BIOFERMENTASI DENGAN JAMUR TIRAM
(Pleurotus ostreatus) DAN AMONIASI
INSUN SANGADJI
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Dengan ini saya menyatakan bahwa disertasi Mengoptimalkan Pemanfaatan Ampas Sagu sebagai Pakan Ruminansia Melalui Biofermentasi dengan Jamur Tiram (Pleurotus ostreatus) dan Amoniasi adalah karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber informasi yang berasal dari atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi.
Bogor, Juli 2009
Insun Sangadji
Pakan Ruminansia Melalui Biofermentasi dengan Jamur Tiram (Pleurotus ostreatus) dan Amoniasi. Dibimbing oleh AMINUDDIN PARAKKASI, KOMANG G WIRYAWAN dan BUDI HARYANTO.
Di beberapa daerah pakan hijauan dirasakan semakin sulit diperoleh terutama di musim kering, sehingga diperlukan usaha mencari alternatif penggantinya. Salah satu alternatif adalah dengan memanfaatkan limbah pengolahan sagu yang di beberapa daerah saat ini belum dimanfaatkan secara optimal. Pemanfaatannya sebagai pakan memerlukan pengolahan awal karena kandungan nutrisinya yang rendah
Bioteknologi yang paling mungkin digunakan untuk perbaikan mutu pakan baik secara skala laboratorium maupun skala di masyarakat adalah teknologi biokonversi. Pada proses ini suatu senyawa akan diubah menjadi senyawa lain yang strukturnya hampir sama oleh kerja enzim mikroba. Dalam proses ini dibutuhkan mikroorganisme yang mampu mendegradasi lignin atau komponen serat lainnya melalui proses fermentasi. Proses ini tidak hanya diharapkan mampu mendegradasi komponen sulit cerna pada ampas sagu bagi mikroba rumen, namun juga dapat meningkatkan nutrisi ampas sagu tersebut sebagai bahan baku pakan. Salah satu mikroorganisme yang dapat digunakan adalah jamur tiram (Pleurotus ostreatus) yang mengandung enzim-enzim pendegradasi lignin. Cara lain untuk perbaikan mutu ampas sagu dapat juga dilakukan melalui proses amoniasi dengan urea.
Tujuan Penelitian ini adalah : (1) memanfaatkan sumber daya pakan lokal hasil ikutan pengolahan tepung sagu rumbiah; (2) mengkaji potensi ampas sagu sebagai media tumbuh jamur tiram; (3) mengkaji nilai nutrisi ampas sagu hasil biofermentasi dan amoniasi; (3) mendapatkan formula ransum dengan bahan dasar ampas sagu untuk produktivitas ternak sapi
Ada beberapa tahap penelitian yang telah dilakukan, di Laboratorium Mikrobiologi dan Biokimia PAU, Laboratorium Biologi Hewan PAU dan Laboratorium Makanan Balitnak Ciawi, kebun jamur rakyat bertempat di Pondok Pesantren Tarbiyatunnisa Kelurahan Bantar Kambing Kabupaten Bogor, kandang Peternakan rakyat di Depok Jakarta. Pelaksanaannya mulai dari bulan Mei 2006 sampai April 2008.
Rancangan yang digunakan adalah Rancangan Acak Kelompok (RAK) dengan 10 perlakuan ransum dan 3 kelompok waktu pengambilan cairan rumen. Perlakuan ransum adalah ransum kontrol, ransum yang mengandung ampas sagu fermentasi 15%, 30% dan 45%, ransum yang mengandung ampas sagu amoniasi 15%, 30% dan 45% dan ransum yang mengandung ampas sagu tanpa pengolahan 15%, 30% dan 45%. Penggunaan ampas sagu baik fermentasi. amoniasi maupun tanpa pengolahan dalam ransum menggantikan rumput rumput lapangan. Uji lanjut menggunakan uji Duncan. Peubah yang diukur adalah kecernaan bahan kering, bahan organik, kadar NH3, kadar VFA total dan pH cairan rumen. Penelitian
tahap III: Pengujian substitusi rumput lapangan dengan ampas sagu dalam ransum sapi Bali. Penelitian ini juga menggunakan hasil penelitian terbaik pada penelitian sebelumnya yaitu ransum kontrol, ransum yang mengandung ampas sagu fermentasi 15%, 30% dan ransum yang mengandung ampas sagu amoniasi 15%, 30%. Ternak yang digunakan adalah 15 ekor sapi Bali jantan muda yang berumur ± 1.5 tahun, yang dikelompokkan berdasarkan bobot badan. Rancangan yang digunakan adalah Rancangan Acak Kelompok dengan 5 perlakuan ransum dan 3 kelompok ternak. Uji lanjut menggunakan uji Duncan. Peubah yang diukur adalah konsumsi bahan kering, bahan organik, pertambahan bobot badan,konversi ransum, kecernaan (bahan kering, bahan organik, protein ADF, NDF dan Selulosa), kadar NH3, kadar VFA total dan parsial, dan income over feed cost (IOFC), retensi N dan kadar kolesterol feses.
Hasil penelitian tahap I; menunjukkan bahan kering, bahan organik, protein dan semua komponen serat hanya dipengaruhi oleh waktu inkubasi dan tidak dipengaruhi oleh dosis Mn. Demikian juga tidak ada interaksi antara keduanya. Hasil penelitian yang digunakan pada penelitian selanjutnya adalah perlakuan dengan waktu inkubasi panen pertama dan panen kedua dengan dosis Mn 0 ppm. Penelilitian tahap II menunjukkan kadar amonia dan pH cairan rumen tidak berbeda antar perlakuan, sedangkan kadar VFA total, kecernaan bahan kering dan bahan organik berbeda (P<0.05) antar perlakuan. Perlakuan terbaik yang digunakan pada penelitian selanjutnya adalah perlakuan dengan menggunakan ampas sagu fermentasi dan amoniasi 15 dan 30 % dibandingkan dengan perlakuan kontrol. Pemilihan ini didasarkan pada hasil tertinggi kecernaan bahan kering dan bahan organik. Penelitian tahap III, menunjukkan bahwa konsumsi bahan kering, bahan organik, pertambahan bobot badan, konversi ransum dan income over feed cost berbeda (P<0.05) antar perlakuan. Kecernaan bahan kering, bahan organik, protein NDF dan selulosa tidak berbeda antar perlakuan, sedangkan kecernaan ADF berbeda antar perlakuan. Demikian juga karakteristik rumen tidak berbeda antar perlakuan. Hasil penelitian ini menunjukkan bahwa ampas sagu terolah dapat digunakan sampai 30% dalam ransum sapi Bali atau 50% menggantikan rumput lapangan.
© Hak Cipta milik IPB, tahun 2009 Hak Cipta dilindungi Undang-Undang
1. Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya.
a. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah
SEBAGAI PAKAN RUMINANSIA MELALUI
BIOFERMENTASI DENGAN JAMUR TIRAM
(Pleurotus ostreatus) DAN AMONIASI
INSUN SANGADJI
Disertasi
sebagai salah satu syarat untuk memperoleh gelar Doktor pada
Program Studi Ilmu Ternak
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
Penguji luar komisi pembimbing :
Nama : Insun Sangadji
NRP : D061030141
Disetujui Komisi Pembimbing
Prof. Dr. Drh. Aminuddin Parakkasi, M.Sc. Ketua
Dr. Ir. Komang G. Wiryawan Prof. Dr. Ir. Budi Haryanto, M.Sc. Anggota Anggota
Mengetahui
Ketua Departemen Dekan Sekolah Pascasarjana Ilmu Nutrisi dan Teknologi Pakan
Dr. Ir. Idat G. Permana, M.Sc.Agr. Prof. Dr. Ir. Khairil Anwar Notodiputro,M.S.
Puji syukur penulis panjatkan kehadirat Allah SWT atas segala rahmat dan karuniaNYA karya tulis ini bisa terselesaikan. Disertasi dengan judul Mengoptimalkan Pemanfaatan Ampas Sagu sebagai Pakan Ruminansia Melalui Biofermentasi dengan Jamur Tiram (Pleurotus ostreatus) dan Amoniasi, disusun sebagai salah satu syarat untuk memperoleh gelar Doktor pada Sekolah Pascasarjana Institut Pertanian Bogor. Penelitian dilaksanakan di Laboratorium Biokimia dan Mikrobiologi PAU, Laboratorium Biologi Hewan PAU, Kebun Jamur rakyat yang bertempat di Pesantren Tarbiyatunnisa Bantar Kambing, Bogor dan kandang peternakan rakyat di Depok Jakarta.
Penelitian dan penulisan ini dapat terselesaikan atas pengarahan serta bimbingan dari Komisi Pembimbing.Ucapan terima kasih dan penghargaan yang sebesar-besarnya penulis sampaikan kepada Prof. Dr. Drh.Aminuddin Parakkasi, M.Sc., Dr. Ir. Komang G. Wiryawan dan Prof. Dr. Ir. Budi Haryanto, M.Sc. yang telah membimbing, memotivasi dan mengarahkan dengan kesabaran yang bersahaja dialiri energi positif dalam setiap menghadapi kesulitan. Ucapan terima kasih juga disampaikan kepada, Dekan, Ketua Program Studi, Ketua Departemen dan seluruh staf dosen Fapet IPB atas kesempatan memperoleh ilmu dan bantuan yang diberikan selama penulis menjalankan studi di IPB. Penghargaan penulis sampaikan kepada Bapak Rektor Unpatti, Dekan Fakultas Pertanian Unpatti, Ketua Jurusan Peternakan, Fakultas Pertanian Unpatti, yang telah memberikan kesempatan kepada penulis untuk menjalankan studi di IPB. Terima kasih juga penulis sampaikan kepada Departemen Pendidikan Nasional melalui DITJEN DIKTI atas bantuan beasiswa yang diberikan, Pemerintah Daerah Maluku, Yayasan Dana Bantuan Maluku, Bantuan Dana Mandiri, Bapak Ketua MUI Ambon, Badan Litbang Departemen Pertanian yang telah memberikan bantuan penelitian kepada penulis.
