INDANA SARAMITA RACHMAN. Potensi
Bacillus
sp. Galur G3,
Bacillus firmus
E65, dan
Bakteri Metanotrof sebagai Penghambat Pertumbuhan Patogen
Xanthomonas oryzae
pv.
oryzae
dan
Rhizoctonia solani
. Dibimbing oleh IMAN RUSMANA dan ALINA AKHDIYA.
Penggunaan mikroba dari genus
Bacillus
dan bakteri metanotrof sebagai agen
biokontrol untuk mengendalikan bakteri patogen tanaman padi
X. oryzae
pv
.
oryzae dan
R. solani
dapat meningkatkan produktifitas padi serta mengurangi emisi metan di lahan
sawah. Sebagian dari bakteri metanotrof dapat melakukan fiksasi nitrogen. Uji
antagonisme dilakukan untuk mengetahui kemampuan dua jenis
Bacillus
(
Bacillus
sp.
galur G3 dan
B. firmus
E65) serta dua bakteri metanotrof (BGM 9 dan SKM 14) dalam
menghambat pertumbuhan
X. oryzae
pv.
oryzae
dan
R. solani
serta mengetahui adanya
penghambatan pertumbuhan di antara kedua kelompok bakteri tersebut. Bakteri
metanotrof
(BGM 9
dan
SKM 14) dapat menghambat pertumbuhan
X. oryzae
pv.
oryzae
dengan nilai indeks penghambatan berturut-turut 2,79 dan 2,27.
Bacillus
sp. galur G3,
B.
firmus
E65 dan bakteri metanotrof SKM 14 dapat menghambat pertumbuhan
R. solani
dengan indeks penghambatan pertumbuhan radial berturut-turut 0.63, 0.64, dan 0.56.
B. firmus
E65 dapat menghambat pertumbuhan bakteri metanotrof (BGM 9 dan SKM 14)
dan
R. solani
. Formulasi agen biokontrol yang mengandung
Bacillus
sp. Galur G3, dan
kedua bakteri metanotrof (SKM 14
dan BGM 9) dapat dilakukan karena tidak ada
antagonisme diantara ketiga bakteri tersebut. Sedangkan kombinasi
B. firmus
E65
dengan kedua metanotrof sebaiknya tidak dilakukan dalam satu formulasi karena
adanya antagonisme antara
B. firmus
E65 dengan kedua bakteri metanotrof tersebut.
Kata kunci: Bakteri metanotrof,
Bacillus firmus
E65,
Bacillus
sp. galur G3,
Rhizoctonia
solani, Xanthomonas oryzae
pv.
oryzae.
INDANA SARAMITA RACHMAN. Potency of
Bacillus
sp. G3,
Bacillus firmus
E65, and
Methanotrophs as Growth Inhibitor of Pathogens
Xanthomonas oryzae
pv.
oryzae
and
Rhizoctonia solani
. Under supervision of IMAN RUSMANA and ALINA AKHDIYA.
The use of bacilli and methanotropic bacteria as biocontrol agents for rice bacterial
pathogens of
X. oryzae
pv
.
oryzae and
R. solani
can enhance rice production and reduce
methane emission from rice fields. Some of methanotrophs had nitrogen fixation activity.
Antagonism methods was tested to determine the ability of two bacilli species (
Bacillus
sp. G3
and
B. firmus
E65) and two methanotrophs (BGM 9 and SKM 14
)
in inhibiting
growth of
X. oryzae
pv.
oryzae
and
R. solani
and inhibition between the two groups of
bacteria. Methanotrophs (BGM 9
and SKM 14) inhibited growth of
X. oryzae
pv.
oryzae
with inhibition index (IP) of 2,27 and 2,79 respectively.
Bacillus
sp. G3,
B. firmus
E65 and
SKM 14
isolates inhibited growth of
R. solani
with inhibition index of 0.63, 0.64, and 0.56
respectively.
B. firmus
E65
inhibited growth of methanotrophs (BGM 9 and SKM 14)
and
R. solani
. Formulation of biocontrol agents
containing
Bacillus
sp. G3
and methanotrophs
(BGM 9 and SKM 14)
can be done because there was no antagonism among the three
bacteria. However the combination of
B. firmus
E65
and the metanotrophs should not be
done in one formulation because of the antagonism between
B. firmus
E65 and the
metanotrophs.
PENDAHULUAN
Latar belakangIndonesia memiliki biodiversitas mikrob yang tinggi termasuk di antaranya berbagai jenis patogen tumbuhan, mikrob yang bersifat antagonis terhadap patogen, penambat nitrogen, dan bakteri metanotrof. Xanthomonas oryzae pv. oryzae merupakan bakteri penyebab penyakit hawar daun bakteri padi, sedangkan Rhizoctonia solani ialah patogen penyebab hawar daun, hawar pelepah, dan rebah semai pada padi. Kedua patogen tersebut tersebar luas di Indonesia dan sulit dikendalikan sehingga sangat merugikan. Pada tahun 2006 luas penularan penyakit hawar daun bakteri mencapai lebih dari 74.000 ha. Angka ini lebih tinggi dibanding kan dengan luas penularan pada tahun 2005 yang baru mencapai 33.800 ha (Kadir 2009).
Pengendalian patogen menggunakan pestisida kimiawi secara berlebihan dalam jangka panjang berbahaya bagi manusia dan lingkungan. Oleh karena itu, perlu dikembangkan teknik dan penyalur pengendali patogen yang efektif dan ramah lingkungan. Salah satu di antaranya ialah memanfaatkan potensi mikrob antagonis sebagai bahan aktif biopestisida.
Bacillus merupakan salah satu genus bakteri yang sebagian anggotanya diketahui mampu memproduksi senyawa antimikrob yang dapat membunuh atau menghambat pertumbuhan mikroorganisme lain. Selain itu, menurut Compant et al. (2005) genus bakteri ini juga dapat menginduksi sistem kekebalan tanaman.
Metanotrof merupakan kelompok bakteri yang dapat menggunakan metan sebagai sumber karbon dan sumber eneginya (Hanson R & Hanson T 1996). Sebagian dari bakteri ini juga dapat melakukan fiksasi nitrogen (Dedysh et al. 2004). Kemampuan tersebut menjadikan bakteri metanotrof berpotensi untuk dikembangkan sebagai penyalur pereduksi emisi metan dan komponen pupuk hayati untuk meningkatkan pertumbuhan tanaman padi di lahan sawah. Berbeda dengan Bacillus, sampai saat ini belum ditemukan laporan penelitian tentang kemampuan bakteri metanotrof dalam menghambat atau mengendalikan mikrob patogen.
Salah satu cara untuk meningkatkan spektrum dan efektivitas suatu formulasi penyalur biokontrol patogen tanaman ialah dengan menggabungkan berbagai mikrob
dalam satu formulasi. Penggunaan beberapa mikrob tersebut dalam satu formulasi dapat berfungsi sebagai penyalur pengendali penyakit dan juga dapat meningkatkan pertumbuhan tanaman. Kombinasi campuran mikrob dalam satu formulasi tidak boleh ada satu organisme yang dapat menghambat atau mematikan organisme lainnya yang merupakan komponen dari formulasi tersebut. Oleh karena itu, kajian terhadap potensi masing-masing mikrob, sinergi serta antagonisme diantara mikrob tersebut merupakan salah satu tahap penting yang harus dilakukan sebelum menggabungkan berbagai mikrob dalam satu formulasi.
Tujuan
Penelitian ini bertujuan mengamati kemampuan dua jenis Bacillus (Bacillus sp. galur G3 dan Bacillus firmus E65) serta dua bakteri metanotrof (BGM 9 dan SKM 14) untuk menghambat pertumbuhan Xabthomonas oryzae pv. oryzae dan Rhizoctonia solani serta mengetahui adanya penghambatan pertumbuhan diantara kedua kelompok bakteri tersebut.
BAHAN DAN METODE
BahanBakteri yang digunakan dalam penelitian ini ialah Bacillus sp. galur G3 berasal dari Laboratorium Mikrobiologi Departemen Biologi FMIPA IPB, B. firmus E65 dari koleksi BB - BIOGEN, dua isolat bakteri metanotrof dari wilayah Bogor dan Sukabumi (BGM 9 dan SKM 14), dan dua jenis patogen (X. oryzae pv. oryzae dan R. solani) dari koleksi BB - BIOGEN.
Metode
Peremajaan Bakteri
1
PENDAHULUAN
Latar belakangIndonesia memiliki biodiversitas mikrob yang tinggi termasuk di antaranya berbagai jenis patogen tumbuhan, mikrob yang bersifat antagonis terhadap patogen, penambat nitrogen, dan bakteri metanotrof. Xanthomonas oryzae pv. oryzae merupakan bakteri penyebab penyakit hawar daun bakteri padi, sedangkan Rhizoctonia solani ialah patogen penyebab hawar daun, hawar pelepah, dan rebah semai pada padi. Kedua patogen tersebut tersebar luas di Indonesia dan sulit dikendalikan sehingga sangat merugikan. Pada tahun 2006 luas penularan penyakit hawar daun bakteri mencapai lebih dari 74.000 ha. Angka ini lebih tinggi dibanding kan dengan luas penularan pada tahun 2005 yang baru mencapai 33.800 ha (Kadir 2009).
Pengendalian patogen menggunakan pestisida kimiawi secara berlebihan dalam jangka panjang berbahaya bagi manusia dan lingkungan. Oleh karena itu, perlu dikembangkan teknik dan penyalur pengendali patogen yang efektif dan ramah lingkungan. Salah satu di antaranya ialah memanfaatkan potensi mikrob antagonis sebagai bahan aktif biopestisida.
