KONSTRUKSI PROMOTOR GEN
LEAFY
KAKAO PADA
VEKTOR EKSPRESI DENGAN SISTEM
GATEWAY
DAN TRANSFORMASI KE
Agrobacterium
sp.
ENDAH RATNA PURI
DEPARTEMEN BIOKIMIA
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
ABSTRAK
ENDAH RATNA PURI. Konstruksi Promotor Gen
LEAFY
Kakao
pada Vektor
Ekspresi dengan Sistem
Gateway
dan Transformasi ke
Agrobacterium
sp.
Dibimbing oleh LAKSMI AMBARSARI dan TETTY CHAIDAMSARI.
Kakao (
Theobroma cacao
L.) merupakan salah satu tanaman perkebunan
yang memiliki prospek cerah karena diperkirakan kebutuhan kakao dunia akan
terus meningkat. Peningkatan permintaan kakao tersebut ternyata disertai
permasalahan besar dalam pembudidayaannya, yaitu layu pentil (
cherelle wilt
)
yang merupakan penyakit fisiologis karena ketidaknormalan gen yang
bertanggung jawab pada proses pembungaan. Salah satu gen yang berperan utama
dalam pembentukan bunga adalah gen
LEAFY
dan promotor gen
LEAFY
diharapkan mengaktifkan kerja gen
LEAFY
. Penelitian ini bertujuan menyisipkan
tiga ukuran promotor gen
LEAFY
(1041 pb, 702 pb, dan 349 pb) ke dalam vektor
ekspresi dengan sistem
gateway
untuk mengetahui ukuran promotor yang efektif
mengekspresikan gen. Promotor gen
LEAFY
yang berukuran 1041 pb, 702 pb, dan
349 pb terlebih dahulu disisipkan ke dalam vektor donor (pDONR
TM221) yang
selanjutnya ke dalam vektor destinasi khusus promotor. Pengujian plasmid
rekombinan setelah transformasi ke dalam
Agrobacterium tumefaciens
galur
AGL-O menghasilkan pita berukuran sekitar 8002 pb, 7663 pb, dan 7310 pb.
Simpulan dari penelitian ini tiga ukuran promotor gen
LEAFY
, yaitu 349 pb, 702
pb, dan 1041 pb telah berhasil disisipkan pada vektor ekspresi dengan sistem
ABSTRACT
ENDAH RATNA PURI
.
Construction of Cacao
LEAFY
Gene Promoter in
Expression Vector with
Gateway
System and Transformation to
Agrobacterium
sp. Under the direction of LAKSMI AMBARSARI and TETTY
CHAIDAMSARI.
Cacao (
Theobroma cacao L.
) is one of the plantations with a good prospect
because the world demand of cocoa is expected to be growing. The increased
demand of cacao is contributed with a major problems related to the cultivation is
cherelle wilt. Cherelle wilt in cocoa is a physiological disease that cause by the
abnormalities of the genes who is responsible to the flowering process. One of the
genes that play a major role in the flower formation is the
LEAFY
gene and
LEAFY
gene promoter is expected to activate
LEAFY
genes. The objective of this
research is to construct three size of
LEAFY
gene promoter (1041 bp, 702 bp, and
349 bp) into the expression vector with
gateway
system by knowing the size of an
effective related to the gene expressed.
LEAFY
gene promoter in size 1041 bp,
702 bp, and 349 bp to be inserted into a donor vector (pDONR
TM221) and then
into the specific destination vector for promoter. The recombinant plasmid has
been analyzed after transformation into the
Agrobacterium tumefaciens
strain
AGL-O give a result in a DNA band about 8002 bp, 7663 bp, and 7310 bp. The
conclusion of this research by the three measures of
LEAFY
gene promoter, which
is 1041 bp, 702 bp, and 349 bp have been successfully inserted into the expression
vector with the
gateway
system and transformed into
Agrobacterium tumefaciens
KONSTRUKSI PROMOTOR GEN
LEAFY
KAKAO PADA
VEKTOR EKSPRESI DENGAN SISTEM
GATEWAY
DAN TRANSFORMASI KE
Agrobacterium
sp.
ENDAH RATNA PURI
Skripsi
sebagai salah satu syarat memperoleh gelar
Sarjana Sains pada
Departemen Biokimia
DEPARTEMEN BIOKIMIA
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
Judul : Konstruksi Promotor Gen
LEAFY
Kakao pada Vektor Ekspresi
dengan Sistem
Gateway
dan Transformasi ke
Agrobacterium
sp.
Nama : Endah Ratna Puri
NIM
: G84070026
Disetujui
Komisi Pembimbing
Diketahui
Tanggal Lulus:
Dr. Ir. I Made Artika, M.App.Sc
Ketua Departemen Biokimia
Dr. Laksmi Ambarsari, M.S
Ketua
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah SWT atas segala
kerunia-Nya, shalawat serta salam semoga selalu tercurah kepada Nabi Muhammad SAW,
keluarga, sahabat, dan para pengikutnya sampai akhir zaman sehingga penulis
dapat menyelesaikan penelitian ini. Penelitian ini berjudul Konstruksi Promotor
Gen
LEAFY
Kakao pada Vektor Ekspresi dengan Sistem
Gateway
dan
Transformasi ke
Agrobacterium
sp. Kegiatan penelitian ini dilakukan dari bulan
Desember hingga April 2011, bertempat di Laboratorium Balai Penelitian
Bioteknologi Perkebunan Indonesia, jalan Taman Kencana, Bogor.
Penulis mengucapkan terima kasih kepada semua pihak yang telah
membantu dalam penyelesaian penelitian ini, antara lain kepada Dr. Laksmi
Ambarsari, MS. selaku pembimbing utama dan Dr. Tetty Chaidamsari, M.Si
selaku pembimbing lapangan yang telah memberikan saran, kritik, dan
bimbingannya serta Mba Nina, Mba Herti, serta segenap staf di Laboratorium
Biologi Molekuler dan Rekayasa Genetika, Balai Penelitian Bioteknologi
Perkebunan Indonesia atas peran dan kerjasamanya yang telah banyak membantu
dalam menyelesaikan penelitian ini.
Ucapan terima kasih juga penulis sampaikan kepada orang tua, kakak, adik,
Andes Dinova, dan Biokimia 44 untuk semua doa, dukungan, dan bimbingan yang
sangat berarti bagi penulis. Serta kepada rekan selama penelitian Yoshita, Riska,
Ismi, Ibrahim, Dewi, Mas Rendi, Rika, dan M. Iqbal Akbar Muttaqin atas saran
dan motivasi yang diberikan. Penulis berharap semoga karya ilmiah ini dapat
memberikan manfaat bagi semua orang yang memerlukannya.
Bogor, April 2011
RIWAYAT HIDUP
Penulis dilahirkan di Bandung pada tanggal 18 Mei 1989 dari ayah Endang
Achyar dan ibu Wiwin Wintarsih. Penulis merupakan anak ketiga dari empat
bersaudara.
Pendidikan penulis dimulai dari TK Bayangkari 18 dan SDN Garuda 5
Bandung, kemudian melanjutkan pendidikan ke SMPN 1 Bandung. Tahun 2007
penulis lulus dari SMAN 7 Bandung dan pada tahun yang sama lulus seleksi
masuk IPB melalui jalur Undangan Seleksi Masuk IPB (USMI). Penulis memilih
mayor Departemen Biokimia, Fakultas Matematika dan Ilmu Pengetahuan Alam.
Selama mengikuti perkuliahan penulis aktif dalam kegiatan organisasi
kemahasiswaan, diantaranya penulis aktif di Himpunan Profesi Biokimia
(CREBs), Himpunan Profesi Komunikasi dan Pengembangan Masyarakat
(HIMASIERA),
AGRIFARMA,
dan
Paguyuban
Mahasiswa
Bandung
(PAMAUNG). Penulis juga aktif sebagai penyiar radio komunitas IPB (AGRI
FM) dan
master ceremony
beberapa acara besar di IPB. Selain itu, penulis juga
aktif mengikuti beberapa acara kepanitiaan seperti masa pengenalan departemen
Biokimia 2008/2009 dan 2009/2010, IPB
Art Contest,
Lomba Karya Ilmiah
Populer, dan beberapa seminar di IPB. Penulis melakukan Praktik Lapang di Balai
Penelitian Bioteknologi Perkebunan Indonesia, Taman Kencana, Bogor. Penulis
juga aktif mengikuti kegiatan kompetisi karya tulis dan kompetisi wirausaha.
Prestrasi yang pernah diraih, yaitu menjadi juara 3 KKTM tingkat nasional Index
2010, juara 2 LKTI Al-Quran 2010, Finalis Wirauasaha Muda Mandiri, Finalis
DAFTAR ISI
Halaman
DAFTAR GAMBAR ... vi
DAFTAR LAMPIRAN ... vii
PENDAHULUAN ………
...
1
TINJAUAN PUSTAKA
Kakao (
Theobroma cacao
L.) ...
1
Promotor Gen ...
3
Pengklonan dengan Sistem
Gateway
...
3
Polymerase Chain Reaction
(PCR) ...
4
Agrobacterium tumefaciens ...
5
BAHAN DAN METODE
Bahan dan Alat ...
6
Metode...
6
HASIL DAN PEMBAHASAN
Hasil Amplifikasi Promotor Gen
LEAFY
dengan Tiga Primer
Spesifik
Gateway
... 9
Hasil Ekstraksi dan Pemurnian Promotor Gen
LEAFY
dari Gel Agarosa ... 10
Hasil Reaksi Rekombinasi BP dan LR Promotor Gen
LEAFY
dengan Sistem
Gateway
... 10
Hasil Konfirmasi Koloni Transforman setelah Reaksi BP
dengan Teknik PCR Koloni ... 11
Hasil Konfirmasi Koloni Transforman setelah Reaksi BP
dengan Isolasi DNA Plasmid Rekombinan (Fermentas) ... 12
Hasil Konfirmasi Koloni Transforman setelah Reaksi LR
dengan Teknik PCR Koloni ... 13
Hasil Konfirmasi Koloni Transforman setelah Reaksi LR
dengan Isolasi DNA Plasmid Rekombinan (Fermentas) ... 14
Hasil Transformasi Klon Ekspresi ke dalam
Agrobacterium tumefaciens
Galur AGL-O ... 15
SIMPULAN DAN SARAN
Simpulan ... 16
Saran ... 16
DAFTAR PUSTAKA ... 16
DAFTAR GAMBAR
Halaman
1 Pohon kakao ... 2
2 Promotor gen prokariot ... 3
3 Promotor gen eukariot ... 3
4 Bagian struktur promotor eukariot ... 3
5 Proses
Polymerase Chain Reaction
(PCR) ... 5
6 Koloni
Agrobacterium tumefaciens
... 6
7 Elektroforegram amplikon promotor gen
LEAFY...
