KEEFEKTIFAN Beauveria bassiana TERHADAP RAYAP
TANAH Coptotermes curvignathus
DEWI RAMDHANIA
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa tesis berjudul Keefektifan Beauveria bassiana terhadap Rayap Tanah Coptotermes curvignathus adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Juli 2015
RINGKASAN
DEWI RAMDHANIA. Keefektifan Beauveria bassiana terhadap Rayap Tanah Coptotermes curvignathus. Dibimbing oleh ACHMAD dan NOOR FARIKHAH HANEDA.
Rayap merupakan hama penting bagi kehidupan manusia yang dapat menyebabkan kerugian ekonomi di Indonesia. Rayap tidak hanya menyerang produk kayu, tetapi juga tanaman hidup dengan sebaran inang yang luas. Pengendalian rayap menggunakan termitisida sintetik telah banyak dilakukan karena dapat mengendalikan rayap secara cepat. Namun banyak dampak negatif yang ditimbulkan oleh termitisida sintetik seperti mengganggu kesehatan manusia, menyebabkan kerusakan lingkungan, dan menyebabkan hama sasaran menjadi resisten. Oleh karena itu penggunaan agensia hayati merupakan salah satu upaya untuk mengurangi implementasi termitisida sintetik karena dinilai lebih ramah lingkungan dan tidak menyebabkan resistensi pada rayap.
Penelitian ini bertujuan untuk: (1) mendapatkan media tumbuh padat dan cair yang paling sesuai untuk pertumbuhan diameter koloni dan biomassa isolat Beauveria bassiana; (2) menganalisis keefektifan tingkat kerapatan konidia B. bassiana dari berbagai jenis media tumbuh terhadap mortalitas rayap Coptotermes curvignathus.
Penelitian mengenai respon pertumbuhan B. bassiana pada media padat dan cair dirancang menggunakan rancangan acak lengkap (RAL) 1 faktor dengan faktor jenis media tumbuh. Sementara itu, aplikasi suspensi konidia B. bassiana terhadap rayap uji dirancang menggunakan rancangan acak lengkap faktorial (RAL Faktorial) dengan mengkombinasikan jenis media dan tingkat kerapatan konidia.
Tiga jenis media padat dan cair digunakan untuk mendapatkan respon pertumbuhan diameter koloni dan biomassa B. bassiana. Pertumbuhan diameter koloni B. bassiana tercepat adalah pada media sabouraud dextrose agar with yeast extract (SDAY). Kemudian berturut-turut adalah koloni B. bassiana pada media potato dextrose agar (PDA) dan malt extract agar (MEA). Produksi biomassa tertinggi adalah isolat yang diinkubasi pada media sabouraud dextrose broth with yeast extract (SDBY), kemudian isolat pada media malt extract broth (MEB) dan potato dextrose broth (PDB). Sementara itu, suspensi B. bassiana dengan kerapatan 108, 106, dan 104 konidia ml-1 dari media PDB, MEB, dan SDBY diaplikasikan pada rayap uji C. curvignathus. Respon mortalitas rayap C. curvignathus akibat aplikasi B. bassiana pada berbagai tingkat kerapatan konidia menunjukkan bahwa, perbedaan jenis media tidak mempengaruhi virulensi suspensi konidia B. bassiana terhadap mortalitas rayap, namun perbedaan kerapatan konidia berpengaruh terhadap mortalitas rayap.
SUMMARY
DEWI RAMDHANIA. Effectiveness of Beauveria bassiana Against Coptotermes curvignathus. Supervised by ACHMAD and NOOR FARIKHAH HANEDA.
Termite is an important pest to human life which can cause severe economic losses in Indonesia. Termite is not only attacking the wood products, but also the living plants with wide host distribution. The use of synthetic termiticide is the most widespread method for termite control because it can controls termite population quickly. But there are a lot of negative impact that caused by synthetic termiticide such as harms the human health, causes the environmental damage and resistance to target termite. Therefore the use of biological agents is one way to reduce the implementation of synthetic termiticide since it is considered to be
more environmentally friendly and doesn’t cause resistance to termites.
This research aimed : (1) to gain the most appropriate solid and liquid media for the growth of colony diameter and biomass of Beauveria bassiana; (2) to analyze the effectiveness of conidial density level of B. bassiana from various media type against Coptotermes curvignathus mortality.
The research on the growth response of B. bassiana in solid and liquid media was designed using completely randomized design with media types as the only factor. Meanwhile, application of B. bassiana conidial suspension against termites was designed using factorial completely randomized design by combining media types with conidial density level.
Three types of solid and liquid media were used to determine the best response of the colony diameter growth and biomass production of Beauveria bassiana. The fastest colony diameter growth was obtained in sabouraud dextrose agar with yeast extract (SDAY) media followed by potato dextrose agar (PDA) and malt extract agar (MEA). The highest biomass production was obtained in sabouraud dextrose broth with yeast extract (SDBY) media followed by malt extract broth (MEB) and potato dextrose broth (PDB). Meanwhile, B. bassiana suspension with density of 108, 106, and 104 conidia ml-1 from PDB, MEB, and SDBY media were applied to C. curvignathus. The results showed that all types of nutrients sources (PDB, MEB and SDBY) did not affect the virulence of B. bassiana conidial suspension against C. curvignathus mortality. Nevertheless there were significant differences among conidial density level from B. Bassiana against C. curvignathus mortality.
© Hak Cipta Milik IPB, Tahun 2015
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
KEEFEKTIFAN Beauveria bassiana TERHADAP RAYAP
TANAH Coptotermes curvignathus
DEWI RAMDHANIA
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Silvikultur Tropika
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Judul Tesis : Keefektifan Beauveria bassiana terhadap Rayap Tanah Coptotermes curvignathus
Nama : Dewi Ramdhania
NIM : E451110011
Disetujui oleh
Komisi Pembimbing
Prof Dr Ir Achmad, MS Ketua
Dr Ir Noor Farikhah Haneda, MS Anggota
Diketahui oleh
Ketua Program Studi Silvikultur Tropika
Prof Dr Ir Sri Wilarso Budi R, MS
Dekan Sekolah Pascasarjana
Dr Ir Dahrul Syah, MScAgr
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah Subhanahu Wa Ta’ala atas segala karunia-Nya sehingga karya ilmiah ini berhasil diselesaikan. Tema yang dipilih dalam penelitian yang dilaksanakan sejak bulan Agustus 2014 ini ialah pengendalian hama tanaman kehutanan, dengan judul Keefektifan Beauveria bassiana terhadap Rayap Tanah Coptotermes curvignathus.
Terima kasih penulis sampaikan kepada Prof Dr Ir Achmad, MS dan Dr Ir Noor Farikhah Haneda, MS selaku pembimbing yang telah membimbing
penulis dalam menyelesaikan studi. Tak lupa penulis sampaikan terima kasih kepada Kepala Balai Besar Perbenihan dan Proteksi Tanaman Perkebunan (BBP2TP) Surabaya dan Kepala Laboratorium Rayap Departemen Hasil Hutan Fakultas Kehutanan IPB yang telah memberikan izin pada penulis untuk melaksanakan kegiatan penelitian di lingkungan kerjanya. Terima kasih pula penulis sampaikan kepada semua pihak yang telah membantu teknis pelaksanaan penelitian ini.
Ungkapan terima kasih yang mendalam juga penulis sampaikan kepada suami tercinta yang senantiasa mendukung serta anakku tersayang yang senantiasa mendampingi penulis. Rasa hormat dan terima kasih penulis sampaikan kepada ayah dan ibu serta seluruh keluarga besar atas segala do’a dan kasih sayangnya.
Semoga karya ilmiah ini bermanfaat.
