KAJIAN PERKEMBANGAN OOSIT KARANG LUNAK
Lobophytum strictum
NON FRAGMENTASI DAN
FRAGMENTASI BUATAN DI PULAU PRAMUKA,
KEPULAUAN SERIBU
MARSHEILLA TJAHJADI
SKRIPSI
DEPARTEMEN ILMU DAN TEKNOLOGI KELAUTAN FAKULTAS PERIKANAN DAN ILMU KELAUTAN
PERNYATAAN MENGENAI SKRIPSI DAN SUMBER INFORMASI
Dengan ini saya menyatakan bahwa Skripsi yang berjudul:
KAJIAN PERKEMBANGAN OOSIT KARANG LUNAK
Lobophytum strictum
NON FRAGMENTASI DAN
FRAGMENTASI BUATAN DI PULAU PRAMUKA,
KEPULAUAN SERIBU
adalah benar merupakan hasil karya sendiri dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Semua sumber data dan informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka dibagian akhir Skripsi ini.
Bogor, Februari 2011
RINGKASAN
MARSHEILLA TJAHJADI. Kajian Perkembangan Oosit Karang Lunak
Lobophytum strictum Non Fragmentasi dan Fragmentasi Buatan di Pulau
Pramuka, Kepulauan Seribu. Dibimbing oleh MUJIZAT KAWAROE dan ADI WINARTO.
Karang lunak belakangan ini mendapat perhatian serius dari para ahli biokimia karena senyawa bioaktif yang dihasilkan sangat bermanfaat terutama dalam dunia kesehatan, diantaranya dapat digunakan untuk anti peradangan, anti bakteri dan anti jamur serta anti kanker. Saat ini pemanfaatan karang lunak untuk memperoleh senyawa bioaktif dilakukan dengan mengambil langsung dari alam sehingga dikhawatirkan akan menimbulkan penurunan populasi karang lunak. Kegiatan budidaya dengan metode fragmentasi yaitu suatu metode yang memanfaatkan kemampuan karang lunak untuk memperbanyak koloni secara aseksual dari potongan atau patahan koloni, merupakan usaha penyediaan karang lunak di alam.
Karang lunak Lobophytum termasuk karang lunak yang memiliki potensi senyawa bioaktif, oleh sebab itu dilakukan kegiatan budidaya Lobophytum strictum. Penelitian mengenai reproduksi seksual karang lunak hasil fragmentasi dengan mengamati perkembangan gametnya perlu dilakukan guna menunjang pengembangan budidaya Lobophytum strictum.
Tujuan penelitian ini adalah mengkaji perkembangan gamet terhadap fase bulan Qomariyah pada karang lunak Lobophytum strictum non fragmentasi dan fragmentasi dan mengkaji pengaruh fragmentasi dan kedalaman lokasi
fragmentasi terhadap reproduksi seksual karang lunak Lobophytum strictum. Pengambilan sampel dilakukan pada bulan Mei dan bulan Juli 2009 saat umur fragmentasi mencapai 8 dan 10 bulan berdasarkan fase bulan Qomariyah. Setiap pengambilan sampel diwakili dengan 3 cabang koloni yang diambil secara acak dari koloni karang lunak non fragmentasi dan hasil fragmentasi pada
kedalaman 3 dan 12 meter. Pembuatan preparat histologis dari sampel karang lunak dilakukan dengan metode parafin, kemudian jaringan karang lunak dipotong dengan ketebalan ± 5μm dan diwarnai dengan pewarna ganda Hematoksilin Eosin (Gunarso, 1989 dan Kiernan, 1990). Pengamatan preparat histologi dilakukan dengan mengamati tahap perkembangan oosit, jumlah rata-rata oosit per lapang pandang, dan diameter oosit. Data jumlah rata-rata oosit kemudian dianalisis secara statistik dengan uji-F dan uji-t pada selang kepercayaan 95%.
Berdasarkan pengamatan terhadap preparat histologi Lobophytum strictum
ditemukan hanya gamet betina atau oosit pada semua sayatan cabang koloni, menunjukkan bahwa semua sampel koloni karang lunak berjenis kelamin betina. Hasil pengamatan yang menunjukkan sampel non fragmentasi hanya memiliki gamet betina dengan tingkat kematangan dari tahap I sampai tahap III sedangkan sampel hasil fragmentasi buatan pada kedalaman 3 dan 12 meter hanya memiliki oosit tahap I dan II membuktikan bahwa baik sampel karang lunak non
fragmentasi maupun karang lunak hasil fragmentasi belum mencapai kematangan reproduksi.
terjadinya pembentukan gamet baru, perkembangan gamet ke tahap
perkembangan yang lebih tinggi atau penyerapan gamet oleh tubuh karang lunak. Pengaruh kedalaman padareproduksi seksual karang lunak Lobophytum strictum
hasil fragmentasi buatan mulai terlihat berbeda nyata pada umur 10 bulan setelah fragmentasi. Reproduksi seksual karang lunak Lobophytum strictum hasil
© Hak cipta milik Marsheilla Tjahjadi, tahun 2011
Hak cipta dilindungi Undang-undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah dengan tidak merugikan kepentingan yang wajar dari Pencipta
KAJIAN PERKEMBANGAN OOSIT KARANG LUNAK
Lobophytum strictum
NON FRAGMENTASI DAN
FRAGMENTASI BUATAN DI PULAU PRAMUKA,
KEPULAUAN SERIBU
MARSHEILLA TJAHJADI
SKRIPSI
Sebagai salah satu syarat untuk memperoleh gelar Sarjana Ilmu Kelautan
Departemen Ilmu dan Teknologi Kelautan Pada Fakultas Perikanan dan Ilmu Kelautan
DEPARTEMEN ILMU DAN TEKNOLOGI KELAUTAN FAKULTAS PERIKANAN DAN ILMU KELAUTAN
Judul : KAJIAN PERKEMBANGAN OOSIT KARANG LUNAK
Lobophytum strictum NON FRAGMENTASI DAN
FRAGMENTASI BUATAN DI PULAU PRAMUKA, KEPULAUAN
SERIBU
Nama : Marsheilla Tjahjadi NRP : C54051224
Disetujui,
Pembimbing Utama Pembimbing Anggota
Dr. Ir. Mujizat Kawaroe, M.Si NIP. 19651213 199403 2 002
drh. Adi Winarto, Ph.D NIP. 19580516 198601 1 001
Mengetahui,
Ketua Departemen Ilmu dan Teknologi Kelautan
Prof.Dr. Ir. Setyo Budi Susilo, M. Sc. NIP. 19580909 1983031 003
UCAPAN TERIMA KASIH
Puji syukur ke hadirat Tuhan Yang Maha Esa atas karunia, rahmat, dan berkatNya sehingga penulis dapat menyelesaikan penelitian dan penulisan skripsi yang berjudul “ Kajian Perkembangan Oosit Karang Lunak Lobophytum strictum Non Fragmentasi dan Fragmentasi Buatan di Pulau Pramuka, Kepulauan Seribu”
sebagai salah satu syarat untuk memperoleh gelar Sarjana Ilmu Kelautan pada Fakultas Perikanan dan Ilmu Kelautan, Institut Pertanian Bogor.
Penulis menyadari bahwa penyelesaian skripsi ini tidak terlepas dari bimbingan, bantuan, dan dukungan, baik moral, spiritual maupun materi dari pihak- pihak di dalam kehidupan sang penulis. Oleh karena itu, pada kesempatan ini, penulis ingin menyampaikan rasa terima kasih dari lubuk hati yang paling dalam kepada :
1. Papa, Mama, kakak dan adik tercinta yang senantiasa memberikan doa dan dukungannya kepada penulis.
2. Prof. Dr. Ir. Dedi Soedharma, DEA selaku Ketua Tim Ristek Penelitian “ Teknologi Fragmentasi Buatan Karang Lunak (Octocorallia :
Alcyonacea) sebagai Penghasil Bioaktif untuk Penyediaan Obat-obatan” dan selaku dosen penguji yang telah memberikan masukan,
saran dan kritiknya demi kesempurnaan penulisan skripsi penulis serta seluruh anggota Tim Ristek.
3. Dr. Ir. Mujizat Kawaroe, M.Si dan drh. Adi Winarto, Ph.D selaku dosen pembimbing yang telah membimbing penulis sehingga dapat
4. Saudari Vidia Chairunnisa, teman seperjuangan selama penelitian, penulisan skripsi hingga seminar dan sidang akhir.
5. Teman-teman yang dengan tulus membantu penulis hingga
terselesaikannya skripsi ini : Iqbal, Fina, Dwito , Ika, Nofaria, Santi, Irsha, dan M. Rizky serta semua teman-teman ITK 42.
6. Teman-teman senasib sepenanggungan semenjak penulis memulai hidupnya di IPB : Marissa, Deliana, Mei Yu, Novi, Pratiwi, Lenny, Novianda, dan rekan-rekan yang tidak dapat disebutkan namanya satu per satu.
7. Teman dan adik tercinta, Olivier Yonathan atas segala bantuan dan dukungan yang diberikan hingga penulis menyelesaikan studinya. 8. Ryan C.Mendoza, yang meskipun dalam ketidakhadirannya senantiasa
menjadi sumber semangat dan inspirasi bagi penulis.
Penulis menyadari bahwa penulisan skripsi ini masih jauh dari kata sempurna, untuk itu saran dan kritik yang membangun diharapkan demi menjadikan skripsi ini lebih baik. Akhir kata, penulis berharap agar skripsi ini dapat bermanfaat bagi pengembangan ilmu pengetahuan di masa yang akan datang.
Bogor, Februari 2011
KATA PENGANTAR
Puji syukur penulis ucapkan kehadirat Tuhan Yang Maha Esa karena atas berkat rahmat dan karunia-Nya, sehingga penulis dapat menyelesaikan penelitian dan penulisan skripsi ini. Penulis mengucapkan terima kasih kepada para dosen pembimbing Dr. Ir. Mujizat Kawaroe, M.Si dan drh. Adi Winarto, Ph.D yang telah memberikan ilmu, bimbingan dan pembekalan dan serta seluruh pihak terkait yang telah membantu hingga penelitian ini dapat terlaksana. Penulis juga mengucapkan terima kasih kepada ayah dan ibu tercinta atas segala doa, nasehat, dan dukungannya.
