pC13-35S-Intron-Sma: VEKTOR BINER FLEKSIBEL UNTUK
EKSPRESI GEN TARGET SECARA KONSTITUTIF PADA
TRANSFORMASI TANAMAN MELALUI
AGROBACTERIUM
NATALIA LUSIANNGSIH SUMANTO
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
RINGKASAN
NATALIA LUSIANINGSIH SUMANTO. pC13-35S-Intron-Sma: Vektor Biner Fleksibel untuk Ekspresi Gen Target Secara Konstitutif pada Transformasi Tanaman Melalui Agrobacterium. Dibimbing oleh Prof. Dr. Ir. Antonius Suwanto, M.Sc dan Dr. Sigit Purwantomo.
Teknik penyisipan gen target ke dalam tanaman menggunakan vektor alami
Agrobacterium telah menjadi hal rutin dalam perakitan tanaman transgenik. Bakteri ini memiliki kemampuan dalam menyalin, memindahkan, dan menyisipkan daerah T-DNA ke dalam kromosom tanaman yang berada pada vektor biner.
Vektor biner pC13-35S-Intron-Sma memungkinkan gen target yang disisipkan pada situs SmaI (CCCGGG) diatur ekspresinya oleh promoter kuat 35S dan terminator Nos untuk membentuk cassette ekspresi dengan cepat. Gen target akan mendapat tambahan intron (cat1) di ujung 5’- dan 6 kali histidin di ujung karboksil. Keberadaan intron untuk memastikan ekspresi gen target tidak berasal dari Agrobacterium. Komponen 6 kali histidin dapat berfungsi sebagai penanda untuk purifikasi dan deteksi pada tingkat protein.
Untuk mengetahui keberhasilan dari konstruksi pC13-35S-Intron-Sma disisipkan gen enhanced green fluorescent protein (eGFP) pada situs SmaI membentuk pC13-35S-Intron-eGFP dan ditransformasi ke padi varietas Nipponbare menggunakan Agrobacterium EHA105. Infeksi dilakukan pada kalus yang berumur lima hari dan ditanam pada media kultivasi. Pengamatan kalus transforman yang berusia satu bulan setelah infeksi menunjukkan adanya sinyal ekspresi eGFP dengan terbentuknya pendaran berwarna hijau di bawah sinar biru. Hasil ini menunjukkan bahwa vektor biner pC13-35S-Intron-Sma dapat digunakan untuk transformasi tanaman dengan Agrobacterium.
SUMMARY
NATALIA LUSIANINGSIH SUMANTO. pC13-35S-Intron-Sma: a flexible binary vector for Agrobacterium-mediated constitutive expression of target gene in plant. Supervised by Prof. Dr. Ir. Antonius Suwanto, M.Sc and Dr. Sigit Purwantomo.
Insertion technique of foreign gene into plants using natural vector
Agrobacterium has become a routine practice to generate transgenic plants.
Agrobacerium has an ability to -copy, -move and -insert a DNA fragment from its T-DNA region located in a binary vector into plant chromosome.
The binary vector pC13-35S-Intron-Sma allows a target gene to be inserted in SmaI site (CCCGGG). The gene will be directed by a strong promoter 35S of CaMV to create an expression cassette and get an intron (cat1) at 5’- end and a hexa histidine at its carboxyl end. The presence of intron will assure the expression of gene is derived from the plant but not Agrobacterium. Hexa histidine could serve as Tag for protein purification and detection.
The enhanced green fluorescent protein (eGFP) was inserted at SmaI site of pC13-35S-Intron-Sma to create pC13-35S-Intron-eGFP. This construct was used to test the expression of the gene with additional intron driven by the 35S promoter. The 5 days of early developed calli of Nipponbare rice were infected with Agrobacterium EHA105 harboring pC13-35S-Intron-eGFP followed by 3 days co-cultivation. Some calli in selection medium containing 50 mg L-1 hygromycin showed eGFP expression at one month after infection. This result indicated that the binary vector pC13-35S-Intron-Sma might be use for Agrobacterium-mediated plant transformation.
DAFTAR ISI
DAFTAR TABEL vi
DAFTAR GAMBAR vi
1 PENDAHULUAN 1
Latar Belakang 1
Perumusan Masalah 2
Tujuan Penelitian 2
Manfaat Penelitian 2
Ruang Lingkup Penelitian 2
2 TINJAUAN PUSTAKA 3
Transfer gen melalui perantaraan Agrobacterium tumefaciens 3
Sistem ekspresi vektor biner 4
Promoter 35S-Intron-Sma 4
Green fluoresence protein (eGFP) 5
3 METODE 5
Waktu dan Tempat Penelitian 5
Bahan 6
Alat 6
Amplifikasi fragmen DNA 6
Kloning frgamen DNA ke pGEM-T Easy 7
Konstruksi plasmid pC-13-35S-Intron-Sma 8
Transformasi Agrobacterium tumefaciens 8
Transformasi kalus padi Nipponbare 9
Isolasi DNA genom tanaman 9
4 HASIL DAN PEMBAHASAN 10
Konstruksi vektor biner pC13-35S-Intron-Sma 10
Konstruksi pC13-35S-Intron-eGFP 11
Transformasi kalus Padi Nipponbare 12
Pembahasan 14
5 SIMPULAN DAN SARAN 17
Simpulan 17 Saran 17
DAFTAR PUSTAKA 18
DAFTAR TABEL
1 Daftar primer yang digunakan dalam konstruksi vektor
pC-35S-Intron-Sma 6
DAFTAR
GAMBAR
1 Struktur dasar T-DNA plasmid pCambia 1301 1
2 Struktur T-DNA plasmid biner fleksibel 2
3 Mekanisme transfer daerah T-DNA dari plasmid Ti Agrobacterium
tumefaciens ke dalam kromosom tanaman 3
4 Sistem ekspresi vektor biner 4
5 Konstruksi umum daerah T-DNA pada plasmid biner 4
6 Peta konstruksi plasmid pC13-35S-Intron-Sma 10
7 Hasil amplifikasi fragmen promoter 35S-Intron-Sma 10 8 Hasil verifikasi koloni yang membawa plasmid pC13-35S-Intron-Sma 11 9 Hasil verifikasi koloni transforman yang membawa pGEM-eGFP 11 10 Hasil verifikasi koloni yang membawa plasmid pC13-35S-Intron-eGFP 12 11 Hasil verifikasi koloni Agrobacterium yang membawa
pC13-35S-Intron-eGFP 12
12 Hasil pengamatan ekspresi gen eGFP 13
13 Hasil pengamataneEkspresi gen eGFP di kalus transforman 13 14 Hasil verifikasi penyisipan T-DNA pC13-35S-Intron-eGFP pada empat
1
PENDAHULUAN
Latar Belakang
Teknik pemindahan dan penyisipan gen target ke dalam tanaman menggunakan vektor alami Agrobacterium telah menjadi hal rutin dalam merakit tanaman transgenik. Teknik ini merupakan teknik yang relatif ekonomis dibandingkan dengan teknik lain seperti penembakan biolistik. Vektor biner dibutuhkan untuk membawa cassette gen target ke inti dan menyisipkannya di kromosom tanaman.
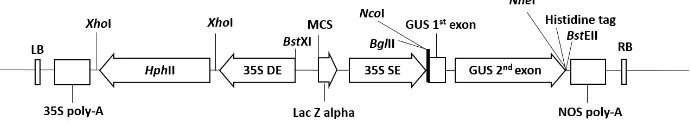
Salah satu vektor biner yang dapat diakses bebas adalah plasmid biner dari Cambia yaitu pCambia 1301 (Gambar 1). Gen GUS (β-Glucuronidase) dapat langsung digantikan oleh gen target untuk diekspresikan secara konstitutif di bawah kendali promoter 35S, akan tetapi penggantian gen GUS mengakibatkan komponen penting dari plasmid ini yaitu intron dan 6 kali histidin menjadi hilang.