Bogor, Juli 2015
DAFTAR ISI
Coptotermes curvignathus Holmgren 3
Beauveria bassiana 5
3 BAHAN DAN METODE
Waktu dan Lokasi 9
Bahan dan Alat 9
Metode 9
A. Pengujian Respon Pertumbuhan Radial Koloni dan
Biomassa B. bassiana 10
Kandungan Nutrisi pada Media Tumbuh 18
Keefektifan B. bassiana terhadap rayap C. curvignathus 20
DAFTAR TABEL
1 Unit percobaan respon pertumbuhan diameter koloni B. bassiana 12
2 Unit percobaan respon pertumbuhan biomassa B. bassiana 12
3 Respon rataan pertumbuhan diameter B. bassiana pada berbagai jenis
media padat 17
4 Komposisi nutrisi yang terdapat pada masing-masing media tumbuh 18 5 Persentase mortalitas kumulatif C. curvignathus yang terinfeksi
cendawan B. bassiana dari tiga jenis media pada berbagai tingkat
kerapatan konidia selama 11 hari 20
6 Respon mortalitas rayap C. curvignathus akibat aplikasi B. bassiana
pada berbagai tingkat kerapatan konidia 22
DAFTAR GAMBAR
1 Laju pertambahan kasus resistensi serangga terhadap insektisida 5 2 Struktur makroskopis (a) dan mikroskopis (b) B. bassiana 6 3 Tipe pertumbuhan dimorfik B. bassiana. (A) fase parasit menyerupai
khamir saat menginfeksi inang. (B) fase saprofit menunjukan filamen
hifa 6
4 Siklus pertumbuhan B. bassiana pada media agar 7
5 Diagram proses infeksi B. bassiana 8
6 Skema penelitian 10
7 Metode pengukuran diameter radial koloni B. bassiana 11
8 Tahapan pengenceran konidia 13
9 Pertumbuhan diameter koloni B. bassiana pada media potato dextrose agar (PDA), malt extract agar (MEA) dan sabouraud dextrose agar
with yeast extract (SDAY) 15
10 Laju pertumbuhan koloni B. bassiana pada media potato dextrose agar (PDA), malt extract agar (MEA), dan sabouraud dextrose agar with yeast extract (SDAY) selama 15 hari setelah inokulasi 16 11 Biomassa B. bassiana pada media potato dextrose broth (PDB), malt
extract broth (MEB), dan sabouraud dextrose broth with yeast extract
(SDBY) selama 4 minggu 17
12 Laju mortalitas C. curvignathus akibat aplikasi B. bassiana dari media tumbuh potato dextrose broth (A), malt extract broth (B), dan sabouraud dextrose with yeast extract (C) pada tingkat kerapatan konidia kontrol, 104, 106, dan 108 konidia ml-1 selama 11 hari
pengamatan 21
DAFTAR LAMPIRAN
1 Proses Pengujian Respon Pertumbuhan Radial Koloni 27
2 Proses Pengujian Respon Produksi Biomassa 28
3 Hasil analisis sidik ragam dan uji selang berganda Duncan uji in vitro pengaruh media kultur terhadap pertumbuhan isolat B. bassiana 29 4 Rekapitulasi sidik ragam jenis media cair dan tingkat kerapatan konidia
1 PENDAHULUAN
Latar Belakang
Rayap merupakan hama penting bagi kehidupan manusia karena aktivitasnya yang kerap menyerang bangunan kayu dan material berselulosa lain. Kerugian ekonomis yang ditimbulkan oleh rayap di Indonesia mencapai 1.67 triliun rupiah pada tahun 1995 dan diperkirakan mencapai 2.8 triliun rupiah pada tahun 2015 (Nandika 2015 dalam Sudiaman 2015).
Rayap tidak hanya menyerang produk kayu, tetapi juga tanaman hidup dengan sebaran inang yang luas. Menurut Nair (2001) terdapat 6 spesies pohon yang berpotensi terserang rayap antara lain Karet, Pinus, Ekaliptus, Akasia, Gmelina, dan Mahoni. Jenis rayap yang dapat menyebabkan kerusakan serius berasal dari genus Coptotermes sp. Coptotermes curvignathus dapat menyebabkan kematian 10 - 50% pancang Acacia mangium berusia kurang dari 1 tahun di Sumatera. Pribadi (2010) menyatakan bahwa, Coptotermes sp. berpotensi menyerang tegakan jabon di areal hutan tanaman industri (HTI) lahan gambut dengan intensitas serangan mencapai 9%. Dengan demikian perlu dilakukan upaya pengendalian populasi rayap karena ancaman bahaya serangan yang tidak hanya pada bangunan namun juga pada tanaman hidup dengan sebaran inang yang luas.
Pengendalian rayap menggunakan termitisida sintetik banyak dilakukan karena dapat mengendalikan rayap secara cepat. Namun banyak dampak negatif yang ditimbulkan oleh termitisida sintetik seperti mengganggu kesehatan manusia, menyebabkan kerusakan lingkungan, dan menyebabkan hama sasaran menjadi resisten. Oleh karena itu penggunaan agensia hayati merupakan salah satu upaya untuk mengurangi implementasi termitisida sintetik karena dinilai lebih ramah lingkungan dan tidak menyebabkan resistensi pada rayap. Beberapa jenis agensia hayati dari golongan mikroorganisme yang pernah diteliti sebagai pengendali rayap Coptotermes sp. di antaranya adalah nematoda (Erningtyas 2006), bakteri Bacillus thuringiensis dan cendawan Metharizium anisopliae (Sukartana et al. 2000), serta cendawan Beauveria bassiana (Desyanti 2007).
2
mengenai fisiologi dasar dan optimalisasi produksi miselia dan spora dalam waktu cepat dengan biaya yang rendah akan sangat bermanfaat untuk produksi massal dan komersialisasi. Penelitian ini menggunakan beberapa jenis media untuk menumbuhkan isolat B. bassiana kemudian mengaplikasikan suspensi konidia B. bassiana pada C. curvignathus dengan beberapa tingkat kerapatan.
Perumusan Masalah
Beberapa permasalahan yang muncul untuk dicarikan solusi, yaitu:
1 Media apakah yang paling sesuai untuk pertumbuhan diameter koloni dan biomassa isolat B. bassiana?
2 Berapakah kerapatan konidia B. bassiana yang paling efektif untuk mengendalikan rayap C. curvignathus?
Tujuan Penelitian
Tujuan penelitian ini adalah:
1 Mendapatkan media tumbuh padat dan cair yang paling sesuai untuk pertumbuhan diameter koloni dan biomassa isolat B. bassiana.
2 Menganalisis keefektifan tingkat kerapatan konidia B. bassiana dari berbagai jenis media tumbuh terhadap mortalitas rayap C. curvignathus.
Manfaat Penelitian
Manfaat penelitian ini antara lain :
1 Memberikan sumbangsih bagi ilmu pengetahuan khususnya di bidang budidaya tanaman hutan dan pengendalian hama tanaman hutan.