Penulis menyadari bahwa penulisan skripsi ini masih memiliki banyak kekurangan. Oleh karena itu, kritik dan saran yang sifatnya membangun sangat diharapkan. Akhirnya penulis berharap semoga skripsi ini dapat bermanfaat dan memberikan kontribusi untuk memperkaya wawasan dan ilmu pengetahuan.
Bogor, Februari 2011
DAFTAR ISI
DAFTAR LAMPIRAN ... vii
1. PENDAHULUAN ... 1
2.5. Faktor-faktor yang Mempengaruhi Reproduksi Karang Lunak ... 15
3.METODOLOGI ... 18
3.1 Waktu dan Lokasi Penelitian ... 18
3.2 Alat dan Bahan ... 19
3.3 Metode Kerja ... 20
3.3.1 Preparat Histologis ... 20
(1) Pengambilan Sampel Karang Lunak ... 20
(2) Pembuatan Preparat Histologis ... 21
3.3.2. Pengamatan Perkembangan Oosit ... 22
3.3.3. Pengukuran Parameter Lingkungan ... 22
3.4 Analisis Data ... 23
4. HASIL DAN PEMBAHASAN ... 25
4.1 Kondisi Lingkungan Perairan Lokasi Penelitian ... 25
4.2 Pengamatan Perkembangan Oosit Karang Lunak Lobophytum strictum ... 27
4.2.1 Kematangan Reproduksi Karang Lunak Lobophytum strictum ... 27
4.2.2 Komposisi Oosit Karang Lunak Lobophytum strictum ... 30
4.3 Pengaruh Kedalaman dan Fragmentasi Buatan Terhadap Reproduksi
Seksual Karang Lunak Lobophytum strictum ... 36
4.3.1 Pengaruh Kedalaman Terhadap Reproduksi Seksual Karang Lunak Lobophytum strictum ... 36
4.3.2 Pengaruh Fragmentasi Buatan Terhadap Reproduksi Seksual Karang Lunak Lobophytum strictum ... 39
5. KESIMPULAN DAN SARAN ... 47
5.1 Kesimpulan ... 47
5.2 Saran ... ... 47
DAFTAR PUSTAKA ... 49
DAFTAR TABEL
DAFTAR GAMBAR
Halaman
Gambar 1. Lobophytum strictum saat polip kontraksi ... 3
Gambar 2. Penampang vertikal polip karang lunak ... 5
Gambar 3. Berbagai macam ukuran spikula pada Lobophytum strictum... 8
Gambar 4. Diagram ilustrasi polip yang mengandung gonad jantan dan betina dari Lobophytum crassum ... 11
Gambar 5. Perkembangan oogenesis Dendronephthya gigantea ... 13
Gambar 6. Perkembangan spermatogenesis Dendronephthya gigantea ... 15
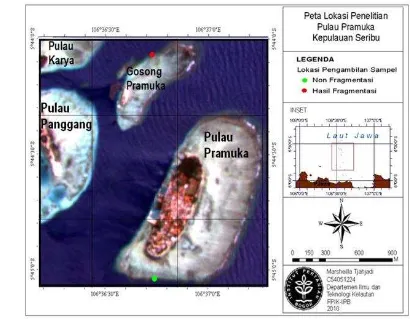
Gambar 7. Peta lokasi penelitian di perairan Pulau Pramuka ... 18
Gambar 8. Komposisi jumlah rata-rata O1 dan O2 pada karang lunak sampel non fragmentasi ... 30
Gambar 9. Komposisi jumlah rata-rata O1 dan O2 pada karang lunak sampel hasil fragmentasi buatan ... 31
Gambar 10. Oosit tahap I (O1) karang lunak Lobophytum strictum ... 33
Gambar 11. Oosit tahap II (O2) karang lunak Lobophytum strictum ... 34
Gambar 12. Persentase jumlah tahap perkembangan oosit pada fase bulan purnama pengambilan sampel tahap 1 ... 35
Gambar 13. Oosit tahap III (O3) karang lunak Lobophytum strictum ... 35
Gambar 14. Jumlah rata-rata oosit tahap I pada karang lunak hasil fragmentasi kedalaman 3 dan 12 meter umur 8 bulan setelah fragmentasi ... 37
Gambar 15. Jumlah rata-rata oosit tahap II pada karang lunak hasil fragmentasi kedalaman 3 dan 12 meter umur 8 bulan setelah fragmentasi ... 37
Gambar 16. Jumlah rata-rata oosit tahap I pada karang lunak hasil fragmentasi kedalaman 3 dan 12 meter umur 10 bulan setelah fragmentasi ... 38
Gambar 18. Jumlah rata-rata oosit tahap I pada karang lunak non fragmentasi dan fragmentasi kedalaman 12 meter umur 8 bulan setelah
fragmentasi ... 40 Gambar 19. Jumlah rata-rata oosit tahap II pada karang lunak non fragmentasi dan fragmentasi kedalaman 12 meter umur 8 bulan setelah
fragmentasi ... 41 Gambar 20. Jumlah rata-rata oosit tahap I pada karang lunak non fragmentasi dan fragmentasi kedalaman 12 meter umur 10 bulan setelah
fragmentasi ... 42 Gambar 21. Jumlah rata-rata oosit tahap II pada karang lunak non fragmentasi dan fragmentasi kedalaman 12 meter umur 10 bulan setelah
fragmentasi ... 43 Gambar 22. Jumlah rata-rata oosit tahap I pada karang lunak non fragmentasi dan fragmentasi kedalaman 3 meter umur 10 bulan setelah
fragmentasi ... 44 Gambar 23. Jumlah rata-rata oosit tahap II pada karang lunak non fragmentasi dan fragmentasi kedalaman 3 meter umur 10 bulan setelah
DAFTAR LAMPIRAN
Halaman Lampiran 1. Metode Pembuatan Preparat Histologis ... 53 Lampiran 2. Baku mutu kualitas air laut untuk biota laut ... 55 Lampiran 3. Jumlah rata-rata oosit pada karang lunak non fragmentasi dan
hasil fragmentasi ... 56 Lampiran 4. Data hasil pengukuran diameter oosit ... 57 Lampiran 5. Tabel hasil pengujian pengaruh kedalaman fragmentasi pada
karang lunak Lobophytum strictum dengan uji - t ... 58 Lampiran 6 Tabel hasil pengujian pengaruh fragmentasi buatan pada karang
1. PENDAHULUAN
1.1Latar Belakang
Karang lunak dalam ekosistem terumbu karang menempati urutan kedua setelah karang keras. Peranannya selain sebagai salah satu hewan penyusun ekosistem terumbu karang, juga sebagai pemasok senyawa karbonat yang berguna bagi pembentukan terumbu. Seperti halnya karang batu, karang lunak termasuk filum Coelenterata, kelas Anthozoa yaitu hewan yang bentuknya seperti bunga dan disebut polip.
Karang lunak sering dikenal sebagai Alcyonaria, yang merupakan nama penggolongan sub-kelas karang lunak (Alcyonaria atau Octocorallia). Tubuh karang lunak disokong oleh sejumlah besar duri-duri yang kokoh, berukuran kecil dan tersusun sedemikian rupa sehingga tubuh karang lunak lentur dan tidak mudah putus. Belakangan ini karang lunak mendapat perhatian serius dari para ahli biokimia karena karang lunak efektif menghasilkan senyawa bioaktif yang diantaranya dapat digunakan untuk anti peradangan, anti bakteri dan anti jamur, anti kanker (Sorokin, 1989 dalam Haris, 2001).
Saat ini pemanfaatan karang lunak untuk memperoleh senyawa bioaktif dilakukan dengan mengambil langsung dari alam sehingga dikhawatirkan akan menimbulkan penurunan populasi karang lunak. Oleh sebab itu, pelestariannya harus segera dilakukan agar didapatkan pemanfaatan yang optimal dan
berkesinambungan. Salah satu upaya dalam menjaga kelestarian karang lunak adalah dengan melakukan pembudidayaan. Budidaya karang lunak dapat dilakukan dengan metode fragmentasi yaitu suatu metode yang memanfaatkan
kemampuan karang lunak untuk memperbanyak koloni secara aseksual dari potongan atau patahan koloni.
Karang lunak Lobophytum termasuk karang lunak yang memiliki potensi senyawa bioaktif, maka dilakukan kegiatan budidaya Lobophytum strictum
sebagai usaha penyediaan karang lunak di alam. Penelitian mengenai reproduksi seksual karang lunak hasil fragmentasi dengan mengamati perkembangan
gametnya perlu dilakukan guna menunjang pengembangan budidaya Lobophytum strictum.
1.2Tujuan Penelitian
Tujuan penelitian ini adalah:
1) Mengkaji perkembangan oosit terhadap fase bulan Qomariyah pada karang lunak Lobophytum strictum non fragmentasi dan fragmentasi buatan 2) Mengkaji pengaruh kedalaman dan fragmentasi buatan terhadap
2. TINJAUAN PUSTAKA
2.1 Klasifikasi karang lunak Lobophytum strictum
Karang lunak Lobophytum strictum dapat diklasifikasikan sebagai berikut : Filum : Coelenterata / Cnidaria
Kelas : Anthozoa
Sub-kelas : Octocorallia Bangsa : Alcyonacea
Sub-bangsa : Alcyoniina Suku : Alcyoniidae Marga : Lobophytum
Jenis : Lobophytum strictum Tixiur- Durivault (Bayer, 1956; Verseveldt, 1983 dalam Haris, 2001)
Sumber : Penelitian Ristek Hibah Bersaing
Gambar 1. Lobophytum strictum saat polip kontraksi. a. Koloni dengan polip menjulur keluar. b. Koloni dengan polip menguncup
b
a
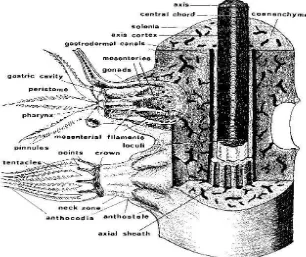
2.2 Morfologi dan Anatomi
Kelas Anthozoa dibagi dalam dua sub-kelas yaitu sub-kelas Zoantharia atau Hexacorallia atau Scleractinia dan sub-kelas Octocorallia atau lebih populer dengan Alcyonaria. Karang lunak termasuk dalam sub-kelas Alcyonaria. Pada prinsipnya yang termasuk dalam kelompok karang lunak ialah anggota
Octocorallia yang memiliki tekstur tubuh yang lunak, disokong oleh spikula yang terdapat di dalam jaringan tubuhnya, memiliki mulut yang membentuk faring yang berupa saluran, rongga gastrovaskuler, serta delapan tentakel, dilanjutkan dengan delapan septa (sekat) tidak berupa kapur yang disebut mesenteri. Polip ada yang dapat ditarik atau dijulurkan, dan ini merupakan ciri morfologi yang dapat membedakan antara jenis yang satu dengan jenis yang lain. (Manuputty, 1996a; Fossa dan Nilsen, 1998 dalam Haris, 2001).