Gambar 1 Struktur dasar T-DNA plasmid pCambia 1301
Keberadaan intron dapat digunakan untuk memastikan ekspresi gen tidak berasal dari Agrobacterium melainkan dari tanaman. Beberapa intron yang telah digunakan antara lain adalah intron hsp70 dari jagung (Pang et al. 1996), intron dari gen katalase cat1 dari Ricinus communis (Ohta et al. 1990), dan intron gen ST-LS1 dari kentang (Vancanneyt et al. 1990). Intron gen katalase cat1 dipilih karena ukurannya yang relatif kecil (190 pb) dan dapat digabungkan dengan gen penanda
GUSpada ujung 5’- (Ohta et al. 1990) atau diletakkan di tengah gen hphII (Wang
et al. 1997). Sehingga, kemungkinan besar intron ini dapat juga ditambahkan pada
ujung 5’- gen yang lain.
Keberadaan 6 kali histidin sebagai penanda penting untuk keperluan purifikasi protein yang diekspresikan gen target. Di samping itu, keberadaan 6 kali histidin sebagai penanda dapat digunakan untuk verifikasi ekspresi gen target pada level protein. Pada protein dengan antibodi yang belum tersedia deteksi akan dapat dilakukan dengan menggunakan antibodi untuk 6 kali histidin.
Gen enhanced green fluorescent protein (eGFP) digunakan untuk memverifikasi proses intron splicing. Berbeda dengan gen GUS dan LUC
2
Perumusan Masalah
Dalam penyisipan cassette gen target ke dalam kromoson tanaman salah satu hal yang paling esensial adalah konstruksi plasmid yang membawa cassette
gen target tersebut. Namun, konstruksi plasmid membutuhkan waktu yang tidak sebentar. Sehingga untuk mempermudah dan mempersingkat waktu dalam konstruksi plasmid dibutuhkan plasmid fleksibel yang komponen-komponen pada
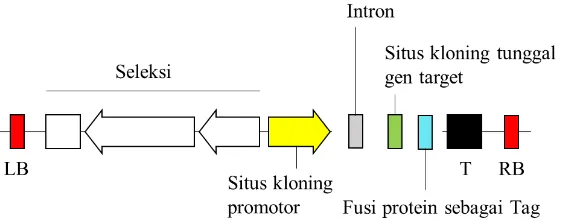
cassette tersebut dapat diubah-ubah secara mudah. Oleh karena itu, perlu dikonstruksi plasmid biner fleksibel yang memiliki situs restriksi yang terbatas pada daerah promoter, situs kloning tunggal untuk menyisipkan gen target serta
dilengkapi intron pada ujung 5’ dan 6 kali histidin pada ujung karboksil (Gambar 2). Adanya plasmid biner fleksibel ini diharapkan dapat mempersingkat waktu dalam mengkonstruksi plasmid dan tidak perlu mengkonstruksi plasmid baru apabila ingin mengganti daerah promoter atau gen target.
Gambar 2 Struktur T-DNA plasmid biner fleksibel
Tujuan Penelitian
Penelitian ini bertujuan untuk mengkonstruksi plasmid biner fleksibel yang dapat mengekspresikan gen target secara konstitutif yang dilengkapi dengan 6 kali histidin sebagai penanda dan intron untuk memastikan bahwa ekspresi gen target tidak berasal dari bakteri.
Manfaat Penelitian
Vektor biner fleksibel pC13-35S-Intron-Sma yang dikonstruksi dalam penelitian ini merupakan vektor dasar yang dapat digunakan untuk mengekspresikan gen target yang bersifat fungsional pada tanaman secara konstitutif di bawah kendali promoter 35S atau promoter konstitutif lainnya seperti OsAct2 dan OsGOS2 dengan menganti promoter 35S pada situs restriksi tertentu.
Ruang Lingkup Penelitian
3
2
TINJAUAN PUSTAKA
Transfer gen melalui perantaraan Agrobacterium tumefaciens
Teknik transformasi gen ke dalam tanaman didasari oleh penemuan bakteri tanah Agrobacterium tumefaciens yang merupakan patogen penyebab penyakit
crown gall di dalam jaringan luka pada berbagai tanaman dikotil. Bagian DNA
A. tumefaciens yang ditransfer ke dalam kromosom tanaman berupa T-DNA yang berada dalam plasmid Ti yang berukuran besar (150-250 kb) (Rossi et al. 1998). Di dalam nukleus T-DNA diintegrasikan ke dalam kromosom tanaman dengan cara
illegitimate recombination yaitu suatu mekanisme bergabungnya dua molekul DNA yang tidak mempunyai homologi secara luas (Offringa et al. 1990). Pada dasarnya A. tumefaciens memberikan respon kemotaksis terhadap senyawa fenol yang dilepaskan oleh jaringan tanaman yang terluka dan bergerak menurut gradien konsentrasi menuju sel yang terluka. Respon kemotaksis merupakan ekspresi konstitutif dari gen-gen kromosomal A. tumefaciens yaitu chvA, chvB, pscA dan att
(Ziemienowicz 2000).
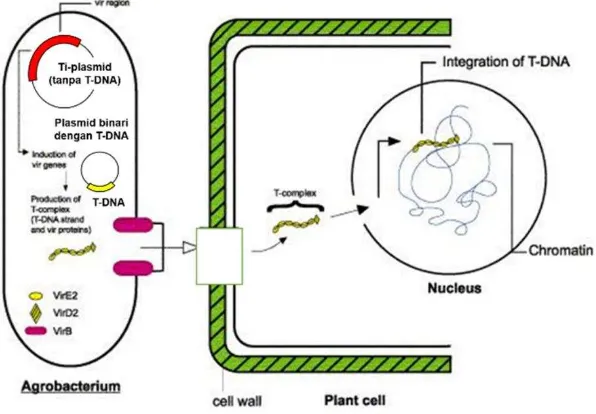
Kontak A. tumefaciens dengan senyawa acetosyringone yang dilepaskan oleh tanaman yang terluka menginduksi transkripsi daerah vir pada plasmid Ti.
Acetosiryngone kemudian berinteraksi dengan virA dan menghasilkan sinyal intraseluler oleh aktivasi virG. Gen virG yang teraktivasi kemudian mengaktifkan gen virulen lainnya (virB, virC, virD dan virE). Induksi gen vir diikuti dengan pengenalan sekuen pembatas 25 pb sebagai imperfect direct repeat/border sequences yang mengapit T-DNA. Pembatas T-DNA kemudian dipotong oleh dua protein yang dihasilkan oleh operon virD yaitu virD1 dan virD2 (Filichkin dan Gelvin 1993), sehingga diperoleh daerah T-DNA yang akan ditransfer ke dalam kromosom tanaman (Gambar 3).
4
Sistem ekspresi vektor biner
Transfer daerah T-DNA A. tumefaciens ke dalam kromosom tanaman dilakukan dengan menggunakan vektor biner yang diperkenalkan oleh Hoekema (1983). Gen-gen vir dan daerah T-DNA berada dalam plasmid yang berbeda (Gambar 4). Selama kedua plasmid tersebut berada dalam Agrobacterium tumefaciens yang sama, protein yang dihasilkan oleh gen vir dapat membantu proses transfer daerah T-DNA ke dalam kromosom tanaman. Plasmid yang mengandung daerah T-DNA disebut dengan vektor biner sedangkan plasmid yang mengandung gen-gen vir disebut dengan vir helper. Pada proses transfer gen, vektor biner lebih efisien karena memiliki ukuran yang lebih kecil, memiliki gen penanda seleksi tanaman, situs pengenalan enzim retriksi, ori E. coli dan A. tumefaciens serta gen resisten antibiotik (Lee et al 2007).
Gambar 4 Sistem ekspresi vektor biner. (A) Gen target berada terletak di daerah
T-DNA pada vektor biner; (B) Gen vir berada pada vir helper
(Lee et al. 2007)
Secara umum daerah T-DNA pada plasmid biner dikonstruksi sehingga mengandung promoter yang mengendalikan ekspresi gen target di tanaman, terminator sebagai daerah pengenalan RNA polimerase dalam menghentikan proses transkripsi dan gen penanda seleksi tanaman (Gambar 5).
