang diinfeksi berbagai isolat VAI H5N1 diadaptasi dari Sturm-Ramirez et al. (2005). Kelompok virus LPAI: A/Thai/1(Kan-1)/04 dan A/Ck/PP/BPPV3/04, HPAI: A/Dk/VN/40D/04, A/Ck/VN/48C/04, A/Dk/Thai/71.1/04, dan A/VN/1203/04.
AI Patogenitas Rendah (low-pathogenic avian influenza, LPAI)
Sasaran utama infeksi virus LPAI pada itik tidak hanya saluran pernafasan
dan jaringan paru-paru. Itik yang diinokulasi intranasal dengan virus LPAI
menunjukkan gejala paru-paru pneumonia ringan dan infiltrasi limfosit dan
makrofag dalam waktu 2 hari. Pewarnaan imunohistokimia nukleoprotein
menunjukkan perubahan pada sel epitel saluran pernafasan namun tidak ada
replikasi virus pada jaringan paru-paru (Cooley et al. 1989). Virus LPAI dapat
melewati saluran pencernaan atas itik dan bereplikasi dalam usus tanpa
menyebabkan manifestasi klinis penyakit (Webster et al. 1978; Kida et al. 1980).
Hasil penelitian yang menginokulasikan virus secara langsung pada tembolok dan
kloaka (Webster et al. 1978) serta ditemukannya titer virus yang tinggi pada feses
merupakan organ target virus LPAI pada itik sebagai tempat replikasi virus tanpa
infeksi pada paru-paru. Lebih spesifik, replikasi virus LPAI diyakini terjadi di
kriptus Lieberkühn usus besar (Kida et al. 1980).
Keragaman jenis itik juga berperan penting dalam patogenisitas virus
influenza. Embrio itik Mallard yang diinokulasi dengan virus LPAI memiliki
tingkat kematian lebih rendah daripada embrio entog. Antigen virus dapat
ditemukan di organ-organ internal seperti sinus hidung, faring, trakea, bronkus,
paru-paru, dan kantung hawa embrio itik Mallard tetapi tidak ditemukan pada
embrio entog. Alasan mortalitas dan replikasi virus pada itik Mallard ini tidak
jelas tetapi mendukung bukti bahwa itik Mallard berperan sebagai reservoir utama
virus LPAI di alam (Mutinelli et al. 2003).
Pemahaman mengenai respon imun itik terhadap VAI masih terbatas
meskipun beberapa penelitian mengenai respon antibodi serum itik yang terinfeksi
secara alami maupun eksperimental telah dilakukan (Suarez dan Schultz-Cherry
2000). Itik Pekin putih yang diinokulasi virus LPAI H7N2 memberikan hasil titer
antibodi HI yang sangat rendah tetapi virus tetap dikeluarkan hingga 7 hari pasca
inokulasi. Inokulasi ulang setelah 46 hari dengan strain virus yang sama memberi
respon antibodi yang lebih tinggi tetapi virus tidak ditemukan pada organ. Hasil
ini disertai rendahnya respon imun sekunder setelah inokulasi menggunakan virus
yang dilemahkan dalam formalin menunjukkan bahwa respon cepat imun pada
itik yang diinfeksi ulang dapat membatasi infeksi influenza untuk rentang waktu
tertentu (Kida et al. 1980). Infeksi yang pernah terjadi tidak dapat melindungi itik
terhadap infeksi berikutnya oleh subtipe virus lain. Sebagai contoh, itik yang
diinfeksi subtipe H4N6 terlindungi dari infeksi ulang dengan virus yang sama
tetapi mengeluarkan virion selama 8 hari setelah ditantang dengan isolat H11N3
(Austin dan Hinshaw 1984).
AI Patogenitas Tinggi (highly pathogenic avian influenza, HPAI)
Beberapa penelitian eksperimental telah dilakukan untuk memahami
patogenisitas virus HPAI H5N1 yang diisolasi sejak 2002 pada itik. Itik Pekin
Cherry Valley yang diinokulasi strain HPAI H5N1 isolat daging itik 2003 dari
22
dan kepala gemetar meskipun tidak mati. Titer virus yang tinggi ditemukan pada
organ pernafasan (paru-paru dan trakea), otak, hati, ginjal, dan usus besar disertai
perubahan mikroskopik pada otak (ensefalitis), jantung (miokarditis dengan
degenerasi dan nekrosis miosit), dan bursa (hiperplasia ringan pada folikel
limfoid) (Kishida et al. 2005).