2 TINJAUAN PUSTAKA
Spesies : Coptotermes curvignathus (Sumber: Nandika et al. 2003)
Karakteristik
Rayap merupakan serangga dengan tubuh kecil, lunak dan hidup dalam koloni sehingga disebut serangga sosial dengan sistem kasta yang berkembang sempurna (Nandika et al. 2003). Ia juga menyatakan bahwa dalam kehidupannya rayap memiliki beberapa sifat penting yaitu:
1 Trophalaxis, yaitu sifat berkumpul saling menjilat serta mengadakan pertukaran makanan
2 Cryptobiotic, yaitu sifat menjauhi cahaya, namun sifat ini tidak berlaku pada imago rayap yang bersayap.
3 Necrophagy, yaitu sifat memakan bangkai sesamanya
4 Cannibalism, yaitu sifat memangsa sesamanya terutama yang lemah dan sakit
5 Polimorfisme, yaitu perbedaan bentuk rayap antar kasta.
Rayap merupakan serangga sosial yang hidup di dalam koloni. Koloni rayap membagi kegiatan-kegiatan utamanya dalam bentuk kasta spesifik. Berdasarkan perbedaan morfologi dan fisiologinya, rayap dibagi menjadi tiga kasta yaitu kasta reproduktif, kasta prajurit, dan kasta pekerja. Rayap merupakan hewan yang paling kompleks secara anatomi dan adaptasi (Eggleton 2011).
Nandika et al. (2003) menjabarkan sistem kasta rayap sebagai berikut: 1 Kasta Reproduktif
Kasta reproduktif terdiri atas raja dan ratu. Raja bertugas membuahi ratu. Kasta ini dapat dibedakan menjadi kasta reproduktif primer dan suplementer atau neoten. Kasta reproduktif suplementer atau neoten terbentuk bila kasta reproduktif primer telah mati atau hilang karena fragmentasi koloni. Selanjutnya neoten menggantikan fungsi kasta reproduktif primer untuk mengembangkan koloni.
2 Kasta Prajurit
4
hidup koloni khususnya semut dan vertebrata predator. Kasta prajurit mampu menyerang musuhnya dengan mandibel.
3 Kasta Pekerja
Kasta pekerja merupakan anggota koloni yang sangat penting dengan jumlah populasi tidak kurang dari 80%. Kasta pekerja umumnya berwarna pucat dengan kutikula mengalami sedikit penebalan sehingga terlihat menyerupai nimfa. Hampir semua tugas koloni dikerjakan oleh kasta pekerja kecuali proses perkembangbiakan dan pertahanan koloni. Kasta pekerja bertugas untuk memelihara telur, memberi makan ratu dan semua anggota koloni, mencari sumber makanan, jenis tertentu menumbuhkan jamur, dan memperbaiki sarang jika terjadi kerusakan. Kasta pekerja menyebabkan kerusakan pada tanaman, kayu, dan bahan berselulosa lainnya.
Hill (2008) menyatakan bahwa, C. curvignathus memiliki sebaran inang yang luas mencakup pepohonan dan tanaman budidaya seperti kopi, cabai, dan tebu. C. curvignathus dapat menyerang pohon sehat kemudian dapat membunuh pohon tersebut dalam beberapa minggu. Pohon yang terserang C. curvignathus menjadi layu, lalu batang dan cabang pohon berlubang karena dimakan oleh rayap, kemudian pohon tersebut tumbang.
Nandika dan Tambunan (1990) menyatakan bahwa, rayap subteran (Coptotermes curvignathus Holmgren) dan rayap tanah (Macrotermes spp.) termasuk rayap yang paling luas serangannya di Indonesia. Menurut Pribadi (2010), Coptotermes sp. merupakan hama kedua tertinggi yang menyerang jabon pada HTI di lahan gambut walaupun dengan tingkat kejadian serangan yang masih tergolong sangat ringan yaitu antara 3.5 - 9.1%.
Pengendalian Hama
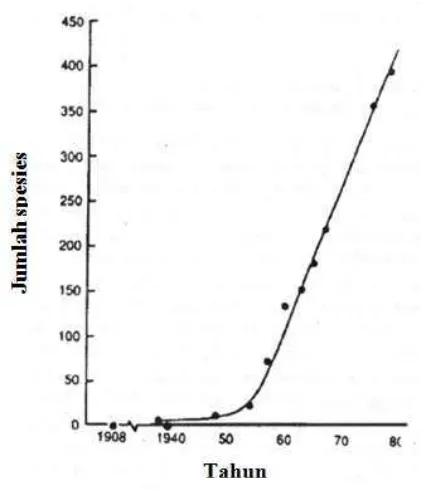
Sudah sejak lama manusia melakukan pengendalian hama secara konvensional menggunakan berbagai jenis insektisida. Akan tetapi efek samping penggunaan insektisida yang tidak terkendali dapat menyebabkan hama menjadi resisten terhadap insektisida tersebut. Resistensi hama terhadap insektisida dapat menimbulkan akibat yang lebih parah yaitu timbulnya resurjensi dan ledakan hama. Georghio (1986) menyatakan bahwa, lebih dari 500 spesies serangga resisten terhadap insektisida. Gambar 1 menunjukkan laju pertambahan kasus resistensi serangga terhadap insektisida selama beberapa tahun.
5
Gambar 1 Laju pertambahan kasus resistensi serangga terhadap insektisida.
Sumber : Georghio (1986)
Beauveria bassiana
Klasifikasi
Kingdom : Fungi
Filum : Ascomycota
Kelas : Sordariomycetes
Ordo : Hypocreales
Famili : Cordycipitaceae
Genus : Beauveria
Spesies : Beauveria bassiana
Sumber: Berman (2012)
Karakteristik
6
a b
Gambar 2 Struktur makroskopis (a) dan mikroskopis (b) B. bassiana
Beberapa keuntungan penggunaan cendawan sebagai insektisida hayati dijelaskan juga oleh Wan (2003), yaitu:
a Tidak berpengaruh pada mamalia dan hal tersebut mengurangi bahaya yang biasa ditemui pada aplikasi insektisida.
b Berkurangnya masalah resisten pada serangga.
c Berpotensi untuk dikembangkan lebih lanjut dengan penelitian bioteknologi.
d Cendawan entomopatogen memiliki ketahanan yang tinggi dalam lingkungan sehingga dapat memberikan efek jangka panjang dalam mengendalikan hama.
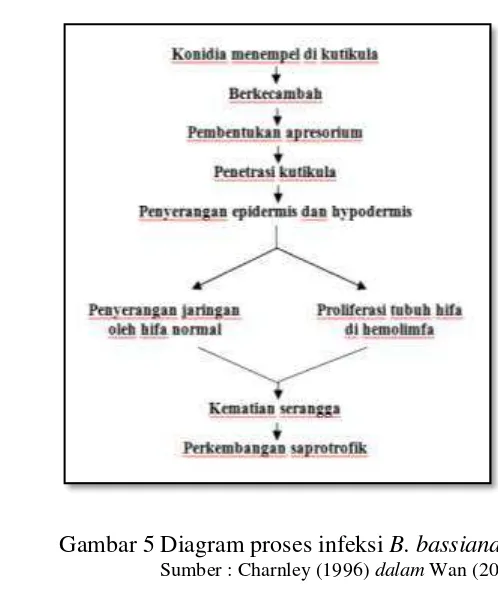
Wan (2003) menyatakan bahwa, B. bassiana memiliki tipe pertumbuhan dimorfik. Pada saat tidak ada serangga inang yang spesifik, B. bassiana mengalami siklus hidup vegetatif aseksual yang meliputi perkecambahan konidia, pertumbuhan filamen hifa, dan pembentukan konidia secara simpodial. Siklus hidup B. bassiana beralih menjadi patogen ketika ada inang (Gambar 3).
A B
7
Alves et al. (2002) menunjukan proses perkembangan B. bassiana pada media agar (Gambar 4). Neves dan Alves (2004) menyatakan bahwa, konidia B. bassiana berkecambah pada kutikula Cornitermes cumulans Kollar (Isoptera: Termitidae) antara 6 sampai 12 jam setelah aplikasi. Penetrasi pada umumnya terjadi 12 sampai 24 jam setelah inokulasi dan kematian serangga terjadi antara 72 sampai 96 jam setelah aplikasi.