Bacillus merupakan salah satu genus bakteri yang sebagian anggotanya diketahui mampu memproduksi senyawa antimikrob yang dapat membunuh atau menghambat pertumbuhan mikroorganisme lain. Selain itu, menurut Compant et al. (2005) genus bakteri ini juga dapat menginduksi sistem kekebalan tanaman.
Metanotrof merupakan kelompok bakteri yang dapat menggunakan metan sebagai sumber karbon dan sumber eneginya (Hanson R & Hanson T 1996). Sebagian dari bakteri ini juga dapat melakukan fiksasi nitrogen (Dedysh et al. 2004). Kemampuan tersebut menjadikan bakteri metanotrof berpotensi untuk dikembangkan sebagai penyalur pereduksi emisi metan dan komponen pupuk hayati untuk meningkatkan pertumbuhan tanaman padi di lahan sawah. Berbeda dengan Bacillus, sampai saat ini belum ditemukan laporan penelitian tentang kemampuan bakteri metanotrof dalam menghambat atau mengendalikan mikrob patogen.
Salah satu cara untuk meningkatkan spektrum dan efektivitas suatu formulasi penyalur biokontrol patogen tanaman ialah dengan menggabungkan berbagai mikrob
dalam satu formulasi. Penggunaan beberapa mikrob tersebut dalam satu formulasi dapat berfungsi sebagai penyalur pengendali penyakit dan juga dapat meningkatkan pertumbuhan tanaman. Kombinasi campuran mikrob dalam satu formulasi tidak boleh ada satu organisme yang dapat menghambat atau mematikan organisme lainnya yang merupakan komponen dari formulasi tersebut. Oleh karena itu, kajian terhadap potensi masing-masing mikrob, sinergi serta antagonisme diantara mikrob tersebut merupakan salah satu tahap penting yang harus dilakukan sebelum menggabungkan berbagai mikrob dalam satu formulasi.
Tujuan
Penelitian ini bertujuan mengamati kemampuan dua jenis Bacillus (Bacillus sp. galur G3 dan Bacillus firmus E65) serta dua bakteri metanotrof (BGM 9 dan SKM 14) untuk menghambat pertumbuhan Xabthomonas oryzae pv. oryzae dan Rhizoctonia solani serta mengetahui adanya penghambatan pertumbuhan diantara kedua kelompok bakteri tersebut.
BAHAN DAN METODE
BahanBakteri yang digunakan dalam penelitian ini ialah Bacillus sp. galur G3 berasal dari Laboratorium Mikrobiologi Departemen Biologi FMIPA IPB, B. firmus E65 dari koleksi BB - BIOGEN, dua isolat bakteri metanotrof dari wilayah Bogor dan Sukabumi (BGM 9 dan SKM 14), dan dua jenis patogen (X. oryzae pv. oryzae dan R. solani) dari koleksi BB - BIOGEN.
Metode
Peremajaan Bakteri
Uji Antagonisme Bacillus sp. Galur G3, B. firmus E65 dan Bakteri Metanotrof terhadap X. oryzae pv. oryzae
Antagonisme Bacillus sp. galur G3, B. firmus E65, SKM 14, dan BGM 9 terhadap X. oryzae pv. oryzae diuji secara in vitro dengan metode agar dua lapis (Lisboa et al. 2006). Keempat bakteri tersebut masing - masing ditumbuhkan pada media cair sambil dicampur dengan kecepatan 80 putaran per menit pada suhu ruang (29°C). Isolat SKM 14 dan BGM 9 diinkubasi selama sepuluh hari, sedangkan isolat yang lainnya diinkubasi selama 3 hari. Sebanyak 1 ml kultur cair X. oryzae pv. oryzae berumur 3 hari diinokulasikan ke dalam 100 ml Trypticase Soy Agar semi padat lalu dituangkan pada permukaan cawan Trypticase Soy Agar padat masing - masing sebanyak 10 ml. Setelah media Trypticase Soy Agar dua lapis memadat, kultur Bacillus sp. Galur G3 dan B. firmus E65 yang berumur 2 hari ditotolkan diatasnya dengan menggunakan tusuk gigi steril lalu diinkubasi pada suhu 30ºC. Pengamatan dan pengukuran zona hambat dilakukan setelah 48 jam untuk uji terhadap Bacillus sp. galur G3 dan B. firmus E65. Uji antagonisme untuk isolat BGM 9 dan SKM 14 dilakukan pada media Nitrat Mineral Salts padat + 1 % metanol yang di lapis dengan media Trypticase Soy Agar semi padat dan telah diinokulasikan dengan X. oryzae pv. oryzae di bagian atasnya. Pengamatan hasil uji antagonis oleh kedua bakteri metanotrof tersebut dilakukan setelah 6 hari inkubasi. Indeks penghambatan dihitung dengan persamaan berikut:
Keterangan:
IP = Indeks Penghambatan Ø zona bening = diameter zona hambat (cm) Ø koloni = diameter koloni (cm)
Uji Antagonisme Bacillus sp. Galur G3 dan
B. firmus E65 terhadap Bakteri Metanotrof (BGM 9 dan SKM 14)
Bacillus sp. galur G3 dan B. firmus E65 diuji antagonismenya terhadap bakteri metanotrof (BGM 9 dan SKM 14) pada media campuran agar Nitrat Mineral Salts + metanol 1 % dan Agar Nutrien (1:1) dengan menggunakan metode cawan sebar. Dua lup inokulum BGM 9 dan SKM 14 masing-masing disuspensikan ke dalam 4 ml air steril lalu dihomogenkan menggunakan vortex. Sebanyak 100 µl suspensi disebar ke atas
cawan berisi media campuran yang telah padat lalu diinkubasi selama 4 hari. Selanjutnya di bagian tengah permukaan media tersebut masing - masing diinokulasi dengan kultur Bacillus sp. galur G3 dan. B. firmus E65 berumur 48 jam menggunakan tusuk gigi steril. Cawan - cawan tersebut kemudian diinkubasi lagi selama 3 hari pada suhu ruang (29°C). Zona hambat di sekitar koloni Bacillus diukur untuk menentukan nilai indeks penghambatannya menggunakan persamaan di atas.
Uji Antagonisme Bacillus sp. Galur G3, B. firmus E65, dan Bakteri Metanotrof (SKM 14 dan BGM9) terhadap R. solani pada Media Padat
Bacillus sp. galur G3 dan B. firmus E65 masing -masing digores lurus sepanjang 3 cm berhadapan dengan inokulum R. solani (d = 0,8 cm) yang diletakkan di tengah cawan. Jarak antara R. solani dan bakteri uji sejauh 2 cm (Anjaiah et.al 1998). Sedangkan isolat BGM 9 dan SKM 14, diuji antagonismenya terhadap R. solani pada media campuran Potato Dextrose Agar dan Nitrat Mineral Salts Padat + metanol 1 % (1:1). BGM 9 dan SKM 14 digores 3 hari lebih awal dari inokulasi R. solani. Cawan - cawan uji untuk Bacillus sp. galur G3 dan B. firmus E65 kemudian diinkubasi pada suhu ruang selama 3 hari, sedangkan untuk bakteri metanotrof diinkubasi selama 5 hari. Penghambatan pertumbuhan radial cendawan oleh bakteri dihitung dengan rumus berikut ini (Dikin et al. 2006):
% Penghambatan Pertumbuhan Radial = 1 – (a/ b)
Keterangan:
a = Jarak antara miselia terluar dengan blok inokulasi pada sisi bakteri.
b = Jarak antara miselia terluar dengan blok inokulasi pada sisi berlawanan (tanpa bakteri).
Uji Antagonisme Bacillus sp. Galur G3, B. firmus E65, dan Isolat Metanotrof SKM 14 terhadap R. solani pada Media Cair
3
ditumbuhkan pada media Nitrat Mineral Salts cair + metanol 1 % selama 14 hari sambil dicampur pada kecepatan yang sama. Masing- masing kultur diambil sebanyak 3 ml dan dibagi ke dalam tiga tabung eppendorf masing-masing 1 ml. Sisa kultur masing - masing disentrifugasi dan supernatannya diambil 3 ml lalu dibagi ke dalam 3 tabung eppendorf masing-masing 1 ml. Kemudian dalam masing - masing eppendorf tadi dimasukkan dua butir sklerotium R. solani yang telah ditimbang, dan dimasukkan ke dalam masing - masing eppendorf lalu diinkubasi selama 15 hari. Sebagai pembanding, sklerotium ditumbuhkan dalam medium Nutrien Cair dan Nitrat Mineral Salts + metanol 1 %. Setelah diinkubasi selama 15 hari, pertumbuhan miselium pada masing - masing perlakuan diamati dan ditimbang berat kering biomassanya.
HASIL
Antagonisme Bacillus sp. Galur G3, B. firmus E65, dan Bakteri Metanotrof terhadap X. oryzae pv. oryzae
Hasil uji antagonisme menunjukkan isolat BGM 9 dan SKM 14 dapat menghambat pertumbuhan X. oryzae pv. oryzae (Tabel 1) dengan indeks penghambatan berturut-turut 2,79 dan 2,27. Sedangkan Bacillus sp. galur G3 dan B. firmus E65 tidak dapat menghambat pertumbuhan X. oryzae pv. oryzae.
Tabel 1 Penghambatan pertumbuhan X. oryzae pv. oryzae oleh Bacillus sp. galur G3, B. firmus E65, dan bakteri metanotrof
Isolat Ø koloni (cm)
Ø zona bening (cm)
IP ± SE
G3 - - -
E65 - - -
BGM 9 0,35 1,33 2,79 ± 0,36 SKM 14 0,43 1,40 2,27 ± 0,40 Keterangan: IP = Indeks Penghambatan; SE =
Standard Error (nilai rataan ± SE, n = 2); (-) = Tidak ada penghambatan.