Gambar 5 Konstruksi umum daerah T-DNA pada plasmid biner
Promoter 35S-Intron-Sma
5
konstitutif pada Arabidopsis sp. (Niwa et al. 1999). Pada penelitian ini sekuen promoter 35S didesain mengandung intron (intron cat1), situs kloning gen target (SmaI) dan 6 kali histidin sebagai penanda.
Ciri khas struktur genom eukariotik adalah gen struktural yang tersusun dari intron dan ekson. Ekson merupakan urutan DNA yang akan ditranskripsi menjadi mRNA kemudian ditranslasi membentuk polipeptida sedangkan intron (intragenic regions) merupakan urutan DNA yang tidak ikut ditranslasi dan akan dibuang pada tahap pasca transkripsi (Bergman 2001). Walaupun intron merupakan bagian yang tidak ditranslasi namun intron berperan dalam pengaturan sintesis protein. Intron yang tidak terpotong pada mRNA mengakibatkan penyimpangan dalam ekspresi gen. Setiap intron pada gen eukariotik memiliki situs pengenalan yakni kedua basa pertama dari intron mengandung GU dan dua basa terakhir dari intron adalah AG serta kotak TACTAAC. Keseluruhan urutan basa tersebut sangat penting sebagai pengenalan enzim spliceosome untuk memotong intron dan menyambung ekson secara tepat (Deutsch dan Long 1999). Penambahan intron dapat mencegah ekspresi gen target di Agrobacterium.
Green Fluoresence Protein (GFP)
Protein GFP yang berasal dari Aequorea victoria merupakan salah satu jenis gen reporter yang umum digunakan sebagai penanda dalam transformasi tanaman (Cinelli et al. 2000). GFP adalah protein yang merupakan polimer dari 238 asam amino dengan berat molekul sekitar 27 kD. GFP mengandung gugus yang disebut
chomophore yang berperan penting dalam menghasilkan pendaran hijau.
Chromophore ini adalah kelompok tiga residu asam amino di posisi 65 (Serin), 66 (Tirosin) dan 67 (Glisin). Ketika dipaparkan pada cahaya biru (395 nm) maka pada gugus ini akan terjadi oksidasi sehingga energi yang diserap menyebabkan elektron-elektron di dalam gugus ini tereksitasi dan menghasilkan pendaran berwarna hijau (Heim et al. 1995). Dalam upaya meningkatkan ekspresi dari GFP maka dilakukan mutasi pada asam amino penyusun protein tersebut. Mutasi residu asam amino nomor 64 dari Fenilalanin menjadi Leusin dan residu asam amino nomor 65 dari Serin menjadi Tirosin membentuk enhanced GFP (eGFP). Gen eGFP memiliki pendaran lebih terang daripada GFP wildtype (Cormarck et al. 1996). Hal ini menyebabkan eGFP banyak digunakan sebagai gen penanda dalam transformasi.
3
METODE
Waktu dan Tempat Penelitian
5
konstitutif pada Arabidopsis sp. (Niwa et al. 1999). Pada penelitian ini sekuen promoter 35S didesain mengandung intron (intron cat1), situs kloning gen target (SmaI) dan 6 kali histidin sebagai penanda.
Ciri khas struktur genom eukariotik adalah gen struktural yang tersusun dari intron dan ekson. Ekson merupakan urutan DNA yang akan ditranskripsi menjadi mRNA kemudian ditranslasi membentuk polipeptida sedangkan intron (intragenic regions) merupakan urutan DNA yang tidak ikut ditranslasi dan akan dibuang pada tahap pasca transkripsi (Bergman 2001). Walaupun intron merupakan bagian yang tidak ditranslasi namun intron berperan dalam pengaturan sintesis protein. Intron yang tidak terpotong pada mRNA mengakibatkan penyimpangan dalam ekspresi gen. Setiap intron pada gen eukariotik memiliki situs pengenalan yakni kedua basa pertama dari intron mengandung GU dan dua basa terakhir dari intron adalah AG serta kotak TACTAAC. Keseluruhan urutan basa tersebut sangat penting sebagai pengenalan enzim spliceosome untuk memotong intron dan menyambung ekson secara tepat (Deutsch dan Long 1999). Penambahan intron dapat mencegah ekspresi gen target di Agrobacterium.
Green Fluoresence Protein (GFP)
Protein GFP yang berasal dari Aequorea victoria merupakan salah satu jenis gen reporter yang umum digunakan sebagai penanda dalam transformasi tanaman (Cinelli et al. 2000). GFP adalah protein yang merupakan polimer dari 238 asam amino dengan berat molekul sekitar 27 kD. GFP mengandung gugus yang disebut
chomophore yang berperan penting dalam menghasilkan pendaran hijau.
Chromophore ini adalah kelompok tiga residu asam amino di posisi 65 (Serin), 66 (Tirosin) dan 67 (Glisin). Ketika dipaparkan pada cahaya biru (395 nm) maka pada gugus ini akan terjadi oksidasi sehingga energi yang diserap menyebabkan elektron-elektron di dalam gugus ini tereksitasi dan menghasilkan pendaran berwarna hijau (Heim et al. 1995). Dalam upaya meningkatkan ekspresi dari GFP maka dilakukan mutasi pada asam amino penyusun protein tersebut. Mutasi residu asam amino nomor 64 dari Fenilalanin menjadi Leusin dan residu asam amino nomor 65 dari Serin menjadi Tirosin membentuk enhanced GFP (eGFP). Gen eGFP memiliki pendaran lebih terang daripada GFP wildtype (Cormarck et al. 1996). Hal ini menyebabkan eGFP banyak digunakan sebagai gen penanda dalam transformasi.
3
METODE
Waktu dan Tempat Penelitian
6
Bahan
Bahan yang digunakan dalam penelitian ini adalah media pertumbuhan bakteri Luria Agar (tripton 1 g, ekstrak khamir 0.5 g, NaCl 1g dan agar bakteriologi 1.5 g) media Luria Bertani (tripton 1 g, yeast ekstrak 0.5 g dan NaCl 1g), agarosa 1% (Choice care), bufer TAE [Tris-Acetate-EDTA] (242 g Tris, 57.1 ml asam asetat glasial, 100 ml EDTA 0.5 M) ethidium bromide, antibiotik ampisilin (100 mg ml-1), kanamisin (25 mg ml-1), sefotaksim (100 mg ml-1), higromisin (15 mg ml-1),
rifampisin (15 mg ml-1), kloramfenikol (25 mg ml-1), QIAPrep Purification Plasmid kit (Qiagen), QIAquick PCR purification gel kit (Qiagen), ddH2O, marker molekuler 1000 pb DNA ladder (Vivantis), VC 100 pb (Vivantis), 6x loading dye (Promega),
Escherichia coli TOP10 (Invitrogen), PCR Go Taq Green Master Mix (Promega), enzim T4 ligase (New England Biolabs), bufer T4 DNA ligase 10x (New England Biolabs), enzim restriksi (New England Biolabs), vektor kloning pGEM-T Easy (Promega), vektor ekspresi pCambia 1301, primer, polybag, tanah berpasir, kalus
padi varietas Nipponbare, media N6D, acetosyringone (20 mg L-1), 2,4-D (2 mg L-1), etanol 70%, natrium hipoklorit dan air steril.
Alat
Alat-alat yang digunakan dalam penelitian ini adalah mesin thermocycler, spektrofotometer, Geldoc UV Transilluminator (Biorad), NanoDrop 2000 (Invitrogen), sentrifus, microwave, inkubator bergoyang, autoklaf, vorteks,
waterbath, laminar air flow, oven, Genetic Analyzer ABI 3130, Tissue lyser (Qiagen), cawan Petri, Erlenmeyer, tabung 1.5 ml, mikropipet dan tip.