Gambar 4 Siklus pertumbuhan B. bassiana pada media agar.
Sumber : Alves et al. (2002)
Hashim (2000) menyatakan bahwa, konidia B. bassiana berkecambah pada kutikula larva Crocidolomia binotalis Zell. 4 sampai 6 jam setelah aplikasi. Kemudian larva terinfeksi akan mati dalam kurun waktu 24 sampai 48 jam. Hifa B. bassiana akan menerobos keluar kutikula dalam waktu 2 sampai 5 hari.
Aplikasi efektif B. bassiana adalah dengan cara menyemprot kanopi tanaman dengan suspensi B. bassiana agar terjadi kontak antara konidia dengan hama sasaran. B. bassiana juga dapat diaplikasikan dengan cara ditaburkan atau disemprotkan pada permukaan tanah, atau dicampur dengan tanah maupun kompos. Konidia B. bassiana mudah diinaktifkan oleh sinar ultraviolet sehingga pencegahannya dapat dilakukan dengan melakukan aplikasi pada pagi hari yaitu sebelum pukul 08.00 atau sore hari setelah pukul 15.00 (Soetopo dan Igaa 2007).
8
Gambar 5 Diagram proses infeksi B. bassiana Sumber : Charnley (1996) dalam Wan (2003)
Khashaveh et al. (2011) menyatakan bahwa, saat ini beberapa formulasi cendawan B. bassiana tersedia secara komersil dan terdaftar untuk digunakan pada fasilitas penyimpanan. Cendawan entomopatogen dapat digunakan untuk mengendalikan sisa hama pada tempat penyimpanan yang kosong sebelum hasil panen yang baru dimasukkan ke dalam tempat penyimpanan tersebut.
Daya simpan produk agensia hayati juga merupakan salah satu hal yang
3 BAHAN DAN METODE
Waktu dan Lokasi
Penelitian dilaksanakan pada bulan Agustus 2014 sampai dengan Maret 2015 di Laboratorium Patologi Hutan dan Entomologi Hutan, Bagian Perlindungan Hutan Departemen Silvikultur, dan Laboratorium Rayap Departemen Hasil Hutan, Fakultas Kehutanan IPB.
Bahan dan Alat
Bahan yang digunakan dalam penelitian antara lain : rayap C. curvignathus kasta pekerja dan prajurit, isolat B. bassiana berasal dari Balai Besar Perbenihan dan Proteksi Tanaman Perkebunan (BBP2TP) Surabaya, potato dextrose agar (HiMediaTM), Malt extract (HiMediaTM), sabouraud dextrose agar (HiMediaTM), yeast extract (HiMediaTM), pepton (DifcoTM), dextrose, pure agar, kentang, antibiotik kloramfenikol, aquades, alkohol dan spirtus. Adapun peralatan yang digunakan antara lain : autoclave, labu erlenmeyer, gelas ukur, botol selai, cawan petri, laminar air flow, lampu spirtus, kertas saring, mikroskop, timbangan elektrik, haemocytometer, test tube, corong plastik, aluminium foil, kapas, mikropipet, gelas piala, korek api, dan kamera.
Metode
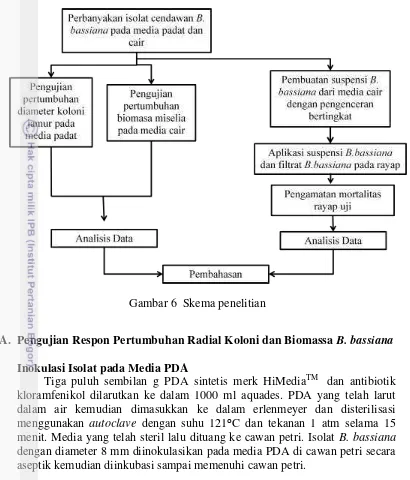
Penelitian yang dilakukan terdiri dari dua aspek yaitu pengujian respon pertumbuhan B. bassiana pada media padat dan cair serta aplikasi B. bassiana terhadap rayap uji pada berbagai tingkat kerapatan konidia. Aspek pertama yaitu pengujian respon pertumbuhan B. bassiana pada media padat dan cair. Pengujian respon pertumbuhan B. bassiana pada media padat dirancang menggunakan rancangan acak lengkap (RAL) 1 faktor dengan faktor jenis media tumbuh potato dextrose agar (PDA), malt extract agar (MEA), dan sabouraud dextrose agar with yeast extract (SDAY) dengan 4 ulangan. Pengujian respon B. bassiana pada media cair dirancang menggunakan rancangan acak lengkap (RAL) 1 faktor dengan faktor jenis media tumbuh potato dextrose broth (PDB), malt extract broth (MEB), dan sabourauddextrosebroth with yeast extract (SDBY) dengan 7 ulangan.
10
Gambar 6 Skema penelitian
A. Pengujian Respon Pertumbuhan Radial Koloni dan Biomassa B. bassiana
Inokulasi Isolat pada Media PDA
Tiga puluh sembilan g PDA sintetis merk HiMediaTM dan antibiotik kloramfenikol dilarutkan ke dalam 1000 ml aquades. PDA yang telah larut dalam air kemudian dimasukkan ke dalam erlenmeyer dan disterilisasi menggunakan autoclave dengan suhu 121⁰C dan tekanan 1 atm selama 15 menit. Media yang telah steril lalu dituang ke cawan petri. Isolat B. bassiana dengan diameter 8 mm diinokulasikan pada media PDA di cawan petri secara aseptik kemudian diinkubasi sampai memenuhi cawan petri.
Inokulasi Isolat pada Media MEA
Pembuatan media MEA mengacu pada Herliyana (2007) yaitu dengan melarutkan 15 g malt extract, 15 g pureagar dan antibiotik kloramfenikol ke dalam 1000 ml aquades. Larutan MEA lalu disterilisasi kemudian diinokulasikan isolat B. bassiana dengan metode yang sama seperti metode inokulasi isolat pada media PDA.
Inokulasi Isolat pada Media SDAY
11
Inokulasi Isolat pada Media PDB
Dua ratus g kentang diiris tipis kemudian direbus dengan 1000 ml aquades lalu diambil ekstraknya. Ekstrak kentang ditambahkan dengan 20 g dextrose dan antibiotik kloramfenikol setelah itu ditambahkan air hingga menjadi 1000 ml larutan. Larutan dimasukkan ke dalam botol selai sebanyak 50 ml lalu disterilisasi menggunakan autoclave dengan suhu 121⁰C dan dan antibiotik kloramfenikol dilarutkan ke dalam 1000 ml aquades. Larutan SDBY lalu disterilisasi kemudian diinokulasikan isolat B. bassiana dengan metode yang sama seperti metode inokulasi isolat pada media PDB.