9
8 Elektroforegram ekstraksi dan pemurnian promotor gen
LEAFY
... 10
9 Koloni setelah reaksi BP ... 11
10 Koloni setelah reaksi LR ... 11
11 Elektroforegram PCR koloni promotor gen
LEAFY
349 pb (pDONR) ... 12
12 Elektroforegram PCR koloni promotor gen
LEAFY
702 pb (pDONR) ... 12
13 Elektroforegram PCR koloni promotor gen
LEAFY
1041 pb (pDONR) ... 12
14 Elektroforegram hasil isolasi plasmid setelah reaksi BP ... 13
15 Elektroforegram PCR koloni promotor gen
LEAFY
349 pb (pDEST) ... 14
16 Elektroforegram PCR koloni promotor gen
LEAFY
702 pb (pDEST) ... 14
17 Elektroforegram PCR koloni promotor gen
LEAFY
1041 pb (pDEST) ... 14
18 Elektroforegram hasil isolasi plasmid setelah reaksi LR ... 15
19 Koloni
Agrobacterium tumefaciens
setelah reaksi LR... 15
DAFTAR LAMPIRAN
Halaman
1 Tahapan konstruksi promotor gen
LEAFY
ke vektor pengklonan ... 19
2 Tahapan konstruksi promotor gen
LEAFY
ke vektor destinasi ... 20
3 Peta marka seleksi pDONR
TM221 ... 21
4 Reaksi BP
Gateway
... 22
5 Reaksi LR
Gateway
... 23
PENDAHULUAN
Kakao (Theobroma cacao L.) merupakan salah satu tanaman perkebunan yang mempunyai nilai ekonomi tinggi sebagai komoditas ekspor. Kakao sebagai komoditas andalan perkebunan berperan penting bagi perekonomian nasional, khususnya sebagai penyedia lapangan kerja, sumber pendapatan, dan devisa negara (Depperin 2007). Indonesia termasuk ke dalam tiga besar produsen kakao dunia bersama Ghana dan Pantai Gading (International Cocoa Organization 2003). Sumbangan devisa terbesar ketiga pada sub sektor perkebunan setelah karet dan minyak sawit yaitu dari kakao sebesar US$ 546 juta. Peranan kakao yang penting di Indonesia menjadi alasan pemerintah untuk menetapkan kakao sebagai salah satu komoditas dalam revitalisasi perkebunan (Deptan 2007).
Tanaman kakao memiliki prospek yang cerah karena diperkirakan kebutuhan kakao dunia akan terus meningkat. Peningkatan permintaan kakao tersebut disertai beberapa masalah besar dalam pembudidayaannya. Masalah besar yang dapat menurunkan produksi kakao tersebut di antaranya serangan penggerek buah kakao (PBK) dan rendahnya persen bunga menjadi buah yang disebabkan oleh layu pentil (cherelle wilt) (Deptan 2007).
Masalah layu pentil dan sedikitnya jumlah bunga yang akan menjadi buah terjadi karena ketidaknormalan gen yang bertanggung jawab pada proses pembungaan. Layu pentil pada kakao merupakan penyakit fisiologis yang diduga karena beberapa faktor, salah satunya yaitu persaingan untuk memperoleh nutrisi yang ketersediaannya terbatas antara buah muda, buah tua, dan berbagai tunas muda. Layu pentil pada kakao juga diduga karena adanya persaingan untuk memperoleh asimilat, terutama karbohidrat. Persaingan ini terjadi antara buah dengan buah dan antara buah dengan pertumbuhan pucuk yang aktif (Alvim 1974). Buah menjadi layu karena rendahnya kandungan fitohormon di dalam biji atau buah sehingga penyerapan asimilat juga menjadi rendah.
Usaha untuk meningkatkan produktivitas tanaman kakao mendorong studi molekuler dan rekayasa genetika dengan mempelajari mekanisme molekuler dari pembungaan kakao. Salah satu gen yang berperan utama dalam pembentukan bunga adalah gen
LEAFY. Promotor gen LEAFY diharapkan
dapat mengaktifkan kerja gen LEAFY
sehingga mencegah penyakit layu pentil, mengatur pembungaan, dan akhirnya meningkatkan produktivitas kakao. Pada penelitian sebelumnya telah berhasil diisolasi promotor gen LEAFY dari daun kakao yang berukuran 1650 pb dengan teknik Genome WalkingTM (Santoso 2010). Hasil menunjukkan bahwa bagian dari promotor gen LEAFY yang efektif untuk mengekspresikan gen masih harus diteliti melalui konstruksi beberapa ukuran promotor gen LEAFY. Pada penelitian ini dilakukan konstruksi promotor gen LEAFY
dengan tiga ukuran berbeda, yaitu 1041 pb, 702 pb, dan 349 pb. Ketiga ukuran tersebut diperoleh dari hasil amplifikasi dengan primer gateway yang spesifik.
Penelitian ini bertujuan menyisipkan tiga ukuran promotor gen LEAFY (1041 pb, 702 pb, dan 349 pb) ke dalam vektor ekspresi dengan sistem gateway untuk mengetahui ukuran promotor yang efektif mengekspresikan gen dan ukuran yang efisien untuk menggabungkannya dengan gen LEAFY atau gen lain. Hipotesis penelitian ini adalah tiga ukuran berbeda dari promotor gen LEAFY, yaitu 1041 pb, 702 pb, dan 349 pb dapat dikonstruksi pada vektor ekspresi dengan sistem gateway. Konstruksi pada beberapa ukuran ini dilakukan karena tidak semua bagian dari
promotor gen LEAFY dapat
mengekspresikan gen dan mengaktifkan gen
LEAFY.
Keberhasilan konstruksi promotor gen
LEAFY pada vektor ekspresi merupakan fokus penelitian ini yang diharapkan dapat digunakan untuk mempelajari sifat, fungsi, dan ukuran efektif promotor gen yang terekspresi. Penyisipan tiga ukuran promotor gen LEAFY yangberbeda (1041 pb, 702 pb, dan 349 pb) pada vektor ekspresi dengan sistem gateway selanjutnya akan ditransformasikan ke dalam tanaman melalui penyisipan ke Agrobacterium tumefaciens
galur AGL-O.
TINJAUAN PUSTAKA
Kakao (Theobroma cacao L.)
air laut. Klasifikasi lengkap tanaman kakao termasuk ke dalam divisi Spermatophyta, subdivisi Angiospermae, kelas Dicotyledone, ordo Malvales, famili Sterculiaceae, genus Theobroma, dan spesies Theobroma cacao L (Tjitrosoepomo 1988). Tanaman kakao bukan tanaman asli Indonesia, melainkan tanaman yang berasal dari lembah hulu sungai Amazon, Amerika Selatan yang dibawa masuk ke Indonesia melalui Sulawesi Utara oleh bangsa Spanyol sekitar tahun 1560. Tanaman kakao ditanam hampir di seluruh pelosok tanah air dengan sentra produksi utama di Sulawesi Selatan, Sulawesi Tenggara, Sulawesi Tengah, Sumatera Utara, Nusa Tenggara Timur, Jawa Timur, Kalimantan Timur, Maluku Utara, dan Irian Jaya (Deptan 2007). Keberhasilan perluasan areal dan peningkatan produksi tersebut telah memberikan hasil nyata bagi peningkatan komoditas kakao Indonesia di dunia. Indonesia berhasil menempatkan diri sebagai produsen kakao terbesar kedua
dunia setelah Pantai Gading (Cote d’Ivoire)
pada tahun 2002, walaupun kembali tergeser ke posisi ketiga oleh Ghana pada tahun 2003 (International Cocoa Organization 2003).
Kakao bagi Indonesia merupakan salah satu komoditas andalan perkebunan yang peranannya cukup penting bagi perekonomian nasional, khususnya sebagai penyedia lapangan kerja, sumber pendapatan, dan devisa negara (Depperin 2007). Kakao juga berperan dalam mendorong pengembangan wilayah dan pengembangan agroindustri. Pada tahun 2004, perkebunan kakao telah menyediakan lapangan kerja dan sumber pendapatan bagi sekitar 1.1 juta kepala keluarga petani yang sebagian besar berada di Kawasan Timur Indonesia (KTI). Perkebunan kakao juga memberikan sumbangan devisa terbesar ketiga pada sub sektor perkebunan setelah karet dan minyak sawit dengan nilai sebesar US$ 546 juta. Atas dasar pentingnya peran kakao di Indonesia, maka pemerintah menetapkan kakao sebagai salah satu komoditas dalam revitalisasi perkebunan (Deptan 2007).
Tanaman kakao di Indonesia memiliki prospek yang cerah karena diperkirakan kebutuhan kakao dunia akan terus meningkat. Peningkatan permintaan kakao tersebut disertai permasalahan besar dalam pembudidayaannya. Masalah besar yang dapat menurunkan produksi kakao adalah serangan penggerek buah kakao (PBK) dan
pembungaan yang tidak konsisten dengan layu pentil (Deptan 2007). Salah satu aspek fisiologis yang penting dalam peningkatan produksi buah kakao adalah tingginya tingkat layu pentil, terutama yang terjadi saat awal pembentukan buah.
Layu pentil merupakan penyakit fisiologis yang disebabkan oleh persaingan nutrisi antara pentil dengan organ lain yang sedang tumbuh aktif yang mengakibatkan kegagalan proses embriogenesis dan perkembangan buah. Hasil penelitian Tjasadihardja (1987), menunjukkan bahwa pertunasan sangat erat hubungannya dengan tingkat layu pentil. Tunas baru yang terbentuk merupakan pesaing yang sangat kuat bagi buah muda dalam menggunakan asimilat.
Kakao merupakan tumbuhan tahunan (perennial) berbentuk pohon, di alam dapat mencapai ketinggian 10 m. Walaupun dalam pembudidayaan tingginya dibuat tidak lebih dari 5 m tetapi dengan tajuk menyamping yang meluas. Hal ini dilakukan untuk memperbanyak cabang produktif (Putri 2004). Pohon kakao menghasilkan buah berbentuk bulat memanjang yang tumbuh dari bunga yang diserbuki. Buah terdiri dari 5 daun buah dan memiliki ruang yang di dalamnya terdapat biji (Gambar 1).