Antagonisme Bacillus sp. Galur G3 dan B. firmus E65 terhadap Bakteri Metanotrof (BGM 9 dan SKM 14)
Informasi yang diperoleh dari hasil uji antagonisme Bacillus sp. galur G3 terhadap bakteri metanotrof diperlukan sebagai salah satu dasar pertimbangan dalam formulasi bakteri tersebut sebagai biokontrol patogen.
Bacillus sp. galur G3 tidak dapat menghambat pertumbuhan kedua bakteri metanotrof BGM 9 dan SKM 14. Sedangkan isolat B. firmus E65 dapat menghambat pertumbuhan kedua bakteri metanotrof ini dengan nilai indeks penghambatan berturut-turut 0,75 dan 1,69 (Tabel 2).
Antagonisme Bacillus sp. Galur G3, B. firmus E65, dan Bakteri Metanotrof(SKM 14 dan BGM 9) terhadap R. solani pada Media Padat
Uji antagonisme yang dilakukan pada media padat menunjukkan B. Firmus E65, Bacillus sp. galur G3, dan SKM 14 dapat menghambat pertumbuhan R. Solani. Gambar 1 dan 2 berikut menunjukkan penghambatan pertumbuhan R. solani oleh Bacillus sp. galur G3 dan B. firmus E65 pada media Potato Dextrose Agar.
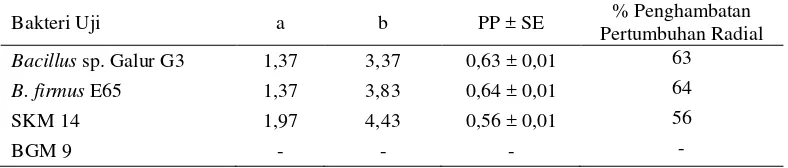
Indeks penghambatan pertumbuhan radial miselia R. solani oleh Bacillus sp. galur G3, B. firmus E65, dan SKM 14 berturut-turut sebesar 0.63, 0.64, dan 0.56 (Tabel 3).
antagonisme diantara ketiga bakteri tersebut. Sedangkan kombinasi B. firmus E65 dengan kedua metanotrof dalam satu formulasi sebaiknya tidak dilakukan karena adanya antagonisme antara B. firmus E65 dengan kedua bakteri metanotrof.
Berdasarkan masing-masing kemampuan-nya, kombinasi antara Bacillus sp. galur G3, SKM 14, dan BGM 9 berpotensi untuk dikembangkan sebagai suatu formulasi penyalur biokontrol untuk mengendalikan patogen padi R. solani dan X. oryzae pv. oryzae. Kemampuan BGM 9 dalam memfiksasi nitrogen dan SKM 14 dalam mengoksidasi metan juga dapat meningkatkan pertumbuhan tanaman padi dan mereduksi emisi metan di lahan sawah sehingga akan memberi nilai lebih dan tambahan manfaat pada formulasi tersebut.
SIMPULAN
Bakteri metanotrof (BGM 9 dan SKM 14) dapat menghambat pertumbuhan X. oryzae pv. oryzae dengan nilai indeks penghambatan berturut - turut 2,79 dan 2,27. Bacillus sp. galur G3, B. firmus E65, dan bakteri metanotrof SKM14 dapat menghambat pertumbuhan R. solani dengan indeks penghambatan pertumbuhan radial berturut - turut 0,63, 0,64, dan 0,56. B. firmus E65 dapat menghambat pertumbuhan bakteri metanotrof (BGM 9 dan SKM 14) dan R. solani.
Pengembangan penyalur biokontrol yang mengandung Bacillus sp. galur G3 dan kedua bakteri metanotrof (SKM 14 dan BGM 9) dapat dilakukan karena tidak ada antagonisme diantara ketiga bakteri tersebut. Sedangkan kombinasi B. firmus E65 dengan kedua metanotrof tersebut dalam satu formulasi sebaiknya tidak dilakukan karena adanya antagonisme antara B. firmus E65 dengan kedua bakteri metanotrof.
DAFTAR PUSTAKA
Anjaiah V, Koedam N, Nawak-Thompson B, Loper JE, Hofte M, Tambong JT, Cornelis P. 1998. Involvement of phenazines and anthranilate in the antagonism of Pseudomonas aeruginosa PNA1 and Tn5 derivattives toward Fusarium spp. and Pythium spp. Mol Plant Microbe Interact 11:847-854.
Bottone EJ, Peluso RW. 2003. Production by Bacillus pumilus (MSH) of an antifungal compound that is active against
Mucoraceae and Aspergillus species: preliminary report. J Med Microbiol 52:69-74.
Crameri R, Davies JE, Hutter R. 1986. Plasmid curing and generation of mutations induced with ethidium bromide in Streptomycetes. J Gen Microbiol 132:819-824.
Compant S, Duffy B, Nowak J, Clement C, Barka EA. 2005. Use of plant growth promoting bacteria for biocontrol of plant diseases: principles, mechanisms of action, and future prospects. Appl Environ Microbiol 71:4951-4959.
Dedysh SN, Ricke P, Liesack W. 2004. NifH and NifD phylogenies: an evolutionary basis for understanding nitrogen fixation capabilities of methanotrophic bacteria. Microbiol 150:1301-1313.
Dikin A, Sijam K, Kadir J, Seman IA. 2006. Antagonistic bacteria against Schizophllum commune FR. In Peninsular Malaysia. Biotropia 13:111-121.
Findy K. 2009. Aktifitas Penghambatan Zat Antimikrob Bacillus sp. asal Tanah terhadap X. oryzae, Pseudomonas fluorescens, dan Pseudomonas syringae. [skripsi]. Bogor: Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Pertanian Bogor.
Hanson R, Hanson TE. 1996. Metanotrophic bacteria. J Microbiol Rev 60:439-471. Hapsary W. 2008. Isolasi dan Karakterisasi
Bakteri Metanotrof Asal Sawah di Bogor dan Sukabumi. [skripsi]. Bogor: Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Pertanian Bogor.
Huang CH, Ano T, Shoda M. 1993. Nucleotide sequence and characteristics of the gene, lpa-14, responsible for biosynthesis of the lipopeptide antibiotics iturin and surfactin from Bacillus subtilis RB14. J Ferment Bioeng 76:445-450. Kadir TS. 2009. Menangkal HBD dengan
menggilir varietas. Warta Litbang Pertanian 31:1-3.
Lisboa MP, Bonatto D, Bizani D, Henriques JAP, Brandelli A. 2006. Characterization of a bacteriocin-like substance produced by Bacillus amyloliquefaciens isolated from the Brazillian atlantic forest. J Intern Microbiol 9:111-118.
3
ditumbuhkan pada media Nitrat Mineral Salts cair + metanol 1 % selama 14 hari sambil dicampur pada kecepatan yang sama. Masing- masing kultur diambil sebanyak 3 ml dan dibagi ke dalam tiga tabung eppendorf masing-masing 1 ml. Sisa kultur masing - masing disentrifugasi dan supernatannya diambil 3 ml lalu dibagi ke dalam 3 tabung eppendorf masing-masing 1 ml. Kemudian dalam masing - masing eppendorf tadi dimasukkan dua butir sklerotium R. solani yang telah ditimbang, dan dimasukkan ke dalam masing - masing eppendorf lalu diinkubasi selama 15 hari. Sebagai pembanding, sklerotium ditumbuhkan dalam medium Nutrien Cair dan Nitrat Mineral Salts + metanol 1 %. Setelah diinkubasi selama 15 hari, pertumbuhan miselium pada masing - masing perlakuan diamati dan ditimbang berat kering biomassanya.
HASIL
Antagonisme Bacillus sp. Galur G3, B. firmus E65, dan Bakteri Metanotrof terhadap X. oryzae pv. oryzae
Hasil uji antagonisme menunjukkan isolat BGM 9 dan SKM 14 dapat menghambat pertumbuhan X. oryzae pv. oryzae (Tabel 1) dengan indeks penghambatan berturut-turut 2,79 dan 2,27. Sedangkan Bacillus sp. galur G3 dan B. firmus E65 tidak dapat menghambat pertumbuhan X. oryzae pv. oryzae.
Tabel 1 Penghambatan pertumbuhan X. oryzae pv. oryzae oleh Bacillus sp. galur G3, B. firmus E65, dan bakteri metanotrof
Isolat Ø koloni (cm)
Ø zona bening (cm)
IP ± SE
G3 - - -
E65 - - -
BGM 9 0,35 1,33 2,79 ± 0,36 SKM 14 0,43 1,40 2,27 ± 0,40 Keterangan: IP = Indeks Penghambatan; SE =
Standard Error (nilai rataan ± SE, n = 2); (-) = Tidak ada penghambatan.
Antagonisme Bacillus sp. Galur G3 dan B. firmus E65 terhadap Bakteri Metanotrof (BGM 9 dan SKM 14)
Informasi yang diperoleh dari hasil uji antagonisme Bacillus sp. galur G3 terhadap bakteri metanotrof diperlukan sebagai salah satu dasar pertimbangan dalam formulasi bakteri tersebut sebagai biokontrol patogen.
Bacillus sp. galur G3 tidak dapat menghambat pertumbuhan kedua bakteri metanotrof BGM 9 dan SKM 14. Sedangkan isolat B. firmus E65 dapat menghambat pertumbuhan kedua bakteri metanotrof ini dengan nilai indeks penghambatan berturut-turut 0,75 dan 1,69 (Tabel 2).