Respon Pertumbuhan Radial Koloni B. bassiana pada Berbagai Jenis Media Padat
Metode pengujian pertumbuhan radial koloni dan biomassa B. bassiana mengacu pada Senthamizhlselvan et al. (2010). Media tumbuh padat yang digunakan dalam penelitian ini adalah PDA, MEA dan SDAY. Isolat B. bassiana berdiameter 8 mm diinokulasikan di tengah masing-masing media pada cawan petri berukuran 9 cm secara aseptik. Respon pertumbuhan radial dihitung sejak hari pertama setelah inokulasi (HSI) selama 15 hari dengan mengukur pola pertumbuhan radial berupa diameter koloni. Diameter koloni ditentukan dengan membuat garis tegak lurus melewati sumbu koloni. Batas terluar dari koloni berupa diameter harian ( ) ditandai dan diukur
Gambar 7 Metode pengukuran diameter radial koloni B. bassiana
Kemudian rataan diameter koloni diperoleh dengan rumus :
12
Respon Pertumbuhan Biomassa B. bassiana pada Berbagai Jenis Media Cair
Biomasa B. bassiana dihitung pada minggu ke-4 setelah inokulasi. Miselia B. bassiana dipisahkan dari media PDB, MEB, dan SDBY menggunakan kertas saring. Kertas saring dan miselia dikeringkan menggunakan oven selama 24 jam pada suhu 60°C. Biomassa B. bassiana ditentukan menggunakan rumus sebagai berikut:
Biomassa (g) = B2 – B1
Keterangan :
B2 = Berat kering miselia dan kertas saring (g) B1 = Berat kering kertas saring (g)
Rancangan Percobaan
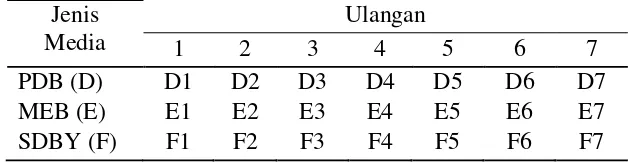
a. Rancangan percobaan pertumbuhan diameter koloni B. bassiana Percobaan respon pertumbuhan diameter koloni B. bassiana terdiri dari 1 faktor perlakuan yaitu perlakuan dengan menggunakan media solid PDA (A), MEA (B) dan SDAY (C). Setiap perlakuan diulang 4 kali sehingga terdapat 12 unit percobaan (Tabel 1).
Tabel 1 Unit percobaan respon pertumbuhan diameter koloni
Jenis Media Ulangan
1 2 3 4
PDA (A) A1 A2 A3 A4
MEA (B) B1 B2 B3 B4
SDAY (C) C1 C2 C3 C4
b. Rancangan percobaan pertumbuhan biomassa B. bassiana
Percobaan respon pertumbuhan biomassa B. bassiana terdiri dari 1 faktor perlakuan yaitu perlakuan dengan menggunakan media cair PDB (D), MEB (E) dan SDBY (F). Setiap perlakuan diulang 7 kali sehingga terdapat 21 unit percobaan (Tabel 2).
Tabel 2 Unit percobaan respon pertumbuhan biomassa
13
Model linear RAL yang digunakan untuk percobaan respon pertumbuhan diameter koloni dan biomassa B. bassiana adalah:
Keterangan:
Yij : pengamatan pada perlakuan ke-i, ulangan ke-j
µ : rataan umum
τi : pengaruh perlakuan ke-i
εij : pengaruh acak pada perlakuan ke-i ulangan ke-j
B. Aplikasi B. bassiana pada Rayap Coptotermes curvignathus
Persiapan Rayap Uji dan Pembuatan Suspensi Larutan B. bassiana dari Media Cair
Serangga yang digunakan dalam penelitian ini adalah rayap C. curvignathus yang diperoleh dari Puslitbang Keteknikan Kehutanan dan Pengolahan Hasil Hutan Gunung Batu Bogor. Sebanyak 30 ekor rayap kasta pekerja dan 3 ekor kasta prajurit C. curvignathus ditempatkan ke dalam cawan petri steril dengan pakan kertas saring steril.
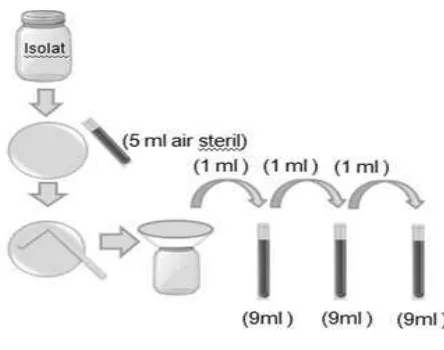
Isolat B. bassiana pada media PDB, MEB, dan SDBY dipisahkan dari media cair, kemudian ditambahkan 5 ml aquades steril. Konidia B. bassiana dipisahkan dari miselium menggunakan batang penyebar atau spatula steril lalu disaring menggunakan corong dan kain saring ke dalam wadah steril. Pengenceran tahap pertama diperoleh dengan mengambil 1 ml suspensi konidia menggunakan mikropipet dan dimasukkan dalam tabung reaksi berisi 9 ml air steril (Gambar 8).
14
Setelah diencerkan, kemudian jumlah konidia dihitung menggunakan haemocytometer agar kerapatan konidia dapat dihitung berdasarkan rumus (Sudibyo 1994) berikut:
Keterangan:
C : Kerapatan konidia
t : total konidia di semua kotak haemocytometer yang diamati n : jumlah semua kotak haemocytometer yang diamati.
Kerapatan konidia B. bassiana yang digunakan adalah 108, 106, dan 104 konidia ml-1, sedangkan untuk perlakuan kontrol digunakan masing-masing media cair tanpa penambahan konidia. Setiap perlakuan diulang 3 kali sehingga terdapat 36 cawan petri dengan total rayap 1188 ekor.
Aplikasi Suspensi B. bassiana dan Perhitungan Mortalitas Rayap Uji
Aplikasi B. bassiana dilakukan dengan cara meneteskan suspensi konidia pada kertas saring yang merupakan pakan rayap dalam cawan petri. Setiap cawan petri berisi 30 ekor rayap kasta pekerja dan 3 ekor kasta prajurit. Mortalitas rayap diamati setiap 24 jam dengan mencatat jumlah rayap yang mati pada masing-masing unit selama 11 hari pengamatan, kemudian
Model linear dari rancangan tersebut adalah
Keterangan:
Yijk : mortalitas rayap dari konidia yang berasal dari jenis media ke-i, kerapatan konidia taraf ke-j dan ulangan ke-k.
µ : rataan umum
αi : pengaruh jenis media ke-i
βj : pengaruh kerapatan konidia taraf ke-j
4 HASIL DAN PEMBAHASAN
Respon Pertumbuhan Radial Koloni B. bassiana pada Media Padat secara
In Vitro
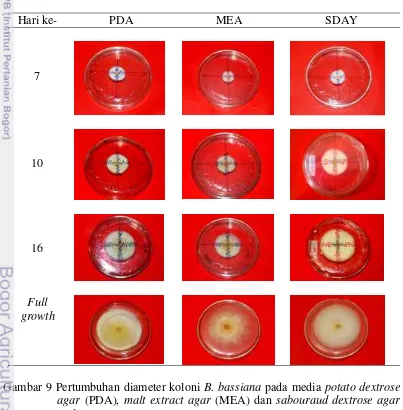
Isolat B. bassiana memiliki kecepatan tumbuh radial yang berbeda pada ketiga jenis media tumbuh. Berdasarkan pengamatan secara makroskopis, pertumbuhan radial koloni B. bassiana tercepat untuk dapat memenuhi cawan petri berukuran 9 cm adalah pertumbuhan koloni pada media SDAY. Pada media SDAY isolat B. bassiana membutuhkan waktu inkubasi selama 26 hari setelah inokulasi (HSI) untuk memenuhi cawan petri, sedangkan koloni B. bassiana pada media PDA dan MEA membutuhkan waktu 27 HSI dan 31 HSI untuk memenuhi cawan petri. Pertumbuhan diameter koloni B. bassiana pada media PDA, MEA dan SDAY disajikan pada Gambar 9.
Hari ke- PDA MEA SDAY
7
10
16
Full growth
16
Isolat B. bassiana pada media PDA dan SDA hampir memenuhi cawan petri berukuran 9 cm pada hari ke-7 (Senthamizhlselvan et al. 2010), sehingga sangat berbeda hasilnya dengan hasil penelitian ini. Hal ini diduga disebabkan oleh perbedaan strain isolat B. bassiana yang diteliti. Chang dan Hayes (1978) dalam Syafiih (2015) menyatakan bahwa, pertumbuhan jamur melibatkan banyak faktor antara lain adalah jenis jamur itu sendiri, komposisi media dan lingkungan.