Gambar 1 Pohon kakao.
yang terbentuk dan keragaman bunganya lebih tinggi dari pada tanaman yang lebih muda. Kontrol lingkungan (lamanya hari dan suhu) mendorong transisi dari sel yang sedang aktif membelah menjadi sel bunga. Pembentukan bunga dimulai dengan mekanisme yang kompleks dari pembentukan kelopak bunga, daun bunga, benang sari, dan putik. Gen penanda meristem mengubah sel yang sedang aktif membelah dengan cepat menjadi sel bunga (Brian 2003).
PromotorGen
Gen prokariot secara umum tersusun atas promotor, bagian struktural, dan terminator. Promotor gen adalah urutan DNA spesifik yang berperan mengendalikan transkripsi gen struktural dan terletak di sebelah hulu (upsteam) dari bagian struktural suatu gen. Bagian promotor menjadi tempat awal pelekatan enzim RNA polimerase yang melakukan transkripsi bagian struktural (Yuwono 2005). Bagian penting promotor pada prokariot disebut sebagai Pribnow Box
pada urutan nukleotida -10 pb dan -35 pb, biasanya berupa TATA-Box. TATA-Box merupakan bagian DNA yang banyak mengandung basa timin dan adenin (Gambar 2) (Murray et al. 2003).
Gambar 2 Promotor gen prokariot.
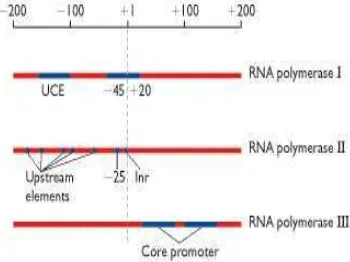
Promotor pada eukariot lebih kompleks dibandingkan dengan promotor pada prokariot. Pada eukariot, promotor digunakan untuk menguraikan semua urutan penting di dalam inisiasi transkripsi suatu gen. Berbagai gen yang ada dalam genom inti ditranskripsi oleh salah satu dari tiga jenis enzim RNA polimerase. RNA polimerase I dan III bertanggung jawab untuk mensintesis rRNA dan tRNA, sedangkan RNA polimerase II bertanggung jawab untuk mentranskripsi gen yang mengkode suatu protein tertentu (Gambar 3) (Brown 2002).
Gambar 3 Promotor gen eukariot.
Promotor gen eukariot tertentu, ada yang memiliki beberapa exon, yaitu bagian yang akan mengkode protein atau biasa disebut
open reading frame (ORF) dan ada yang tidak ditranslasi menjadi protein tetapi berperan dalam proses transkripsi (untranslated region). Hasil transkripsi seringkali juga membawa sekuen yang disebut sebagai intron, yang terdapat pada bagian ORF atau pada bagian untranslated region. Secara bertahap intron akan dibuang dari hasil transkripsi yang terbentuk dan exon yang ada akan digabungkan membentuk mRNA yang siap untuk ditranslasi menjadi protein (Gambar 4).
Gambar 4 Bagian struktur promotor eukariot.
Pengklonan dengan Sistem Gateway
Sistem pengklonan dengan sistem
Gateway pada dasarnya bergantung pada dua reaksi yaitu, rekombinasi BP dan LR. Sistem pengklonan Gateway Site-Specific Recombination (SSR) dikomersialisasikan oleh Invitrogen. Metode ini mengkonstruksi gen sisipan yang terlebih dahulu diklon ke dalam vektor donor (pDONR) dan kemudian ditransfer ke dalam vektor destinasi (pDEST) dengan rekombinasi situs spesifik. Teknologi ini merupakan penerapan reaksi rekombinasi situs spesifik yang dapat balik (reversible). Penamaan reaksi BP dan LR didasarkan pada singkatan B dari bakteri dan P (phage) sedangkan L (left) dan R (right).
pb dari faga lamda dan situs attB yang berukuran 25 pb dari rekombinasi
Escherichia coli dan genom faga lamda yang disambungkan ke genom E. coli. Hasilnya genom faga diikat oleh attL yang berukuran 100 pb dan attR yang berukuran 168 pb (Lampiran 4). Kebalikannya faga lamda dipotong dari genom E. coli dengan rekombinasi di antara situs attL dan attR dalam reaksi LR. Reaksi ini merekombinasikan gen sisipan yang diikat oleh situs attL dengan vektor destinasi yang membawa situs attR. Hasilnya gen sisipan diikat oleh dua situs yakni situs attB1 dan attB2 yang selanjutnya disebut klon ekspresi (Lampiran 5) (Hartley et al. 2000).
Reaksi BP dikatalisis oleh enzim BP klonase yang terdiri atas integrase faga(Int) dan IHF (Integration Host Factor). Campuran BP klonase memindahkan DNA sisipan ke dalam vektor donor yang menghasilkan suatu klon entri (pENTR) dan diikat oleh dua situs attL. Entri klon merupakan substrat kunci dalam reaksi LR yang dikatalisis oleh campuran LR klonase yang terdiri atas integrase faga (Int), IHF (Integration Host Factor), dan eksisionase (Xis). Campuran LR klonase mentransfer DNA sisipan yang diikat oleh situs attL ke dalam vektor destinasi (pDEST) yang membawa situs attR (Karimi et al. 2007). Setelah pencocokan rekombinasi situs attL dan attR, DNA sisipan dimasukkan ke dalam klon ekspresi (pEXPR) dan diikat kembali oleh situs attB1 dan attB2. Vektor donor (pDONR), klon entri (pENTR), vektor destinasi (pDEST), dan klon ekspresi (pEXPR) merupakan bagian yang diadopsi oleh pengguna gateway untuk memasukkan dan mengeluarkan plasmid di dalam reaksi klonase (Karimi et al. 2007).
Hasil rekombinasi dengan sistem
gateway dapat dengan mudah diseleksi dengan seleksi positif (resisten terhadap antibiotik) dan seleksi negatif (gen sitotoksik ccdB) yang akan menghambat pertumbuhan
E. coli yang telah disisipi salah satu vektor (Magnani et al. 2006). Gen sitotoksik ccdB terdapat pada vektor donor dan vektor destinasi. Pada saat reaksi rekombinasi BP, ccdB akan tertukar dengan gen sisipan sehingga vektor yang tidak tersisipi gen akan mati karena adanya ccdB. Pada reaksi rekombinasi LR, gen sisipan yang telah diikat oleh situs attL akan tertukar dengan gen ccdB yang telah diikat oleh situs attR pada vektor destinasi (Bernard & Couturier 1992).
Sistem gateway memiliki banyak keuntungan diantaranya kloning berlangsung cepat dengan efisiensi yang tinggi dalam mentransfer sekuen DNA ke dalam berbagai sistem vektor untuk ekspresi protein dan analisis protein. Multisite gateway memungkinkan penggunaan dan ekspresi berbagai tipe DNA sekuen (misalnya hasil PCR, klon cDNA, dan fragmen restriksi) (Lampiran 6). Keuntungan lain sistem
gateway, yaitu memudahkan akomodasi transfer sejumlah besar sekuen DNA ke dalam berbagai vektor destinasi (Invitrogen 2008). Modifikasi pengklonan gateway juga sering dilakukan untuk tujuan memperkirakan Open Reading Frame
(ORF) yang menyandi protein dalam vektor entri untuk mencegah keberadaan atau penambahan muatan sekuen. Penambahan muatan sekuen yang sering kali tinggi setelah rekombinasi dapat mempengaruhi fungsi protein (Dubin et al. 2008).
Polymerase Chain Reaction (PCR)
Polymerase Chain Reaction (PCR) adalah suatu metode enzimatis untuk melipatgandakan secara eksponensial suatu sekuen nukleotida tertentu dengan cara in
vitro. Metode ini pertama kali
dikembangkan pada tahun 1985 oleh Kary B. Mullis dan sekarang telah banyak digunakan untuk berbagai macam manipulasi dan analisis genetik (Yuwono 2006). Metode yang dikembangkan oleh Kary B. Mullis ini akhirnya mendapatkan hadiah nobel di bidang kimia pada tahun 1993. Metode PCR sangat sensitif, sensitivitas tersebut dapat digunakan untuk melipatgandakan satu molekul DNA (Jonas 2003). Instrumen PCR digunakan untuk menggandakan jumlah molekul DNA pada target tertentu dengan mensintesis molekul DNA baru yang berkomplemen dengan molekul DNA target dengan bantuan enzim dan oligonukleotida sebagai primer dalam suatu thermocycler. Primer yang berada sebelum daerah target disebut sebagai primer forward dan primer yang berada setelah daerah target disebut primer reverse. Enzim yang digunakan sebagai pencetak rangkaian molekul DNA baru dikenal sebagai enzim polimerase (Muladno 2002).
untuk mengawali sintesis rantai DNA, (3) deoksiribonukleotida trifosfat (dNTP), terdiri atas dATP, dCTP, dGTP, dTTP, dan (4) enzim DNA polimerase, yaitu enzim yang melakukan katalis reaksi sintesis rantai DNA. Komponen lain yang juga penting adalah senyawa bufer (Yuwono 2006). Bufer berfungsi menjaga kestabilan pH selama proses PCR berlangsung, menjaga kekuatan ionik, dan suhu penempelan primer (annealing) pada DNA target. Bufer yang umum digunakan terdiri dari KCL dengan konsentrasi 50 mM dan Tris-HCL 10 mM. Bufer memiliki pH 8.3 pada suhu 25oC (Kolmodin & Williams 1997).
