BIOLOGI, SIKLUS HIDUP DAN POTENSI PARASITOID
TELUR
Trissolcus
sp. PADA
Chrysocoris javanus
Westw., HAMA
TANAMAN JARAK PAGAR (
Jatropha curcas
L.)
YULIUS DIKA CIPTADI
DEPARTEMEN PROTEKSI TANAMAN
FAKULTAS PERTANIAN
INSTITUT PERTANIAN BOGOR
BOGOR
ABSTRAK
YULIUS DIKA CIPTADI. Biologi, Siklus Hidup, dan Potensi Parasitoid Telur Trissolcus sp. Pada Chrysocoris javanus Westw., Hama Tanaman Jarak Pagar (Jatropha curcas L.). Dibimbing oleh DHAMAYANTI ADIDHARMA.
Beberapa tahun terakhir, jarak pagar (Jatropha curcas L.) mulai diusahakan dalam skala luas dan ditanam secara monokultur karena potensinya sebagai sumber bahan bakar nabati. Cara budi daya seperti ini dapat menyebabkan perubahan keadaan agroekosistem yaitu pengurangan keanekaragaman hayati sehingga menyediakan lingkungan yang sesuai untuk perkembangan hama. Salah satunya adalah hama Chrysocoris javanus. Hama ini menyerang buah sehingga menyebabkan buah rusak dan tidak dapat dipanen. Musuh alami seperti parasitoid telah ditemukan menyerang telur C. javanus. Tujuan dari penelitian ini adalah untuk mengetahui biologi, siklus hidup dan potensi Trissolcus sp. sebagai parasitoid telur pada inang C. javanus. Telur C. javanus direkatkan pada pias kertas dengan menggunakan gom arab kemudian dimasukkan ke dalam tabung gelas yang telah berisi parasitoid betina. Penelitian tentang bentuk, ukuran, warna, dan ciri-ciri lain setiap stadium pradewasa dilakukan dengan membedah telur setiap 24 jam kecuali stadia larva setiap 12 jam dan diamati menggunakan mikroskop stereo. Penelitian tentang biologi, siklus hidup dan potensi Trissolcus sp. sebagai parasitoid telur pada inang C. javanus dilakukan tanpa pembedahan telur dan pengamatan dilakukan terhadap keperidian, produksi telur harian, potensi produksi telur, lama hidup parasitoid, lama perkembangan, kemunculan imago, nisbah kelamin dan lama masa reproduksi. Hasil penelitian menunjukkan bahwa telur parasitoid Trissolcus sp. bertipe stalked, larva instar pertama bertipe teleaform, larva instar ketiga bertipe hymenopteriform. Lama stadium telur Trissolcus sp. satu hari, larva empat hari, prapupa satu hari, pupa lima hari, lama hidup imago jantan 23,70 ± 9,49 hari dan betina 17,40 ± 7,38 hari. Keperidian 95,70 ± 18,34 butir, produksi telur harian 15,76 ± 2,63 butir, potensi produksi telur 129,30 ± 25,96 butir, lama masa reproduksi 6,10 ± 0,88 hari, nisbah kelamin 1 : 3,83. Tingkat parasitisasi 19,14 ± 3,67 % dan keberhasilan hidup mencapai 86,71 ± 3,77 %. Parasitoid telur Trissolcus sp. berpotensi dikembangkan sebagai parasitoid telur untuk mengendalikan kepik jarak pagar C. javanus.
BIOLOGI, SIKLUS HIDUP DAN POTENSI PARASITOID
TELUR
Trissolcus
sp. PADA
Chrysocoris javanus
Westw., HAMA
TANAMAN JARAK PAGAR (
Jatropha curcas
L.)
YULIUS DIKA CIPTADI
Skripsi
sebagai salah satu syarat untuk memperoleh gelar Sarjana Pertanian pada
Departemen Proteksi Tanaman
DEPARTEMEN PROTEKSI TANAMAN
FAKULTAS PERTANIAN
INSTITUT PERTANIAN BOGOR
BOGOR
Judul Skripsi : Biologi, Siklus Hidup, dan Potensi Parasitoid Telur Trissolcus sp. Pada Chrysocoris javanus Westw., Hama Tanaman Jarak Pagar (Jatropha curcas L.)
Nama : Yulius Dika Ciptadi
NIM : A34070044
Disetujui, Pembimbing
Dhamayanti Adidharma, Ph.D NIP. 19481006 197903 2 001
Diketahui, Ketua Departemen
Prof. Dr. Ir. Dadang, MSc NIP 19640204 199002 1 002
RIWAYAT HIDUP
Penulis dilahirkan di Yogyakarta pada tanggal 16 Maret 1989. Penulis merupakan anak pertama dari dua bersaudara, dari pasangan Stefanus Padmono Ciptadi dan Henrica Lastyandari. Tahun 2007 penulis menyelesaikan pendidikannya di SMA Pangudi Luhur Van Lith Muntilan dan pada tahun yang sama penulis diterima di Institut Pertanian Bogor melalui jalur Undangan Seleksi Masuk IPB (USMI). Penulis mengambil mayor Proteksi Tanaman dan Minor Agronomi dan Hortikultura.
