STRUKTUR KOMUNITAS PARASITOID TELUR
PADA PERTANAMAN KEDELAI DAN IMPLIKASINYA
TERHADAP TINGKAT SERANGAN DAN POPULASI HAMA
Oleh :
Okti Aryani Hapsari
A44101059
PROGRAM STUDI HAMA DAN PENYAKIT TUMBUHAN
FAKULTAS PERTANIAN
ABSTRAK
OKTI ARYANI HAPSARI. Struktur Komunitas Parasitoid Telur pada Pertanaman Kedelai dan Implikasinya terhadap Tingkat Serangan dan Populasi Hama. Dibimbing oleh PUDJIANTO.
Penelitian ini bertujuan untuk mengetahui jenis-jenis parasitoid telur yang terdapat pada pertanaman kedelai serta me mpelajari hubungannya dengan serangan hama. Penelitian dilaksanakan pada lahan pertanaman kedelai di Cikabayan, Bogor dan di Laboratorium Bioekologi Parasitoid, Departemen Proteksi Tanaman, Institut Pertanian Bogor, mulai bulan Desember 2004 sampai bulan Juli 2005.
Struktur komunitas parasitoid telur diamati dengan dua cara, yaitu pemasangan telur perangkap dan pengamatan langsung terhadap telur serangga yang terkumpul. Larva Lepidoptera dan gejala serangannya pada pertanaman kedelai juga diamati. Pemasangan telur perangkap dan pengamatan secara langsung dilakukan secara serentak. Lahan seluas 394, 25 m 2 dibagi menjadi tiga petak dengan jarak antara petak satu dengan yang lainnya sejauh 5 m. Setiap petak lahan dibagi lagi menjadi empat kuadran. Telur S. litura dan C.
cephalonica dipasang pada 20 titik contoh pertanaman kedelai yang telah
ditentukan mengikuti pola X setiap kuadrannya. Telur perangkap dipasang sekali dalam seminggu dan dengan lama pemasangan 6 jam pada setiap kali pemasangan. Pengamatan langsung dilakukan dengan menyisir seluruh luasan lahan. Telur perangkap yang telah dipasang di lapang beserta hasil pengamatan langsung, dipelihara dan diamati perkembangannya di laboratorium. Parasitoid telur yang muncul diidentifikasi sampai tingkat spesies melalui pengamatan terhadap genitalia jantan.
Struktur komunitas parasitoid telur pada pertanaman kedelai terdiri atas
Trichogrammatoidea armigera, Trichogrammatoidea cojuangcoi, Trichogramma
chilonis, Trichogramma japonicum, Trichogramma sp., Telenomus remus, Gryon
sp., dan satu spesies dari famili Encyrtidae. Dalam struktur komunitas pertanaman kedelai, Gryon sp. merupakan parasitoid dengan jumlah individu terbanyak, yaitu mencapai 179 individu dari 265 individu parasitoid telur yang ditemukan atau sebesar 36,23%. T. armigera merupakan spesies parasitoid telur yang paling sering ditemukan. Parasitoid telur ini memiliki jumlah individu terbanyak dari hasil pemasangan telur perangkap, yaitu mencapai 28 individu atau sebesar 10, 57%. Pada minggu-minggu awal, parasitoid muncul dari telur C. cephalonica sedangkan pada minggu-minggu akhir parasitoid muncul dari telur S. litura.
Pada pertanaman kedelai umur 0-12 MST ditemukan pula telur, larva, pupa, dan gejala serangan hama penjalin daun Lamprosema indicata
STRUKTUR KOMUNITAS PARASITOID TELUR
PADA PERTANAMAN KEDELAI DAN IMPLIKASINYA
TERHADAP TINGKAT SERANGAN DAN POPULASI HAMA
Skripsi
Sebagai salah satu syarat untuk memperoleh gelar Sarjana Pertanian pada Fakultas Pertanian
Institut Pertanian Bogor
Oleh :
Okti Aryani Hapsari A44101059
PROGRAM STUDI HAMA DAN PENYAKIT TUMBUHAN
FAKULTAS PERTANIAN
Judul : STRUKTUR KOMUNITAS PARASITOID TELUR PADA PERTANAMAN KEDELAI DAN IMPLIKASINYA TERHADAP TINGKAT SERANGAN DAN POPULASI HAMA
Nama : Okti Aryani Hapsari
NRP : A44101059
Menyetujui,
Dr. Ir. Pudjianto, MSi NIP. 131475578
Mengetahui, Dekan Fakultas Pertanian
Prof. Dr. Ir. H. Supiandi Sabiham, M Agr NIP. 130422698
RIWAYAT HIDUP
Penulis dilahirkan di Rembang, Jawa Tengah pada tanggal 13 Oktober 1982, sebagai anak pertama dari tiga bersaudara dari pasangan bapak Arief Djatmiko dan ibu Puji Handayani. Pada tahun 2001, penulis menyelesaikan pendidikan di Sekolah Menengah Umum Negeri 1 Rembang.
PRAKATA
Bismillahirrahmanirrahim
Puji syukur penulis panjatkan kehadirat Allah SWT. atas limpahan rahmat, berkah dan ridho-Nya dapat menyelesaikan skripsi yang berjudul “Struktur Komunitas Parasitoid Telur pada Pertanaman Kedelai dan Implikasinya terhadap Tingkat Populasi dan Serangan Hama”. Penelitian ini dilakukan di Bogor pada Desember 2005 – Juli 2006 dengan sumber dana penelitian dari Hibah Bersaing Perguruan Tinggi.
Penelitian ini tidak bisa lepas dari peran serta, dukungan dan bantuan dari berbagai pihak. Pada kesempatan ini, penulis menyampaikan ucapan terima kasih kepada:
1. Dr. Ir. Sri Hendrastuti Hidayat, MSc selaku dosen pembimbing akademik yang telah memberikan bimbingan dan masukan selama penulis aktif kuliah.
2. Dr. Ir. Pudjianto, MSi selaku dosen pembimbing skripsi yang telah memberikan saran dan petunjuk selama penelitian dan penyusunan skripsi. 3. Dr. Ir. Supramana, MSi selaku dosen penguji tamu yang telah memberikan
masukan dalam penyusunan skripsi.
4. Bapak, Ibu, Randy, Ita, Mba Yanik, Mba Dhani atas cinta, perhatian, doanya selama ini.
5. Seluruh staf pengajar Departemen Proteksi Tanaman, Fakultas Pertanian yang telah memberikan ilmunya selama penulis duduk di bangku kuliah. 6. Mba Adha, Mas Bandung, Mas Jalu, Mba Atik, Pak Slamet, Pak Ucup,
Pak Yanto, Mba Danur, Bang Hertab, Bang Alal, Mba Wika, Nita yang selama ini telah banyak memberikan dorongan, saran dan bantuannya. 7. Mba Nita (almh.), Mba Ikha, Uci, Saras, Mba Rury, Nda, Dewi, Dian,
Tito, Muslik atas kebersamaannya selama ini.
8. Lia, Mu’min, Ita, Mia, Asti, Nita, Iis, Mela, Winta, Nia, Indah yang telah memberikan arti persahabatan yang sebenarnya.
9. Andalebers 1 dan 2 terimakasih atas kebersamaannya selama ini.
10. Sopian, Ridwan, Deni, Zul, Duan, Budi juga teman-teman HPT 38 yang telah memberikan dukungan, bantuan dan persahabatan yang tulus selama ini.
11. Semua pihak yang telah membantu dan tidak bisa disebutkan satu persatu.
Penulis berharap semoga karya tulis ini dapat bermanfaat bagi semua pihak.
Bogor, Februari 2006
DAFTAR ISI
Sejarah Hidup, Morfologi, dan Biologi... 5
Ulat Penggerek Polong Kedelai ... 6
Taksonomi ... 6
Sejarah Hidup, Morfologi, dan Biologi ... 6
Ulat Tanah ... 7
Taksonomi ... 7
Sejarah Hidup, Morfologi, dan Biologi ... 7
Lalat Bibit... 8
Taksonomi ... 8
Sejarah Hidup, Morfologi, dan Biologi ... 8
Ulat Penjalin Daun Kedelai ... 9
Taksonomi ... 9
Sejarah Hidup, Morfologi, dan Biologi ... 9
Kepik Hijau ... 9
Taksonomi ... 9
Sejarah Hidup, Morfologi, dan Biologi ... 9
Parasitoid Telur yang Berasosiasi dengan Tanaman Kedelai ... 10
Trichogrammatoidea spp. dan Trichogramma spp... ... 12
Taksonomi ... 12
Sejarah Hidup, Morfologi, dan Biologi ... 12
Taksonomi ... 13
Sejarah Hidup, Morfologi, dan Biologi ... 13
BAHAN DAN METODE ... 15
Tempat dan Waktu ... 15
Bahan dan Alat ... 15
Budidaya Tanaman Kedelai ... 15
Analisis Struktur Komunitas Parasitoid Telur ... 16
Pemasangan Telur Perangkap di Lapang ... 17
Pengumpulan Telur Serangga dari Lapang ... 18
Pengamatan Larva Lepidoptera dan Gejala Serangannya ... 18
Identifikasi... 18
HASIL DAN PEMBAHASAN ... 20
Struktur Komunitas Parasitoid Telur ... 20
Telur Serangga Alami, Larva Lepidoptera dan Gejala pada Pertanaman Kedelai ... 26
Hubungan Antara Struktur Komunitas Parasitoid Telur dengan Tingkat Serangan dan Populasi Hama ... 31
KESIMPULAN DAN SARAN ... 33
Kesimpulan ... 33
Saran... 33
DAFTAR PUSTAKA ... 35
DAFTAR TABEL
Teks
No. Halaman 1 Parasitoid Telur yang Pernah Digunakan di Indonesia ... 11 2 Stadia Hidup dan Gejala Serangan Lamprosema indicata pada
DAFTAR GAMBAR
Teks
No. Halaman
1. Penentuan Titik Contoh Setiap Petak Lahan ... 17
2. Struktur Komunitas Parasitoid Telur pada Pertanaman Kedelai di Cikabayan, Bogor... 21
3. Kekayaan dan Kelimpahan Parasitoid Telur C. cephalonica, S. litura dan Serangga Alami pada Pertanaman Kedelai Umur 0-12 MST ... 21
4. Kelimpahan Parasitoid Telur pada Pertanaman Kedelai Asal Telur Perangkap C. cephalonica, dan S. litura serta Telur
12. Telur serangga tidak teridentifikasi ... 44
13. Lamprosema indicata ... 44
14. Pupa parasitoid larva L. indicata ... 45
15. Parasitoid larva L. indicata (famili Braconidae) ... 45
16. Parasitoid larva L. Indicata (famili Elasmidae) ... 45
STRUKTUR KOMUNITAS PARASITOID TELUR
PADA PERTANAMAN KEDELAI DAN IMPLIKASINYA
TERHADAP TINGKAT SERANGAN DAN POPULASI HAMA
Oleh :
Okti Aryani Hapsari
A44101059
PROGRAM STUDI HAMA DAN PENYAKIT TUMBUHAN
FAKULTAS PERTANIAN
ABSTRAK
OKTI ARYANI HAPSARI. Struktur Komunitas Parasitoid Telur pada Pertanaman Kedelai dan Implikasinya terhadap Tingkat Serangan dan Populasi Hama. Dibimbing oleh PUDJIANTO.