Ucapan terima kasih kepada teman-temanku tersayang Nani, usi Popy, pak Dede, Pak Dedi Rahmat, Hary Uhie, ibu Maritje, Ari Borel yang telah banyak memberikan saran dan masukan yang sangat bermanfaat dalam penulisan ini. Ungkapan terima kasih juga penulis sampaikan kepada pak Iwa, Endang, Rahmat, mas Abdi, Budi, Iwan dan Bram atas segala bantuan selama penulis mengadakan penelitian baik di Laboratorium maupun di lapangan. Kepada teman-teman kos Ibu Dety, Mita, Eli, Eka, Mala dan Tika terima kasih untuk kenyamanan kebersamaan dan bantuan yang diberikan selama ini. Terima kasih yang tak terhingga penulis sampaikan kepada kakak-kakak, adik-adik dan ponakan-ponakan tercinta, tulisan ini kupersembahkan kepada kalian untuk semua doa dan kasih sayang yang telah diberikan.
Semoga tulisan ini bermanfaat.
Bogor, Juli 2009
Penulis dilahirkan di Namlea Kabupaten Buru, Propinsi Maluku 4 Juli 1961 sebagai anak ke 9 dari 11 bersaudara dari pasangan Alm. H. Ali Sangadji dan Alm. Hj. Nun Malawat. Program sarjana ditempuh di Jurusan Peternakan Fakultas Pertanian Unpatti, lulus pada tahun 1988. Pada tahun 1995-1996 penulis mengikuti Bridging program untuk tenaga pengajar kimia dasar pada Institut Teknologi Bandung. Pada tahun 1996, penulis diterima di program studi Ilmu Ternak pada Program Pascasarjana IPB dan menamatkannya pada tahun 1998. Kesempatan untuk melanjutkan ke program doktor pada program studi ilmu dan perguruan tinggi yang sama diperoleh pada tahun 2003 dengan beasiswa BPPS Ditjen Dikti Depdiknas.
Saat ini penulis terdaftar sebagai dosen tetap pada Jurusan Peternakan, Fakultas Pertanian Universitas Pattimura Ambon.
Sebuah artikel telah diterbitkan dengan judul Perubahan Nilai Nutrisi Ampas Sagu pada Fase Pertumbuhan Jamur Tiram (Pleurotus ostreatus) yang Berbeda pada jurnal Ilmu ternak, Fakultas Peternakan Universitas Padjadjaran, Bandung tahun 2008. Artikel lain yang berjudul Produktivitas sapi Bali yang diberi Pakan Berbahan Dasar Ampas Sagu Hasil Fermentasi Jamur Tiram
DAFTAR GAMBAR ……….
xiv
DAFTAR LAMPIRAN ………...
xv
1 PENDAHULUAN ………
1
1.1 Latar Belakang ………..
1
1.2 Tujuan Penelitian ………...
4
1.3 Manfaat Penelitian ……….
4
1.4 Hipotesis Penelitian ………...
4
2 TINJAUAN PUSTAKA ………..
5
2.1 Tanaman Sagu ………...
5
2.2 Potensi Ampas Sagu sebagai Komponen Pakan ………...
6
2.3 Komposisi Kimia dan Komponen Ampas Sagu ...
8
2.4 Pengolahan Pakan Berserat ...
2.4.1 Pengolahan Pakan Berserat Secara fisik ...
2.4.2 Pengolahan Pakan Berserat Secara Kimiawi ...
2.4.3 Pengolahan Pakan Berserat Secara Biologis ...
16
17
18
19
2.5
Jamur Tiran (Pleurotus ostreatus) dan Peranannya dalam
Perbaikan Mutu Pakan ...
21
2.6 Peranan Amoniasi terhadap Perbaikan Mutu Pakan ...
26
2.7
Sistem Pencernaan dan Penyerapan Nutrien pada
Ruminansia ...
27
2.8 Peranan Serat terhadap Kolesterol ...
31
3 MATERI DAN METODE ...
33
3.1 Waktu dan Lokasi Pelaksanaan ...
33
3.2 Materi Percobaan ...
33
3.3 Tahap I. Biofermentasi Ampas Sagu dengan Jamur Tiram
(Pleurotus ostreatus) tanpa atau dengan Mineral Mn
dengan Masa Inkubasi Berbeda ...
33
3.4 Tahap II. Evaluasi In Vitro Ampas Sagu yang Difermentasi
dengan Jamur Tiram (Pleurotus ostreatus) dan
Amoniasi ………
36
3.5
Tahap III. Pengujian Substitusi Rumput Lapangan dengan
Ampas Sagu Hasil Pengolahan dalam Ransum Sapi Bali ...
40
3.6 Metode Pengukuran Peubah ...
43
4 HASIL DAN PEMBAHASAN ...
49
4.1
Tahap I Biofermentasi Ampas Sagu dengan Jamur Tiram
(Pleurotus ostreatus) tanpa atau dengan Mineral Mn
dengan Masa Inkubasi Berbeda ...
49
4.1.1
Potensi Ampas Sagu sebagai Media Tumbuh Jamur
Tiram (Pleurotus ostreatus) ...
49
4.1.2 Pertumbuhan Miselium ...
50
4.1.3
Kandungan Bahan kering, Bahan Organik dan
Protein Ampas Sagu Hasil Biofermentasi ...
53
Amoniasi ... ...
61
4.2.1 Fermentabilitas rumen In Vitro ...
61
4.2.2
Kecernaan Bahan Kering dan Bahan Organik
in vitro ...
64
4.3. Tahap III. Pengujian Substitusi Rumput Lapangan dengan
Ampas Sagu Hasil Pengolahan dalam Ransum Sapi Bali ...
67
4.3.1
Konsumsi, Pertambahan Bobot Badan dan
Konversi Ransum ...
67
4.3.2
Kecernaan Nutrien, Retensi Nitrogen dan
Kolesterol Feses ...
71
4.3.3 Karakteristik Fermentasi Mikroba dalam Rumen...
75
4.3.3.1. Amonia ...
75
4.3.3.2. Asam Lemak Terbang (VFA) ...
76
5 SIMPULAN DAN SARAN ...
80
5.1 SIMPULAN ...
80
5.2 SARAN ...
80
DAFTAR PUSTAKA ...
81
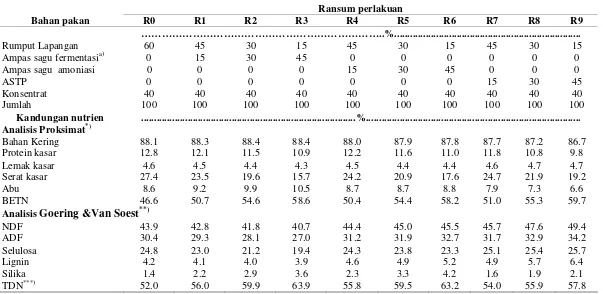
1 Komposisi nutrisi ampas sagu dan rumput lapangan ………
9
2 Pembagian bahan organic tanaman melalui sistem analisis
deterjen ………...
10
3 Pengaruh metode pengolahan terhadap konsumsi pakan dan
Perbaikan kecernaan bahan kering ……….
17
4 Perlakuan kimiawi untuk meningkatkan mutu bahan pakan
lokal ………
18
5 Beberapa jenis jamur yang dapat digunakan untuk
mendegradasi lignin ………
20
6 Kandungan nutrien bahan pakan penyusun ransum
percobaan in vitro ...
37
7 Susunan ransum dan kandungan nutrien ransum percobaan
in vitro ...
38
8 Kandungan nutrien bahan pakan penyusun ransum
percobaan in vivo ...
41
9 Susunan bahan pakan dan kandungan nutrien ransum
percobaan in vivo ...
42
10 Komposisi larutan penyangga ...
44
11 Kandungan nutrien ampas sagu bahan penelitian ...
50
12 Kandungan bahan kering, bahan organik dan protein
media tumbuh jamur dengan waktu fermentasi dan
dosis Mn yang berbeda . ...
54
13 Komponen serat media tumbuh jamur pada
waktu fermentasi dan dosis Mn yang berbeda ...
58
14 Rataan pH, konsentrasi amonia dan VFA total cairan
rumen in vitro ...
62
15 Kecernaan bahan kering dan bahan organik in vitro ...
65
16 Rataan konsumsi bahan kering, bahan organik, bobot badan awal,
akhir, pertambahan bobot badan dan konversi ransum ...
69
17 Rataan perhitungan ekonomis pakan terhadap pertambahan
bobot badan sapi ...
71
18 Rataan kecernaan nutrien pada sapi Bali...
72
19 Konsumsi nitrogen, nitrogen feses, nitrogen urine dan
retensi nitrogen pada sapi Bali ...
74
20 Rataan konsentrasi Amonia, VFA total dan parsial cairan
rumen sapi Bali ...
77
21 Rataan proporsi molar VFA parsial cairan rumen sapi
2 Skema pengolahan tepung sagu dengan hasil sisa
ampas sagu (Dinas Pertanian Maluku 1984) ...
7
3 Monomer-monomer penyusun hemiselulosa ...
11
4 Struktur
selulosa
tanaman...
12
5 Tiga monomer utama penyusun lignin...
14
6 Struktur
lignin
tanaman...
15
7 Mekanisme degradasi lignin...
25
8 Diagram cara isolasi dengan kultur jaringan untuk memperoleh
bibit tanam ...
34
9 (a) ampas sagu sebelum fermentasi, (b) pembentukan miselium
selesai, (c) pertumbuhan tubuh buah, (d) ampas sagu hasil
fermentasi ...
35
10 (a) Miselium jamur tiram putih pada hari ke 50 pada media
tumbuh dengan pembesaran 1000x, (b) media tumbuh jamur
sebelum fermentasi dengan pembesaran 500x, (c) media tumbuh
jamur sesudah fermentasi dengan pembesaran 500x ...