Antagonisme Bacillus sp. Galur G3, B. firmus E65, dan Bakteri Metanotrof(SKM 14 dan BGM 9) terhadap R. solani pada Media Padat
Uji antagonisme yang dilakukan pada media padat menunjukkan B. Firmus E65, Bacillus sp. galur G3, dan SKM 14 dapat menghambat pertumbuhan R. Solani. Gambar 1 dan 2 berikut menunjukkan penghambatan pertumbuhan R. solani oleh Bacillus sp. galur G3 dan B. firmus E65 pada media Potato Dextrose Agar.
Indeks penghambatan pertumbuhan radial miselia R. solani oleh Bacillus sp. galur G3, B. firmus E65, dan SKM 14 berturut-turut sebesar 0.63, 0.64, dan 0.56 (Tabel 3).
Tabel 2 Penghambatan pertumbuhan bakteri metanotrof BGM 9 dan SKM 14 oleh Bacillus sp. galur G3 dan B. firmus E65
Isolat
BGM 9 SKM 14
Ø koloni (cm)
Ø zona
hambat (cm) IP ± SE
Ø koloni (cm)
Ø zona hambat
(cm) IP ± SE
G3 - - - -
E65 0,37 0,63 0,75 ± 0,14 0,43 1,13 1,69 ± 0,64
Keterangan: IP = Indeks Penghambatan; (-) = Tidak ada penghambatan; SE = Standard Error (nilai rataan ± SE, n = 3). Uji dilakukan pada media Nitrat Mineral Salts + 1 % metanol dan Natrium Agar.
Gambar 2 Penghambatan pertumbuhan Rhizoctonia solani oleh B. firmus E65 pada media Potato Dextrose Agar.
Antagonisme Bacillus sp. Galur G3, B. firmus E65, dan Isolat Metanotrof SKM 14 terhadap R. solani pada Media Cair
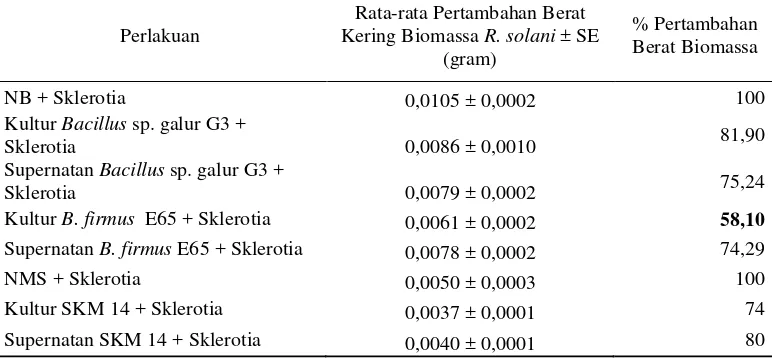
Sklerotium yang diinkubasi dalam Nutrien Cair terlihat tumbuh lebih baik dan membentuk lebih banyak miselia dibandingkan dengan sklerotia yang diinkubasi di dalam kultur B. firmus E65 dan Bacillus sp. galur G3. Data berat kering biomassa R. Solani dalam Nutrien Cair juga menunjukkan pertambahan berat biomassa yang lebih tinggi dibandingkan biomassa R. solani yang ditumbuhkan dalam kultur ataupun supernatan Bacillus sp. galur G3, B. firmus E65, dan SKM 14 maupun media Nitrat Mineral Salts cair + 1 % metanol (Tabel 4).
Data berat kering biomassa R. Solani menunjukkan bahwa inkubasi sklerotium dalam kultur B. Firmus E65 menghasilkan pertambahan berat biomassa R. Solani paling rendah sebesar 58,10% dibandingkan dengan perlakuan lainnya.
Tabel 3 Uji antagonisme bakteri dan cendawan Rhizoctonia solani
Keterangan: a = Jarak antara miselia terluar dengan koloni bakteri; b = Jarak antara miselia terluar pada sisi berlawanan (tanpa bakteri); PP = Penghambatan Pertumbuhan Radial; SE = Standard error (nilai rataan ± SE, n= 3); (-) = Tidak ada penghambatan.
Bakteri Uji a b PP ± SE % Penghambatan
Pertumbuhan Radial
Bacillus sp. Galur G3 1,37 3,37 0,63 ± 0,01 63
B. firmus E65 1,37 3,83 0,64 ± 0,01 64
SKM 14 1,97 4,43 0,56 ± 0,01 56
5
Tabel 4
Berat kering biomassa R. solani di akhir inkubasi selama 15 hari pada
masing-masing media cair (29° C)
Perlakuan
Rata-rata Pertambahan Berat Kering Biomassa R. solani ± SE
(gram)
% Pertambahan Berat Biomassa
NB + Sklerotia 0,0105 ± 0,0002 100
Kultur Bacillus sp. galur G3 +
Sklerotia 0,0086 ± 0,0010 81,90
Supernatan Bacillus sp. galur G3 +
Sklerotia 0,0079 ± 0,0002 75,24
Kultur B. firmus E65 + Sklerotia 0,0061 ± 0,0002 58,10
Supernatan B. firmus E65 + Sklerotia 0,0078 ± 0,0002 74,29
NMS + Sklerotia 0,0050 ± 0,0003 100
Kultur SKM 14 + Sklerotia 0,0037 ± 0,0001 74
Supernatan SKM 14 + Sklerotia 0,0040 ± 0,0001 80
Keterangan: SE = Standard Error (nilai rataan ± SE, n = 3)
PEMBAHASAN
Potensi suatu mikrob untuk mengendalikan mikrob patogen dapat diketahui melalui uji antagonis. Antagonisme suatu mikrob terhadap mikrob lainnya dapat disebabkan oleh produksi senyawa antimikrob. Senyawa antimikrob merupakan senyawa yang mampu membunuh atau menghambat pertumbuhan mikroorganisme lain.
Uji antagonis Bacillus sp. galur G3, B. firmus E65, BGM 9, dan SKM 14 yang dilakukan terhadap X. oryzae pv. oryzae menunjukkan hanya BGM 9 dan SKM 14 yang mampu menghambat pertumbuhan X. oryzae pv. oryzae. Bacillus sp. galur G3 tidak dapat menghambat pertumbuhan X. oryzae pv. oryzae. Hal ini berbeda dengan hasil penelitian sebelumnya yang menunjukkan bahwa Bacillus sp. galur G3 menghambat sangat kuat pertumbuhan X. oryzae pv. oryzae dengan nilai IP sebesar 2,00 (Findy 2009). Perbedaan hasil uji antagonis ini diduga disebabkan di antaranya oleh adanya penurunan atau hilangnya kemampuan produksi senyawa antimikrob yang dapat menghambat X. oryzae pv. oryzae akibat peremajaan atau pengkulturan berulang yang dilakukan selama penyimpanan kultur.
Pengkulturan atau peremajaan berulang dapat menimbulkan hilangnya plasmid pembawa gen - gen untuk produksi senyawa antimikrob. Hilangnya plasmid diduga dapat menyaring atau menutupi klon antibiotik yang sensitif (Crameri et al. 1986) dan ketidakmampuan sel dalam mempertahankan replikasi DNA (Toh & Wickner 1981).
Hasil pengamatan penghambatan pertumbuhan R. solani oleh Bacillus sp. galur G3, B. firmus E65, SKM 14, dan BGM 9 menunjukkan ketiga bakteri (Bacillus sp. galur G3, B. firmus E65, dan SKM 14) mampu menghambat pertumbuhan miselia R. solani. B. firmus E65 paling baik kemampuannya dalam menghambat pertumbuhan miselia R. solani baik pada media padat maupun media cair.
Spesies Bacillus menghasilkan sedikitnya 66 jenis antibiotik (Nishijima et al. 2005). Kelompok Bacillus, khususnya Bacillus cereus, Bacillus subtilis dan Bacillus licheniformis juga menghasilkan beberapa senyawa antifungi terutama golongan peptida (Silo et al. 1994). Salah satu senyawa antifungi yang dihasilkan oleh bakteri ialah iturin (Huang et al. 1993). Bottene dan Peluso (2003) melaporkan bahwa beberapa Mucoraceae dan Aspergillus dihambat perkecambahan spora dan pemanjangan hifanya oleh B. pumilus. Berbeda dengan Bacillus, laporan penelitian tentang penghambatan pertumbuhan bakteri dan cendawan patogen oleh bakteri metanotrof sampai saat ini belum ditemukan.
antagonisme diantara ketiga bakteri tersebut. Sedangkan kombinasi B. firmus E65 dengan kedua metanotrof dalam satu formulasi sebaiknya tidak dilakukan karena adanya antagonisme antara B. firmus E65 dengan kedua bakteri metanotrof.
Berdasarkan masing-masing kemampuan-nya, kombinasi antara Bacillus sp. galur G3, SKM 14, dan BGM 9 berpotensi untuk dikembangkan sebagai suatu formulasi penyalur biokontrol untuk mengendalikan patogen padi R. solani dan X. oryzae pv. oryzae. Kemampuan BGM 9 dalam memfiksasi nitrogen dan SKM 14 dalam mengoksidasi metan juga dapat meningkatkan pertumbuhan tanaman padi dan mereduksi emisi metan di lahan sawah sehingga akan memberi nilai lebih dan tambahan manfaat pada formulasi tersebut.
SIMPULAN
Bakteri metanotrof (BGM 9 dan SKM 14) dapat menghambat pertumbuhan X. oryzae pv. oryzae dengan nilai indeks penghambatan berturut - turut 2,79 dan 2,27. Bacillus sp. galur G3, B. firmus E65, dan bakteri metanotrof SKM14 dapat menghambat pertumbuhan R. solani dengan indeks penghambatan pertumbuhan radial berturut - turut 0,63, 0,64, dan 0,56. B. firmus E65 dapat menghambat pertumbuhan bakteri metanotrof (BGM 9 dan SKM 14) dan R. solani.