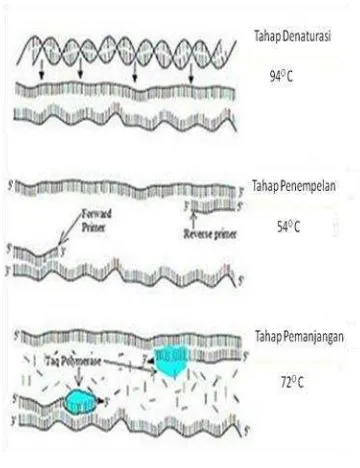
Reaksi PCR pada dasarnya adalah tiruan dari proses replikasi DNA, yaitu adanya pembukaan rantai DNA utas ganda, penempelan primer, dan pemanjangan rantai DNA baru oleh DNA polimerase dari arah
5’ ke 3’. Satu siklus pada teknik PCR terdiri
atas tiga tahap (Gambar 5), yaitu denaturasi,
annealing, dan ekstensi. Denaturasi dilakukan pada suhu 90o-95oC sehingga terjadi pemisahan utas ganda DNA menjadi dua utas tunggal DNA yang menjadi cetakan (template) tempat penempelan primer dan tempat kerja DNA polimerase. Selanjutnya tahap annealing, suhu diturunkan untuk penempelan primer oligonukleotida pada sekuens yang komplementer pada molekul DNA cetakan. Suhu annealing tiap sekuens DNA bersifat spesifik dan merupakan faktor penentu keberhasilan suatu reaksi PCR, sedangkan tahap terakhir adalah ekstensi. Tahap ekstensi dilakukan pada suhu 72oC. Suhu ini merupakan suhu optimum untuk kerja enzim Taq DNA polimerase (Yuwono 2006). Pada tahap ini terjadi sintesis DNA komplemen dengan DNA cetakan.
Ketiga tahapan tersebut dilakukan berulang kali dalam mesin PCR, pada umumnya antara 25-30 kali (siklus) bergantung dari jumlah DNA yang diinginkan sehingga pada akhir siklus akan diperoleh molekul-molekul DNA rantai ganda yang baru hasil polimerisasi dalam jumlah yang jauh lebih banyak dibandingkan dengan jumlah DNA cetakan yang digunakan (Yuwono 2006). Banyaknya siklus amplifikasi tergantung pada konsentrasi DNA target di dalam campuran reaksi. Sedikitnya diperlukan 25 siklus untuk melipatgandakan satu kopi sekuen DNA target di dalam genom mamalia agar hasilnya dapat dilihat secara langsung, misalnya dengan elektroforesis gel agarosa (Sambrook et al. 1989).
Gambar 5 Proses Polymerase Chain Reaction (PCR).
Agrobacterium tumefaciens
Bakteri Agrobacterium tumefaciens
merupakan bakteri tanah Gram negatif yang termasuk famili Rhizobiaceae dan anggota dari genus Agrobacterium. Agrobacterium
memiliki tiga komponen genetik yang digunakan untuk menginfeksi tanaman. Komponen yang pertama adalah komponen T-DNA yaitu fragmen yang ditransfer ke sel tanaman. T-DNA terletak di plasmid Ti (tumour-inducing) dari Agrobacterium. Komponen kedua adalah virulance (vir) region yang mensintesis protein vir dan komponen ketiga adalah gen chomosomal virulance (chv) yang berfungsi dalam pelekatan bakteri ke dalam sel tanaman (Tzafira & Citovsky 2002). Langkah awal dalam induksi tumor adalah ketika A. tumefaciens menempel pada permukaan sel tanaman yang disebut kolonisasi bakteri (Gambar 6) (Matthysse1986).
Gen asing dapat disisipkan ke dalam plasmid Ti dengan menggunakan teknik DNA rekombinan. Plasmid rekombinan ditransformasikan ke A. tumefaciens yang dapat digunakan untuk menginfeksi sel tumbuhan. Plasmid rekombinan akan menyelipkan dirinya ke dalam kromosom tumbuhan. Hal ini memungkinkan menghasilkan tumbuhan yang mengandung dan mengekspresikan gen asing yang dapat diwariskan ke keturunannya. Plasmid Ti ditemukan pada semua galur A. tumefaciens
Kemampuan T-DNA untuk mentransfer DNA ke organisme eukariot dan gen yang berada di T-DNA dapat digantikan dengan gen apa saja, menjadikan A. tumefaciens
sebagai vektor yang ideal untuk mentransfer gen ke dalam organisme eukariotik, seperti tanaman untuk menghasilkan suatu tanaman transgenik (Tzfira & Citovsky 2002).
Gambar 6 Koloni Agrobacterium tumefaciens.
BAHAN DAN METODE
Bahan dan Alat
Bahan-bahan yang digunakan pada proses amplifikasi dengan kit platinum dari Invitrogen, yaitu memerlukan bahan-bahan seperti promotor gen LEAFY, primer
forward spesifik (promLFY F349, promLFY F702, dan promLFY F1041), primer promTcLFY reverse, larutan bufer, MgCl2, dNTPs, Taq polimerase, dan molecular water. Bahan yang digunakan pada proses
rekombinasi dengan sistem
Gateway®Technology yakni BP dan LR Recombination Reaction Protocol (Kit Invitrogen). Proses BP rekombinasi memerlukan bahan-bahan seperti bufer TE 1x pH 8, insert berupa hasil elusi,
pDONR™vektor, dan 5X BP
Clonase™bufer reaksi. Bahan-bahan yang digunakan pada proses LR rekombinasi adalah bufer TE 1x pH 8, klon entri, vektor destinasi khusus promotor gen, dan 5X LR
Clonase™bufer reaksi.
Bahan-bahan yang digunakan dalam elektroforesis yaitu gel agarosa (Sigma), bufer tris-borate-EDTA (TBE) 0.5x, Etidium bromida (EtBr) 5 µg/100ml, loading buffer
(Bromfenol biru 2.5%, sukrosa 40%), dan marker DNA 1 kb plus ladder (Invitrogen). Tahapan transformasi ke dalam sel kompeten memerlukan bahan media Luria Bertani (LB) agar yang sudah ditambahkan kanamisin 50 ppm, 1 µL proteinase K, dan
sel kompeten Escherichia coli galur XL-1 Blue. Tahap transformasi ke dalam
Agrobacterium tumefaciens membutuhkan media Luria Bertani (LB) agar yang sudah ditambah kanamisin 50 ppm dan rimfapisin 50 ppm, media yeast extract pepton (YEP), dan sel kompeten A. tumefaciens galur AGLO. Tahap konfirmasi dengan PCR koloni setelah reaksi BP, bahan yang dibutuhkan adalah larutan bufer, MgCl2,
dNTPs, Taq polimerase, primer M13
Forward, primer M13 Reverse, dan
molecular water.
Alat yang digunakan untuk elektroforesis adalah sisir dan cetakan agar, bak elektroforesis, mikropipet, tabung mikro,
mircowave, adaptor 100 Volt, transluminator ultraviolet (UV) T2201 (Sigma), dan gel documenter. Selain itu alat yang digunakan adalah mesin PCR (ESCO Swift max), DNA
speed vacum 110 savant, inkubator bergoyang (shaker incubator), laminar air flow cabinet, segitiga penyebar, pinset, pisau skalpel, autoklaf, Eppendorf sentrifus 5417R, neraca analitik, dan peralatan-peralatan gelas seperti cawan petri, gelas piala, labu Erlenmeyer, labu takar, dan gelas ukur.
Metode
Amplifikasi promotor Gen LEAFY dengan Primer Spesifik Gateway
Amplifikasi promotor gen LEAFY
menggunakan kit dari Invitrogen (Platinum® Taq DNA Polymerase). Amplifikasi tiga ukuran promotor gen
LEAFY menggunakan tiga pasang primer spesifik gateway yang berbeda yaitu,
Gateway promLFY F1041 dan Gateway
promTcLFY R; Gateway promLFY F702 dan Gateway promTcLFY R; dan Gateway
promLFY F349 dan Gateway promTcLFY R. Proses amplifikasi dimulai dengan menyiapkan campuran reaksi yang terdiri atas 2.5 µL bufer, 1 µL MgCl2, 1 µL dNTPs,
0.2 µL Taq polimerase, dan 14.3 µL
molecular water. DNA cetakan (template) dimasukkan ke dalam tabung mikro sebanyak 1 µL. Reverse primer ditambahkan sebanyak 1 µL ke dalam tabung mikro kemudian ditambahkan pula sebanyak 1 µL
forward primer spesifik ke dalam masing-masing tabung.
penempelan primer (annealing) pada suhu 55oC selama 45 detik, pemanjangan primer (extension) pada suhu 72oC selama 2 menit, dan pascapemanjangan pada suhu 72oC selama 5 menit. Verifikasi produk PCR pada gel agarosa 1%.
Membuat Gel Agarosa Konsentrasi 1 % dan Elektroforesis Hasil Amplifikasi
Serbuk agarosa sebanyak 0.6 gram dilarutkan dalam 60 ml larutan TBE 0.5x. Larutan agarosa dipanaskan dalam
microwave sampai larut selama ± 120 detik. Larutan didiamkan sampai terasa hangat, kemudian ditambahkan larutan EtBr sebanyak 3 µL. Hal ini bertujuan agar EtBr tidak rusak karena panas. Larutan dituang ke dalam cetakan dan dibiarkan hingga mengeras membentuk gel. Gel agarosa yang telah mengeras digunakan untuk elektroforesis hasil amplifikasi.
Promotor gen yang telah diamplifikasi dengan metode PCR selanjutnya dielektroforesis untuk mengetahui hasil amplifikasi. Promotor gen hasil amplifikasi yang akan dielektroforesis dalam gel agarosa terlebih dahulu dicampurkan dengan loading buffer. Setelah dicampurkan dengan loading buffer kemudian campuran dimasukkan ke dalam sumur gel agarosa dan dielektroforesis dengan tegangan 75 volt selama ± 1 jam.
Ekstraksi dan Pemurnian Promotor Gen
LEAFY
Elektroforegram positif ditunjukkan dengan adanya pita pada gel agarosa setelah diamati dengan transluminator UV. Proses ekstraksi dan purifikasi promotor gen
LEAFY dari gel menggunakan kit dari Invitrogen. Pita yang terang pada gel agarosa dipotong di bawah transluminator UV. Potongan gel tersebut dimasukkan ke dalam tabung mikro dan ditambahkan 3x volume solubilization bufer (L3). Potongan gel diinkubasi pada suhu 50oC selama 10 menit sampai gel tersebut larut dan diinkubasi kembali pada suhu ruang selama 5 menit. Setelah potongan gel larut kemudian dimasukkan ke dalam kolom (Quick Gel Extraction Column) yang berada di atas tube pencuci (Wash Tube) dan disentrifus 1 menit pada 12000 rpm dengan suhu ruang. Sebanyak 500 µL bufer pencuci (Wash buffer) ditambahkan ke dalam kolom dan disentrifus 1 menit pada 12000 rpm, suhu ruang. Supernatan dibuang dan disentrifus dalam keadaan kosong pada
12000 rpm, 25oC selama 1 menit. Setelah itu, ditambahkan bufer elusi sebanyak 30 µL dan diinkubasi selama 2 menit pada suhu ruang. Larutan tersebut disentrifus kembali pada 12000 rpm, 25oC selama 2 menit. Hasil ekstraksi dan pemurnian gel dilihat dengan elektroforesis gel agarosa 1%.