Prosedur Penelitian Amplifikasi fragmen DNA
Amplifikasi fragmen DNA menggunakan metode PCR dengan denaturasi awal dilakukan selama 5 menit pada suhu 98oC, dilanjutkan dengan siklus PCR yang terdiri dari denaturasi (30 detik 98oC), annealing (55oC selama 30 detik dan pemanjangan primer (72oC dengan perhitungan 1 kb/menit) sebanyak 40 siklus). Daftar primer yang digunakan untuk amplifikasi fragmen terlampir pada tabel 1. Visualisasi produk PCR dilakukan dengan elektroforesis pada gel agarosa 1% (b/v) dilanjutkan dengan pengamatan di bawah sinar UV.
Tabel 1 Daftar primer yang digunakan dalam konstruksi vektor pC-35S-Intron-Sma
No Primer Situs
35S-Intron-Sma pC1301 pGem-35S-Intron-Sma 1020 2 Intron-extraAA-SmaI-R BstEII
7
Kloning fragmen DNA ke pGEM-T Easy Purifikasi fragmen
Fragmen DNA hasil PCR terlebih dahulu dikloning pada vektor pGEM-T Easy (Promega). Fragmen hasil amplifikasi dipurifikasi menggunakan QIAquick PCR purification gel kit (Qiagen). Gel agarosa yang mengandung fragmen DNA yang diingiinkan dipotong dan dimasukkan ke dalam tabung 1.5 ml dan ditimbang. Ditambahkan bufer QG sebanyak 3 kali berat gel kemudian diinkubasi 60oC, 10 menit. Sebanyak 500 µl campuran dimasukkan ke dalam spin column, disentrifugasi 13000 rpm, 1 menit. Berturut-turut dimasukkan 750 µl bufer PE dan 500 µl bufer PB kemudian masing-masing disentrifugasi 13000 rpm, 1 menit. Sebanyak 30 µl bufer EB ditambahkan dan disentrifugasi 13000 rpm, 1 menit.
Ligasi fragmen DNA ke pGEM-T Easy
Penyambungan fragmen menggunakan enzim ligase yang dilakukan mengikuti Sambrook et al. (2001). Komposisi ligasi terdiri dari bufer T4 ligase, vektor pGEM-T Easy, fragmen DNA, enzim T4 ligase dan ddH2O dengan volume total 10 µl. Campuran diinkubasi pada suhu ruang semalaman.
Pembuatan Eschericia coli TOP10 kompeten
Pembuatan E. coli TOP10 kompeten menggunakan perlakuan CaCl2 dan MgCl2 sesuai metode Tang et al. (1994) dengan beberapa modifikasi. Koloni tunggal E. coli TOP10 dikultur di dalam 3 ml LB dan diinkubasi dalam inkubator bergoyang 200 rpm, 37oC semalaman. Sebanyak 200 µl kultur dimasukkan ke dalam 10 ml LB yang baru kemudian diinkubasi 200 rpm, 37oC sampai OD600=0.4. Sebanyak 1.5 ml kultur dimasukkan ke dalam tabung 1.5 ml, disentrifugasi 5000 rpm, 4oC 1 menit dan dibuang supernatan. Pelet diresuspensi dengan 1 ml CaCl
2 (20 mM CaCl2 dan 80 mM MgCl2) dan diinkubasi 20 menit dalam es. Campuran disentrifugasi 5000 rpm, 4oC 1 menit dan dibuang supernatan. Pelet diresuspensi kembali dengan 250 µl CaCl2 0.1 M kemudian diinkubasi 10 menit dalam es. Sebanyak 100 µl gliserol 50% ditambahkan ke dalam campuran.
TransformasiEschericia coli TOP10
Transformasi E. coli TOP10 kompeten menggunakan metode kejut panas sesuai dengan Sambrook et al. (2001). Sebanyak 10 µl plasmid dimasukkan ke dalam 100 µl sel kompeten dan diinkubasi 15 menit di dalam es. Campuran diberikan perlakukan kejut panas 42oC 90 menit kemudian diinkubasi 15 menit di dalam es. Media Luria Bertani (LB) 1 ml ditambahkan ke dalam campuran kemudian diinkubasi dalam inkubator bergoyang 200 rpm, 37oC 1 jam. Campuran disentrifugasi 13000 rpm selama 1 menit dan supernatan dibuang. Sebanyak 25 µl campuran disebar pada media agar LB yang telah mengandung antibiotik dan diinkubasi 37oC semalaman.
Sekuensing dan analisis hasil kloning
Fragmen DNA sisipan disekuensing menggunakan ABI 3130 Genetic Analyzer. Siklus sequencing dilakukan menggunakan primer yang sesuai dan hasil
8
Isolasi plasmid yang mengandung fragmen sisipan
Isolasi plasmid dilakukan menggunakan QIAPrep Purification Plasmid kit. Koloni tunggal bakteri transforman ditumbuhkan dalam 10 ml LB dan diinkubasi dalam inkubator bergoyang 200 rpm, 37oC semalaman. Sebanyak 1.5 ml kultur dimasukkan ke dalam tabung 1.5 ml dan disentrifugasi 6000 rpm, 1 menit. Supernatan dibuang lalu pelet diresuspensi berturut-turut dengan 250 µl bufer P1, 250 µl bufer P2 dan 300 µl bufer N3. Sebanyak 750 µl campuran dimasukkan ke dalam spin column dan disentrifugasi 6000 rpm 1 menit. Berturut-turut dimasukkan 500 bufer PB dan 750 bufer PE kemudian masing-masing disentrifugasi 13000 rpm 1 menit. Sebanyak 30 µl bufer EB ditambahkan dan disentrifugasi 13000 rpm 1 menit.
Konstruksi plasmid pC13-35S-Intron-Sma
Konstruksi plasmid yang melibatkan pemotongan utas DNA dengan enzim restriksi dan penyambungan utas DNA dengan enzim ligase dilakukan mengikuti Sambrook et al. (2001). Fragmen promoter 35S-Intron-Sma diisolasi dengan pemotongan fragmen menggunakan enzim restriksi yang sesuai. Komposisi restriksi terdiri dari bufer enzim restriksi, plasmid rekombinan, enzim restriksi dan H2O dengan volme total 20 µl. Campuran diinkubasi sesuai dengan suhu inkubasi enzim yang digunakan selama 1 jam. Hasil pemotongan divisualisaasi pada gel agarosa 1% dan dilakukan purifikasi menggunakan QIAquick PCR purification gel kit (Qiagen).
Plasmid biner pCambia 1301 dipotong dengan EcoRI dan BstEII untuk menghilangkan fragmen promoter 35S, GUS dan multiple cloning site (MCS).
Backbone diligasi dengan promoter 35S-Intron-Sma untuk mendapatkan plasmid pC13-35S-Intron-Sma. Gen eGFP disisipkan pada situs SmaI yang terdapat pada plasmid pC13-35S-Intron-Sma sebagai gen reporter untuk mengetahui aktivitas promoter 35S.
Transformasi Agrobacterium tumefaciens
Sel kompeten A. tumefaciens EHA105 (Hood et al. 1993) dibuat dengan perlakuan CaCl2 dan transformasi vektor biner dilakukan menggunakan metode
freeze-thaw (Weigel dan Glazebrook 2002). Koloni tunggal A. tumefaciens
EHA105 digores pada LB padat yang mengandung antibiotik rifampisin 25 mg ml-1 diinkubasi 28oC selama 3 hari. Koloni yang tumbuh discrab lalu
dimasukkan ke dalam 10 ml LB yang mengandung antibiotik dan diinkubasi dalam inkubator bergoyang 200 rpm, 28oC semalaman. Kultur dimasukkan ke dalam 10 ml LB yang mengandung antibiotik sampai OD600=0.6. Sebanyak 1.5 ml kultur dimasukkan ke dalam tabung 1.5 ml lalu disentrifugasi 4000 rpm, 4oC 10 menit. Supernatan dibuang dan pelet diresuspensi dengan 1 ml CaCl2 20 mM kemudian disentrifugasi 4000 rpm, 4oC 1 menit. Supernatan dibuang dan pelet diresuspensi kembali dengan 200 µl CaCl2 20 mM.
