POTENSI Trichoderma sp. SEBAGAI BAHAN
ANTIBAKTERIAL DAN IMUNOSTIMULAN PADA
UDANG VANAME, Litopenaeus vannamei
CATUR AGUS PEBRIANTO
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS
DAN SUMBER INFORMASI
Dengan ini saya menyatakan bahwa tesis dengan judul Potensi Trichoderma sp. Sebagai Bahan Antibakterial dan Imunostimulan pada Udang Vaname,
Litopenaeus vannamei adalah karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini
Bogor, Oktober 2009
ABSTRACT
CATUR AGUS PEBRIANTO. The potential of Trichoderma sp. as antibacterial and immunostimulant in white shrimp, Litopenaeus vannamei. Under guidance of SUKENDA and WIDANARNI
We studied antibacterial and immunostimulatory effects of Trichoderma sp. on white shrimp, Litopenaeus vannamei. First experiment was conducted to evaluate inhibitory effect of Trichoderma sp. againts Vibrio harveyi, a pathogenic bacteria caused vibriosis disease on shrimp. Second experiment was conducted to evaluate immunostimulatory effect of Trichoderma sp. on shrimp immunity as well as protective effect againts V. harveyi. A group of shrimp was injected with a minimum inhibitory concentration obtained at first experiment, and a week after shrimps were challenged with V. harveyi (prophylactic). Another group was challenged with V. harveyi previously, and then injected with Trichoderma sp. two fold of MIC a day after (therapeutic). Positive control, that was received only
V. harveyi, and negative controls, that was received neither Trichoderma sp. nor
V. harveyi were included in this experiment. Results of first experiment showed a concentration of 600 ppm was a minimal inhibitory concentration of Trichoderma
sp. to inhibit V. harveyi. While in the second experiment, groups received
Trichoderma sp., either prophylactic or therapeutic, showed protective against V. harveyi significantly higher than positive control and lower compared with negative control. Total haemocyte count (THC), differential haemocity count (DHC), phagocytic index and phenoloxidase activity were different among the groups of prophylactic treatment or therapeutic treatment compared to positive control and negative control. In conclusion, Trichodema sp. should be used in prophylactic and therapeutic treatment to combat infection of V. harveyi on L. vannamei
RINGKASAN
CATUR AGUS PEBRIANTO. Potensi Trichoderma sp. Sebagai Bahan Antibakterial dan Imunostimulan pada Udang Vaname, Litopenaeus vannamei
Dibimbing oleh SUKENDA dan WIDANARNI.
Perkembangan sistem imun pada udang sangat primitif bila dibandingkan dengan ikan dan vertebrata lainnya, karena udang tidak memproduksi antibodi spesifik. Sistem imun pada udang merupakan sistem imun alami (innate immunity). Udang tidak memproduksi limfosit dan tidak memiliki sistem imun
adaptive seperti yang dimiliki vertebrata lain. Salah satu strategi yang digunakan pembudidaya udang dalam mengendalikan penyakit pada budidaya udang adalah dengan meningkatkan sistem imun udang dengan pemberian imunostimulan.
Cendawan laut merupakan sumber kekayaan alam yang kaya akan produk bioaktif alami. Beberapa cendawan laut telah diisolasi memiliki kemampuan sebagai bahan antimikrobial, antitumor, antivirus, anti kanker dan antiperadangan. Mikroorganisme ini mampu tumbuh pada habitat yang unik dan ekstrim karena mereka mampu menghasilkan produk metabolik sekunder yang unik dan tidak biasa. Berbagai cendawan yang terdapat pada serasah mangrove yang telah mengalami dekomposisi antara lain Aspergillus sp., Penicillum sp., Trichoderma
sp., dan Fusarium sp.
Trichoderma sp. yang terdapat pada serasah mangrove memiliki peranan penting bagi kehidupan biota perairan seperti meningkatkan aktivitas metabolisme biota perairan, sebagai bioactive metabolite, agen biokontrol dan sebagai antibiotik alami di perairan. Trichoderma sp. dapat menghasilkan beberapa enzim seperti -1,3-glukanase, kitinase, protease, serta lipase yang memiliki kemampuan untuk mendegradasi dinding dan membran sel patogen. Selain itu Trichoderma
sp. juga menghasilkan peptida berupa peptaibols dan polyketide. Peptaibols
merupakan peptida linier pendek yang dapat berfungsi sebagai anti bakteri, anti cendawan, dan anti virus. Polyketide merupakan kandidat potensial yang dapat berfungsi sebagai antioksidan, antibiotik, anti kanker.
Pada kegiatan perikanan budidaya Trichoderma sp. belum diketahui manfaatnya tetapi dalam dunia pertanian sudah banyak dimanfaatkan sebagai agen biokontrol seperti menghambat pertumbuhan cendawan patogen pada tanaman dan pengendali penyakit layu bakteri pada tomat. Penelitian ini bertujuan untuk menguji potensi Trichoderma sp. sebagai anti bakteri untuk menghambat Vibrio harveyi dan menguji potensi Trichoderma sp. sebagai imunostimulan pada udang vaname.
Metode yang digunakan dalam penelitian ini adalah eksperimental di Laboratorium. Penelitian ini dilaksanakan selama 6 bulan di Laboratorium Kesehatan Ikan, Departemen Budidaya Perairan, dan Laboratorium Fisiologi Hewan Air, Departemen Manajemen Sumberdaya Perairan, FPIK IPB. Udang uji yang digunakan dalam penelitian ini adalah udang vaname (Litopenaeus vannamei) dengan bobot rata-rata 9,41 ± 0,47 g yang diambil dari lokasi budidaya udang vaname di Bakauheni, Lampung. Cendawan yang diisolasi adalah
Vibrio harveyi MR 5339. Wadah perlakuan yang digunakan berupa akuarium dengan ukuran 60 x 33 x 50 cm yang dilengkapi dengan aerasi
Penelitian dilakukan secara in vitro dan in vivo. Penelitian in vitro
dilakukan untuk mengetahui potensi Trichoderma sp. sebagai bahan anti bakterial, sedangkan in vivo dilakukan untuk mengetahui potensi Trichoderma sp. sebagai bahan imunostimulan pada udang vaname. Penelitian ini terdiri dari 4 (empat) perlakuan dengan masing masing 3 (tiga) kali ulangan, kontrol positif, kontrol negatif, pengobatan, pencegahan. Pemeriksaan parameter imun terdiri dari total hemosit (THC), diferensial hemosit (DHC), aktifitas fagositosis, dan aktifitas Phenoloxidase (PO). Pengamatan parameter sistem imun dilakukan 24, 48, 72, dan 96 jam pasca infeksi. Sedangkan pengamatan kelangsungan hidup dilakukan diakhir penelitian.
Trichoderma sp. yaitu koloni pada media Potatoe Dextrose Agar (PDA) awalnya berwarna putih, tiga hari kemudian sebagian berwarna hijau yang dikelilingi miselium berwarna putih dan selanjutnya seluruh koloni berwarna hijau. Secara mikroskopis cendawan yang diperoleh memiliki konidiofor percabangan berupa segitiga/piramid dan konidia berbentuk bulat sehingga diidentifikasi sebagai Trichoderma sp. Dari 100 ml Potatoe Dextrose Broth
(PDB) yang berisi cendawan Trichodermasp. menghasilkan berat kering 0,522 g. Semakin tinggi dosis cendawan Trichoderma sp., semakin kuat daya hambatnya terhadap V. harveyi. Pengujian dengan dosis 100-200 ppm berbeda nyata dengan dosis 300-500 ppm, serta 600-1.000 ppm. Dari data tersebut diketahui bahwa pada konsentrasi ekstrak 600 ppm memiliki daya hambat yang berbeda nyata dengan konsentrasi 0-500 ppm, tetapi tidak berbeda nyata terhadap konsentrasi 700-1.000 ppm.
Pada akhir penelitian menunjukkan sintasan pada kontrol negatif tidak berbeda nyata (p>0,05) terhadap perlakuan pengobatan maupun perlakuan pencegahan. Walaupun demikian sintasan tertinggi diperoleh dari kontrol negatif sebesar 85,71 ± 0,00% dan selanjutnya diikuti perlakuan pencegahan (71,43 ± 14,29%) dan pengobatan (57,14 ± 14,29%). Semua udang pada kontrol positif mengalami kematian total sampai 24 jam pasca uji tantang dengan V. harveyi. Pemberian Trichoderma sp. sebelum uji tantang (pencegahan) mampu meningkatkan nilai parameter immunologis sehingga sel hemosit dapat mengendalikan V. harveyi dalam tubuh udang melalui aktifitas fagositosis. Pemberian Trichoderma sp. setelah uji tantang (pengobatan) mampu mengendalikan populasi V. harveyi karena sifat anti bakterialnya dan juga efek dari immunostimulasinya. Dalam hal ini pemberian Trichoderma sp. setelah uji tantang dapat memberikan proteksi terhadap udang vaname dari infeksi V. harveyi.
Peranan total hemosit pada krustasea dalam resistensi patogen, apabila terjadi penurunan total hemosit maka dapat terjadi infeksi akut yang dapat menyebabkan kematian. Dengan terjadinya peningkatan total hemosit maka akan meningkatkan kemampuan untuk memfagositosis. Peningkatan total hemosit juga menjadikan daya peningkatan sel granular untuk melakukan aktifitas phenoloxidase sehingga udang dapat bertahan terhadap serangan bakteri.