Reaksi Rekombinasi BP dan LR Promotor Gen LEAFY dengan Sistem Gateway
Reaksi rekombinasi BP promotor gen
LEAFY dengan sistem gateway dimulai dengan menyiapkan bufer TE sebanyak 5 µL, 2 µL hasil elusi, dan vektor Donor (pDONRTM221) sebanyak 1 µL. Setelah itu, ditambahkan sebanyak 2 µL BP ClonaseTM II dan diinkubasi selama 2 jam pada suhu 250C. Setelah inkubasi selesai, ke dalam larutan ditambahkan sebanyak 2 µL Proteinase K dan diinkubasi kembali pada suhu 370C selama 15 menit. Hasil rekombinasi promotor gen LEAFY pada vektor Donor (pDONR) kemudian ditransformasikan ke dalam sel kompeten
E. coli untuk perbanyakkan plasmid yang telah membawa promotor gen LEAFY. Koloni bakteri E. coli yang tumbuh pada media LB agar yang mengandung antibiotik kanamisin dikonfirmasi dengan metode PCR koloni. Koloni bakteri rekombinan ditandai adanya pita setelah pengujian dengan metode PCR. Koloni bakteri rekombinan kemudian diisolasi DNA plasmidnya untuk direkombinasikan ke vektor destinasi (pDEST).
Reaksi rekombinasi LR, sebanyak 5 µL bufer TE, 2 µL hasil BP reaksi (klon entri), dan vektor destinasi (pDEST) sebanyak 1 µL. Setelah itu, ditambahkan sebanyak 2 µL LR ClonaseTM II dan diinkbuasi selama 2 jam pada suhu 250C. Setelah inkubasi selesai, ke dalam larutan ditambahkan sebanyak 2 µL Proteinase K dan diinkubasi lagi pada suhu 370C selama 15 menit. Hasil rekombianasi pada vektor destinasi kemudian ditransformasikan ke dalam E. coli. Koloni bakteri yang tumbuh diisolasi DNA plasmid dan selanjutnya dikonfirmasi dengan metode PCR Koloni. Plasmid rekombinan selanjutnya ditransformasikan ke A. tumefaciens galur AGL-O.
Transformasi Promotor Gen LEAFY ke Escherichia coli
perlahan hingga tercampur rata kemudian diinkubasi di dalam es selama 30 menit. Larutan hasil rekombinasi dan sel kompeten diberi kejut panas (heat shock) pada suhu 42oC selama 50 detik yang selanjutnya segera dimasukkan ke dalam es selama 10 menit. Ke dalam larutan hasil rekombinasi dan sel kompeten ditambahkan 800 µL Luria Bertani (LB) + glukosa 20 mM kemudian diinkubasi di dalam shaker incubator selama 1.5 jam pada suhu 37oC dengan kecepatan 150 rpm. Campuran tersebut diambil sebanyak 100 µL (1/10 volume) untuk ditumbuhkan dalam media LB agar yang terdiri atas tripton 10 g/l, yeast extract 5 g/l, NaCl 5 g/l, dan bakto agar 15 g/l. Sisanya sebanyak 9/10 volume disentrifus pada 3500 rpm, 25oC selama 5 menit. Supernatan dibuang ± 800 µL sedangkan sisa supernatan 100 µL dan pelet diresuspensi kemudian ditumbuhkan dalam media LB agar yang mengandung antibiotik kanamisin 50 ppm dan meratakannya dengan segitiga penyebar steril. Media diinkubasi dalam kondisi 37oC selama 16-20 jam kemudian dilihat koloni yang tumbuh.
Konfirmasi Koloni Transforman dengan Teknik PCR Koloni
Koloni yang tumbuh dipindahkan ke dalam media LB agar yang baru untuk membuat duplikat koloni. Setelah proses duplikasi, koloni yang tumbuh juga dipindahkan ke dalam tabung mikro yang berisi molecular water (MW) untuk persiapan PCR koloni. Pemindahan koloni dilakukan secara steril dalam laminar air flow cabinet. PCR koloni setelah reaksi rekombinasi BP dilakukan dengan menyiapkan komponen mix yang terdiri atas, 2.75 µL molecular water (MW), 1.5 µL
buffer complete, 0.3 µL dNTP’s, 0.15 µL
M13F, 0.15 µL M13R, dan 0.15 µL taq polymerase.
PCR koloni setelah reaksi rekombinasi LR dilakukan dengan menyiapkan komponen mix yang terdiri atas, 2.75 µL
molecular water (MW), 1.5 µL buffer complete, 0.3 µL dNTP’s, 0.15 µL masing -masing primer forward spesifik gateway
yang berbeda yaitu, Gateway promLFY F1041, Gateway promLFY F702, Gateway
promLFY F349, 0.15 µL masing-masing primer Gateway promTcLFY R, dan 0.15 µL taq polymerase. Tahap pertama PCR adalah program lisis 96oC selama 5 menit, 50oC selama 1 menit 30 detik, 96oC selama 1 menit 30 detik, 45oC selama 1 menit 30
detik, 96oC selama 1 menit, 40oC selama 1 menit. Program dihentikan sejenak untuk penambahan sebanyak 5 µL komponen mix
ke dalam masing-masing tabung. Kemudian program PCR dilanjutkan kembali dengan program PCR yakni, 94oC selama 30 detik, 45oC selama 1 menit, 72oC selama 2 menit. Setelah perbanyakan koloni transforman dengan teknik PCR selesai kemudian pengujian dengan elektroforesis gel agarosa 1 %.
Isolasi DNA Plasmid Rekombinan (Fermentas)
Isolasi DNA plasmid dilakukan berdasarkan hasil PCR koloni yang diketahui mengandung plasmid terinsersi fragmen yang diinginkan. Plasmid diisolasi dengan Fermentas GeneJet Plasmid Miniprep Kit. Sampel yang positif mengandung plasmid terinsersi disentrifus 12000 rpm terlebih dahulu. Pelet dihomogenisasi dengan penambahan 250 µL
resuspension solution. Setelah itu, ditambahkan 250 µL lisis solution, 350 µL
neutralization solution, dan setiap penambahan larutan dalam tabung dibolak-balik sebanyak 6x, kemudian disentrifus selama 5 menit, 12000 rpm pada suhu ruang. Pengikatan DNA dilakukan dengan memindahkan supernatan ke dalam kolom dan disentrifus selama 1 menit. Kolom dicuci dengan 500 µL wash solution dan disentrifus selama 1 menit (dilakukan sebanyak 2x). Kolom dalam keadaan kosong disentrifus kembali selama 1 menit. Kolom dipindahkan ke dalam tabung mikro baru dan ditambahkan 30 µL elution buffer tepat di tengah membran kolom. Setelah itu, diinkubasi selama 2 menit dan disentrifus kembali selama 2 menit. Hasil isolasi DNA plasmid diverifikasi dengan visualisasi elektroforesis gel agarosa 1%.
Transformasi ke dalam Agrobacterium
tumefaciens galur AGL-O
Sejumlah 10 µL DNA plasmid dimasukkan ke dalam A. tumefaciens stain AGL-O, didiamkan di dalam es selama 15 menit. Selanjutnya diinkubasi kembali di dalam N2 cair selama 5 menit dan pada suhu
37oC selama 5 menit. Sejumlah 1 ml YEP (Yeast Exctract Pepton) ditambahkan ke dalamnya dan dikocok selama 3 jam pada suhu 28oC. Larutan kemudian disentrifugasi pada kecepatan 6000 rpm selama 3 menit.
dipipet, dan dimasukkan ke dalam media LB yang telah diberi antibiotik kanamisin 50 ppm dan rimfapisin 50 ppm. Inkubasi dilakukan selama 2 hari pada suhu 28oC dan dalam kondisi gelap. Hasil yang diperoleh dikonfirmasi dengan PCR koloni dan isolasi plasmid. Setelah itu, promotor gen LEAFY
siap untuk dintransformasikan ke tanaman melalui A. tumefaciens.
HASIL DAN PEMBAHASAN
Hasil Amplifikasi Promotor Gen LEAFY dengan Tiga Primer Spesifik Gateway
Tahap pertama yang dilakukan dalam percobaan untuk mengkonstruksi promotor gen LEAFY ke dalam vektor ekspresi adalah dengan mendesain tiga buah primer gateway
yang spesifik. Primer tersebut disusun berdasarkan perancangan primer pada sistem
gateway yang didasarkan pada sekuen promotor gen LEAFY yang telah berhasil diisolasi dari daun kakao dengan metode
Genome Walking™ pada penelitian sebelumnya. Ukuran promotor gen LEAFY
yang diperoleh adalah 1650 pb (Santoso 2010). Dari ukuran 1650 pb, akan dibagi kembali ke dalam tiga ukuran yang berbeda melalui amplifikasi promotor gen LEAFY
dengan primer gateway yang spesifik, yaitu
Gateway F1041, Gateway F702, Gateway
F349, dan satu buah primer reverse
(promTcLFY R). Primer tersebut dirancang berdasarkan aturan perancangan sistem
gateway. Empat basa nukleotida GGGG diikuti situs attB kemudian ditambahkan 18-25 urutan basa nukleotida spesifik promotor gen LEAFY (Invitrogen 2010). Situs attB ini disebut sebagai tempat pengikatan lamda (lamda attachment site) yaitu tempat integrasi DNA lamda ke dalam kromosom
E. coli yang terletak di antara operon gal dan operon biotin (Yuwono 2005).
Pemilihan tiga ukuran yang akan dikonstruksi (1041 pb, 702 pb, dan 349 pb) didasarkan pada pencarian panjang ukuran promotor yang efektif dapat mengekspresikan gen. Pencarian tersebut dilakukan dengan pengujian pada beberapa ukuran promotor gen LEAFY. Pengujian ukuran yang diharapkan adalah ½ panjang promotor gen LEAFY (± 825 pb), ½ dari panjang promotor 825 pb (± 415 pb), dan ½ dari panjang promotor 415 (± 210 pb). Namun, setelah dirancang dengan ketentuan aturan sistem gateway, diperoleh primer yang dapat menempel pada proses
amplifikasi promotor gen LEAFY yang berukuran 1650 pb menjadi 1041 pb, 702 pb, dan 349 pb.