Genus Lobophytum merupakan koloni besar bertangkai pendek,
merambat, sepintas nampak seperti mengerak (encrusting). Memiliki kapitulum lebar, permukaan atas dapat berupa lobata yaitu berbentuk jari atau juga
Gambar 2. Penampang vertikal polip karang lunak (Bayer, 1956 dalam
Manuputty, 2002)
Polip dapat dibagi menjadi tiga bagian besar yaitu antokodia, kaliks dan antostela. Antokodia merupakan bagian yang terdapat di permukaan koloni dan bersifat retraktil, yaitu dapat ditarik masuk ke dalam jaringan tubuh. Apabila antokodia ditarik ke dalam, maka yang nampak dari atas adalah pori-pori kecil seperti bintang. Bangunan luar dari pori-pori inilah yang disebut kaliks. Pada antokodia ditemukan tentakel yang berjumlah delapan dengan deretan duri-duri di sepanjang sisinya. Duri-duri ini disebut pinnula, fungsinya untuk membantu mengalirkan air dan zat-zat makanan ke dalam mulut. Selain tentakel, ditemukan mulut yang melanjutkan diri membentuk septa. Antokodia juga mengandung spikula yang letaknya berderet sampai ke ujung masing-masing tentakel. (Manuputty, 1996a; Manuputty, 2002; Fossa dan Nilsen, 1998 ).
Pada pangkal tentakel terdapat mulut yang berbentuk kepingan yang disebut stomodeum. Lanjutan mulut berupa saluran pendek disebut faring. Bagian dalam faring disusun oleh sel-sel epitel kelenjar dan sel-sel epitel kolumnar yang berflagela. Fungsi flagela untuk membantu mengalirkan air ke dalam rongga perut pada proses respirasi. Sel-sel epitel tadi tersusun sedemikian rupa sehingga bagian dalam faring berbentuk alur-alur yang disebut sifonoglifa. Bagian polip dimana sifonoglifa terletak disebut bagian ventral, sebaliknya yang berseberangan dengannya disebut bagian dorsal. Pada daerah kaliks ditemukan rongga gastrovaskuler atau rongga perut, terusan dari faring yang terbagi menjadi delapan dan disebut septa, benang-benang septa dan organ reproduksi atau gonad.
membantu menyalurkan air dan sisa-sisa makanan ke atas untuk dibuang ke luar. Sedangkan enam septa lainnya pendek-pendek, mengandung sel-sel kelenjar yang fungsinya membantu proses pencernaan makanan. Masing-masing septa
mempunyai otot retraktor yang fungsinya membantu kontraksi antokodia (Manuputty, 2002).
Tubuh Alcyonaria lunak karena tidak mempunyai kerangka kapur luar yang keras, tetapi disokong oleh sejumlah besar duri-duri yang kokoh, berukuran kecil dan tersusun sedemikian rupa sehingga tubuh Alcyonaria lentur dan tidak mudah putus. Duri-duri ini disebut spikula. Spikula berasal dari kata spiculus
atau spicus yang berarti bulir dengan ujung yang tumpul maupun runcing. Spikula terdiri dari kandungan kalsium karbonat yang padat dan keras, berfungsi sebagai penyokong seluruh bagian tubuh karang lunak mulai dari bagian basal tempat melekat sampai ke ujung tentakel. Bentuk dasar spikula bagi bangsa Octocorallia ialah bentuk kumparan sederhana (spindle) dengan permukaan mempunyai tonjolan-tonjolan namun ada juga yang memiliki permukaan licin (Manuputty, 2002).
Gambar 3. Berbagai macam ukuran spikula pada Lobophytum strictum Tixier Durivault, 1957 (Manuputty, 2002)
2.3 Reproduksi Karang Lunak
Karang lunak dapat bereproduksi secara seksual dan aseksual. Kontribusi dari tiap bentuk reproduksi terhadap pertumbuhan populasi keseluruhan bervariasi antar spesies dan lingkungan (Lasker 1983; Lasker 1996 dalam Simpson, 2008).
2.3.1 Reproduksi Aseksual
Reproduksi aseksual atau sering juga disebut reproduksi vegetatif merupakan suatu mekanisme penting dalam peningkatan jumlah individu dalam koloni karang yang dapat terjadi secara alami maupun buatan. Reproduksi aseksual yang umum terjadi adalah dengan formasi stolon, fragmentasi,
pertunasan (budding), dan pembelahan ( fission) (Fabricius dan Alderslade, 2001). Reproduksi aseksual dengan membentuk tunas terjadi pada jaring-jaring solenia yang terdapat dibagian basal tubuhnya. Solenia merupakan saluran yang
menghubungkan polip karang lunak yang satu dengan yang lainnya Polip baru muncul dalam jaringan solenia ini sebagi polip sekunder yang bentuk dan ukurannya berbeda dengan polip primer.
2.3.2 Reproduksi Seksual
Reproduksi seksual karang lunak dilakukan dengan cara kawin. Sebagian besar karang lunak bersifat dioceous dimana kelamin jantan dan betina letaknya terpisah. Mekanisme reproduksi seksual karang lunak beragam yang didasari oleh penghasil gamet dan fertilisasi (Richmond, 2001). Berdasarkan individu
penghasil gamet, reproduksi seksual dapat dikategorikan sebagai berikut (Harrison dan Wallace, 1990) :
a. Gonokhorik, yaitu tipe karang lunak yang menghasilkan hanya gamet jantan atau betina dalam satu koloni selama hidupnya (dioceous atau kelamin terpisah).
Berdasarkan mekanisme pertemuan telur dan sperma, dapat dibedakan menjadi : a. Brooding/ planulator yaitu telur dan sperma yang dihasilkan tidak
dilepaskan ke kolom perairan. Ada dua tipe brooding yaitu internal brooding dan external brooding. Bila fertilisasi terjadi secara internal dimana zigot berkembang menjadi larva planula di dalam polip untuk kemudian dilepaskan disebut internal brooding (Richmond, 2001). Ketika telur dikeluarkan di permukaan koloni karang lunak dan menunggu hingga terjadi fertilisasi disebut external brooding (Hwang dan Song, 2007).
b. Spawning yaitu melepaskan telur dan sperma ke kolom perairan sehingga terjadi fertilisasi (broadcast spawning).
Octocorallia yang memijah dengan menghasilkan telur dan dibuahi secara eksternal (broadcast spawning) adalah marga-marga Lobophytum, Sarcophyton, dan Sinularia, dan kebanyakan dari marga Alcyonium (Yamazato et al., 1981; Babcock et al., 1986 dalam Sorokin, 1989). Marga-marga tersebut memiliki koloni-koloni biseksual.
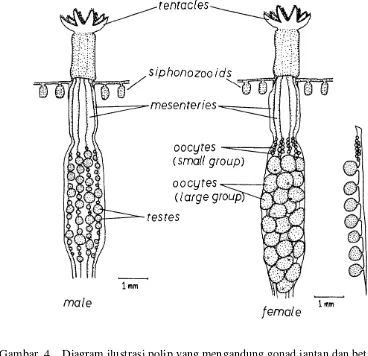
Autozoid sebagai polip yang fertil, memiliki alat kelamin atau gonad yang terpisah, masing-masing polip memiliki salah satu sel kelamin jantan atau betina. Gonad terdapat pada masing-masing mesenteri. Proses reproduksi seksual
Gambar 4. Diagram ilustrasi polip yang mengandung gonad jantan dan betina dari Lobophytum crassum. Mesenteri yang terdiri dari dua kelompok oosit ditunjukkan di sebelah kanan polip yang mengandung gonad betina (Yamazato et al., 1981).
2.4 Gametogenesis
Sel-sel kelamin berasal dari gastrodermis yang ditandai dengan lapisan mesoglea yang mengelilinginya. Pada gastrodermis terdapat sel folikel yang nantinya akan berasosiasi dengan sel kelamin yang sedang berkembang. Kehadiran sel folikel dalam gametogenesis inilah yang membedakan proses perkembangan Anthozoa dari kelas-kelas lain dalam Cnidaria (Fautin dan Mariscal, 1991 dalam Simpson, 2008).
2.4.1 Oogenesis
Perkembangan oosit dimulai ketika kumpulan gamet primordial terbagi membentuk oogonia yang akan menjadi oosit primer. Seiring dengan
perkembangan oosit, terjadi proses vitelogenesis yang merupakan proses sintesis lemak heterosintetik secara ekstensif ( Eckelbarger et al., 1998 dalam Simpson, 2008). Oosit dengan kandungan lemak tinggi telah ditemukan pada karang lunak termasuk diantaranya adalah karang Alcyonacean, Alcyonium glomeratum.
Permukaan oosit ditutupi oleh banyak mikrovili yang dapat berguna untuk proteksi mekanik atau fungsi nutrisi.
telah matang dan mencapai ukuran maksimum, butiran lemak yang melimpah dan terbungkus lapisan folikel.
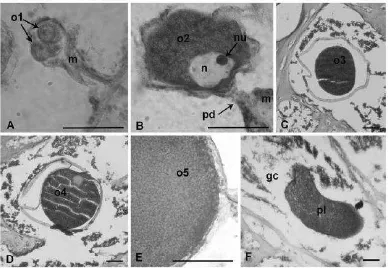
Gambar 5. Perkembangan oogenesis Dendronephthya gigantea. A. Kumpulan oosit tahap 1 yang melekat pada mesenteri. B. Oosit tahap 2 dengan nukleus terlihat jelas yang terhubung ke mesenteri dengan pedikel. C. Oosit tahap 3 yang terbungkus lapisan folikel. D. Oosit tahap 4
dengan nukleus yang terletak di bagian pinggir. E. Oosit tahap 5 dengan butiran lemak melimpah. F. Planula yang terbentuk di saluran gastrodermal (Hwang dan Song, 2007).