Penelitian ini bertujuan untuk mengetahui jenis-jenis parasitoid telur yang terdapat pada pertanaman kedelai serta me mpelajari hubungannya dengan serangan hama. Penelitian dilaksanakan pada lahan pertanaman kedelai di Cikabayan, Bogor dan di Laboratorium Bioekologi Parasitoid, Departemen Proteksi Tanaman, Institut Pertanian Bogor, mulai bulan Desember 2004 sampai bulan Juli 2005.
Struktur komunitas parasitoid telur diamati dengan dua cara, yaitu pemasangan telur perangkap dan pengamatan langsung terhadap telur serangga yang terkumpul. Larva Lepidoptera dan gejala serangannya pada pertanaman kedelai juga diamati. Pemasangan telur perangkap dan pengamatan secara langsung dilakukan secara serentak. Lahan seluas 394, 25 m 2 dibagi menjadi tiga petak dengan jarak antara petak satu dengan yang lainnya sejauh 5 m. Setiap petak lahan dibagi lagi menjadi empat kuadran. Telur S. litura dan C.
cephalonica dipasang pada 20 titik contoh pertanaman kedelai yang telah
ditentukan mengikuti pola X setiap kuadrannya. Telur perangkap dipasang sekali dalam seminggu dan dengan lama pemasangan 6 jam pada setiap kali pemasangan. Pengamatan langsung dilakukan dengan menyisir seluruh luasan lahan. Telur perangkap yang telah dipasang di lapang beserta hasil pengamatan langsung, dipelihara dan diamati perkembangannya di laboratorium. Parasitoid telur yang muncul diidentifikasi sampai tingkat spesies melalui pengamatan terhadap genitalia jantan.
Struktur komunitas parasitoid telur pada pertanaman kedelai terdiri atas
Trichogrammatoidea armigera, Trichogrammatoidea cojuangcoi, Trichogramma
chilonis, Trichogramma japonicum, Trichogramma sp., Telenomus remus, Gryon
sp., dan satu spesies dari famili Encyrtidae. Dalam struktur komunitas pertanaman kedelai, Gryon sp. merupakan parasitoid dengan jumlah individu terbanyak, yaitu mencapai 179 individu dari 265 individu parasitoid telur yang ditemukan atau sebesar 36,23%. T. armigera merupakan spesies parasitoid telur yang paling sering ditemukan. Parasitoid telur ini memiliki jumlah individu terbanyak dari hasil pemasangan telur perangkap, yaitu mencapai 28 individu atau sebesar 10, 57%. Pada minggu-minggu awal, parasitoid muncul dari telur C. cephalonica sedangkan pada minggu-minggu akhir parasitoid muncul dari telur S. litura.
Pada pertanaman kedelai umur 0-12 MST ditemukan pula telur, larva, pupa, dan gejala serangan hama penjalin daun Lamprosema indicata
STRUKTUR KOMUNITAS PARASITOID TELUR
PADA PERTANAMAN KEDELAI DAN IMPLIKASINYA
TERHADAP TINGKAT SERANGAN DAN POPULASI HAMA
Skripsi
Sebagai salah satu syarat untuk memperoleh gelar Sarjana Pertanian pada Fakultas Pertanian
Institut Pertanian Bogor
Oleh :
Okti Aryani Hapsari A44101059
PROGRAM STUDI HAMA DAN PENYAKIT TUMBUHAN
FAKULTAS PERTANIAN
Judul : STRUKTUR KOMUNITAS PARASITOID TELUR PADA PERTANAMAN KEDELAI DAN IMPLIKASINYA TERHADAP TINGKAT SERANGAN DAN POPULASI HAMA
Nama : Okti Aryani Hapsari
NRP : A44101059
Menyetujui,
Dr. Ir. Pudjianto, MSi NIP. 131475578
Mengetahui, Dekan Fakultas Pertanian
Prof. Dr. Ir. H. Supiandi Sabiham, M Agr NIP. 130422698
RIWAYAT HIDUP
Penulis dilahirkan di Rembang, Jawa Tengah pada tanggal 13 Oktober 1982, sebagai anak pertama dari tiga bersaudara dari pasangan bapak Arief Djatmiko dan ibu Puji Handayani. Pada tahun 2001, penulis menyelesaikan pendidikan di Sekolah Menengah Umum Negeri 1 Rembang.
PRAKATA
Bismillahirrahmanirrahim
Puji syukur penulis panjatkan kehadirat Allah SWT. atas limpahan rahmat, berkah dan ridho-Nya dapat menyelesaikan skripsi yang berjudul “Struktur Komunitas Parasitoid Telur pada Pertanaman Kedelai dan Implikasinya terhadap Tingkat Populasi dan Serangan Hama”. Penelitian ini dilakukan di Bogor pada Desember 2005 – Juli 2006 dengan sumber dana penelitian dari Hibah Bersaing Perguruan Tinggi.
Penelitian ini tidak bisa lepas dari peran serta, dukungan dan bantuan dari berbagai pihak. Pada kesempatan ini, penulis menyampaikan ucapan terima kasih kepada:
1. Dr. Ir. Sri Hendrastuti Hidayat, MSc selaku dosen pembimbing akademik yang telah memberikan bimbingan dan masukan selama penulis aktif kuliah.
2. Dr. Ir. Pudjianto, MSi selaku dosen pembimbing skripsi yang telah memberikan saran dan petunjuk selama penelitian dan penyusunan skripsi. 3. Dr. Ir. Supramana, MSi selaku dosen penguji tamu yang telah memberikan
masukan dalam penyusunan skripsi.
4. Bapak, Ibu, Randy, Ita, Mba Yanik, Mba Dhani atas cinta, perhatian, doanya selama ini.
5. Seluruh staf pengajar Departemen Proteksi Tanaman, Fakultas Pertanian yang telah memberikan ilmunya selama penulis duduk di bangku kuliah. 6. Mba Adha, Mas Bandung, Mas Jalu, Mba Atik, Pak Slamet, Pak Ucup,
Pak Yanto, Mba Danur, Bang Hertab, Bang Alal, Mba Wika, Nita yang selama ini telah banyak memberikan dorongan, saran dan bantuannya. 7. Mba Nita (almh.), Mba Ikha, Uci, Saras, Mba Rury, Nda, Dewi, Dian,
Tito, Muslik atas kebersamaannya selama ini.
8. Lia, Mu’min, Ita, Mia, Asti, Nita, Iis, Mela, Winta, Nia, Indah yang telah memberikan arti persahabatan yang sebenarnya.
9. Andalebers 1 dan 2 terimakasih atas kebersamaannya selama ini.
10. Sopian, Ridwan, Deni, Zul, Duan, Budi juga teman-teman HPT 38 yang telah memberikan dukungan, bantuan dan persahabatan yang tulus selama ini.
11. Semua pihak yang telah membantu dan tidak bisa disebutkan satu persatu.
Penulis berharap semoga karya tulis ini dapat bermanfaat bagi semua pihak.
Bogor, Februari 2006
DAFTAR ISI
Sejarah Hidup, Morfologi, dan Biologi... 5
Ulat Penggerek Polong Kedelai ... 6
Taksonomi ... 6
Sejarah Hidup, Morfologi, dan Biologi ... 6
Ulat Tanah ... 7
Taksonomi ... 7
Sejarah Hidup, Morfologi, dan Biologi ... 7
Lalat Bibit... 8
Taksonomi ... 8
Sejarah Hidup, Morfologi, dan Biologi ... 8
Ulat Penjalin Daun Kedelai ... 9
Taksonomi ... 9
Sejarah Hidup, Morfologi, dan Biologi ... 9
Kepik Hijau ... 9
Taksonomi ... 9
Sejarah Hidup, Morfologi, dan Biologi ... 9
Parasitoid Telur yang Berasosiasi dengan Tanaman Kedelai ... 10
Trichogrammatoidea spp. dan Trichogramma spp... ... 12
Taksonomi ... 12
Sejarah Hidup, Morfologi, dan Biologi ... 12
Taksonomi ... 13
Sejarah Hidup, Morfologi, dan Biologi ... 13
BAHAN DAN METODE ... 15
Tempat dan Waktu ... 15
Bahan dan Alat ... 15
Budidaya Tanaman Kedelai ... 15
Analisis Struktur Komunitas Parasitoid Telur ... 16
Pemasangan Telur Perangkap di Lapang ... 17
Pengumpulan Telur Serangga dari Lapang ... 18
Pengamatan Larva Lepidoptera dan Gejala Serangannya ... 18
Identifikasi... 18
HASIL DAN PEMBAHASAN ... 20
Struktur Komunitas Parasitoid Telur ... 20
Telur Serangga Alami, Larva Lepidoptera dan Gejala pada Pertanaman Kedelai ... 26
Hubungan Antara Struktur Komunitas Parasitoid Telur dengan Tingkat Serangan dan Populasi Hama ... 31
KESIMPULAN DAN SARAN ... 33
Kesimpulan ... 33
Saran... 33
DAFTAR PUSTAKA ... 35
DAFTAR TABEL
Teks
No. Halaman 1 Parasitoid Telur yang Pernah Digunakan di Indonesia ... 11 2 Stadia Hidup dan Gejala Serangan Lamprosema indicata pada
DAFTAR GAMBAR
Teks
No. Halaman
1. Penentuan Titik Contoh Setiap Petak Lahan ... 17
2. Struktur Komunitas Parasitoid Telur pada Pertanaman Kedelai di Cikabayan, Bogor... 21
3. Kekayaan dan Kelimpahan Parasitoid Telur C. cephalonica, S. litura dan Serangga Alami pada Pertanaman Kedelai Umur 0-12 MST ... 21
4. Kelimpahan Parasitoid Telur pada Pertanaman Kedelai Asal Telur Perangkap C. cephalonica, dan S. litura serta Telur
12. Telur serangga tidak teridentifikasi ... 44
13. Lamprosema indicata ... 44
14. Pupa parasitoid larva L. indicata ... 45
15. Parasitoid larva L. indicata (famili Braconidae) ... 45
16. Parasitoid larva L. Indicata (famili Elasmidae) ... 45
PENDAHULUAN
Latar Belakang
Kedelai merupakan komoditas pertanian yang sangat penting karena bersifat multiguna, baik untuk dikonsumsi langsung maupun untuk keperluan yang lain. Sebagai sumber protein nabati yang utama dan penghasil minyak, kedelai banyak dimanfaatkan manusia. Sejak ribuan tahun yang lalu, masyarakat Cina, Korea, Jepang, dan Indonesia telah memanfaatkannnya sebagai tanaman pertanian. Di Indonesia, tanaman kedelai banyak ditanam di Jawa (± 80 %), Madura, Nusa Tenggara, dan beberapa daerah di Sumatera (Oerke et al. 1999).