PRAKATA
Puji syukur penulis panjatkan kepada kehadirat Tuhan yang Maha Esa atas limpahan rahmat-Nya, sehingga penulis dapat menyelesaikan tugas akhir yang berjudul Biologi, Siklus Hidup dan Potensi Parasitoid Telur Trissolcus sp. Pada Chrysocoris javanus Westw., Hama Tanaman Jarak Pagar (Jatropha curcas L.).
Penulis sangat menyadari bahwa tidak dapat berbuat maksimal dalam menyelesaikan tugas akhir ini tanpa bimbingan, dukungan, dan bantuan dari berbagai pihak. Penulis menyampaikan ucapan terima kasih kepada :
1. Dhamayanti Adidharma, Ph.D sebagai pembimbing yang telah
membimbing dan mengarahkan penulis dalam penyusunan tugas akhir 2. Dr. Ir. Gede Suastika, MSc. selaku dosen penguji tamu atas
masukkannya dalam skripsi ini
3. Dr. Ir. Nina Maryana, Msi. selaku moderator seminar dan atas bantuannya dalam identifikasi parasitoid
4. Keluarga di Cianjur yang telah memberikan kasih sayang, perhatian, nasehat dan doa
5. Anggota laboratorium Bioekologi Parasitoid dan Predator: mbak Adha, mbak Nita, kak Putri, kak Ai, kak Eldy, Gama
6. Teman-teman seperjuangan HPT’ers 44: eter, latip, harwan, taher, lutfi, doli, sista, anik, sherli, mia, osmond, iky, sanny, nelly dan semua yang tidak bisa disebutkan satu persatu
7. Semua pihak yang telah membantu dalam penyelesaian skripsi ini Penulis menyadari bahwa tugas akhir ini jauh dari kesempurnaan. Oleh karena itu, penulis mengharapkan kritik dan saran yang membangun sehingga tugas akhir ini bermanfaat bagi masyarakat secara umum maupun perkembangan IPTEK khususnya dalam bidang proteksi tanaman.
Bogor, Desember 2011
DAFTAR ISI
Halaman
DAFTAR TABEL ... viii
DAFTAR GAMBAR ... ix
DAFTAR LAMPIRAN ... x
PENDAHULUAN... 1
Latar Belakang ... 1
Tujuan Penelitian ... 3
Manfaat Penelitian ... 3
TINJAUAN PUSTAKA... 4
Tanaman Jarak Pagar (Jatropha curcas L.) ... 4
Hama Chrysocoris javanus Westw. ... 5
Parasitoid Trissolcus sp. ... 6
Tahap Perkembangan Parasitoid Famili Scelionidae ... 6
Telur ... 6
Larva ... 7
Prapupa dan Pupa ... 7
Imago ... 8
BAHAN DAN METODE ... 9
Tempat dan Waktu Penelitian ... 9
Metode Penelitian ... 9
Survei Awal Bioekologi C. javanus dan Trissolcus sp. ... 9
Perbanyakan C. javanus ... 9
Perbanyakan Parasitoid Telur Trissolcus sp. ... 10
Penelitian Biologi dan Siklus Hidup Parasitoid Trissolcus sp. pada Inang C. javanus ... 11
Penelitian Potensi Trissolcus sp. Sebagai Parasitoid Telur pada Inang C. javanus ... 12
HASIL DAN PEMBAHASAN ... 15
vii
Biologi dan Siklus Hidup Parasitoid Trissolcus sp. pada Inang
C. javanus ... 15
Telur ... 16
Larva ... 16
Prapupa... 18
Pupa ... 18
Imago ... 20
Potensi Trissolcus sp. Sebagai Parasitoid Telur pada Inang C. javanus ... 22
Lama Perkembangan ... 22
Keberhasilan Hidup... 23
Parameter Kehidupan Parasitoid ... 24
Kemunculan Imago Trissolcus sp. ... 26
Nisbah Kelamin... 27
Kemampuan Reproduksi ... 28
Persentase Parasitisasi ... 29
KESIMPULAN DAN SARAN ... 32
Kesimpulan ... 32
Saran ... 32
DAFTAR PUSTAKA ... 33
DAFTAR TABEL
Halaman
Tabel 1 Parasitoid yang ditemukan di lapang ... 15
Tabel 2 Lama perkembangan parasitoid Trissolcus sp. ... 20
Tabel 3 Jumlah imago Trissolcus sp. jantan dan betina yang muncul ... 23
Tabel 4 Keberhasilan hidup imago Trissolcus sp. ... 24
Tabel 5 Parameter kehidupan imago betina Trissolcus sp. ... 25
Tabel 6 Nisbah kelamin keturunan F1 dari imago betina Trissolcus sp. ... 28
Tabel 7 Kemampuan reproduksi imago betina Trissolcus sp. ... 29
DAFTAR GAMBAR
Halaman
Gambar 1 Lahan jarak pagar di Leuwikopo ... 9
Gambar 2 Perbanyakan C. javanus ... 10
Gambar 3 Mikroskop stereo dengan kamera digital tipe OLYMPUS 11D, digunakan untuk pemotretan semua stadia Trissolcus sp... 12