51
1 Pembuatan Inokulum ……….. 94 2 Diagram alir fermentasi ampas sagu dengan
jamur tiram ... ... 96 3 Hasil sidik ragam dan uji Duncan bahan kering media
tumbuh jamur ... ... 97 4 Hasil sidik ragam dan uji Duncan kandungan bahan
kering (g) media tumbuh jamur……….. 98
5 Hasil sidik ragam dan uji Duncan bahan organik (%)
media tumbuh jamur... 99 6 Hasil sidik ragam dan uji Duncan kandungan bahan
organik (g) media tumbuh jamur... 100 7 Hasil sidik ragam dan uji Duncan kandungan protein (%)
media tumbuh jamur... 101 8 Hasil sidik ragam dan uji Duncan kandungan protein (g)
media tumbuh jamur... 102 9 Hasil sidik ragam dan uji Duncan NDF (%) media
tumbuh jamur………. 103
10 Hasil sidik ragam dan uji Duncan NDF (g) media tumbuh
jamur………... 104 11 Hasil sidik ragam dan uji Duncan NDF (g) media tumbuh
jamur………... 105 12 Hasil sidik ragam dan uji Duncan ADF (g) media tumbuh
jamur………... 106 13 Hasil sidik ragam dan uji Duncan Hemiselulosa (%)
media tumbuh jamur……….. 107
14 Hasil sidik ragam dan uji Duncan Hemiselulosa (g)
media tumbuh jamur……….. 108
15 Hasil sidik ragam dan uji Duncan Selulosa (%) media
tumbuh jamur………. 109
16 Hasil sidik ragam dan uji Duncan Selulosa (g) media
tumbuh jamur………. 110
17 Hasil sidik ragam dan uji Duncan Lignin (%) media
tumbuh jamur………... 111
18 Hasil sidik ragam dan uji Duncan Lignin (g) media
tumbuh jamur………. 112
19 Hasil sidik ragam dan uji Duncan pH cairan rumen in
vitro... 113 20 Hasil sidik ragam dan uji Duncan Amonia cairan rumen
in vitro……… 114
21 Hasil sidik ragam dan uji Duncan VFA total cairan
rumen in vitro... 115 22 Hasil sidik ragam dan uji Duncan KcBK in vitro... 116 23 Hasil sidik ragam dan uji Duncan KcBO in vitro... 117 24 Hasil sidik ragam dan uji Duncan konsumsi bahan kering
organik (%BB)... 119
26 Hasil sidik ragam dan uji Duncan pertambahan bobot badan harian (kg/ekor/hari) ... 120
27 Hasil sidik ragam dan uji Duncan konversi ransum... 121
28 Hasil sidik ragam dan uji Duncan Income over feed cost (IOFC) ……….. 122
29 Hasil sidik ragam dan uji Duncan Kecernaan bahan kering ... 123
30 Hasil sidik ragam dan uji Duncan Kecernaan bahan organik ... 124
31 Hasil sidik ragam dan uji Duncan Kecernaan Protein ………... 125
32 Hasil sidik ragam dan uji Duncan kecernaan NDF... 126
33 Hasil sidik ragam dan uji Duncan Kecernaan ADF……... 127
34 Hasil sidik ragam dan uji Duncan Kecernaan Selulosa ……… 128
35 Hasil sidik ragam dan uji Duncan konsumsi nitrogen ... 129
36 Hasil sidik ragam dan uji Duncan nitrogen feses ... 130
37 Hasil sidik ragam dan uji Duncan nitrogen urine... 131
38 Hasil sidik ragam dan uji Duncan retensi nitrogen ... 132
39 Hasil sidik ragam dan uji Duncan kolesterol feses ... 133
40 Hasil sidik ragam dan uji Duncan amonia cairan rumen sapi Bali ……… 134
41 Hasil sidik ragam dan uji Duncan VFA total cairan rumen sapi Bali ... 135
42 Hasil sidik ragam dan uji Duncan asam asetat cairan rumen sapi Bali ……….. 136
43 Hasil sidik ragam dan uji Duncan asam propionat cairan rumen sapi Bali ... 137
44 Hasil sidik ragam dan uji Duncan asam butirat cairan rumen sapi Bali ……….. 138
45 Hasil sidik ragam dan uji Duncan asam isobutirat cairan rumen sapi Bali ……….. 139
46 Hasil sidik ragam dan uji Duncan asam valerat cairan rumen sapi Bali ... 140 47 Hasil sidik ragam dan uji Duncan asam isovalerat cairan rumen sapi Bali ... 141
48 Hasil sidik ragam dan uji Duncan proporsi molar asam asetat cairan rumen sapi Bali ………. 142
49 Hasil sidik ragam dan uji Duncan proporsi molar asam propionat cairan rumen sapi Bali ……….. 143
valerat cairan rumen sapi Bali ... 146 53 Hasil sidik ragam dan uji Duncan proporsi molar asam
Peningkatan populasi dan produktivitas ternak ruminansia belum memperlihatkan hasil yang optimal. Sementara itu permintaan akan daging terus meningkat dari tahun ke tahun terutama pada hari-hari raya. Konsumsi daging sapi di Indonesia mengalami peningkatan, dari 330 300 ton pada tahun 2002 menjadi 389 300 ton pada tahun 2006. Sebaliknya populasi ternak sapi pedaging cenderung mengalami penurunan dari 11 297 625 ekor pada tahun 2002 menjadi 10 835 686 ekor pada tahun 2006 (DITJENAK 2006). Ketidakseimbangan ini disebabkan antara lain oleh produktivitas ternak yang sangat rendah. Hal ini perlu mendapat perhatian pemerintah karena akan berdampak negatif terhadap ketersediaan protein hewani.
Salah satu kendala yang dihadapi oleh usaha peternakan adalah belum tercukupinya kebutuhan nutrisi terutama protein pakan, sehingga ternak belum dapat tumbuh dan berkembang dengan baik. Rumput di daerah tropis kebanyakan bermutu rendah dengan serat kasar yang tinggi. Sementara itu penanaman rumput unggul seperti rumput gajah dan sebagainya juga mendapat kendala karena terbatasnya lahan, yang kebanyakan sudah digunakan untuk pemukiman dan lahan pertanian. Keadaan ini merupakan tantangan bagi sektor peternakan, karena perlu mencari pakan alternatif untuk meningkatkan produksi ternak.
Pemanfaatan sumber daya lokal secara optimal merupakan langkah strategis dalam upaya mencapai efisiensi usaha produksi ternak ruminansia di Indonesia. Hal ini akan semakin nyata, apabila sumberdaya tersebut bukan merupakan kebutuhan langsung bagi kompetitor, seperti manusia atau jenis ternak lain. Oleh karena pakan sangat erat kaitannya dengan produktivitas dan biaya produksi, maka pemanfaatan bahan baku lokal secara efisien akan berpengaruh nyata terhadap perkembangan ternak.
dalam jumlah yang besar, ada sepanjang tahun dan terkonsentrasi. Bahan baku yang mempunyai karakter tersebut umumnya terkait dengan industri, yang menghasilkan berbagai produk baik yang bersifat sampingan maupun limbah.
Bahan baku lokal untuk tiap daerah berbeda tergantung pada kondisi daerah tersebut. Maluku yang dikenal sebagai daerah produsen sagu mempunyai limbah dari industri pengolahan tepung sagu yang berlimpah. Sagu merupakan salah satu sumber daya nabati di Indonesia yang mulai akhir tahun tujuh puluhan makin meningkat pemanfaatannya, sebagai akibat dari program pemantapan swasembada pangan nasional dan permintaan akan bahan baku industri dan energi. Potensi sagu di Maluku cukup besar, walaupun pada beberapa wilayah telah terjadi pengalihan status pemanfaatan lahan sagu untuk pemanfaatan lain (Louhenapessy 1998).
Ampas sagu merupakan limbah yang didapatkan pada proses pengolahan tepung sagu, dimana dalam proses tersebut diperoleh tepung dan ampas sagu dalam perbandingan 1 : 6 (Rumalatu 1981). Jumlah limbah yang banyak tersebut, sampai saat ini belum dimanfaatkan sebagaimana mestinya hanya dibiarkan menumpuk pada tempat - tempat pengolahan tepung sagu sehingga menyebabkan pencemaran lingkungan. Kalaupun ada ternak yang memanfaatkannya, hanya ternak-ternak yang berada di sekitar lokasi pengolahan tepung sagu, yang langsung mengkonsumsi di tempat penumpukan ampas tanpa dikontrol.
Pemanfaatan limbah atau ampas sagu sebagai pakan alternatif merupakan suatu hal yang baik, walau disadari bahwa pemanfaatannya perlu mendapat sentuhan teknologi, karena ampas sagu mempunyai keterbatasan untuk digunakan sebagai pakan yaitu kandungan serat kasarnya tinggi dan proteinnya rendah. Oleh karena itu perlu dilakukan pengolahan pendahuluan sebelum diberikan kepada ternak. Cara pengolahan limbah yang sudah dikenal antara lain pengolahan fisik, kimia dan biologi.
et al. 2003), disamping budidayanya membutuhkan biaya relatif murah dan mampu memperbaiki pakan bermutu rendah menjadi pakan bermutu tinggi (Rai dan Saxena 1990)
Jamur tiram dalam biokonversi jerami padi diketahui mampu mendegradasi lignin, dan setelah dicobakan pada ternak ternyata dapat meningkatkan kecernaan bahan kering dan bahan organiknya (Jafari et al. 2007) sehingga apabila jamur tiram putih tersebut digunakan untuk ampas sagu diharapkan dapat memberi hasil yang baik pula. Disamping itu penerapan bioteknologi ini dapat bersifat dwiguna, yaitu dapat mengembangkan agribisnis budidaya jamur tiram putih (Pleurotus ostreatus). Diharapkan dengan biofermentasi ampas sagu dengan jamur tiram ini dapat memberikan dua keuntungan sekaligus yaitu disatu pihak berupa jamur tiram sebagai komoditi pangan bernilai ekonomi tinggi. Dilain pihak substrat media tumbuh jamur dapat digunakan sebagai pakan ruminansia, sehingga terbuka peluang untuk penggemukan sapi potong didaerah-daerah penghasil tepung sagu. Dengan demikian pemanfaatan ampas sagu dapat dioptimalkan, sehingga dapat meningkatkan pendapatan petani peternak, sekaligus mencegah pencemaran lingkungan.
Salah satu faktor penting yang mempengaruhi proses biodegradasi adalah adanya mineral dalam substrat, dimana mineral dapat mempengaruhi pertumbuhan fungi dan aktifitas enzim lignin peroksidase, mangan peroksidase dan lakase (Badrian et al. 2000). Penelitian yang dilakukan oleh Karem dan Hadar (1997) menunjukkan bahwa penambahan Mn kedalam substrat dapat meningkatkan degradasi lignin oleh Pleurotus ostreatus hingga 50% dan meningkatkan kecernaan bahan kering.
Perlakuan kimia juga sering digunakan untuk meningkatkan mutu pakan berserat yang berkualitas rendah. Amoniasi merupakan salah satu perlakuan kimia yang sering digunakan, karena urea yang ditambahkan pada pakan akan mengalami proses ureolitik menjadi NH3 dan CO2 oleh urease bakteri pakan.
bahwa perlakuan amoniasi pada jerami padi dengan menggunakan urea dapat meningkatkan kadar protein kasar, kecernaan bahan kering dan bahan organik secara in vitro.