Neurotropisme dan pankreatotropisme virus terlihat pada penelitian lain
yang menggunakan isolat virus HPAI. Itik yang ditantang dengan virus HPAI
H5N1 pada dosis letal menunjukkan gejala neurologis berat, seperti tortikolis,
inkoordinasi, tremor, dan kejang (Sturm-Ramirez et al. 2004; Vascellari et al.
2007). Imunohistokimia positif yang ditemukan pada otak dan batang otak serta
hibridisasi in situ virus yang terlihat pada neuron dan sel glia materi abu-abu otak
menunjukkan neurotropisme isolat setelah tahun 2002 (Sturm-Ramirez et al.
2004; Vascellari et al. 2007).
Meskipun rute masuknya virus ke dalam sistem saraf pusat belum dapat
dipastikan, setidaknya dua hipotesis dapat menjelaskan. Hipotesis pertama yaitu
transmisi virus dapat menjalar melalui serabut saraf vagus, olfaktorius, dan
trigeminus, dan hipotesis kedua yaitu virus dapat melakukan penetrasi melewati
blood-brain barrier (Silvano et al. 1997; Park et al. 2002).
Ciri-ciri lain virus HPAI H5N1 pada itik adalah titer virus yang sering lebih
tinggi pada usap orofaringeal dibandingkan usap kloaka (Sturm-Ramirez et al.
2004; Keawcharoen et al. 2008). Ekskresi virus HPAI H5N1 pada faring diduga
berasal dari paru-paru dan/atau kantung hawa karena hanya kedua jaringan ini
yang menunjukkan bukti replikasi virus secara imunohistokimia. Kecenderungan
ekskresi pada faring ini menunjukkan bahwa usap faring juga harus diambil ketika
melakukan surveilans VAI pada bebek liar selain usap kloaka yang selalu
dilakukan (Keawcharoen et al. 2008). Jika tidak, prevalensi HPAI H5N1 dapat
disalahperhitungkan.
Hasil penelitian FKH-IPB tahun 2006 menujukkan bahwa bebek yang tidak
dikandangkan memiliki resiko terinfeksi HPAI lebih tinggi (OR = 6,87; SK 95%;
1.29-36.54) dibandingkan dengan bebek yang dipelihara dalam kandang tertutup.
kecenderungan risiko positif AI yang lebih tinggi (OR = 4,05) dibandingkan
dengan yang tidak dicampur. Isolat virus HPAI H5N1 FKH/IPB/Duck/NG29 yang
ditemukan pada bebek sehat dapat menginfeksi ayam yang berkontak sehingga
pemeliharaan yang dicampur antara bebek dan ayam berpotensi meningkatkan
shedding virus dimana bebek berperan sebagai bank virus dan ayam sebagai
media propagasi (FKH-IPB 2006).
Teknik Diagnostik Avian Influenza
Diagnosa AI dilakukan dengan isolasi virus atau melalui deteksi dan
karakterisasi segmen genom virus karena gejala klinis yang ditimbulkan sangat
beragam menurut spesies inang, strain virus, status kekebalan inang, keberadaan
infeksi lain dan kondisi lingkungan (OIE 2009). Identifikasi VAI diawali dengan
isolasi virus pada ruang alantois telur ayam berembrio (TAB) specific pathogen
free (SPF). Selanjutnya cairan alantois diuji tapis dengan hemagglunation test
(HA) untuk mendeteksi keberadaan virus yang mampu mengaglutinasi sel darah
merah, kemudian diuji dengan agar gel immunodiffusion test (AGID) atau enzyme
linked immunosorbent assay (ELISA) yang masing-masing untuk mendeteksi tipe
dan subtipe virus. Pengujian subtipe virus juga dapat dilakukan dengan
hemagglutination inhibition test (HI) dan neuraminidase inhibition test (NI).
Alternatif lain untuk mendeteksi keberadaan VAI adalah dengan
reverse-transcription polymerase chain reaction (RT-PCR) ataureal time RT-PCR
(RRT-PCR) menggunakan primer spesifik matriks atau nukleoprotein. Selanjutnya
subtipe virus ditentukan dengan menggunakan primer spesifik hemaglutinin dan
neuraminidase. Uji serologis seperti AGID, HI, dan ELISA juga digunakan untuk
mendeteksi antibodi dalam serum inang.