Gambar 4 Penelitian di laboratorium ... 14
Gambar 5 Telur Trissolcus sp. ... 16
Gambar 6 Larva Trissolcus sp... 17
Gambar 7 Prapupa Trissolcus sp. ... 18
Gambar 8 Pupa Trissolcus sp. ... 19
Gambar 9 Imago Trissolcus sp. ... 21
Gambar 10 Kemunculan imago Trissolcus sp. ... 27
Gambar 11 Reproduksi harian imago betina Trissolcus sp. ... 30
DAFTAR LAMPIRAN
Tabel
Halaman
Tabel Lampiran 1 Ukuran pradewasa dan imago Trissolcus sp. ... 38
Gambar Halaman Gambar Lampiran 1 Perbanyakan C. javanus ... 38
Gambar Lampiran 2 Imago parasitoid Trissolcus sp. ... 39
Gambar Lampiran 3 Telur C. javanus terparasit ... 40
Gambar Lampiran 4 Telur dan larva C. javanus tidak terparasit ... 41
PENDAHULUAN
Latar Belakang
Kebutuhan bahan bakar minyak Indonesia saat ini cukup tinggi. Konsumsi energi terbesar adalah minyak bumi yang mencapai 54,4 % (Krisnamurthi 2005). Menurut data Direktorat Lalu Lintas Angkutan Jalan – Ditjen Perhubungan Darat tahun 2009, kebutuhan solar mencapai 12.382.242 liter per hari dan jumlah kendaraan akan mencapai 110 juta kendaraan pada tahun 2020. Sedangkan cadangan energi fosil hanya akan bertahan kurang lebih 18 tahun (Krisnamurthi 2005).
Melihat kenyataan tersebut, perlu adanya tindakan untuk mengurangi ketergantungan terhadap bahan bakar minyak dan menanggulangi masalah harga minyak yang makin meningkat dan cadangan yang makin menipis. Pemerintah telah menerbitkan Peraturan Presiden Republik Indonesia Nomor 5 Tahun 2006 tentang Kebijakan Energi Nasional untuk mengembangkan sumber energi alternatif sebagai pengganti bahan bakar minyak. Presiden Indonesia juga menerbitkan Instruksi Presiden Nomor 1 Tahun 2006 tentang penyediaan dan pemanfaatan bahan bakar nabati (biofuel) sebagai energi alternatif.
Hambali (2005) menyatakan salah satu sumber minyak nabati yang sangat prospektif sebagai bahan baku biodiesel adalah biji jarak pagar yang bernama latin Jatropha curcas Linn. Program pemerintah untuk mensubtitusi 5% minyak bumi sampai tahun 2025 dapat memacu penanaman jarak pagar dalam skala luas, monokultur dan intensif. Cara budi daya seperti ini dapat menyebabkan perubahan keadaan ekosistem yaitu berkurangnya keanekaragaman hayati. Kondisi ini akan menyediakan lingkungan yang sesuai untuk perkembangan hama (Dadang 2005).
Kepik C. javanus menjadi salah satu hama penting pada pertanaman jarak pagar karena menyerang tanaman jarak fase generatif yaitu pada saat pembungaan, menjelang pembentukan buah dan saat pembentukan buah. C. javanus menyerang tanaman dengan cara menghisap buah jarak sehingga menimbulkan kerusakan pada kapsul buah yang sedang berkembang. Gejala yang ditimbulkan pada buah jarak pagar yaitu adanya bekas tusukan pada buah dan buah berwarna coklat kehitaman kemudian membusuk dan mengering (Rumini dan Karmawati 2007; Qodir 2010). Berbeda dengan di Indonesia, serangga anggota famili Scutelleridae yang menyerang jarak pagar di India adalah Scutellera nobilis Fabr. Sedangkan Pachycoris klugii Burmeister dan Agonosoma trilineatum Fabr. menyerang jarak pagar di Nicaragua. Serangga ini dilaporkan dapat menyebabkan gugurnya bunga, aborsi buah dan kerusakan benih. (Shanker dan Dhyani 2006). Bahkan A. trilineatum menjadi agens hayati untuk mengendalikan gulma famili Euphorbiaceae, Jatropha gossypiifolia (Bebawi et al. 2007; Heard et al. 2009).
3
Tujuan Penelitian
Penelitian ini bertujuan mengetahui biologi, siklus hidup, dan potensi parasitoid telur Trissolcus sp. pada inang C. javanus.
Manfaat Penelitian
Tanaman Jarak Pagar (Jatropha curcas Linn.)