Peningkatan aktifitas fagositik udang mengindikasikan bahwa Trichoderma
dalam tubuh udang direspon pada sel semi granular dan granular dengan melepaskan sistem proPO yang diaktifkan. Selanjutnya sel semi granular menstimulasi fagositosis oleh sel hialin atau enkapsulasi. Pemberian ekstrak dapat meningkatkan daya tahan terhadap penyakit infeksi, karena meningkatnya mekanisme respon imun pertahanan non spesifik. Mekanisme proses fagositosis melalui tahapan-tahapan sebagai berikut : (1) perlekatan partikel pada permukan sel, (2) penelanan, (3) penghancuran dan pencernaan bakteri.
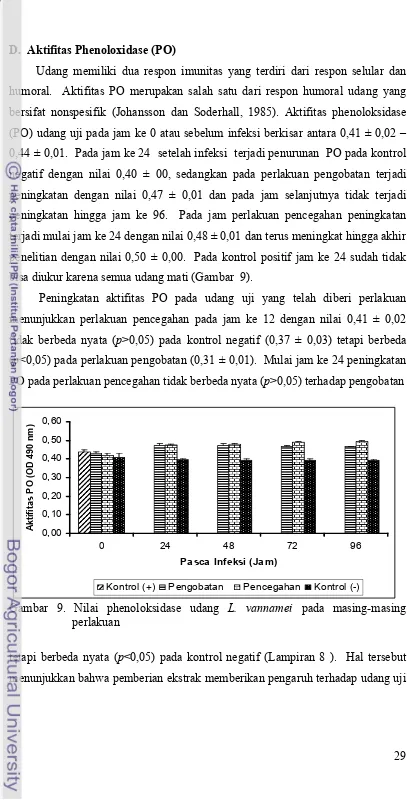
Enzim phenoloxidase dihasilkan melalui sistem proPO yang dapat diaktifkan oleh adanya imunostimulan dan enzim yang berperan dalam proses melanisasi. Pemberian ekstrak Trichoderma sp. mampu meningkatkan aktifitas phenoloksidase menunjukkan bahwa ekstrak dapat menstimulasi hemosit udang vaname hingga aktifitas phenoloksidase terbentuk. Dengan meningkatnya aktifitas phenoloksidase maka kemampuan udang vaname untuk mengenali partikel asing yang masuk kedalam tubuh semakin baik dan untuk selanjutnya proses fagositosis terjadi. Proses ini akan mengurangi partikel asing dalam tubuh sehingga daya tahan udang dapat meningkat.
© Hak cipta milik IPB, tahun 2009
Hak cipta dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan yang wajar IPB
POTENSI Trichoderma sp. SEBAGAI BAHAN
ANTIBAKTERIAL DAN IMUNOSTIMULAN PADA
UDANG VANAME,
Litopenaeus vannamei
CATUR AGUS PEBRIANTO
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
Program Studi Ilmu Perairan
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Judul Tesis : Potensi Trichoderma sp. Sebagai Bahan Antibakterial dan Imunostimulan pada Udang Vaname, Litopenaeus vannamei
Nama : Catur Agus Pebrianto
NIM : C151060121
Disetujui
Komisi Pembimbing
Dr. Ir. Sukenda, M.Sc Dr. Ir. Widanarni, M.Si
Ketua Anggota
Diketahui
Ketua Program Studi Ilmu Perairan
Dekan Sekolah Pascasarjana
Prof. Dr. Ir. Enang Harris, M.S Prof. Dr. Ir. Khairil A. Notodiputro, M.S
PRAKATA
Puji dan syukur kepada Allah SWT atas segala karunia-Nya sehingga
penulis dapat menyelesaikan penelitian dan penulisan tesis yang berjudul Potensi
Trichoderma sp. Sebagai Bahan Antibakterial dan Imunostimulan pada Udang Vaname,Litopenaeus vannamei.
Terima kasih penulis ucapkan kepada Bapak Dr. Ir. Sukenda, M.Sc dan Ibu
Dr. Ir. Widanarni, M.Si selaku komisi pembimbing atas saran dan pengarahannya
dalam penyusunan tesis ini, Bapak Dr. Ir. Tatag Budiardi, M.Si sebagai penguji
yang telah memberikan saran untuk melengkapi tulisan ini. Disamping itu ucapan
terima kasih penulis sampaikan kepada Bapak Dr. Ridwan Affandi atas ijinnya
memanfaatkan Laboratorium Fisiologi Hewan Air dan Bapak Ranta dari
Laboratorium Kesehatan Ikan Departemen Budidaya Perairan IPB, yang telah
membantu selama selama penelitian dilaksanakan. Ungkapan terima kasih juga
disampaikan kepada seluruh keluarga khususnya kepada Bapak Heryanto S (alm),
Ibu Nasriah, kakak, adik serta istriku Siti Zakiyah atas segala doa dan
dukungannya selama ini. Ucapan terima kasih juga penulis sampaikan kepada
Yudiana J, Diana Y. Syahailatua, Pak Hengky M serta rekan-rekan mahasiswa
program studi Ilmu Perairan, Sekolah Pascasarjana IPB angkatan 2006 dan
rekan-rekan di LKI atas kekompakan, kerjasama yang baik dan bantuannya dalam
penyelesaian tesis ini.
Dalam penyusunan tesis ini penulis menyadari masih terdapat kekurangan
yang perlu disempurnakan. Oleh karena itu segala segala saran untuk
perbaikannya akan sangat dihargai demi kesempurnaan hasil penelitian ini di
kemudian hari. Penulis berharap agar hasil penelitian ini dapat bermanfaat.
Bogor, Oktober 2009
RIWAYAT HIDUP
Penulis dilahirkan di Loa Janan pada tanggal 14 Februari 1978 dari ayah
Heryanto.S (Alm) dan ibu Nasriah. Penulis merupakan anak ke empat dari lima
bersaudara.
Tahun 1997 penulis lulus dari SMA Negeri 4 Samarinda dan pada tahun
1998 lulus seleksi masuk pada Program Studi Budidaya Perairan, Jurusan
Perikanan, Fakultas Pertanian, Universitas Mulawarman dan lulus pada tahun
2004 pada Fakultas Perikanan dan Ilmu Kelautan, Universitas Mulawarman. Pada
tahun 2005 diterima sebagai staf pengajar pada Program Studi Budidaya Perairan,
Fakultas Perikanan dan Ilmu Kelautan, Universitas Mulawarman. Penulis
melanjutkan studi di Program Studi Ilmu Perairan, Sekolah Pascasarjana Institut
Pertanian Bogor dengan bantuan dana Beasiswa Pendidikan Pascasarjana (BPPs)
yang diperoleh dari Departemen Pendidikan Nasional Republik Indonesia pada
DAFTAR ISI
Halaman
DAFTAR TABEL ... xv
DAFTAR GAMBAR ... xvi
DAFTAR LAMPIRAN... xvii
PENDAHULUAN ... 1
Latar Belakang ... 1
Tujuan Penelitian ... 3
Hipotesis ... 3
TINJAUAN PUSTAKA Cendawan Trichoderma sp. ... 4
Darah Udang ... 6
Mekanisme Pertahanan Tubuh Pada Udang ... 7
Imunostimulan Udang ... 8
Vibrio harveyi ... 9
METODE PENELITIAN Waktu dan Tempat Penelitian ... 11
Bahan Penelitian... 11
Pelaksanaan Penelitian ... 11
Uji In Vitro ... 11
Uji In Vivo ... 12
Pemeriksaan Parameter Penelitian ... 12
Sistem Imun ... 12
Total Hemosit... 12
Diferensial Hemosit ... 13
Aktifitas Fagositosis ... 13
Aktifitas Phenoloxidase ... 14
Sintasan ... 14
Rancangan Percobaan ... 15
HASIL DAN PEMBAHASAN Trichoderma sp ... 16
Uji In Vitro ... 16
Uji In Vivo ... 19
Sintasan Udang ... 19
Total Hemosit ... 20
Diferensial Hemosit ... 22
Aktifitas Fagositosis... 26
xiv KESIMPULAN DAN SARAN
Kesimpulan ... 31
Saran ... 31
DAFTAR PUSTAKA ... 32
DAFTAR TABEL
Halaman
xvi
DAFTAR GAMBAR
Halaman
1. Trichoderma sp. pada cawan petri dan tabung reaksi ... 16 2. Sintasan udang L. vannamei pada masing-masing perlakuan ... 19 3. Total hemosit udang L. vannameipada masing-masing perlakuan 20 4. Persentase sel hialin udang L. vannameipada masing-masing
perlakuan ... 23
5. Persentase sel semi granulosit udang L. vannameipada
masing-masing perlakuan ... 24
6. Persentase sel granulosit udang L. vannameipada masing-masing
perlakuan ... 25
7. Sel yang melakukan aktifitas fagositosis ... 26
8. Aktifitas fagosistosis udang L. vannamei pada masing-masing
perlakuan ... 27
9. Nilai phenoloksidase udang L. vannamei pada masing-masing
DAFTAR LAMPIRAN
Halaman
1. Uji daya hambat fungi Trichoderma sp. terhadap bakteri Vibrio harveyi 36 2. Sintasan udang vaname (Litopenaeus vannamei) selama masa
penelitian ... 37
3. Total Hemosit udang vaname dengan beberapa perlakuan... 38
4. Sel Hialin (%) udang vaname dengan beberapa perlakuan ... 39
5. Sel Semi Granulosit (%) udang vaname dengan beberapa perlakuan 40
6. Sel Granulosit (%) udang vaname dengan beberapa perlakuan ... 41
7. Aktifitas Fagositik udang vaname dengan beberapa perlakuan ... 42
8. Aktifitas Phenoloksidase udang vaname dengan beberapa
perlakuan ... 43
I. PENDAHULUAN
Latar Belakang
Permasalahan yang sering muncul pada budidaya udang vaname adalah
penyakit yang disebabkan oleh virus, bakteri, parasit dan cendawan. Kehadiran
penyakit ini mempengaruhi dan membatasi budidaya udang (Reed et al., 2003) . Sistem pertahanan tubuh utama pada udang terdiri dari dua bagian yaitu
sistem pertahanan tubuh seluler dan sistem pertahanan humoral. Sistem
pertahanan seluler meliputi fagosit sel-sel hemosit, nodulasi dan encapsulasi.