9
ditambahkan ke dalam campuran dan diinkubasi di inkubator bergoyang 200 rpm, 28oC 4 jam. Campuran disentrifugasi 13000 rpm 1 menit dan sebanyak 25 µl disebar pada LB padat yang mengandung antibiotik.
Transformasi kalus padi Nipponbare
Transformasi kalus padi dilakukan menggunakan metode Toki et al. (2006). Kulit biji padi dibuka kemudian biji padi disterilkan menggunakan air sabun sambil digoyang 200 rpm selama 15 menit. Biji padi dibilas dengan air steril sampai busa hilang. Sterilisaasi dilanjutkan dengan memasukkan biji padi berturut-turut ke dalam EtOH 70% selama 1 menit dan natrium hipoklorit 4% sambil digoyang 200 rpm selama 30 menit. Biji padi dibilas menggunakan air steril sebanyak 5 kali masing-masing selama 15 menit sambil digoyang 200 rpm. Selanjutnya biji padi dikeringkan dengan kertas saring steril dan ditanam dalam media N6D, 32oC, 5 hari pada kondisi terang.
Tahapan infeksi Agrobacterium dilakukan dengan memasukkan kalus yang berusia 5 hari ke dalam kultur Agrobacterium (OD600=0.1) yang mengandung
acetosyringone lalu didiamkan 1.5 menit. Kalus dikeringkan dengan kertas saring steril dan ditanam dalam media N6D yang mengandung acetosyringone, 20oC, 3 hari pada kondisi gelap. Kalus dibilas dengan air steril sebanyak dua kali 2 menit digoyang. Selanjutnya kalus dibilas kembali dengan air steril yang mengandung antibiotik sebanyak 5 kali masing-masing selama 15 menit. Kalus dikeringkan dengan kertas saring steril dan ditanam dalam media N6D yang mengandung antibiotik 250 mg L-1 sefotaksim dan 200 mg L-1 ampisilin, 32oC, pada kondisi terang dan disubkultur setiap 2 minggu. Seleksi kalus putative transgenik menggunakan media yang mengandung 50 mg L-1 higromisin, 250 mg L-1 sefotaksim dan 200 mg L-1 ampisilin dan dilakukan subkultur setiap 2 minggu.
Isolasi DNA genom tanaman
10
4
HASIL DAN PEMBAHASAN
Konstruksi vektor biner pC13-35S-Intron-Sma
Langkah awal yang dilakukan untuk mengkonstruksi plasmid pC13-35S-Intron-Sma adalah melakukan amplifikasi fragmen promoter 35S-Intron-pC13-35S-Intron-Sma. Pada bagian N-terminal gen target terdapat penambahan sepuluh asam amino dan diharapkan dengan penambahan tersebut intron pada pC13-35S-Intron-Sma akan mengalami proses splicing
seperti pada pC1301. Modifikasi ini juga diharapkan tidak mengganggu fungsi gen target yang disisipkan. Fragmen promoter 35S-Intron-Sma mengandung intron yang memiliki masing-masing lima asam amino gen GUS dari pC1301 pada ujung 5’ dan 3’, situs
kloning gen target (SmaI), 6 kali kodon histidin dan stop kodon TGA. Pada fragmen ini
ditambahkan situs restriksi EcoRI pada ujung 5’ dan BstEII pada ujung 3’ promoter 35S (Gambar 6)
Gambar 6 Peta konstruksi plasmid pC13-35S-Intron-Sma
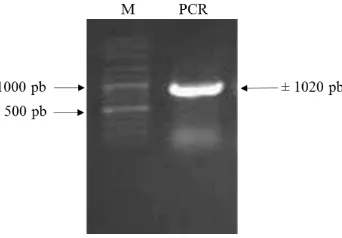
Fragmen promoter 35S-Intron-Sma yang diamplifikasi dengan PCR menghasilkan pita berukuran 1020 pb pada gel agarosa (Gambar 7). Fragmen ini dipurifikasi, diligasi ke pGEM-T Easydan diintroduksikan pada sel kompeten E. coli TOP10. Koloni yang membawa pGEM-35S-Intron-Sma diseleksi pada media yang mengandung ampisilin 100 mg L-1. Verifikasi koloni yang membawa plasmid rekombinan diverifikasi dengan PCR menggunakan primer khusus 35S-Intron-Sma_F dan 35S-Intron-Sma_R dan verifikasi keberadaan fragmen promoter 35S-Intron-Sma juga dilakukan dengan restriksi plasmid rekombinan menggunakan EcoRI dan BstEII. Hasil verifikasi PCR koloni dan restriksi plasmid rekombinan menghasilkan pita berukuran 1020 pb.
11
Fragmen 35S-Intron-Sma dari pGemT-35S-Intron-Sma disubkloning ke pC1301 yang telah dipotong dengan enzim restriksi EcoRI dan BstEII untuk menghilangkan daerah MCS, promoter 35S dan gen GUS. Plasmid pC1301 yang telah dimodifikasi disebut dengan Intron-Sma. Plasmid pC13-35S-Intron-Sma diintrodukasi pada sel kompeten E. coli TOP10. Koloni yang membawa pC13-35S-Intron-Sma ditapiskan pada media yang mengandung kanamisin 50 mg L-1 dan diverifikasi dengan PCR menggunakan primer khusus dan menghasilkan pita berukuran 1020 pb (Gambar 8).
Gambar 8 Hasil verifikasi koloni yang membawa plasmid pC13-35S-Intron-Sma. Koloni No. 4 positif membawa plasmid pC13-35S-Intron-Sma (M= penanda 1000 pb DNA ladder; K(-)= kontrol negatif)
Konstruksi pC13-35S-Intron-eGFP
Gen eGFP diamplifikasi dengan PCR dari plasmid pST2 dan dikloning ke pGEM-T Easy. Plasmid pGEM-eGFP diintroduksi pada sel kompeten E. coli
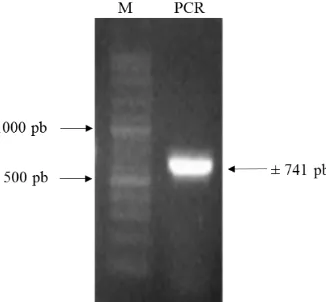
TOP10. Koloni yang membawa plasmid ini ditapiskan pada media yang mengandung ampisilin 100 mg L-1 dan diverifikasi dengan PCR menggunakan primer eGFP_F dan eGFP_R. Hasil verifikasi PCR menghasilkan pita berukuran 741 pb (Gambar 9).
Gambar 9 Hasil verifikasi koloni transforman yang membawa pGEM-eGFP
(M= penanda 100 pb)
12
dilakukan dengan 35S-Intron-Sma_F dan eGFP_R untuk melihat orientasi dari eGFP. Hasil verifikasi PCR menghasilkan pita berukuran 1761 pb (Gambar 10).
Gambar 10 Hasil verifikasi koloni yang membawa plasmid pC13-35S-Intron-eGFP (M= penanda 1000 pb DNA ladder; K(-)= kontrol negatif)
Transformasi kalus padi Nipponbare
Plasmid pC13-35S-Intron-eGFP diintroduksi pada Agrobacterium
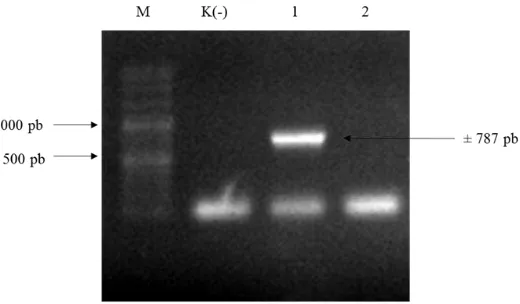
kompeten dan diseleksi pada media yang mengandung antibiotik rifampisin 25 mg L-1, klorampenikol 35 mg L-1 dan kanamisin 200 mg L-1. Koloni Agrobacterium yang membawa pC13-35S-Intron-eGFP diverifikasi dengan PCR menggunakan primer ujung intron_F dan 6xHis-TGA-BstEII-R dan menghasilkan pita berukuran 787 pb (Gambar 11).