Keterangan : Skala 50 μm. (m mesenteri, n nukleus, nu nukleolus, o1 oosit tahap 1, o2 oosit tahap 2, o3 oosit tahap 3, o4 oosit tahap 4, o5 oosit tahap 5,gc saluran gastrodermal, pd pedikel, pl planula)
Ketika oosit dewasa mencapai ukuran akhirnya, terjadi beberapa
Ukuran telur yang lebih besar berhubungan dengan periode perkembangan oosit yang lebih lama pada karang, meskipun tidak semua karang dengan siklus oogenesis lebih panjang menghasilkan telur yang besar (Harrison dan Wallace, 1990) .
2.4.2 Spermatogenesis
Spermatogenesis terjadi pada anthozoa, dengan proses pematangan di dekat folikel sperma hingga susunan lumen intinya terisi ekor spermatozoa (Harrison, 1991). Serupa dengan oogenesis, spermatogenesis terjadi di dalam mesentarial mesoglea. Sperma matang dapat ditunjukkan secara makroskopik, terlihat dengan menonjolnya bagian samping dari mesenteri. Pada Octocorallia, ketika spawning kantung sperma akan pecah ketika melewati rongga polip, melepaskan sperma melalui bukaan mulut polip.
Pada tahap awal spermatogenesis, spermatosit mengandung nukleus dengan nukleolus dan sitoplasma yang tipis. Ketika spermatosit berkembang menjadi spermatid, sel sperma akan memiliki flagela dan membentuk susunan yang radial di sepanjang dinding cyst dengan flagela mengarah ke pusat cyst. Tahap akhir spermatogenesis adalah spermatid berubah menjadi spermatozoa dewasa (Simpson, 2008). Proses spermatogenesis terjadi lebih cepat
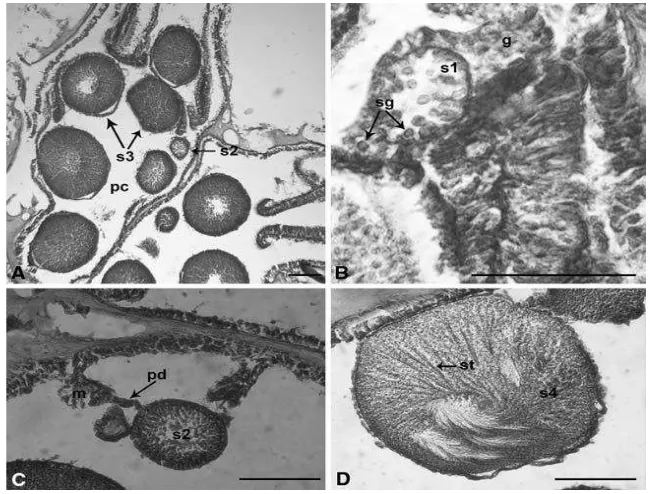
Gambar 6. Perkembangan spermatogenesis Dendronephthya gigantea. A. Berbagai macam tahap spermatogenesis dalam satu polip. B.
Sperma dengan spermatogonia dalam gastrodermis. C. Sperma tahap 2 yang terhubung pada mesenteri dengan pedikel. D. Sperma tahap 4 yang mengandung spermatozoa dengan ekor dalam jumlah besar (Hwang dan Song, 2007).
Keterangan : Skala 100 μm (g gastrodermis, m mesenteri, pc rongga polip, pd pedikel, s1 sperma tahap 1, s2 sperma tahap 2, s3 sperma tahap 3, s4
sperma tahap 4, sg spermatogonia, st kumpulan sperma berekor).
2.5 Faktor-faktor yang Mempengaruhi Reproduksi Karang Lunak Reproduksi karang lunak dipengaruhi oleh faktor internal dan eksternal. Faktor internal yang mempengaruhi reproduksi adalah umur dan ukuran koloni dan faktor genetik seperti jam biologis (Benayahu dan Loya 1984; Coma et al., 1995 dalam Simpson, 2008). Di dalam suatu spesies, ukuran koloni merupakan indikator yang baik untuk menunjukkan kematangan reproduktif.
reproduksi apabila volume koloninya belum mencapai 10 cm3( Achituv dan Benayahu, 1990)
Faktor eksternal yang mempengaruhi reproduksi karang lunak antara lain suhu, fase bulan, siklus pasang surut, arus dan ketersediaan makanan. Variasi dari berbagai faktor lingkungan diketahui berpengaruh terhadap siklus reproduksi invertebrata laut diantaranya suhu, salinitas, makanan, cahaya bulan, siklus pasang surut, dan siklus penyinaran harian (Harrison dan Wallace, 1990).
Suhu merupakan faktor penting bagi kehidupan karang lunak. Distribusi global karang keras maupun lunak yang bersimbiosis dengan zooxanthellae umumnya terdapat pada daerah perairan yang hangat dikarenakan toleransi suhu alga simbiosisnya yang terbatas (Fabricius dan Alderslade, 2001).
Biasanya karang dapat tumbuh pada suhu 18oC sampai 36oC dan pertumbuhan optimum terjadi di perairan dengan suhu rata-rata 26oC sampai 28oC. Suhu mempengaruhi kecepatan metabolisme organisme. Efek perubahan suhu pada karang dapat menyebabkan berkurangnya tingkat reproduksi, turunnya respon makan, banyak mengeluarkan lendir dan proses fotosintesis atau respirasi berkurang.
Adanya persamaan waktu dalam pengeluaran gamet ke kolom perairan pada saat surut di wilayah barat dan timur Australia mengindikasikan bahwa
pengeluaran gamet dipengaruhi oleh pola pasang surut yang mengikuti fase bulan purnama. Pola pasang surut dapat menjadi faktor penting untuk menentukan periode reproduksi yang tepat dan karang menggunakan penyinaran cahaya bulan sebagai kondisi lingkungan untuk memperkirakan pasang surut (Babcock et al., 1986 dalam Harrison dan Wallace, 1990).
Perairan yang berarus atau berombak memiliki pertumbuhan karang yang baik dibanding dengan karang yang hidup di perairan yang tidak berarus atau bergelombang. Hal ini disebabkan pada perairan yang berarus memungkinkan karang untuk memperoleh asupan oksigen yang cukup, mendapatkan sumber nutrien dan plankton sebagai sumber makanan bagi koloni karang serta menghalangi terjadinya pengendapan sedimen pada koloni (Nybakken, 1992).
Gametogenesis dan spawning yang terjadi pada kurang lunak sangat dipengaruhi oleh periode ketersediaan makanan. Pada Alcyonium digitatum
3. METODOLOGI
3.1 Waktu dan lokasi penelitian
Penelitian dilakukan dari bulan Februari - September 2009 dan Oktober 2010 - Desember 2010. Sampel karang lunak Lobophytum strictum diperoleh dari rak fragmentasi buatan yang berada di Area Perlindungan Laut pada koordinat 05o 44’03,7” LS dan 106o 36’42,5” BT dan daerah tubir laut di selatan Pulau Pramuka, Kepulauan Seribu. Pembuatan dan pengamatan preparat histologis dari sampel dilakukan di Laboratorium Histologi, Fakultas Kedokteran Hewan, Institut Pertanian Bogor.
3.2 Alat dan bahan
Alat dan bahan yang digunakan dalam penelitian mengenai reproduksi karang lunak ini dibagi menjadi dua bagian, yaitu alat dan bahan yang digunakan untuk pengambilan sampel di lapang dan untuk pembuatan preparat dan
pengamatan histologis di laboratorium. Alat dan bahan yang digunakan selama penelitian ini disajikan dalam Tabel 1.
Tabel 1. Alat dan bahan yang digunakan dalam penelitian
Alat Bahan
Pengambilan Sampel
Peralatan SCUBA Formalin 10%
Kapal motor Karang lunak Lobophytum
strictum
Pisau Selam Botol sampel
Pembuatan Preparat dan Pengamatan Histologis Alat pemanas air Eyela Water Bath SB-650 Formalin 10%
Botol film Karang lunak Lobophytum
strictum
Basket jaringan Air kran
Blok kayu Air destilasi
Pisau dan cutter Formalin 37%
Pinset Asam Formik
Jarum Asam Klorida
Lemari Asam Etanol (70%, 80%, 90%,
95% dan 100%) Embedding Console Tissue-Tek TEC Parafin
Inkubator Eyela Soft Incubator SLI-450N Enthellan
Inkubator Memmert Pewarna Hematoxylin dan
Eosin
Mikrotom Spencer 820 Xylol
Gelas objek dan cover glass Mikrometer objektif dan okuler
Mikroskop cahaya dan stereo Olympus CH20
Tutup Pagoda Kertas Label Tisu
3.3. Metode kerja
3.3.1 Preparat Histologis
(1) Pengambilan Sampel Karang Lunak
Sampel karang lunak Lobophytum strictum diambil dari karang non fragmentasi (alam) dan hasil fragmentasi buatan pada dua kedalaman berbeda yaitu 3 dan 12 meter. Pengambilan sampel karang lunak dilakukan dengan memotong koloni karang lunak secara acak menggunakan pisau atau cutter
kemudian disimpan dalam botol sampel dan difiksasi dengan formalin 10 %. Sampel karang lunak hasil fragmentasi diambil pada usia 8 bulan (pengambilan sampel tahap 1) dan 10 bulan (pengambilan sampel tahap 2). Pengambilan sampel dilakukan berdasarkan siklus bulan menurut penanggalan bulan Qomariyah yang dibagi dalam empat fase bulan, yaitu :
a. Fase bulan baru sampai bulan ¼ pertama (hari 1-7) b. Fase bulan ¼ pertama sampai bulan purnama (hari 8-14) c. Fase bulan purnama sampai bulan ¼ ketiga (hari 15-21) d. Fase bulan ¼ ketiga sampai bulan gelap (hari 22-28)
(2) Pembuatan preparat histologis
Pembuatan preparat histologis untuk pengamatan dilakukan dengan metode parafin (Gunarso, 1989 dan Kiernan, 1990). Tahapan-tahapan pembuatan preparat adalah sebagai berikut : (1) Pemotongan sampel karang lunak dengan ukuran yang sesuai untuk dijadikan preparat; (2) Dekalsifikasi sampel yang telah difiksasi dengan menggunakan larutan campuran antara HCl absolut, Formalin 37%, Asam Formik dengan komposisi 0,5 : 0,5 : 0,25 yang ditambahkan air destilasi hingga 100 ml selama ± 12 jam ; (3) Dehidrasi menggunakan alkohol bertingkat (70 – 100% ); (4) Penjernihan (clearing) menggunakan xylol sebanyak tiga kali ulangan; (5) Infiltrasi parafin ke dalam jaringan menggunakan parafin cair dalam inkubator bersuhu 650 C selama tiga kali ulangan; (6) Embedding atau penanaman sampel karang lunak menggunakan parafin cair; (7) Pendinginan dan penempelan parafin pada blok kayu; (8) Pemotongan jaringan (sectioning) menggunakan mikrotom putar dengan ketebalan ± 5μm; (9) Penempelan sayatan jaringan lunak pada kaca preparat; (10) Deparaffinisation menggunakan xylol sebanyak tiga kali ulangan masing-masing selama ± 3menit ; (11) Rehidrasi menggunakan alkohol bertingkat (100-70%) masing-masing selama 3 menit dan air (kran dan akuades) masing-masing selama 5-10 menit ; (12)Pewarnaan menggunakan pewarna Hematoksilin-Eosin; (13) Dehidrasi dengan alkohol bertingkat; (14) Penjernihan dengan xylol sebanyak tiga kali ulangan; (15)
3.3.2 Pengamatan perkembangan oosit
Pengamatan preparat histologi dilakukan di Laboratorium Histologi, Fakultas Kedokteran Hewan, Institut Pertanian Bogor. Pengamatan dilakukan menggunakan mikroskop dengan perbesaran lensa sebesar 400x untuk
memperoleh data :
a. Tahap perkembangan oosit
Perkembangan oosit diamati berdasarkan ciri morfologi dan ukurannya. b. Jumlah rata-rata oosit per lapang pandang
Jumlah oosit ditentukan berdasarkan jumlah rata-rata telur terbanyak yang ditemukan dalam lima lapang pandang pada tiga sayatan berseri dari tiap cabang koloni.
