RESISTENSI UDANG VANAME
(Litopenaeus vannamei)
HENKY MANOPPO
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
ii
PERNYATAAN MENGENAI DISERTASI DAN
SUMBER INFORMASI
Dengan ini saya menyatakan bahwa disertasi Peran Nukleotida sebagai Imunostimulan terhadap Respon Imun Nonspesifik dan Resistensi Udang Vaname (Litopenaeus vannamei) adalah karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun yang tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Bogor, Februari 2011
i
PERAN NUKLEOTIDA SEBAGAI IMUNOSTIMULAN
TERHADAP RESPON IMUN NONSPESIFIK DAN
RESISTENSI UDANG VANAME
(Litopenaeus vannamei)
HENKY MANOPPO
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
iii ABSTRACT
HENKY MANOPPO. Role of Nucleotides as Immunostimulant on Non Specific Immune Response and Resistance of Whiteleg Shrimp (Litopenaeus vannamei). Supervised by SUKENDA, DANIEL DJOKOSETIYANTO, M. FATUCHRI SUKADI and ENANG HARRIS
Whiteleg shrimp (Litopenaeus vannamei) culture in Indonesia began in 2000 to partly replace the tiger prawn (Penaeus monodon) that was seriously affected by disease. However, losses due to disease still remain an important concern till now. Therefore, a series of experiments was conducted to evaluate the efficacy of nucleotides in enhancing nonspecific immune response, resistance and growth of whiteleg shrimp. In experiment 1, juveniles were fed commercial pellet previously supplemented with nucleotides at A:0, B:100, C:200, D:300, E:400, F:500 mg.kg-1 pellet, each with three replication. At the end of feeding, shrimp was challenged intramuscularly with 0.1 mlVibrio harveyi solution 1x106 cfu.shrimp-1. Mean total haemocyte count (THC) of shrimp in treatment D (300 mg nucleotides) increased up to 76% higher than control. PO activity in treatment D (300 mg nucleotides) was also significantly different (p=0.005) compared to control. Fourteen days post-challenge, shrimp in treatment E (400 mg nucleotides) had higher resistance to V. harveyi infection (p=0.005). In treatment E, shrimp growth was significantly higher (p=0.001) than control as well as other treatments.Experiment 2 evaluated the effect of administration time of nucleotides on the nonspecific immune response and growth of shrimp. Shrimps were fed pellet supplemented with nucleotides at 400 mg.kg-1 diet. Research result showed that oral administration of nucleotides for four weeks successively significantly enhanced nonspecific immune response and shrimp growth. Experiment 3 evaluated the effect of β–glucan on total haemocyte, PO activity, resistance and growth of shrimp as comparison to nucleotides. THC of shrimp fed nucleotides diet increased up to 87% higher than control. PO activity also increased significantly (p=0.01). Supplementation of β–glucan could increase THC and PO activity, but the increase was not different compared to control. Administration of nucleotides and β–glucan for four consecutive weeks significantly increased resistance of shrimp to disease (p<0.01) where the highest resistance was observed on shrimp fed nucleotides diet. Growth of shrimp fed nucleotide-diet was significantly different compared to control (p<0.01), as well as to β–glucan diet. Experiment 4 was conducted to apply nucleotide-supplemented diet directly in brackishwater pond. Survival rate of shrimp fed nucleotide diet was higher (83.24%) than shrimp fed basal diet (81.71%) but statistically not different.Growth of shrimps fed nucleotide diet was significantly different (p<0.01) compared to shrimp fed basal diet. After 4 weeks of feeding, final weight of shrimp fed nucleotide diet was 11.98±1.08 g and weight gain was 7.48±1.08 g or 35.75% and 68.85% heavier than shrimp fed basal diet and shrimp raised in pond, respectively. As conclusion, application of nucleotides at 400 mg.kg-1
iv
RINGKASAN
HENKY MANOPPO. Peran Nukleotida sebagai Imunostimulan terhadap Respon Imun Nonspesifik dan Resistensi Udang Vaname (Litopenaeus vannamei). Dibimbing oleh SUKENDA, DANIEL DJOKOSETIYANTO, M. FATUCHRI SUKADI dan ENANG HARRIS
Udang vaname (Litopenaeus vannamei) pertama kali diimpor ke Indonesia pada tahun 2000 untuk mengganti udang windu (Penaeus monodon) yang banyak terserang penyakit terutama bakteri dan virus. Namun dalam perjalanan usaha ini, kerugian-kerugian akibat serangan penyakit terus saja berlanjut.Sejumlah metoda telah diterapkan dalam upaya mengontrol penyakit seperti penggunaan antibiotik/bahan kimia, vaksin, probiotik, SPR, dan sistim produksi biosekuriti. Belakangan ini, penggunaan imunostimulan semakin mendapat perhatian untuk dikembangkan dalam kontrol penyakit. Nukleotida merupakan imunostimulan yang menawarkan alternatif bagi penggunaan antibiotik atau bahan-bahan kimia sebab bahan ini tidak meninggalkan residu dalam tubuh ikan serta tidak mengakibatkan kerusakan lingkungan. Oleh karena itu, serangkaian penelitian telah dikerjakan untuk mengevaluasi potensi aplikasi nukleotida dalam meningkatkan respon imun nonspesifik, resistensi dan pertumbuhan udang vaname.
Penelitian pertama mengevaluasi pengaruh dosis nukleotida terhadap jumlah hemosit, aktivitas PO, resistensi dan pertumbuhan udang vaname. Udang dipelihara dalam akuarium dan diberi pakan yang telah ditambahkan nukleotida dengan dosis 0, 100, 200, 300, 400, dan 500 mg.kg-1 pakan, masing-masing dengan 3 ulangan. Hasil analisis ragam menunjukkan bahwa setelah diberikan selama 4 minggu, penambahan nukleotida dalam pakan berpengaruh nyata terhadap peningkatan respon imun udang (p<0.01). Total haemocyte count (THC) tertinggi teramati pada udang yang diberi suplementasi nukleotida 300 mg.kg-1 pakan, kemudian 400 mg.kg-1
Empat belas hari setelah uji tantang, resistensi udang yang diberi pakan dengan penambahan nukleotida 400 mg.kg
yang masing-masing mencapai 76% dan 73% lebih tinggi dari udang kontrol. Peningkatan ini dapat terjadi karena nukleotida merupakan nutrien semi esensial yang dibutuhkan untuk pertumbuhan dan perbanyakan sel, termasuk sel-sel imun. Pada kedua perlakuan ini, aktivitas PO juga mencapai nilai tertinggi yakni >0.35 yang berarti memiliki aktivitas tinggi. Bagaimana kerja nukleotida dalam meningkatkan aktivitas PO belum diketahui dan perlu diteliti secara lebih detil.Diduga, nukleotida yang ditambahkan dalam pakan selain digunakan sebagai nutrien untuk proses-proses biosintesa, juga akan berfungsi dalam cellular signals. Dalam penelitian ini terlihat bahwa udang yang memiliki THC yang tinggi memiliki aktivitas PO yang tinggi pula. Kondisi ini terjadi karena hemosit berperan dalam produksi dan pelepasan proPO ke dalam hemolim. Dalam keadaan normal, jumlah hemosit yang tinggi akan diikuti pula oleh aktivitas PO yang tinggi.
-1
v Sekalipun demikian, mekanisme imun mana yang paling penting bagi resistensi penyakit belum dapat ditetapkan. Pertumbuhan udang yang diberi nukleotida 400 mg.kg-1 pakan berbeda nyata jika dibandingkan dengan kontrol maupun dengan perlakuan lainnya (p=0.001). Udang dengan berat awal 6 g dapat tumbuh mencapai berat 11.05±0.40 g dengan perolehan 5.05±0.40 g atau mencapai 50.75% lebih berat dari perolehan berat udang kontrol. Penambahan nukleotida dalam pakan akan meningkatkan napsu makan udang sehingga efisiensi dan pengambilan pakan meningkat. Hal ini terjadi karena beberapa nukleotida seperti IMP, AMP dan guanine merupakan perangsang napsu makan ikan. Dalam penelitian ini juga teramati bahwa pada dosis yang lebih tinggi (500 mg.kg-1
Penelitian kedua mengevaluasi pengaruh protokol pemberian nukleotida terhadap jumlah hemosit, aktivitas PO dan pertumbuhan udang vaname. Udang diberi pakan dengan penambahan nukleotida 400 mg.kg
pakan), penambahan nukleotida tidak akan memacu pertumbuhan tetapi sebaliknya menekan pertumbuhan.