1.2 Tujuan Penelitian
Rangkaian percobaan dalam penelitian bertujuan untuk :
1. Memanfaatkan sumber daya pakan lokal hasil samping pengolahan sagu. 2. Mengkaji potensi ampas sagu sebagai media tumbuh jamur tiram. 3. Mengkaji nilai nutrisi ampas sagu hasil biofermentasi dan amoniasi.
4. Mendapatkan formula ransum dengan bahan baku ampas sagu untuk produktivitas ternak ruminansia.
1.3 Manfaat Penelitian
Manfaat penelitian ini adalah :
1. Memberi informasi baru tentang pendayagunaan limbah atau ampas sagu rumbiah sebagai bahan pakan potensial.
2. Mencegah pencemaran lingkungan, akibat menumpuknya ampas sagu pada sentra-sentra industri pengolahan tepung sagu.
1.4 Hipotesis Penelitian
1. Ampas sagu dapat digunakan sebagai media tumbuh jamur.
2. Pengolahan biofermentasi dan amoniasi dapat meningkatkan penggunaan pakan lokal ampas sagu.
3. Pengolahan biofermentasi dan amoniasi dapat meningkatkan nilai nutrisi ampas sagu
Sagu termasuk tumbuhan monokotil dari famili Palmae, ordo
Spadiciflorae dan genus Metroxylon (Sastrapradja dan Mogen 1976). Tanaman
sagu terdiri atas sagu berduri dan sagu tidak berduri. Sagu berduri adalah sagu
Tuni ( M. rumpii), sagu Ihur (M. sylvestre), sagu Makanaru (M.longispinum) dan
sagu duri rotan (M. microcanthum) serta satu jenis sagu yang tidak berduri yaitu
sagu molat (M. sagu) (Bintoro 2008). Selanjutnya Papilaya (2009) menyatakan
bahwa kelima jenis sagu ini mempunyai nilai ekonomis yang tinggi di Maluku.
Tanaman sagu dapat tumbuh pada berbagai kondisi hidrologi dari yang
terendam sepanjang masa sampai ke lahan jalan yang tidak terendam air (Bintoro
2008). Bentuk pohon yang tegak dan kuat dengan ukuran tinggi dan diameter
batang yang berbeda-beda menurut jenis dan umurnya. Pohon sagu yang mulai
berbunga mempunyai tinggi yang bervariasi antara 10-15 m dan diameter
batangnya mencapai 75 cm dengan berat berkisar satu ton (Flach 1977).
Metroxylon berasal dari bahasa Yunani, yaitu Metra dan Xylon. Metra
artinya empulur dan Xylon artinya Xylem atau pembuluh kayu (Flach 1983). Di
Indonesia dikenal dengan beberapa nama diantaranya rumbia (Minangkabau,
Makassar dan Bugis), lapia atau napia (Ambon), kirai (Jawa Barat), sedangkan
Jawa Timur dan Jawa Tengah dikenal dengan nama bilung atau kresula.
Perbanyakan tanaman sagu dapat dilakukan dengan benih (biji sagu) untuk
pembibitan dengan cara generatif dan anakan untuk pembibitan vegetatif. Biji
atau buah yang digunakan berasal dari pohon sagu yang sudah tua atau
mengering, sedangkan anakan berasal dari tunas yang melekat pada pangkal
batang pohon induknya atau anakan yang sudah menjalar di atas permukaan tanah
(Papilaya 2009).
Secara kasar sagu dapat dibagi dalam dua golongan yaitu sagu yang
berbunga atau berbuah satu kali dan yang berbunga atau berbuah lebih dari satu
kali (Rumalatu 1981). Golongan sagu yang berbunga atau berbuah satu kali
mempunyai karbohidrat yang tinggi dibandingkan dengan sagu yang berbunga
umur antara 10-12 tahun pada waktu tinggi tanaman sudah mencapai 10-15
meter. Batang sagu banyak mengandung pati. Pamanenan pati sagu hendaknya
pada saat inisiasi pembentukan bunga. Saat pembentukan bunga, meskipun masih
terjadi akumulasi pati tetapi laju pati yang digunakan untuk pembuatan buah lebih
cepat daripada laju akumulasi pati. Pati yang terdapat pada batang bagian bawah
akan lebih dahulu digunakan untuk pertumbuhan bunga dan buah padahal
sebenarnya pati lebih banyak terdapat pada batang bagian bawah (Bintoro 2008).
Gambar 1 Pohon sagu spesies Metroxylon sago
Pati sagu diperoleh dengan cara memeras empulur batang tanaman sagu.
Skema pengolahan batang sagu menjadi tepung sagu dengan hasil ikutan ampas
sagu dapat dilihat pada Gambar 2.
2.2 Potensi Ampas Sagu Sebagai Komponen Pakan ternak
Schuling dan Flach (1993) memprediksi bahwa potensi luas areal sagu
31 690.4 ton dan tahun 1995 sebesar 36 198.2 ton (Biro pusat statistik 1995).
Menurut Louhenapessy (1998) potensi luasan sagu di Maluku adalah sekitar
47 600 ha. Luasan ini masih mungkin bertambah karena umumnya sagu tersebar
di seluruh wilayah Maluku walaupun dalam luasan yang sempit dan mungkin juga
berkurang karena terjadi konversi lahan sagu untuk kepentingan lainnya.
Batang Sagu
Pengupasan kulit dan pemotongan
Kulit batang --- Pemarutan
--- ditambah air
Peremasan
--- ditambah air
Penyaringan
Ampas sagu ---
Pengendapan
Air sisa ---
Pengeringan
Tepung sagu
Gambar 2 Diagram pengolahan tepung sagu dengan hasil sisa ampas sagu ().
Jumlah produksi tepung sagu di Maluku adalah 4 400 kg/ha. Dimana
rata-rata pohon masak tebang untuk hutan sagu di Maluku dari berbagai jenis sagu
adalah 20 pohon/ha dan rata-rata produksi tiap pohon 220 kg tepung sagu.
ampas sagu berpotensi dapat digunakan sebagai komponen pakan ternak. Di
Ambon atau Maluku ampas sagu disebut ela sagu.
Banyak penelitian telah dilakukan dalam melihat pemanfaatan ampas
sagu sebagai komponen pakan, baik dalam ransum ruminansia maupun
monogastrik. Pantjawidjaja et al. (1984) melaporkan bahwa substitusi rumput
lapangan dengan ampas sagu (Metroxylon sp) sampai pada level 45% dengan urea
3% dari bahan kering ampas sagu tidak memperlihatkan perbedaan yang nyata
dalam efisiensi penggunaan makanan dan pertambahan bobot badan sapi
peranakan ongole (PO), sedangkan menurut Nurkurnia (1989) penggunaan 40%
ampas sagu dalam ransum tidak mempengaruhi produksi VFA total atau parsial,
imbangan asetat/propionat dan produksi NH3.
Hangewa (1992) menyatakan bahwa dengan penggunaan
kompleks-NPN-karbohidrat yang terbuat dari urea dan ampas sagu dengan waktu pemasakan 116
menit dan dosis urea 5.4% dari bahan kering ampas sagu dicapai sintesis protein
yang optimal yaitu 890 mg/g/4jam. Penggunaan ampas sagu aren 50%
memberikan bobot badan akhir ayam broiler yang lebih baik dari penggunaan
50% ampas sagu kirai (Nawal 1995). Pada penelitian lain didapatkan bahwa
ampas sagu dapat digunakan dengan komposisi nutrisi yang seimbang sampai
taraf 12.5% pada ransum ayam pedaging dan pada ransum ayam kampung sampai
taraf 25% (Kompiang et al. 1995).
Hasil penelitian yang dilaporkan oleh Ralahalu (1998) menjelaskan bahwa
penggunaan ampas sagu hasil fermentasi dengan Aspergillus niger sampai taraf
15% dalam ransum ternak babi memberikan pertambahan bobot badan yang baik.
Biyatmoko (2002), menyatakan bahwa penggunaan ampas sagu fermentasi dalam
ransum itik alabio jantan hingga sekitar 10.6% ternyata mampu meningkatkan
aktifitas selulolitik tanpa menyebabkan kerusakan organ pencernaan itik.
2.3. Komposisi Kimia dan Komponen Ampas sagu
Dilihat dari segi kuantitas, ampas sagu cukup tersedia untuk digunakan
sebagai pakan ternak terutama pada daerah-daerah produsen tepung sagu seperti
Maluku dan Papua, tetapi dari segi kualitas, ampas sagu mempunyai nilai gizi
yang rendah karena kadar serat kasarnya yang tinggi dan kadar proteinnya yang
dilihat dalam Tabel 1, dari tabel tersebut terlihat bahwa ada perbedan komposisi
dari 2 penelitian. Hal ini mungkin disebabkan oleh adanya perbedaan spesies,
umur, tempat hidup dan proses pengolahannya.
Bila dibandingkan dengan komponen lain dari tanaman sagu, maka ampas
sagu merupakan komponen terbesar. Disayangkan dari jumlah yang besar
tersebut pemanfaatannya sebagai pakan (ruminansia sekalipun) dibatasi oleh
kadar seratnya yang relatif tinggi, tanpa mendapat perlakuan khusus terlebih
dahulu (Preston dan Leng 1987). Dari hasil analisa kimia (Tabel 1) terlihat
bahwa ampas sagu sebagian besar terdiri dari komponen serat.
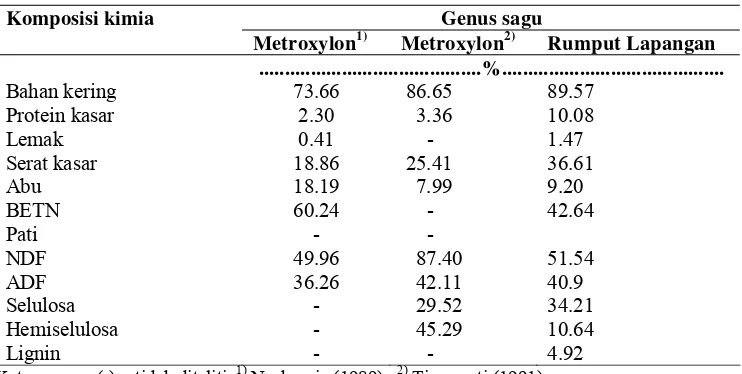
Tabel 1 Komposisi nutrisi ampas sagu dan rumput lapangan
Komposisi kimia Genus sagu
Metroxylon1) Metroxylon2) Rumput Lapangan
...%...