c. Diameter oosit
Panjang oosit diukur pada bagian oosit yang paling panjang dan lebarnya diukur secara tegak lurus terhadap panjangnya. Data panjang dan lebar oosit kemudian dirata-rata menjadi data diameter oosit. Untuk setiap tahap
perkembangan oosit diambil 50 sampel secara acak untuk diukur diameternya.
3.3.3 Pengukuran Parameter Lingkungan
Parameter lingkungan yang diukur adalah parameter fisika dan kimia yang dilakukan secara in situ dan pengamatan melalui analisis laboratorium. Parameter fisika – kimia yang diukur, satuan, dan alat/metode yang digunakan dalam
Tabel 2. Parameter fisika dan kimia perairan yang diukur
No Parameter Satuan Pengukuran Alat/Metode Parameter Fisika
keasaman pH laboratorium pH meter 3 Oksigen terlarut mg/l in situ Titrasi Winkler 4 Fosfat mg/l laboratorium Spektrofotometer 5 Nitrat mg/l laboratorium Spektrofotometer
3.4. Analisis data
Perkembangan dan jumlah oosit karang lunak Lobophytum strictum
dianalisis dengan melakukan pengamatan preparat histologi secara visual dengan mikroskop dan gambar hasil mikrofotografi berdasarkan fase bulan, pengaruh fragmentasi dan non fragmentasi, serta kedalaman fragmentasi dan
membandingkannya dengan pustaka terbaru atau jurnal.
Jumlah oosit diukur dengan menghitung rata –rata jumlah oosit terbanyak yang ditemukan pada lima lapang pandang di tiga sayatan berseri koloni sampel. Data jumlah oosit sampel yang diperoleh kemudian dianalisis secara statistik menggunakan uji-F dua sampel. Langkah pertama adalah melakukan pengujian mengenai asumsi homogenitas ragam populasi menggunakan uji-F dengan selang kepercayaan 95% untuk mengetahui jenis uji-t yang harus digunakan.
Jika Fhit > Ftabel maka tolak Ho yang berarti ragam populasi diasumsikan
homogen. Jika Fhit < Ftabel maka gagal tolak Ho yang berarti ragam populasi
diasumsikan sama maka digunakan uji-t dengan asumsi ragam homogen. Kedua uji-t yang dipakai menggunakan selang kepercayaan 95%. Apabila nilai thit > ttabel maka tolak Ho yang berarti ada perbedaan nyata pada
4. HASIL DAN PEMBAHASAN
4.1 Kondisi Lingkungan Perairan Lokasi Penelitian
Pengukuran parameter fisika dan kimia yang dilakukan di lapangan dan di laboratorium secara rinci disajikan dalam Tabel 3. Parameter fisika dan kimia yang diukur menunjukkan nilai dengan kisaran yang tidak terlalu jauh berbeda pada tahap 1 dan tahap 2 pengambilan sampel.
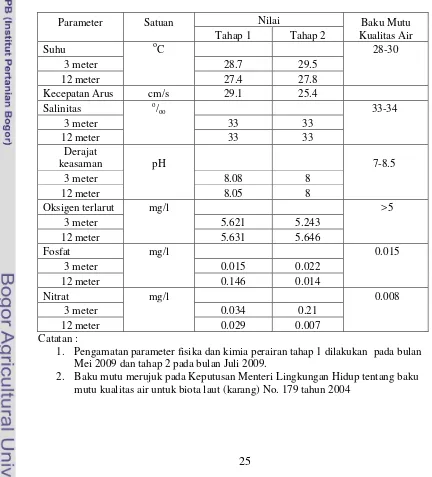
Tabel 3. Hasil pengukuran parameter fisika dan kimia perairan
Parameter
1. Pengamatan parameter fisika dan kimia perairan tahap 1 dilakukan pada bulan
Mei 2009 dan tahap 2 pada bulan Juli 2009.
2. Baku mutu merujuk pada Keputusan Menteri Lingkungan Hidup tentang baku
mutu kualitas air untuk biota laut (karang) No. 179 tahun 2004
Berdasarkan data yang diperoleh, pengukuran suhu di lokasi penelitian pada kedalaman 3 meter sebesar 28.7 oC dan 29.5 oC sedangkan data suhu pada kedalaman 12 meter sebesar 27.4 oC dan 27.8 oC pada tahap 1 dan 2 pengukuran. Suhu tersebut berada pada kisaran suhu yang baik untuk kehidupan karang sesuai dengan baku mutu kualitas air untuk karang (Lampiran 2). Birkeland (1997) menyatakan bahwa karang dapat tumbuh optimum pada kisaran suhu 26 - 28 oC. Suhu mempengaruhi kecepatan metabolisme, reproduksi dan pembentukan morfologi luar dari karang (Sorokin, 1989).
Banyak Octocorallia membutuhkan arus yang tetap, searah dan cukup kuat untuk asupan makanan yang maksimum. Arus membawa makanan dan
mengangkut limbah dari koloni dan merangsang fotosintesis (Fabricius dan Alderslade, 2001). Arus di lokasi penelitian pada kedalaman 3 meter sebesar 29.1 cm/s dan pada kedalaman 12 meter sebesar 25.4 cm/s. Kecepatan arus di
kedalaman yang lebih dangkal umumnya lebih besar karena besar kecilnya kecepatan arus dipengaruhi oleh kecepatan angin yang bertiup di permukaan perairan.
Data salinitas yang diperoleh pada kedua tahap pengambilan di kedua kedalaman memiliki nilai yang sama yaitu sebesar 33o/oo. Nilai ini dapat
dikatakan baik karena masih berada pada kisaran baku mutu salinitas untuk karang yaitu 33-34 o/oo . Secara fisiologis, salinitas mempengaruhi kehidupan
hewan karang karena adanya tekanan osmosis pada jaringan hidup. Salinitas optimal bagi kehidupan karang berkisar 30-35 o/oo (Sorokin, 1989). Meningkatnya
jarang mencapai 2-3 ppt, namun menurunnya kadar salinitas karena curah hujan tinggi atau masukan air dari sungai dapat mengganggu.
Nilai derajat keasaman yang diukur pada lokasi penelitian memiliki kisaran dengan fluktuasi yang sangat kecil yaitu antara 8 – 8.08 yang masih berada pada kisaran baku mutu kualitas air yaitu 7 – 8.5. Nilai pH yang optimum untuk pertumbuhan dan kelangsungan hidup karang lunak adalah 8.1-8.5 (Ellis dan Sharon, 2005). Hal serupa juga terjadi dalam nilai pengukuran oksigen terlarut yang seluruhnya berada di atas 5 mg/l dan sesuai dengan baku mutu kualitas air untuk karang. Nutrien seperti nitrat dan fosfat mempengaruhi
pertumbuhan alga pada karang. Alga cenderung tumbuh lebih baik pada perairan yang kaya akan nutrien.
Berdasarkan Tabel 3, nilai kandungan fosfat pada kedalaman 12 meter di pengukuran tahap1 jauh dari nilai baku mutu dibandingkan nilai pengukuran lainnya. Fosfat dalam bentuk senyawa orto-P ( PO43-) merupakan nutrien yang
dibutuhkan oleh fitoplankton untuk melakukan produktivitas primer. Nilai kandungan nitrat pada kedalaman 3 meter pada tahap 1 dan 2 sebesar 0.034 dan 0.21 mg/ l jauh di atas nilai baku mutu yang hanya sebesar 0.008 mg/l. Secara garis besar nilai kandungan nitrat di kedalaman 3 meter lebih tinggi dibandingkan dengan kedalaman 12 meter.
4.2. Pengamatan perkembangan oosit karang lunak Lobophytum strictum
4.2.1 Kematangan reproduksi karang lunak Lobophytum strictum
gamet serta jumlah dan diameter gametnya. Pengamatan terhadap sayatan menegak cabang koloni karang lunak Lobophytum strictum menunjukkan bahwa gamet ditemukan sendirian (soliter) atau berkelompok pada rongga
gastrovaskuler, menempel atau cenderung dekat dengan mesenteri yang menyebar dalam sayatan cabang koloni.