RRT-PCR untuk Deteksi Avian Influenza
Perkembangan teknologi yang pesat memberikan berbagai pilihan teknik
dan produk yang dapat digunakan untuk mendukung pengujian diagnostik yang
telah ada atau menjadi landasan untuk pengujian diagnostik yang baru. Teknik
reverse transcriptase PCR (RT-PCR) secara konvensional telah dikembangkan
24
real-time RT-PCR (RRT-PCR) dalam rangka pengawasan rutin, selama wabah,
dan untuk penelitian karena lebih menguntungkan dari segi sensitivitas dan
spesifisitas yang lebih tinggi, memerlukan waktu yang lebih singkat, bersifat
kuantitatif, lebih ramah lingkungan, dan meskipun biaya yang diperlukan untuk
investasi peralatan lebih tinggi namun untuk operasional dan pengamanan
lingkungan teknik ini memerlukan biaya yang lebih sedikit.
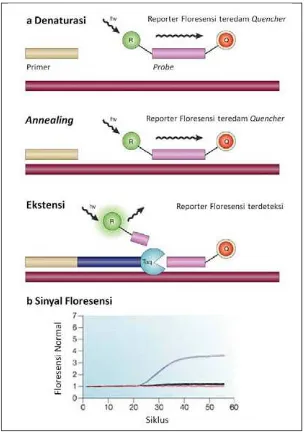
Pembacaan hasil RRT-PCR tidak memerlukan elektroforesis gel melainkan
dapat dilihat secara langsung berupa grafik intensitas pendaran zat warna
floresens yang meningkat secara eksponensial, linier, kemudian mendatar seiring
siklus amplifikasi (Gambar 4). Hasil RRT-PCR berupa nilai Ct (cycle threshold)
yang merupakan perpotongan antara kurva amplifikasi dengan garis threshold
yang menggambarkan konsentrasi relatif target PCR.Aktivitas nuklease ujung 5'-
polimerase yang terdapat dalam polymerase chain reaction (PCR) memecah
probe hidrolisis saat ekstensi amplikon sehingga memisahkan reporter (R)
florofor dari quencher (Q). Sinyal floresens yang dihasilkan ketika tereksitasi oleh
cahaya dari luar di setiap siklus PCR sebanding dengan jumlah produk yang
dihasilkan (Koch 2004).
Beberapa peningkatan yang penting demi perbaikan pengujian RRT-PCR
telah tersedia untuk VAI, antara lain: pengembangan dan penggunaan kontrol
internal untuk mengurangi negatif palsu reaksi dan pengembangan reagen kering
beku (lyophilized) untuk meningkatkan kualitas kontrol (Das et al. 2006; Di Trani
et al. 2006); penggunaan robot untuk meningkatkan keluaran laboratorium agar
mampu menangani peningkatan jumlah sampel selama wabah meskipun mungkin
tidak memberikan sensitivitas yang lebih baik (Spackman et al. 2002; Spackman
dan Suarez 2005); dan protokol baru untuk pengolahan sampel sulit seperti
sampel kloaka atau jaringan (Das et al. 2006).
Semua uji diagnostik molekuler bertujuan untuk memperoleh hasil yang
cepat dengan sensitifitas yang sebanding dengan isolasi virus, dan
mempertahankan tingkat spesifisitas yang tinggi. Oleh karena itu tiga titik kritis
RT-PCR, dan urutan basa primer dan probe. Ketiga faktor tersebut harus diperhatikan
agar pengujian menjadi sensitif dan spesifik.
Gambar 4. Prinsip probe hidrolisis TaqMan diadaptasi dari Koch (2004) (a)
proses hidrolisis probe saat ekstensi memisahkan reporter floresensi dari quencher (b) sinyal floresensi meningkat secara eksponensial, linier, kemudian mendatar seiring siklus amplifikasi.
Ekstraksi RNA merupakan tahap yang penting dalam setiap uji diagnostik
molekuler karena kualitas RNA akan mempengaruhi efisiensi amplifikasi.
Berbagai teknologi ekstraksi RNA yang ada seperti ekstraksi organik, ekstraksi
kolom silika, dan ekstraksi manik (beads) magnetik (Hale et al. 1996; Petrich et al.
26
Metode ekstraksi organik yang menggunakan fenol dan guanidinium
memberikan efisiensi ekstraksi yang baik untuk sampel kompleks tetapi metode
ini memungkinkan inhibitor PCR juga ikut terekstraksi sehingga dapat
menyebabkan hasil negatif palsu (Das et al. 2006). Metode ekstraksi juga
bervariasi dalam hal kemudahan penggunaan dan skalabilitas. Metode ekstraksi
organik relatif memerlukan tenaga intensif dan sulit untuk pengerjaan sampel
skala besar.