Sistem pertahanan humoral mencakup phenoloxidase (PO), lectin dan aglutinin.
Kedua sistem pertahanan ini bekerja sama memberikan perlindungan tubuh
terhadap infeksi organisme patogen (Itami, 1994).
Meningkatnya ketahanan tubuh udang dapat diketahui dengan meningkatnya
aktifitas sel fagosit dari hemosit. Fagositosis merupakan mekanisme pertahanan
non spesifik yang secara umum dapat melindungi infeksi patogen. Mekanisme
aktifitas hemosit pada udang terdiri dari mekanisme penjeratan (encapsulasi)
terhadap suatu materi asing, mekanisme fagositosis gabungan terbentuk dari
beberapa hemosit yang membentuk kumpulan lebih besar, dan kumpulan dari
banyak hemosit membentuk suatu lapisan terpigmentasi (Fontaine dan Lightner,
1974). Sedangkan ProPO diaktifkan oleh prophenoloxidase activating enzim
(PPA). ProPO dan PPA ini merupakan protein yang berlokasi di granular hemosit.
Akibat dari pengaktifan proPO menjadi PO dihasilkan protein faktor opsonin
yang merangsang fagositosis hialosit (Johansson dan Soderhall, 1989).
Perkembangan sistem imun pada udang sangat primitif bila dibandingkan
dengan ikan dan vertebrata lainnya, karena udang tidak memproduksi antibodi
spesifik. Sistem imun pada udang merupakan sistem imun alami (innate immunity) (Kwang, 1996). Udang tidak memproduksi limfosit dan tidak memiliki sistem imun adaptive seperti yang dimiliki vertebrata lain (Van de Braak, 2002). Salah satu strategi yang digunakan pembudidaya udang dalam mengendalikan
penyakit pada budidaya udang adalah dengan meningkatkan sistem imun udang
dengan pemberian imunostimulan (Dugger dan Jory, 1999).
Penggunaan immunostimulan sebagai pakan suplemen dapat meningkatkan
(Kumari and Sahoo, 2006). Strategi yang digunakan pembudidaya udang dalam
mengendalikan penyakit pada budidaya udang adalah dengan meningkatkan
sistem imun udang dengan pemberian immunostimulan, terutama -glucan
(Dugger dan Jory, 1999). Immunostimulan mengaktifkan mekanisme non
spesifik, sel perantara imunitas dan respon immun spesifik (Swicki et al., 1998). Ditambahkan oleh Sakai (1999), bahwa immunostimulan dapat meningkatkan
daya tahan tubuh terhadap infeksi, serta dapat meningkatkan mekanisme
pertahanan non spesifik. Pemanfaatan immunostimulan tidak memperlihatkan
efek samping negatif pada udang, tidak seperti pemberian antibiotik (Anderson,
1992; Secombes 1994). Bahan immunostimulan yang telah digunakan antara lain
glukan, chitin, chitosan serta levamisole, selain itu vitamin C dan B serta hormon
pertumbuhan dan prolaktin.
Hutan mangrove menghasilkan bahan pelapukan sebagai serasah menjadi
sumber makanan bagi udang, ikan, kepiting, zooplankton, invertebrate kecil dan
organisme pemakan bahan-bahan hasil pelapukan lainnya. Serasah mangrove
diuraikan oleh bakteri dan cendawan menjadi detritus terlarut yang dapat
dimanfatkan organisme perairan sebagai bahan makanan. Bakteri dan cendawan
merupakan mikroorganisme primer yang berperan dalam proses dekomposisi
berbagai komponen serasah.
Cendawan laut merupakan sumber kekayaan alam yang kaya akan produk
bioaktif alami. Beberapa cendawan laut telah diisolasi memiliki kemampuan
sebagai bahan antimikrobial, antitumor, antivirus, anti kanker dan antiperadangan.
Mikroorganisme ini mampu tumbuh pada habitat yang unik dan ekstrim karena
mereka mampu menghasilkan produk metabolik sekunder yang unik dan tidak
biasa. Yunasfi (2006), melaporkan berbagai cendawan yang terdapat pada serasah
mangrove yang telah mengalami dekomposisi antara lain Aspergillus sp.,
Penicillum sp., Trichoderma sp., dan Fusarium sp.
Trichoderma sp. yang terdapat pada serasah mangrove memiliki peranan penting bagi kehidupan biota perairan seperti meningkatkan aktivitas metabolisme
3 lipase yang memiliki kemampuan untuk mendegradasi dinding dan membran sel
patogen (El ad et al. 1983 dalam Tomia, 2005). Selain itu Trichoderma sp. juga menghasilkan peptida berupa peptaibols dan polyketide. Peptaibols merupakan peptida linier pendek yang dapat berfungsi sebagai anti bakteri, anti cendawan,
dan anti virus (Ruiz et al. 2007). Polyketide merupakan kandidat potensial yang dapat berfungsi sebagai antioksidan, antibiotik, anti kanker (Saleem et al. 2007)
Pada kegiatan perikanan budidaya Trichoderma sp. belum diketahui manfaatnya tetapi dalam dunia pertanian sudah banyak dimanfaatkan sebagai agen
biokontrol seperti menghambat pertumbuhan cendawan patogen pada tanaman
dan pengendali penyakit layu bakteri pada tomat.
Tujuan Penelitian
Penelitian ini bertujuan untuk :
1. Menguji potensi Trichoderma sp. sebagai anti bakteri pada udang vaname. 2. Menguji potensi Trichoderma sp. sebagai bahan immunostimulan udang
vaname.
Hipotesis
Hipotesis yang diajukan pada penelitian ini adalah jika pemberian ekstrak
cendawan Trichoderma sp. pada udang vaname bersifat antibakteri dan dapat berperan dalam meningkatkan imunitas udang maka udang akan resisten terhadap
II. TINJAUAN PUSTAKA
Cendawan Trichoderma sp.
Hutan mangrove adalah suatu komunitas tumbuhan yang secara alami
dipengaruhi oleh pasang surut air laut, yang tergenang pada saat pasang naik dan
bebas dari genangan pada saat pasang rendah. Ekosistem mangrove merupakan
suatu sistem yang terdiri dari lingkungan biotik dan abiotik yang saling
berinteraksi didalam suatu habitat mangrove. Jenis mangrove yang banyak
dijumpai di Indonesia antara lain Bakau (Rhizophora spp.), Api-api (Avicennia
spp.), Pedada (Sonneratia spp.), Tanjang (Bruguiera spp.), Nyirih (Xylocarpus
spp.), Tenger (Ceriops spp.), dan Buta-buta (Exoecaria spp.). Secara ekologi mangrove menghasilkan bahan organik unsur hara di perairan, sehingga memiliki
peranan bagi kelangsungan hidup berbagai biota perairan. Perairan yang terdapat
didekat hutan mangrove merupakan daerah subur sehingga banyak dimanfaatkan
sebagai tempat berlindung dan mencari makan bagi biota perairan seperti ikan dan
udang yang masih muda (Leswara et al. 1987).
Hutan mangrove menghasilkan bahan pelapukan yang menjadi sumber
makanan bagi udang, ikan, kepiting, zooplankton, invertebrate kecil dan
organisme pemakan bahan-bahan hasil pelapukan lainnya. Bahan pelapukan
mangrove berasal dari berbagai organ pohon mangrove seperti daun, ranting,
cabang, bunga dan bagian pohon lainnya yang jatuh dan lazim disebut serasah.
Agar dapat dimanfaatkan oleh organisme perairan lainnya, serasah tersebut perlu
didekomposisi terlebih dahulu menjadi bahan lain yang dapat menjadi sumber
makanan bagi organisme tersebut.
Sebagian serasah mangrove diuraikan oleh bakteri dan cendawan menjadi
detritus terlarut yang dapat dimanfatkan organisme perairan sebagai bahan
makanan. Bakteri dan cendawan merupakan mikroorganisme primer yang
berperan dalam proses dekomposisi berbagai komponen serasah. Yunasfi (2006),
melaporkan berbagai cendawan yang terdapat pada serasah mangrove yang telah
mengalami dekomposisi antara lain Aspergillus sp., Penicillum sp., Trichoderma
5
Trichoderma sp. merupakan mikroba yang terdapat dihampir semua jenis tanah dan habitat yang berbeda. Genus ini termasuk kedalam divisi
Amastigoycota, subdivisi Deuteromycotina, kelas Deuteromicetes, sub kelas
Hypomycetidae, ordo Moniliales, family moniliaceae (Alexopoulos dan Mims,
1979 dalam Rossiana, 1992).