Berdasarkan pengamatan terhadap preparat histologis Lobophytum strictum ditemukan hanya gamet betina atau oosit pada semua sayatan cabang koloni. Hal ini menunjukkan bahwa semua sampel koloni karang lunak berjenis kelamin betina. Keseragaman jenis kelamin ini dikarenakan karang lunak
fragmentasi buatan pada kedalaman 3 dan 12 meter berasal dari koloni induk yang sama, dan sampel karang lunak non fragmentasi diambil dari lokasi yang sama dengan koloni induk untuk fragmentasi buatan. Dengan demikian dapat dikatakan bahwa pada lokasi asal koloni sampel untuk penelitian ini didominasi oleh
kumpulan koloni karang lunak dengan jenis kelamin betina. Tidak ditemukannya gamet jantan atau sperma selama penelitian membuktikan terjadinya reproduksi aseksual pada karang lunak Lobophytum strictum. Lobophytum pauciflorum
umumnya membentuk kumpulan koloni dimana semua koloninya memiliki jenis kelamin yang sama. Hal ini dapat terjadi disebabkan koloni-koloni dalam kumpulan koloni kemungkinan besar dibentuk dengan pembelahan secara aseksual (Fan et al., 2005).
karang lunak non fragmentasi dan fragmentasi buatan, dari pengambilan sampel tahap 1 dan 2 serta pada semua fase bulan Qomariyah yang menunjukkan terjadinya reproduksi seksual pada karang lunak Lobophytum strictum. Namun hasil pengamatan yang menunjukkan sampel non fragmentasi hanya memiliki gamet betina dengan tingkat kematangan dari tahap I sampai tahap III sedangkan sampel fragmentasi buatan pada kedalaman 3 dan 12 meter hanya memiliki oosit tahap I dan II membuktikan bahwa baik sampel karang lunak non fragmentasi maupun karang lunak hasil fragmentasi buatan belum mencapai kematangan reproduksi.
Octocorallia sangat bervariasi dalam umur dan ukuran untuk mencapai kematangan reproduksinya. Pada beberapa spesies karang, ada ukuran minimal koloni, tanpa memperhatikan umur fisiologisnya, yang harus dicapai sebelum gametogenesis dapat terjadi (Kojis dan Quinn, 1985 dalam Harrison dan Wallace, 1990). Lobophytum crassum dengan ukuran koloni dan berat kurang dari 18 cm dan 83 gram tidak memiliki gonad. Spesies ini mencapai kematangan reproduksi seksual pada saat diameter koloninya mencapai 25cm (Yamazato et al., 1981) sedangkan Lobophytum pauciflorum pada saat diameternya mencapai 10-15 cm untuk betina dan 5-10 cm untuk jantan ( Fan et al., 2005).
bulan terhadap reproduksi karang lunak Lobophytum strictum non fragmentasi dan hasil fragmentasi buatan tidak dapat diamati.
4.2.2 Komposisi oosit karang lunak Lobophytum strictum
Pengamatan yang dilakukan terhadap preparat histologis karang lunak
Lobophytum strictum menunjukkan bahwa oosit yang ditemukan dalam cabang koloni memiliki jumlah dan tahap perkembangan yang berbeda-beda. Gambar 8 memperlihatkan komposisi jumlah rata-rata O1 dan O2 pada karang lunak non fragmentasi berdasarkan fase bulan Qomariyah. Oosit tahap III (O3) ditemukan pada sampel non fragmentasi dalam jumlah sangat sedikit sehingga tidak
dimasukkan dalam penghitungan komposisi jumlah rata-rata tahap perkembangan oosit.
Gambar 8. Komposisi jumlah rata-rata O1 dan O2 pada karang lunak sampel non fragmentasi
Keterangan : (O1) Oosit tahap I, (O2) Oosit tahap II, (P) Fase bulan Purnama, (T) Fase bulan Tiga per empat, (M) Fase bulan Mati, (S) Fase bulan Seperempat
Berdasarkan grafik komposisi rata-rata tahap perkembangan oosit karang lunak terlihat adanya fluktuasi dari jumlah O1 dan O2 baik pada pengambilan
sampel tahap 1 dan 2. Jumlah rata-rata oosit terbanyak ditemukan pada fase bulan mati atau gelap baik pada pengambilan sampel tahap 1 maupun tahap 2. Jumlah O2 terlihat jelas mengalami peningkatan pada pengambilan sampel tahap 2 dibandingkan dari pengambilan sampel tahap 1.
Jumlah oosit mengalami peningkatan dari fase bulan purnama sampai fase bulan mati dan menurun pada fase bulan seperempat pengambilan sampel tahap 1, sedangkan pada pengambilan sampel tahap 2, jumlah oosit mengalami penurunan dari fase bulan purnama ke fase bulan tiga per empat, mengalami peningkatan signifikan pada fase bulan mati dan sedikit menurun kembali jumlahnya pada fase bulan seperempat.
Gambar 9 merupakan gambar komposisi jumlah rata-rata O1 dan O2 pada karang lunak Lobophytum strictum hasil fragmentasi buatan. Terjadinya fluktuasi pada komposisi jumlah rata-rata oosit dapat terlihat pada karang lunak hasil fragmentasi buatan umur 8 dan 10 bulan pada kedua kedalaman lokasi fragmentasi.
Gambar 9. Komposisi jumlah rata-rata O1 dan O2 pada karang lunak sampel hasil fragmentasi buatan
Keterangan : (O1) Oosit tahap I, (O2) Oosit tahap II, (P) Fase bulan Purnama, (T) Fase bulan Tiga per empat, (M) Fase bulan Mati, (S) Fase bulan Seperempat 3 meter 12 Meter 3 Meter 12 Meter Tahap 1 (8 bulan setelah
fragmentasi)
Jumlah rata-rata oosit terlihat lebih banyak pada sampel fragmentasi kedalaman 3 meter dibandingkan dengan 12 meter. Karang lunak fragmentasi buatan pada kedalaman 3 meter mengalami peningkatan jumlah oosit dari umur 8 bulan setelah fragmentasi ke umur 10 bulan setelah fragmentasi, terlihat jelas terutama dari peningkatan jumlah O2. Hal berbeda terjadi pada karang lunak fragmentasi buatan kedalaman 12 meter yang mengalami penurunan jumlah O1 dari umur 8 bulan setelah fragmentasi ke umur 10 bulan setelah fragmentasi, namun mengalami peningkatan jumlah O2.
Meningkatnya jumlah O1 dan O2 dikarenakan terjadinya pembentukan oosit baru dan perkembangan oosit menuju ke tingkat kematangan yang lebih tinggi. Semenjak koloni-koloni sampel diduga tidak mengalami spawning maka faktor penyebab menurunnya jumlah oosit kemungkinan besar dikarenakan terjadinya penyerapan oleh koloni karang itu sendiri sehingga jumlah oosit berkurang selama tahap perkembangannya. Dalam beberapa spesies, sejumlah oosit dapat diserap kembali selama oogenesis, agar dapat menyediakan nutrien bagi oosit yang tersisa (Harrison dan Wallace, 1990). Informasi mengenai fenomena terjadinya penyerapan oosit masih sedikit karena tidak diketahui bagaimana penyerapan itu terjadi, faktor penentu sel mana yang dikorbankan, ataukah adanya keterbatasan lingkungan, nutrisi atau fisiologi yang memicu degenerasi oosit (Harrison dan Wallace, 1990).
4.2.3 Oogenesis dan ukuran oosit karang lunak Lobophytum strictum
tahap perkembangan gamet yang dapat terlihat hanya perkembangan gamet betina atau oogenesis. Penentuan tahap perkembangan gamet mengacu pada penelitian yang dilakukan oleh Schleyer et al. (2004) terhadap spesies karang lunak
Sarcophyton glaucum dan Choi dan Song (2007) terhadap spesies karang lunak
Dendronephthya suensoni.
Berdasarkan pengamatan yang dilakukan, ditemukan oosit tahap I dan oosit tahap II tersebar di semua bagian sayatan histologis sampel. Oosit tahap I cenderung berkumpul dengan oosit lainnya, melekat pada mesoglea di mesenteri dengan nukleus yang sudah mulai terlihat dengan batas sitoplasma yang belum jelas. Ukuran diameter oosit tahap I terbilang kecil yaitu 7.24 ± 0.83 μm ( n = 50).
Gambar 10. Oosit tahap I (O1) karang lunak Lobophytum strictum
Keterangan : O1 oosit tahap I, n nukleus, batas antara nukleus dan sitoplasma belum terlalu jelas
Oosit tahap II dicirikan dengan gamet yang cenderung sudah mulai terpisah atau soliter. Umumnya terhubung pada mesenteri dengan pedikel, memiliki nukleus dan nukleolus, serta batas sitoplasmanya mulai terlihat jelas. Meskipun terkadang sulit dibedakan, oosit tahap II cenderung memiliki ukuran
n
diameter rata-rata yang lebih besar dari oosit tahap I yaitu 10.45 ± 1.35 μm ( n = 50).
Gambar 11. Oosit tahap II (O2) karang lunak Lobophytum strictum
Keterangan : O2 oosit tahap II, n nukleus, nu nukleolus
Oosit tahap III (O3) ditemukan hanya pada karang lunak non fragmentasi pengambilan sampel tahap 1 dalam jumlah sangat sedikit pada fase bulan tiga per empat, mati dan purnama dengan jumlah oosit tahap III pada fase bulan purnama paling tinggi dibandingkan dengan fase-fase lainnya. Persentase jumlah O3 berturut-turut pada fase bulan tiga per empat dan fase bulan mati hanya sebesar 0.2 % dan 0.3%. Gambar 12 merupakan persentase jumlah tahap perkembangan oosit pada fase bulan purnama dengan persentase jumlah O3 sebesar 1.4% dari total jumlah oosit yang dihitung.
O2 nu n
Gambar 12. Persentase jumlah tahap perkembangan oosit pada fase bulan purnama pengambilan sampel tahap 1
Keterangan : O1 oosit tahap I, O2 oosit tahap 2, O3 oosit tahap 3
Oosit tahap III memiliki ukuran yang jauh lebih besar, batas nukleus dan nukleolus yang jelas, mulai muncul butiran-butiran lemak yang menggantikan butiran- butiran sitoplasma dan umumnya telah terlepas dari mesenteri menuju rongga gastrovaskuler. Gambar oosit tahap III secara jelas ditampilkan pada Gambar 13.