Tanaman jarak pagar (Jatropha curcas Linn.) termasuk dalam famili Euphorbiaceae merupakan salah satu tanaman yang baik sebagai sumber bahan bakar nabati (Prihandana dan Hendroko 2006; Hambali et al. 2007). Jarak pagar berasal dari Amerika Tengah dan didistribusikan oleh pelaut Portugis melalui pulau Cape Verde ke berbagai negara di Afrika dan Asia (Hambali 2005). Jarak pagar mempunyai 4 varietas, yaitu varietas Cape Verde, Nicaragua, Ife-Nigeria, dan varietas tidak beracun Mexico. Varietas Cape Verde merupakan varietas yang umum terdapat di seluruh dunia dan bersifat toksik karena mengandung senyawa lektine dan ester forbol (Henning 2005 dalam Nurcholis dan Sumarsih 2007).
5 Hama Chrysocoris javanus Westw.
C. javanus memiliki nama umum kepik buah jarak (Sosromarsono et al. 2007). Tubuhnya berbentuk perisai yang khas dengan skutelum yang berkembang dengan baik menutupi abdomen, warna tubuh imago cerah dan mencolok dengan elitra berwarna merah dan corak hitam. Serangga ini memiliki antena tiga ruas, lebih panjang dari kepala (Dadang 2006; Kalshoven 1981). Telur berbentuk silinder seperti drum, bagian bawah datar sedangkan bagian atas cembung. Telur berdiameter 1,24 mm dan tinggi 1,34 mm. Telur yang baru diletakkan berwarna krem terang dan ada yang berwarna agak kehijauan. Telur diletakkan secara berkelompok di bawah permukaan daun, pada batang atau ranting dan pada permukaan buah jarak pagar saat pagi hingga menjelang siang. Nimfa tubuhnya berwarna hitam dengan bintik merah, kuning, dan hijau mengkilat, sementara bagian dorsal toraks berwarna hijau metalik. Ukuran tubuh imago betina relatif lebih besar dari pada imago jantan, yaitu panjang 17,65 mm dan lebar 9,55 mm sedangkan ukuran tubuh jantan panjang 15,95 mm dan lebar 8,1 mm. Kemampuan reproduksi cukup tinggi, satu kelompok telur terdiri dari 28 sampai 126 butir dengan rerata 84,80 butir (Qodir 2010). Siklus hidupnya berkisar 60-80 hari (Rumini dan Karmawati 2007).
2006; Rumini dan Karmawati 2007).
Parasitoid Trissolcus sp.
Trissolcus sp. merupakan parasitoid yang termasuk ke dalam Superfamili Platygastroidea Famili Scelionidae Subfamili Telenominae (Rajmohana 2006). Semua spesies yang telah diketahui dalam famili ini merupakan parasitoid telur dan hidup pada berbagai habitat (Hagen 1973; Austin et al. 2005; Driesche et al.. 2008). Trissolcus sp. telah menjadi spesies yang penting dalam penelitian maupun praktek pengendalian hayati serangga hama khususnya serangga ordo Hemiptera (Awan et al. 1990; Weber et al. 1996; Justo et al. 1997; Koçak dan Kilinçer 2003; Arakawa et al. 2004; Tohir 2004; Laumann et al. 2008)
Tahap Perkembangan Parasitoid Famili Scelionidae
Parasitoid famili Scelionidae terbagi pada tiga kelompok subfamili yaitu Scelioninae, Telesinae, dan Telenominae (Krombein 1979; Masner 1993). Parasitoid ini dapat menyerang telur serangga ordo Lepidoptera, Hemiptera, Orthoptera, Diptera (Tabanidae) dan Arachnida. Adakalanya beberapa spesies menyerang telur serangga ordo Coleoptera dan Neuroptera (Clausen 1940).
Imago parasitoid famili Scelionidae hidup secara soliter (Clausen 1940), umumnya berada pada lingkungan terbuka dan terpapar sinar matahari seperti padang rumput, gurun pasir, hutan, tanah, dan air (Masner 1993).
Telur
7 Larva
Hagen (1973) menyatakan hampir semua parasitoid yang meletakkan telur dan berkembang di dalam inang mengalami perkembangan hipermetamorfosis. Artinya, larva instar awal memiliki perbedaan bentuk dengan larva instar akhir. Jumlah instar larva sangat beragam antar genus dan spesies. Pada larva ektoparasit terdapat lima instar dan pada larva endoparasit kurang dari lima instar.
Clausen (1940) menyebutkan ada 14 tipe larva instar awal pada parasitoid dari ordo Hymenoptera. Larva instar awal memiliki variasi bentuk yang terbesar dalam perkembangan pradewasa parasitoid. Tipe larva instar awal tersebut adalah sacciform, hymenopteriform, caudate, vesiculate, encyrtiform, mandibulate, teleaform, microtype, mymariform, planidium, agriotypiform, polypodeiform, eucoiliform dan cyclopiform.
Larva instar pertengahan tidak memilki perbedaan karakteristik yang mendasar dari bentuk larva instar awalnya (Hagen 1973). Bentuk sebenarnya dari larva instar kedua serangga famili Scelionidae masih dalam perdebatan. Pada beberapa spesies hanya ditemukan dua instar larva, ini ditentukan berdasarkan persamaan bentuk antara larva instar kedua dan ketiga (Clausen 1940). Larva instar akhir pada parasitoid dari ordo Hymenoptera adalah hymenopteriform (Hagen 1973).