Kedelai merupakan tanaman pertanian penghasil minyak yang paling penting di dunia (Oerke et al. 1999). Kedelai mengandung 30-50 % protein, 25 %
karbohidrat dan 15-30 % minyak. Lebih dari 80 % hasil panen kedelai digunakan masyarakat dalam produksi minyak untuk konsumsi, memproduksi cat, varnis,
resin, dan plastik (Oerke et al. 1999; Smith 1994); kayu lapis dan kosmetik (Smith 1994); serta pakan ternak (Umar 1996).
Melihat potensi kedelai dalam pemenuhan berbagai kebutuhan, upaya peningkatan produksi kedelai dalam negeri melalui perluasan areal dan
peningkatan produktivitas telah dilakukan. Namun, upaya tersebut belum berhasil memenuhi kebutuhan kedelai dalam negeri setiap tahunnya (Umar 1996). Upaya
perluasan areal dari 643.000 ha menjadi 1,2 juta ha dilakukan di dalam kurun waktu 1969-1971. Produktivitas kedelai saat itu hanya meningkat dari 728 kg/ ha menjadi 1.105 kg/ ha (Oerke et al. 1999). Indonesia sendiri pernah mengekspor kedelai pada tahun 1972. Namun pada tahun-tahun berikutnya, volume ekspor kedelai menurun bahkan volume impor kedelai menjadi naik (Sihombing 1985). Sampai tahun 2005, produksi kedelai nasional rata-rata 700.000 ton, sedangkan kebutuhan nasional mencapai 1,4-1,5 juta ton, sehingga diperlukan kedelai impor (Kusdirianto 2005). Ini merupakan peluang sekaligus tantangan bagi para petani Indonesia untuk meningkatkan produksi kedelai dalam negeri.
spp., Etiella zinckenella (Suryana 1999; Oerke et al. 1999), Agrotis spp., dan
Spodoptera litura merupakan hama-hama penting pada pertanaman kedelai yang
menyebabkan kerugian ekonomi secara signifikan (Oerke etal. 1999). Kalshoven (1981) menunjuk S. litura sebagai hama penting pada pertanaman kedelai. Melihat beragamnya hama pada tanaman kedelai, Sihombing (1985) menyarankan dilakukan program-program pengendalian hama secara efektif dan efisien.
Pengendalian hama tanaman kedelai yang biasa dilakukan adalah dengan aplikasi insektisida (Suryana 1999; Turnipseed & Kogan 1994). Namun, aplikasi insektisida yang kurang bijaksana berakibat buruk terhadap upaya pengendalian itu sendiri, terhadap faktor biotik lingkungan dan manusia serta konsumen komoditi pertanian (Sosromarsonono 2002). Penggunaan insektisida yang berlebih menimbulkan dampak negatif terhadap lingkungan sekitar, sehingga
mendorong pemerintah mengeluarkan kebijaksanaan tentang penetapan konsep PHT (Pengandalian Hama Terpadu).
Salah satu prinsip PHT adalah insektisida hanya digunakan untuk mengendalikan hama yang statusnya telah mencapai ambang ekonomi.
Penggunaan musuh alami (parasitoid, predator, dan patogen) menjadi komponen utama dalam PHT. Parasitoid telur sebagai agens pengendali hayati hama
merupakan trend dalam pengendalian hama yang sering diusahakan saat ini (Sosromarsono 2002). Menurut Alba (1988), penggunaaan parasitoid telur
sebagai agens pengendali hayati hama dinilai efektif karena hama yang dikendalikan berada pada stadia awal, sehingga usaha pengendalian terjadi sebelum hama merusak tanaman. Penentuan spesies parasitoid telur yang tepat dalam mengendalikan hama pada suatu komoditas merupakan hal yang sangat penting sebelum upaya pengendalian dilakukan, sehingga perlu diketahui jenis-jenis parasitoid telur yang terdapat pada komoditas tersebut.
Tujuan
Manfaat
TINJAUAN PUSTAKA
Tanaman Kedelai
Taksonomi
Kedelai (Glycine max L.) termasuk famili Leguminosae. Kedelai merupakan tanaman semusim yang tumbuh tegak, batangnya bercabang dan pada
beberapa kultivar permukaannya berambut halus (Oerke et al.1999). Tanaman kedelai merupakan tanaman perdu yang berbatang (Umar 1996) dan memiliki ketinggian tanaman antara 30 sampai 150 cm (Oerke et al. 1999). Daun yang terbentuk terdiri dari tiga anak daun yang bentuknya lonjong atau bulat telur, berwarna hijau pucat, namun pada beberapa varietas berwarna hijau gelap (Umar 1996).
Budidaya Tanaman Kedelai
Kedelai dapat tumbuh pada berbagai jenis tanah yang memiliki drainase dan aerasi yang cukup baik. Pengairan yang cukup selama pertumbuhan tanaman juga menjadi syarat tumbuh kedelai (Deptan 2005). Kedelai dapat tumbuh baik pada tanah alluvial, regosol, grumosol, latosol atau andosol. Pada tanah yang kurang subur dan jenis tanah podsolik merah-kuning, perlu diberi pupuk organik dan pengapuran.
Kedelai dapat tumbuh subur pada curah hujan optimal 100-200 mm/
bulan; temperatur 25-27 °C dengan penyinaran penuh minimal 10 jam/ hari;
ketinggian tempat 0-900 m di atas permukaan laut. Ketinggian tempat 600 m di atas permukaan laut merupakan ketinggian optimal bagi pertumbuhan tanaman kedelai. Curah hujan yang cukup selama pertumbuhan, berkurang saat pembungaan dan menjelang pemasakan biji, akan meningkatkan hasil kedelai (Deptan 2005).
Hama-hama Tanaman Kedelai
Ophiomyia spp., Etiellazinckenella (Treitsche) (Suryana 1999; Oerke et al. 1999),
Agrotis spp., dan Spodoptera litura (Fabricius) (Oerke et al. 1999), dan kepik
Nezara viridula (Linnaeus) (Hemiptera: Pentatomidae) (Indayani 2002).
Ulat Grayak Taksonomi
Ulat grayak, Spodoptera litura (Fabricius), tergolong dalam ordo
Lepidoptera dan famili Noctuidae (Borror et al. 1992). Sebelumnya serangga ini dikenal dengan nama ilmiah Laphygma frugiperda (Baldwin 1994). S. litura
merupakan serangga yang berasal dari Afrika dan Eropa, namun sekarang bersifat kosmopolitan dan dapat menyerang berbagai tanaman pertanian (Kalshoven 1981).
Sejarah Hidup, Morfologi, dan Biologi
Telur diletakkan dalam kelompok pada permukaan bawah daun dan ditutupi oleh sisik. Sisik yang dihasilkan ujung abdomen betina ini berfungsi sebagai pelindung telur-telur terhadap tusukan ovipositor parasitoid. Menurut Kalshoven (1981), satu kelompok telur mengandung kira-kira 350 butir telur. Menurut Baldwin (1994) telur diletakkan pada malam hari dengan stadia telur
berkisar antara dua sampai empat hari pada suhu 21-27 °C. Larva instar awal
bersifat gregarius, memakan daun pada bagian permukaan bawah. Larva instar satu, dua, dan tiga menyebabka n kerusakan kurang dari 2%. Baldwin (1994) melaporkan berdasarkan penelitian yang dilakukan, menunjukkan rata-rata konsumsi larva instar satu, dua, tiga berturut-turut mencapai 5, 16, dan 77%. Ini berarti bahwa dalam beberapa hari saja dapat terjadi penggundulan daun (Baldwin 1994). Menjelang masa berpupa, larva instar akhir menuju tanah dan pemupaan terjadi di dalam tanah. Imago merupakan ngengat berumur pendek, berkisar antara tujuh sampai sembilan hari, tergantung pada suhu dan musim.
S. litura melewati enam instar larva. Larva instar awal memiliki ciri kepala hitam, abdomen hijau kekuningan, dan hidup gregarius di bawah
permukaan daun. Larva instar dua memiliki kepala berwarna coklat muda, abdomen hijau kekuningan kemudian hijau coklat dengan garis-garis putih
titik hitam pada kedua sisinya. Larva instar tiga memiliki ciri khas pada ‘leher’ terdapat lingkaran berwarna hitam menyerupai kalung. Memasuki instar ini, larva mulai hidup memencar. Larva instar empat memiliki warna dasar abu-abu. Pada bagian dorsal abdomen terdapat tiga garis kuning memanjang dan terdapat bintik-bintik kuning berbentuk setengah lingkaran pada garis tersebut. Larva instar lima dan instar ena m memiliki ciri yang sama. Keduanya dibedakan berdasarkan ukurannya. Larva tersebut memiliki ciri abdomen berwarna hitam, garis-garis kuning pada bagian dorsal abdomen yang kemudian berubah warna menjadi jingga. Bintik hitam dan kuning pada ruas kedua dan ketiga abdomen serta garis jingga dan putih pada ruas ke-11. Pupa berwarna coklat gelap, ukuran pupa calon betina lebih besar dan panjang dari pupa calon jantan. Imago jantan dan betina dapat dibedakan dari pola sayapnya. Ngengat betina tidak memiliki
pola warna biru keabu-abuan pada bagian belakang sayap depan (Evan & Crossley 2004).