Amplifikasi promotor gen LEAFY
dengan metode PCR bertujuan menggandakan promotor gen tersebut secara
in vitro. Hasil amplifikasi tiga ukuran promotor gen LEAFY dengan primer spesifik
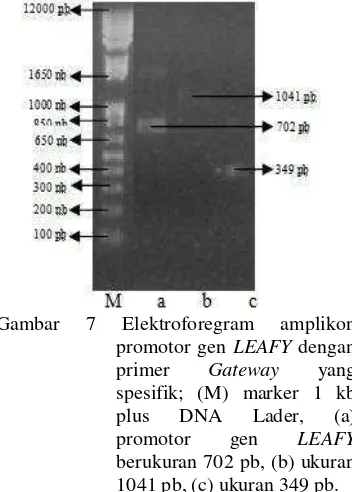
gateway kemudian dielektroforesis dengan konsentrasi gel agarosa 1%. Prinsip elektroforesis adalah memisahkan molekul berdasarkan muatannya. DNA yang bermuatan negatif akan bergerak ke arah kutub positif selama elektroforesis karena adanya gugus fosfat. Fragmen DNA mempunyai muatan negatif yang sama untuk tiap-tiap ukuran panjang, sehingga pergerakan DNA ini akan memiliki kecepatan yang sama untuk mencapai kutub positif. Pergerakan yang sama antar molekul DNA ini tidak akan dapat digunakan untuk memisahkan DNA berdasarkan ukurannya. Hal inilah yang menyebabkan digunakannya gel untuk memperlambat pergerakan DNA. Gel yang digunakan adalah agarosa yang berasal dari ekstrak rumput laut yang telah dimurnikan. Marka atau penanda yang digunakan pada proses running merupakan campuran molekul dengan ukuran berbeda-beda yang digunakan untuk menentukan ukuran molekul dalam pita sampel (Clark & Christopher 2008). Hasil pengujian dengan elektroforesis agarosa menunjukkan pita berukuran sekitar 1041 pb, 702 pb, dan 349 pb (Gambar 7).
Gambar 7 Elektroforegram amplikon promotor gen LEAFY dengan primer Gateway yang spesifik; (M) marker 1 kb plus DNA Lader, (a)
promotor gen LEAFY
Ketiga pita tersebut menunjukkan bahwa primer gateway yang dirancang sudah benar. Hal ini didasarkan pada hasil amplifikasi yang sesuai dengan ukuran yang diharapkan yaitu 1041 pb, 702 pb, dan 349 pb. Promotor gen LEAFY yang telah teramplifikasi selanjutnya diekstraksi dan dimurnikan.
Hasil Ekstraksi dan Pemurnian Promotor
Gen LEAFY dari Gel Agarosa
Pita yang terang pada gel agarosa dipotong dengan pisau scalpel di bawah transluminator UV, kemudian dilanjutkan proses ekstraksi dan pemurnian. Ekstrasi dan pemurnian bertujuan untuk memurnikan DNA dari berbagai pengotor yang tidak diinginkan seperti protein dan RNA. Ukuran pita DNA hasil ekstraksi dan pemurnian sama dengan ukuran pita setelah amplifikasi karena proses esktraksi dan pemurnian hanya menghilangkan pengotor yang ada pada DNA. Hasil ekstraksi dan pemurnian divisualisasi dengan elektroforesis gel agarosa 1 % dan diperoleh pita berukuran 1041 pb, 702 pb, dan 349 pb (Gambar 8). Ukuran pita yang diperoleh sama dengan hasil amplifikasi, sehingga proses ekstraksi dan pemurnian sudah berhasil dilakukan. Hasil ekstraksi dan pemurnian ini kemudian direkombinasikan ke dalam vektor donor menggunakan sistem gateway.
Gambar 8 Elektroforegram ekstraksi dan pemurnian promotor gen
LEAFY dari gel; (a) promotor gen LEAFY berukuran 1041 pb, (b) ukuran 702 pb, (c) ukuran 349 pb, (M) marker 1 kb plus DNA Ladder.
Hasil Reaksi Rekombinasi BP dan LR Promotor Gen LEAFY dengan Sistem Gateway
Pengklonan bertujuan memperbanyak DNA yang tersisip pada sel kompeten (E.coli). Tahapan pengklonan pada sistem
gateway pada dasarnya bergantung pada dua
reaksi, yaitu BP rekombinasi dan LR rekombinasi. Hasil ekstraksi dan pemurnian (elusi) sebanyak 2 µL dicampurkan dengan 1 µL vektor donor (pDONRTM221) melalui reaksi BP. Reaksi BP adalah upaya menyisipkan hasil elusi (fragmen promotor gen LEAFY) ke dalam vektor donor. Promotor gen LEAFY hasil amplifikasi yang memiliki situs attB1 dan attB2 akan direaksikan dengan vektor donor (pDONRTM221) yang memiliki situs attP1 dan situs attP2 sebagai tempat rekombinasi. Adanya situs untuk rekombinasi ini menyebabkan kemungkinan tidak adanya kesalahan orientasi gen yang direkombinasikan. Reaksi ini dikatalisis oleh enzim campuran BP klonase yang memindahkan DNA sisipan ke dalam vektor donor yang menghasilkan suatu klon entri (pENTR) dan diikat oleh dua situs attL. Reaksi BP pada sistem gateway dapat dilihat pada lampiran 4. Setelah BP rekombinasi, selanjutnya di transformasikan ke dalam sel kompeten E. coli galur XL1-Blue. Sel kompeten adalah sel yang mampu menyerap DNA karena telah mendapat perlakuan fisik maupun kimia. Sel dapat dibuat kompeten dengan perlakuan garam CaCl2, LiCl2, atau
RbCl2 (Howe 1995). Garam CaCl2 dapat
meningkatkan porositas dinding sel sehingga afinitas sel terhadap DNA juga meningkat.
Hasil rekombinasi promotor gen LEAFY
pada vektor donor (pDONRTM221) kemudian ditransformasikan ke dalam sel kompeten E. coli galur XL-1 Blue dengan perlakuan kejut panas (heat shock). Heat Shock menyebabkan pori membran sel terbuka dan protein Hsp (heat shock protein) aktif. Kejut panas yang diberikan menyebabkan membran sel kompeten menjadi permeabel terhadap molekul asing sehingga memudahkan plasmid rekombinan masuk ke dalam E. coli. Bakteri E. coli
merupakan bakteri yang paling sering digunakan sebagai inang dalam kloning gen baik sebagai sel inang sementara maupun sel inang tetap. Bakteri ini dipilih karena sifat-sifat fisiologi dan genetika jasad ini telah banyak diketahui. Selain itu, pertumbuhan
Lampiran 3. Koloni yang tumbuh dengan sistem pengklonan gateway semuanya berwarna putih (Gambar 9). Koloni yang diduplikasi setelah reaksi BP sebanyak 10 koloni dari setiap cawan petri. Koloni putih yang tumbuh pada media seleksi diduga kuat klon entri yang membawa promotor gen
LEAFY, sedangkan by product yang tersisipi ccdB akan mati dan tidak tumbuh sebagai koloni putih. Koloni berwarna putih yang tumbuh diduplikasi dan dikultur untuk memperbanyak jumlah plasmid rekombinan. Selanjutnya koloni hasil duplikasi yang tumbuh diambil dengan menggunakan tusuk gigi steril untuk PCR koloni.
Plasmid rekombinan yang telah diikat oleh situs attL1 dan attL2 direkombinasikan ke dalam vektor destinasi yang membawa situs attR1 dan attR2. Reaksi ini dikatalisis oleh enzim campuran LR klonase sehingga dalam sistem gateway reaksi rekombinasi ini disebut reaksi LR (LR reaction). Mekanisme reaksi LR dapat dilihat pada Lampiran 5. Vektor destinasi yang telah tersisipi promotor gen LEAFY (klon ekspresi) ditransformasikan ke E. coli galur XL-1 Blue.
Gambar 9 Koloni yang tumbuh setelah reaksi BP pada promotor gen
LEAFY yang berukuran (a) 349 pb, (b) 702 pb, (c) 1041 pb.
Hasil rekombinasi ke dalam vektor destinasi yang ditransformasikan ke sel kompeten E. coli galur XL-1 Blue ditumbuhkan pada media LB agar. Ke dalam media tersebut ditambahkan antibiotik kanamisin 50 ppm. Hal ini dikarenakan pada vektor destinasi mengandung marka seleksi resisten terhadap antibiotik tersebut sehingga seleksi transforman dilakukan
dengan melihat koloni yang dapat tumbuh pada media tersebut. Koloni putih yang tumbuh setelah reaksi LR lebih sedikit jumlahnya dibandingkan dengan koloni yang tumbuh setelah reaksi BP. Setelah reaksi LR, pada cawan petri promotor gen
LEAFY ukuran 349 pb hanya 3 koloni, pada 702 pb sebanyak 5 koloni, dan pada 1041 pb sebanyak 4 koloni yang diduplikasi dan di PCR koloni. Hal ini terjadi karena koloni rekombinan telah mengalami penyeleksian pada tahap sebelumnya (Gambar 10). Koloni putih yang tumbuh pada media LB agar setelah reaksi LR juga diduplikasi untuk menyimpan duplikat koloni yang telah membawa fragmen promotor gen sisipan agar koloni tidak terkontaminasi dan menghindari terjadinya pertumbuhan bertumpuk (overgrowth) yang mengakibatkan koloni tidak tunggal lagi.
Gambar 10 Koloni yang tumbuh setelah reaksi LR pada promotor gen
LEAFY yang berukuran (a) 349 pb, (b) 702 pb, (c) 1041 pb.