Prapupa dan Pupa
(Borror et al. 1996). Serangga ordo Hymenoptera khususnya parasitoid berpupa di dalam inang. Seringkali larva parasitoid berpupa di dalam kokon atau puparium inang atau berpupa di dalam terowongan yang dibuat inang dimana inang tersebut terlindungi. Parasitoid yang hidup di pada inang yang tersembunyi biasanya tidak berkokon (Hagen 1973).
Imago
BAHAN DAN METODE
Tempat dan Waktu Penelitian
Penelitian dilaksanakan di lapang dan di Laboratorium Bioekologi Parasitoid dan Predator Departemen Proteksi Tanaman Institut Pertanian Bogor, pada bulan Mei sampai Agustus 2011.
Metode Penelitian
Survei Awal Bioekologi C. javanus dan Trissolcus sp.
Survei awal tentang bioekologi serangga uji parasitoid Trissolcus sp dan C. javanus dilakukan di kebun jarak pagar di Kebun Percobaan IPB desa Leuwikopo, kecamatan Darmaga, kabupaten Bogor. Informasi tentang cara hidup, perilaku dan tempat peletakkan telur serangga uji ini diperlukan sebagai dasar dalam teknik perbanyakannya.
Gambar 1 Lahan jarak pagar di Leuwikopo Perbanyakan C. javanus
tanaman jarak pagar dalam polibag berukuran 18 cm x 25 cm dengan media tanah dan selanjutnya dimasukkan ke dalam sangkar berkerangka kayu berdinding kain kasa dengan ukuran 45 cm x 45 cm x 60 cm. Setiap hari tanaman disiram untuk menjaga kesegaran tanaman. Sebagai pakan C. javanus, buah jarak pagar segar digantungkan pada tanaman jarak. Pada setiap kaki-kaki sangkar diberi alas wadah plastik yang diisi oli untuk mencegah semut dan serangga lain masuk kedalam sangkar. Telur-telur C. javanus yang dihasilkan diambil setiap hari sebagai bahan penelitian.
Gambar 2 Perbanyakan C. javanus, (a) sangkar pemeliharaan nimfa dan imago, (b) kelompok telur C. javanus yang diletakkan di bawah permukaan daun tanaman jarak pagar dalam sangkar.
Perbanyakan Parasitoid Telur Trissolcus sp.
Parasitoid Trissolcus sp. diperoleh dengan cara mengumpulkan kelompok telur kepik C. javanus terparasit dari pertanaman jarak pagar di desa Leuwikopo, Darmaga dan desa Lulut, Citeureup. Telur-telur C. javanus diambil beserta tempat melekatnya telur-telur tersebut (daun, ranting atau buah), dimasukkan ke dalam kantung plastik dan diberi label keterangan asal telur dan waktu pengambilan. Di laboratorium, telur tersebut dimasukkan ke dalam tabung reaksi berukuran 0,5 cm x 10 cm yang ditutup kapas. Imago parasitoid yang keluar dipelihara dengan diberi pakan larutan madu 10% yang dioleskan pada permukaan dinding dalam tabung. Imago-imago yang baru muncul tersebut akan digunakan untuk perbanyakan.
11
parasitoid Trissolcus sp. berumur satu hari dan telah mengalami kopulasi. Telur inang yang telah terparasit dipindahkan ke dalam tabung reaksi lainnya. Imago parasitoid yang keluar dipelihara dengan diberi pakan larutan madu 10% yang dioleskan pada permukaan dinding dalam tabung. Tabung-tabung tersebut disusun rapi diatas papan kayu yang ditopang wadah plastik yang diisi oli untuk mencegah semut dan serangga lain mencapai tabung. Seluruh tabung disimpan dalam ruangan dengan suhu 28,18 ± 0,59 0C dan kelembaban relatif 56,33 ± 4,27 %. Imago-imago yang baru muncul tersebut akan digunakan untuk penelitian.
Penelitian Biologi dan Siklus Hidup Parasitoid Trissolcus sp. pada Inang C. javanus
Telur C. javanus umur satu hari sebanyak 5 butir yang diambil dari pembiakan massal direkatkan pada pias kertas karton ukuran 0,5 x 2 cm dengan perekat gom arab. Pias ini dimasukkan ke dalam tabung gelas yang berisi sepasang parasitoid Trissolcus sp. berumur satu hari dan sudah mengalami kopulasi. Jumlah lima butir telur C. javanus yang digunakan dimaksudkan untuk mendapatkan waktu peletakan telur yang bersamaan sehingga mendapatkan perkembangan pradewasa yang sama tiap harinya. Parasitoid diberi pakan madu 10% yang dioles pada dinding tabung gelas yang ditutup dengan kapas. Tabung-tabung tersebut disusun rapi diatas papan kayu yang ditopang wadah plastik yang diisi oli untuk mencegah semut dan serangga lain mencapai tabung. Seluruh tabung disimpan dalam ruangan dengan suhu 28,18 ± 0,59 0C dan kelembaban relatif 56,33 ± 4,27 %.