-1
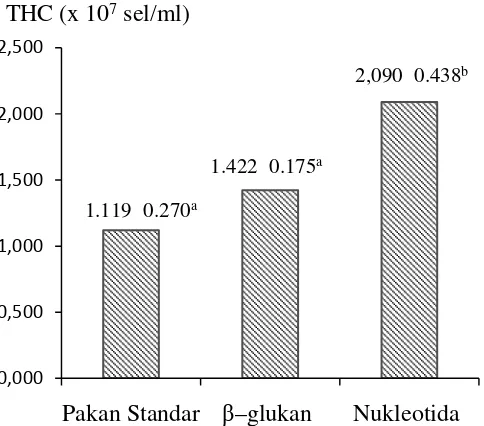
Penelitian ketiga mengevaluasi pengaruh suplementasi β–glukanterhadap total haemocyte count, aktivitas PO, resistensi dan pertumbuhan udang vaname sebagai pembanding terhadap suplementasi nukleotida. Udang diberi pakan dengan penambahan β–glukan dan nukleotida.Setelah 4 minggu pemberian pakan, THC udang yang diberi pakan bersuplemen nukleotida secara signifikan meningkat mencapai 87% lebih tinggi dari udang kontrol(p=0.02). Penambahan
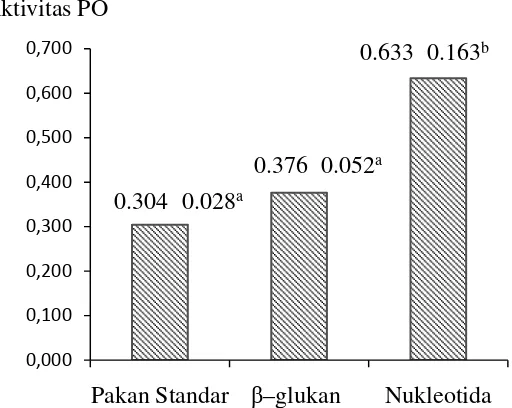
β–glukan juga dapat meningkatkan THC, namun tidak berbeda nyata jika dibandingkan dengan kontrol. Aktivitas PO juga meningkat secara sangat nyata (p=0.01) dibandingkan dengan udang yang diberi pakan standar. Aktivitas PO udang yang diberi suplementasi β–glukan juga meningkat namun dibandingkan dengan kontrol namun peningkatan yang terjadi tidak berbeda nyata. Dalam penelitian ini, nilai aktivitas PO udang, baik yang diberi suplementasi nukleotida
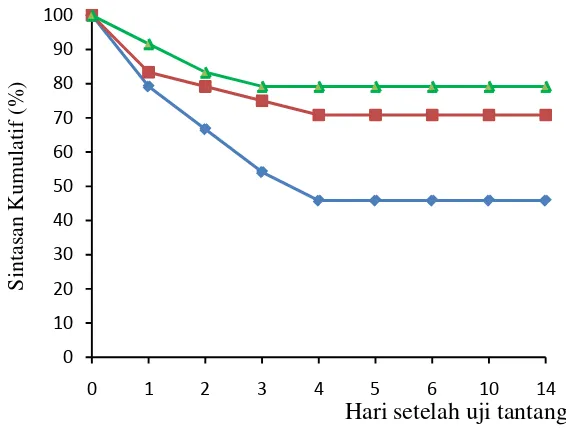
maupun β–glukan mencapai >0.35, yang berarti memiliki aktivitas tinggi sedangkan udang yang hanya diberi pakan standar memiliki aktivitas PO normal (0.20–035).Penambahan nukleotida dan β–glukan secara nyata (p=0.003) meningkatkan resistensi udang terhadap infeksi vibrio namun secara statistik, resistensi pada kedua perlakuan ini tidak berbeda nyata. Hasil ini konsisten dengan hasil penelitian pertama dimana udang yang diberi pakan yang ditambahkan nukleotida 400 mg.kg
pakan dengan protokol pemberian 7 hari pakan yang ditambahkan nukleotida dan 7 hari pakan standar secara bergantian selama 49 hari. Hasil penelitian memperlihatkan bahwa suplementasi nukleotida dengan protokol pemberian yang ditetapkan tidak berpengaruh terhadap peningkatan parameter imun maupun pertumbuhan udang vaname. Hal ini mungkin terjadi karena: 1) lama waktu pemberian nukleotida dalam percobaan ini mungkin belum cukup untuk dapat menghasilkan peningkatan respon imun dan pertumbuhan udang, 2) udang mungkin membutuhkan suplementasi nukleotida secara kontinyu untuk meningkatkan respon imun dan pertumbuhannya. Hal ini terlihat pada hasil penelitian pertama dimana pemberian nukleotida secara berlanjut selama 4 minggu mampu meningkatkan respon imun, resistensi dan pertumbuhan udang vaname.
-1
vi yang tertinggi pula. Pertumbuhan udang yang diberi pakan bersuplemen nukleotida juga berbeda nyata (p<0.01) dibandingkan dengan kontrol. Udang dengan berat rata-rata 5.39±0.56g dapat tumbuh mencapai 10.12±0.57g dengan perolehan berat 4.73±0.57g atau mencapai 65.38% lebih berat dari udang kontrol. Hasil ini dapat mengkonfirmasi hasil penelitian pertama dimana perolehan berat udang yang diberi suplementasi nukleotida mencapai 50.74% lebih berat dari kontrol. Penambahan β–glukan juga berpengaruh nyata terhadap pertumbuhan udang vaname, namun bagaimana mekanisme kerja bahan ini dalam meningkatkan pertumbuhan belum diketahui dengan jelas.
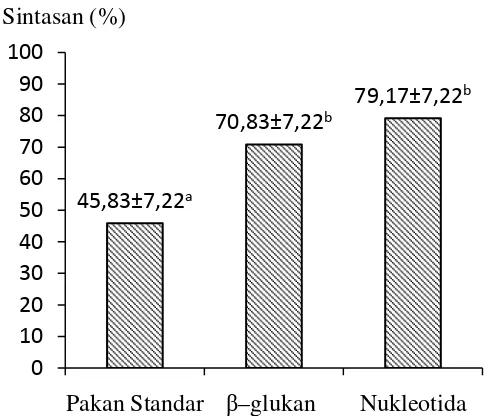
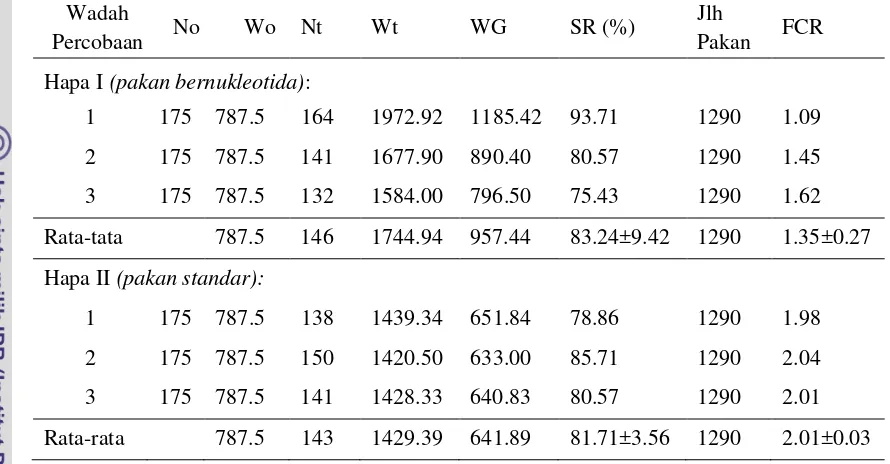
Penelitian keempat bersifat demonstratif dengan maksud untuk mengaplikasikan nukleotida secara langsung dalam manajemen kesehatan budidaya udang vaname di tambak. Udang dipelihara selama 4 minggu dalam dua rangkaian 3-hapa yang ditempatkan dalam tambak dimana usaha pemeliharaan sedang berlangsung. Pada rangkaian Hapa I, udang diberi pakan bersuplemen nukleotida 400 mg.kg-1 pakan sedangkan pada rangkaian Hapa II diberi pakan standar. Masing-masing hapa berukuran 2x1x1m dengan padat tebar 175 ekor/hapa. Hasil penelitian memperlihatkan bahwa sintasan udang yang diberi pakan bersuplemen nukleotida mencapai 83.24±9.42%, namun tidak berbeda nyata jika dibandingkan dengan sintasan udang yang diberi pakan standar (81.71±3.56%). Kematian udang selama masa pemeliharaan terjadi disebabkan oleh adanya kanibalisme terhadap udang molting dan penyakit myo (Infectious Myonecrosis Virus, IMNV). Selama masa percobaan berlangsung tidak terjadi wabah myo sehingga sulit untuk menjelaskan pengaruh nukleotida terhadap resistensi udang percobaan. Pemberian pakan dengan suplementasi nukleotida secara nyata meningkatkan pertumbuhan udang.Setelah 4 minggu pemeliharaan, udang dengan berat awal 4.5 g/ekor dapat tumbuh mencapai berat akhir 11.98±1.08 g jika diberi pakan dengan suplementasi nukleotida dan 10.01±1.36 g jika hanya diberi pakan standar. Udang yang dipelihara dalam tambak pada umur yang sama memiliki berat akhir rata-rata 8.93±0.21 g. Perolehan berat udang yang diberi suplementasi nukleotida mencapai 7.48±1.08 g atau 35.75% lebih besar dari perolehan berat udang kontrol (5.51±1.36 g) dan 68,85% lebih tinggi dari perolehan berat udang yang dipelihara di tambak (4.43±0.21 g). Perolehan berat harian rata-rata udang yang diberi pakan dengan suplementasi nukleotida mencapai 0.277±0.039 g dengan food conversion ratio (FCR) 1.35. Pada udang yang diberi pakan standar, perolehan berat harian rata-rata sebesar 0.204±0.049 g dengan FCR 2.01. Sebagai kesimpulan, penambahan nukleotida pada dosis 400 mg.kg-1
vii
©Hak Cipta milik IPB, tahun 2011
Hak Cipta dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan yang wajar IPB
viii
PERAN NUKLEOTIDA SEBAGAI IMUNOSTIMULAN
TERHADAP RESPON IMUN NONSPESIFIK DAN
RESISTENSIUDANG VANAME
(Litopenaeus vannamei)
HENKY MANOPPO
Disertasi
sebagai salah satu syarat untuk memperoleh gelar Doktor pada
Departemen Budidaya Perairan
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
ix Penguji pada Ujian Tertutup : Prof. Dr. drh. Retno Damayanti S, MS
Dr. Ir. Nur Bambang PU, MS
Penguji pada Ujian Terbuka : Dr. drh.Angela Mariana Lusiastuti, MSi
x Judul Disertasi : Peran Nukleotida sebagai Imunostimulan terhadap Respon
Imun Nonspesifik dan Resistensi Udang Vaname (Litopenaeus vannamei)
Nama : Henky Manoppo
NRP : C161060071
Disetujui
Komisi Pembimbing
Dr.Ir.Sukenda, M.Sc
Ketua Anggota
Prof.Dr.Ir.Daniel Djokosetiyanto,DEA
Prof.Dr.Ir. M.Fatuchri Sukadi, M.S
Anggota Anggota
Prof.Dr.Ir. Enang Harris, M.S
Diketahui
Ketua Program Studi Dekan Sekolah Pascasarjana Ilmu Perairan
Prof.Dr.Ir. Enang Harris, M.S Prof.Dr.Ir. Khairil A.Notodiputro, M.S
xi
PRAKATA
Puji syukur kehadirat Allah yang Maha Kuasa karena atas kasih dan limpahan karunia-Nya sehingga penyusunan laporan Disertasi dengan judul “Peran Nukleotida sebagai Imunostimulan terhadap Respon Imun Nonspesifik dan Resistensi Udang Vaname (Litopenaeus vannamei)” dapat diselesaikan. Laporan disertasi ini disusun sebagai salah satu syarat yang harus dipenuhi mahasiswa untuk menyelesaikan studi Doktor di Sekolah Pascasarjana Institut Pertanian Bogor.
Dengan tersusunnya laporan ini, penulis menghaturkan banyak terima kasih kepada Bapak Dr. Ir. Sukenda, MSc sebagai ketua komisi pembimbing,Bapak Prof. Dr.Ir. Daniel Djokosetiyanto, DEA, Bapak Prof.Dr.M.Fatuchri Sukadi, MS dan Bapak Prof.Dr.Ir. Enang Harris,MS masing-masing sebagai anggota komisi pembimbing yang telah memberikan bimbingan, petunjuk dan arahan mulai dari penyusunan proposal, pelaksanaan penelitian sampai pada penulisan laporan ini. Ungkapan terima kasih juga disampaikan kepada keluarga terkasih, istri dan anak-anak atas segala dukungan dan doanya. Terima kasih juga disampaikan kepada Bapak Ranta, Bapak Aris, Ibu Esti, Ibu Rosa, dan Bapak Atna yang telah membantu selama pelaksanaan penelitian.Disadari bahwa mungkin masih banyak kekurangan yang terdapat dalam tulisan ini. Oleh karena itu kritik dan saran perbaikan demi penyempurnaan laporan ini sangat diharapkan penulis.