Pengembangan penyalur biokontrol yang mengandung Bacillus sp. galur G3 dan kedua bakteri metanotrof (SKM 14 dan BGM 9) dapat dilakukan karena tidak ada antagonisme diantara ketiga bakteri tersebut. Sedangkan kombinasi B. firmus E65 dengan kedua metanotrof tersebut dalam satu formulasi sebaiknya tidak dilakukan karena adanya antagonisme antara B. firmus E65 dengan kedua bakteri metanotrof.
DAFTAR PUSTAKA
Anjaiah V, Koedam N, Nawak-Thompson B, Loper JE, Hofte M, Tambong JT, Cornelis P. 1998. Involvement of phenazines and anthranilate in the antagonism of Pseudomonas aeruginosa PNA1 and Tn5 derivattives toward Fusarium spp. and Pythium spp. Mol Plant Microbe Interact 11:847-854.
Bottone EJ, Peluso RW. 2003. Production by Bacillus pumilus (MSH) of an antifungal compound that is active against
Mucoraceae and Aspergillus species: preliminary report. J Med Microbiol 52:69-74.
Crameri R, Davies JE, Hutter R. 1986. Plasmid curing and generation of mutations induced with ethidium bromide in Streptomycetes. J Gen Microbiol 132:819-824.
Compant S, Duffy B, Nowak J, Clement C, Barka EA. 2005. Use of plant growth promoting bacteria for biocontrol of plant diseases: principles, mechanisms of action, and future prospects. Appl Environ Microbiol 71:4951-4959.
Dedysh SN, Ricke P, Liesack W. 2004. NifH and NifD phylogenies: an evolutionary basis for understanding nitrogen fixation capabilities of methanotrophic bacteria. Microbiol 150:1301-1313.
Dikin A, Sijam K, Kadir J, Seman IA. 2006. Antagonistic bacteria against Schizophllum commune FR. In Peninsular Malaysia. Biotropia 13:111-121.
Findy K. 2009. Aktifitas Penghambatan Zat Antimikrob Bacillus sp. asal Tanah terhadap X. oryzae, Pseudomonas fluorescens, dan Pseudomonas syringae. [skripsi]. Bogor: Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Pertanian Bogor.
Hanson R, Hanson TE. 1996. Metanotrophic bacteria. J Microbiol Rev 60:439-471. Hapsary W. 2008. Isolasi dan Karakterisasi
Bakteri Metanotrof Asal Sawah di Bogor dan Sukabumi. [skripsi]. Bogor: Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Pertanian Bogor.
Huang CH, Ano T, Shoda M. 1993. Nucleotide sequence and characteristics of the gene, lpa-14, responsible for biosynthesis of the lipopeptide antibiotics iturin and surfactin from Bacillus subtilis RB14. J Ferment Bioeng 76:445-450. Kadir TS. 2009. Menangkal HBD dengan
menggilir varietas. Warta Litbang Pertanian 31:1-3.
Lisboa MP, Bonatto D, Bizani D, Henriques JAP, Brandelli A. 2006. Characterization of a bacteriocin-like substance produced by Bacillus amyloliquefaciens isolated from the Brazillian atlantic forest. J Intern Microbiol 9:111-118.
POTENSI
Bacillus
sp. GALUR G3,
Bacillus firmus
E65, DAN
BAKTERI METANOTROF SEBAGAI PENGHAMBAT
PERTUMBUHAN PATOGEN
Xanthomonas oryzae
pv.
oryzae
DAN
Rhizoctonia solani
INDANA SARAMITA RACHMAN
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
DAN
Rhizoctonia solani
INDANA SARAMITA RACHMAN
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
ABSTRAK
INDANA SARAMITA RACHMAN. Potensi
Bacillus
sp. Galur G3,
Bacillus firmus
E65, dan
Bakteri Metanotrof sebagai Penghambat Pertumbuhan Patogen
Xanthomonas oryzae
pv.
oryzae
dan
Rhizoctonia solani
. Dibimbing oleh IMAN RUSMANA dan ALINA AKHDIYA.
Penggunaan mikroba dari genus
Bacillus
dan bakteri metanotrof sebagai agen
biokontrol untuk mengendalikan bakteri patogen tanaman padi
X. oryzae
pv
.
oryzae dan
R. solani
dapat meningkatkan produktifitas padi serta mengurangi emisi metan di lahan
sawah. Sebagian dari bakteri metanotrof dapat melakukan fiksasi nitrogen. Uji
antagonisme dilakukan untuk mengetahui kemampuan dua jenis
Bacillus
(
Bacillus
sp.
galur G3 dan
B. firmus
E65) serta dua bakteri metanotrof (BGM 9 dan SKM 14) dalam
menghambat pertumbuhan
X. oryzae
pv.
oryzae
dan
R. solani
serta mengetahui adanya
penghambatan pertumbuhan di antara kedua kelompok bakteri tersebut. Bakteri
metanotrof
(BGM 9
dan
SKM 14) dapat menghambat pertumbuhan
X. oryzae
pv.
oryzae
dengan nilai indeks penghambatan berturut-turut 2,79 dan 2,27.
Bacillus
sp. galur G3,
B.
firmus
E65 dan bakteri metanotrof SKM 14 dapat menghambat pertumbuhan
R. solani
dengan indeks penghambatan pertumbuhan radial berturut-turut 0.63, 0.64, dan 0.56.
B. firmus
E65 dapat menghambat pertumbuhan bakteri metanotrof (BGM 9 dan SKM 14)
dan
R. solani
. Formulasi agen biokontrol yang mengandung
Bacillus
sp. Galur G3, dan
kedua bakteri metanotrof (SKM 14
dan BGM 9) dapat dilakukan karena tidak ada
antagonisme diantara ketiga bakteri tersebut. Sedangkan kombinasi
B. firmus
E65
dengan kedua metanotrof sebaiknya tidak dilakukan dalam satu formulasi karena
adanya antagonisme antara
B. firmus
E65 dengan kedua bakteri metanotrof tersebut.
Kata kunci: Bakteri metanotrof,
Bacillus firmus
E65,
Bacillus
sp. galur G3,
Rhizoctonia
solani, Xanthomonas oryzae
pv.
oryzae.
Methanotrophs as Growth Inhibitor of Pathogens
Xanthomonas oryzae
pv.
oryzae
and
Rhizoctonia solani
. Under supervision of IMAN RUSMANA and ALINA AKHDIYA.
The use of bacilli and methanotropic bacteria as biocontrol agents for rice bacterial
pathogens of
X. oryzae
pv
.
oryzae and
R. solani
can enhance rice production and reduce
methane emission from rice fields. Some of methanotrophs had nitrogen fixation activity.
Antagonism methods was tested to determine the ability of two bacilli species (
Bacillus
sp. G3
and
B. firmus
E65) and two methanotrophs (BGM 9 and SKM 14
)
in inhibiting
growth of
X. oryzae
pv.
oryzae
and
R. solani
and inhibition between the two groups of
bacteria. Methanotrophs (BGM 9
and SKM 14) inhibited growth of
X. oryzae
pv.
oryzae
with inhibition index (IP) of 2,27 and 2,79 respectively.
Bacillus
sp. G3,
B. firmus
E65 and
SKM 14
isolates inhibited growth of
R. solani
with inhibition index of 0.63, 0.64, and 0.56
respectively.
B. firmus
E65
inhibited growth of methanotrophs (BGM 9 and SKM 14)
and
R. solani
. Formulation of biocontrol agents
containing
Bacillus
sp. G3
and methanotrophs
(BGM 9 and SKM 14)
can be done because there was no antagonism among the three
bacteria. However the combination of
B. firmus
E65
and the metanotrophs should not be
done in one formulation because of the antagonism between
B. firmus
E65 and the
metanotrophs.
POTENSI
Bacillus
sp. GALUR G3,
Bacillus firmus
E65, DAN BAKTERI
METANOTROF SEBAGAI PENGHAMBAT PERTUMBUHAN PATOGEN
Xanthomonas oryzae
pv.
oryzae
DAN
Rhizoctonia solani
Sarjana Sains pada
Departemen Biologi
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
BOGOR
Judul
: Potensi
Bacillus
sp. Galur G3,
Bacillus firmus
E65, dan Bakteri Metanotrof
sebagai Penghambat Pertumbuhan Patogen
Xanthomonas oryzae
pv.
oryzae
dan
Rhizoctonia solani
Nama
: Indana Saramita Rachman
NIM
: G34063511
Menyetujui:
Mengetahui:
Ketua Departemen Biologi
Fakultas Matematika dan Ilmu Pengetahuan Alam
Institut Pertanian Bogor
NIP 196410021989031002
Dr. Ir. Ence Darmo Jaya Supena, M.Si.
PRAKATA
Segala puji bagi Allah SWT, berkat rahmat dan izin-Nya penulis dapat menyelesaikan karya ilmiah ini. Shalawat serta salam tak lupa penulis haturkan kepada junjungan nabi besar Muhammad SAW yang selalu membimbing kita. Karya ilmiah ini merupakan hasil penelitian yang dilaksanakan mulai bulan Januari hingga bulan November 2010 dengan judul Potensi Isolat
Bacillus sp. Galur G3, Bacillus firmus Galur E65, dan Bakteri Metanotrof sebagai PenghambatPertumbuhan Patogen Xanthomonas oryzae pv. oryzae dan Rhizoctonia solani. Penelitian ini disusun berdasarkan hasil penelitian di Laboratorium Mikrobiologi, Departemen Biologi, Institut Pertanian Bogor.