Gambar 3 Mikroskop stereo dengan kamera digital tipe OLYMPUS 11D, digunakan untuk pemotretan semua stadia Trissolcus sp.
Penelitian Potensi Trissolcus sp. Sebagai Parasitoid Telur pada Inang C. javanus
13
dilakukan terhadap keperidian dan produksi telur harian, potensi produksi telur, lama hidup, lama perkembangan, jumlah imago Trissolcus sp. yang muncul, nisbah kelamin dan lama masa reproduksi.
Keperidian dan Produksi Telur Harian. Keperidian dihitung dari jumlah total telur yang diletakkan oleh imago betina Trissolcus sp. selama masa hidupnya dalam telur inang C. javanus. Produksi telur harian dihitung berdasarkan jumlah telur yang dihasilkan oleh imago Trissolcus sp. betina setiap harinya selama masa oviposisi.
Potensi Produksi Telur. Potensi produksi telur merupakan total jumlah telur yang dihasilkan dan sisa telur dalam ovari Trissolcus sp.
Lama Hidup. Lama hidup Trissolcus sp. diamati mulai dari hari pertama imago muncul sampai imago tersebut mati.
Lama Perkembangan. Lama perkembangan dihitung dari hari pertama Trissolcus sp. meletakkan telur sampai waktu pemunculan imago.
Nisbah Kelamin. Perbandingan jumlah imago jantan dan betina Trissolcus sp. dihitung dari total seluruh imago keturunan F1 yang muncul.
Lama Masa Reproduksi. Masa reproduksi dihitung mulai hari pertama imago betina Trissolcus sp. meletakkan telur sampai hari terakhir imago betina meletakkan telur.
Persentase Parasitisasi. Persentase parasitisasi diketahui dengan cara menghitung banyaknya telur inang yang terparasit dengan menggunakan rumus berikut :
Persentase Keberhasilan Hidup. Keberhasilan hidup Trissolcus sp. diketahui dengan cara menghitung banyaknya imago yang muncul dari kelompok telur terparasit dengan menggunakan rumus :
HASIL DAN PEMBAHASAN
Parasitoid yang ditemukan di Lapang
Selama survei pendahuluan, telah ditemukan tiga jenis parasitoid yang tergolong dalam famili Eupelmidae, Pteromalidae dan Scelionidae. Data pada Tabel 1 menunjukkan bahwa parasitoid famili Scelionidae selalu ditemukan pada kelompok telur C. javanus yang terparasit. Hasil ini menunjukkan bahwa parasitoid ini cukup dominan di lapang. Hal yang sama juga dikemukakan oleh Qodir (2010) yaitu bahwa parasitoid ini selalu ditemukan di setiap pengamatan dan selalu berperan dalam memarasit telur C. javanus. Oleh karena itu pemahaman biologi, siklus hidup dan potensi parasitoid Trissolcus sp. dari famili Scelionidae ini penting untuk membangun strategi pengelolaan hama C. javanus yang efektif.
Tabel 1 Parasitoid yang ditemukan di lapang
Kelompok Telur Eupelmidae Pteromalidae Scelionidae
1 0 0 43
2 22 0 23
3 10 24 11
Jumlah 32 24 77
Biologi dan Siklus Hidup Trissolcus sp. pada Inang C. javanus
Telur
Telur Trissolcus sp. dalam ovari berbentuk lonjong, berwarna putih susu berukuran panjang 0,25 – 0,34 mm dan lebar 0,05 – 0,12 mm sedangkan telur Trissolcus sp. yang ditemukan pada telur C. javanus yang dibedah berbentuk bulat telur, warna telur putih susu dengan ukuran panjang 0,35 - 0,43 mm dan lebar 0,15 - 0,2 mm. Pada kedua telur, terdapat tangkai (stalk) berbentuk lonjong meruncing dengan panjang 0,14 ± 0,01 mm pada salah satu ujungnya (Gambar 5). Menurut Clausen (1940) telur tersebut bertipe stalked.
Gambar 5 Telur Trissolcus sp.; (a) dalam ovari, (b) 12 jam setelah peletakkan telur (SPT).
Larva
Larva Trissolcus sp. dijumpai pada hari pertama setelah telur diletakkan. Perkembangan larva dapat dibedakan berdasarkan bentuk dan ukuran. Larva instar pertama memiliki abdomen berbentuk bulat seperti bola hingga agak lonjong menyerupai buah pir, berwarna putih keruh agak transparan, aktif bergerak, dan memilki mandibel besar berbentuk seperti kait yang mengarah ke bagian ventral (Gambar 6a, 6b, dan 6c). Segmen tubuh tidak terlihat jelas namun pembagian tubuh larva menjadi dua bagian terlihat jelas. Mandibel terdapat pada bagian anterior yang disebut kepala (Henriksen, Bakkendorf dan Pagden 1934 dalam Clausen 1940) atau cephalothorax (Noble dan Kamal dalam Clausen 1940). Larva instar pertama berukuran panjang 0,37 - 0,85 mm dan lebar 0,14 - 0,57 mm. Umur larva instar pertama adalah 36 jam. Clausen (1940) menggolongkan larva ini ke dalam tipe teleaform.