Pada siang hari, S. litura cenderung berlindung dan bersembunyi di dalam tanah maupun di balik daun. Aktivitas penyerangan baru dimulai setelah malam
hari (Kalshoven 1981). Sebelum kopulasi terjadi, imago betina selalu mengeluarkan feromon sex untuk menarik imago jantan. Respon imago jantan
terhadap feromon sex dapat berlangsung selama empat hari setelah feromon tersebut dihasilkan imago betina (Baldwin 1994).
Ulat Penggerek Polong Kedelai Taksonomi
Ulat penggerek polong kedelai, Etiella zinckenella (Treitsche), tergolong dalam ordo Lepidoptera, subordo Microlepidoptera, superfamili Pyralidoidae, dan
famili Pyralidae (Borror et al. 1992). E. zinckenella dikenal sebagai penggerek polong Leguminosae di hampir semua negeri beriklim tropis (Kalshoven 1981).
Sejarah Hidup, Morfologi, dan Biologi
Imago mampu meletakkan 7-15 butir telur secara berkelompok pada sisi
bawah calyx dan sepal. Telur yang baru diletakkan berwarna putih dan berubah menjadi merah jambu, berukuran rata-rata 0,55 x 0,34 mm dengan masa inkubasi
1992). Larva instar awal akan segera menggerek melalui sebuah lubang kecil pada polong dan memakan biji polong (Kalshoven 1981). Menurut Yuanivar (1992), larva instar awal dicirikan oleh kepala berwarna hitam. Lebih lanjut dijelaskan, pada instar dua dan tiga memiliki warna tubuh kehijauan dengan garis horizontal merah dan ditumbuhi rambut-rambut. Larva instar empat memiliki kepala berwarna kuning. Memasuki instar kelima, warna tubuh berubah menjadi merah kebiruan. Menurut Kalshoven (1981), larva instar akhir ini meninggalkan polong sebelum biji polong masak. Pemupaan terjadi dalam tanah. Pupa terbungkus dalam kokon dan berukuran panjang kira-kira 14 mm (Yuanivar 1992).
Imago spesies ini merupakan ngengat dengan ukuran jantan lebih kecil dari betina. Lama hidup imago jantan mencapai 4,8 hari, sedangkan imago betina
mencapai 5,2 hari pada suhu rata-rata 28 °C. Pada suhu yang sama, siklus hidupnya mencapai 25 hari (Yuanivar 1992).
Ulat Tanah Taksonomi
Ulat tanah, Agrotisipsilon (Hufnaker), tergolong dalam ordo Lepidoptera, subordo Heterocera dan famili Noctuidae (Kalshoven 1981). Dalam bahasa
inggris, hama ini dikenal dengan nama umum cutworm. Menurut Wintersteen (1994), Agrotis ipsilon dan spesies ulat tanah yang lain merupakan spesies asli
Amerika Utara dan telah tersebar luas di dunia.
Sejarah Hidup, Morfologi, dan Biologi
Larva Agrotis spp. berada dalam tanah, bersifat polifag dan rakus (Kalshoven 1981). Kerusakan tanaman akibat aktvitas Agrotis spp. ini terlihat
melalui terpotongnya batang tanaman di atas permukaan tanah (Kalshoven 1981). Pada pembibitan kedelai, larva merusak dengan memotong bibit tanaman kedelai
pada bagian bawah kotiledon (Wintersteen 1994). A. ipsilon (Lepidoptera: Noctuidae) berukuran panjang 6-38 mm (Wintersteen 1994); berwarna gelap
hingga coklat kehitaman, agak licin dan berkilat, serta terdapat pita berwarna coklat pada kedua sisi abdomennya (Kalshoven 1981). Pupa berwarna coklat dan
imago berwarna abu pada bagian abdomen, sedangkan sayap berwarna abu-abu dengan spot pada sayap depan brwarna hitam kecoklatan (Kalshoven 1981; Wintersteen 1994). Imago merupakan penerbang yang baik dan mampu menghasilkan telur sebanyak 970 butir (Kalshoven 1981). Namun, di Bogor, imago betina spesies ini mampu memproduksi telur sebanyak 1430-2775 butir (Kalshoven 1981). Lama perkembangannya (telur sampai imago) di Bogor mencapai 36 hari (Kalshoven 1981).
Imago betina A. ipsilon mampu menghasilkan feromon pada hari keempat setelah waktu pemunculan (Kalshoven 1981). Feromon ini me rupakan bentuk komunikasi interspesies dan secara kontinyu dikeluarkan betina yang belum kawin.
Lalat Bibit Taksonomi
Lalat bibit, Ophiomyia phaseoli (Tryon.), tergolong dalam ordo Diptera, subordo Cyclorrapha, dan famili Agromyzidae (Kalshoven 1981). Spesies ini dikenal dengan nama umum bean fly dan merupakan hama penting di pembibitan (Kalshoven 1981). Saat ini telah terdistribusi di Filipina, Malaysia, Indonesia, Taiwan, Srilanka, India, Afrika (Kalshoven 1981), Australia, Mesir, Israel, dan
Hawai (Rusamsi 1982).
Sejarah Hidup, Morfologi, dan Biologi
Imago meletakkan telur dekat kotiledon di pembibitan (Kalshoven 1981) atau pada permukaan daun (Rusamsi 1982). Telur berwarna putih, berbentuk lonjong, berukuran panjang 0,31 mm dan lebar 0,15 mm (Awaludin 1997). Stadium telur mencapai dua hari (Rusamsi 1982). Awaludin (1997) melaporkan
bahwa stadium larva berkisar antara tujuh sampai sepuluh hari. Pupa yang terbentuk berwarna kuning kecoklatan, berukuran panjang 3 mm dengan stadia
pupa berkisar antara 7-13 hari. Di Jawa dengan ketinggian tempat di atas permukaan laut, stadia larva berkisar antara 17-22 hari, sedangkan stadia pupa
berkisar antara 13-20 hari (Rusamsi 1982). Imago merupakan lalat berukuran kerukuran kecil, berwarna hitam mengkilat (Rusamsi 1982). Imago betina
Ulat Penjalin Daun Kedelai Taksonomi
Ulat penjalin daun kedelai, Lamprosema indicata (Fabricius), tergolong dalam ordo Lepidoptera dan famili Pyralidae (Kalshoven 1981).
Sejarah Hidup, Morfologi, dan Biologi
Telur diletakkan soliter pada permukaan daun kedelai, terutama pada daun
yang belum membuka (Awaludin 1997), berwarna bening seperti embun. Larva berwarna hijau mengkilap dengan kepala dan toraks berwarna ke kuningan, serta terdapat sepasang bercak hitam pada protoraks (Awaludin 1997). Dilaporkan juga bahwa larva instar akhir berukuran panjang 20 mm dan imago merupakan ngengat berwarna kuning kecoklatan dengan tiga garis coklat kehitaman pada sayapnya.
Kepik Hijau Taksonomi
Kepik hijau, Nezara viridula (Linnaeus), tergolong dalam ordo Hemiptera dan famili Pentatomidae. Kepik ini dikenal dengan nama lain Green Stink Bug dan kepik ijo (Kalshoven 1981). Sampai saat ini telah ditemukan menyerang tanaman pertanian dan hortikultura di Ethiopia, Australia, Eropa Selatan (Waterhouse & Sands 2001), dan Indonesia (Kalshoven 1981).
Sejarah hidup, Morfologi, dan Biologi
Telur kepik ini diketahui dapat diparasit oleh parasitoid telur Gryon sp. (Wardani 2001), Ooencyrtus malayensis Ferr., dan Telenomus sp. (Kalshoven 1981).
Parasitoid Telur yang Berasosiasi dengan Tanaman Kedelai
Berbagai jenis parasitoid telur telah dilaporkan pernah diaplikasikan dalam
pengendalian hayati terhadap hama -hama pada berbagai tanaman pertanian. Namun, penggunaan parasitoid telur untuk mengendalikan hama -hama
pertanaman kedelai masih jarang.
Parasitoid telur yang pernah dilaporkan digunakan untuk pengendalian hama pertanaman kedelai adalah Trichogramma chilonis terhadap Helicoverpa
armigera (Hubner) (Lepidoptera: Noctuidae) (Helinda et al. 1999),
Trichogrammatoidea bactrae bactrae terhadap Etiella sp. (Marwoto et al. 1996, 1999), Trichogrammatoidea armigera terhadap Etiella zinckenella (Meilin 1999) dan Telenomus remus terhadap Spodoptera litura (Yuliarti 2002). Informasi mengenai parasitoid telur lain yang pernah dilaporkan penggunaannya pada pertanaman lain di Indonesia (Meilin 1999, Yuliarti 2002, Moy 2005) disajikan dalam Tabel 1.
Trichogramma chilonis dilaporkan pernah digunakan untuk
mengendalikan hama H. armigera pada pertanaman kedelai di Indonesia. Hasil
penelitian terhadap pelepasan parasitoid telur secara inundasi dua kali seminggu sebanyak 350.000 ekor per ha sejak 23 Hari Setelah Tanam (HST) ternyata telah
berhasil mengendalikan populasi H. armigera. Tingkat parasitisasi T. chilonis
terhadap telur H. armigera mencapai 50 % sedangkan pada petak kontrol, tingkat
parasitisasinya hanya 11 %. (Herlinda et al. 1999).
Trichogrammatoidea bactrae bactrae juga pernah dilaporkan
penggunaannya utuk mengendalikan hama penggerek polong kedelai, Etiella zinckenella, H. armigera, dan Chrysodeixis chalcites (Esper) (Lepidoptera:
Tabel 1 Parasitoid telur yang pernah digunakan di Indonesia
No. Spesies Telur inang Tanaman Pustaka
1. Trichogramma
Chillo auricilius Tebu Samoedi et al.