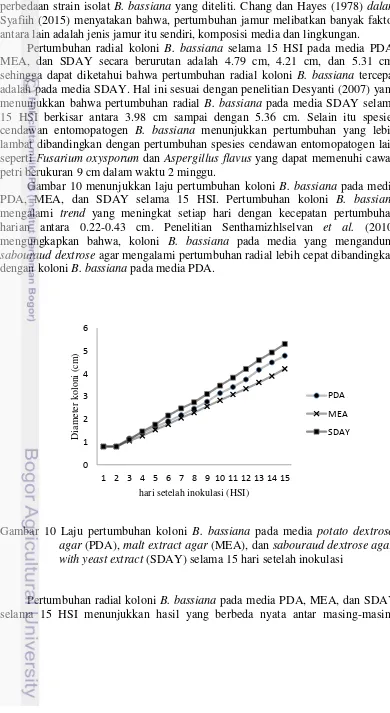
Pertumbuhan radial koloni B. bassiana selama 15 HSI pada media PDA, MEA, dan SDAY secara berurutan adalah 4.79 cm, 4.21 cm, dan 5.31 cm, sehingga dapat diketahui bahwa pertumbuhan radial koloni B. bassiana tercepat adalah pada media SDAY. Hal ini sesuai dengan penelitian Desyanti (2007) yang menunjukkan bahwa pertumbuhan radial B. bassiana pada media SDAY selama 15 HSI berkisar antara 3.98 cm sampai dengan 5.36 cm. Selain itu spesies cendawan entomopatogen B. bassiana menunjukkan pertumbuhan yang lebih lambat dibandingkan dengan pertumbuhan spesies cendawan entomopatogen lain seperti Fusarium oxysporum dan Aspergillus flavus yang dapat memenuhi cawan petri berukuran 9 cm dalam waktu 2 minggu.
Gambar 10 menunjukkan laju pertumbuhan koloni B. bassiana pada media PDA, MEA, dan SDAY selama 15 HSI. Pertumbuhan koloni B. bassiana mengalami trend yang meningkat setiap hari dengan kecepatan pertumbuhan harian antara 0.22-0.43 cm. Penelitian Senthamizhlselvan et al. (2010) mengungkapkan bahwa, koloni B. bassiana pada media yang mengandung sabourauddextrose agar mengalami pertumbuhan radial lebih cepat dibandingkan dengan koloni B. bassiana pada media PDA.
Gambar 10 Laju pertumbuhan koloni B. bassiana pada media potato dextrose agar (PDA), malt extract agar (MEA), dan sabouraud dextrose agar with yeast extract (SDAY) selama 15 hari setelah inokulasi
17
media. Pertumbuhan radial koloni B. bassiana tercepat adalah pada media SDAY. Kemudian berturut-turut adalah koloni B. bassiana pada media PDA dan MEA (Tabel 3). Hal ini diduga karena kandungan pepton dan yeast extract pada media SDAY merupakan nutrisi optimal bagi pertumbuhan radial koloni B. bassiana pada media padat secara in vitro.
potato dextrose agar (PDA) 2.59125b
malt extract agar (MEA) 2.34125c
* Nilai diameter yang diikuti huruf yang berbeda menunjukkan beda nyata pada
taraf α=0.05 berdasarkan uji jarak berganda Duncan.
Respon Pertumbuhan Biomassa B. bassiana pada Media Cair secara In Vitro Biomassa B. bassiana yang diinkubasi pada media PDB, MEB dan SDBY selama 4 minggu secara berturut-turut adalah 0.18, 0.19, dan 0.24 g. Sehingga dapat diketahui bahwa biomassa tertinggi diperoleh isolat yang diinkubasi pada media SDBY, kemudian isolat pada media MEB, dan biomassa yang terendah diperoleh isolat yang diinkubasi pada media PDB ( Gambar 11).
Gambar 11 Biomassa B. bassiana pada media potato dextrose broth (PDB), malt extract broth (MEB), dan sabouraud dextrose broth with yeast extract (SDBY) selama 4 minggu
18
bahwa, terdapat korelasi antara nilai produksi biomassa B. bassiana pada media cair SDBY dengan pertumbuhan radial koloni B. bassiana pada media padat SDAY.
Media cair SDBY menghasilkan biomassa B. bassiana terbesar dan media padat SDAY menghasilkan pertumbuhan radial koloni tercepat. Dengan demikian, media SDBY dan SDAY merupakan media yang paling sesuai untuk produksi biomassa dan pertumbuhan radial B. bassiana. Kandungan nutrisi media tersebut menyebabkan B. bassiana menjadi optimal. Menurut Sharma (2002) dalam Senthamizhlselvan et al. (2010) bahwa, pertumbuhan optimal B. bassiana pada medium cair sabouraud diduga karena kandungan pepton sebagai sumber nitrogen.
Kandungan Nutrisi pada Media Tumbuh
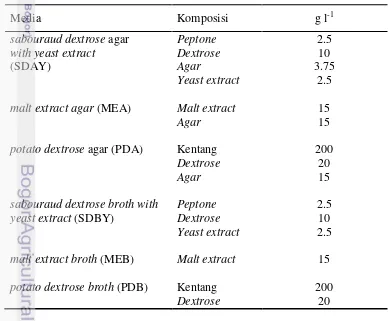
Cendawan adalah organisme heterotrof, sehingga mereka memerlukan senyawa organik sebagai sumber nutrisi. Pertumbuhan fungi melibatkan banyak faktor yang menentukan seperti faktor organisme itu sendiri dan lingkungan (Griffin 1991). Media kultur dapat dijadikan sebagai sarana untuk mengukur pertumbuhan fungi berdasarkan fungsi waktu baik pada media kultur padat maupun media kultur cair (Chang dan Miles 2004). Komposisi masing-masing media tumbuh disajikan pada Tabel 4.
Tabel 4 Komposisi nutrisi yang terdapat pada masing-masing media tumbuh
Media Komposisi g l-1
malt extract agar (MEA) Malt extract Agar
15 15
potato dextrose agar (PDA) Kentang Dextrose
malt extract broth (MEB) Malt extract 15
potato dextrosebroth (PDB) Kentang Dextrose
19
Menurut Chang dan Miles (2004), sumber karbon merupakan unsur yang penting bagi pembentukan struktur dan sumber energi pertumbuhan sel cendawan. Ketiga jenis media tumbuh yang digunakan dalam penelitian ini (potato dextrose, malt extract, dan sabouraud dextrosewith yeast extract) memiliki sumber nutrisi yang berbeda.
Potato dextrose terdiri dari ekstrak kentang yang merupakan sumber karbohidrat, vitamin dan mineral dengan tambahan dextrose yang juga merupakan sumber karbohidrat. Kentang tersusun atas pati dengan kandungan amilosa dan amilopektin. Amilosa dan amilopektin termasuk molekul kompleks karbohidrat golongan polisakarida. Proses penguraian karbohidrat kompleks golongan polisakarida tidak semudah pada molekul karbohidrat yang lebih sederhana seperti halnya glukosa yang tergolong ke dalam monosakarida. Dengan demikian cendawan membutuhkan waktu untuk menguraikan senyawa karbohidrat yang terkandung dalam kentang. Akan tetapi cendawan mendapat tambahan dextrose yang merupakan molekul karbohidrat sederhana dan mudah diurai, sehingga aktivitas seluler dapat berlangsung dari energi yang dihasilkan dari penguraian dextrose terlebih dahulu.