Terima kasih penulis ucapkan kepada Bapak Iman Rusmana selaku pembimbing I, Ibu Alina Akhdiya selaku pembimbing II dan Ibu Agustin Wydia Gunawan yang telah banyak memberi saran. Di samping itu, penghargaan penulis sampaikan kepada Ibu Endang dari Balai Besar Penelitian dan Pengembangan Bioteknologi dan Sumberdaya Genetik Pertanian (BB - BIOGEN) yang telah membantu selama pengumpulan data. Ungkapan terima kasih juga disampaikan kepada Mama, Papa, Wira, Riri, Meutia, dan Kiki atas doa, dukungan dan kasih sayangnya.
Semoga karya ilmiah ini bermanfaat.
Bogor, Januari 2011
Andhy Rachmansyah dan ibunda Nurbaity. Penulis merupakan anak pertama dari tiga bersaudara. Tahun 2006 penulis lulus dari SMA Negeri 4 Medan dan lolos seleksi masuk IPB melalui Undangan Seleksi Masuk IPB (USMI) pada Departemen Biologi Fakultas Matematika dan Ilmu Pengetahuan Alam.
Selama mengikuti perkuliahan penulis aktif menjadi staf divisi Bioworld Himpunan Mahasiswa Biologi (HIMABIO) pada tahun 2008-2009 dan anggota KPH (Komisi Pengawas Himpunan Mahasiswa Biologi) pada tahun 2009-2010. Selain itu, penulis juga aktif mengikuti beberapa kegiatan kepanitian seperti Pesta Sains Nasional, Revolusi Sains tahun 2009 dan kegiatan-kegiatan Himpunan Mahasiswa Biologi serta aktif di organisasi eksternal kampus yaitu HMI Cabang Bogor.
Pada saat mengikuti perkuliahan, penulis melakukan studi lapang di Kawasan Wisata Alam Situgunung Sukabumi, dengan judul makalah “Isolasi Bacillus Selulolitik”. Penulis juga melakukan praktek lapangan di Balai Besar BIOGEN Kelti Biokimia dengan judul makalah “Antagonisme Isolat Pseudomonas sp. B dan Bacillus sp. terhadap Pseudomonas syringae,
DAFTAR ISI
Halaman
DAFTAR TABEL………...
DAFTAR GAMBAR………...
PENDAHULUAN………... Latar belakang ……….………... Tujuan ………...
BAHAN DAN METODE………... Bahan………... Metode………... Peremajaan Bakteri………... Uji Antagonisme Bacillus sp. galur G3, B. firmus E65danBakteri Metanotrof terhadap X. oryzae pv. oryzae.......... Uji Antagonisme Bacillus sp. galur G3 dan B. firmus E65 terhadap Bakteri Metanotrof (BGM 9 dan SKM 14)... Uji Antagonisme Bacillus sp. galur G3, B. firmus E65, Bakteri Metanotrof (SKM 14 dan BGM 9) terhadap R. solani pada Media Padat... Uji Antagonisme Bacillus sp. galur G3, B. firmus E65, dan Isolat Metanotrof SKM 14terhadap R. solani pada Media Cair...
HASIL………... Antagonisme Bacillus sp. Galur G3, B. firmus E65 dan Bakteri Metanotrof terhadap
X. oryzae pv. oryzae…………... Antagonisme Bacillus sp. Galur G3 dan B. firmus E65 terhadap Bakteri Metanotrof (BGM 9 dan SKM 14)………...... Antagonisme Bacillus sp. Galur G3, B. firmus E65, Bakteri Metanotrof (SKM 14dan BGM 9) terhadap R. solani pada Media Padat..………... Antagonisme Bacillus sp. Galur G3, B. firmus E65, dan Isolat Metanotrof SKM 14 terhadap R. solani pada Media Cair...
PEMBAHASAN………...
SIMPULAN………...
DAFTAR PUSTAKA………... viii
ix 1 1 1 1 1 1 1 2
2
2
2
3 3 3
3
Halaman
1 Penghambatan pertumbuhan X. oryzae pv. oryzae oleh Bacillus sp. galur G3, B. firmus E65 dan bakteri metanotrof... 4 2 Penghambatan pertumbuhan bakteri metanotrof BGM 9 dan SKM 14 oleh Bacillus
sp. galur G3dan B. Firmus E65………...... 4 3 Uji antagonisme bakteri dan cendawan Rhizoctonia solani………. 4 4
Berat kering biomassa
R. solani
di akhir inkubasi selama 15 hari pada
masing - masing media cair (29 °C) ...
5DAFTAR GAMBAR
Halaman
1 Penghambatan pertumbuhan Rhizoctonia solani oleh Bacillus sp. galur G3 pada media Potato Dextrose Agar ………... 3 2 Penghambatan pertumbuhan Rhizoctonia solani oleh B. firmus E65 pada media
1
PENDAHULUAN
Latar belakangIndonesia memiliki biodiversitas mikrob yang tinggi termasuk di antaranya berbagai jenis patogen tumbuhan, mikrob yang bersifat antagonis terhadap patogen, penambat nitrogen, dan bakteri metanotrof. Xanthomonas oryzae pv. oryzae merupakan bakteri penyebab penyakit hawar daun bakteri padi, sedangkan Rhizoctonia solani ialah patogen penyebab hawar daun, hawar pelepah, dan rebah semai pada padi. Kedua patogen tersebut tersebar luas di Indonesia dan sulit dikendalikan sehingga sangat merugikan. Pada tahun 2006 luas penularan penyakit hawar daun bakteri mencapai lebih dari 74.000 ha. Angka ini lebih tinggi dibanding kan dengan luas penularan pada tahun 2005 yang baru mencapai 33.800 ha (Kadir 2009).
Pengendalian patogen menggunakan pestisida kimiawi secara berlebihan dalam jangka panjang berbahaya bagi manusia dan lingkungan. Oleh karena itu, perlu dikembangkan teknik dan penyalur pengendali patogen yang efektif dan ramah lingkungan. Salah satu di antaranya ialah memanfaatkan potensi mikrob antagonis sebagai bahan aktif biopestisida.
Bacillus merupakan salah satu genus bakteri yang sebagian anggotanya diketahui mampu memproduksi senyawa antimikrob yang dapat membunuh atau menghambat pertumbuhan mikroorganisme lain. Selain itu, menurut Compant et al. (2005) genus bakteri ini juga dapat menginduksi sistem kekebalan tanaman.
Metanotrof merupakan kelompok bakteri yang dapat menggunakan metan sebagai sumber karbon dan sumber eneginya (Hanson R & Hanson T 1996). Sebagian dari bakteri ini juga dapat melakukan fiksasi nitrogen (Dedysh et al. 2004). Kemampuan tersebut menjadikan bakteri metanotrof berpotensi untuk dikembangkan sebagai penyalur pereduksi emisi metan dan komponen pupuk hayati untuk meningkatkan pertumbuhan tanaman padi di lahan sawah. Berbeda dengan Bacillus, sampai saat ini belum ditemukan laporan penelitian tentang kemampuan bakteri metanotrof dalam menghambat atau mengendalikan mikrob patogen.
Salah satu cara untuk meningkatkan spektrum dan efektivitas suatu formulasi penyalur biokontrol patogen tanaman ialah dengan menggabungkan berbagai mikrob
dalam satu formulasi. Penggunaan beberapa mikrob tersebut dalam satu formulasi dapat berfungsi sebagai penyalur pengendali penyakit dan juga dapat meningkatkan pertumbuhan tanaman. Kombinasi campuran mikrob dalam satu formulasi tidak boleh ada satu organisme yang dapat menghambat atau mematikan organisme lainnya yang merupakan komponen dari formulasi tersebut. Oleh karena itu, kajian terhadap potensi masing-masing mikrob, sinergi serta antagonisme diantara mikrob tersebut merupakan salah satu tahap penting yang harus dilakukan sebelum menggabungkan berbagai mikrob dalam satu formulasi.
Tujuan
Penelitian ini bertujuan mengamati kemampuan dua jenis Bacillus (Bacillus sp. galur G3 dan Bacillus firmus E65) serta dua bakteri metanotrof (BGM 9 dan SKM 14) untuk menghambat pertumbuhan Xabthomonas oryzae pv. oryzae dan Rhizoctonia solani serta mengetahui adanya penghambatan pertumbuhan diantara kedua kelompok bakteri tersebut.
BAHAN DAN METODE
BahanBakteri yang digunakan dalam penelitian ini ialah Bacillus sp. galur G3 berasal dari Laboratorium Mikrobiologi Departemen Biologi FMIPA IPB, B. firmus E65 dari koleksi BB - BIOGEN, dua isolat bakteri metanotrof dari wilayah Bogor dan Sukabumi (BGM 9 dan SKM 14), dan dua jenis patogen (X. oryzae pv. oryzae dan R. solani) dari koleksi BB - BIOGEN.