(1988)
S. innotata Padi Kalshoven (1981)
2. Trichogramma chilonis
Helicoverpa armigera
Bawang Shepard & Barrion (1998)
3. Trichogrammatoidea armigera
H. armigera Kapas Nurindah & Bindra (1989)
E. zinckenella Tebu Nagarkatti & Nagaraja (1997)
S. incertulas Tebu Meilin (1999)
H. armigera Kapas Meilin (1999)
Plutella
Trichogrammatoidea armigera diketahui memarasit E. zinckenella pada pertanaman kedelai (Meilin 1999). Namun, belum ada laporan mengenai keefektifannya dalam mengendalikan E. zinckenella.
Telenomus remus dilaporkan dapat memarasit telur S. litura pada
Trichogrammatoidea spp. dan Trichogramma spp. Taksonomi
Trichogrammatoidea dan Trichogramma termasuk dalam ordo
Hymenoptera, subfamili Apocrita, superfamili Chalcidoidea, dan famili Trichogrammatidae (Borror et al. 1992). Berdasarkan cara penyerangannya, tergolong parasitoid dalam (endoparasitoid) karena Trichogrammatoidea spp. dan
Trichogramma sp. berkembang di dalam inangnya. Trichogrammatoidea spp. dan
Trichogramma spp. menyelesaikan perkembangannya pada stadia inang yang
diletaki telur atau dikenal idiobiont (Godfray 1994).
Dalam penelitian terhadap pengendalian hama ordo Lepidoptera yang pernah dilakukan di Philipina, Alba (1988) melaporkan bahwa enam spesies
Trichogrammatoidea diketahui efektif memarasit telur hama Lepidoptera pada
berbagai inang tanaman.
Sejarah Hidup, Morfologi, dan Biologi
Hassan (1993) melaporkan 18 spesies Trichogramma dan 3 spesies
Trichogrammatoidea dapat memarasit hama pada jagung, tebu, padi, kedelai,
kapas, bit gula, sayuran, dan cemara dari 24 negara. Sedangkan Pinto et al.
(1989) menyebutkan terdapat ±130 spesies Trichogramma di dunia. Kalshoven
(1981) melaporkan bahwa di Bogor, Trichogrammatoidea nana pernah memarasit telur H. armigera.
Trichogrammatidae dikenal sebagai tabuhan kecil (< 0,5 mm), sayapnya
memiliki fringe setae (rumbai), mata berwarna merah, bersifat polifag, dan terdistribusi luas (Kalshoven 1981). Selanjutnya juga disebutkan bahwa
klasifikasi Trichogrammatoidea spp. dan Trichogramma spp. sangat sulit karena perbedaan morfologi keduanya sangat kecil (Kalshoven 1981; Nagarkatti &
Nagaraja 1977). Namun, Nagarkatti & Nagaraja (1977) dan Alba (1988) menyatakan bahwa fringe setae Trichogrammatoidea spp. lebih panjang
dibanding Trichogramma spp.
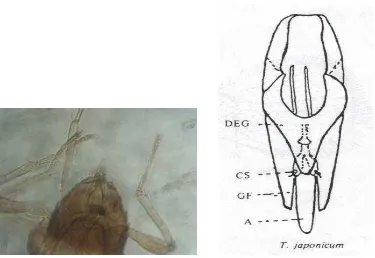
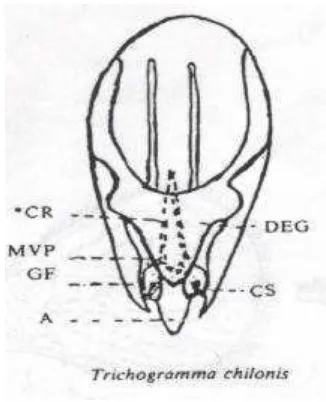
Karakter morfologi seperti genitalia jantan dapat digunakan sebagai
memiliki lekukan gonobase (GB) yang terlihat jelas dan Chelate Structure (CS), tidak memiliki Lateral Tubercle (LT), serta posisi Gonoforcep (GF) tidak menutup Aedeagus (A) sampai bawah (Alba 1988). Lebih lanjut dijelaskan bahwa Trichogrammatoidea bactrae bactrae memiliki Gonobase (GB) yang tidak berlekuk, Lateral Tubercle (LT) dan Chelate Structural (CS) terlihat jelas,
Gonoforcep (GF) menutup Aedaeagus (A) sampai bawah, serta Aedeagus tidak sebesar Trichogrammatoidea armigera.
Trichogrammatidae memiliki tipe perkembangan arrhenothoky (Nagarkatti & Nagaraja 1977). Terkait dengan proses penemuan inang, Noldus & Van Lenteren (1995 dalam Van Alphen dan Vet 1986) melaporkan bahwa
Trichogramma spp. merespon feromon yang dihasilkan inang dewasa. Respon
yang dilakukan parasitoid beragam (Waage 1978 dalam Van Alphen dan Vet
1986), misalnya ditunjukkan oleh perubahan kecepatan terbang (orthokinesis) (Va n Alphen dan Vet 1986).
Telenomus spp. Taksonomi
Telenomus spp. diklasifikasikan dalam ordo Hymenoptera, subfamili
Apocrita, superfamili Proctotrupoidea, dan famili Scelionidae (Borror et al. 1992).
Dalam penggolongannya berdasarkan cara penyerangan, Telenomus spp. termasuk parasitoid dalam (endoparasitoid). Telenomus spp. merupakan
parasitoid telur dari berbagai ordo serangga (Kalshoven 1981).
Polaszek et al. (1993) melaporkan, genus Telenomus terdiri atas 500
spesies dimana semua spesies tersebut merupakan parasitoid telur dan sebagian besar memarasit ordo Lepidoptera dan Hemiptera. Anggara (2005) melaporkan,
spesies Telenomus remus merupakan parasitoid spesifik telur S. litura.
Sejarah Hidup, Morfologi, dan Biologi
Telenomus spp. merupakan tabuhan yang terdistribusi luas meliputi
Indonesia (Jawa, Bangka) dan Jepang (Kalshoven 1981). Siklus hidupnya terdiri
atas empat stadia, yaitu telur, larva, pupa, dan imago (Anggara 2005). Telenomus
remus hanya mengoviposisi telur inang yang berumur kurang dari 72 jam atau
juga bahwa stadia telur berkisar 10 jam pada suhu 30 °C dan 18-24 jam pada suhu 15,5 °C. Larva T.remus melewati dua stadia instar dan memerlukan waktu empat
hari pada suhu 30 °C dan tujuh hari pada suhu 15,5 °C untuk menyelesaikan
stadia larva (Anggara 2005). Stadia pupa memerlukan waktu 6 hari dan imago T. remus keluar dari telur inang melalui lubang kecil yang telah dibuat (Anggara 2005). Kalshoven (1981) melaporkan, lama perkembangan Telenomus spp. pada
telur Chilo sp. berkisar 8-14 hari dan pada sebagian besar spesies Telenomus, hanya satu imago yang berkembang atau muncul dari setiap telur inang. Selanjutnya, disebutkan bahwa tipe perkembangannya adalah arrhenothoky
dengan lama hidup mencapai 20 hari. Di laboratorium, lama hidup T. remus
betina pada inang Spodoptera exigua dan Spodoptera litura dengan pakan tambahan madu 10%, berkisar 6-16 hari (Anggara 2005).
Telur T. remus berbentuk lonjong dan berwarna bening (Anggara 2005). Larva instar pertama dicirikan dengan tubuh yang tidak bersegmen, memiliki sepasang mandible yang bergerak vertikal, serta memiliki dua spina ekor
(Anggara 2005). Selanjutnya disebutkan bahwa larva instar dua memiliki tubuh bersegmen, mandible lebih pendek dan lurus, serta tidak memliki spina ekor.
Polaszek et al. (1993) melakukan pengamatan terhadap bentuk genitalia jantan
Telenomus spp. untuk mengetahui spesies Telenomus.
Cave (2000 dalam Anggara 2005) menggambarkan perilaku perkawinan T. remus bahwa imago jantan yang muncul lebih dahulu dari telur inang akan tetap
BAHAN DAN METODE
Tempat dan Waktu
Penelitian ini dilakukan pada lahan pertanaman kedelai di Cikabayan, Bogor dan di Laboratorium Bioekologi Parasitoid, Departemen Proteksi Tanaman, Institut Pertanian Bogor, mulai bulan Desember 2004 sampai bulan Juli 2005.
Bahan dan Alat
Benih kedelai yang digunakan adalah varietas Wilis yang diperoleh dari Balitro. Kebutuhan benih kedelai tersebut sebesar 31 kg/ha. Telur perangkap S. litura (Fabricius) (Lepidoptera: Noctuidae) dan C. cephalonica (Stainton) (Lepidoptera: Pyralidae) diperoleh dari hasil rearing di laboratorium. Telur perangkap yang dibutuhkan dalam setiap minggunya, masing-masing sebanyak 3600 butir. Telur perangkap direkatkan pada pias (karton manila 1 cm x 5 cm) dan dimasukkan dalam kulkas selama sehari. Untuk telur C. cephalonica, perlu difreezer dulu selama dua jam untuk menghambat penetasan telur bila telur tidak terparasit.Untuk memenuhi kebutuhan telur perangkap S.litura setiap minggunya, selain didapat dari hasil perbanyakan di laboratorium, setiap jangka waktu dua minggu juga dilakukan pengambilan larva S. litura (berbagai instar) dari lapang.
Bahan lain yang digunakan adalah kertas buram, daun talas, pakan buatan, pakan jagung dan pur, madu 10%, gum arabic, karton manila, isolatip, larutan
Hoyer, dan KOH 10%.
Alat-alat yang digunakan dalam proses penelitian ini adalah kotak plastik
untuk pemeliharaan S. litura dan C. cephalonica, pinset, kuas, tabung reaksi, gunting, wadah plastik, stoples, silinder peneluran, cawan petri, mikroskop, lampu
duduk, jarum bertangkai, ajir bambu sebagai tempat pemasangan telur di lapang, gelas obyek dan gelas penutup, gelas ukur Pyrex, kompor listrik, dan kunci
identifikasi Alba 1988, Nixon 1937, Barrion & Litsinger 1989.
Metode
Budidaya tanaman kedelai dilakukan pada lahan seluas 394, 25 m 2. Lahan dibagi menjadi tiga petakan dengan ukuran 10,5 m x 9,5 m untuk masing-masing petak. Jarak antara satu petak dengan petak lain sejauh 5 m. Ketiga petak tersebut diolah dengan cara ditraktor, selanjutnya dilakukan pengapuran dan pemberian pupuk kandang secara bersamaan setelah pengolahan lahan. Dosis pemberian kapur dan pupuk kandang masing-masing sebesar 2 ton/ha dan 20 ton/ ha.