Gambar 11 Hasil verifikasi koloni Agrobacterium yang membawa pC13-35S-Intron-eGFP. Koloni No.1 positif membawa pC13-35S-Intron-eGFP (M= penanda 1000 pb DNA ladder; K(-)= kontrol negatif)
Sebelum dilakukan transformasi pada tanaman model, koloni
13
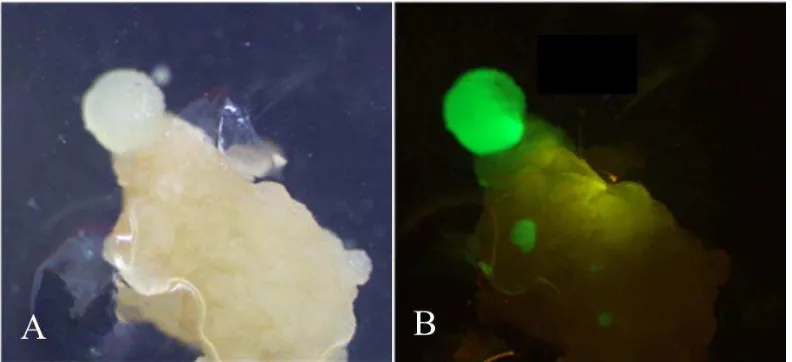
Agrobacterium yang mengandung pC-35S-Intron-eGFP tidak menunjukkan ekspresi gen eGFP (Gambar 12).
Gambar 12 Hasil pengamatan ekspresi gen eGFP di bawah sinar putih (A) dan sinar biru (B)
Transformasi dilakukan pada kalus padi Nipponbare. Transformasi dilakukan melalui ko-kultivasi kalus selama 3 hari. Kalus hasil ko-kultivasi diseleksi pada media seleksi menggandung antibiotik higromisin 50 mg L-1. Hasil transformasi menunjukkan bahwa terdapat empat galur kalus yang membawa sisipan T-DNA plasmid pC-35S-Intron-eGFP. Hal ini diketahui dari pengamatan ekspresi gen eGFP pada kalus di bawah sinar biru. Ekspresi eGFP diamati pada kalus padi yang berumur empat minggu setelah infeksi (Gambar 13).
Gambar 13 Hasil pengamatan ekspresi gen eGFP di bawah sinar putih (A) dan sinar biru (B)
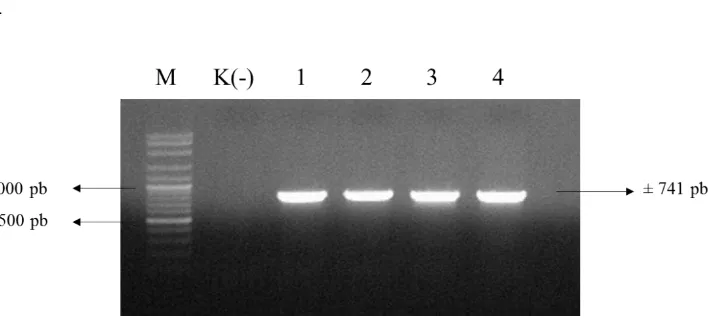
Verifikasi keberadaan T-DNA pC13-35S-Intron-eGFP pada empat galur kalus transgenik dilakukan dengan mengamplifikasi genom kalus transgenik menggunakan primer eGFP_F dan eGFP_R menghasilkan pita berukuran 741 pb (Gambar 14).
14
Gambar 14 Hasil verifikasi keberadaan T-DNA pC13-35S-Intron-Sma pada empat galur kalus transgenik (M= penanda 100 pb)
Pembahasan
Penyisipan gen ke dalam kromoson tanaman melalui perantaraan
Agrobacterium merupakan suatu metode transformasi yang telah berlangsung lama di alam. Hal ini disebabkan karena A. tumefaciens merupakan bakteri tanah yang secara alami mampu melakukan transfer DNA ke dalam kromoson tanaman. Bakteri ini mampu menyisipkan daerah T-DNA yang merupakan bagian dari Ti-plasmid. Di dalam T-DNA terdapat gen onkogenik penyandi fitohormon dan gen penyandi opin. Secara alami gen-gen yang berada di dalam T-DNA tersebut tidak dapat diekspresikan oleh Agrobacterium karena enzim RNA polimerase
Agrobacterium tidak mengenali daerah promoter gen-gen tersebut. Oleh karena itu,
Agrobacterium menyisipkan daerah T-DNA tersebut ke dalam kromoson tanaman sehingga gen-gen tersebut diekspresikan oleh tanaman karena RNA polimerase tanaman dapat mengenali daerah promoter gen-gen tersebut.
15
Oleh karena itu untuk mempermudah proses konstruksi daerah T-DNA tersebut dilakukan modifikasi pada plasmid pCambia 1301.
Plasmid hasil modifikasi pCambia 1301 disebut dengan pC13-35S-Intron-Sma. Plasmid ini didesain agar memiliki struktur yang lebih sederhana dan fleksibel sehingga akan mempermudah proses konstruksi kaset ekspresi. Daerah T-DNA pC13-35S-Intron-Sma tersusun dari kaset ekspresi gen target yang terdiri dari promoter 35S yang memiliki situs restriksi yang terbatas. Situs pengenalan enzim restriksi EcoRI dan BstXI terdapat pada daerah upstream promoter 35S serta NcoI dan BglII pada daerah downstream promoter 35S. Selain itu, terdapat intron pertama dari gen katalase (cat1), situs restriksi tunggal sebagai tempat penyisipan gen target (SmaI) dan 6 kali kodon histidin sebagai penanda. Promoter 35S merupakan promoter kuat yang dapat mengendalikan ekspresi gen target secara konstitutif. Manipilasi promoter tersebut relatif lebih mudah karena memiliki ukuran sekuen yang pendek (343 pb), tidak memiliki situs pengenalan enzim restriksi dan aktif pada tanaman monokotil maupun dikotil (Benfey et al. 1989). Promoter 35S merupakan promoter yang sangat kuat dan merupakan promoter yang mengendalikan ekspresi gen patogen yang menyerang tanaman sehingga promoter tersebut aktif di Agrobacterium. Vancanneyt et al. (1990) menyebutkan bahwa beberapa promoter yang mengendalikan ekspresi gen patogen tanaman dapat dikenali oleh sistem transkripsi Agrobacterium. Hal ini akan mengakibatkan gen target dapat terekspresi dibawah kendali promoter 35S pada Agrobacterium
sehingga menimbulkan bias pada saat transformasi.
Subkloning gen target pada pC13-35S-Intron-Sma dilakukan dengan metode kloning konvensional yang melibatkan PCR, enzim restriksi dan ligase. Proses insersi pada situs SmaI yang menghasilkan ujung tumpul memang lebih sulit dibanding dengan situs restriksi yang sticky-end dan membutuhkan verifikasi lanjutan dengan PCR untuk mengetahui posisi orientasi gen target. Akan tetapi teknik konvensional ini relatif feasible untuk dilakukan pada banyak laboratorium karena hanya membutuhkan satu jenis inang E. coli seperti DH5 atau TOP10. Teknik kloning dengan metode gateway mungkin menjadi menarik karena verifikasi orientasi tidak perlu dilakukan seperti pada seri vektor biner pGWB (Nakagawa et al. 2007). Menariknya, jenis vektor yang memanfaatkan intron cat1
tidak ada dalam daftar seri pGWB. Di samping itu, aplikasi metode gateway lebih kompleks daripada teknik kloning konvensional. Walaupun demikian, ada kemungkinan untuk mengubah pC13-35S-Intron-Sma untuk menjadi vektor destinasi dengan sistem gateway.