Metode
Peremajaan Bakteri
Uji Antagonisme Bacillus sp. Galur G3, B. firmus E65 dan Bakteri Metanotrof terhadap X. oryzae pv. oryzae
Antagonisme Bacillus sp. galur G3, B. firmus E65, SKM 14, dan BGM 9 terhadap X. oryzae pv. oryzae diuji secara in vitro dengan metode agar dua lapis (Lisboa et al. 2006). Keempat bakteri tersebut masing - masing ditumbuhkan pada media cair sambil dicampur dengan kecepatan 80 putaran per menit pada suhu ruang (29°C). Isolat SKM 14 dan BGM 9 diinkubasi selama sepuluh hari, sedangkan isolat yang lainnya diinkubasi selama 3 hari. Sebanyak 1 ml kultur cair X. oryzae pv. oryzae berumur 3 hari diinokulasikan ke dalam 100 ml Trypticase Soy Agar semi padat lalu dituangkan pada permukaan cawan Trypticase Soy Agar padat masing - masing sebanyak 10 ml. Setelah media Trypticase Soy Agar dua lapis memadat, kultur Bacillus sp. Galur G3 dan B. firmus E65 yang berumur 2 hari ditotolkan diatasnya dengan menggunakan tusuk gigi steril lalu diinkubasi pada suhu 30ºC. Pengamatan dan pengukuran zona hambat dilakukan setelah 48 jam untuk uji terhadap Bacillus sp. galur G3 dan B. firmus E65. Uji antagonisme untuk isolat BGM 9 dan SKM 14 dilakukan pada media Nitrat Mineral Salts padat + 1 % metanol yang di lapis dengan media Trypticase Soy Agar semi padat dan telah diinokulasikan dengan X. oryzae pv. oryzae di bagian atasnya. Pengamatan hasil uji antagonis oleh kedua bakteri metanotrof tersebut dilakukan setelah 6 hari inkubasi. Indeks penghambatan dihitung dengan persamaan berikut:
Keterangan:
IP = Indeks Penghambatan Ø zona bening = diameter zona hambat (cm) Ø koloni = diameter koloni (cm)
Uji Antagonisme Bacillus sp. Galur G3 dan
B. firmus E65 terhadap Bakteri Metanotrof (BGM 9 dan SKM 14)
Bacillus sp. galur G3 dan B. firmus E65 diuji antagonismenya terhadap bakteri metanotrof (BGM 9 dan SKM 14) pada media campuran agar Nitrat Mineral Salts + metanol 1 % dan Agar Nutrien (1:1) dengan menggunakan metode cawan sebar. Dua lup inokulum BGM 9 dan SKM 14 masing-masing disuspensikan ke dalam 4 ml air steril lalu dihomogenkan menggunakan vortex. Sebanyak 100 µl suspensi disebar ke atas
cawan berisi media campuran yang telah padat lalu diinkubasi selama 4 hari. Selanjutnya di bagian tengah permukaan media tersebut masing - masing diinokulasi dengan kultur Bacillus sp. galur G3 dan. B. firmus E65 berumur 48 jam menggunakan tusuk gigi steril. Cawan - cawan tersebut kemudian diinkubasi lagi selama 3 hari pada suhu ruang (29°C). Zona hambat di sekitar koloni Bacillus diukur untuk menentukan nilai indeks penghambatannya menggunakan persamaan di atas.
Uji Antagonisme Bacillus sp. Galur G3, B. firmus E65, dan Bakteri Metanotrof (SKM 14 dan BGM9) terhadap R. solani pada Media Padat
Bacillus sp. galur G3 dan B. firmus E65 masing -masing digores lurus sepanjang 3 cm berhadapan dengan inokulum R. solani (d = 0,8 cm) yang diletakkan di tengah cawan. Jarak antara R. solani dan bakteri uji sejauh 2 cm (Anjaiah et.al 1998). Sedangkan isolat BGM 9 dan SKM 14, diuji antagonismenya terhadap R. solani pada media campuran Potato Dextrose Agar dan Nitrat Mineral Salts Padat + metanol 1 % (1:1). BGM 9 dan SKM 14 digores 3 hari lebih awal dari inokulasi R. solani. Cawan - cawan uji untuk Bacillus sp. galur G3 dan B. firmus E65 kemudian diinkubasi pada suhu ruang selama 3 hari, sedangkan untuk bakteri metanotrof diinkubasi selama 5 hari. Penghambatan pertumbuhan radial cendawan oleh bakteri dihitung dengan rumus berikut ini (Dikin et al. 2006):
% Penghambatan Pertumbuhan Radial = 1 – (a/ b)
Keterangan:
a = Jarak antara miselia terluar dengan blok inokulasi pada sisi bakteri.
b = Jarak antara miselia terluar dengan blok inokulasi pada sisi berlawanan (tanpa bakteri).
Uji Antagonisme Bacillus sp. Galur G3, B. firmus E65, dan Isolat Metanotrof SKM 14 terhadap R. solani pada Media Cair
3
ditumbuhkan pada media Nitrat Mineral Salts cair + metanol 1 % selama 14 hari sambil dicampur pada kecepatan yang sama. Masing- masing kultur diambil sebanyak 3 ml dan dibagi ke dalam tiga tabung eppendorf masing-masing 1 ml. Sisa kultur masing - masing disentrifugasi dan supernatannya diambil 3 ml lalu dibagi ke dalam 3 tabung eppendorf masing-masing 1 ml. Kemudian dalam masing - masing eppendorf tadi dimasukkan dua butir sklerotium R. solani yang telah ditimbang, dan dimasukkan ke dalam masing - masing eppendorf lalu diinkubasi selama 15 hari. Sebagai pembanding, sklerotium ditumbuhkan dalam medium Nutrien Cair dan Nitrat Mineral Salts + metanol 1 %. Setelah diinkubasi selama 15 hari, pertumbuhan miselium pada masing - masing perlakuan diamati dan ditimbang berat kering biomassanya.
HASIL
Antagonisme Bacillus sp. Galur G3, B. firmus E65, dan Bakteri Metanotrof terhadap X. oryzae pv. oryzae
Hasil uji antagonisme menunjukkan isolat BGM 9 dan SKM 14 dapat menghambat pertumbuhan X. oryzae pv. oryzae (Tabel 1) dengan indeks penghambatan berturut-turut 2,79 dan 2,27. Sedangkan Bacillus sp. galur G3 dan B. firmus E65 tidak dapat menghambat pertumbuhan X. oryzae pv. oryzae.
Tabel 1 Penghambatan pertumbuhan X. oryzae pv. oryzae oleh Bacillus sp. galur G3, B. firmus E65, dan bakteri metanotrof
Isolat Ø koloni (cm)
Ø zona bening (cm)
IP ± SE
G3 - - -
E65 - - -
BGM 9 0,35 1,33 2,79 ± 0,36 SKM 14 0,43 1,40 2,27 ± 0,40 Keterangan: IP = Indeks Penghambatan; SE =
Standard Error (nilai rataan ± SE, n = 2); (-) = Tidak ada penghambatan.
Antagonisme Bacillus sp. Galur G3 dan B. firmus E65 terhadap Bakteri Metanotrof (BGM 9 dan SKM 14)
Informasi yang diperoleh dari hasil uji antagonisme Bacillus sp. galur G3 terhadap bakteri metanotrof diperlukan sebagai salah satu dasar pertimbangan dalam formulasi bakteri tersebut sebagai biokontrol patogen.
Bacillus sp. galur G3 tidak dapat menghambat pertumbuhan kedua bakteri metanotrof BGM 9 dan SKM 14. Sedangkan isolat B. firmus E65 dapat menghambat pertumbuhan kedua bakteri metanotrof ini dengan nilai indeks penghambatan berturut-turut 0,75 dan 1,69 (Tabel 2).
Antagonisme Bacillus sp. Galur G3, B. firmus E65, dan Bakteri Metanotrof(SKM 14 dan BGM 9) terhadap R. solani pada Media Padat
Uji antagonisme yang dilakukan pada media padat menunjukkan B. Firmus E65, Bacillus sp. galur G3, dan SKM 14 dapat menghambat pertumbuhan R. Solani. Gambar 1 dan 2 berikut menunjukkan penghambatan pertumbuhan R. solani oleh Bacillus sp. galur G3 dan B. firmus E65 pada media Potato Dextrose Agar.
Indeks penghambatan pertumbuhan radial miselia R. solani oleh Bacillus sp. galur G3, B. firmus E65, dan SKM 14 berturut-turut sebesar 0.63, 0.64, dan 0.56 (Tabel 3).
Tabel 2 Penghambatan pertumbuhan bakteri metanotrof BGM 9 dan SKM 14 oleh Bacillus sp. galur G3 dan B. firmus E65
Isolat
BGM 9 SKM 14
Ø koloni (cm)
Ø zona
hambat (cm) IP ± SE
Ø koloni (cm)
Ø zona hambat
(cm) IP ± SE
G3 - - - -
E65 0,37 0,63 0,75 ± 0,14 0,43 1,13 1,69 ± 0,64
Keterangan: IP = Indeks Penghambatan; (-) = Tidak ada penghambatan; SE = Standard Error (nilai rataan ± SE, n = 3). Uji dilakukan pada media Nitrat Mineral Salts + 1 % metanol dan Natrium Agar.
Gambar 2 Penghambatan pertumbuhan Rhizoctonia solani oleh B. firmus E65 pada media Potato Dextrose Agar.
Antagonisme Bacillus sp. Galur G3, B. firmus E65, dan Isolat Metanotrof SKM 14 terhadap R. solani pada Media Cair
Sklerotium yang diinkubasi dalam Nutrien Cair terlihat tumbuh lebih baik dan membentuk lebih banyak miselia dibandingkan dengan sklerotia yang diinkubasi di dalam kultur B. firmus E65 dan Bacillus sp. galur G3. Data berat kering biomassa R. Solani dalam Nutrien Cair juga menunjukkan pertambahan berat biomassa yang lebih tinggi dibandingkan biomassa R. solani yang ditumbuhkan dalam kultur ataupun supernatan Bacillus sp. galur G3, B. firmus E65, dan SKM 14 maupun media Nitrat Mineral Salts cair + 1 % metanol (Tabel 4).
Data berat kering biomassa R. Solani menunjukkan bahwa inkubasi sklerotium dalam kultur B. Firmus E65 menghasilkan pertambahan berat biomassa R. Solani paling rendah sebesar 58,10% dibandingkan dengan perlakuan lainnya.