Gambar 13. Oosit tahap 3 (O3) karang lunak Lobophytum strictum
Keterangan : O3 oosit tahap 3, rg rongga gastrovaskuler, n nukleus, nu nukleolus O3
rg
n
nu
Pengambilan sampel tahap 1 dilakukan pada bulan Mei yang merupakan musim peralihan dari musim barat ke musim timur. Ditemukannya oosit tahap III hanya pada sampel non fragmentasi pengambilan sampel tahap 1 diduga sebagai respon terhadap faktor lingkungan yang kurang mendukung pada saat itu yaitu mematangkan gamet untuk memaksimalkan reproduksinya. Hal ini didukung dengan jumlah rata-rata oosit tahap II yang mengalami penurunan pada saat yang bersamaan.
4.3 Pengaruh kedalaman dan fragmentasi buatan terhadap reproduksi seksual karang lunak Lobophytum strictum
4.3.1 Pengaruh kedalaman terhadap reproduksi seksual karang lunak
Lobophytum strictum
Fragmentasi karang lunak dilakukan pada dua kedalaman berbeda yaitu pada kedalaman 3 dan 12 meter. Perlakuan ini dimaksudkan untuk melihat pengaruh kedalaman berbeda terhadap reproduksi karang lunak, dengan asumsi bahwa perbedaan kedalaman maka terdapat perbedaan dalam faktor-faktor yang mempengaruhi reproduksi karang tersebut.
Gambar 14. Jumlah rata-rata oosit tahap I pada karang lunak hasil fragmentasi kedalaman 3 dan 12 meter umur 8 bulan setelah fragmentasi
Jumlah rata-rata oosit tahap II pada kedalaman 3 dan 12 meter yang terlihat pada Gambar 15 memiliki pola fluktuasi yang serupa, yaitu mengalami peningkatan seiring dengan pergantian fase bulan, dari fase bulan purnama sampai seperempat. Pada grafik jumlah rata-rata oosit 8 bulan setelah fragmentasi , terlihat bahwa jumlah rata-rata oosit tahap I dan tahap II pada kedalaman 3 meter lebih besar dibandingkan dengan kedalaman 12 meter.
Gambar 15. Jumlah rata-rata oosit tahap II pada karang lunak hasil fragmentasi kedalaman 3 dan 12 meter umur 8 bulan setelah fragmentasi
Uji t dengan selang kepercayaan 95% yang dilakukan untuk melihat pengaruh perbedaan perlakuan kedalaman 3 dan 12 meter menunjukkan hasil yang tidak berbeda nyata pada jumlah rata-rata oosit tahap I dan tahap II,
dibuktikan dengan nilai t hit < t tab (Lampiran 5.1 dan 5.2 ) yang berarti gagal tolak
Ho. Hal ini menjelaskan bahwa pada umur 8 bulan setelah fragmentasi ,
perkembangan reproduksi karang lunak yang diwakilkan dengan jumlah rata-rata oosit tahap I dan II menunjukkan hasil yang sama pada dua kedalaman.
Berdasarkan Gambar 16 terlihat bahwa jumlah rata-rata oosit tahap I pada kedalaman 3 dan 12 meter memiliki pola fluktuasi yang sama yaitu menurun dari fase bulan purnama sampai fase bulan mati kemudian meningkat pada fase bulan seperempat. Pada Gambar 17 terlihat bahwa fluktuasi yang sama juga terjadi pada kedalaman 3 dan 12 meter dimana jumlah oosit mengalami peningkatan dari fase bulan purnama ke fase bulan tiga per empat dan selanjutnya mengalami
penurunan. Pola fluktuasi pada grafik jumlah rata-rata oosit tahap I berbeda dengan tahap II. Pada kedua grafik terlihat bahwa jumlah rata-rata oosit tahap I dan II pada kedalaman 3 meter cukup jauh melebihi jumlah rata-rata oosit di kedalaman 12 meter.
Gambar 16. Jumlah rata-rata oosit tahap I pada karang lunak hasil fragmentasi kedalaman 3 dan 12 meter umur 10 bulan setelah fragmentasi
Gambar 17. Jumlah rata-rata oosit tahap II pada karang lunak hasil fragmentasi kedalaman 3 dan 12 meter umur 10 bulan setelah fragmentasi
Hasil uji statistik yang dilakukan menunjukkan bahwa perbedaan
kedalaman memberikan hasil yang berbeda nyata baik pada oosit tahap I dan II, dibuktikan dengan nilai t hit > t tab (Lampiran 5.3 dan 5.4 ) yang berarti tolak Ho.
Hal ini menjelaskan bahwa reproduksi seksual karang lunak pada kedalaman 3 meter dan 12 meter berbeda nyata, dengan hasil reproduksi seksual pada karang lunak fragmentasi buatan kedalaman 3 meter lebih baik dari reproduksi karang lunak fragmentasi buatan kedalaman 12 meter. Dengan demikian pada umur 10 bulan setelah fragmentasi, kedalaman lokasi fragmentasi mulai memberikan pengaruh berbeda terhadap reproduksi karang lunak Lobophytum strictum.
4.3.2 Pengaruh fragmentasi buatan terhadap reproduksi seksual karang lunak Lobophytum strictum
Reproduksi memiliki toleransi yang lebih sempit dibandingkan dengan fungsi kehidupan lainnya sehingga dapat digunakan sebagai indikator terjadinya stres pada karang (Harrison dan Wallace, 1990). Ada atau tidaknya pengaruh fragmentasi pada reproduksi seksual karang lunak Lobophytum strictum dalam
penelitian ini dilihat dengan membandingkan reproduksi seksual karang lunak non fragmentasi dan hasil fragmentasi buatan.
Karang memiliki sumber daya energi terbatas yang harus dibagi antara berbagai macam fungsi kehidupan termasuk reproduksi seksual dan aseksual, pertumbuhan, pemeliharaan dan perbaikan. Interaksi antara reproduksi dan pertumbuhan penting untuk diketahui karena mereka berkompetisi untuk mendapatkan energi setelah kebutuhan untuk perbaikan dan pemeliharaan telah terpenuhi (Harrison dan Wallace, 1990).
Gambar 18 menunjukkan bahwa jumlah rata-rata oosit tahap I pada karang lunak non fragmentasi secara garis besar memiliki nilai yang lebih tinggi dari karang lunak fragmentasi buatan kedalaman 12 meter, sedangkan pada Gambar 19 jumlah rata-rata oosit tahap II lebih tinggi pada karang lunak
fragmentasi buatan kedalaman 12 meter dibandingkan dengan karang lunak non fragmentasi.
Gambar 18. Jumlah rata-rata oosit tahap I pada karang lunak non fragmentasi dan fragmentasi kedalaman 12 meter umur 8 bulan setelah
Purnama Tiga perempat Mati /baru Seperempat
Gambar 19. Jumlah rata-rata oosit tahap II pada karang lunak non fragmentasi dan fragmentasi kedalaman 12 meter umur 8 bulan setelah
fragmentasi
Adanya variasi faktor lingkungan dan gangguan manusia dapat menyebabkan stress pada karang yang berakibat pada rendahnya fekunditas (Harrison dan Wallace, 1990). Terlihat pada Gambar 18 bahwa karang lunak hasil fragmentasi buatan kedalaman 12 meter belum dapat menghasilkan oosit sebanyak karang lunak non fragmentasi. Sebaliknya, karang lunak hasil fragmentasi buatan kedalaman 12 meter diduga selain menggunakan energinya untuk pertumbuhan juga mengalokasikan energi yang ada untuk pematangan oosit daripada pembentukan oosit baru.
Meskipun demikian, uji statistik menggunakan uji-t yang dilakukan memperlihatkan bahwa jumlah rata-rata oosit tahap I dan II pada karang lunak non fragmentasi dan fragmentasi buatan kedalaman 12 meter pada umur 8 bulan setelah fragmentasi tidak berbeda nyata pada selang kepercayaan 95%. Hal ini dibuktikan dengan nilai t hit < t tab (Lampiran 6.1 dan 6.2) yang berarti gagal tolak
Ho, maka tidak terdapat pengaruh fragmentasi buatan terhadap reproduksi seksual
karang lunak Lobophytum strictum umur 8 bulan setelah fragmentasi.
Jumlah rata-rata oosit tahap I pada karang lunak non fragmentasi cukup berbeda jauh dengan hasil fragmentasi buatan kedalaman 12 meter pada umur 10 bulan setelah fragmentasi yang terlihat pada Gambar 20. Bila dibandingkan dengan jumlah rata-rata oosit tahap I karang lunak fragmentasi buatan kedalaman 12 meter pada umur 8 bulan setelah fragmentasi, karang lunak hasil fragmentasi buatan kedalaman 12 meter pada umur 10 bulan mengalami penurunan jumlah oosit. Diduga, hal ini disebabkan adanya pengaruh faktor lingkungan yang kurang mendukung proses reproduksi seksual karang lunak hasil fragmentasi buatan kedalaman 12 meter saat berumur 10 bulan setelah fragmentasi.
Gambar 20. Jumlah rata-rata oosit tahap I pada karang lunak non fragmentasi dan fragmentasi kedalaman 12 meter umur 10 bulan setelah fragmentasi
Uji t dilakukan dengan selang kepercayaan 95 % menunjukkan hasil bahwa reproduksi seksual karang lunak fragmentasi buatan kedalaman 12 meter berbeda nyata dengan karang lunak non fragmentasi pada jumlah rata-rata oosit tahap I. Diperoleh nilai nilai t hit > t tab (Lampiran 6.3 ) yang berarti tolak Ho
dengan hasil reproduksi seksual karang lunak non fragmentasi lebih baik dari karang lunak hasil fragmentasi buatan kedalaman 12 meter untuk oosit tahap I.
Gambar 21 menunjukkan jumlah rata-rata oosit tahap II karang lunak non fragmentasi dan hasil fragmentasi buatan kedalaman 12 meter yang hampir sama pada fase bulan purnama dan tiga perempat, namun berbeda signifikan pada fase bulan mati dan seperempat dengan jumlah oosit non fragmentasi jauh melebihi jumlah oosit fragmentasi buatan kedalaman 12 meter.