Keberadaan intron pada konstruksi ini merupakan hal yang penting karena mampu meningkatkan ekspresi gen target dan intron juga dapat digunakan untuk memastikan bahwa ekspresi gen target tidak berasal dari Agrobacterium melainkan
16
penyisipan gen target yang diamplifikasi dengan menggunakan enzim DNA polimerase yang tidak menambahkan basa nitrogen A pada bagian ujung, misalnya menggunakan DNA polimerase Pfu yang diikuti dengan seleksi bakteri transforman yang mengandung plasmid rekombinan dengan arah orientasi gen target yang sesuai.
Promoter 35S pada pC13-35S-Intron-Sma dapat diganti dengan promoter konstitutif lain pada situs EcoRI dan BstXI pada ujung 5’ serta NcoI dan BglII pada
ujung 3’. Intron dapat dihilangkan dengan menyisipkan gen target pada situs NcoI atau BglII dan SmaI. Komponen tambahan 6 kali histidin pada pC13-35S-Intron-Sma secara langsung akan ditambahkan jika gen target disisipkan pada situs tersebut. Komponen 6 kali histidin tidak akan diekspresikan apabila fragmen gen target mengandung stop kodon. Vektor pC13-35S-Intron-Sma telah didesain mengandung stop kodon TGA setelah 6 kali kodon histidin sehingga fragmen gen target diamplifikasi tanpa mengandung stop kodon. Komponen 6 kali histidin dapat digunakan sebagai penanda untuk purifikasi di tingkat protein serta dapat digunakan untuk purifikasi protein yang belum memiliki antibodi. Kemungkinan lain yang digunakan apabila tidak menginginkan penambahan intron dan 6 kali histidin pada gen target adalah menyisipkan gen target pada situs EcoRI atau BstXI dan BstEII.
Gen eGFP merupakan gen reporter yang digunakan untuk mengetahui aktivitas promoter, pengaruh penambahan asam amino dan proses splicing intron
cat1 pada pC13-35S-Intron-Sma. Gen eGFP diamplifikasi dari plasmid pST2 mulai dari start kodon tetapi tidak mengikutsertakan stop kodon dengan menambahkan situs enzim restriksi SmaI dan EcoRV pada bagian forward dan reverse. Gen eGFP merupakan gen repoter yang digunakan untuk seleksi positif kalus trasngenik yang telah tersisipi gen target karena ekspresi gen eGFP tidak menyebabkan kematian pada kalus transgenik.
Pengujian ekspresi intron-eGFP dengan transformasi kalus padi Nipponbare melalui Agrobacterium
Transformasi plasmid pC-35S-Intron-eGFP pada kalus padi Nipponbare dilakukan dengan menggunakan A. tumefaciens karena secara alami mampu mentransfer T-DNA dari Ti-plasmid ke dalam genom tanaman sehingga transformasi dengan perantara A. tumefaciens relatif lebih efisien dibandingkan dengan metode artificial (Sahoo et al. 2011). Di samping itu, keuntungan transformasi melalui perantaraan A. tumefaciens adalah dapat menyisipkan T-DNA yang berukuran besar, salinan T-DNA di dalam kromoson tanaman sedikit dan ekspresi gen targetdi tanaman stabil (Lopez et al. 2004). Strain A. tumefaciens yang digunakan adalah strain EHA105 (Hood 1993) yang menghasilkan yang bersifat supervirulensi sehingga lebih efisien dalam menginfeksi kalus padi Nipponbare.
Padi Nipponbare merupakan varietas japonica yang umum digunakan sebagai tanaman model dalam transformasi tanaman karena memiliki masa pertumbuhan singkat yaitu sekitar tiga bulan, mudah ditransformasi karena memiliki ukuran genom yang relatif kecil (430 Mpb) dan genom padi ini sudah dipetakan dengan lengkap sehingga mudah dimanipulasi (Nishimura et al. 2006).
17
intron splicing tidak terjadi pada Agrobacterium dapat meyakinkan bahwa ekspresi yang terjadi adalah berasal dari sel tanaman. Apabila terdapat intron pada gen target maka Agrobacterium tidak mampu melakukan intron splicing sehingga mengakibatkan gen target tidak terekspresi pada Agrobacterium (Tanaka et al. 1990).
Proses kokultivasi kalus dan Agrobacterium yang mengandung plasmid pC13-35S-Intron-eGFP dilakukan pada media yang mengandung Acetosyringone. Penambahan senyawa fenolik ini bertujuan untuk meningkatkan efisiensi transfer T-DNA oleh Agrobacterium (Amoah et al. 2001). Setelah proses kokultivasi kemudian dilanjutkan dengan proses kultivasi yang menggunakan media dengan penambahan antibiotik ampisilin, sefotaksim dan higromisin. Kombinasi antibiotik ampisilin dan sefotaksim efektif untuk membunuh Agrobacterium sehingga tidak terjadi overgrowth sedangkan higromisin digunakan untuk menyeleksi kalus yang telah mengandung T-DNA. Jumlah kalus yang tahan dengan sistem seleksi berjenjang ini akan terus menurun sampai pada tahap regerasi karena hanya kalus yang benar-benar mengandung gen ketahanan terhadap higromisin yang dapat tumbuh pada media seleksi (Chakrabarty et al. 2002).
Keberadaan plasmid biner pC13-35S-Intron-Sma diharapkan dapat digunakan untuk mempermudah ekspresi gen secara konstitutif. Walaupun plasmid ini tidak ditujukan untuk mengkloning promoter, plasmid ini juga memiliki kemungkinan untuk mengubah promoter konstitutif 35S dengan promoter lain menggunakan enzim restriksi yang tersedia.
5
SIMPULAN DAN SARAN
Simpulan
Vektor biner pC35S-Intron-Sma yang mengandung gen eGFP intron untuk transformasi tanaman telah berhasil dikonstruksi. Signal pada kalus padi Nipponbare yang diamati di bawah sinar birumenunjukkan bahwa gen eGFP yang mengandung intron terekspresi di bawah kendali promoter 35S. Ekspresi gen eGFP tidak tampak pada A.tumefaciens yang diamati di bawah sinar biru sehingga dapat disimpulkan gen eGFP hanya terekspresi pada kalus transgenik dan terdapat proses
intron splicing pada konstruksi gen eGFP.
Saran
17
intron splicing tidak terjadi pada Agrobacterium dapat meyakinkan bahwa ekspresi yang terjadi adalah berasal dari sel tanaman. Apabila terdapat intron pada gen target maka Agrobacterium tidak mampu melakukan intron splicing sehingga mengakibatkan gen target tidak terekspresi pada Agrobacterium (Tanaka et al. 1990).
Proses kokultivasi kalus dan Agrobacterium yang mengandung plasmid pC13-35S-Intron-eGFP dilakukan pada media yang mengandung Acetosyringone. Penambahan senyawa fenolik ini bertujuan untuk meningkatkan efisiensi transfer T-DNA oleh Agrobacterium (Amoah et al. 2001). Setelah proses kokultivasi kemudian dilanjutkan dengan proses kultivasi yang menggunakan media dengan penambahan antibiotik ampisilin, sefotaksim dan higromisin. Kombinasi antibiotik ampisilin dan sefotaksim efektif untuk membunuh Agrobacterium sehingga tidak terjadi overgrowth sedangkan higromisin digunakan untuk menyeleksi kalus yang telah mengandung T-DNA. Jumlah kalus yang tahan dengan sistem seleksi berjenjang ini akan terus menurun sampai pada tahap regerasi karena hanya kalus yang benar-benar mengandung gen ketahanan terhadap higromisin yang dapat tumbuh pada media seleksi (Chakrabarty et al. 2002).
Keberadaan plasmid biner pC13-35S-Intron-Sma diharapkan dapat digunakan untuk mempermudah ekspresi gen secara konstitutif. Walaupun plasmid ini tidak ditujukan untuk mengkloning promoter, plasmid ini juga memiliki kemungkinan untuk mengubah promoter konstitutif 35S dengan promoter lain menggunakan enzim restriksi yang tersedia.