Penanaman benih dilakukan dengan cara ditugal dimana dua benih ditanam dalam satu lubang dan menggunakan jarak tana m 20 cm x 40 cm. Benih yang tidak tumbuh disulam paling lambat pada saat tanaman berumur 1 MST (minggu setelah tanam).
Pemeliharaan yang dilakukan meliputi pemupukan, pengairan, dan penyiangan. Pupuk Urea, TSP, dan KCl diberikan secara bersamaan di sekitar
lubang benih (10 cm dari lubang benih) dengan dosis masing-masing 76 kg/ha, 152 kg/ha, dan 152 kg/ha. Setelah tanaman mulai berbunga, pupuk Urea
diberikan kembali, yaitu sebesar 76 kg/ha. Pengairan dilakukan menggunakan
sprinkle. Sampai dengan tana man berumur 1 MST, pengairan dilakukan setiap
hari. Sampai dengan tanaman berumur 2 MST, frekuensi pengairan sebanyak satu kali dalam dua hari. Selanjutnya, frekuensi pengairan hanya dua kali dalam
seminggu. Penyiangan dilakukan sebanyak dua kali, yaitu pada saat 2-3 minggu pertama dan saat tanaman mulai berbunga. Penyemprotan insektisida untuk
mengendalikan hama sengaja tidak dilakukan karena serangga hama dan parasitoid telur pada pertanaman kedelai merupakan obyek penting dalam penelitian ini.
Analisis Struktur Komunitas Parasitoid Telur
Struktur komunitas parasitoid telur diamati melalui dua cara, yaitu melalui pemasangan telur perangkap dan pengamatan langsung terhadap telur serangga
yang terkumpul dari pertanaman kedelai.
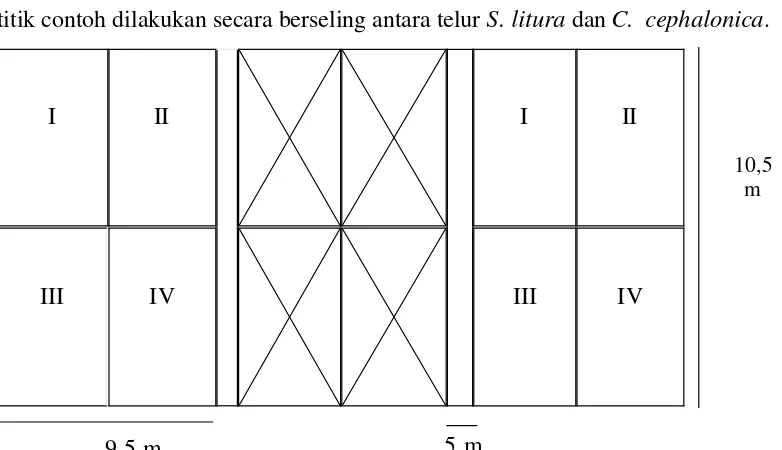
contohnya mengikuti pola X (Gambar 1). Pemasangan telur perangkap pada titik-titik contoh dilakukan secara berseling antara telur S. litura dan C. cephalonica.
I II
I II
III IV
III IV
9,5 m
Gambar 1 Penentuan titik contoh setiap petak lahan
Pengamatan langsung dilakukan terhadap telur serangga yang berhasil dikumpulkan dari pertanaman kedelai. Pengumpulan telur-telur tersebut
dilakukan dengan menyisir seluruh luasan lahan. Tujuan pengumpulan telur-telur ini adalah untuk mengetahui jenis-jenis serangga yang berasosiasi dengan
tanaman kedelai sehingga hubungan antara struktur komunitas parasitoid telur dengan serangan hama pertanaman kedelai dapat dipelajari.
Pemasangan Telur Perangkap di Lapang. Telur S. litura dan C.
cephalonica dipasang secara serentak pada pertanaman kedelai sejak tanaman
berumur 0 sampai 12 MST. Frekuensi pemasangan dilakukan satu kali dalam satu
minggu. Dalam setiap pemasangan, kebutuhan telur S. litura dan C. cephalonica
masing-masing 120 kelompok telur. Setiap satu kelompok telur, masing-masing
berisi ± 30 butir telur yang direkatkan pada pias. Hari pemasangan telur perangkap dalam setiap minggu diusahakan konsisten, namun tidak menutup
kemungkinan terjadinya percepatan atau penundaan waktu pemasangan karena faktor cuaca dan ketersediaan telur. Pemasangan telur perangkap selama enam
jam pada pertanaman. Telur kemudian diambil dan dipelihara di laboratorium sampai muncul parasitoidnya. Parasitoid yang muncul dibiarkan berkopulasi
5 m
selama 24 jam dan diparasitkan pada telur C. cephalonica sebagai inangnya di laboratorium. Tujuan perbanyakan parasitoid ini adalah untuk menyediakan parasitoid telur sebanyak-banyaknya sehingga apabila terjadi kerusakan pada saat proses identifikasi dapat diganti dengan spesies yang sama dari hasil perbanyakan. Pemeliharaan parasitoid dilakukan dengan memberikan madu 10% sebagai sumber nutrisi bagi kelangsungan hidupnya di laboratorium.
Pengumpulan Telur Serangga dari Lapang. Pengumpulan telur serangga dari lapang dilakukan bersamaan dengan waktu pemasangan telur
perangkap. Telur alami yang terkumpul, dipelihara dan diamati di laboratorium. Pemeliharaan parasitoid dilakukan dengan memberikan madu 10% sebagai
sumber nutrisi bagi kelangsungan hidupnya di laboratorium.
Pengamatan Larva Lepidoptera dan Gejala Serangannya
Pengumpulan larva Lepidoptera dan gejala serangannya dilakukan saat tanaman berumur 0-12 MST dengan frekuensi pengumpulan sekali dalam satu
minggu. Pengumpulan dilakukan secara bersamaan dengan pengumpulan telur serangga alami, yaitu dengan menyisir seluruh petakan lahan pertanaman kedelai.
Hasil pengumpulan larva dan gejala serangan larva Lepidoptera dibawa ke laboratorium untuk diidentifikasi. Ini dilakukan untuk mengetahui jenis-jenis
serangga yang berasosiasi dengan pertanaman kedelai sehingga hubungan antara struktur komunitas parasitoid telur dengan serangan hama dapat dipelajari.
Identifikasi
Identifikasi parasitoid dilakukan dengan tahapan-tahapan yang terdiri dari
perebusan, penataan parasitod pada kaca obyek, dan identifikasi. Dalam proses perebusan, parasitoid jantan dimasukkan dalam tabung reaksi berisi KOH 10% (±
5 ml). Tabung reaksi kemudian direbus dalam gelas ukur besar yang berisi air (± 1/3 volum) sampai air mendidih. Parasitoid jantan tersebut ditata di atas kaca
obyek yang telah ditetesi larutan Hoyer (satu tetes), kemudian ditutup dengan kaca penutup dan diberi label. Identifikasi terhadap genitalia jantan
bawah mikroskop stereo (Olympus BX 51), berdasarkan kunci identifikasi Alba (1988), Nixon (1937), dan Barrion & Litsinger (1989).
HASIL DAN PEMBAHASAN
Struktur Komunitas Parasitoid Telur
Struktur komunitas parasitoid telur pada pertanaman kedelai di Cikabayan, Bogor tersusun atas lima spesies parasitoid telur famili
Trichogrammatidae, dua spesies famili Scelionidae dan satu spesies famili Encyrtidae. Semua spesies parasitoid telur famili Trichogrammatidae dan satu
spesies famili Scelionidae diperoleh melalui pemasangan telur perangkap, sedangkan satu spesies famili Scelionidae lainnya dan satu famili Encyrtidae
diperoleh dari pengamatan telur serangga alami yang diperoleh dari pertanaman kedelai. Spesies parasitoid famili Trichogrammatidae yang diperoleh terdiri dari
Trichogrammatoidea cojuangcoi (Gambar lampiran 1), Trichogramma japonicum
(Gambar lampiran 2), Trichogrammatoidea armigera (Gambar lampiran 3),
Trichogramma chilonis (Gambar lampiran 4), dan satu spesies Trichogamma yang tidak teridentifikasi (Trichogramma sp.) (Gambar lampiran 5). Parasitoid famili Scelionidae terdiri dari Telenomus remus (Gambar lampiran 6), dan Gryon sp. (Gambar lampiran 7). Parasitoid famili Encyrtidae yang ditemukan belum teridentifikasi sampai tingkat spesies.
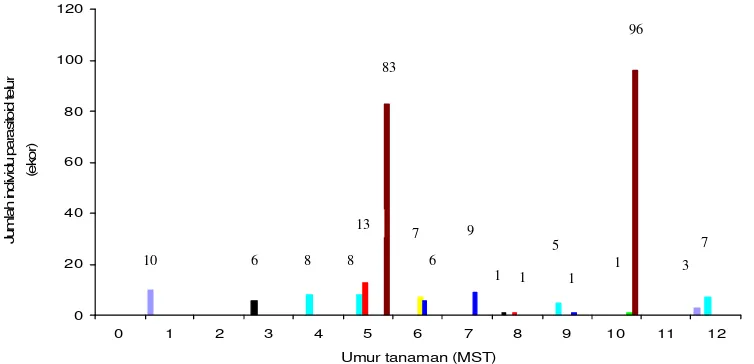
Dalam struktur komunitas pertanaman kedelai parasitoid, Gryon sp. merupakan parasitoid dengan jumlah individu terbanyak, yaitu mencapai 179 individu dari 265 individu parasitoid telur yang ditemukan atau sebesar 36,23%.
T. armigera merupakan spesies parasitoid telur dengan jumlah individu terbanyak dari hasil pemasangan telur perangkap, yaitu mencapai 28 individu atau sebesar
10, 57%. Satu individu parasitoid ditemukan tergolong dalam famili Encyrtidae. Dari hasil pemasangan telur perangkap diperoleh T. japonicum dan T. chilonis
dengan jumlah individu terendah, yaitu masing-masing sebanyak 7 individu atau sebesar 2, 64% (Gambar 2).