Malt extract mengandung sumber karbon maltosa yang tinggi (Vanderzant dan Splittstoesser 1992 dalam Syafiih 2015). Maltosa merupakan kelompok karbohidrat disakarida yang tersusun atas 2 molekul glukosa. Pertumbuhan miselium B. bassiana lebih lambat pada media malt extract dibandingkan dengan media potato dextrose diduga karena struktur maltosa yang merupakan disakarida lebih kompleks dibandingkan dengan dextrose. Cendawan membutuhkan waktu lebih lama dalam menyederhanakan struktur malt dibandingkan dengan struktur dextrose, sehingga energi yang dihasilkan pada media malt lebih lambat dibandingkan dengan media potato dextrose.
Media sabouraud dextrose with yeast extract tersusun atas pepton dan yeast extract yang merupakan sumber nitrogen serta dextrose sebagai sumber karbon. Nitrogen berperan dalam pembentukan molekul protein dan asam nukleat. Salah satu fungsi nitrogen adalah sebagai sumber energi, sehingga isolat B. bassiana yang ditumbuhkan pada media yang lebih banyak mengandung molekul nitrogen memiliki sumber energi lebih besar untuk membentuk sel-sel baru. Selain itu, karbon dalam dextrose merupakan unsur penting sebagai penyusun komponen dinding sel.
20
Keefektifan B. bassiana terhadap rayap C. curvignathus
Tabel 5 menunjukkan bahwa, mortalitas tertinggi rayap C. curvignathus disebabkan oleh suspensi konidia B. bassiana dari media MEB (44.4%), kemudian suspensi konidia dari media PDB (40.4%) dan SDBY (21.2%) pada
tingkat kerapatan konidia yang sama yaitu 108 konidia ml-1. Mortalitas C. curvignathus mengalami peningkatan seiring dengan meningkatnya kerapatan
konidia B. bassiana.
Tabel 5 Persentase mortalitas kumulatif C. curvignathus yang terinfeksi cendawan B. bassiana dari tiga jenis media pada berbagai tingkat kerapatan konidia selama 11 hari
Pengamatan terhadap rayap uji menunjukkan bahwa kematian rayap dicirikan dengan gejala bertahap. Gejala awal menunjukkan kondisi rayap terinfeksi semakin melemah, dicirikan dengan mobilitas rayap terinfeksi tidak gesit seperti halnya pada rayap sehat. Selanjutnya, sebagian rayap yang lemah dan sakit tersebut dimangsa oleh rayap sehat. Nandika et al. (2003) menyatakan bahwa, salah satu sifat rayap adalah kanibalisme, yaitu sifat memangsa sesama rayap terutama yang lemah dan sakit. Sebagian rayap terinfeksi mengalami melanisasi kutikula yang dicirikan dengan perubahan warna kutikula menjadi hitam dan keras, kemudian setelah beberapa hari tubuh rayap ditumbuhi miselium yang dapat menyelimuti seluruh tubuh rayap. Adanya bercak hitam pada kutikula menunjukkan lokasi terjadinya penetrasi (Boucias dan Pendland 1998).
Aplikasi suspensi B. bassiana pada berbagai tingkat kerapatan konidia mengakibatkan mortalitas C. curvignathus yang terus meningkat setiap hari. Gambar 12 menunjukkan mortalitas tertinggi pada tiap jenis media disebabkan oleh suspensi B. bassiana dengan kerapatan 108 konidia ml-1, kemudian kerapatan 106 dan 104 konidia ml-1.
Kerapatan konidia (konidia ml-1)
21
Gambar 12 Laju mortalitas C. curvignathus akibat aplikasi B. bassiana dari media tumbuh potato dextrose broth (A), malt extract broth (B), dan sabouraud dextrose with yeast extract (C) pada tingkat kerapatan konidia kontrol, 104, 106, dan 108 konidia ml-1 selama 11 hari pengamatan
Respon mortalitas rayap C. curvignathus akibat aplikasi B. bassiana pada berbagai tingkat kerapatan konidia menunjukkan bahwa, perbedaan jenis media tidak mempengaruhi virulensi suspensi konidia B. bassiana terhadap mortalitas
22
rayap, namun perbedaan kerapatan konidia berpengaruh terhadap mortalitas rayap. Semakin tinggi kerapatan konidia maka semakin tinggi mortalitas rayap. Mortalitas rayap pada kerapatan 108 konidia ml-1 tertinggi dibandingkan dengan kerapatan konidia 106 dan 104 konidia ml-1 serta kontrol (Tabel 6).
Tabel 6 Respon mortalitas rayap C. curvignathus akibat aplikasi B. bassiana pada berbagai tingkat kerapatan konidia
Kerapatan konidia (konidia mL-1) Rataan mortalitas (%)*
108 28.94a
106 19.54b
104 16.72b
Kontrol 4.03c
*Nilai mortalitas yang diikuti huruf yang berbeda menunjukkan perbedaan nyata
pada taraf α=0.05 berdasarkan uji jarak berganda Duncan.
Desyanti (2007) menyatakan bahwa, semakin tinggi kerapatan konidia yang diaplikasikan terhadap rayap menyebabkan konidia lebih banyak melakukan kontak dengan tubuh rayap, sehingga memberi peluang lebih besar bagi konidia untuk berkecambah dan mempenetrasi tubuh rayap. Boucias dan Pendland (1998) menyatakan bahwa, semakin banyak kontak antara cendawan dengan serangga, maka proses kematian larva terinfeksi akan semakin cepat.
5 SIMPULAN DAN SARAN
Simpulan
Media tumbuh padat maupun cair yang paling sesuai untuk pertumbuhan diameter koloni dan biomassa isolat B. bassiana adalah media yang mengandung pepton yaitu media padat SDAY dan media cair SDBY.
Mortalitas rayap meningkat seiring meningkatnya kerapatan konidia. Aplikasi suspensi B. bassiana dengan kerapatan 108 konidia ml-1 paling efektif menyebabkan mortalitas rayap, sedangkan ketiga jenis sumber nutrisi (PDB, MEB dan SDBY) tidak efektif mempengaruhi virulensi isolat B. bassiana terhadap mortalitas rayap C. curvignathus.
Saran
DAFTAR PUSTAKA
Alves SB, Rossi LS, Lopes RB, Tamai MA, Pereira RM. 2002. Beauveria bassiana Yeast Phase on Agar Medium and Its Pathogenicity against Diatraea saccharalis (Lepidoptera:Crambidae) and Tetranychus urticae (Acari:Tetranychidae). Journal of Invertebrate Pathology. 81(2) : 70-77 Barron GL. 2001. Beauveria bassiana [Internet]. [Diunduh 12 April 2013].