Hasil Konfirmasi Koloni Transforman setelah Reaksi BP dengan Teknik PCR Koloni
Koloni bakteri yang tumbuh setelah transformasi ke dalam E. coli dikonfirmasi dengan teknik PCR koloni untuk memastikan koloni tersebut mengandung plasmid rekombinan. Pengujian koloni yang mengandung plasmid rekombinan setelah reaksi BP dengan metode PCR koloni menggunakan sepasang primer universal M13. Penggunaan primer ini karena pada peta pDONRTM221 terlihat bahwa amplifikasi dengan PCR koloni memerlukan primer M13 Forward dan M13 Reverse
setelah reaksi BP pada promotor gen LEAFY
yang berukuran 349 menunjukkan bahwa hanya tujuh dari sepuluh koloni yang diujikan mengandung promotor gen LEAFY. Hal ini ditunjukkan dengan adanya pita berukuran sekitar 349 pb setelah elektroforesis (Gambar 11). Berbeda dengan hasil PCR koloni pada promotor berukuran 702 pb, dari sepuluh koloni hanya tiga koloni yang positif (Gambar 12), sedangkan promotor yang berukuran 1041 pb, dari sepuluh koloni hanya dua koloni yang positif mengandung promotor gen LEAFY
ukuran 1041 pb (Gambar 13).
Gambar 11 Elektroforegram PCR koloni rekombinasi promotor gen
LEAFY ukuran 349 pb pada vektor donor; (1-10) koloni bakteri, (M) marker 1 kb plus DNA Ladder.
Gambar 12 Elektroforegram PCR koloni rekombinasi promotor gen
LEAFY ukuran 702 pb pada vektor donor; (M) marker 1 kb plus DNA Ladder, (1-10) koloni bakteri.
Gambar 13 Elektroforegram PCR koloni rekombinasi promotor gen
LEAFY ukuran 1041 pb pada vektor donor; (1-10) koloni bakteri, (M) marker 1 kb plus DNA Ladder.
Keberhasilan proses PCR koloni dapat diamati dari ukuran klon transforman. Ukuran pita yang ditunjukkan ketiga gambar hasil PCR koloni menunjukkan ukuran sekitar 349 pb, 702 pb, dan 1041 pb, maka dapat dipastikan bahwa proses penyisipan fragmen promotor gen LEAFY sudah berhasil dilakukan ke dalam vektor donor (pDONRTM221). Koloni yang positif sudah tersisip fragmen promotor gen LEAFY
dikultur dalam media LB cair yang mengandung kanamisin 50 ppm untuk perbanyakkan koloninya, setelah itu dilakukan proses isolasi plasmid rekombinan untuk konfirmasi penyisipan fragmen promoter gen pada vektor donor. Media Luria Bertani (LB) cair sebagai sumber nutrisi untuk membantu pertumbuhan bakteri. Media LB merupakan media kompleks yang terdiri atas tripton, ekstrak khamir, dan NaCl. Tripton berfungsi menyediakan asam amino dan peptida pendek sedangkan ekstrak khamir sebagai sumber nitrogen, gula, dan nutrisi organik maupun anorganik.
Hasil Konfirmasi Koloni Transforman setelah Reaksi BP dengan Teknik Isolasi DNA Plasmid Rekombinan (Fermentas)
Hasil kultur bakteri yang ditumbuhkan dari media LB cair kemudian diisolasi DNA plasmidnya untuk rekombinasi ke vektor destinasi. Isolasi DNA plasmid dilakukan berdasarkan hasil PCR DNA koloni yang diketahui mengandung plasmid terinsersi fragmen yang diinginkan. Plasmid diisolasi dengan Fermentas GeneJet Plasmid Miniprep Kit. Hasil isolasi DNA plasmid dielektroforesis pada gel agarosa 1% sehingga dapat diamati pita-pita DNA-nya.
M1 2 3 4 5 6 7 8 9 10
1 2 3 4 5 6 7 8 9 10M
Elektroforegram menunjukkan bahwa pita berada sekitar 5110 pb, 5463 pb, dan 5802 pb (Gambar 14). Elektroforegram ini menunjukkan ukuran promotor gen sisipan ditambah dengan ukuran vektor donor (pDONRTM221) sebesar 4761 pb yang berada pada plasmid rekombinan. Fragmen promotor gen LEAFY yang berukuran 349 pb ditambah dengan ukuran vektor pDONRTM221 sebesar 4761 pb sehingga ukuran plasmid rekombinan menjadi sekitar 5110 pb, fragmen promotor gen LEAFY
ukuran 702 pb ditambah ukuran pDONRTM221 menjadi sekitar 5463 pb, dan pada fragmen ukuran 1041 pb ditambah ukuran vektor donor diperoleh plasmid rekombinan sekitar 5802 pb. Isolasi plasmid rekombinan dari ketiga koloni telah berhasil dilakukan. Hal ini dibuktikan dengan terlihat pita pada ukuran yang sudah sesuai dengan teori perhitungan plasmid rekombinan. Hasil positif dari konfirmasi dengan isolasi DNA plasmid rekombinan menunjukkan bahwa proses transformasi ke dalam E.coli telah berhasil dilakukan dengan baik.
Gambar 14 Elektroforegram hasil isolasi plasmid rekombinan setelah reaksi BP; (1) plasmid rekombian berukuran sekitar 5110 pb, (2) ukuran sekitar 5463 pb, (3) ukuran sekitar 5802 pb, (M) marker 1 kb plus DNA Ladder.
Isolasi DNA plamid dengan kit ini, pada prinsipnya sama dengan pemisahan plasmid berdasarkan ukuran. Proses pemisahan plasmid berdasarkan ukuran dimulai dengan pembuatan ekstrak sel. Jika sel dipecah secara perlahan pada kondisi terkendali DNA kromosom yang putus hanya sebagian kecil, sehingga fragmen DNA kromosom berukuran jauh lebih besar dari ukuran plasmid dan dapat dipisahkan dari plasmid dengan cara mengendapkan DNA kromosom bersama serpihan sel (debris) melalui sentrifugasi. Cara ini dapat meminimalkan kerusakan DNA kromosom
dan supernatan (larutan jernih) hasil sentrifugasi yang hampir seluruhnya mengandung DNA plasmid. DNA plasmid yang telah berhasil diisolasi selanjutnya direkombinasikan ke dalam vektor destinasi khusus untuk promotor, yaitu vektor destinasi yang memiliki marka gen.
Hasil Konfirmasi Koloni Transforman setelah Reaksi LR dengan Teknik PCR Koloni
PCR koloni dapat mengkonfirmasi sel transforman yang sudah tersisipi DNA rekombinan. Melalui tahapan ini, promotor gen LEAFY yang disisipkan ke dalam vektor destinasi diamplifikasi menggunakan primer spesifik. Program PCR koloni yang terlebih dahulu harus dilakukan adalah program lisis sel yang diawali dengan pemanasan pada suhu 96oC selama 5 menit, 50oC selama 1 menit 30 detik, 96oC selama 1 menit 30 detik, 45oC selama 1 menit 30 detik, 96oC selama 1 menit, dan 40oC selama 1 menit. Program lisis sel E. coli dilakukan untuk memecah sel sehingga DNA sel transforman dapat digunakan dalam proses PCR untuk konfirmasi sel transforman. Setelah program lisis dihentikan, komponen mix ditambahkan ke dalam masing-masing tabung. Kemudian program PCR dilanjutkan kembali dengan program PCR yaitu, 94oC selama 30 detik, 45oC selama 1 menit, 72oC selama 2 menit. Hasil visualisasi PCR koloni dapat dilihat dengan elektroforesis gel agarosa 1 %.
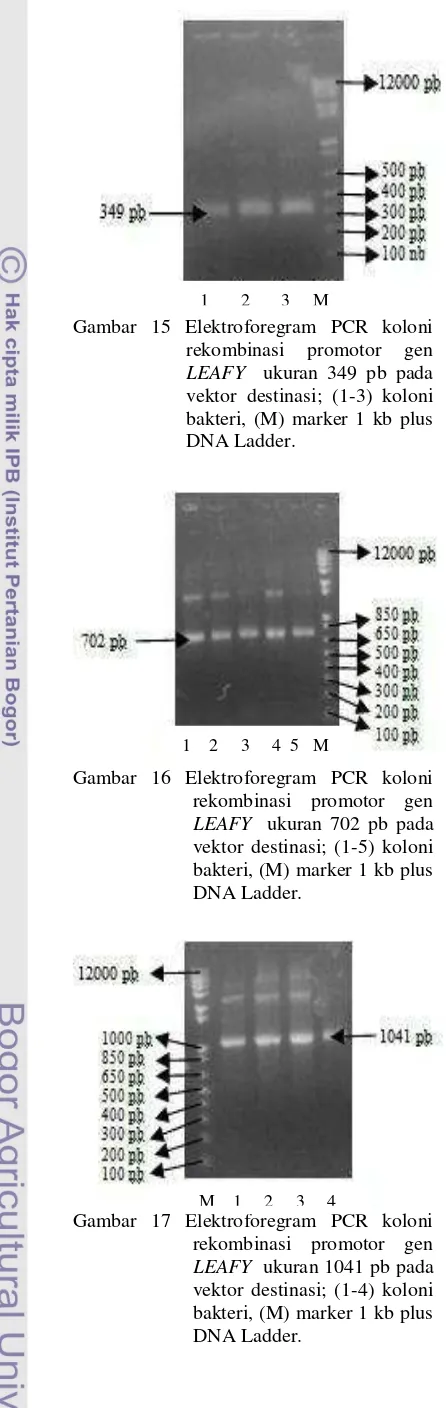
Gambar 15 Elektroforegram PCR koloni rekombinasi promotor gen
LEAFY ukuran 349 pb pada vektor destinasi; (1-3) koloni bakteri, (M) marker 1 kb plus DNA Ladder.
Gambar 16 Elektroforegram PCR koloni rekombinasi promotor gen
LEAFY ukuran 702 pb pada vektor destinasi; (1-5) koloni bakteri, (M) marker 1 kb plus DNA Ladder.
Gambar 17 Elektroforegram PCR koloni rekombinasi promotor gen
LEAFY ukuran 1041 pb pada vektor destinasi; (1-4) koloni bakteri, (M) marker 1 kb plus DNA Ladder.