Kekayaan dan kelimpahan parasitoid telur tertinggi sekaligus melalui pemasangan telur perangkap diperoleh saat 5 MST sedangkan melalui
sekaligus diperoleh saat 10 MST (Gambar 3). Kekayaan dan kelimpahan parasitoid telur pada 10 MST diperoleh melalui pengamatan terhadap telur alamiah yang ditemukan di lapang sedangkan melalui pemasangan telur perangkap tidak ditemukan parasitoid telur. Tidak ditemukannya parasitoid telur melalui telur perangkap pada 10 MST disebabkan cuaca yang mendung selama pemasangan telur perangkap di lapang. Steenburg (1930 dalam Smith 1994) melaporkan, saat cuaca mendung aktivitas penerbangan parasitoid berkurang. Ditemukannya parasitoid melalui telur alami Piezodorus sp. (Hemiptera: Pentatomidae) dan telur Riptortus sp. (Hemiptera: Alydidae) diduga karena proses parasitisasi telah terjadi sebelumnya, yaitu ketika cuaca mendukung Gryon sp. dan parasitoid famili Encyrtidae untuk memarasit.
67.55%
Gambar 3 Kekayaan dan kelimpahan parasitoid telur C. cephalonica, S. litura dan serangga alami pada pertanaman kedelai umur 0-12 MST ( T. cojuangcoi, T. japonicum, T . armigera, Trichogramma sp., Gryon sp., T. chilonis, Telenomus remus, Encyrtidae)
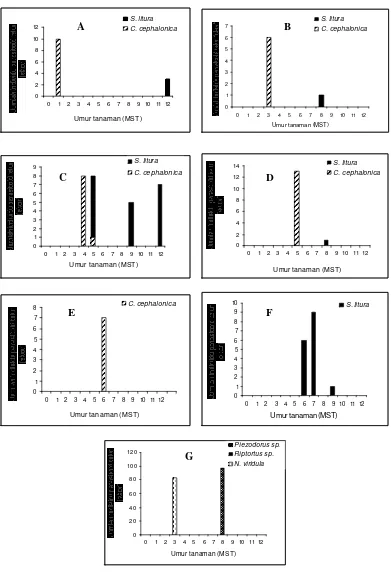
Parasitoid telur yang pertama kali ditemukan adalah Trichogrammatoidea
cojuangcoi (Gambar 4a). Parasitoid ini ditemukan pada pertanaman kedelai
ketika tanaman berumur 1 dan 12 MST. Kelimpahannya pada 1 MST diperoleh dari telur C. cephalonica sedangkan kelimpahan pada 12 MST diperoleh dari telur
S. litura. Fenomena kemunculan T. cojuangcoi sebagai satu-satunya parasitoid yang muncul saat pertama kali, yaitu saat umur tanaman 1 MST kemungkinan karena parasitoid ini menyukai habitat dengan tanaman yang masih kecil dan belum rimbun. Pada minggu berikutnya spesies ini tidak ditemukan lagi dan ini diduga karena kondisi lingkungan yang mungkin tidak sesuai bagi aktivitas penerbangan dan peletakan telur menyebabkan spesies-spesies parasitoid ini tidak selalu ditemukan pada setiap minggunya. Saat 12 MST, T. cojuangcoi kembali ditemukan. Ini dapat dijelaskan bahwa kemungkinan kondisi tanaman yang sudah tua sehingga tidak rimbun lagi karena banyaknya daun rontok dan diduga kondisi seperti ini disukai T. cojuangcoi. Namun, fenomena seperti ini masih perlu dikaji ulang untuk mengetahui pengaruh kondisi tanaman terhadap perilaku memarasit
T. cojuangcoi dan mengungkap penyebab penurunan kelimpahan parasitoid ini.
Pada umur tanaman 3 dan 8 MST diperoleh Trichogramma japonicum
(Gambar 4b). Pada 3 MST parasitoid diperoleh dari telur C. cephalonica,
sedangkan pada 8 MST diperoleh dari telur S.litura. Kelimpahan T. japonicum
pada 8 MST cenderung menurun dibandingkan dengan kelimpahannya pada 3
MST. Ditemukannya T. japonicum hanya pada 3 dan 8 MST kemungkinan disebabkan oleh berbagai hal, seperti distribusinya yang dipengaruhi oleh suhu
dan kelembaban optimal. Studi lebih lanjut terhadap distribusi dan penurunan kelimpahan T. japonicum pada 8 MST perlu dilakukan untuk mengetahui
penyebab terjadinya fenomena ini.
Trichogrammatoidea armigera adalah parasitoid telur yang paling sering
individu dari telur S. litura. Hal ini kemungkinan terkait dengan ditemukannya kelompok telur alami S. litura pada pertanaman kedelai pada 6 MST dan diduga imago S. litura telah eksis sejak 5 MST. Nordlund (1994) melaporkan bahwa
feromon sex yang dikeluarkan imago S. litura dapat menarik parasitoid untuk datang mendekat. Dugaan adanya feromon sex yang dikeluarkan imago S. litura
pada 5 MST inilah yang mungkin menyebabkan T. armigera diperoleh dari telur perangkap S. litura. Pengkajian lebih lanjut mengenai dugaan pengaruh feromon sex terhadap perilaku T. armigera yang memarasit telur perangkap pada 4, 5, 9, dan 12 MST tersebut perlu dilakukan.
Pada 12 MST, T. armigera kembali muncul dengan kelimpaha n yang lebih tinggi daripada saat 9 MST. Ditemukannya T. armigera dengan kelimpahan dan frekuensi penemuan yang tinggi menunjukkan parasitoid ini memiliki kamampuan
bertahan dan kemapanan yang baik. Bahagiawati (2003) dan Moy (2005) melaporkan bahwa T. armigera merupakan parasitoid yang dominan ditemukan
pada pertanaman jagung di Jawa Barat. Tingginya kelimpahan T. armigera
dibanding parasitoid telur famili Trichogrammatidae lainnya pada pertanaman
kedelai di Bogor juga menunjukkan bahwa parasiotid ini mungkin telah mendominasi struktur komunitas parasitoid telur pada beberapa pertanaman di
Jawa Barat, khususnya Bogor.
Trichogramma chilonis pada pertanaman kedelai hanya ditemukan sekali,
yaitu pada 6 MST (Gambar 4e). Ditemukannya parasitoid tersebut pada 6 MST diduga bahwa T. chilonis menyukai terbang di bagian atas kanopi pada habitat yang lebih rimbun. Perilaku seperti ini pernah dilaporkan Stinner et al. (1974
dalam Smith 1994) yang menyatakan bahwa Trichogramma minutum lebih
menyukai terbang di bagian atas kanopi daun sedangkan Trichogramma pretiosum lebih memilih terbang di bagian bawah kanopi daun.
0
Telenomus remus hanya diperoleh dari telur S. litura saat 6, 7, dan 9 MST (Gambar 4f). Kecenderungan T. remus terhadap telur S. litura karena T. remus
adalah parasitoid telur spesialis genus Spodoptera (Cave 2000). Ditemukannya parasitoid ini pada minggu-minggu terakhir karena Spodoptera litura sebagai inang spesifik T. remus hanya ditemukan pada minggu ke-6 sampai ke-9. Kelimpahan T. remus meningkat pada 7 MST. Ketersediaan dan kesesuaian inang bagi perkembangan T. remus menyebabkannya meleta kkan telur lebih banyak lagi daripada pada 6 MST. Pada 8 MST tidak ditemukan T. remus karena kondisi cuaca mendung sehingga aktivitas penerbangannya berkurang (Steenburg 1930
dalam Smith 1994). Kelimpahan T. remus menurun pada 9 MST, yaitu hanya 1
individu. Penurunan kelimpahan parasitoid T. remus setelah tidak ditemukan pada minggu sebelumnya merupakan fenomena yang perlu dikaji lebih lanjut.
Gryon sp. dan parasitoid famili Encyrtidae ditemukan pada pertanaman
kedelai melalui pengamatan langsung (Gambar 4g). Parasitod tersebut ditemukan
pada 5 dan 10 MST. Gryon sp. diperoleh dari telur Nezara viridula pada 5 MST sebanyak 83 individu dan dari telur Piezodorus sp. pada 10 MST sebanyak 96
individu. Gryon sp. ditunjukkan oleh Gambar lampiran 7. Telur N. viridula dan telur Piezodorus sp. terparasit Gryon sp. masing-masing ditunjukkan Gambar
lampiran 8 dan 9. Parasitoid famili Encyrtidae ditemukan dari telur Riptortus sp. dengan kelimpahan 1 individu. Telur Riptortus sp. ditunjukkan oleh Gambar
lampiran 10. Diduga parasitoid famili Encyrtidae dan Gryon sp. yang ditemukan dari telur Riptortus sp. dan Piezodorus sp. pada 10 MST telah memarasit pada minggu sebelumnya. Ini didukung oleh keluarnya kedua jenis parasitoid tersebut yang lebih cepat dari lama perkembangan kedua telur yang semestinya. Lama perkembangan Gryon sp. pada telur Piezodorus hybneri adalah 14 hari (Wardani 2001). Susila (1993) melaporkan bahwa lama perkembangan Ooencyrtus malayensis (Ferr.) (Hymenoptera: Encyrtidae) pada telur Riptortus linearis
berkisar 13-14 hari.
feromon sex yang dikeluarkan imago S. litura pada 5 MST. Diduga feromon sex
ini lebih kuat menarik parasitoid telur untuk datang mendekat dan memarasit telur
S. litura dibandingkan kairomon dari telur perangkap C. cephalonica. Pengkajian lebih lanjut terhadap fenomena ini perlu dilakukan untuk mengetahui faktor yang mempengaruhi perilaku parasitoid telur tersebut.
Pada 0 MST ketika tanaman kedelai belum ada di lapangan, tidak ditemukan parasitoid baik dari telur C. cephalonica maupun telur S. litura. Nordlund (1994) menjelaskan, keberadaan tanaman inang mampu menarik parasitoid dalam menemukan inang karena rangsangan kimia yang diproduksi tanaman inang tersebut. Saat 2 MST juga tidak ditemukan parasitoid karena saat pemasangan telur perangkap dilakukan, hujan turun. Smith (1994) dan Yu et al.