Semoga karya ilmiahini dapat bermanfaat bagi pembaca sekalian.
Bogor, Februari 2011
xii
RIWAYAT HIDUP
Penulis dilahirkan di Manado sebagai anak ke empat dari enam bersaudara. Pendidikan sarjana ditempuh di Program Studi Budidaya Perairan, Fakultas Perikanan, Universitas Sam Ratulangi Manado, lulus pada tahun 1983. Pada tahun 1991, penulis diterima untuk melanjutkan ke program master di Department of Biological Science, Study Program of Aquaculture, Faculty of Science, Simon Frazer University, Canada dan lulus pada akhir tahun 1994. Beasiswa pendidikan diperoleh dari Canadian Indonesian Development Agency (CIDA) Project. Tahun 2006 penulis mendapat kesempatan untuk melanjutkan ke program doktor pada Program Studi Ilmu Perairan, Fakultas Perikanan dan Ilmu Kelautan, Institut Pertanian Bogor. Beasiswa pendidikan pascasarjana diperoleh dari Departemen Pendidikan Nasional Republik Indonesia.
Penulis bekerja sebagai staff pengajar pada Fakultas Perikanan dan Ilmu Kelautan Universitas Sam Ratulangi Manado sejak tahun 1984. Bidang penelitian yang menjadi tanggung jawab penulis adalah parasit dan penyakit ikan.
xiii
DAFTAR ISI
Halaman
DAFTAR TABEL ……… xv
DAFTAR GAMBAR ……… xvi
DAFTAR LAMPIRAN ……… xvii
PENDAHULUAN ……… 1
Latar Belakang ………. 1
Pendekatan Masalah ………. 3
Perumusan Masalah ………. 4
Tujuan Penelitian ………. 5
Manfaat Penelitian ………. 6
Hipotesis ………. 6
Kebaharuan ………. 6
TINJAUAN PUSTAKA ………. 7
JUDUL 1 RESPON IMUN NONSPESIFIK DAN RESISTENSI …….. 19
UDANG VANAME (Litopenaeus vannamei) YANG DIBERI PAKAN BERSUPLEMENNUKLEOTIDA Abstrak ………. 19
Abstract ………. 19
Pendahuluan ………. 20
Bahan dan Metode ………. 22
Hasil dan Pembahasan ……… 27
Kesimpulan ………. 35
JUDUL 2 RESPON IMUN NONSPESIFIK DAN PERTUMBUHAN … 36 UDANG VANAME (Litopenaeus vannamei)YANG DIBERI PAKAN YANG DITAMBAHKAN NUKLEOTIDA DENGAN LAMA PEMBERIAN BERBEDA Abstrak ……….. 36
Abstract ……….. 36
Pendahuluan ………. 37
Bahan dan Metode ………. 38
Hasil dan Pembahasan ……… 42
Kesimpulan ………. 45
JUDUL 3 KOMPARASI RESPON IMUN NONSPESIFIK, ………….. 46
RESISTENSI DANPERTUMBUHAN UDANGVANAME (Litopenaeus vannamei) YANG DIBERI
β
-
GLUKAN DAN NUKLEOTIDA Abstrak ………. 46Abstract ………. 46
xiv
Bahan dan Metode ………. 49
Hasil dan Pembahasan ……… 54
Kesimpulan ………. 62
JUDUL 4 APLIKASI NUKLEOTIDA DALAM BUDIDAYA ………. 63
INTENSIF UDANG VANAME(Litopenaeus vannamei) DI TAMBAK Abstrak ………. 63
Abstract ………. 63
Pendahuluan ………. 64
Bahan dan Metode ………. 66
Hasil dan Pembahasan ……… 71
Kesimpulan ………. 76
PEMBAHASAN UMUM ……… 77KESIMPULAN DAN SARAN………. 86 DAFTAR PUSTAKA ……… 87
xv
DAFTAR TABEL
Halaman
1 THC rata-rata L.vannameiyang diberi pakan yang ditambahkan nukleotida dengan dosis berbeda selama 4 minggu……… 28 2 Aktivitas PO L. vannamei yang diberi pakan yang ditambahkan
nukleotida dengan dosis berbeda selama 4 minggu………. 29 3 Resistensi L. vannamei yang diberi pakan yang ditambahkannukleotida
dengan dosis berbeda dan diuji tantang dengan bakteri Vibrio harveyi….. 31 4 Pertumbuhan L. vannamei yang diberi pakan yang
ditambahkannukleotida dengan dosis berbeda selama 4 minggu……….
33
5 Protokol administrasi pakan dengan dan tanpa suplementasi nukleotidapadaudang
vaname……….
39
6 THC, aktivitas PO dan perolehan berat udang vaname setelah diberipakan yang ditambahkan nukleotida dengan lama pemberian berbeda…...
43
7 THC, aktivitas PO dan perolehan berat udang vaname setelah diberi
pakan yang ditambahkan nukleotida selama 4x7 hari berturut-turut…….. 45 8 Pertumbuhan L. vannameisetelah diberi β-glukan dan nukleotida selama
4 minggu……….. 61
9 Sintasan dan efisiensi pakan udang vaname yang diberi pakan
yangditambahkan nukleotida selama 4 minggu………...
72
10 Performa pertumbuhan udang vaname yang dipelihara dalam Hapa
xvi
DAFTAR GAMBAR
Halaman
1 Skema pendekatan masalah……… 5
2 Morfologi Litopenaeusvannamei……… 8
3 Siklus hidup Litopenaeus vannamei……… 11
4 Proses pagositosis……… 14
5 Sintasan kumulatif udang vanameyang diberi pakan yang ditambahkan nukleotida selama 4 minggu dan diuji tantang dengan Vibrio harveyi 31 6 THC rata-rata L. vannamei setelahdiberiβ–glukan dan nukleotida selama 4 minggu………... 55
7 Aktivitas PO L.vannamei setelahdiberi β–glukandan nukleotida selama 4 minggu……… 57
8 Sintasan kumulatif udang vaname setelah diberi β–glukan dannukleotidadan diuji-tantang dengan Vibrio harveyi ……… 59
9 SintasanL.vannamei setelah diberi β–glukan dan nukleotidadan dan diuji-tantang dengan bakteri V. Harveyi……… 60
10 Tata letak hapa percobaan dalam tambak udang vaname……… 68
11 Posisi dasar hapa 10-30 cm di atas dasar tambak………. 68
12 Penghitungan dan penebaran udang dalam hapa percobaan……… 69
13 Pemberian pakan pada udang percobaan………. 70
14 Berat akhir udang vaname yang dipelihara dalam Hapa I, Hapa II, dan Tambak………... 75
15 Perolehan berat udang vaname yang dipelihara dalam Hapa I, Hapa II dan Tambak……… 76
xvii
DAFTAR LAMPIRAN
Halaman 1 Total haemocyte count udang vaname setelah diberi pakan yang
ditambahkan nukleotida dengan dosis berbeda selama 4 minggu … 95 2 Deskriptif THC udang vaname setelah diberi pakan
yangditambahkan nukleotida dengan dosis berbeda selama 4 minggu…
96
3 Analisis Ragam THC udang vaname setelah diberi pakan yangditambahkan nukleotida dengan dosis berbeda selama 4 minggu….
97
4 Uji Duncan untuk THC udang vaname setelah diberi pakan yang
ditambahkan nukleotida dengan dosis berbeda selama 4 minggu … 97 5 Aktivitas PO udang vaname setelah diberi pakan yang …………...
ditambahkan nukleotida dengan dosis berbeda selama 4 minggu
99
6 Deskriptif Aktivitas PO udang vaname setelah diberi pakan yangditambahkan nukleotida dengan dosis berbeda selama 4 minggu….
100
7 Analisis Ragam Aktivitas PO udang vaname setelah diberi pakan
yang ditambahkan nukleotida dosis berbeda selama 4 minggu …... 101 8 Uji Duncan aktivitas PO udang vaname setelah diberi pakan yang
ditambahkan nukleotida dengan dosis berbeda selama 4 minggu…. 101 9 Kelangsungan hidup kumulatif (%) udang vaname 14 hari setelah
uji tantang dengan bakteri Vibrio harveyi………. 103 10 Deskriptif tingkat resistensi (%) udang vaname 14 hari setelah uji
tantang dengan bakteri Vibrio harveyi……….. 103 11 Analisis Ragam tingkat resistensi udang vaname 14 hari setelah uji
tantang dengan bakteri Vibrio harveyi……….. 104 12 Uji Duncan untuk resistensi udang vaname 14 hari setelah uji
tantang dengan bakteri Vibrio harveyi……….. 104 13 Berat akhir dan perolehan berat udang vaname setelah diberi
pakan yang ditambahkan nukleotida dengan dosis berbeda selama
4 minggu ……… 105
14 Deskriptif perolehan berat udang vaname setelah diberi pakan
yang ditambahkan nukleotida dosis berbeda selama 4 minggu …... 106 15 Analisis Ragam perolehan berat udang vaname setelah diberi
xviii
4 minggu ……… 106
16 Uji Duncan untuk perolehan berat udang vaname setelah diberi pakan yang ditambahkan nukleotida dengan dosis berbeda selama
4 minggu ………... 107
17 THC, aktivitas PO, berat akhir (Wt) dan perolehan berat (WG) udang vaname yang diberi pakan bersuplemen nukleotida dengan
lama pemberian berbeda ……… 108
18 THC, aktivitas PO dan resistensi udang vaname yang diberi pakan
bersuplemen beta glukan dan nukleotida selama 4 minggu ………. 109 19 Deskriptif THC, aktivitas PO dan resistensi udang vaname vaname
yang diberi pakan bersuplemen beta glukan dan nukleotida selama
4 minggu ………... 109
20 Analisis Ragam THC, aktivitas PO dan resistensi udang vaname yang diberi pakan bersuplemen beta glukan dan nukleotida selama
4 minggu ………... 110
21 Uji Duncan untuk THC, aktivitas PO dan resistensi udang vaname yang diberi pakan bersuplemen beta glukan dan nukleotida selama
4 minggu ………... 110
22 Kelangungan hidup kumulatif (%) udang vaname yang diberi pakan bersuplemen beta glucan dan nukleotida dan diuji tantang
dengan Vibrio harveyi ……….. 111 23 Pertumbuhan udang vaname yang diberi pakan bersuplemen beta
glucan dan nukleotida selama 4 minggu ……….. 112 24 Deskriptif berat akhir (g) udang vaname yang diberi pakan
bersuplemen beta glucan dan nukleotida selama 4 minggu ………. 113 25 Deskriptif perolehan berat (g) udang vaname yang diberi pakan
bersuplemen beta glucan dan nukleotida selama 4 minggu ………. 113 26 Analisis Ragam berat akhir (g) udang vaname yang diberi pakan
bersuplemen beta glucan dan nukleotida selama 4 minggu ………. 114 27 Analisis Ragam perolehan berat (g) udang vaname yang diberi
pakan bersuplemen beta glucan dan nukleotida selama 4 minggu ... 114 28 Uji Duncan untuk berat akhir udang vanameyang diberi pakan
bersuplemen beta glucan dan nukleotida selama 4 minggu ………. 114 29 Uji Duncan Perolehan berat (g) udang vaname yang diberi pakan
bersuplemen beta glucan dan nukleotida selama 4 minggu ………. 114 30 Berat akhir (g) udang vaname setelah 2 minggu dipelihara dalam
Hapa I dan Hapa II ………... 116 31 Berat akhir (g) udang vaname setelah 4 minggu dipelihara dalam
Hapa I dan Hapa II ……….. 116 32 Analisis Ragam perolehan berat (g) udang vaname setelah 2
xix 33 Analisis Ragam perolehan berat (g) udang vaname setelah 4
minggu dipelihara dalam Hapa I dan Hapa II ……….. 117 34 Perolehan berat harian (g) udang vaname setelah 4 minggu
PENDAHULUAN
Latar Belakang
Sebagai suatu aktivitas industri, budidaya udang mulai berkembang sejak
awal tahun 1970-an dengan spesies budidaya utama adalah udang penaeid
(Bachere 2000). Menjelang pertengahan tahun 1970-an, produksi budidaya udang
dunia mulai meningkat dengan cepat dan mencapai 22600 ton pada tahun 1975
(Briggs et al. 2004). Pada satu dekade berikutnya, produksi mencapai 200000 ton
dimana 75% dari total produksi tersebut berasal dari Asia Tenggara dan Asia
Timur. Pada tahun 1988, produksi meningkat secara cepat melebihi 560000 ton
terutama sebagai hasil peningkatan produksi dari Cina, Taiwan, Ecuador,
Indonesia, Thailand, dan Filipina (Rosenberry 2001).