Bahan kering 73.66 86.65 89.57
Protein kasar 2.30 3.36 10.08
Lemak 0.41 - 1.47
Serat kasar 18.86 25.41 36.61
Abu 18.19 7.99 9.20
BETN 60.24 - 42.64
Pati - -
NDF 49.96 87.40 51.54
ADF 36.26 42.11 40.9
Selulosa - 29.52 34.21
Hemiselulosa - 45.29 10.64
Lignin - - 4.92
Keterangan : (-) = tidak diteliti, 1) Nurkurnia (1989), 2) Tisnowati (1991),
Pada umumnya sel tanaman digolongkan dalam dua golongan berdasarkan
kelarutannya dalam larutan deterjen yaitu : 1) isi sel (Neutral Detergent Solubles
[NDS]), merupakan bagian yang kecernaanya tinggi; terdiri dari gula-gula , pati,
pektin, protein, asam-asam organik; larut dalam larutan deterjen netral; dan 2)
dinding sel (Neutral Detergent Fiber [NDF]), merupakan bagian yang
kecernaannya rendah; terdiri dari hemiselulosa, selulosa, lignin, silika; tidak larut
dalam larutan deterjen netral (Van Soest 1982). Dinding sel tanaman dibentuk
dari serat selulosa yang terikat dengan karbohidrat non-selulolitik, hemiselulosa
dan pektin (metil ester dari asam poligalakturonat) serta beberapa komponen
asam-asam fenolat yang juga dapat berkontribusi dalam stabilitas struktur.
terlignifikasi dengan menggantikan pektin oleh deposisi lignin dan hemiselulosa
di lamela tengah dan dinding sel sekunder. Dengan demikian residu tanaman,
sebagaimana halnya ampas sagu tersusun atas matriks serat selulosa yang terdiri
dari lignin dan biopolimer hemiselulosa. Akibat dari sifat-sifat residu tanaman
tersebut, biodegradasinya dibatasi oleh ketiga polimer, yaitu selulosa, lignin dan
hemiselulosa (Paterson 1989). Pembagian bahan organik tanaman melalui sistem
analisis Van Soest disajikan pada Tabel 2.
Tabel 2 Pembagian bahan organik tanaman melalui sistem analisis deterjen
Fraksi Komponen Manfaat
1. Isi sel
(larut dalam detergen Netral)
Lemak, gula, asam-asam Organik dan bahan yang larut dalam air, pektin, pati, NPN, protein terlarut
Hampir seluruhnya dapat dicerna, tidak terikat oleh lignin
2. Dinding sel
(serat yang tidak larut Dalam deterjen netral) :
a. Larut dalam
deterjen asam
b. Tidak larut dalam
deterjen asam
Hemiselulosa, serat pengikat protein Selulosa, kitin, lignin, Silika, nitrogen yang terikat oleh lignin
Sebagian dapat dicerna Tergantung dari derajat lignifikasi
Sumber : Van Soest (1977)
Kandungan selulosa dan hemiselulosa yang cukup tinggi pada pakan serat
merupakan sumber energi untuk ruminansia, tetapi dari beberapa kajian
didapatkan kecernaannya rendah. Rendahnya kecernaan selulosa dan
hemiselu-losa disebabkan oleh keterikatannya dengan lignin dalam bentuk ikatan kompleks.
Ikatan fisik dan kimia ketiga komponen tersebut akan menghambat
mikroorganisme rumen dalam mencerna serat kasar (Winugroho 1986). Fraksi
hemiselulosa umumnya lebih mudah dicerna dari selulosa tetapi lignin praktis
tidak dapat dicerna karena lignin mempunyai proporsi karbon, hidrogen, dan
oksigen yang berbeda dari polisakarida lainnya (Morison 1980).
Hemiselulosa merupakan polimer kompleks yang terdiri atas campuran
berbagai polimer monosakarida yang berbeda dimana gula penyusunnya
dikelompokkan sebagai berikut : 1) heksosa (glukosa, manosa dan galaktosa);
(glukoronat, metilglukoronat, galaktoronat); 4) deoksiheksosa (ramnosa, fukosa)
(Fengel dan Wegener 1989). Monomer-monomer penyusun hemiselulosa dapat
dilihat pada Gambar 3. Komponen terbesar dari hemiselulosa adalah siloglukan
yang tersusun dari unit-unit glukosa dengan ikatan β-1-4 yang ujungnya
membentuk cabang dengan unit-unit silosa pada ikatan α-1-6. Molekul ini terikat
secara kovalen dengan fraksi pektin dinding sel serta ikatan hidrogen terhadap
mikrofibril selulosa sehingga memperkuat sel-sel tanaman.
Gambar 3 Monomer-monomer penyusun hemiselulosa
Gambar 3 Monomer-monomer penyusun hemiselulosa
Bila dibandingkan dengan selulosa, hemiselulosa lebih mudah didegradasi
menjadi gula sederhana dan produk lainnya serta lebih siap dicerna daripada
selulosa (Crowder dan Chheda 1982), sehingga seringkali disebut juga sebagai
karbohidrat yang larut dalam alkali lemah (Maynard et al. 1984). Tetapi menurut
Doyle et al. (1986), hemiselulosa berbeda dengan selulosa dalam hubungannya
dengan keterikatan dengan lignin. Hemiselulosa terikat lebih erat dengan lignin
dibandingkan selulosa. Pada kondisi ini selulosa lebih mudah dicerna
dibandingkan hemiselulosa. Dilaporkan juga bahwa kecernaan selulosa dan
hemiselulosa ini diakibatkan oleh kandungan lignin yang berubah-ubah.
Kandungan hemiselulosa di dalam sel tanaman berbeda tergantung pada spesies
tanaman, jenis jaringan dan tingkat perkembangan sel (Blanchette 1995). Lebih
lanjut dilaporkan bahwa sel primer tanaman mengandung 24% hemiselulosa, pada
Selulosa adalah unsur utama yang membungkus kerangka tumbuhan yang
dihasilkan dari proses fotosintesa tumbuh-tumbuhan (Harjo et al. 1989 ; Mayes et
al. 1995). Bentuk ini tidak larut dalam pelarut biasa dan terdiri dari sejumlah unit
β-D-glukopiranosa yang dihubungkan lewat ikatan β-1-4 untuk membentuk rantai lurus dan panjang yang dikuatkan oleh ikatan hidrogen berikatan silang yang
memberikan struktur kristalin dan tersusun dalam mikrofibril (Gambar 4).
Ada dua macam ikatan hidrogen pada struktur selulosa yaitu : 1) ikatan hidrogen
intra molekul yang mempertahankan kekuatan rantai selulosa; dan 2) ikatan inter
molekul yang menyebabkan rantai selulosa saling berikatan membentuk suatu
mikrofibril. Secara alami selulosa umumnya berstruktur kristalin. Hal ini
diketahui dengan analisis difraksi sinar-X, dan dengan analisis spektroskopi infra
merah menunjukkan adanya beberapa gugus hidroksil bebas yang saling berikatan
melalui ikatan hidrogen membentuk struktur kristalin dan berikatan dengan lignin
melalui ikatan kovalen membentuk lignoselulosa yang kuat sehingga sulit
dihidrolisis dengan enzim (Tsao et al . 1978).
Struktur kristal yang dibungkus oleh lignin yang berperan dalam pencegahan
terhadap serangan enzim pemecah selulosa. Komposisi kimia dan struktur yang
demikian mengakibatkan bahan mengandung selulosa menjadi kuat dan keras
(Browning 1967).
Molekul selulosa umumnya tidak ditemukan secara bebas pada sel
tanaman, tetapi dalam bentuk ikatan membentuk jalinan berdiameter 0,3 µ yang
disebut mikrofibril dan diameter > 0,3 µ disebut fibril (Clark dan Swan 1975).
Setiap mikrofibril tersusun oleh rantai selulosa berupa unit-unit glukosa.
Fibril-fibril tersebut membentuk ikatan lebih besar dengan ikatan hidrogen menjadi
struktur kristalin. Selulosa banyak terdapat pada roughage yaitu pada dinding sel
sekundernya. Kesempurnaan pemecahannya tergantung pada jenis ternak, yaitu
ada atau tidaknya enzim selulase.
Mayes et al. (2003), menyatakan bahwa pada hewan pemamahbiak dan
hewan herbivora terdapat mikroorganisme yang menghasilkan enzim selulase
yang dapat menyerang ikatan β sehingga selulosa dapat digunakan sebagai sumber
penghasil kalori yang penting. Selulosa dicerna secara mantap didalam rumen
dan retikulum ternak menjadi selobiosa dan selanjutnya oleh enzim selobiose
diubah menjadi glukosa. Hasil akhir pencernaan oleh jasad renik adalah asam
lemak terbang yang terdiri dari asam asetat, asam propionat dan asam butirat
dengan hasil sampingan berupa gas CO2 dan metana (Tillman et al. 1991).
Tanaman mengandung bagian yang tidak dapat dicerna. Analisis kimia
pakan telah menggolongkan bagian tersebut ke dalam kategori lignin kasar (Van
Soest 1977). Lignin adalah fraksi non-karbohidrat yang merupakan polimer
aromatik polifenolat yang terdiri atas unit-unit oksifenilpropana yang tergabung
dalam struktur ikatan silang yang kompleks, sehingga sangat sukar dihidrolisis
(McDonald et al. 2002). Pada dinding sel tanaman yang tua lignin berikatan
dengan hemiselulosa dan disebut glikolignin atau lignohemiselulosa.
Pentosan-pentosan seperti silosa, arabinosa, asam uronat, glukosa dan galaktosa adalah
hemiselulosa yang selalu terikat dengan lignin membentuk kompleks
lignin-karbohidrat (LCC) (Paterson 1989).
Wahyu (1995), menyatakan bahwa lignin sebenarnya bukan karbohidrat
struktur selulosa-hemiselulosa dan jaringan tanaman selama pertumbuhannnya
untuk mencegah serangan bakteri. Dari segi nutrisi selalu dihubungkan dengan
selulosa dan hemiselulosa. Jumlah lignin dan penempatannya tidak bermanfaat
sebagai zat makanan bahkan mempunyai efek yang merugikan terutama dalam hal
ketersediaan zat makanan untuk diabsorbsi.