Trichoderma sp. membentuk koloni berwarna putih dengan miselia yang longgar atau kompak. Warna koloni biasanya dipengaruhi oleh pigmentasi
fialosfor, jumlah spora maupun pH media (Rifai,1969). Trichoderma sp. merupakan mikoparasitik aktif yang dapat digunakan sebagai agen biokontrol. T. harzianum mampu menyerang sejumlah cendawan patogen penyebab utama penyakit tanaman (Papavizas 1985; Chet 1987). Mekanisme antagonis utama T. harzianum terhadap cendawan patogen melalui pengeluaran enzim seperti β-1,3 glukanase, kitinase, dan protease yang memiliki kemampuan untuk mendegradasi
dinding dan membran sel patogen (El ad et al. 1983 dalam Tomia, 2005).
Sifat antagonistik yang dimiliki Trichoderma sp. adalah antibiosis, lisis, kompetisi dan mikoparasit. Wells (1986), mengemukakan bahwa Trichoderma
sp. dapat tumbuh diberbagai tempat, mudah diisolasi dan dibiakkan, serta kisaran
parasitisme terhadap patogen tumbuhan sangat luas. Trichoderma sp. mempunyai kemampuan berkompetisi akan makanan dan tempat, umumnya menghasilkan
antibiotik serta memiliki kerja enzim yang memungkinkan kerusakan pada
berbagai cendawan patogen.
Trichoderma sp. yang terdapat pada serasah mangrove memiliki peranan penting bagi kehidupan biota perairan seperti meningkatkan aktivitas metabolisme
biota perairan, sebagai bioactive metabolite, agen biokontrol dan sebagai antibiotik alami di perairan (Sun et al. 2008). Selain itu Trichoderma sp. juga menghasilkan peptida berupa peptaibols dan polyketide. Peptaibols merupakan peptida linier pendek yang dapat berfungsi sebagai anti bakteri, anti cendawan,
Darah Udang
Darah udang disebut hemolim memiliki dua komponen yakni plasma dan sel
darah dengan komponen organik dan anorganik. Komponen organik terdiri dari
gula, lemak dan protein sedangkan komponen anorganik terdiri dari natrium dan
klorida serta sedikit kalium, kalsium dan magnesium. Darah udang tidak
berwarna merah karena tidak memiliki Hemoglobin (Hb), tetapi jika berikatan
dengan oksigen akan berwarna biru muda. Fungsi Hb pada udang digantikan oleh
hemosianin sebagai transport oksigen, yaitu suatu protein yang mengandung Cu
dan bisa berikatan dengan oksigen. Hemosianin berfungsi dalam transport
oksigen, sebagai buffer dalam darah krustasea dan berperan penting dalam
osmotik darah (Maynard, 1960).
Keberadaan hemosit dalam darah crayfish (arthropoda) menentukan tingkat
kekebalan tubuh udang terhadap serangan penyakit patogen. Apabila terjadi
penurunan total hemosit akan menyebabkan kerusakan pada inang yang ditandai
dengan reaksi melanisasi oleh jamur Aphanomyces astaci pada jaringan kutikula, penetrasi pada jaringan tubuh dan disusul kematian organisme (Person et al.,
1987).
Maynard (1960) menyatakan bahwa sel hemolim terdiri dari granulosit dan
hialosit. Sel hialosit mempunyai nukleus yang besar terletak ditengah dikelilingi
oleh sitoplasma basophilik. Pada sitoplasma tidak terlihat adanya retikulum
endoplasmik serta ribosom dan juga tidak ditemukan badan golgi, granul hampir
tidak ada atau terlihat sangat sedikit (Amirante, 1986).
Bentuk hemosit Penaeid dibedakan menjadi bentuk yang tidak bergranula (agranolucyte), granulanya sedikit (semigranulocyte) dan bergranula banyak
(granulocyte) (Martin dan Graves, 1985). Dari analisa flow cytometer oleh
Owens dan O'Neill (1996) bahwa persentase hialosit udang windu (Penaeus monodon Fab.) yang normal terdiri dari 60% - 93% dari total hemosit, sedangkan persentase granulosit berjumlah 17 % - 40 %. Berdasarkan analisa Cell-Dyn
3000, total hemosit rata-rata berjumlah 2,1 x 107. Sel hialosit bersifat fagositik,
yakni kemampuan untuk menerkam partikel-partikel asing seperti bakteri dan
virus. Apabila hialosit masuk ke dalam jaringan, hialosit berkembang menjadi
7 di dalam sitoplasma. Granulosit merupakan jaringan pertama untuk sistem
pertahanan seluler melawan infeksi dengan migrasi ke daerah-daerah yang sedang
mengalami infeksi, menembus dinding pembuluh dan memfagosit partikel asing
untuk dihancurkan.
Johansson dan Soderhall (1989), menyatakan bahwa hialosit juga berperan
dalam sistem pertahanan udang. Sel hialosit ini diaktifkan oleh faktor opsonin
yang dihasilkan dari aktifnya proPO menjadi PO pada sel granular, sehingga
dapat memfagositosis material asing baik bakteri maupun virus, tetapi yang paling
berperan dalam sistem pertahanan udang adalah granulosit.
Mekanisme Pertahanan Tubuh pada Udang
Tidak seperti ikan atau vertebrata lain yang mempunyai antibodi spesifik
atau komplemen, sistem kekebalan udang masih primitif dan tidak memiliki sel
memori. Pada invertebrata seperti udang tidak memiliki imonoglobulin yang
berperan dalam mekanisme kekebalan tubuh (Kwang, 1996).
Sistem pertahanan tubuh utama pada udang terdiri dari dua bagian yaitu
sistem pertahanan tubuh seluler dan sistem pertahanan humoral. Sistem
pertahanan seluler meliputi fagosit sel-sel hemosit, nodulasi dan encapsulasi.
Sistem pertahanan humoral mencakup phenoloxidase (PO), propenoloxidase
(ProPO), lectin dan aglutinin. Kedua sistem pertahanan ini bekerja sama
memberikan perlindungan tubuh terhadap infeksi organisme patogen dari
lingkungan (Itami, 1994). ProPO diaktifkan oleh prophenoloxidase activating enzim (PPA). Sedangkan PPA ini bisa diaktifkan oleh lipopolisakarida. ProPO dan PPA ini merupakan protein yang berlokasi di granular hemosit. Akibat dari
pengaktifan proPO menjadi PO dihasilkan protein faktor opsonin yang
merangsang fagositosis hialosit (Johansson dan Soderhall, 1989).
Meningkatnya ketahanan tubuh udang dapat diketahui dengan meningkatnya
aktifitas sel fagosit dari hemosit. Fagositosis merupakan mekanisme pertahanan
non spesifik yang secara umum dapat melindungi adanya serangan penyakit yang
bersifat patogen. Hemosit sebagai faktor yang sangat penting dalam sistem
pertahanan seluler yang bersifat non spesifik. Kemampuan hemosit dalam
pertahanan tubuh yang bersifat seluler. Dengan adanya infeksi tersebut akan
merangsang sistem pertahanan non spesifik seluler sehingga diharapkan dapat
menangkal serangan penyakit. Mekanisme aktifitas hemosit pada udang terdiri
dari mekanisme penjeratan (encapsulasi) terhadap suatu materi asing, mekanisme
fagositosis gabungan terbentuk dari beberapa hemosit yang membentuk kumpulan
lebih besar, dan kumpulan hemosit membentuk suatu lapisan terpigmentasi
(Fontaine dan Lightner, 1974).
lmunostimulan
Imunisasi udang adalah suatu usaha untuk meningkatkan ketahanan udang
dengan jalan memasukkan antigen ke dalam tubuh, imunisasi yang telah dikenal
secara umum adalah vaksinasi (Anderson, 1992). Sedangkan menurut Ellis
(1988), bahwa vaksin adalah produk yang dihasilkan dari suspensi
mikroorganisme hidup maupun mati yang dapat menghasilkan kekebalan
(immunity). Pada hewan invertebrata misalnya kekebalan udang ini berupa
aglutinin, bakterisidin, fagositosis dan presipitin (Paterson et al. 1992).
Dalam budidaya udang penggunaan antibiotik untuk menghambat bakteri
saat ini sudah tidak efektif lagi, dikarenakan beberapa bakteri sebagai contoh V. harveyi telah memiliki sifat resistensi terhadap beberapa antibiotik antara lain
chloramphenicol, erytrhromimycin, neomycin, oxytetracycline, furazolidone, nifurpirinol dan gentamycin (Karunasagar et al. 1994 ; Muliani, 2002). Selain itu penggunaan antibiotik memiliki dampak terhadap lingkungan akuatik dan
residunya dapat membahayakan kesehatan manusia yang mengkonsumsinya
(Reed et al. 2003).
ltami et al. (1996) mengatakan bahwa pemberian immunostimulan dapat meningkatkan aktivitas fagosit hemosit dan meningkatkan aktivitas ProPO,
sehingga mencegah infeksi dari Vibrio. Taslihan (1991), menambahkan terdapat penyebaran hemosit diasumsikan sebagai bentuk dari mekanisme respon seluler
terhadap masuknya benda asing ke dalam tubuh udang terutama pada
hepatopankreas pasca larva udang windu setelah dilakukan vaksinasi. Sejauh ini
diketahui bahwa pemberian immunostimulan tidak mempunyai efek samping dan
9 dalam sistem kekebalannya, seperti golongan krustase dengan cara merangsang
atau memaksimalkan respon kebal non-spesifiknya (Kwang, 1996).