Tabel 3 Uji antagonisme bakteri dan cendawan Rhizoctonia solani
Keterangan: a = Jarak antara miselia terluar dengan koloni bakteri; b = Jarak antara miselia terluar pada sisi berlawanan (tanpa bakteri); PP = Penghambatan Pertumbuhan Radial; SE = Standard error (nilai rataan ± SE, n= 3); (-) = Tidak ada penghambatan.
Bakteri Uji a b PP ± SE % Penghambatan
Pertumbuhan Radial
Bacillus sp. Galur G3 1,37 3,37 0,63 ± 0,01 63
B. firmus E65 1,37 3,83 0,64 ± 0,01 64
SKM 14 1,97 4,43 0,56 ± 0,01 56
5
Tabel 4
Berat kering biomassa R. solani di akhir inkubasi selama 15 hari pada
masing-masing media cair (29° C)
Perlakuan
Rata-rata Pertambahan Berat Kering Biomassa R. solani ± SE
(gram)
% Pertambahan Berat Biomassa
NB + Sklerotia 0,0105 ± 0,0002 100
Kultur Bacillus sp. galur G3 +
Sklerotia 0,0086 ± 0,0010 81,90
Supernatan Bacillus sp. galur G3 +
Sklerotia 0,0079 ± 0,0002 75,24
Kultur B. firmus E65 + Sklerotia 0,0061 ± 0,0002 58,10
Supernatan B. firmus E65 + Sklerotia 0,0078 ± 0,0002 74,29
NMS + Sklerotia 0,0050 ± 0,0003 100
Kultur SKM 14 + Sklerotia 0,0037 ± 0,0001 74
Supernatan SKM 14 + Sklerotia 0,0040 ± 0,0001 80
Keterangan: SE = Standard Error (nilai rataan ± SE, n = 3)
PEMBAHASAN
Potensi suatu mikrob untuk mengendalikan mikrob patogen dapat diketahui melalui uji antagonis. Antagonisme suatu mikrob terhadap mikrob lainnya dapat disebabkan oleh produksi senyawa antimikrob. Senyawa antimikrob merupakan senyawa yang mampu membunuh atau menghambat pertumbuhan mikroorganisme lain.
Uji antagonis Bacillus sp. galur G3, B. firmus E65, BGM 9, dan SKM 14 yang dilakukan terhadap X. oryzae pv. oryzae menunjukkan hanya BGM 9 dan SKM 14 yang mampu menghambat pertumbuhan X. oryzae pv. oryzae. Bacillus sp. galur G3 tidak dapat menghambat pertumbuhan X. oryzae pv. oryzae. Hal ini berbeda dengan hasil penelitian sebelumnya yang menunjukkan bahwa Bacillus sp. galur G3 menghambat sangat kuat pertumbuhan X. oryzae pv. oryzae dengan nilai IP sebesar 2,00 (Findy 2009). Perbedaan hasil uji antagonis ini diduga disebabkan di antaranya oleh adanya penurunan atau hilangnya kemampuan produksi senyawa antimikrob yang dapat menghambat X. oryzae pv. oryzae akibat peremajaan atau pengkulturan berulang yang dilakukan selama penyimpanan kultur.
Pengkulturan atau peremajaan berulang dapat menimbulkan hilangnya plasmid pembawa gen - gen untuk produksi senyawa antimikrob. Hilangnya plasmid diduga dapat menyaring atau menutupi klon antibiotik yang sensitif (Crameri et al. 1986) dan ketidakmampuan sel dalam mempertahankan replikasi DNA (Toh & Wickner 1981).
Hasil pengamatan penghambatan pertumbuhan R. solani oleh Bacillus sp. galur G3, B. firmus E65, SKM 14, dan BGM 9 menunjukkan ketiga bakteri (Bacillus sp. galur G3, B. firmus E65, dan SKM 14) mampu menghambat pertumbuhan miselia R. solani. B. firmus E65 paling baik kemampuannya dalam menghambat pertumbuhan miselia R. solani baik pada media padat maupun media cair.
Spesies Bacillus menghasilkan sedikitnya 66 jenis antibiotik (Nishijima et al. 2005). Kelompok Bacillus, khususnya Bacillus cereus, Bacillus subtilis dan Bacillus licheniformis juga menghasilkan beberapa senyawa antifungi terutama golongan peptida (Silo et al. 1994). Salah satu senyawa antifungi yang dihasilkan oleh bakteri ialah iturin (Huang et al. 1993). Bottene dan Peluso (2003) melaporkan bahwa beberapa Mucoraceae dan Aspergillus dihambat perkecambahan spora dan pemanjangan hifanya oleh B. pumilus. Berbeda dengan Bacillus, laporan penelitian tentang penghambatan pertumbuhan bakteri dan cendawan patogen oleh bakteri metanotrof sampai saat ini belum ditemukan.
antagonisme diantara ketiga bakteri tersebut. Sedangkan kombinasi B. firmus E65 dengan kedua metanotrof dalam satu formulasi sebaiknya tidak dilakukan karena adanya antagonisme antara B. firmus E65 dengan kedua bakteri metanotrof.
Berdasarkan masing-masing kemampuan-nya, kombinasi antara Bacillus sp. galur G3, SKM 14, dan BGM 9 berpotensi untuk dikembangkan sebagai suatu formulasi penyalur biokontrol untuk mengendalikan patogen padi R. solani dan X. oryzae pv. oryzae. Kemampuan BGM 9 dalam memfiksasi nitrogen dan SKM 14 dalam mengoksidasi metan juga dapat meningkatkan pertumbuhan tanaman padi dan mereduksi emisi metan di lahan sawah sehingga akan memberi nilai lebih dan tambahan manfaat pada formulasi tersebut.
SIMPULAN
Bakteri metanotrof (BGM 9 dan SKM 14) dapat menghambat pertumbuhan X. oryzae pv. oryzae dengan nilai indeks penghambatan berturut - turut 2,79 dan 2,27. Bacillus sp. galur G3, B. firmus E65, dan bakteri metanotrof SKM14 dapat menghambat pertumbuhan R. solani dengan indeks penghambatan pertumbuhan radial berturut - turut 0,63, 0,64, dan 0,56. B. firmus E65 dapat menghambat pertumbuhan bakteri metanotrof (BGM 9 dan SKM 14) dan R. solani.
Pengembangan penyalur biokontrol yang mengandung Bacillus sp. galur G3 dan kedua bakteri metanotrof (SKM 14 dan BGM 9) dapat dilakukan karena tidak ada antagonisme diantara ketiga bakteri tersebut. Sedangkan kombinasi B. firmus E65 dengan kedua metanotrof tersebut dalam satu formulasi sebaiknya tidak dilakukan karena adanya antagonisme antara B. firmus E65 dengan kedua bakteri metanotrof.
DAFTAR PUSTAKA
Anjaiah V, Koedam N, Nawak-Thompson B, Loper JE, Hofte M, Tambong JT, Cornelis P. 1998. Involvement of phenazines and anthranilate in the antagonism of Pseudomonas aeruginosa PNA1 and Tn5 derivattives toward Fusarium spp. and Pythium spp. Mol Plant Microbe Interact 11:847-854.
Bottone EJ, Peluso RW. 2003. Production by Bacillus pumilus (MSH) of an antifungal compound that is active against
Mucoraceae and Aspergillus species: preliminary report. J Med Microbiol 52:69-74.
Crameri R, Davies JE, Hutter R. 1986. Plasmid curing and generation of mutations induced with ethidium bromide in Streptomycetes. J Gen Microbiol 132:819-824.
Compant S, Duffy B, Nowak J, Clement C, Barka EA. 2005. Use of plant growth promoting bacteria for biocontrol of plant diseases: principles, mechanisms of action, and future prospects. Appl Environ Microbiol 71:4951-4959.
Dedysh SN, Ricke P, Liesack W. 2004. NifH and NifD phylogenies: an evolutionary basis for understanding nitrogen fixation capabilities of methanotrophic bacteria. Microbiol 150:1301-1313.
Dikin A, Sijam K, Kadir J, Seman IA. 2006. Antagonistic bacteria against Schizophllum commune FR. In Peninsular Malaysia. Biotropia 13:111-121.
Findy K. 2009. Aktifitas Penghambatan Zat Antimikrob Bacillus sp. asal Tanah terhadap X. oryzae, Pseudomonas fluorescens, dan Pseudomonas syringae. [skripsi]. Bogor: Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Pertanian Bogor.
Hanson R, Hanson TE. 1996. Metanotrophic bacteria. J Microbiol Rev 60:439-471. Hapsary W. 2008. Isolasi dan Karakterisasi
Bakteri Metanotrof Asal Sawah di Bogor dan Sukabumi. [skripsi]. Bogor: Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Pertanian Bogor.
Huang CH, Ano T, Shoda M. 1993. Nucleotide sequence and characteristics of the gene, lpa-14, responsible for biosynthesis of the lipopeptide antibiotics iturin and surfactin from Bacillus subtilis RB14. J Ferment Bioeng 76:445-450. Kadir TS. 2009. Menangkal HBD dengan
menggilir varietas. Warta Litbang Pertanian 31:1-3.
Lisboa MP, Bonatto D, Bizani D, Henriques JAP, Brandelli A. 2006. Characterization of a bacteriocin-like substance produced by Bacillus amyloliquefaciens isolated from the Brazillian atlantic forest. J Intern Microbiol 9:111-118.
7
Silo S, Lethbridge LA, Raffel BJ, Clardy SJ, Handelsman J. 1994. Biological activities of two fungistatic antibiotics produced by Bacillus cereus UW 85. Appl Environ Microbiol 60:2023-2030.