17
6e). Bentuk tubuh larva instar kedua bulat, berwarna putih keruh agak kekuningan. Perubahan bentuk, warna dan ukuran terjadi pada 72 jam setelah peletakkan telur (SPT). Larva berukuran panjang 0,85 - 1,23 mm dan lebar 0,57 - 1,08 mm. Umur larva instar kedua adalah 24 jam. Larva instar kedua parasitoid dari ordo Hymenoptera biasanya menyerupai bentuk larva instar akhirnya yaitu bagian kepala dan mandibel menghilang (Hagen 1973). Pada famili Scelionidae bentuk larva instar kedua yang sebenarnya masih dalam perdebatan (Clausen 1940).
Gambar 6 Larva Trissolcus sp.; (a) 24 jam SPT, mandibel terlihat jelas, (b) 36 jam SPT, (c) 48 jam SPT (d) 60 jam SPT mandibel mulai menghilang, (e) 72 jam SPT mandibel menghilang (f) 84 jam SPT, (g) 96 jam SPT, dan (h) 108 jam SPT.
1940). Stadia larva berlangsung selama empat hari (Tabel 2). Memasuki hari ke 5 setelah peletakkan telur, larva parasitoid berkembang menjadi prapupa.
Prapupa
Prapupa dijumpai pada pembedahan 120 jam setelah peletakkan telur (Gambar 7). Stadium ini dimulai ketika larva instar akhir telah berhenti makan dan hampir tidak menunjukkan pergerakan tubuh. Tubuh prapupa Trissolcus sp. berwana putih susu agak kekuningan, segmen tubuh terlihat dan struktur tubuh tidak selunak stadium larva. Tubuh prapupa Trissolcus sp. memilki ukuran panjang 1,15 - 1,38 mm dan lebar 0,85 - 1,08 mm. Pada saat pengamatan, larva memasuki fase eonymph dengan ciri-ciri bentuknya masih menyerupai larva instar terakhir namun lebih mengembang atau menggelembung, dan seringkali ditandai perubahan warna larva dari putih kekuningan menjadi putih buram (Morris 1937 dalam Hagen 1973). Stadium prapupa berlangsung selama satu hari (Tabel 2).
Gambar 7 Prapupa Trissolcus sp.; (a) lateral, (b) ventral.
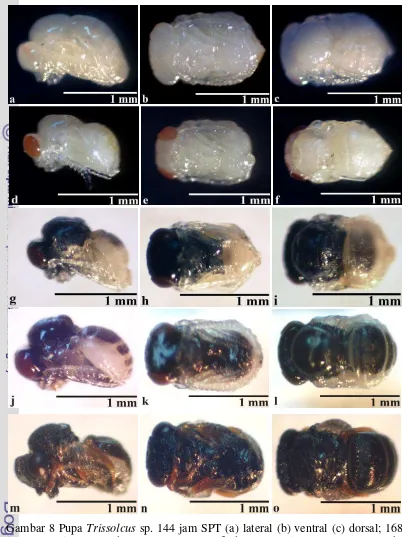
Pupa
19
Gambar 8 Pupa Trissolcus sp. 144 jam SPT (a) lateral (b) ventral (c) dorsal; 168 jam SPT (d) lateral (e) ventral (f) dorsal; 192 jam SPT (g) lateral (h) ventral (i) dorsal; 216 jam SPT (j) lateral (k) ventral (l) dorsal; 240 jam SPT (m) lateral (n) ventral (o) dorsal.
Trissolcus sp. pada pengamatan 168 jam setelah peletakkan telur 1,31 - 1,38 mm dan lebar 0,85 - 0,88 mm.
Pupa Trissolcus sp. mulai berubah warna menjadi hitam 192 jam setelah peletakkan telur. Perubahan warna dimulai dari bagian kepala dan torak yang semula putih keruh menjadi hitam namun bagian abdomen belum berwarna hitam (Gambar 8g, 8h, dan 8i). Organ tubuh sudah lengkap dan bakal sayap mulai terlihat jelas. Pupa berukuran panjang 1,37 - 1,38 mm dan lebar 0,91 - 0,92 mm.
Pada pengamatan 216 jam setelah peletakkan telur, tubuh pupa Trissolcus sp. mulai menghitam hingga bagian abdomen (Gambar 8j, 8k, dan 8l). Tungkai dan antena berwarna putih bening, sayap berwarna putih keruh dan warna mata menjadi merah kehitaman. Pupa berukuran panjang 1,37 - 1,40 mm dan lebar 0,91 - 0,92 mm.