Vibrio harveyi
Bakteri Vibrio tergolong dalam divisi Bakteria, klas Shyzomycetes, ordo Eubacterial, family Vibrionaceae dan genus Vibrio. Bakteri ini merupakan bakteri gram negatif, berbentuk sel tunggal, berbentuk batang pendek yang bengkok
(koma) atau lurus, bersifat motil, ukuran sel 1-4 mikron, berpendar dan
mempunyai flagella disalah satu kutubnya (Kreig dan Peter, 1984). Sedangkan
sifat biokimianya adalah oksidase positif (kecuali V. metschnikovii dan V. gazogenes), fermentatife terhadap glukosa, sensitife terhadap uji 0/129. DNA genomnya mengandung 51 % mol G+C (guanin dan sitosin) (Logan, 1994), tidak
membentuk gas pada produksi asam dari glukosa dan dapat menggunakan sukrosa
sebagai sumber energi (Lavilla-Pitogo et al. 1990). V. harveyi menghasilkan katalase, indol, lysine, dekarboksilase, nitrat reduktase dan oksidase, tapi tidak
menghasilkan arginin dihidrolase (Alvares et al. 1998)
Menurut Liu, Lee, dan Cheng (1996), dalam Zhang et al. 2001), protease, phospolipase, haemolisin atau exotoxin merupakan patogenitas penting untuk
Vibrio harveyi. Bakteri penghasil cahaya ini sangat patogen dan akut sehingga dapat menyebabkan kematian larva udang sampai 100% dalam waktu singkat.
Larva udang yang terserang cepat mati, tidak berbangkai dan hancur (Rukyani et al. 1992).
Penularan penyakit bakterial dalam lingkungan perairan, dapat terjadi
melalui kontak langsung dengan inang yang sakit, alat-alat yang digunakan,
bagian sisa tubuh ikan, melalui hewan dan tumbuhan air serta air bekas ikan sakit.
Pada larva yang terinfeksi V harveyi terjadi penyusutan hepatopankreas dan perubahan warna menjadi coklat kehitaman (Roza et al. 1997). Larva yang terinfeksi pada tingkat parah terlihat bercahaya pada kondisi gelap dan
penyebabnya diidentifikasi sebagai V. harveyi. V. harveyi dapat menyebabkan bercak merah pada dasar bak pemeliharaan larva (Lavilla Pitogo et al. 1990; Roza
Menurut Zafran dan Roza (1993), V. harveyi akan bersifat patogen bagi larva udang windu apabila kepadatannya dalam air pemeliharaan mencapai 8,35 x
104 cfu/ml. Hal senada dengan Prajitno (1995) yang mengatakan bahwa dengan
III. METODE PENELITIAN
Waktu dan Tempat Penelitian
Penelitian ini dilaksanakan selama 6 bulan dari bulan Maret - Agustus 2008.
Pelaksanan penelitian dilakukan di Laboratorium Kesehatan Ikan, Departemen
Budidaya Perairan, dan Laboratorium Fisiologi Hewan Air, Departemen
Manajemen Sumberdaya Perairan, Fakultas Perikanan dan Ilmu Kelautan IPB.
Bahan Penelitian
Udang uji yang digunakan dalam penelitian ini adalah udang vaname
(Litopenaeus vannamei) dengan bobot rata-rata 9,41 ± 0,47 g diambil dari lokasi budidaya udang vaname di Bakauheni, Lampung. Bahan yang digunakan sebagai
imunostimulan adalah fungi Trichoderma sp. sedangkan bakteri yang digunakan untuk uji tantang adalah Vibrio harveyi MR5339. Wadah perlakuan yang digunakan berupa akuarium dengan ukuran 60x33x50 cm yang dilengkapi dengan
peralatan aerasi
Pelaksanaan Penelitian
Sampel cendawanTrichoderma sp. diperoleh dari lingkungan tambak udang di Bakauheni, Lampung dengan mengisolasi secara bertahap dari serasah
mangrove pada media Potatoe Dextrose Agar (PDA). Setelah diidentifikasi berdasarkan Fassatiova, 1986, kultur murni Trichoderma sp. yang diperoleh diambil sebanyak 1 ose untuk dikultur pada Potatoe Dextrose Broth (PDB) dan diinkubasi pada inkubator shaker selama 5 hari. Selanjutnya dilakukan
penyaringan dengan menggunakan kertas saring Whatman nomor 41 pada labu
erlenmeyer lain sehingga hanya diperoleh filtratnya (Tomia, 2005). Hasil
saringan dikeringkan pada freeze dryer yang sebelumnya sudah dibekukan.
Uji In Vitro
kering. Pengujian dilakukan dengan meletakkan kertas cakram berdiameter 6 mm
yang telah dicelupkan pada larutan Trichoderma sp. dengan konsentrasi 0, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1.000 ppm diatas agar cawan yang telah
disebari V. harveyi. Biakan bakteri V. harveyi kemudian diinkubasi pada suhu kamar selama 24 jam. Respon antibakterial ditentukan dengan mengukur zona
bebas bakteri disekeliling kertas cakram yang kelihatan bening dan diukur
diameter daerah hambatnya (Lay, 1994).
Uji In Vivo
Penelitian ini terdiri dari 4 perlakuan dengan masing masing 3 kali ulangan,
sebagai berikut :
1. Kontrol positif : udang uji diinjeksi V. harveyi106 CFU/ekor.
2. Kontrol negatif : udang uji diinjeksi larutan fisiologis (NaCl 0,85%).
3. Perlakuan pengobatan : udang uji diinjeksi dengan V. harveyi 106 CFU/ekor, kemudian diinjeksi Trichoderma sp. sebanyak 0,1 ml/ekor dengan dosis 1200 ppm (11,3 mg/ekor) sehari kemudian.
4. Perlakuan pencegahan : udang uji diinjeksi dengan Trichoderma sp. sebanyak 600 ppm (5,6 mg/ekor) kemudian diinjeksi dengan V. harveyi 106 CFU/ekor sehari kemudian.
Pengamatan parameter sistem imun dilakukan 24, 48, 72, 96, jam pasca
infeksi.
Pemeriksaan Parameter Penelitian
1. Sistem Imun
Parameter imun yang diukur adalah total hemosit, diferensial hemosit, aktifitas
fagositosis, dan aktifitas phenoloksidase.
a. Total Hemosit (Blaxhall dan Daishley, 1973)
Hemolim diambil sebanyak 0,1 ml dibagian pangkal kaki jalan ke 5 dengan
syringe 1 ml yang sudah berisi antikoagulan sebanyak 0,3 ml, kemudian
dihomogenkan selama 5 menit. Tetesan pertama hemolim pada syringe dibuang,
13 per ml dibawah mikroskop cahaya dengan pembesaran 40 kali. Total hemosit
dihitung dengan menggunakan rumus :
Total Hemosit (sel/ml) = Rata-rata sel terhitung x 25 x FP x 10.000
Keterangan :
FP = faktor pengenceran
b. Diferensial Hemosit (Martin dan Graves , 1995)
Hemolim yang telah diambil dari udang uji diteteskan pada gelas objek dan
dibuat ulasan, kemudian dikeringkan di udara dan difiksasi dengan metanol 100%
selama 5 menit. Setelah itu dikeringkan udara kembali dan diwarnai dengan
larutan giemsa 10% selama 10 menit, dicuci dalam air mengalir selama 30 detik
dan dibiarkan kering. Preparat diamati menggunakan mikroskop cahaya dengan
pembesaran 100 kali dan dibedakan menurut jenisnya yaitu sel hialin, semi
granular dan granular. Persentase jenis hemosit dihitung dengan menggunakan
rumus :
c. Aktifitas Fagositosis (Anderson dan Siwicki , 1993)
Hemolim 0,1 ml dimasukkan kedalam mikroplate dan dicampur secara
merata dengan 25 μl bakteri Staphylococcus aureus dan diinkubasi selama 20 menit. Hemolim sebanyak 5 μl diteteskan pada objek glas dan dibuat preparat
ulas lalu dikeringkan. Fiksasi dengan metanol 100% selama 5 menit dan diwarnai
dengan giemsa selama 15 menit. Aktivitas fagositik diukur berdasarkan
persentase sel-sel fagosit yang melakukan fagositosis. Aktifitas fagositosis
d. Aktifitas Phenoloxidase (PO) (Liu and Chen, 2004)
Aktifitas phenoloxidase diukur menggunakan spektrofotometer.
Pengamatan dilakukan dengan melihat perekaman pembentukan dopachrome
yang dihasilkan dari L-dihydroxyphenylalanine (L-DOPA). Hemolim disentrifuse
pada 1000 rpm pada suhu 4oC selama 20 menit. Cairan supernatan dibuang dan
pellet dibilas dengan cacodylate-citrate buffer hingga 1 ml (sodium cacodylate 0,01 M, sodium chloride 0,45 M, trisodium citrate 0,10 M, pH 7,0) kemudian
disentrifuse ulang. Pellet selanjutnya dicampur dengan cacodylate buffer hingga 200 l (sodium cacodylate 0,01 M, sodium chloride 0,45 M, calcium chloride
0,01 M dan magnesium chloride 0,26 M, pH 7,0). Larutan kemudian dibagi dua.:
Larutan pertama sebanyak 100 l diinkubasi selama 10 menit pada suhu 25oC
dengan 50 l trypsin (1 mg /ml), sebagai elicitor kemudian ditambahkan 50 l L -DOPA, dan 5 menit kemudian dicampur 800 l cacodylate buffer. Larutan kedua sebanyak 100 l suspensi sel dicampur dengan 50 l cacodylate buffer (untuk menggantikan trypsin) dan 50 l L-DOPA yang digunakan sebagai kontrol untuk background aktifitas phenoloxidase pada semua kondisi uji. Optikal density (OD) diukur menggunakan spektrofotometer pada panjang gelombang 490 nm.