Lignin merupakan bentuk polimer dari senyawa aromatik yang berfungsi
memberi kekuatan dan kekakuan pada struktur tanaman. Karena adanya
hubungan kedua fungsi ini maka lignin bertindak sebagai penghalang fisik dan
penghalang pemecahan oleh mikroba dari senyawa-senyawa polisakarida tersebut
(Jackson 1978). Pendapat ini didukung oleh Liyama (2000) yang menyatakan
bahwa lignin merupakan biopolimer yang lain seperti polisakarida dan protein.
Selain itu lignin berikatan dengan selulosa dan hemiselulosa dalam
jaringan tanaman, dan lignin tidak pernah ditemui dalam bentuk sederhana
diantara polisakarida-polisakarida dinding sel tetapi selalu berikatan dengan
polisakarida lainnya (Fengel dan Wegener 1989). Lignin disusun dari unit-unit
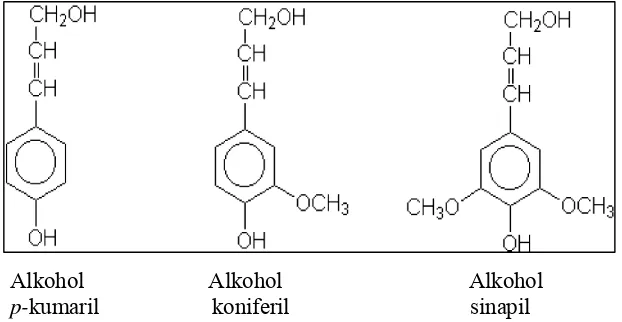
fenilpropen yaitu koniferil alkohol, sinapil alkohol dan p-kumaril alkohol melalui
proses polimerisasi dehidrogenasi, (Nolan et al. 1989). Gambar 5 dan 6
menjelaskan monomer-monomer penyusun lignin (fenilpropana) serta struktur
lignin yang berasal dari tanaman konifer.
[image:31.612.153.464.453.614.2]
Alkohol Alkohol Alkohol p-kumaril koniferil sinapil
Gambar 5 Tiga monomer utama penyusun lignin (Crawford 1981)
Pada tanaman lignin bervariasi dari 2-15% (Paturau 1982). Lignin
yang lebih tinggi daripada yang terdapat pada karbohidrat. Struktur khas lignin
dapat dijelaskan dengan baik karena bentuknya yang sangat variatif dari satu
tanaman ke tanaman lainnya, sehingga dapat membentuk ikatan atom C - C dan
C-O-C. Pada monomernya mempunyai gugus hidroksil (OH) dan metoksi
(OCH3) dan secara umum polimer amorf tidak membentuk kristal seperti pada
selulosa (Maynard et al. 1980). Struktur kompleks lignin mengandung
rantai-rantai karbon dan ikatan-ikatan eter yang tahan terhadap asam dan alkali.
Dilihat dari struktur dasar unit-unit monomernya, lignin digolongkan pada
tiga kelompok utama yaitu : 1) lignin pada Gymnospermae terdiri dari 8% polimer
alkohol koniferil, 14% alkohol p-kumaril dan 6% alkohol sinapil alkohol; 2)
lignin pada Angiospermae tersusun dari campuran 56% polimer alkohol koniferil,
40% alkohol sinapil dan 4% alkohol p-kumaril; dan 3) lignin dari Gramineae
merupakan esterifikasi p-asam kumarat menjadi Cŷ-gugus hidroksil pada gugus
samping polimer lignin yang terdiri dari koniferil, sinapil dan p-kumaril
(McDonald et al. 2002).
Gambar 6 Struktur lignin tanaman (Crawford 1981)
Ikatan rangkap antara lignin dengan selulosa dan hemiselulosa merupakan
dengan lignin adalah pentosan-pentosan seperti silosa, arabinosa, asam uronat,
glukosa dan galaktosa yang akan membentuk lignohemiselulosa (Harkin 1973).
Menurut Liyama (2000) bahwa selama masa pemasakan tanaman lignin
akan bertambah secara berangsur-angsur dan kecernaan dinding sel secara cepat
akan menurun. Penurunan kecernaan dinding sel ini ditentukan oleh deposisi
lignin. Lignin memiliki kandungan energi yang potensial, tetapi tidak dapat
digunakan sebagai sumber energi oleh mikroorganisme rumen terutama pada
cincin aromatiknya. Cincin aromatik hanya dapat dipecah pada keadaan aerob,
sedangkan rumen dalam keadaan yang anaerob (Orpin 1984).
Lignin terdiri dari sebuah struktur padat dengan ikatan karbon ke karbon
dan ikatan-ikatan lainnya yang resisten terhadap hidrolisis asam atau alkali.
Ikatan-ikatan ini berada dalam bentuk tiga dimensi dan merupakan jawaban
mengapa lignin begitu sukar untuk dihidrolisis, meskipun demikian seperti
senyawa polifenol lainnya lignin labil terhadap oksidasi (Van Soest 1977).
Lignin mengurangi kecernaan karbohidrat melalui ikatan hidrogen pada
sisi kritis, sehingga membatasi aktivitas selulase (Arora 1995). Van Soest (1977)
menjelaskan keterikatan lignin dengan substansi bukan lignin terjadi karena tiga
hal yaitu : 1) ikatan antara komponen bukan lignin dengan lignin dimana R dapat
berupa selulosa atau kelompok amino protein (ikatan benzil ini sukar dihidrolisis
dengan alkali); 2) ikatan antara komponen bukan lignin dengan lignin dimana X
menunjukkan sebuah ester silan (Wilkie 1979; Hartley 1981); dan 3) sebuah ester
fenolitik dengan sebuah silan (komponen hemiselulosa).
Houghton et al. (1987) menyatakan bahwa perombakan lignin oleh
mikroorganisme melibatkan enzim ligninolitik yang menguraikan lignin menjadi
karbondioksida yaitu peroksidase, mangan peroksidase, lakase dan oksidase. Laju
perombakan lignin akan meningkat seiring dengan meningkatnya hidrogen
peroksida yang berfungsi sebagai oksidan ekstraseluler dan perangsang aktivitas
lignilisis. Hal serupa juga dikemukakan oleh Leisola dan Garsia (1989) bahwa
degradasi lignin masih mungkin terjadi akibat proses jamur.
2.4 Pengolahan Pakan Berserat
Mikroba rumen sangat sulit mencerna pakan dengan serat kasar yang
untuk meningkatkan asas manfaat dari bahan-bahan pakan berserat maka perlu
dilakukan perlakuan terlebih dahulu. Tujuan perlakuan bahan pakan adalah untuk
menurunkan ukuran partikel sehingga meningkatkan luas permukaan jaringan
bahan agar terbuka terhadap serangan mikroorganisme rumen (Winugroho 1986).
Terdapat beberapa proses perlakuan yang telah diketahui dapat
memperbaiki nilai nutrisi pakan dengan kandungan serat yang tinggi yaitu secara
fisik, kimia dan biologis (Ryu 1989). Beberapa hasil penelitian dengan berbagai
metode pengolahan pakan berserat terhadap perbaikan nilai nutrisinya disajikan
pada Tabel 3. Kecernaan bahan kering dengan perlakuan fisik lebih rendah
[image:34.612.137.506.334.396.2]dibandingkan perlakuan kimia dan biologis.
Tabel 3 Pengaruh metode pengolahan terhadap konsumsi pakan dan perbaikan kecernaan bahan kering.
Perlakuan
Rata-rata perbaikan (%)
Konsumsi pakan Kecernaan bahan kering
1. Fisik (tekanan uap) 32 31
2. Kimia (alkali) 31 36
3. Biologis (biokonversi) 41 36
Sumber : Ryu (1989)
2.4.1. Pengolahan Pakan Serat Secara Fisik.
Pengolahan secara fisik banyak macamnya, anatara lain : pencacahan,
penggilingan, peleting, pembasahan, dan pemasakan (sterilisasi), pemanasan
(tekanan uap). Pencincangan 1-3 cm dapat meningkatkan konsumsi. Penggilingan
(ukuran 0.1-0.3 cm), peleting, pemasakan (sterilisasi), pemanasan di bawah
tekanan, pemanasan dengan sinar radiasi gamma 25-50 M, tekanan uap 21.1
kg/cm2 selama 10-30 detik dan sinar X bertujuan untuk memecah ikatan lignin
dan merubah sifat fisiko-kimia dinding sel, memperkecil ukuran, memperluas
ukuran partikel, meningkatkan akseptabilitas terhadap selulase, meningkatkan
pembengkakan partikel, menurunkan derajat polimerisasi, tetapi tidak merubah
komposisi kimianya (Doyle et al. 1986; Ryu 1989). Rusaknya dinding sel
menyebabkan aktivitas mikroba meningkat (Winugroho 1986; Usri 1987).
penggilingan akan mengurangi bahan organik dan sebagai konsekuensinya protein
kasar dan mineral meningkat (Komar 1984).
Pakan yang bentuk fisiknya lebih halus dapat meningkatkan konsumsi,
efisiensi, pertambahan berat badan dan kecernaan (Anggorodi 1979). Hangewa
(1992) melaporkan bahwa pemasakan urea-ampas sagu dengan tekanan uap dapat
meningkatkan kecernaan bahan kering, bahan organik, konsentrasi total dan
individual VFA serta konsentrasi amonia secara in vitro.
2.4.2. Pengolahan Pakan Serat Secara Kimiawi
Pengolahan secara kimiawi terhadap pakan telah banyak diungkapkan para
peneliti, dan nyata dapat memperbaiki kualitas pakan, walaupun ada yang
berpendapat bahwa residu zat kimia yang digunakan dapat membahayakan.
Kecernaan dan intake beberapa komponen dinding sel dapat meningkat dengan
perlakuan kimia. Bahan-bahan yang digunakan untuk meningkatkan degradasi
dinding sel dapat diklasifikasikan dalam tiga kelompok yaitu alkali, asam, reagen
oksidatif (NaOH, urine sapi, urea dan amonia encer) (Doyle et al 1986; Ryu
1989). Bahan kimia yang sangat efisien digunakan untuk pengolahan pakan serat
adalah alkali (NaOH, KOH, Ca[OH]2), amoniak anhidrase (NH3), larutan amoniak
(NH4OH)(Komar 1984). Berbagai perlakuan terhadap lignin juga telah
dikembangkan untuk mengkaji efek pengolahan terhadap peningkatan nilai nutrisi
[image:35.612.133.508.503.621.2]yang disajikan pada Tabel 4.