5
SIMPULAN DAN SARAN
Simpulan
Vektor biner pC35S-Intron-Sma yang mengandung gen eGFP intron untuk transformasi tanaman telah berhasil dikonstruksi. Signal pada kalus padi Nipponbare yang diamati di bawah sinar birumenunjukkan bahwa gen eGFP yang mengandung intron terekspresi di bawah kendali promoter 35S. Ekspresi gen eGFP tidak tampak pada A.tumefaciens yang diamati di bawah sinar biru sehingga dapat disimpulkan gen eGFP hanya terekspresi pada kalus transgenik dan terdapat proses
intron splicing pada konstruksi gen eGFP.
Saran
18
DAFTAR PUSTAKA
Amoah BK, Wu H, Sparks C, Jones HD. 2001. Factors influencing Agrobacterium-mediated transient expression of uidA in wheat inflorescence tissue. J. Exp. Bot. 52:1135-1142.
Benfey PN, Ren L, Chua NH. 1989. The CaMV 35S enhancer contains at least two domains which can confer different developmental and tissue-specific expression patterns. EMBO J. 8(8):2195-202.
Bergman CM, Kreitman M. 2001. Analysis of conserved noncoding DNA in drosophila reveals similar constraints in intergenic and intronic sequences cases. Cold Spring Harbor Laboratory Press. 11:1335-1344.
Chakrabarty R, Viswakarma N, Bhat SR, Kirti PB, Singh BD, Chopra VL. 2002.
Agrobacterium-mediated transformation of cauliflower: optimization of protocol and development of Bt-transgenic cauliflower. J. Biosci. 27:495-502.
Cormarck PB, Valdivia RH, Falkow S. 1996. FACS optimized mutants of the green fluorescent protein (GFP). Gene. 173:33-38.
Deutsch M, Long M. 1999. Intron-exon structures of eukaryotic model organisms.
Nucleid Acids Research. 27(15):3219-3228.
Elliot AR, Campbell JA, Dugdale B, Brettell RIS, Grof CPL. 1999. Green-fluorescent protein facilitates rapid in vivo detection of genetically transformed plant cells. Plant Cell Rep. 18:707-714.
Filchkin SA, Gelvin SB. 1993. Formation a putative relaxation intermediate during T-DNA processing directed by the Agrobacterium tumefaciens virD1, virD2 endonuclease. Mol. Microbiol 8:915-926.
Heim RA, Cubitt B, Tsien RY. 1995. Improved greenfluorescence. Nature. 373:663-664.
Hoekema A, Hirsch PR, Hooykaas PJJ, Schilperoort RA. 1983. A binary plant vector strategy based on separation of vir- and T-region of the
Agrobacterium tumefaciens Ti-plasmid. Nature. 303: 179-180.
Holme IB, Pedersen HB, Lange M, Holm PB. 2006. Transformation of barley (Hordeum vulgare L.) by Agrobacterium tumefaciens infection of in vitro cultured ovules. Plant Cell Rep. 25:1325-1335.
Hood EE, Gelvin SB, Melchers LS, Hoekema A. 1993. New Agrobacterium helper plasmids for gene transfer to plants. Transgenic Res. 2:208-218.
Jefferson RA. 1987. Assaying chimeric genes in plants: the GUS gene fusion gene.
Plant Mol Biol Rep. 4(5):387-405.
Jordan MC. 2000. Green fluorescent protein as a visual marker for wheat transformation. Plant Cell. 19:1069-1075.
Lee LY, Kononov ME, Bassuner B, Frame BR, Wang K, Gelvin SB. 2007. Novel plant transformation vectors containing the superpromoter. Plant Physiol
19
Nakagawa T, Kurose T, Hino T, Tanaka K, Kawamukai M, Niwa Y, Toyooka K, Matsuoka K, Jinbo T, Kimura T. 2007. Development of series of gateway binary vectors, pGWBs, for realizing efficient construction of fusion genes for plant transformation. Journal of Bioscience and Bioengineering. 1(104): 34-41.
Nishimura A, Aichi I, Matsioka M. 2006. A protocol for Agrobacterium-mediated transformation in rice. Nature. 6(1):2796-2802.
NiwaY, Hirano T, Yoshimoto K, Shimizu M, Kobayashi H. 1999. Non-invasive quantitative detection and application of non-toxic, S65T-type green fluorescent protein in living plants. The Plant J. 18:455-463.
Ohta S, Mita S, Hattori T, Nakamura K. 1990. Construction and expression in
tobacco of a β-glucuronidase (GUS) reporter gene containing an intron within the coding sequence. Plant and Cell Physiol. 31:805-813.
Offringa R, de Groot MJ, Haagsman HJ, Does MP, Van den Elzen, Hooykaas PJJ. 1990. Extrachromosomal homologous recombination and gene targetting in plant cells after Agrobacterium mediated transformation. EMBO J 9: 3077-3084.
Pang SZ, DeBoer DL, Wan Y, Ye G, Layton JG, Neher MK, Amstrong CL, Fry EJ, Hinchee MAW, Fromm ME. 1996. An improved green fluorescent protein gene as a vital marker in plants. Plant Physiol. 112:893-900.
Rachmawati S. 2006. Studi perkembangan perbaikan sifat genetik padi menggunakan transformasi Agrobacterium. AgroBiogen. 2(1):36-44. Rossi L, Hohn B, Tinland B. 1996. Integration of complete T-DNA units is
dependent on the activity of virE protein of Agrobacterium tumefaciens.
Proc. Natl. Acad. Sci. USA 93: 126-130.
Sahoo KK, Tripathi AK, Pareek A, Sopory SK, Pareek-Singla SL. 2011. An improved protocol for efficient transformation and regeneration of diverse indica rice cultivars. Plant Methods. 49(7):1-11.
Sambrook J, Fritsch EF, Maniatis T. 2001. Molecular Cloning: A laboratory Manual. Ed ke-3. New York (US): Cold Spring Harbor.
Tanaka A, Mita S, Ohta S, Kyozuka J, Shimamoto K, Nakamura K. 1990. Enhancement of foreign gene expression by a dicot intron in rice but not in tobacco is correlated with an increased level of mRNA and an efficient splicing of the intron. NucAcids Res. 23(18):6767-6781.
Tang X, Nakata Y, Li HO, Zhang M, Gao H, Fujita A, Sakatsume O, Ohta T, Yokoyama K. 1994. The optimization of preparations of competent cells for transformation of Escherichia coli. Nucleic Acids Res. 22(14): 2857-2858. Toki S, Hara N, Ono K, Onodera, Tagiri, Oka S, Tanaka H. 2006. Early infection
of scutellum tissue with Agrobacterium allows high-speed transformation of rice. Plant J. 47:969-976.
20
Vancanneyt G, Schmidt R, Sanchez AO, Willmitzer L, Sosa MR. 1990. Construction an intron-containing marker gene: Splicing of the intron in transgenic plants and its use in monitoring early events in Agrobacterium -mediated plant transformation. Mol Gen Genet. 220:245-250.
Wang MB, Brettell RIS, Upadhyaya NM, Waterhouse PM. 1997. Intron-mediated improvement of a selectable marker gene for plant transformation using
Agrobacterium tumefaciens. J. Genet. Breed. 51:325-334.
Weigel D, Glazebrook J. 2002. Arabidopsis Laboratory Manual. New York (US): Cold Spring Harbor.
21
RIWAYAT HIDUP
Penulis dilahirkan di Beringin (Kabupaten Simalungun, Sumatera Utara), pada tanggal 17 Desember 1989 dan merupakan puteri pertama dari pasangan Antonius Sumanto dan Prudentina Widyastuti. Penulis memperoleh gelar Sarjana Sains (S.Si) dari Departemen Biologi, Fakultas Matematika dan Ilmu Pengetahuan Alam, Universitas Sumatera Utara pada tahun 2011 dan pada tahun 2013 penulis melanjutkan pendidikan Magister Sains di Program Studi Bioteknologi, Sekolah Pascasarjana Institut Pertanian Bogor.