Tersedia pada : http://www.uoguelph.ca/~gbarron/MISCELLANEOUS/ nov01.htm
Berman JJ. 2012. Taxonomic Guide to Infectious Diseases: Understanding The Biologic Classes of Pathogenic Organisms. Waltham: Academic Press Boucias DG, Pendland JC. 1998. Principle of Insect Pathology. Kluwer Academic
Publisher. London
Chang ST, Miles PG. 2004. Mushroom: Cultivation, Nutritional Value, Medicinal Effect and Environmental Impact. 2 nd Ed. Florida (US): CRC Press LLC Desyanti. 2007. Kajian Pengendalian Rayap Tanah Coptotermes spp. (Isoptera:
Rhinotermitidae) Dengan Menggunakan Cendawan Entomopatogen Isolat Lokal [Disertasi]. Bogor(ID): Institut Pertanian Bogor
Eggleton P. 2011. An Introduction to Termites: Biology, Taxonomy and Functional Morphology. In Biology of Termites: A Modern Synthesis. Bignell DE, Roisin Y, Nathan Lo, editor. London: Springer Dordrecht Heidelberg
Erningtyas T. 2006. Efikasi Nematoda Entomopatogen Heterorhabditis sp. dan Steinernema sp. Isolat Bogor Sebagai Bioinsektisida terhadap Rayap Tanah Coptotermes curvignathus Holmgren (Isoptera : Rhinotermitidae) [Tesis]. Bogor(ID): Institut Pertanian Bogor
Faria M, Joseph HH, Stephen PW. 2012. Application of Modified Atmosphere Packaging (Gas Flushing And Active Packaging) for Extending The Shelf Life of Beauveria bassiana Conidia at High Temperatures. Biological Control 61(1):78-88
Gao L, Xingzhong L. 2010. Nutritional Requirements of Mycelial Growth and Sporulation of Several Biocontrol Fungi in Submerged and Solid Culture. Mikrobiologiia 79(5):622-9
Georghio GP. 1986. The Magnitude of The Resistance Problem. In Pesticide Resistance: Strategies and Tactics for Management. Washington DC: National Academy Press
Griffin DH. 1991. Fungal Physiology. New York (US): J Willey
Hashim N. 2000. Bioefficacy and Infection Mechanism of Several Entomopathogenic Fungi on Crocidolomia Binotalis Zell. [Thesis]. Universiti Putra Malaysia
Herliyana EN. 2007. Potensi Ligninolitik Jamur Pelapuk Kayu Kelompok Pleurotus [Disertasi]. Bogor (ID): Institut Pertanian Bogor
Hill DS. 2008. Pests of Crops in Warmer Climates and Their Control. New York: Springer
24
Mehta J, Madhulika J, Sudha C, Jishan M, Dilip S, Pawan K, Priyanka G, Meenu M N. 2012. Impact of Carbon and Nitrogen Sources on The Trichoderma viridae (Biofungicide) and Beauveria bassiana (entomopathogenic fungi). European Journal of Experimental Biology 2(6):2061-2067
Nair KSS. 2001. Pest Outbreaks in Tropical Forest Plantations: Is There a Greater Risk for Exotic Tree Species?. Bogor (ID): Center for International Forestry Research
Nandika D, Tambunan B. 1990. Biodeteriorasi Kayu oleh Faktor Biologis. Bogor: Pusat Antar Universitas Bioteknologi Institut Pertanian Bogor
Nandika D, Yudi R, Farah D. 2003. Rayap : Biologi dan Pengendaliannya. Harun JP, ed. Surakarta : Muhammadiyah University Press
Neves PMOJ, Alves SB. 2004. External Events Related to The Infection Process of Cornitermes cumulans (Kollar) (Isoptera: Termitidae) by The Entomopathogenic Fungi Beauveria bassiana and Metarhizium anisopliae. Neotropical Entomology 33(1):051-056
Pribadi A. 2010. Serangan Hama dan Tingkat Kerusakan Daun Akibat Hama Defoliator pada Tegakan Jabon (Anthocephalus cadamba Miq.). Jurnal Penelitian Hutan dan Konservasi Alam 7(4):451-458. Riau (ID): Balai Penelitian Teknologi Serat Tanaman Hutan Kuok
Purnomo H. 2010. Pengantar Pengendalian Hayati. Yogyakarta: Andi Offset Sahayaraj K, Namasivayam SKR. 2008. Mass Production of Entomopathogenic
Fungi Using Agricultural Products and By Products. African Journal of Biotechnology 7 (12):1907-1910
Santoro PH, Janaina Z, Kelly C, Pedro MOJN. 2014. Conidial Production, Virulence, and Stress Tolerance of Beauveria bassiana Conidia After Successive In Vitro Subculturing. Revista Colombiana de Entomología 40(1):85-90
Senthamizhlselvan P, Sujeetha JARP, Jeyalakshmi C. 2010. Growth, Sporulation and Biomass Production of Native Entomopathogenis Fungal Isolates on A Suitable Medium. Journal of Biopesticides 3(2):466-469
Soetopo D, Igaa I. 2007. Status Teknologi dan Prospek Beauveria bassiana Untuk Pengendalian Serangga Hama Tanaman Perkebunan yang Ramah Lingkungan. Perspektif 6:29-46
Sudiaman M. 2015. Peneliti IPB: Kerugian Akibat Rayap di Indonesia Capai Rp 2,8 Triliun. Republika Online. 13 Maret 2015. [Diakses 22 April 2015]. Tersedia pada :http://www.republika.co.id/berita/nasional/umum/15/03/13/ nl5fq0-peneliti-ipb-kerugian-akibat-rayap-di-indonesia-capai-rp-28-triliun Sudibyo D. 1994. Petunjuk Praktis Cara Menghitung Jumlah, Kerapatan dan
Viabilitas Spora Jamur. Laboraturium Utama Pengendalian Hayati Dinas Perkebunan Propinsi Jawa Timur
Sukartana P, Rushelia R, Rumini W. 2000. Pathogenicity of Two Species of Entomopathogenic Agents to Subterranean Termite Coptotermes gestroi Wasmann (Isoptera:Rhinotermitidae) in Laboratory. Buletin Hama dan Penyakit Tumbuhan 12(1):17-20
25
27
La
mpi
ra
n
1 P
rose
s P
eng
uji
an Respon P
ertumbuha
n Radial K
29
Lampiran 3 Hasil analisis sidik ragam dan uji selang berganda Duncan uji in vitro pengaruh media kultur terhadap pertumbuhan isolat B. bassiana
1. Pertumbuhan isolat pada media padat
General Linier Model Procedure
Duncan's Multiple Range Test for Pertumbuhan Diameter
Duncan Grouping Mean N MEDIA A 2.85042 60 SDAY B 2.59125 60 PDA C 2.34125 60 MEA
2. Pertumbuhan isolat pada media cair
30
Source DF Sum of Squares Mean Square F Value Pr > F Model 2 0.01357143 0.00678571 7.90 0.0034 Error 18 0.01545714 0.00085873
Corrected Total 20 0.02902857
R-Square Coeff Var Root MSE DM Mean 0.467520 14.14681 0.029304 0.207143
Source DF Sum of Squares Mean Square F Value Pr > F media 2 0 0.01357143 0.00678571 7.90 0.0034
Duncan's Multiple Range Test for Biomassa
Duncan Grouping Mean N MEDIA A 0.24286 7 SDBY B 0.19286 7 MEB B
B 0.18571 7 PDB
Lampiran 4 Rekapitulasi sidik ragam jenis media cair dan tingkat kerapatan konidia terhadap mortalitas rayap uji
Sumber DB KT Fhitung Pr > F
Jenis Media 2 138.301111 2.29 0.1227
Tingkat Kerapatan 3 950.905185 15.76 < 0.0001
Interaksi 6 135.777407 2.25 0.0728
RIWAYAT HIDUP
Penulis dilahirkan pada tanggal 30 Mei 1986 di Jakarta, sebagai anak kedua dari tiga bersaudara dari pasangan Bapak Abdurrahim Idris dan Ibu Endah Listyarini. Penulis telah berkeluarga dengan suami Febrie Subhan, S.Komp dan dikarunia 1 orang anak, yaitu Sofia Zahidah (1 tahun).
Penulis menyelesaikan sekolah tingkat menengah di SMUN 1 Bekasi pada tahun 2004 dan melanjutkan ke jenjang S1 di Departemen Hasil Hutan, Fakultas Kehutanan IPB melalui jalur SPMB dan lulus pada tahun 2010. Pada tahun 2011, penulis melanjutkan studi jenjang S2 Sekolah Pascasarjana IPB pada Program Studi Silvikultur Tropika.