Aktifitas phenoloxidase udang diekspresikan sebagai pembentukan dopachrome
per 50 l hemolim.
2. Sintasan (Effendie, 1997)
Sintasan udang uji dihitung mulai infeksi dilakukan dan dihitung dengan
menggunakan rumus :
100
(%) x
No Nt
SR
Keterangan :
SR : Sintasan
Nt : Jumlah udang uji yang hidup pada akhir pengamatan (ekor)
15
Rancangan Percobaan
Rancangan percobaan yang digunakan dalam ujicoba ini adalah rancangan
acak lengkap (RAL) dengan dua perlakuan (pencegahan dan pengobatan) dan dua
kontrol dengan masing-masing tiga ulangan. Kontrol yang digunakan dalam
penelitian ini adalah kontrol positif (tanpa pemberian ekstrak dengan uji tantang),
dan kontrol negatif ( tanpa pemberian ekstrak dan tanpa uji tantang).
Parameter yang diamati dianalisa keragamannya dengan ANOVA dan untuk
mengetahui pengaruh perlakuan dilakukan uji lanjut Fisher’s LSD test
IV. HASIL DAN PEMBAHASAN
Trichoderma sp.
Trichoderma sp. memiliki ciri koloni berwarna putih, beberapa hari kemudian bagian tengah berwarna hijau yang dikelilingi oleh miselium berwarna
putih bersih untuk selanjutnya koloni tersebut berubah menjadi warna hijau.
Secara mikroskopis Trichoderma sp. memiliki konidiofor dengan percabangan menyerupai piramid yaitu cabang yang lebih panjang dibawahnya, fialid tersusun
pada kelompok-kelompok yang berbeda, terdapat 2-3 fialid perkelompok.
Konidia berbentuk bulat dengan diameter 3,5-4,5 µm (Fassaitova, 1986). Dari
isolasi fungi yang berasal dari serasah mangrove diperoleh ciri-ciri Trichoderma
sp. pada media PDA pada awalnya koloni berwarna putih, tiga hari kemudian
sebagian berwarna hijau yang dikelilingi miselium berwarna putih. Untuk
selanjutnya seluruh koloni berwarna hijau (Gambar 1). Secara mikroskopis
Trichoderma sp. memiliki konidiofor percabangan berupa segitiga/piramid dan konidia berbentuk bulat. Dari 100 ml PDB yang berisi fungi Trichoderma sp. menghasilkan berat kering 0,522 g.
Gambar 1. Trichoderma sp. pada cawan petri dan tabung reaksi
Uji In Vitro
Berdasarkan hasil uji daya hambat difusi agar untuk mengetahui sensitifitas
bakteri V. harveyi terhadap bahan aktif dari fungi Trichoderma sp. terlihat adanya potensi untuk menghambat pertumbuhan bakteri V. harveyi. Hal tersebut dapat dilihat dari zona bening yang terbentuk di sekitar kertas cakram yang mengandung
17 melalui pengamatan daerah jernih disekeliling kertas cakram, membuktikan
adanya daya kerja antimikrobial (Lay, 1994).
Daya hambat cendawan Trichoderma sp. terhadap pertumbuhan bakteri
V.harveyi setelah 24 jam masa inkubasi ditunjukkan pada Tabel 1. Semakin tinggi dosis cendawan Trichoderma sp., semakin kuat daya hambatnya terhadap
V. harveyi. Pengujian dengan dosis 100-200 ppm berbeda nyata (p<0,05) dengan dosis 300-500 ppm, serta 600-1.000 ppm. Dari data tersebut diketahui bahwa
pada konsentrasi ekstrak 600 ppm memiliki daya hambat yang berbeda nyata
dengan konsentrasi 0-500 ppm, tetapi tidak berbeda nyata terhadap konsentrasi
700-1.000 ppm (Lampiran 1).
Tabel 1. Rata-rata diameter zona hambat bebas bakteri V. harveyi yang diberi perlakuan ekstrak fungi Trichoderma sp.
Konsentrasi Ekstrak Fungi perbedaan nyata pada taraf 95% (p < 0,05)
Daya hambat dari fungi Trichoderma sp. mulai kelihatan pada konsentrasi 100 ppm, tetapi diameter zona hambat bebas bakteri yang ditunjukkan hingga
konsentrasi 1000 ppm jauh lebih rendah bila dibandingkan dengan nilai standar
resisten bahkan intermediate (Tabel 2). Nilai zona hambat bebas bakteri yang
berada dibawah nilai resisten tersebut mengindikasikan bahwa Trichoderma sp. memiliki antibakterial yang lemah terutama terhadap bakteri V. harveyi. Rendahnya daya hambat Trichoderma sp. terhadap bakteri V. harveyi dapat disebabkan perbedaan kemurnian ekstrak serta jenis antibiotik. Selain itu dapat
diduga pula bahwa V. harveyi telah resisten terhadap bahan dengan konsentrasi tersebut (Tabel 1).
Pemanfaatan Trichoderma sp. juga telah dilaporkan oleh Nurjanani (2001), dimana Trichoderma sp. cukup efektif untuk mengendalikan penyakit layu bakteri (Ralstonia solanacearum) pada tomat. Pengendalian penyakit layu bakteri tersebut ditunjukkan dengan bobot buah tomat yang baik pada akhir musim tanam
bila dibandingkan dengan bahan lainnya.
Tabel 2. Diameter zona hambat beberapa antibiotik
Antibiotik
Dalam budidaya udang penggunaan antibiotik untuk menghambat bakteri
saat ini sudah tidak efektif lagi, dikarenakan beberapa bakteri sebagai contoh V. harveyi telah memiliki sifat resistensi terhadap beberapa antibiotik antara lain
chloramphenicol, erytrhromimycin, neomycin, oxytetracycline, furazolidone, nifurpirinol dan gentamycin (Karunasagar et al. 1994 ; Muliani, 2002). Selain itu penggunaan antibiotik memiliki dampak terhadap lingkungan akuatik dan
residunya dapat membahayakan kesehatan manusia yang mengkonsumsinya
19
Kontrol (+) Pengobatan Pencegahan Kontrol (-)
Uji In Vivo
Sintasan Udang
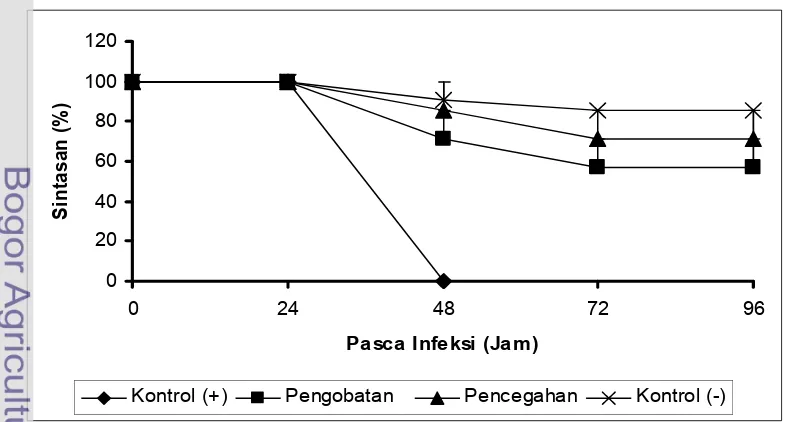
Sintasan udang uji untuk semua perlakuan hingga akhir penelitian disajikan
pada Gambar 2. Pada akhir penelitian menunjukkan sintasan pada kontrol negatif
tidak berbeda nyata (p>0,05) terhadap perlakuan pengobatan maupun perlakuan pencegahan. Walaupun demikian sintasan tertinggi diperoleh dari kontrol
negatif sebesar 85,71 ± 0,00% dan selanjutnya diikuti perlakuan pencegahan
(71,43 ± 14,29%) dan pengobatan (57,14 ± 14,29%). Semua udang pada kontrol
positif mengalami kematian total sampai 24 jam pasca uji tantang dengan V. harveyi ( Lampiran 2).
Pemberian cendawan Trichoderma sp. setelah uji tantang mampu mengendalikan populasi V. harveyi karena sifat anti bakterialnya dan juga efek dari immunostimulasinya. Dalam hal ini pemberian Trichoderma sp. setelah uji tantang (pengobatan) ternyata dapat memberikan proteksi terhadap udang vaname
dari infeksi V. harveyi. Walaupun pada uji in vitro menunjukkan bahwa
Trichoderma sp. memiliki daya bakterial rendah tetapi pada uji in vivo
menunjukkan pengaruh yang besar. Pemberian Trichoderma sp. sebelum uji tantang (pencegahan) mampu meningkatkan nilai parameter immunologis
sehingga sel hemosit dapat mengendalikan V. harveyi dalam tubuh udang melalui aktifitas fagositosis.
0,00
Kontrol (+) Pengobatan Pencegahan Kontrol (-)
A. Total Hemosit
Darah udang dikenal dengan hemosit. Darah udang tidak berwarna merah
karena tidak memiliki Hemoglobin (Hb), tetapi jika berikatan dengan oksigen
akan berwarna biru muda. Fungsi Hb pada udang digantikan oleh hemosianin
yang berfungsi dalam transport oksigen, sebagai buffer dalam darah krustasea dan
berperan penting dalam osmotik darah (Maynard, 1960). Total hemosit udang uji
untuk semua perlakuan hingga akhir penelitian disajikan pada Gambar 3.