Dalam perkembangan budidaya udang, masalah kegagalan produksi
pertama terjadi di Taiwan pada tahun 1987–1989 disebabkan terjadinya degradasi
lingkungan serta meningkatnya wabah penyakit infeksius terutama bakteri dan
virus. Produksi udang windu (Penaeus monodon)Taiwan menurun drastis dari
78500 ton menjadi 16600 ton (Briggs et al. 2004). Masalah kegagalan produksi
kedua terjadi di Cina sebagai akibat munculnya penyakit White Spot Syndrome
Virus (WSSV) pada tahun 1992.Produksi menurun dari 207000 ton pada tahun
1992 menjadi 64000 ton pada tahun 1993-1994.Masalah yang sama juga terjadi di
Thailand, Filipina, dan Indonesia yang disebabkan oleh penyakit Yellow Head
Virus (YHV) dan WSSV pada awal 1990-an.
Untuk meningkatkan produksi budidaya udang maka pada tahun 2000
Indonesia mengintroduksi udang vaname (Litopenaeusvannamei) sebagai
pengganti udang winduyang terserang WSSV (DKP 2007). Selanjutnya pada 11
Juni 2005, Presiden Republik Indonesia mencanangkan Program Revitalisasi
Pertanian, Perikanan dan Kehutanan secara nasional. Upaya-upaya yang
dilakukan antara lain revitalisasi tambak intensif dengan udang vaname pada
lahan tambak seluas 7000 ha dengan rata-rata produksi 30 ton/ha/tahun,
revitalisasi tambak tradisional dengan udang vaname pada lahan tambak seluas
vaname bebas patogen (Specific Pathogen Free), domestikasi udang vaname
menjadi induk yang bebas penyakit dan tahan penyakit (Specific Pathogen
Resistance) sehingga mengurangi ketergantungan dari impor, melakukan
revitalisasi teknik pembenihan udang skala rumah tangga (backyard hatchery),
penerapan sertifikasi perbenihan dan pembudidayaan udang, pengembangan
laboratorium lingkungan dan penyakit, penyediaan sarana dan prasarana
budidaya, dan membantu penguatan permodalan bagi pembudidaya udang.
Berdasarkan data Kementerian Kelautan dan Perikanan Indonesia, total
produksi budidaya udang nasional Indonesia yang tercatat tahun 2005 sebesar
295000 ton (DKP 2007). Tahun 2008, produksi udang meningkat mencapai
410000 ton namun pada tahun 2009, produksi menurun menjadi 350000 ton atau
turun sekitar 14.6%. T
Agar program peningkatan produksi udang dapat berkesinambungan, baik
secara ekologi maupun ekonomi, maka kontrol penyakit harus menjadi prioritas
utama. Beberapa metoda yang telah diterapkan dalam mengontrol penyakit antara
lain penggunaan antibiotik atau bahan kimia, vaksin, probiotik, penggunaan
SPF/SPR, dan biosekuriti. Penggunaan antibiotik merupakan metoda kontrol
penyakit yang telah lama dan paling banyak diterapkan dalam aktivitas budidaya.
Namun demikian, telah ditemukan bahwa pemberian antibiotik dalam tambak
telah mengakibatkan munculnya patogen yang tahan terhadap antibiotik
(antibiotic-resistant pathogen). Selain itu, pemberian antibiotik dalam tambak
membutuhkan sejumlah besar bahan yang mahal dan dapat terakumulasi dalam
tubuh ikan/udang atau lingkungan budidaya dan membahayakan kesehatan
konsumen. Vaksin telah digunakan pada beberapa spesies ikan dan
memperlihatkan hasil positif. Pada udang, penggunaan vaksin
(formalin-inactivated WSSV, recombinant protein WSSV) telah mulai diteliti dan
memperlihatkan hasil yang menjanjikan meskipun udang tidak memiliki sistem
imun spesifik (Namikoshi et al. 2004; Witteveldt et al. 2003). Namun demikian, ahun 2010 produksi berkisar 352000 ton atau turun dari
target semula 400300 ton. Volume ekspor berkisar 144410 ton atau turun 4.36%
dengan nilai ekspor 1.03 miliar dollar AS. Menurunnya produksi disebabkan
vaksin sangat mahal dan proteksi yang dihasilkan bersifat spesifik sehingga tidak
efektif melawan beberapa patogen secara simultan.
Penggunaan udang SPR dan sistem biosekuriti yang diterapkan beberapa
tahun terakhir ini secara signifikan mampu meningkatkan kelangsungan hidup dan
produksi. Dalam jangka panjang, kedua metoda inipun nampaknya belum mampu
mencegah munculnya wabah penyakit yang terjadi secara berulang (Moss et al.
2006). Dengan adanya mutasi virus, maka udang yang awalnya resisten terhadap
patogen tertentu menjadi rentan terhadap virus baru. Lebih lanjut, kini terdapat
bukti bahwa udang SPR ternyata memiliki pertumbuhan yang lebih kecil
dib&ingkan dengan udang bukan SPR (Henryon et al. 2002; Parenrengi 2010).
Saat ini, penggunaan imunostimulan semakin mendapat perhatian untuk
dikembangkan sebagaimetoda kontrol penyakit dalam budidaya udang. Banyak
bukti telah memperlihatkan bahwa imunostimulan yang ditambahkan dalam pakan
dapat meningkatkan resistensi ikan dan udang terhadap infeksi penyakit melalui
peningkatan respon imun nonspesifik (Pais et al. 2008; Welker et al. 2007).
Udang tidak memiliki respon imun spesifik dan sepenuhnya tergantung
pada respon imun nonspesifik. Nukleotida merupakan imunostimulan yang sudah
diketahui dapat meningkatkan respon imun nonspesifik ikan (Burrels et al. 2001).
Pada udang, belum banyak penelitian tentang penggunaan nukleotida sebagai
imunostimulan.Nukleotida dapat menawarkan alternatif bagi penggunaan
antibiotik atau bahan-bahan kimia sebab bahan ini tidak meninggalkan residu
dalam tubuh ikan serta tidak mengakibatkan kerusakan lingkungan. Oleh karena
itu, penelitian terhadap penggunaan nukleotida dalam upaya mengontrol penyakit
dalam aktivitas budidaya udang dirasa sangat diperlukan.
Pendekatan Masalah
Nukleotida merupakan nutrient semi esensial yang mulai mendapat
perhatian serius untuk dikembangkan penggunaannya dalam budidaya ikan dan
krustase dalam beberapa tahun terakhir ini. Meskipun penelitian tentang
penggunaan nukleotida masih berada pada tahap awal, laporan-laporan penelitian
biosintesa asam amino non esensial, eksogenous nukleotida dapat meningkatkan
imunitas dan resistensi ikan terhadap infeksi virus, bakteri dan parasit. Selain itu,
pemberian nukleotida juga dapat meningkatkan efikasi vaksinasi yang
ditunjukkan oleh peningkatan titer antibodi setelah ikan divaksinasi (Burrells et
al. 2001); meningkatkan oxidative radical neutrofil darah dan sintasan ikan;
meningkatkan aktivitasfagositosis, respiratory burst, serum komplemen dan
aktivitas lisosim serta menurunkan infeksi A. hydrophilapada ikan mas.
Pemberian nukleotida juga dapat memperbaiki pertumbuhan pada fase
perkembangan awal, meningkatkan kualitas larva, serta meningkatkan toleransi
terhadap stres. Pada krustase, pemberian nukleotida dapat meningkatkan
pertumbuhan udang vaname (Li et al. 2007). Sebaliknya, defisiensi nukleotida
dapat merusak fungsi hati, usus, dan fungsi imun (Li & Galtin 2006).
Hasil-hasil penelitian di atas menunjukkan bahwa penambahan nukleotida
dalam pakan dapat meningkatkan imunitas dan resistensi ikan terhadap sejumlah
patogen yang berbeda. Namun demikian, penelitian-penelitian menyangkut
sumber dan tipe nukleotida yang tepat, dosis, dan lama waktu pemberian, masih
sangat terbatas. Pada udang, laporan-laporan penelitian tentang penggunaan
nukleotida masih sangat terbatas. Sehubungan dengan itu, serangkaian penelitian
telah dilakukan untuk mengkaji peranan nukleotida dalam meningkatkan respon
imun nonspesifik dan resistensi udang vaname. Skema pendekatan masalah
disajikan pada Gambar 1.