Tabel 4 Perlakuan kimiawi untuk meningkatkan mutu bahan pakan lokal
Perlakuan Agen perlakuan Target perlakuan Substrat perlakuan
Hidrolitik NaOH Urea
H2SO4
Alkali H2O2
Lignin-Karbohidrat Lignin-Karbohidrat Selulosa-selulosa Modifikasi fenolik Lignin
Jerami, pucuk tebu, limbah tanaman, kulit buah kakao
Oksidatif SO2
O3
Modifikasi fenolik lignin
Modifikasi fenolik lignin
Jerami,
Limbah tanaman Sawit
Sumber : Chesson (1993)
Hasil penelitian Usri (1987), menunjukkan bahwa jerami padi yang diolah
dengan NaOH dan abu sekam dapat meningkatkan konsumsi protein ransum dan
kualitas karkas, walaupun tidak nyata dalam pertambahan bobot badan dan
KOH dan NaOH] dapat melemahkan atau memutuskan ikatan lignoselulosa
dinding sel pakan, sehingga memudahkan penetrasi enzim mikroba dalam rumen
yang akhirnya kecernaan pakan dapat ditingkatkan (Komar 1984).
Terjadi perubahan pada ikatan antar lignin, antar lignin-karbohidrat atau
antar karbohidrat dengan perlakuan kimiawi yang bersifat hidrolitik. Perlakuan
yang bersifat oksidatif menghasilkan perubahan pada komposisi fenolik penyusun
rantai polimer lignin (Chesson 1993). Pengaruh mekanisme oksidatif terhadap
kualitas nutrisi bahan pakan lebih tinggi dibandingkan dengan cara hidrolitik,
namun penggunaan di lapangan sangat terbatas akibat pertimbangan ekonomi.
2.4.3. Pengolahan Pakan Serat Secara Biologis
Prinsip perlakuan biologis adalah pemutusan ikatan kompleks
selulosa-lignin dengan cara mengekstraksi atau mendekomposisi selulosa-lignin. Menurut Zadrazil
dan Kurtzman (1984), jerami, kayu serta limbah pertanian lainnya pada umumnya
akan mengalami mineralisasi setelah dilakukan pengolahan secara biologis
dengan menggunakan jamur. Beberapa penelitian melaporkan bahwa perlakuan
secara biologis nampaknya memberikan harapan yang lebih baik dibandingkan
perlakuan secara fisik dan kimia. Ryu (1989) meringkas beberapa laporan
penelitian mengenai pengaruh berbagai perlakuan terhadap perbaikan nilai nutrisi
bahan pakan, ternyata berdasarkan kecernaan maupun konsumsi bahan kering,
perlakuan biologis lebih baik dari perlakuan fisik maupun kimia.
Dekomposisi lignin secara biologis merupakan salah satu cara untuk
memecah ikatan selulosa-lignin dalam jaringan tanaman limbah pertanian,
sehingga dapat meningkatkan energi tersedia bagi ternak ketika digunakan sebagai
pakan. Mikroba yang ideal untuk mendegradasi lignin adalah yang memiliki
potensi kuat untuk mendegradasi selulosa dan hemiselulosa. Selama proses
dekomposisi lignin, jamur juga akan mendegradasi karbohidrat (selulosa),
sehingga waktu panen merupakan faktor kritis dalam mengoptimalkan manfaat
dekomposisi lignin untuk meningkatkan kecernaan bahan. Tuomela (2002),
menyatakan bahwa jamur akar putih merupakan salah satu jenis mikroba pengurai
lignin yang paling efisien yang tersedia di alam dan mempunyai kemampuan
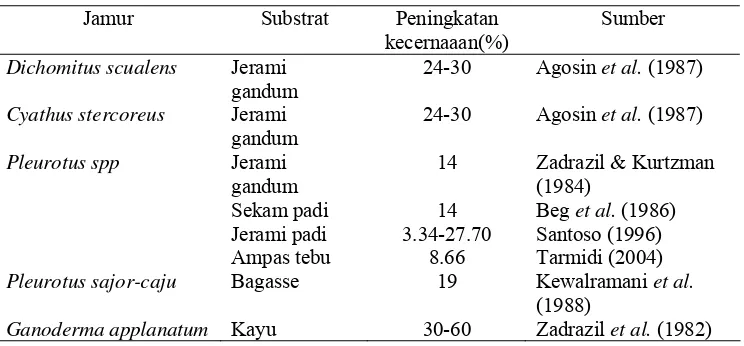
untuk mengurai lignin. Pada Tabel 5 disajikan beberapa jenis jamur yang telah
Tabel 5 Beberapa jenis jamur yang dapat digunakan untuk mendegradasi lignin
Jamur Substrat Peningkatan
kecernaaan(%)
Sumber
Dichomitus scualens Jerami gandum
24-30 Agosin et al. (1987)
Cyathus stercoreus Jerami gandum
24-30 Agosin et al. (1987)
Pleurotus spp Jerami gandum
14 Zadrazil & Kurtzman
(1984) Sekam padi
Jerami padi Ampas tebu
14 3.34-27.70
8.66
Beg et al. (1986) Santoso (1996) Tarmidi (2004)
Pleurotus sajor-caju Bagasse 19 Kewalramani et al.
(1988)
Ganoderma applanatum Kayu 30-60 Zadrazil et al. (1982)
Beberapa organisme selulolitik dapat menghasilkan protein sel tunggal
yang berasal dari substrat limbah pertanian. Dilaporkan bahwa ampas tebu yang
dikonversi dengan bakteri dapat menghasilkan biomassa protein sel tunggal
sebesar 26% (Linko 1977). Badve et al. (1987) melaporkan bahwa kandungan
dinding sel ampas tebu yang dibiofermentasi dengan jamur Pleurotus sajor-caju
nilainya menurun dari 88.4% menjadi 77.2% dengan bertambahnya waktu
inkubasi. Selanjutnya Tarmidi (1999) melaporkan bahwa Nilai kecernaan
komponen serat dan TDN ampas tebu hasil biokonversi jamur tiram putih lebih
tinggi dibandingkan dengan ampas tebu tanpa biokonversi.
Ralahalu (1998) menyatakan bahwa penggunaan ampas sagu yang
difermentasi dengan Aspergillus niger dapat meningkatkan pertambahan bobot
badan ternak babi, sedangkan menurut Biyatmoko (2002), fermentasi ampas sagu
dengan Aspergillus niger dapat memperbaiki nilai nutrisi ampas sagu terutama
meningkatkan protein murni dari 2.16 menjadi 9.55%. Proses fermentasi juga
terbukti dapat meningkatkan nilai gizi bahan asalnya karena selain terjadi
perombakan bahan kompleks menjadi lebih sederhana, didalam proses fermentasi
juga terbentuk beberapa vitamin misalnya riboflavin, vitamin B12 dan provitamin
A (Santoso 1987).
Winarno (1992) menyatakan bahwa substrat yang mengalami fermentasi
dikarenakan sifat katabolik dan anabolik mikroorganisme sehingga mampu
memecah komponen yang lebih kompleks menjadi mudah tercerna. Proses
biofermentasi diharapkan akan merombak struktur jaringan kimia dinding sel,
pemutusan ikatan lignosellulosa dan penurunan kadar lignin.
2.5 Jamur Tiram Putih (Pleurotus ostreatus) dan Peranannya dalam Perbaikan Mutu Pakan
Telah dijelaskan sebelumnya bahwa perbaikan mutu pakan dapat
dilakukan dengan bantuan mikroba dalam proses fermentasi. Keberhasilannya
ditentukan oleh jenis mikroba yang digunakan dalam metode fermentasi.
Umumnya jenis mikroba yang digunakan adalah yang mempunyai aktifitas
selulolitik yang tinggi serta mampu meningkatkan kadar protein bahan.
Sedangkan proses fermentasi yang banyak digunakan adalah fermentasi padat atau
semi padat.
Ada tiga jenis jamur yang dapat mendegradasi komponen serat (lignin,
selulosa dan hemiselulosa), yaitu jamur pembusuk putih (white-rot fungi), jamur
pembusuk coklat (brown-rot fungi), dan jamur pelapuk lunak (soft-rot fungi).
Dari ketiga jenis jamur tersebut, jamur pembusuk putih, yang sebagian besar
termasuk kelas Basidiomycetes, lebih efektif dalam mendegradasi lignin
(Crawford 1981).
Pleurotus ostreatus atau jamur pembusuk putih merupakan jamur genus
Pleurotus, famili Agaricaceae (Tricholomata), ordo Agaricales, subdivisi
Basidiomycotae, divisi Amastigomycota. Jamur ini biasanya disebut jamur tiram
atau jamur mutiara. Budidayanya relatif mudah dan tidak memerlukan lahan yang
luas, serta tidak membutuhkan modal yang besar. Secara alami jamur ini tumbuh
dan berkembang pada kayu yang sudah lapuk dengan kandungan nutrisi dan
mineral yang sedikit. Bahan-bahan seperti bongkol jagung, jerami, merang, serbuk
gergajian, kayu dan sisa pemintalan kapas dapat dijadikan substrat
pertumbuhannya (Lukitasari 2003). Selanjutnya dikatakan bahwa jamur tiram
putih dapat tumbuh pada kayu karena dapat menguraikan lignin yang terdapat
pada substrat yang terikat dengan polisakarida pada dinding sel kedua lamella
intraselluler.
nutrisi yang sesuai. Suhu yang dikehendaki berkisar antara 20-30 oC, namun suhu
optimum untuk pertumbuhan miselium adalah 22 oC Kelembaban relatif tidak
begitu penting bagi pertumbuhan miselium selama substrat difermentasikan
didalam kantong plastik, karena kelembaban minimum sebesar 80% dapat dicapai
apabila difermentasikan dalam kantong plastik dan cukup untuk menjaga
permukaan substrat dari kekeringan (Tripathi & Yadav 1992).
Selanjutnya Tripathi dan Yadav (1992) menyatakan bahwa faktor-faktor
yang saling berhubungan terhadap pertumbuhan miselium antara lain ukuran
partikel dan kadar air substrat. Besarnya ruang udara antara partikel-partikel
substrat akan mengganggu pertumbuhan miselium. Oleh sebab itu untuk limbah
pertanian ukuran partikel substrat diusahakan 2-3 cm, dan kadar air sekitar 75%
merupakan kondisi ideal bagi budidaya jamur tiram putih.