Total hemosit udang vaname yang diberi Trichoderma sp. pada akhir penelitian secara statistik menunjukkan perbedaan nyata (p<0,05) terhadap perlakuan. Sedangkan antar perlakuan menunjukkan perbedaan nyata (p<0,05) antara kontrol negatif dengan perlakuan pengobatan dan pencegahan. Untuk
perlakuan pencegahan juga memberikan perbedaan nyata (p<0,05) terhadap perlakuan pengobatan. Hal tersebut mengindikasikan perlakuan ekstrak memberi
pengaruh yang lebih baik dibandingkan kontrol negatif dan juga berpengaruh
terhadap pemberian perlakuan sebelum (pengobatan) maupun sesudah infeksi
(pencegahan) terjadi. Total hemosit terendah terjadi pada jam ke 0 sebelum
perlakuan infeksi yaitu untuk kontrol negatif rata-rata 2,20 ± 0,05 x 106 sel/ml dan
meningkat pada jam ke 24. Total hemosit mencapai puncak pada jam ke 24 yaitu
2,27 ± 0,08 x 106 sel/ml kemudian turun kembali hingga jam ke 96 yaitu 2,23 ±
0,03 x 104 sel/ml. Untuk perlakuan pencegahan total hemosit terendah terjadi
pada jam ke 0 sebelum infeksi V.harveyi yaitu 2,23 ± 0,09 x 106 sel/ml dan selanjutnya meningkat hingga jam ke 48 yaitu 2,67 ± 0,03 x 106 sel/ml (Lampiran
3).
21 Hal ini menunjukkan bahwa perlakuan pemberian ekstrak Trichoderma sp. akan meningkatkan total hemosit udang uji hingga jam ke 48 dan untuk
pengamatan jam berikutnya akan turun kembali. Peningkatan total hemosit pada
jam ke 24 setelah infeksi menunjukkan adanya kemampuan bahan aktif dari
ekstrak untuk meningkatkan imunnitas dengan cepat. Hal tersebut berbeda
dengan pengaruh yang ditunjukkan oleh lipopolisakarida, dimana total hemosit
udang menurun pada saat 4 dan 24 jam setelah perlakuan dan baru meningkat
kembali setelah 48 jam (Van de Braak, 2002).
Peningkatan total hemosit udang uji yang diinjeksi ekstrak Trichoderma sp. mengindikasikan bahwa bahan aktif tersebut mampu merangsang pembentukan
sel-sel hemosit yang kemudian dilepaskan ke dalam hemolimph. PBS steril yang
diinjeksikan pada udang uji ternyata dapat juga meningkatkan total hemosit. Hal
ini didukung pendapat Lorenzo et al. (1999) bahwa total hemosit pada krustasea menurun dengan cepat setelah diinjeksi dengan bahan asing, sementara total
hemosit seringkali meningkat setelah diinjeksi PBS.
Penurunan total hemosit setelah uji tantang berhubungan dengan aktifitas
pertahanan yang berbeda. Hemosit akan bermigrasi ke tempat injeksi
menyebabkan berkurangnya konsentrasi sel dalam hemolimph (Van de Braak
2002). Hemosit beragregat menjadi gumpalan setelah infeksi bakteri yang akut
(Johnson 1976, diacu dalam Van de Braak 2002). Sel-sel fagositik hemosit mungkin meninggalkan sirkulasi setelah memfagositosis dan masuk ke dalam
jantung, jaringan penunjang, insang atau sinus hemallain (Factor dan Beekman
1990, diacu dalam Van de Braak 2002). Degranulasi dapat diikuti oleh lisis sel (Soderhall et al. 1986), oleh karena itu sejumlah hemosit dapat juga hilang selama proses degranulasi.
Total hemosit merupakan salah satu parameter status kesehatan udang.
Beberapa informasi menunjukkan pentingnya total hemosit dalam ketahanan
terhadap patogen. Person et al. (1987) melaporkan hubungan antara jumlah hemosit dan ketahanan Pacifastacus leniusculus terhadap parasit jamur
hemosit bebas dapat bervariasi dan dapat menurun secara drastis selama infeksi.
Le Moullac et al. (1998) mengemukakan bahwa Penaeus stylorostris dengan total hemosit yang rendah akibat hypoxia, menjadi lebih sensitif terhadap infeksi
dengan V. alginolyticus yang sangat virulen. Hypoxia yang menyebabkan menurunnya total hemosit yang signifikan disebabkan oleh penurunan sel-sel semi
granular dan hialin yang signifikan.
Peningkatan total hemosit udang uji setelah perlakuan ekstrak Trichoderma
sp. berarti meningkatkan status kesehatan organisme tersebut karena dengan
peningkatan total hemosit berarti meningkatkan peluang terbentuknya sel-sel
fagositik yang sangat berperan dalam mengendalikan serangan mikroorganisme
dalam tubuh. Dalam hal ini ketahanan tubuh akan semakin meningkat dan
kerentanan tubuh terhadap penyakit infeksi akan menurun. Total hemosit diyakini
mempengaruhi kemampuan inang untuk bereaksi melawan bahan asing (Person et al. 1987) dan berbagai respon terhadap infeksi, perubahan lingkungan pada sebagian besar crustacea (Tsing et al. 1989). Jumlah hemosit yang rendah sangat berkorelasi dengan sensitifitas terhadap patogen yang lebih tinggi (Person et al.
1987; Le Moullac dan Haffner. 2000) dan oleh karena itu, total hemosit yang
rendah mengindikasikan kerentanan terhadap penyakit infeksi tinggi.
Peranan total hemosit pada krustasea dalam resistensi patogen, apabila
terjadi penurunan total hemosit maka dapat terjadi infeksi akut yang dapat
menyebabkan kematian (Rondriguez dan Le Moullac, 2000). Dengan terjadinya
peningkatan total hemosit maka akan meningkatkan kemampuan untuk
memfagositosis. Peningkatan total hemosit juga menjadikan daya peningkatan sel
granular untuk melakukan aktifitas phenoloxidase sehingga udang dapat bertahan
terhadap serangan bakteri
B. Diferensial Hemosit
Berdasarkan bentuk, warna, ukuran relatif dan rasio sitoplasma serta inti
selnya, hemosit dapat dibedakan atas tiga jenis sel yaitu hialin, granular dan semi
granular. Hialin merupakan sel dengan perbandingan inti sel yang lebih tinggi
dibandingkan sitoplasma, granulosit memiliki perbandingan inti sel yang lebih
23
Kontrol (+) Pengobatan Pencegahan Kontrol (-)
dengan perbandingan inti sel yang lebih rendah dibandingkan sitoplasmanya dan
beberapa granul submikron dan mikron. Diferensial hemosit udang uji untuk
semua perlakuan hingga akhir penelitian disajikan pada Gambar 4, 5 dan 6.
Sebelum infeksi V. harveyi persentase hialin berkisar antara 60,39 ± 0,58% -61,47 ± 0,58%, semigranulosit 8,52 ± 1,53% – 10,68 ± 1,53%, dan sel granulosit
28,93 ± 2,65% – 30,01 ±4,36%. 24 jam setelah infeksi pada perlakuan kontrol
positif semua udang vaname mati, sedangkan perlakuan lainnya terjadi penuruan
persentase pada hialin yaitu antara 24,23 ± 1,75% – 53,70 ± 1,17%, tetapi terjadi
peningkatan pada semigranulosit dan granulosit secara berturut turut 7,53 ± 2,06%
- 12,39 ± 1,15% dan 35,35 ± 2,08% – 64,79 ± 1,63%.
Persentase hialin pada perlakuan pencegahan pada jam ke 24 setelah infeksi
menunjukkan penurunan dengan nilai 41,98 ± 0,30% dibandingkan pada jam ke 0
sesaat sebelum infeksi dan pada jam ke 48 persentase hialin mulai meningkat
hingga jam ke 96. Semua waktu pengamatan menunjukkan beda nyata (p<0,05) terhadap perlakuan pengobatan maupun kontrol negatif, kecuali pada jam ke 0.
Persentase perlakuan pengobatan setelah jam ke 0 antara 24,23 ± 1,75% hingga
37,93 ± 5,47%. Nilai terendah terjadi pada jam ke 48 dengan nilai 24,23 ± 1,75%
kemudian meningkat kembali pada jam ke 72 dan 96 jam dengan nilai
berturut-turut 25,95 ± 4,39% dan 26,48 ± 2,75%. Sedangkan pada kontrol negatif nilai
hialin pasca infeksi antara 26,47 ± 2,91% hingga 26,48 ± 2,75% (Lampiran 4).
0
Kontrol (+) Pengobatan Pencegahan Kontrol (-)
Persentase semigranulosit pasca infeksi V. harveyi menunjukkan bahwa kontrol negatif tidak berbeda nyata (p>0,05) terhadap semua perlakuan pencegahan dan pengobatan pada semua waktu pengamatan. Persentase
semigranular terendah pasca infeksi terjadi pada perlakuan pencegahan yaitu pada
jam ke 24 yaitu 7,53 ± 2,06% diikuti kontrol negatif (10,03 ± 2,54%) dan
perlakuan pengobatan dengan nilai 10,37 ± 2,36% .