Perumusan Masalah
Eksogenous nukleotida sangat penting untuk digunakan dalam kontrol
penyakit karena selain dapat memperbaiki pertumbuhan, pemberian nukleotida
dapat meningkatkan imunitas dan resistensi udang yang dibudidayakan. Namun
demikian, informasi menyangkut tipe nukleotida yang tepat, dosis, serta lama
waktu pemberian yang menghasilkan respon optimal pada udang vaname masih
Gambar 1. Skema pendekatan masalah
1. Mengevaluasi pengaruh penambahan nukleotida dengan dosis berbeda dalam pakan terhadap jumlah hemosit, aktivitas PO, resistensi udang vaname terhadap infeksi Vibrio harveyidan pertumbuhan.
Tujuan Penelitian
2. Mengevaluasi pengaruh protokol pemberian nukleotida terhadap jumlah hemosit, aktivitas PO dan pertumbuhan udang vaname.
3. Mengevaluasi pengaruh suplementasi β-glukan dalam pakan terhadap jumlah hemosit, aktivitas PO,resistensi dan pertumbuhan udang vanamesebagai pembanding terhadap nukleotida.
4. Mengevaluasi pengaruh pemberian nukleotida terhadap resistensi dan performa pertumbuhan udang vaname yang dipelihara di tambak.
Respon humoral
Respon imun nonspesifik
Respon selular (+)
(-)
(-)
(+)
(+)
Produksi
Resistensi (-)
Pertumbuhan
Udang
Nukleotida
Manfaat Penelitian
Hasil penelitian diharapkan dapat dijadikan bahan aplikasi yang penting
bagi manajemen kesehatan dalam industri budidaya udang vaname di Indonesia.
Hipotesis
• Pemberian nukleotida sebagai imunostimulan dalam pakan sangat potensial meningkatkan respon imun nonspesifik dan resistensi udang vaname.
• Respon imun nonspesifik udang vaname dipengaruhi oleh dosis dan lama waktu pemberian imunostimulan nukleotida.
Kebaharuan
Kebaharuan dari penelitian ini adalah pemanfaatan nukleotida sebagai
sumber alternatif imunostimulan dan peranannya dalam sistem imun nonspesifik
udang yaitu meningkatkan jumlahhemosit, aktivitas phenoloxidase (PO) dan
resistensi terhadap infeksi patogen. Nukleotida juga berperan memacu
pertumbuhan. Dengan demikian maka aplikasi nukleotida dalam pakan udang
diharapkan mampu mengatasi atau meminimalisir permasalahan penyakit dan
TINJAUAN PUSTAKA
Udang vaname termasuk krustase dalam ordo dekapoda dimana di
dalamnya juga termasuk udang, lobster dan kepiting. Klasifikasi udang vaname
adalah sebagai berikut (Wyban & Sweeney 1991):
Biologi Litopenaeus vannamei (Boone 1931)
Phylum :Anthropoda
Subphylum : Krustase
Class : Malacostraca
Subclass : Eumalacostraca
Superorder : Eucarida
Order : Decapoda
Suborder : Dendrobranchiata
Super Family : Penaeidea
Family : Penaeidae
Genus : Litopenaeus
Spesies : L. vannamei
Nama umum udang vaname adalah Pasific white shrimp, West Coast white
shrimp, Camaron blanco, Langostino. Nama FAO adalah whiteleg shrimp,
Crevette pattes blanches, Camaron patiblanco (Elovaara 2001; Rosenberri 2006).
Ciri-ciri udang vanameadalah rostrum bergigi, biasanya 2-4
(kadang-kadang 5-8) pada bagian ventral yang cukup panjang dan pada udang muda
melebihi panjang antennular peduncle (Gambar 2).Karapaks memiliki
pronounced antenal dan hepatic spines. Pada udang jantan dewasa, petasma
symmetrical, semi-open, dan tidak tertutup. Spermatofora sangat kompleks yang
terdiri atas masa sperma yang dibungkus oleh suatu pembungkusyang
mengandung berbagai struktur perlekatan (anterior wing, lateral flap, caudal
flange, dorsal plate) maupun bahan-bahan adhesif dan glutinous. Udang betina
dewasa memiliki open thelycumdan sternit ridges, yang merupakan pembeda
Gambar 2 Morfologi Litopenaeus vannamei (Sumber: Wyban & Sweeney 1991)
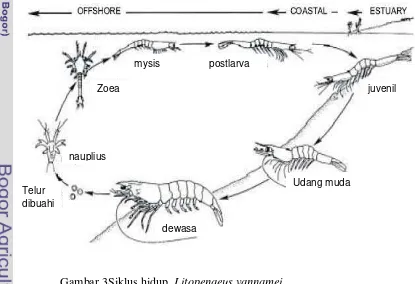
Udang vaname memiliki 6 fase nauplii, 3 fase protozoea dan 3 fase mysis
dalam siklus hidupnya. Fase larva (panjang karapaks 1,95 – 2,73 mm) dapat
dikenal melalui kurangnya spine pada sternit ke 7, dan panjang rostrum relatif
terhadap panjang mata termasuk tangkai mata.Ciri morfologi yang paling dapat
dikenal adalah perkembangan supraorbital spine pada fase zoea ke 2 dan ke 3.
Tubuh berwarna putih transparan sehingga lebih umum dikenal sebagai
“white shrimp”. Tubuh sering berwarna kebiruan karena lebih dominannya
kromatofor biru. Panjang tubuh dapat mencapai 23 cm. Udang vaname dapat
dibedakan dengan spesies lainnya berdasarkan pada eksternal genitalnya.
Distribusi
Udang vaname tersebar di bagian timur pantai Pasifik Amerika Tengah
dan Selatan dari Mexico sampai Peru(Elovaara 2001; Rosenberry 2006), dimana
& Sweeney 1991). Karena spesies ini relatif mudah dibudidayakan, maka udang
ini telah tersebar keseluruh dunia.
Habitat
Di alam udang ini menyukai dasar berlumpur pada kedalaman dari garis
pantai sampai sekitar 72 m. Hewan ini juga telah ditemukan menempati daerah
mangrove yang masih belum terganggu. Udang ini nampaknya dapat beradaptasi
dengan perubahan temperatur dan tekanan di alam. Hasil penelitian menunjukkan
bahwa udang vanamedapat beradaptasi dengan baik pada level salinitas yang
sangat rendah sehingga menjadikan udang ini sebagai udang yang paling banyak
dibudidayakan di kolam air tawar (salinitas sangat rendah dimana udang ini dapat
beradaptasi (Elovaara 2001).
Molting dan Pertumbuhan
Pertumbuhan udang vaname, seperti halnya arthropoda lainnya, tergantung
pada dua faktor yaitu frekuensi molting (waktu antara molting) dan pertumbuhan
yaitu berapa besar pertumbuhan pada setiap molting baru (Wyban & Sweeney
1991). Karena tubuh udang ditutupi oleh karapaks yang keras, maka untuk
tumbuh, karapaks yang lama harus dilepas dan diganti dengan yang baru dan lebih
besar. Saat molting, terjadi pemisahan kulit antara karapaks dan intercalary
sclerite, dimana sepalotoraks dan appendic anterior dikeluarkan. Karapaks baru
pada awalnya lunak, tetapi akan mengeras kembali pada laju yang proporsional
terhadap ukuran udang.
Molting merupakan fungsi dari ukuran udang, jika udang tumbuh maka
waktu antar molting meningkat. Pada fase larva, molting terjadi setiap 30-40 jam
pada temperatur 28°C. Juvenil udang ukuran 1–5 gram akan molting setiap 4-6
hari, tetapi udang berukuran 15 gram akan molting setiap 2 minggu.
Kondisi lingkungan dan faktor nutrisi juga mempengaruhi frekuensi
molting. Pada temperatur yang lebih tinggi, frekuensi molting meningkat.
Selama molting, absorpsi oksigen menjadi kurang efisien dan udang yang mati
molting, karapaks masih lunak dan udang menjadi rentan terhadap predasi dari
sesamanya. Udang yang baru molting dengan kulit yang masih lunak sering
membenamkan diri dalam endapan detritus yang lunak.
Reproduksi dan Siklus Hidup
Karapaks udang vanameberwarna transparan sehingga memungkinkan
untuk mengamati warna perkembangan ovari. Pada betina, gonad pertama-tama
berwarna keputih-putihan, selanjutnya berkembang menjadi coklat emas atau
coklat kebiru-biruan pada saat akan memijah (Rosenberry 2006). Udang jantan
menyimpan spermatophora pada betina berkulit keras. Tingkah laku kawin
dimulai pada sore hari dimana hal ini berkaitan dengan ketersediaan intensitas
cahaya. Proses pemijahan dimulai dengan lompatan secara tiba-tiba dan udang
betina aktif berenang. Seluruh proses pemijahan berakhir selama sekitar satu
menit. Jumlah telur yang dapat dilepaskan seekor induk betina bervariasi menurut
ukuran individu. Udang berukuran 30–45 gr dapat melepaskan 100000–250000
butir telur. Ukuran diameter telur sekitar 0.22 mm.
Udang betina memiliki open thelycum dan inilah yang membedakannya
dengan dengan udang penaeid lainnya (Elovaara 2001). Udang jantan melekatkan
spermatophora berjeli (berisi sperma) pada open thelycum pada saat kawin.
Perkawinan terjadi pada saat udang betina berada pada fase intermolt pada saat
ovari telah mencapai kematangan. Pelepasan telur terjadi pada malam hari
beberapa jam setelah perkawinan, biasanya kurang dari tiga jam. Proses pelepasan
telur berlangsung selama 1-3 menit dimana selama proses pelepasan telur, induk
betina melindungi telur yang baru dilepaskan. Hal ini memungkinkan sperma
untuk membuahi telur sebanyak mungkin. Segera setelah semua bahan genetik
dari jantan maupun betina bersatu maka pembuahanpun selesai.
Telur akan menetas menjadi nauplii dalam waktu sekitar 16-17 jam setelah
pembuahan. Jika diamati di bawah mikroskop,nauplii secara fisik nampak seperti
laba-laba air. Selama beberapa hari nauplii makan dari makanan cadangan dari
telur sampai nauplii bermetamorfosa menjadi zoeae sebagai tahap larva yang
mysis. Pada fase mysis, larva sudah mulai nampak seperti bentuk udang dewasa.
Selain mikro algae, mysis memakan diatom dan zooplankton, terutama di alam.