Beberapa metode ekstraksi berbasis kolom atau beads magnetik dapat
digunakan untuk keluaran besar misalnya dengan pemrosesan pada plat 96
sumuran atau menggunakan robot. Banyak sistem robotik yang tersedia secara
komersial dengan format reagen dan perlengkapan sendiri atau terbuka. Platform
kerja robotik juga sangat bervariasi dalam hal biaya tergantung pada kerumitan
dan fitur mesin. Platform kerja robotik cukup menjanjikan untuk meningkatkan
efisiensi laboratorium diagnostik, tetapi setiap robot dengan teknologi ekstraksi
RNA masih memerlukan validasi sebelum dapat digunakan secara rutin pada
sampel diagnostik. Kecil kemungkinan bagi sebuah mesin atau kit ekstraksi untuk
dapat memiliki fleksibilitas dalam menangani berbagai jenis sampel yang dibawa
ke laboratorium diagnostik dengan efisiensi ekstraksi RNA dan kemurnian yang
diperlukan untuk memperoleh hasil yang konsisten (Aguero et al. 2007; Tewari et
al. 2007). Diagnosis AI untuk ayam dan kalkun sebaiknya menggunakan sampel
usap trakea atau orofaringeal karena tropisme virus pada kedua spesies tersebut
adalah saluran pernafasan. Namun untuk spesies lain seperti itik
direkomendasikan sampel usap kloaka karena pada spesies tersebut virus LPAI
memiliki tropisme enterik. Sampel usap trakea/orofaringeal relatif mudah
digunakan untuk ekstraksi RNA karena mengandung sedikit sekali sel. Sampel
RNA lebih sulit diekstraksi dari usap kloaka dan jaringan karena keduanya
mengandung bahan organik lebih tinggi, komposisi kimia yang kompleks, dan
berpotensi mengandung inhibitor PCR (Cone et al. 1992; Buonagurio et al. 1999;
Petrich et al. 2006).
Reagen amplifikasi RT-PCR merupakan area kritis lain yang dapat
mempengaruhi hasil pengujian. Berbagai macam kit komersial dengan enzim dan
dilengkapi dengan tahap reverse transkripsi dan amplifikasi PCR, biasanya
dengan enzim yang berbeda untuk setiap tahapan. Kedua tahap tersebut sangat
penting untuk pengujian diagnostik yang sensitif. Secara umum RT-PCR dapat
dijalankan dengan prosedur dua tahap atau satu tahap (onestep). Pada prosedur
dua tahap, reverse transkripsi RNA dan amplifikasi DNA dijalankan secara
terpisah sehingga optimasi dilakukan di kedua reaksi. Sedangkan pada RT-PCR
onestep, semua reagen untuk tahapan reverse transkripsi RNA dan amplifikasi
DNA dimasukkan kedalam tabung yang sama sehingga pengujian dapat selesai
tanpa membuka tabung untuk memasukkan reagen tambahan. Prosedur dua tahap
dianggap lebih sensitif daripada metode onestep karena kedua tahapan dilakukan
pada kondisi yang optimal namun amplifikasi onestep menyederhanakan prosedur
dan mengurangi kemungkinan kontaminasi silang sampel sehingga prosedur
onestepini lebih baik untuk berbagai situasi (OIE 2008b).
Enzim-enzim untuk RT-PCR onestep dapat dibeli secara terpisah atau dalam
bentuk kit yang mencakup hampir semua reagen yang perlukan untuk pengujian.
Keuntungan penggunaan kit adalah peningkatan kontrol kualitas yang didapatkan
dari produk komersial selain lebih mudah untuk dipesan dan digunakan
dibandingkan dengan penggabungan reagen dari berbagai sumber. Meskipun
banyak kit diagnostik yang tersedia secara komersial, tidak semua memiliki
kinerja yang sama di setiap aplikasi. Prosedur diagnostik resmi RRT-PCR AI
yang diterapkan oleh jaringan laboratorium kesehatan hewan nasional (National
Animal Health Laboratory Network, NAHLN) Amerika Serikat yang dikelola
oleh layanan inspeksi kesehatan hewan dan tumbuhan (Animal and Plant Health
Inspection Service, APHIS) Departemen Pertanian Amerika Serikat (United States
Department of Agriculture, USDA) menggunakan kit untuk ekstraksi RNA dan
amplifikasi RT-PCR (Suarez et al. 2007). Kit alternatif kadang dapat bekerja
dengan baik sehingga ketentuan yang dibuat dalam protokol resmi NAHLN
memberi ruang bagi penggunaan metode alternatif untuk ekstraksi RNA atau
reagen amplifikasi RT-PCR (selain primer atau probe) namun pengguna harus
memiliki data yang cukup untuk menunjukkan bahwa protokol modifikasi sama
sensitifnya dengan protokol resmi. Perubahan protokol dapat terjadi bila prosedur