Gambar 21. Jumlah rata-rata oosit tahap II pada karang lunak non fragmentasi dan fragmentasi kedalaman 12 meter umur 10 bulan setelah fragmentasi
Uji statistik yang dilakukan dengan uji t pada selang kepercayaan 95% untuk melihat pengaruh fragmentasi buatan kedalaman 12 meter terhadap jumlah rata-rata oosit tahap II pada umur 10 bulan setelah menunjukkan jumlah rata-rata oosit tahap II pada karang lunak non fragmentasi dan fragmentasi buatan
kedalaman 12 meter tidak berbeda nyata dengan nilai t hit < t tab (Lampiran 6.4)
yang berarti gagal tolak Ho. Hasil ini berbeda dengan hasil yang didapat dari
Berdasarkan Gambar 22 terlihat bahwa jumlah rata-rata oosit tahap I pada karang lunak fragmentasi buatan kedalaman 3 meter dan karang lunak non
fragmentasi memiliki nilai yang tidak berbeda jauh. Jumlah rata-rata oosit tahap I pada karang lunak hasil fragmentasi buatan kedalaman 3 meter lebih tinggi pada semua fase bulan kecuali pada fase bulan mati yang jumlahnya menurun, sedikit di bawah jumlah rata-rata oosit karang lunak non fragmentasi.
Gambar 22. Jumlah rata-rata oosit tahap I pada karang lunak non fragmentasi dan fragmentasi kedalaman 3 meter umur 10 bulan setelah fragmentasi
Terlihat dari Gambar 23, jumlah rata-rata oosit tahap II pada karang lunak non fragmentasi dan hasil fragmentasi buatan kedalaman 3 meter umur 10 bulan setelah fragmentasi memiliki nilai yang tidak berbeda jauh, kecuali pada fase bulan tiga perempat dimana jumlah rata-rata oosit pada fragmentasi buatan kedalaman 3 meter jauh melebihi jumlah oosit karang lunak non fragmentasi. Gambar 22 dan 23 menunjukkan bahwa kondisi karang lunak hasil fragmentasi buatan kedalaman 3 meter sudah pulih terlihat dari kemampuan reproduksinya yang hampir sama bahkan melebihi karang lunak non fragmentasi.
Gambar 23. Jumlah rata-rata oosit tahap II pada karang lunak non fragmentasi dan fragmentasi kedalaman 3 meter umur 10 bulan setelah
fragmentasi
Uji t dengan selang kepercayaan 95% yang dilakukan untuk menganalisa pengaruh fragmentasi buatan kedalaman 3 meter terhadap reproduksi seksual karang lunak pada umur 10 bulan setelah fragmentasi menunjukkan hasil yang tidak berbeda nyata dengan karang lunak non fragmentasi. Hal ini dibuktikan dengan nilai t hit < t tab (Lampiran 6.5 dan 6.6) yang berarti gagal tolak Ho untuk
oosit tahap I dan II.
Secara umum jumlah rata-rata oosit pada umur fragmentasi yang lebih lama yaitu umur 10 bulan setelah fragmentasi lebih besar dibandingkan dengan jumlah rata-rata oosit umur 8 bulan setelah fragmentasi karena lama umur fragmentasi terkait dengan periode yang lebih lama pula untuk memulihkan kondisi akibat fragmentasi kembali seperti normal. Namun terdapat pengecualian untuk jumlah rata-rata oosit tahap I pada fragmentasi kedalaman 12 meter yang jumlah rata-rata oositnya lebih kecil pada umur fragmentasi yang lebih lama. Fragmentasi dapat menyebabkan berkurangnya fekunditas pada karang. Fragmen karang Goniastrea favulus umur 6 bulan memiliki fekunditas yang lebih rendah
dibandingkan dengan fragmen berukuran sama berumur 2 tahun ( Kojis dan Quinn, 1981, 1985 dalam Harrison dan Wallace, 1990).
5. KESIMPULAN DAN SARAN
5.1. Kesimpulan
Karang lunak Lobophytum strictum non fragmentasi dan hasil
fragmentasi buatan belum mencapai kematangan reproduksi seksual. Pengaruh kedalaman padareproduksi seksual karang lunak Lobophytum strictum hasil fragmentasi buatan mulai terlihat berbeda nyata pada umur 10 bulan setelah fragmentasi. Reproduksi seksual karang lunak Lobophytum strictum hasil fragmentasi buatan sudah pulih dari pengaruh fragmentasi dan mampu
bereproduksi seperti karang lunak Lobophytum strictum non fragmentasi pada umur 8 bulan setelah fragmentasi. Penggunaan metode fragmentasi buatan untuk pembudidayaan karang lunak Lobophytum strictum lebih baik dilakukan pada kedalaman 3 meter karena memberikan hasil yang lebih maksimal.
5.2. Saran
Beberapa saran yang ingin disampaikan agar penelitian terkait di masa yang akan datang dapat lebih baik diantaranya :
1. Perlu dilakukan adanya tagging koloni untuk pengambilan sampel histologis untuk menghindari efek pemotongan koloni yang tidak merata terutama pada karang lunak hasil fragmentasi buatan.
2. Pencatatan ukuran diameter koloni sampel sebagai bukti kematangan reproduksi dari sampel yang diambil.
3. Sebaiknya lakukan pengambilan sampel dari bagian tengah koloni untuk menghindari terambilnya cabang koloni yang reproduksinya belum matang.
4. Pengamatan sampel menggunakan dissecting microscope untuk melihat morfologi karang lunak termasuk perkembangan gametnya sebelum diolah secara histologis.
5. Sebaiknya dilakukan penelitian yang kontinyu sampai karang lunak non fragmentasi dan hasil fragmentasi buatan memiliki siklus gametogenesis yang lengkap.
DAFTAR PUSTAKA
Achituv, Y. dan Y. Benayahu. 1990. Polyp dimorphism and functional, sequential hermaphroditism in the soft coral Heteroxenia fuscescens
(Octocorallia). Mar. Ecol. Prog. Ser. 64: 263-269.
Birkeland, C. 1997. Symbiosis, fisheries and economic development on coral reefs. Trends in Ecology and Evolution. 12: 364-367.
Choi, E.J. dan J.I. Song. 2007. Reproductive Biology of the Temperate Soft Coral Dendronephthya suensoni (Alcyonacea: Nephtheidae). Integrative Biosciences. 11: 215-225.
Ellis, E dan L. Sharron. 2005. The culture of soft corals (Order: Alcyonacea) for the marine aquarium trade. Center for Tropical and Subtropical
Aquaculture Publication No. 137.
Fabricius, K. and P. Alderslade. 2001. Soft Corals and Sea Fans A Comprehensive Guide to the Tropical Shallow Water Genera of the Central-West Pacific, the Indian Ocean and the Red Sea. Australian Institute of Marine Science. Townsville.
Fan T.Y, Y.H. Chou, dan C.F. Dai. 2005. Sexual Reproduction of the
Alcyonacean Coral Lobophytum pauciflorum in Southern Taiwan. Bull. Mar. Sci. 76(1): 143-154.
Fossa, S.A. dan A.J. Nilsen. 1998. The Modern Coral Reef Aquarium. Volume 2. Birgit Schmettkamp Verlag. Bornheim.
Gunarso, W. 1989. Bahan Pengajaran Mikroteknik. Pusat Antar Universitas Ilmu Hayat. Institut Pertanian Bogor. Bogor.
Haris, A. 2001. Laju pertumbuhan dan tingkat kelangsungan hidup fragmentasi buatan karang lunak (Octocorallia: Alcyonacea) Sarcophyton
trocheliophorum Von Marenzeller dan Lobophytum strictum Tixier-Durivault di Perairan Pulau Pari, Kepulauan Seribu. Tesis. Program Pascasarjana, Institut Pertanian Bogor. Bogor.
Harrison, F. W. 1991. Microscopic Anatomy of Invertebrates. Volume 2: Placozoa, Porifera, Cnidaria and Ctenophora. Wiley-Liss. New York. Harrison, P. L. dan C. C. Wallace. 1990. Reproduction, dispersal and
recruitment of scleractinian corals in Z. Dubinsky (ed.), Coral Reefs. Ecosystems of the world. Volume 25. Elsevier. Amsterdam. Hal 133-207.
Hartnoll, R.G. 1975. The Annual Cycle of Alcyonium digitatum. Estuarine and Coastal Marine Science. 3: 71-78.
Hwang, S.J. dan J.I. Song. 2007. Reproductive Biology and Larval Development of temperate soft coral Dendronephthya gigantea (Alcyonacea :
Neptheidae). Marine Biology. 152: 273-284.
Kiernan, J.A. 1990. Histological and Histochemical Methods: Theoryand Practice. 2nd Edition. Pergamon Press. Oxford.
Manuputty, A.E.W. 1996a. Pengenalan beberapa karang lunak (Octocorallia, Alyonacea) di lapangan. Oseana. Vol.XXI, Nomor 4: 1-11. Puslitbang-Oseanologi-LIPI, Jakarta.
Manuputty, A. E. W. 2002. Karang lunak (Soft Coral) Perairan Indonesia (Buku I, Laut Jawa & Selat Sunda). Lembaga Ilmu Pengetahuan Indonesia. Pusat Penelitian Oseanografi. Jakarta.
Mawardi, W. 2003. Ekosistem Terumbu Karang Peranan, Kondisi dan Konservasinya.
http : //www.google.co.id/ [12 Maret 2009]
Michalek-Wagner, K. dan B. L. Willis. 2001. Impacts of bleaching on the soft coral Lobophytum compactum. I. Fecundity, fertilization and off spring viability. Coral Reefs. 19: 231–239.
Nybakken, J. W. 1992. Biologi Laut : Suatu Pendekatan Ekologis.
Diterjemahkan oleh H. M. Eidman, Koesoebiono, D. G. Bengen, M. Hutomo dan S. Subarjo. PT. Gramedia Pustaka Utama. Jakarta.
Richmond, R.H. 2001. Reproduction and Recruitment in Corals: Critical Links in the Persistance of Reefs. In Birkeland, C. (ed.), Life and Death of Coral Feef. Chapman and Hall. New York. p 175 -197.
Sandy, R. E. 2000. Penempelan fragmen buatan Sinularia sp. pada pecahan karang. Tesis. Program Pascasarjana, Institut Pertanian Bogor. Bogor. Schleyer, M. H., A. Kruger, dan Y. Benayahu. 2004. Reproduction and the
unusual condition of hermaphroditism in Sarcophyton glaucum
(Octocorallia, Alcyoniidae) in KwaZulu-Natal, South Africa.
Hydrobiologia. 530(531): 399–409.
Simpson, A. Reproduction in Octocorals (Subclass Octocorallia): A Review of Published Literature. Darling Marine Center, University of Maine. http://www.google.co.id/search [12 Maret 2008]