Fase mysis berlangsung selama 4 hari sampai mysis bermetamorfosa kembali
menjadi postlarva. Post larva telah berbentuk seperti udang dewasa, memakan
zooplankton, detritus dan berbagai formula makanan buatan jika dipelihara dalam
hatchery.
Di alam, udang dewasa mencapai matang gonad, kawin dan bertelur di laut
terbuka sampai pada kedalaman sekitar 70 m pada temperatur 26-28oC dan salinitas sekitar 35 ppt. Setelah menetas, larva berkembang di perairan lepas
pantai ini dan setelah mencapai post larva, udang bermigrasi ke perairan pantai
dan menetap di dasar estuari yang dangkal. Setelah beberapa bulan di daerah
estuari, udang dewasa kembali bermigrasi ke perairan laut terbuka dimana
[image:31.595.89.504.417.701.2]selanjutnya terjadi kematangan gonad, perkawinan, dan pemijahan (Gambar 3).
Gambar 3Siklus hidup Litopenaeus vannamei (Sumber: diadaptasikan dari Braak 2002)
postlarva
Zoea
mysis
nauplius
dewasa
Udang muda Telur
dibuahi
Makanan dan Kebiasaan Makan
Di alam, udang penaeid bersifat karnivor yang memangsa krustase kecil,
ampipoda, polikaeta. Namun dalam tambak, udang ini makan makanan tambahan
atau detritus. Udang vanamebersifat nokturnal. Udang muda tetap membenamkan
diri dalam substrat selama siang hari dan tidak makan atau tidak mencari
makanan. Tingkah laku makan ini dapat diubah dengan pemberian pakan ke
dalam tambak. Hasil penelitian di Ocean Institute Honolulu menunjukkan bahwa
udang yang diberi pakan beberapa kali sehari tumbuh lebih cepat dibandingkan
dengan udang yang hanya diberi pakan sekali dalam satu hari (Wyban & Sweeney
1991).
Krustase tidak memiliki respon imune spesifik (adaptive) dan nampak
bergantung pada berbagai respon imun nonspesifik (innate). Meskipun dianggap
tidak begitu memuaskan, respon imun nonspesifik mampu dengan cepat dan
efisien mengenal dan menghancurkan material asing, termasuk patogen
(Vargas-Albores & Yepiz-Plascencia 2000;Witteveldt et al. 2003). Respon imun
nonspesifik terdiri atas respon selular dan respon humoral. Respon Imun Krustase
Hemositkrustase, dan invertebrata lain, memainkan peranan penting dalam
sistem pertahanan tubuh terhadap patogen seperti virus, bakteri, fungi, protozoa,
dan metazoa (Johansson et al. 2000; Sindermann 1990; Rodriquez & Le Moullac
2000). Pertama, hemosit mengeluarkan partikel asing dalam hemocoel melalui
fagositosis, enkapsulasi dan aggregasi nodular. Kedua, hemosit berperan dalam
penyembuhan luka melalui cellular clumpingserta membawa dan melepaskan
prophenoloxidase system (proPO). Hemosit juga berperan dalam sintesa dan
pelepasan molekul penting hemolim seperti α Respon Selular
2-macroglubulin (α2
Klasifikasi tipe hemositkrustase terutama didasarkan pada keberadaan
granula sitoplasma, yaitu sel hyalin, semigranular, dan granular (Johansson et al.
2000; Le Moullac & Haffner 2000). Sel hyalin merupakan tipe sel yang paling M), agglutinin,
kecil dengan ratio nukleus sitoplasma tinggi dan tanpa atau hanya sedikit granula
sitoplasma; sel granular merupakan tipe sel paling besar dengan nukleus yang
lebih kecil dan terbungkus dengan granula; sel semi granular merupakan tipe sel
diantara hyalin dan granular. Masing-masing tipe sel aktif dalam reaksi kekebalan
tubuh, sebagai contoh, sel hyalin terlibat dalam fagositosis, sel semigranular aktif
dalam enkapsulasi, sel granular aktif dalam penyimpanan dan pelepasan proPO
system dan sitotoksisiti.
Jumlah hemosit dapat sangat bervariasi berdasarkan spesies, respon
terhadap infeksi, stres lingkungan, aktivitas endokrin selama siklus molting
(Johansson et al. 2000), seks, fase perkembangan, status reproduksi dan nutrisi
(Song et al. 2003). Pada kuruma shrimp (Marsupenaeus japonicus), total
hemocyte count (THC) sebanyak 1.7x107 sel ml-1, pada L. stylirostris 1.84x107 sel ml-1, P. monodon berkisar 2.10x107 sel ml-1 (flow cytometry) sampai 2.33x107 sel.ml-1 (hemacytometer). Hasil penelitian Song et al. (2003) menunjukkan bahwa setelah 3-5 hari diinfeksi dengan Taura Syndrome Virus (TSV), THC L.
vannamei berukuran 10-20 g mengalami penurunan sebesar 70% menjadi
0.345x107 sel ml-1 dibandingkan dengan kontrol 1.64x107 sel.ml-1
Pada M. japonicus dan L. stylirostris, jumlah hemosit terbesar ditemukan
pada fase postmoult dan terendah pada fase intermoult. Sel granular tertinggi
dilepaskan pada fase postmoult pada L. stylirostris dan S. ingentis sedangkan sel
hyalinmencapai puncak selama proses ganti kulit pada S. ingentis dan M.
japonicus. Adanya sel hyalin yang tinggi selama proses ganti kulit nampaknya
penting sebab mereka mengawali koagulasi dan mungkin terlibat dalam
pembentukan kulit. Konsentrasi sel granular yang tinggi dalam hemolimL.
stylirostris selama fase intermoult berhubungan dengan aktivitasphenoloxidase
(PO) yang tinggi dan resistensi terhadap vibriosis (Le Moullac et al. 1997)
, dengan
mortalitas mencapai 80%. Dalam kondisi hypoxia, THC L. styloristris turun
menjadi rendah serta udang menjadi lebih sensitif terhadap infeksi V.
aglinolyticus. Differential hemosit count (DHC) juga berubah (sel granular, semi
granular, dan hyalin) dengan perubahan besar terjadi pada sel hyalindan
Fagositosis merupakan reaksi yang paling umum dalam pertahanan selular
udang. Proses fagositosis dimulai dengan perlekatan (attachment) dan penelanan
(ingestion) partikel mikroba ke dalam sel fagosit. Sel fagosit kemudian
membentuk vacuola pencernaan (digestive vacuola) yang disebut
fagosom(Rodriquez & Le Moullac 2000). Lisosom(granula dalam sitoplasma
fagosit) kemudian menyatu dengan fagosom membentuk fagolisosom.
Mikroorganisme selanjutnya dihancurkan dan debris mikroba dikeluarkan dari
dalam sel melalui proses egestion (Gambar 4). Pemusnahan partikel mikroba
yang difagosit melibatkan pelepasan enzim ke dalam fagosom dan produksi ROI
(reactive oxygen intermediate) yang kini disebut respiratory burst (Rodriquez &
Le Moullac 2000; Sindermann 1990).
Gambar 4 Proses fagositosis
(Sumber:http://www.cliffsnotes.com/WileyCDA/CliffsReviewTopic/P hagocytosis.topicArticleId-8524)
Hemosit berfungsi dalam enkapsulasi. Hal ini terjadi pada organisme
yang memiliki tubuh terlalu besar untuk fagositosis. Pada saat hemosit
mengelilingi tubuh benda asing yang besar, bagian sel terluar dari hemosit tetap
berbentuk oval atau bulat sedangkan bagian tengah sel menjadi datardan pada fase
tersebut tidak diserap kembali dan tetap sebagai tanda enkapsulasi meskipun
sudah tidak ada hemosit yang dikenal disitu. Hemosit juga berfungsi dalam
formasi melanin pada fase akhir penyembuhan atau perbaikan luka. Enzim yang
terlibat dalam formasi melanin adalah phenoloxidase (PO) dan telah ditemukan
terdapat dalam hemolim dan kulit arthropoda (Sritunyalucksana & Söderhäll
2000).
Proses imun pertama pada krustase adalah pengenalan mikroorganisme
penyerang yang dimediasi oleh hemosit dan plasma protein (Bachere 2000).
Beberapa tipe modulator protein telah diketahui dapat mengenal komponen
dinding sel mikroorganisme seperti β-1,3-glucan-binding protein (BGBP),
lipopolysaccharide-binding protein (LPS-BP), hemosit receptor yang mengikat
plasmatic glucan-binding protein (PGBP) setelah PGBP bereaksi dengan β -1,3-glukan; peptidoglycan recognition protein yang mampu mengaktifkan
phenoloxidase. Respon Humoral
Enzim phenoloxidase (PO) terdapat dalam hemolim sebagai inactive
pro-enzyme yang disebut proPO. Transformasi proPO menjadi PO melibatkan
beberapa reaksi dikenal sebagai proPO activating system (sistem aktivasi proPO).
Sistem ini terutama diaktifkan oleh beta glukan, dinding sel bakteri dan LPS.
Sistem aktivasi proPO dipertimbangkan sebagai bagian dari sistem imun yang
mungkin bertanggung jawab terhadap proses pengenalan benda asing dalam
sistem pertahanan krustase dan insekta. Sistem proPO dapat digunakan sebagai
marker kesehatan udang dan lingkungan karena perubahan sistemproPO
berkorelasi dengan tahap infeksi dan variasi lingkungan.
Enzim phenoloxidase (PO) bertanggung jawab terhadap proses melanisasi
pada artropoda (Rodriquez & Le Moullac 2000). Enzim ini mengkatalis
hidroksilasi monophenol dan oksidasi phenol menjadi quinones yang diperlukan
untuk proses melanisasi sebagai respon terhadap penyerang asing dan selama
proses penyembuhan (Sritunyalucksana & Söderhäll 2000; Vargas-Albores &
non-enzymatic menjadi melanin dan sering dideposit pada benda yang dienkapsulasi,
dalam nodulhemosit, dan pada daerah kulit yang terinfeksi jamur. Produksi
reactive oxygen species seperti superoxide anion dan hydroxyl radical selama
pembentukan quonoid juga memainkan peranan penting sebagai antimikroba.
Reaksi biologi seperti fagositosis, enkapsulasi dan nodulasi juga diaktifkan.