Hasil Konfirmasi Koloni Transforman setelah Reaksi LR dengan Teknik Isolasi DNA Plasmid Rekombinan (Fermentas)
Koloni E. coli yang positif mengandung sisipan promotor gen LEAFY, selanjutnya dikonfirmasi kembali dengan teknik isolasi DNA plasmid rekombinan. Secara umum, isolasi plasmid mirip dengan isolasi DNA total, yaitu melibatkan proses pembiakan sel pembawa plasmid, pemanenan sel, pembuatan ekstrak sel, penghilangan protein dan RNA, dan pemekatan DNA dengan presipitasi etanol. Pembiakan sel pembawa plasmid rekombinan dilakukan dengan membiakkannya dalam media LB cair yang sudah ditambahkan antobiotik kanamisin 50 ppm. Penambahan antibiotik kanamisin 50 ppm dilakukan untuk menyeleksi koloni transforman, maka hanya koloni yang mengandung DNA rekombinan yang akan tumbuh dan dapat dibiakkan. Pembiakan sel dilakukan dalam inkubator bergoyang dengan suhu 37oC selama 16 jam. Hasil dari proses pembiakkan, yaitu larutan dalam botol yang diinkubasi dan dikocok menjadi keruh. Setelah inkubasi, dilakukan pemanenan sel untuk selanjutnya diisolasi DNA plasmidnya. Pemanenan bakteri dilakukan secara steril di dalam laminar air flow cabinet. Sebanyak 1-5 ml E. coli hasil pemanenan siap diisolasi DNA plasmid.
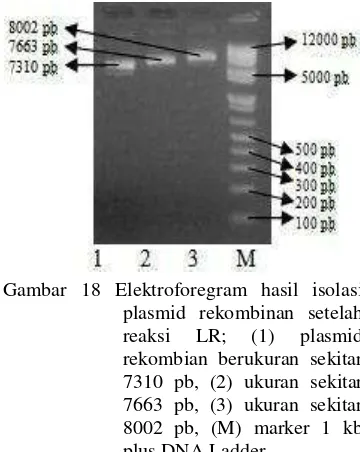
Isolasi DNA plasmid rekombinan setelah reaksi LR dengan menggunakan kit dari Fermentas. Elektroforegram DNA plasmid menunjukkan pita berukuran sekitar 7310 pb, 7663 pb, dan 8002 pb (Gambar 18). Ukuran ini menunjukkan ukuran promotor gen sisipan (349 pb, 702 pb, dan 1041 pb) ditambah dengan ukuran vektor destinasi khusus promotor (pDEST) yang berukuran 6961 pb. Penambahan ukuran fragmen promotor gen LEAFY 349 ditambah dengan 6961 pb ukuran vektor menghasilkan plasmid rekombinan sebesar 7310 pb. Perhitungan pada promotor gen LEAFY
ukuran 702 pb dan 1041 pb juga sudah sesuai dengan teori, yaitu 702 pb ditambah dengan 6961 pb menjadi 7663 pb dan 1041 ditambah dengan vektor menjadi 8002 pb. Hasil visualisasi isolasi plasmid menunjukkan bahwa proses isolasi plasmid rekombinan dari ketiga koloni yang mengandung DNA rekombinan telah berhasil dilakukan. Hasil positif dari konfirmasi dengan isolasi DNA plasmid rekombinan menunjukkan bahwa proses transformasi ke dalam sel kompeten E.coli
telah berhasil dilakukan dengan baik.
Gambar 18 Elektroforegram hasil isolasi plasmid rekombinan setelah reaksi LR; (1) plasmid rekombian berukuran sekitar 7310 pb, (2) ukuran sekitar 7663 pb, (3) ukuran sekitar 8002 pb, (M) marker 1 kb plus DNA Ladder.
Hasil Transformasi Klon Ekspresi ke dalam Agrobacterium tumefaciens Galur AGL-O
Transformasi ke dalam Agrobacterium tumefaciens (A. tumefaciens) bertujuan untuk menguji ekspresi pengaruh promotor gen tersebut ke tanaman. Sel Agrobacterium
yang membawa plasmid rekombinan digunakan untuk menginfeksi protoplast tanaman. Penggunaan A. tumefaciens galur AGL-0 dikarenakan galur ini mempunyai virulensi yang tinggi dibandingkan galur
Agrobacterium lainnya. DNA plasmid setelah rekombinasi pada vektor destinasi diisolasi kemudian ditransformasikan ke dalam A. tumefaciens dengan metode kejut dingin. Hal ini dikarenakan terjadi lonjakan suhu es 0oC ke nitrogen cair yang bersuhu sekitar -196oC. Lonjakan suhu tersebut akan menjadikan membran sel Agrobacterium
menjadi tidak selektif terhadap molekul asing sehingga DNA sisipan dapat masuk. Hasil transformasi ke dalam A. tumefaciens
dapat dilihat setelah inkubasi selama ± 2 hari. Pada saat proses inkubasi, koloni dalam cawan petri dibungkus dengan kertas agar kondisi menjadi gelap. Hal ini dilakukan agar koloni A. tumefaciens dapat menyesuaikan kondisinya selama hidup di dalam tanah (akar). Koloni yang tersisipi oleh promotor gen LEAFY ukuran 349 pb hanya ada dua koloni, sedangkan koloni yang tersisipi oleh promotor ukuran 702 dan 1041 hanya satu koloni saja (Gambar 19).
Seleksi transforman yang sudah ditransformasikan ke dalam A. tumefaciens
dilakukan dengan cara menumbuhkannya di
dalam media LB agar yang mengandung antibiotik kanamisin 50 ppm dan rifampisin 50 ppm. Sel transforman akan tumbuh membentuk koloni berwarna putih. Antibiotik kanamisin digunakan karena vektor destinasi (pDEST) yang digunakan membawa marka seleksi berupa gen resistensi terhadap antobiotik kanamisin, sedangkan rifampisin digunakan untuk membunuh E. coli. Koloni yang tumbuh dimedia seleksi lebih sedikit (1 sampai 2 koloni) dibandingkan dengan koloni yang tumbuh saat transformasi ke E. coli (sepuluh koloni). Hal ini dikarenakan A. tumefaciens
memiliki jumlah salinan yang lebih sedikit (low copy number) dibandingkan E. coli.
Gambar 19 Koloni Agrobacterium tumefaciens yang tumbuh setelah reaksi LR pada promotor gen LEAFY
yang berukuran (a) 349 pb, (b) 702 pb, (c) 1041 pb.
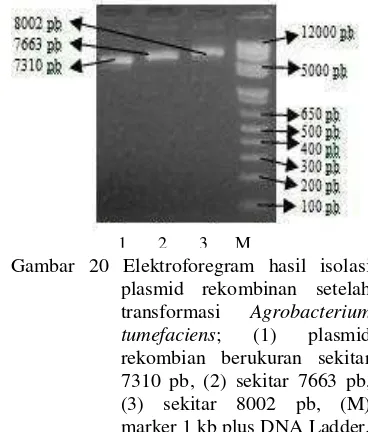
Koloni yang tumbuh diisolasi DNA plasmidnya untuk menguji plasmid rekombinan yang sudah tersisipi di dalam koloni A. tumefaciens yang tumbuh tersebut. Hasil isolasi DNA plasmid menunjukkan pita yang terlihat berukuran sekitar 7310 pb, 7663 pb, dan 8002 pb (Gambar 20). Ukuran tersebut sama dengan ukuran hasil isolasi plasmid rekombinan setelah reaksi LR. Ukuran hasil isolasi DNA plasmid menunjukkan ukuran promotor gen sisipan (349 pb, 702 pb, dan 1041 pb) ditambah dengan ukuran vektor destinasi khusus promotor (pDEST) yang berukuran 6961 pb, oleh karena itu dapat disimpulkan bahwa plasmid rekombinan di dalam vektor ekspresi dapat ditransformasikan ke dalam
Gambar 20 Elektroforegram hasil isolasi plasmid rekombinan setelah transformasi Agrobacterium tumefaciens; (1) plasmid rekombian berukuran sekitar 7310 pb, (2) sekitar 7663 pb, (3) sekitar 8002 pb, (M) marker 1 kb plus DNA Ladder.
SIMPULAN DAN SARAN
Simpulan
Tiga ukuran promotor gen LEAFY, yaitu 349 pb, 702 pb, dan 1041 pb telah berhasil disisipkan pada vektor ekspresi dengan sistem gateway. Ukuran promotor gen
LEAFY pada plasmid rekombinan setelah reaksi BP sekitar 5110 pb, 5463 pb, dan 5802 pb. Setelah reaksi LR hasil isolasi DNA plasmid menunjukkan pita yang terlihat berukuran sekitar 7310 pb, 7663 pb, dan 8002 pb. DNA plasmid dalam vektor ekspresi telah berhasil ditransformasikan ke
Agrobacterium tumefaciens galur AGL-O dan siap ditransfer ke dalam tanaman.
Saran
Penelitian lebih lanjut perlu dilakukan untuk mengetahui ekspresi promotor gen
LEAFY melalui tanaman tembakau sebagai tanaman model dengan menggunakan reporter gen seperti GUS (β-glukoronidase).
DAFTAR PUSTAKA
Alvim PT, AD Machado, F Vello. 1974. Physiological responses of cacao to environt factors. J Revista Theobroma 4(3):3-12.
Bernard P, Couturier M. 1992. Cell killing by the F plasmid ccdB protein involves poisoning of DNA-topoisomerase II complexes. J Mol Biol 226:735-745.
Brian. 2003. Developmental biology. [terhubung berkala]. http://www. mun.ca/biology/desmid/BIOL3530W 2003/DB Ch07/DBNPlant.html [23 April 2011].
Brown TA. 2000. Essential Molecular Biology: A Practical Approach. New York: Oxford University Press Inc.
Brown TA. 2002. Genomes Second Editions. New York: Joh Wiley and Sons Inc.
Clark W, Chistpher K. 2008. An
Introduction to DNA:
Spectrophotometry, Degradation, and The “Frangekel” Eksperimen. Alberta: University of Alberta.
[Depperin] Departemen Perindustrian. 2007. Gambaran sekilas industri kakao. [terhubung berkala]. http://www. depperin.go.id [10 April 2011].
[Deptan] Departemen Pertanian. 2007. Prospek dan arah pengembangan agribisnis kakao. Ed ke-2. [terhubung berkala]. http://www. deptan.go.id [10 April 2011].
Dubin MJ, Bowler C, Benvenuto G. 2008. A modified gateway cloning strategy for overexpressing tagged proteins in plants. Plant methods 4:1-11.
Fermentas. 2006. GeneJetTM plasmid miniprep kit. Life Science.
Hartley JL, Temple GF, Brasch MA. 2000. DNA cloning using in vitro site-specific recombination. Genome Res
10:1788-1795.
Howe C. 1995. Gene Cloning an
Manipulation. Cambridge:
Cambridge University Press.
International Cocoa Organization (ICCO). 2003. Quarterly Bulletin of Cocoa Statistics. Vol: XXIX.