(1984) menjelaskan bahwa hujan merupakan salah satu komponen cuaca yang
berpengaruh terhadap keberhasilan pelepasan parasitoid karena aktivitas parasitoid sangat dipengaruhi oleh suhu. Parasitoid juga tidak ditemukan melalui
pemasangan telur perangkap pada 10 dan 11 MST karena cuaca mendung selama telur dipasang di lapangan. Steenburg (1930 dalam Smith 1994) melaporkan, saat
cuaca mendung aktivitas penerbangan parasitoid berkurang.
Telur Serangga Alami, Larva Lepidoptera dan Gejala Serangannya pada Pertanaman Kedelai
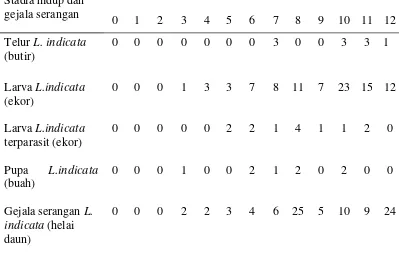
Pada pertanaman kedelai umur 0-12 MST banyak ditemukan telur, larva, pupa, dan gejala serangan hama penjalin daun Lamprosema indicata
(Lepidoptera: Pyralidae) hampir pada setiap minggu (Tabel 2). Tingginya frekuensi penemuan L. indicata menunjukkan bahwa hama ini merupakan hama dominan pada pertanaman kedelai di Cikabayan, Bogor. Larva L. indicata
ditunjukka n oleh Gambar lampiran 13. Selain L. indicata, ditemukan juga larva
dan gejala serangan ulat jengkal, telur dan larva S. litura, larva dan gejala serangan E. zinckenella, telur Riptortus sp. sehat dan terparasit, telur dan larva
ulat bulu, telur Hemiptera, dan telur Halticus bractatus (Hemiptera: Miridae) meskipun frekuensi penemuan dan jumlahnya tidak setinggi L. indicata.
Keberadaan L. indicata dapat dikenali dengan adanya jalinan beberapa daun secara bersama dan jalinan ini direkatkan oleh sutera yang diproduksi kelenjar
indicata biasanya terjadi pada 4-6 MST. Hasil pengamatan di lapangan dalam penelitian ini menunjukkan bahwa serangan terjadi ketika tanaman berumur 3 MST hingga 12 MST. Serangan larva instar akhir menyebabkan defoliasi pada daun dan hanya menyisakan tulang daun (Kalshoven 1981), sehingga apabila serangan berat sejak awal hingga menjelang panen dapat menurunkan produktivitas tanaman kedelai. Habisnya jaringan daun mengakibatkan proses fotosintesis terganggu. Hasil pengumpulan dari lapang juga menemukan larva L. indicata terparasit (Tabel 2). Pupa parasitoid tersebut ditunjukkan oleh Gambar lampiran 14. Identifikasi terhadap parasitoid yang telah dilakukan belum sampai tingkat spesies, namun diketahui bahwa parasitoid tersebut tergolong dalam famili Braconidae, Elasmidae, dan Ceraphronidae (Gambar lampiran 15; 16; 17). Laporan Rahardjo dan Supeno (1999) menyatakan bahwa Apanteles sp.
(Hymenoptera: Braconidae) merupakan parasitoid larva Lamprosema sp. yang umum ditemukan pada pertanaman kedelai di Pulau Lombok dan Sumbawa.
Tabel 2 Stadia hidup dan gejala serangan Lamprosema indicata pada pertanaman kedelai umur 0-12 MST
kelompok telur dan larva S. litura (Tabel 3). Larva dan gejala serangan ulat jengkal sering ditemukan, namun jumlahnya tidak setinggi L. indicata, dan frekuensi penemuannya tidak pada setiap minggu. Gejala serangan ulat ini dikenali dengan adanya kekhasan tulang daun yang ditinggalkannya. Serangan berat menyebabkan gangguan terhadap fotosintesis akibat habisnya jaringan daun yang dimakan sehingga dapat menurunkan hasil. Sullivan et al. (1994) melaporkan bahwa pada umumnya serangan larva ini berada di bawah tingkat kerusakan ekonomi.
Tabel 3 Stadia hidup dan gejala serangan serangga perusak daun kedelai umur 0-12 MST
Telur dan larva S. litura ditemukan secara berturut-turut pada 6-9 MST.
pembungaan dan pembentukan polong merupakan stadia kritis dari tanaman kedelai, sehingga jika serangan berat terjadi pada masa ini dapat menurunkan produktivitas tanaman kedelai.
Serangga hama perusak daun lain yang ditemukan adalah telur dan larva ulat bulu. Larva ditemukan pada 7 dan 9 MST masing-masing sebanyak satu ekor. Telurnya ditemukan pada 10 MST sebanyak satu kelompok telur (Gambar lampiran 11). Larva ini memakan jaringan daun dan menyebabkan defoliasi. Berdasarkan waktu penemuannya di lapangan, serangga ini ditemukan saat pembungaan dan pembentukan polong kedelai yang merupakan stadia kritis tanaman kedelai. Serangan berat pada masa tersebut dapat mengurangi produktivitas. Telur Halticus bractatus sebanyak 11 butir (dalam kelompok) juga ditemukan pada 8 MST. Nimfa dan imago serangga ini menyerang tanaman
kedelai dengan menghisap cairan daun dan batang tanaman. Hisapan pada bagian tersebut meninggalkan luka berupa spot kecil berwarna putih. Serangga ini
ditemukan pada 8 MST, dan bertepatan dengan saat pembungaan kedelai dan ini merupakan stadia kritis tanaman kedelai. Serangan berat saat 8 MS T dapat
menurunkan produksi kedelai.
Ketika tanaman berumur 8-11 MST, ditemukan serangga hama perusak
polong kedelai, yaitu larva dan gejala serangan E.zinckenella serta telur Riptortus
sp. (Tabel 4). Gejala E.zinckenella yang berupa bekas gerekan larva pada polong
kedelai ditemukan pada 8, 9, 11, dan 12 MST, sedangkan larvanya ditemukan pada 11 MST. Menurut Kalshoven (1981), E. zinckenella merupakan hama penting pada kedelai selain S. litura, Agrotis spp., dan Ophiomyia spp. Serangan berat mampu menurunkan produksi kedelai karena kerusakan yang ditimbulkan merupakan kerusakan secara langsung.
Telur serangga lain juga ditemukan sebanyak satu kelompok, berwarna putih agak pink, diletakkan secara berderet dalam dua barisan pada polong kedelai (Gambar lampiran 12). Telur berbentuk silinder pada bagian bawahnya, sedangkan bagian atas (tutup) datar dan berambut-rambut pendek.
Tabel 4 Stadia hidup dan gejala serangan serangga perusak polong kedelai umur 0-12 MST
Populasi dan serangan S. litura yang tidak setinggi L. indicata, ulat jengkal, dan E. zinckenella diperkirakan karena parasitoid yang berasosiasi dengan S. litura mampu me nekan populasinya. Berdasarkan perolehan parasitoid melalui pemasangan telur perangkap, T. armigera merupakan parasitoid dominan.
T. remus juga ditemukan dari pemasangan telur S. litura. Hal ini diperkuat oleh
hasil penelitian Yuliarti (2002) yang menunjukkan bahwa T. remus memarasit telur S. litura pada pertanaman kedelai. Ditemukannya T. armigera dari telur C.
cephalonica dan S. litura menunjukkan luasnya kisaran inang T. armigera. Keterangan ini juga dilaporkan Meilin (1999) yang menyatakan T. armigera
mampu memarasit Plutella xylostella (Linnaeus) (Lepidoptera: Plutellidae),
Crocidolomia binotalis (Zeller) (Lepidoptera: Pyralidae), Scirpophaga incertulas
(Walker) (Lepidoptera: Pyralidae), dan Etiella zinckenella (Treitsche) (Lepidoptera: Pyralidae). Tingginya populasi E. zinckenella diduga karena tidak
Terkait dengan hasil penemuan berbagai jenis parasitoid yang berhasil diperangkap serta hubungannya dengan serangan hama, maka jenis-jenis inang yang berasosiasi dengan tanaman kedelai beserta parasitoid telur yang pernah menyerangnya perlu disampaikan. Berdasarka n laporan Herlinda et al. (1999) bahwa T. chilonis menyerang H. armigera pada pertanaman kedelai. Herlinda et al. (1999) juga melaporkan pelepasan T. bactrae-bactrae untuk mengendalikan penggerek polong kedelai Etiella spp. pada pertanaman kedelai di Cianjur.
Yuliarti (2002) melaporkan bahwa, T. remus memarasit telur S. litura pada pertanaman kedelai di Cianjur dengan persentase parasitisasi mencapai 53, 80%.
Hubungan Antara Struktur Komunitas Parasitoid Telur dengan Tingkat Serangan dan Populasi Hama
Trichogramma spp. dan Trichogrammatoidea spp. sebagai parasitoid yang bersifat generalis, memiliki kisaran inang yang luas, sedangkan Telenomus remus
bersifat spesifik telur inang. Trichogrammatoidea spp. dan Trichogramma spp. selain dapat memarasit hama Lepidoptera, juga pernah dilaporkan mampu memarasit hama Diptera (Meilin 1999). T. remus merupakan parasitoid spesifik telur S. litura (Cave 2000). Rendahnya tingkat serangan hama-hama dari ordo Lepidoptera, kecuali L. indicata, kemungkinan besar disebabkan oleh adanya
parasitoid-parasitoid telur tersebut.
Tingginya frekuensi penemuan Lamprosema indicata (Tabel 2) dan tidak
ditemukannya telur L. indicata terparasit menunjukkan bahwa hama ini cukup dominan karena tidak ada parasitoid yang berasosiasi dengan telur L. indicata.
Belum pernah ada laporan sebelumnya yang menyebutkan bahwa
Trichogrammatoidea spp. dan Trichogramma spp. memarasit telur L. indicata.
Laporan sebelumnya menyebutkan bahwa di Pulau Lombok dan Sumbawa, larva
Lamprosema sp. dapat diparasit oleh Apanteles sp. (Hymenoptera: Braconidae)
(Rahardjo & Supeno 1999).
S. litura sebagai hama penting pada pertanaman kedelai ditemukan dengan
populasi dan serangan yang lebih rendah dari tingkat populasi dan serangan L.
indicata. Hal ini dapat dipaha mi bahwa pada pertanaman kedelai ditemukan