Vaksinasi mungkin dapat meningkatkan aktivitashemosit, fagositosis dan
aktivitas opsonin. Pada invertebrata yang tidak memiliki antibodi, lektin
berfungsi sebagai molekul pengenal (recognition molecules) untuk aktivitas
pertahanan seperti agregasi dan opsonisasi (Wittevelt et al. 2003). Lektin
merupakan suatu set protein yang secara spesifik mengikat pada molekul gula
termasuk glikoprotein dan glikolipid. Hasil uji coba Namikoshi et al. (2004)
menunjukkan bahwa penggunaan formalin-inactivatedWSSV vaksin dapat
meningkatkan resistensi P. japonicus terhadap WSSV sepuluh hari setelah
divaksinasi dengan metoda vaksinasi intramuskular. Hasil penelitian yang
dilakukan oleh Wittevelt et al. (2003) juga memperlihatkan bahwa penggunaan
WSSV subunit vaksin dapat meningkatkan resistensi udang windu terhadap
WSSV meskipun udang tidak memiliki respon imun spesifik.
Imunostimulan
Sejumlah substan biologi dan sintetik telah ditemukan dapat meningkatkan
sistemimun nonspesifik ikan. Bahan tersebut dapat meningkatkan resistensi ikan
dan udang terhadap infeksi sejumlah patogen secara simultan(Kumari et al. 2003;
Raa et al. 1992). Oleh karena itu maka imunostimulan sangat penting untuk
digunakan dalam kontrol penyakit karena menawarkan suatu alternatif terhadap
penggunaan antibiotik yang saat ini banyak digunakan dalam budidaya ikan dan
krustase (Cook et al. 2003; Yin et al. 2006), serta tanpa efek samping (Yin et al.
2006).
Secara sederhana, imunostimulan merupakan suatu substan yang
merangsang atau meningkatkan sistem imun dengan berinteraksi secara langsung
dengan sel-sel yang mengaktifkan sistem imun (Gannam & Schrok 2001).
dengan cara meningkatkan aktivitas sel-sel fagosit(Yin et al. 2006). Jadi
imunostimulan meningkatkan resistensi ikan atau udang terhadap patogensecara
simultan dengan cara merangsang respon imun nonspesifik (Gannam & Schrok
2001). Imunostimulan dapat berupa bakteri dan produk bakteri, yeast, kompleks
karbohidrat, faktor nutrisi, ekstrak hewan, ektrak tumbuhan, dan obat-obatan
sintetik (Cook et al. 2003; Sakai 1999; Sealey & Gatlin 2001).
Nukleotida merupakan nutrien semi esensial bagi ikan dan krustase.
Nukleotida memiliki peranan penting dalam fisiologi dan biokimia seperti
penandaan (encoding) dan penerusan informasi genetik, memediasi energi
metabolisme dan cell signalling serta sebagai komponen koensim, allosteric
effectors, dan cellular agonist (Li & Galtin 2006). Nukleotida terdiri atas basa
purin atau pirimidin,ribosa atau 2’-deoksiribosa dan satu atau lebih grup fosfat.
Basa purin yang utama terdiri atas adenin, guanin, hiposantin dan santin.
Nukleosida purinmengandung ribosa atau 2-deoksiribosa yang berikatan dengan
cicin purin melalui ikatan glikosidik (glycosidic bond) pada N-9. Nukleotida
merupakan fosfat ester dari nukleosida. Basa pirimidinyang utama terdiri atas
urasil, timin, dan sitosin. Urasil dan sitosin merupakan komponen pirimidin utama
dari RNA.Nukleosida pirimidin atau nukleotida mengandungribosa atau
2’-deoksiribosayang berikatan dengan pirimidin melalui ikatan glikosidik pada N-1.
Fosfat esterdari nukleosida pirimidin adalah UMP, CMP, dan TMP (Devlin 2002;
Li & Galtin 2006).Secara alami nukleotida terdapat dalam semua makanan yang
berasal dari hewan dan tumbuhan dalam bentuk nukleotida bebas dan asam
nukleat.
Purin dan pirimidin disintesa dari de novo pathway atau diperoleh dari
salvage pathway.Purin disintesa dalam sitosol sel mamalia dari glycine, aspartate,
glutamine, turunan tetrahydrofolate dan CO2 dengan input energi yang besar. Pirimidin disintesa dari aspartate, glutamine, dan CO2
Kebutuhan nukleotida untuk fungsi fiosiologi hewan dapat dipenuhi dari
sintesa de novo. Namun demikian, suplai nukleotida dari sintesa tersebut tidak
cukup untuk menjalankan fungsi fisiologi secara optimal terutama pada dalam sitosol dan
sistemimun pada saat berada dalam kondisi stres (Li et al. 2004). Dalam
akuakultur, stres akibat penanganan (handling), penyortiran (grading),
pengangkutan, kepadatan tinggi, penyakit, dan kualitas air yang kurang baik
merupakan masalah yang umum terjadi dan karenanya penambahan nukleotida
dalam pakan mungkin diperlukan (Burrells et al. 200; Li et al. 2004).
Hasil-hasil penelitian pada manusia dan hewan ternak memperlihatkan
bahwa penambahan nukleotida dalam pakan dapat meningkatkan cell-mediated
immunity(CMI), proliferasi limfosit, interleukin-2, dan meningkatkan resistensi
terhadap infeksi bakteri (Li et al. 2004). Sebaliknya, hewan yang diberi pakan
yang tidak mengandung nukleotida menderita gangguan pada fungsi imun selular
dan humoral seperti penurunan aktivitasNK-cell dan makrofag, produksi sitokin
rendah, penurunan produksi antibodi, dan suseptibilitas terhadap infeksi
meningkat (Field et al. 2002). Penambahan nukleotida dalam pakan dapat
memperbaiki kondisi tersebut. Pada bayi yang diberi susu ibu atau makanan yang
ditambahkan nukleotida memiliki aktivitasNK-cell dan produksi IL-2 yang lebih
tinggi dibandingkan dengan bayi yang diberi makanan tanpa suplementasi
nukleotida. Makanan anak-anak yang tersedia saat ini umumnya telah
ditambahkan nukleotida. Bagaimana mekanisme nukleotida dalam meningkatkan
fungsi imun belum diketahui dan perlu diteliti lebih lanjut.
Perhatian terhadap suplementasi nukleotida sebagai imunostimulan pada
pakan ikan mulai meningkat sejak adanya laporan Burrels et al. (2001) yang
memperlihatkan bahwa pakan yang ditambahkan nukleotida dapat meningkatkan
resistensi ikan terhadap infeksi virus, bakteri dan parasit. Nukleotida dapat juga
meningkatkan pertumbuhan serta meningkatkan toleransi ikan terhadap stres.
Pada udang, nukleotida merupakan nutrient kunci (key nutrient) bagi sistem imun
udang dan pemberian nukleotida seperti yeast atau ekstrak yeast dapat
JUDUL 1 RESPON IMUN NONSPESIFIK DAN RESISTENSI
UDANG VANAME (
Litopenaeus vannamei
) YANG DIBERI
PAKAN BERSUPLEMEN NUKLEOTIDA
Abstrak
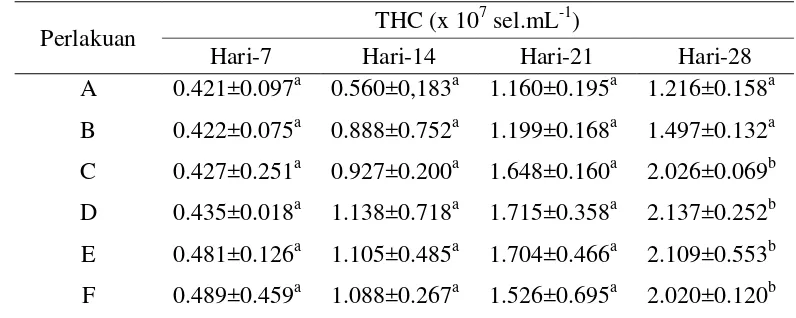
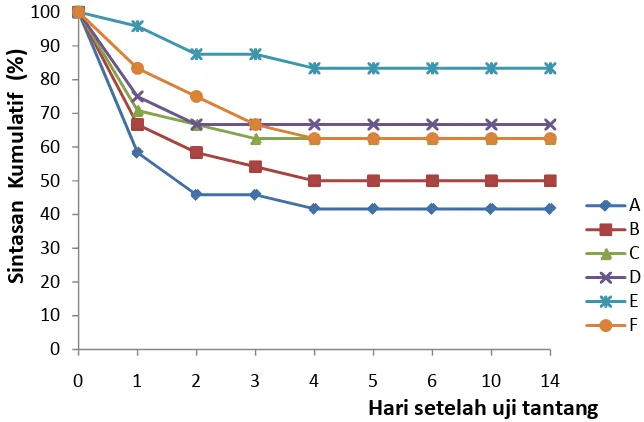
Efikasi nukleotida yang ditambahkan dalam pakan untuk meningkatkan respon imun nonspesifik, resistensi dan pertumbuhan udang vaname(Litopenaeus vannamei) telah diteliti. Juvenil udang berukuran rata-rata 6.0±0.5 g diberi pakan pelet komersial yang sebelumnya telah ditambahkan nukleotida dengan 6 dosis berbeda yakni A: 0, B:100, C:200, D:300, E:400, dan F:500 mg.kg-1 pelet masing-masing dengan tiga ulangan. Udang diberi pakan 3 kali per hari selama 4 minggu berturut-turut dengan tingkat pemberian 3%/bb/hari.Pada akhir periode pemberian pakan, udang diuji tantang melalui injeksi intramuskular dengan Vibrio harveyi 0.1x106 cfu.udang-1. Nukleotida yang ditambahkan dalam pakan memperlihatkan pengaruh signifikan pada udang setelah diberikan selama empat minggu. Jumlah total hemosit(THC) udang pada perlakuan D (300 mg nukleotida) meningkat 76% lebih tinggi dibandingkan dengan kontrol. Aktivitas PO udang pada perlakuan D (300 mg nukleotida) juga berbeda nyata (p=0.005) dibandingkan dengan kontrol namun tidak berbeda dibandingkan dengan perlakuan E (400 mg nukleotida) dan F (500 mg nukleotida). Resistensi yang diukur berdasarkan sintasan rata-rata (83.33±7.21%) pada perlakuan E(400 mg nukleotida) empat belas hari setelah uji tantang berbeda nyata (p=0.005) dibandingkan dengan kontrol, namun tidak berbeda nyata jika dibandingkan dengan perlakuan D (300 mg nukleotida).Pertumbuhan udang yang diamati sampai akhir periode pemberian pakan pada perlakuan E (400 mg nukleotida) berbeda nyata (p=0.001) dibandingkan dengan kontrol maupun dengan perlakuan lainnya. Sebagai kesimpulan, penambahan nukleotida pada dosis300 mg.kg-1 pelet selama 4 minggu dapat meningkatkan jumlah hemosit dan aktivitas PO, sedangkan resistensi dan pertumbuhan terbaik dicapai pada dosis 400 mg.kg-1 pakan.