Penurunan persentase sel hialin berdampak pada peningkatan sel semi
granulosit dan sel granulosit. Pada perlakuan pengobatan persentase tertinggi
pada jam ke 48 dengan nilai sel semi granulosit 12,39 ± 1,15% berbeda nyata
(p<0,05) dengan perlakuan pencegahan tetapi tidak berbeda nyata (p> 0,05) terhadap kontrol negatif. Untuk perlakuan pencegahan nilai tertinggi terjadi pada
jam ke 72 denga nilai 11,10 ± 1,95% dan tidak berbeda nyata (p>0,05) terhadap perlakuan pengobatan dan kontrol negatif. Perlakuan kontrol negatif tertinggi
pada jam ke 48 dengan nilai 12,32 ± 1,56% (Lampiran 5).
Sel granulosit tertinggi pada perlakuan pengobatan jam ke 24 dengan nilai
64,79 ± 1,63% di ikuti dengan kontrol negatif (61,56 ± 2,00%) dan perlakuan
pencegahan dengan nilai 50,58 ± 1,76%.
Gambar 5. Persentase sel semi granulosit udang L. vannamei pada masing-masing perlakuan
Sel granulosit setelah jam ke 24 pasca infeksi terus mengalami penurunan hingga
akhir penelitian begitu pula dengan kontrol negatif dan perlakuan pencegahan
25
Kontrol (+) Pengobatan Pencegahan Kontrol (-)
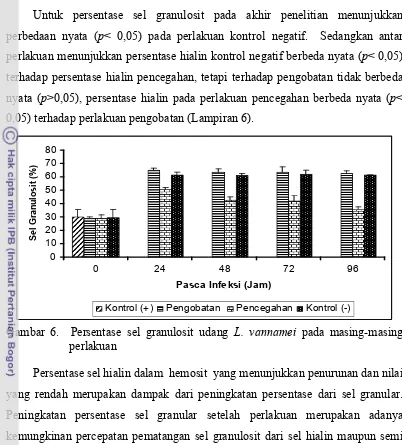
Untuk persentase sel granulosit pada akhir penelitian menunjukkan
perbedaan nyata (p< 0,05) pada perlakuan kontrol negatif. Sedangkan antar perlakuan menunjukkan persentase hialin kontrol negatif berbeda nyata (p< 0,05) terhadap persentase hialin pencegahan, tetapi terhadap pengobatan tidak berbeda
nyata (p>0,05), persentase hialin pada perlakuan pencegahan berbeda nyata (p< 0,05) terhadap perlakuan pengobatan (Lampiran 6).
Gambar 6. Persentase sel granulosit udang L. vannamei pada masing-masing perlakuan
Persentase sel hialin dalam hemosit yang menunjukkan penurunan dan nilai
yang rendah merupakan dampak dari peningkatan persentase dari sel granular.
Peningkatan persentase sel granular setelah perlakuan merupakan adanya
kemungkinan percepatan pematangan sel granulosit dari sel hialin maupun semi
granular. Soderhall dan Cerenius (1992), menjelaskan sebagian besar hemosit
crustacea tidak membelah sehingga kemungkinan besar terjadi proses pergantian
sel tua ke sel yang dilepas kedalam hemolim. Granulosit merupakan tahap
terakhir dari suatu perkembangan sel dalam hemolim. Sel hialin merupakan sel
termuda dibandingkan dengan semi granulosit maupun granulosit dan merupakan
cell line yang dilepas dari hepatopoeietik (Van de Braak,2002). Peningkatan sel-sel granular pada udang setelah perlakuan memberi pengaruh positif terhadap
ketahanan tubuh udang yang dapat mengendalikan populasi V. harveyi dalam tubuh sehingga jenis sel tersebut merupakan sel hemosit yang besar peranannya
dalam meningkatkan ketahanan tubuh udang. Dengan demikian, peningkatan
kesehatan udang yang tentunya tidak lepas dari peran dan fungsi dari jenis sel lain
dalam hemosit. Menurut Soderhall dan Cerenius (1992), kerjasama dan
komunikasi sel pada beberapa reaksi pertahanan terjadi ketika mikroorganisme
atau parasit dikenali dan respon immun dibentuk. Bachere (2000),
mengemukakan berdasarkan karakteriktisasi morfologi dan sitokimia, beberapa
fungsi dan keterlibatan dalam reaksi pertahanan yang berbeda berhubungan
dengan jenis sel yang berbeda, misalnya sel hialin dengan fagositosis, sel granular
dan semi granular dengan encapsulasi dan sistem ProPO (Soderhall dan Cerenius,
1992).
C. Aktifitas Fagositosis
Aktifitas fagositosis merupakan reaksi pertahanan seluler paling umum pada
udang. Respon ini sebagai upaya untuk mempertahankan diri dari masuknya
antigen, dengan cara menghancurkan antigen secara nonspesifik melalui proses
fagositosis (Le Moullac dan Haffner, 2000). Aktifitas fagositik udang uji untuk
semua perlakuan hingga akhir penelitian disajikan pada Gambar 8.
10 µm 10 µm
10 µm
27
Kontrol (+) Pengobatan Pencegahan Kontrol (-)
Aktifitas fagosistosis udang uji pada jam ke 0 atau sebelum infeksi berkisar
antara 40,02 ± 2,54% – 42,56 ± 2,03%. Pada kontrol negatif aktifitas fagosistosis
mencapai puncaknya pada jam ke 24 dengan nilai 42,35 ± 3,11%, kemudian turun
kembali hingga jam ke 72 dengan nilai 41,75 ± 2,42%. Pada jam ke 96 terjadi
peningkatan aktifitas fagosistosis dengan nilai 42,06 ± 3,79%.
Untuk perlakuan pengobatan puncak aktifitas fagosistosis terjadi pada jam
ke 48 dengan nilai 47,44 ± 5,25%, kemudian terjadi penurunan hingga jam ke 96
dengan nilai aktifitas fagosistosis 44,34 ± 4,62%. Perlakuan pencegahan aktifitas
fagosistosis dari jam ke 0 sebelum infeksi dan jam ke 24 pasca infeksi terus
meningkat hingga jam ke 96 dengan nilai 56,55 ± 0,52% (Gambar 8 ).
Aktifitas fagositosis pada jam ke 24 menunjukkan peningkatan tetapi tidak
berbeda nyata (p>0,05) pada semua perlakuan. Pada jam ke 48 perlakuan pencegahan terjadi peningkatan aktifitas fagositosis (54,15 ± 1,83% ) dan berbeda
nyata (p<0,05) terhadap perlakuan pengobatan maupun kontrol negatif.
Gambar 8. Aktifitas fagosistosis udang L. vannamei pada masing-masing perlakuan
Aktifitas fagositosis pada perlakuan pencegahan terus meningkat hingga jam
ke 96 dengan nilai 56,55 ± 0,52%. Perlakuan pengobatan menunjukkan puncak
aktifitas fagositosis pada jam ke 48 (47,44 ± 5,25%) dan menurun kembali pada
jam ke 72. Pada kontrol negatif nilai tertinggi pada jam ke 24 pasca infeksi dan
menurun hingga jam ke 72, untuk jam 96 terjadi peningkatan dengan nilai 42,06 ±
Penurunan aktifitas fagosistosis udang uji setelah uji tantang disebabkan
oleh berkurangnya sel-sel fagositik dalam hemolimph. Sel-sel fagositik dapat
berkurang dalam hemolimph karena dalam aktifitas fagositosis, sel-sel fagositik
sebagian akan hancur bersama dengan bakteri setelah melewati berbagai proses
fagositosis. Menurut Secombes (1994), mekanisme proses fagositosis melalui
tahapan-tahapan sebagai berikut : (1) perlekatan partikel pada permukan sel, (2)
penelanan, (3) penghancuran dan pencemaan bakteri. Degranulasi dapat diikuti
oleh lisis sel (Soderhall et al. 1986), dan oleh karena itu sejumlah hemosit dapat juga hilang selama proses degranulasi, meninggalkan kandungannya dengan
bahan yang terdegradasi dalam matrix fibrous ekstraseluler (Van de Braak, 2002).
Sel-sel fagositik yang tidak hancur bersama patogen dalam proses fagositosis akan
meninggalkan hemolimph. Menurut Faktor dan Beekman (1990), diacu dalam
Van de Braak (2002), sel-sel fagositik hemosit dapat meninggalkan sirkulasi
setelah melakukan fagositosis dan masuk ke jantung, jaringan penunjang, insang,
dan sinus haemal lainnya. Immuno-staining menunjukkan bahwa banyak
haemosit yang terdegranulasi dalam organ limfoid, menghasilkan lapisan bahan
fibrous pada bagaian luar dinding tubule. Ini dapat berkontribusi terhadap
konsentrasi hemosit yang berkurang dalam hemolimph hewan yang sakit atau
setelah injeksi dengan bahan asing.
Peningkatan aktifitas fagositik udang mengindikasikan bahwa Trichoderma
sp. mampu meningkatkan aktifitas fagositik sel-sel fagosit. Sel yang berperan
besar dalam proses fagositik ini adalah sel hialin. Bakteri patogen yang masuk ke
dalam tubuh udang direspon pada sel semi granular dan granular dengan
melepaskan sistem proPO yang diaktifkan. Selanjutnya sel semi granular
menstimulasi fagositosis oleh sel hialin atau enkapsulasi (Söderhäll dan Cerenius,
1992). Pemberian ekstrak dapat meningkatkan daya tahan terhadap penyakit
infeksi, karena meningkatnya mekanisme respon imun pertahanan non spesifik