Pleurotus ostreatus dapat diidentifikasi dengan memperhatikan ciri-cirinya
yaitu pertumbuhan miseliumnya cepat dan berkolonisasi tinggi. Miselium yang
berumur 30-40 hari warnanya seputih kapas, padat, kompak dan mengeluarkan
enzim kedalam substrat. Lapisan miselium pada media yang akan membentuk
tubuh buah menunjukkan konsistensi seperti lemak. Tubuh buah dimulai dari
bintil-bintil bakal tubuh buah, mula-mula membentuk batang, kemudian ujungnya
membulat (Chang & Miles 1989).
Untuk meningkatkan aktifitas beberapa enzim yang dikeluarkan oleh
miselium jamur tiram putih diperlukan beberapa mineral tambahan seperti
potasium, sodium, fosfor, kalsium, magnesium dan mangan (Chang & Miles
1989; Kerem et al. 1995). Mineral-mineral tersebut juga penting bagi stabilitas
pH substrat dimana pH substrat yang optimum untuk pertumbuhan miselium
adalah 5.5 (Suriawiria 1997). Menurut Tripathi & Yadav (1992) gipsum juga
dibutuhkan untuk meningkatkan struktur substrat yang lebih bergranular sehingga
dapat mengikat air lebih baik untuk keperluan pertumbuhan miselium.
Beberapa jenis vitamin yang essensial diperlukan untuk pertumbuhan
jamur tiram putih diantaranya thiamin (vitamin B1), biotin (vitamin H atau
vitamin B7), dan vitamin B kompleks. Sumber-sumber vitamin tersebut dapat
Jamur tiram putih mensekresikan enzim-enzim ekstraseluler dan
intraseluler yang berperan dalam degradasi lignin, selulosa dan hemiselulosa,
terutama enzim-enzim endoglukonase, silanase, fenoloksidase yang terdiri atas
lakase dan peroksidase, enzim aril alkohol oksidase, Mn-oksidase, aril alkohol
dehidrogenase yang sebelumnya dikenal sebagai aril aldehida reduktase, dan
veratril alkohol oksidase (Sannia et al. 1991; Kerem et al. 1992).
Kerusakan substrat dimulai pada saat disekresikannya enzim yang dapat
mengubah substansi dalam bahan berselulosa yang tidak larut menjadi bentuk
yang larut. Akibatnya miselium jamur akan terpenetrasi ke dalam dinding sel
melalui lubang-lubang kecil yang terbentuk. Lignin dapat didegradasi tanpa
terjadinya kehilangan selulosa, tetapi secara simultan hemiselulosa juga akan
didegradasi (Crawford 1981).
Lignoselulosa terutama lignin sukar dihidrolisis baik oleh asam maupun
enzim disebabkan : 1) struktur kristalin selulosa; 2) asosiasinya dengan molekul
lignin dan hemiselulosa yang membuat sisi serang enzim menjadi terbatas. Kedua
hambatan tersebut juga akan mengurangi penetrasi enzim ke dalam
molekul-molekul selulosa, sehingga enzim tidak dapat menghidrolisis selulosa dengan
sempurna (Marsden & Gray 1986). Beberapa jenis jamur penghasil enzim
selulolitik dapat mendegradasi senyawa selulosa pada daerah kristalin. Rantai
selulosa mengikat air dan mengembang sehingga mudah didegradasi oleh selulase
yang dihasilkan oleh jamur atau fungi. Jamur tiram putih juga menghasilkan
enzim hidrolitik dan enzim oksidatif untuk mendegradasi selulosa. Selain itu juga
dihasilkan enzim selobiose quinon oksidoreduktase (Hollaender 1981).
Degradasi selulosa secara enzimatis terjadi karena adanya selulase sebagai
agen perombak bersifat spesifik untuk menghidrolisis ikatan β-1,4-glikosidik dari
rantai selulosa dan derivatnya. Enzim selulase kompleks umumnya terdiri dari
tiga unit yaitu : 1) endo-β-1,4 glukanase (Cx), berperan secara acak menghidrolisis
ikatan glikosida-β-1,4 sepanjang rantai selulosa. Enzim ini tidak menghidrolisis
selobiosa, tetapi menghidrolisis selodekstrin yang telah direnggangkan oleh asam
fosfat. Terbukanya ujung terminal selulosa memberi kesempatan kepada ekso-β-
1,4 glukanase mereduksi ujung rantai selulosa non-pereduksi untuk menghasilkan
pemecahan selodekstrin yaitu selulosa yang telah direnggangkan oleh asam fosfat.
Enzim ini mereduksi ujung rantai selulosa non-pereduksi dan melepaskan satu
unit selobiosa. Enzim C1 bekerja pada daerah kristalin dari serat, tidak
menghidrolisis selobiosa dan selulosa yang tersubstitusi, tetapi dapat mereduksi
selodekstrin; 3) β-1,4 glukosidase menurunkan unit enzim yang penting untuk
mereduksi selobiosa dan selodekstrin menghasilkan produk glukosa serta asam
selobionat menjadi glukosa dan glukanolakton (Hollaender 1981).
Hemiselulosa didegradasi oleh enzim: 1) silanase, merupakan kelompok
enzim endo- dan ekso- β-1,4-D-silanase yang menyerang rantai silan secara acak,
menyebabkan turunnya derajat polimerisasi dari substrat. Hasil utamanya adalah
silosa, silobiosa termasuk oligomer silosa dan L-arabinosa; 2) β-silosidase,
mereduksi silooligosakarida serta mengeluarkannya dari satu ujung rantai polimer
menjadi silosa; 3) α-glukonase, dibutuhkan untuk memecahkan terminal 4-0-asam
metil-glikoronik rantai sisi menyebabkan oligomer mudah direduksi oleh β
-silosidase; 4) mannase, mereduksi rantai β-1,4-D-mannapiranosil dan manna; 5)
esterase, merupakan asetil silan esterase yang membebaskan kelompok O-asetil
dari posisi C2 dan C3 pada silosa di dalam silooligomer (Puls & Poutanen 1981).
Degradasi hemiselulosa seperti halnya pada selulosa dan pati, yaitu dengan
memutuskan ikatan kimia diantara gugus gula dan menghasilkan silosa, arabinosa
dan glukosa ( Linko 1977).
Pada prinsipnya degradasi lignin oleh jamur pembusuk putih terdiri dari
tiga proses utama yaitu : 1) oksidasi rantai samping dengan membebaskan cincin
aromatik, terutama asam vanilat; 2) oksidasi karbon-α pada rantai samping
fenilpropana; dan 3) pemutusan cincin aromatik yang terikat pada polimer.
Ketiga proses tersebut dilakukan oleh enzim-enzim fenoloksidase, yaitu enzim
peroksidase dan lakase, serta enzim aril alkohol oksidase (AAO) (Kerem et al.
1992)
Putusnya ikatan lignin kristalin selulosa (dalam bentuk ikatan ester)
dengan koniferil, sinapil dan p-kumaril alkohol memudahkan penetrasi selulase
yang dihasilkan mikroba rumen. Terputusnya ikatan tersebut ditandai dengan
meningkatnya kelarutan masing-masing komponen serat (hemiselulosa, selulosa,
kecernaan in vitro komponen serat pakan meningkat (Maynard et al. 1980).
Mekanisme degradasi lignin dijelaskan pada Gambar 7.
Gambar 7 Mekanisme degradasi lignin (Gutierrez et al. 1996)
Menurut Nout (1995) substrat bekas dari penanaman jamur tiram putih
pada media jerami gandum setelah 12 minggu biokonversi dapat menurunkan
lignin sekitar separuhnya, dan dapat meningkatkan kecernaan. Pada penelitian
lain yang menggunakan substrat jerami gandum, media bekas penanaman jamur
tiram putih dapat meningkatkan kecernaan nitrogen sebesar 84.4% dan substansi
organik sebesar 74.4%. Disamping itu miselium jamur tersebut dapat menyum-
bangkan asam-asam amino yang lengkap, kecuali fenilalanin dan metionin yang
agak rendah. Media bekas penanaman jamur tiram putih menunjukkan
peningkatan kandungan protein kasar sebesar 22.4%, lemak 0.06%, dan abu 4.7%.
Peningkatan yang besar pada protein tersebut akibat meningkatnya kandungan
asam-asam amino (Sova & Cibuka 1990).
Penelitian yang dilakukan oleh Adamovic et al. (1998) menunjukkan
bahwa jerami yang didegradasi oleh enzim-enzim yang diproduksi selama
produksi jamur dapat lebih mudah dicerna oleh ruminansia, sehingga
meningkatkan bobot badan, konsumsi bahan kering dan efisiensi penggunaan
sedikit selulosa dan lignin, dengan peningkatan kandungan mineral dibandingkan
dengan jerami sebelum perlakuan dengan jamur.
2.6 Peranan Amoniasi terhadap Perbaikan Mutu Pakan
Perlakuan amoniasi hijauan pakan, baik berupa rumput maupun jerami
dapat meningkatkan kandungan protein kasar sekitar 6-8% (Yulistiani et al. 2003;
Weiss dan Underwood 2002; Stewart dan Silcox 2001). Selanjutnya Weiss dan
Underwood 2002 menyatakan bahwa amoniasi dapat meningkatkan kecernaan
bahan organik (KcBO) in vitro 20-30% dan konsumsi bahan kering 15-20%.
Pemberian hijauan hasil amoniasi pada sapi jantan dapat meningkatkan
pertambahan bobot hidup harian (PBHH) antara 272–544 gram (Brown 1991).
Amoniasi hijauan pakan dengan menggunakan urea, amonia padat,
maupun amonia gas memperlihatkan hasil yang tidak berbeda nyata antar
perlakuan (Joy et al. 1992; Hadjipanayiotou et al. 1993). Hasil penelitian
MacDearmid et al. (1988) memperlihatkan nitrogen yang teretensi dalam jaringan
hijauan pakan lebih tinggi pada amoniasi dengan urea daripada dengan amonia
padat. Urea dapat memperbaiki nilai nutritif beragam hijauan, meskipun
keefektifannya tergantung pada kandungan bahan kering dari pakan yang diberi
perlakuan urea tersebut (Mason et al. 1990; Zaman et al. 1993).
Penurunan kandungan NDF (Neutral Detergent Fibre) hijauan diduga
akibat rusaknya hemiselulosa oleh perlakuan amoniasi yang merusak ikatan kimia
antara lignin dan hemiselulosa (Weiss & Underwood 2002). Lignin menghambat
pencernaan serat, karena itu rusaknya ikatan menyebabkan hemiselulosa dan
selul