Kata kunci: Aktivitas PO, Litopenaeus vannamei, nukleotida, resistensi, total hemocyte count
TOPIC 1 NONSPECIFIC IMMUNE RESPONSE AND RESISTANCE OF WHITELEG SHRIMP (Litopenaeus vannamei) FED
NUCLEOTIDE- SUPPLEMENTED DIET
Abstract
with 0.1 mlVibrio harveyi solution containing 1x106 cfu. Nucleotides diet showed significant effect on shrimp after four weeks of feeding. Mean THC per ml of hemolymph in treatment D(300 mg nucleotides) increased up to 76% higher than control. PO activity of shrimp in treatment D (300 mg nucleotides) was also significantly different (p=0.005) compared to control shrimp, but not different as compared to those of treatments E (400 mg nucleotides) and F (500 mg nucleotides). Resistancemeasured as survival rate (83.33±7.21%) of shrimp in treatment E (400 mg nucleotides) fourteen days post-challenge with V. harveyi was significantly different (p=0.005) from that of control, but not different compared to treatment D (300 mg nucleotides). Shrimp growth observed in treatment E (400 mg nucleotides) was significantly higher (p=0.001) than those of other treatments. As conclusion, oral administration of nucleotides at 300 mg.kg-1 pellet for four weeks enhanced total hemocyte count dan PO activity while the highest disease resistance and growth were obtained at 400 mg.kg-1 diet.
Keywords: resistance, Litopenaeus vannamei, nucleotides, total hemocyte count, PO activity
PENDAHULUAN
Budidaya udang mulai berkembang dengan pesat di Indonesia sejak tahun
1980 setelah pemerintah mengeluarkan larangan terhadap pengoperasian alat
tangkap trawler di beberapa daerah di Indonesia, sekaligus mencanangkan
Program Budidaya Udang Nasional. Namun sejak awal tahun 1990, budidaya
udang menghadapi kegagalan produksi akibat munculnya serangan penyakit
terutama virus (MBV) dan vibriosis. Kondisi ini lebih diperburuk lagi dengan
mewabahnya WSSV di Indonesia sejak tahun 1992/1993. Sejak ditemukan pada
tahun 1992, WSSV telah menyebabkan masalah penyakit yang serius dan telah
menyebabkan kerugian ekonomi yang signifikan bagi industri budidaya udang
dengan mortalitas kumulatif mencapai 100% (Wang et al. 1999; Witteveldt et al.
2003). Masalah ini terjadi bukan hanya di Indonesia tetapi juga di negara-negara
produsen lain seperti di Thailand dan Filipina. Pada tahun 2006, penyakit virus
yang baru, infectious myonecrosis virus (IMNV), terdeteksi menyerang budidaya
udang vaname di Indonesia (Taukhid dan Nuraini 2008).
Sejumlah pendekatan telah diterapkan dalam upaya mengatasi masalah
penyakit. Belakangan ini, berbagai bahan kimia, polisakarida, ekstrak tumbuhan
sebagai imunostimulan (Misra et al. 2006;Pais et al. 2008). Nukleotida
merupakan nutrient semi esensial yang mulai mendapat perhatian serius untuk
dikembangkan penggunaannya sebagai imunostimulan dalam budidaya ikan dan
krustase dalam beberapa tahun terakhir ini. Nukleotida memiliki fungsi penting
dalam fisiologi dan biokimia seperti penandaan (encoding) dan penerusan
informasi genetik, memediasi energi metabolisme dan cell signalling maupun
sebagai koensim, allosteric effectors, dan cellular agonist (Galtin& Li 2007).
Evaluasi terhadap penggunaan nukleotida sebagai suplemen pakan ikan
telah mulai dilakukan sejak awal tahun 1970-an. Penelitian-penelitian pada saat
itu terutama bertujuan untuk mengevaluasi kemungkinan penggunaan nukleotida
sebagai atraktan pakan (Li & Galtin 2006). Pada hewan ternak, nukleotida sudah
lama dimanfaatkan sebagai atraktan pakan. Beberapa produk nukleotida komersil
untuk ternak telah tersedia di pasaran dengan merek dagang ascogen dan optimun
(Chemoforma, Switzerland).
Beberapa laporan penelitian yang sudah ada menunjukkan bahwa selain
terlibat dalam palatabilitas pakan dan biosintesa asam amino non esensial,
eksogenous nukleotida dapat meningkatkan respon imun dan resistensi ikan
terhadap sejumlah patogen secara simultan (Burrels et al. 2001). Selain itu,
pemberian nukleotida juga dapat meningkatkan efikasi vaksinasi, meningkatkan
pertumbuhan, meningkatkan kualitas larva, serta meningkatkan toleransi terhadap
stres. Hasil pengamatan Leonardi et al. (2003) menunjukkan bahwa pemberian
nukleotida (optimun) pada atlantik salmon menurunkan level serum cortisol dan
menghasilkan peningkatan resistensi terhadap penyakit.
Pada udang, laporan-laporan penelitian tentang penggunaan nukleotida
masih belum tersedia, atau jika ada, masih sangat terbatas. Disisi lain, nukleotida
sangat aman digunakan dalam kontrol penyakit sebab bahan ini selain tidak
meninggalkan residu dalam tubuh, juga tidak mengakibatkan terjadinya kerusakan
lingkungan. Penelitian tentang penggunaan nukleotida dalam budidaya udang
juga dirasa sangat diperlukan untuk menyediakan solusi praktis guna mengurangi
resiko terjadinya serangan penyakit infeksius maupun kerusakan lingkungan
nukleotida dengan dosis berbeda dalam pakan terhadap jumlah hemosit, aktivitas
PO, resistensi terhadap infeksi Vibrio harveyidan pertumbuhan udang vaname.
BAHAN DAN METODE
Tempat dan Waktu Penelitian
Penelitian dilaksanakan di laboratorium Kesehatan Ikan, Departemen
Budidaya Perairan, Fakultas Perikanan dan Ilmu Kelautan, Institut Pertanian
Bogor. Penelitian dilaksanakan pada bulan Januari – Maret 2009.
Hewan Uji
Hewan uji adalah juvenil udang vaname yang diperoleh dari fasilitas
pembesaran udang vaname di areal pertambakan Bakauheni, Lampung
Selatan.Udang yang dikumpulkan dimasukkan dalam kotak styrofoam yang
dilengkapi aerator baterei kemudian diangkut ke Laboratorium Kesehatan Ikan
Institut Pertanian Bogor menggunakan mobil.
Nukleotida
Nukleotida yang digunakan adalah nukleotida murni (Sigma-Aldrich)
yang terdiri atas adenosine monophosphate (AMP), guanosine monophosphate
(GMP), cytidine monophosphate (CMP), uridinemonophosphate (UMP), dan
inosinemonophosphate (IMP).
Persiapan Pakan Uji
Sebelum dicampurkan ke dalam pakan, kelima jenis nukleotida dalam
jumlah yang sama (1:1:1:1:1) dicampur terlebih dahulu secara merata.
Selanjutnya campuran nukleotida ditimbang sesuai dosis yang dibutuhkan
(perlakuan), dilarutkan dalam sedikit air, dan dicampurkan ke dalam pakan
komersilsecara merata. Pakan kemudian dikering-anginkan dalam temperatur
ruang. Setelah kering, putih telur (sebagai coater) dicampurkan secara merata ke
dalam campuran pakan-nukleotida, dan dikering-anginkan kembali. Pelet
selanjutnya dimasukkan dalam kantong plastik kemudian disimpan dalam lemari
memiliki komposisi: protein 38%, lemak 6%, serat kasar 3%, abu 13% dan kadar
air 11%.
Sebelum pakan yang telah ditambahkan nukleotida ini digunakan dalam
percobaan maka dilakukan uji ketahanan dalam air (water stability). Uji ini
dilakukan dengan cara memasukkan 5 g pakan ke dalam sebuah kotak berukuran
10x10x10 cm. Rangka kotak terbuat dari kawat sedangkan dindingnya berupa
kain saring berdiameter 0.5 mm. Pakan yang telah ditambahkan nukleotida
dimasukkan ke dalam kotak, kemudian kotak dimasukkan dalam akuarium dalam
posisi menggantung dekat dasar. Akuarium berisi 50 l air, diberi aerasi dan
menggunakan resirkulasi air sehingga kotak akan bergerak-gerak mengikuti
gerakan air. Kondisi ini dibuat sama seperti pada akuarium percobaan. Setelah 3
jam, kotak dikeluarkan, pakan sisa dikeringkan pada 60o
Rancangan Percobaan
C selama 4 jam,
kemudian ditimbang. Water stability(Ketahanan dalam air) merupakan ratio antara
berat kering pakan sisa setelah direndam dalam air dalam waktu tertentu dan berat
kering pakan sebelum direndam di kali 100%. Water stability yang diperoleh
adalah 77.4% (3096 g/4 g x 100).
Penelitian dikerjakan dengan menggunakan rancangan acak lengkap
(RAL) dengan enam perlakuan dimana masing-masing perlakuan memiliki 3
ulangan. Penempatan perlakuan ke dalam unit-unit percobaan dilakukan secara
acak. Perlakuan yang digunakan adalah sebagai berikut:
A: Nukleotida 0 mg.kg-1 B:
pakan (pakan standar)
Nukleotida 100 mg.kg-1 C:
pakan
Nukleotida 200 mg.kg-1 D:
pakan
Nukleotida 300 mg.kg-1 E:
pakan
Nukleotida 400 mg.kg-1 F:
pakan
Prosedur Penelitian dan Pengambilan Data
Juvenil udang vaname sebanyak 270 ekor dipelihara selama dua minggu
dalam bak fiberglas berkapasitas 1000 l yang dilengkapi dengan aerator untuk
proses aklimatisasi. Selama periode aklimatisasi, udang diberi pakan standar
sebanyak 3%/bb/hari dan diberikan tiga kali sehari yakni pada pukul 09.00, 13.00,
dan 17.00. Kualitas air dipertahankan stabil dan penggantian air dilakukan setiap
3-4 hari sekali.
Selanjutnya udang (berat rata-rata 6.0±0.5 g) didistribusikan ke dalam 18
unit akuarium percobaan masing-masing berukuran 60x30x30 cm. Setiap
akuarium berisi 50 l air dengan 15 ekor udang, dilengkapi dengan aerasi dengan
airlift system serta menggunakan sistem resirkulasi air. Kualitas air media
pemeliharaan adalah temperatur air 28-29o
Pakan perlakuan diberikan selama 4 minggu berturut-turut dengan tingkat
pemberian 3%/bb/hari dan frekuensi pemberian 3 kali sehari yakni pukul 09.00,
13.00, dan 17.00. Selama masa percobaan, kualitas air dimonitor setiap hari
untuk menjamin agar kondisi