Warna tubuh pupa Trissolcus sp. menghitam sempurna 240 jam setelah peletakkan telur (Gambar 8m, 8n, dan 8o). Mata berwarna hitam. Femur, tibia dan tarsus berwarna coklat terang. Sayap belum terbentuk sempurna dan masih berwarna putih keruh. Pupa masih belum bergerak namun sudah memasuki fase akhir perkembangan. Pupa berukuran panjang 1,45 - 1,46 mm dan lebar 0,69 - 0,71 mm. Stadium pupa berlangsung selama lima hari (Tabel 2).
Tabel 2 Lama perkembangan parasitoid Trissolcus sp.
Stadia Perkembangan Lama Stadium (hari)
Telur 1
Larva 4
Prapupa 1
Pupa 5
Imago
21
panjang 1,33 ± 0,06 mm dan lebar 0,69 ± 0,03 mm, lebih kecil dibandingkan dengan imago betina yang berukuran panjang 1,51 ± 0,06 mm dan lebar 0,73 ± 0,03 mm (Tabel lampiran 1).
Gambar 9 Imago Trissolcus sp. betina (a) lateral (b) ventral (c) dorsal; jantan (d) lateral (e) ventral (f) dorsal.
Setelah keluar dari telur inang, imago Trissolcus sp. jantan akan berputar-putar di sekitar telur inang yang berisi parasitoid betina yang belum keluar (Gambar Lampiran 2a). Imago jantan tersebut juga akan mengusir imago jantan lain yang mendekati telur inang yang berisi parasitoid betina yang belum keluar. Segera setelah imago betina keluar, imago jantan akan menghampiri dan melakukan kopulasi. Kopulasi berlangsung sangat singkat hanya sekitar 2 – 3 detik (Gambar Lampiran 2b).
[image:32.595.114.505.152.373.2]dibagian dinding samping telur untuk telur yang berada di pinggir kelompok. Trissolcus sp. juga teramati melakukan penandaan telur yang telah diparasit. Tanda dibuat dengan cara menyentuhkan ovipositor pada permukaan telur. Tanda yang dibuat berupa pola yang menyerupai angka delapan. Perilaku ini juga teramati pada spesies Trissolcus yang digunakan Weber et al. (1996) sebagai materi penelitian. Penempelan ovipositor pada telur inang yang terparasit bertujuan untuk menempelkan senyawa berupa feromon penanda inang (host marking pheromone). Senyawa ini disekresikan oleh kelenjar aksesoris yaitu kelenjar Dufour (Rosi et al. 2001).
Potensi Trissolcus sp. Sebagai Parasitoid Telur pada Inang C. javanus
Lama Perkembangan
23
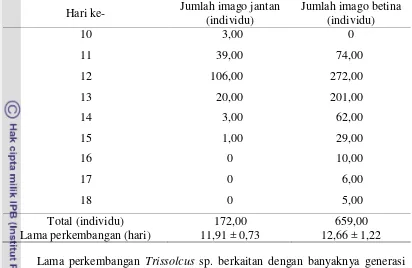
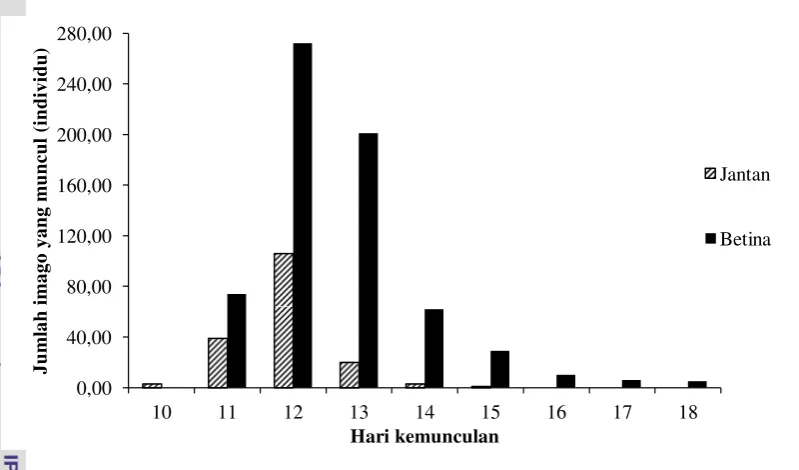
Tabel 3 Jumlah imago Trissolcus sp. jantan dan betina yang muncul Hari ke- Jumlah imago jantan
(individu)
Jumlah imago betina (individu)
10 3,00 0
11 39,00 74,00
12 106,00 272,00
13 20,00 201,00
14 3,00 62,00
15 1,00 29,00
16 0 10,00
17 0 6,00
18 0 5,00
Total (individu) 172,00 659,00
Lama perkembangan (hari) 11,91 ± 0,73 12,66 ± 1,22 Lama perkembangan Trissolcus sp. berkaitan dengan banyaknya generasi yang dapat dihasilkan. Parasitoid yang efektif memiliki lama perkembangan pradewasa yang singkat dan keperidian yang tinggi (Doutt dan DeBach 1973). Semakin singkat waktu perkembangan Trissolcus sp., semakin banyak generasi yang dihasilkan dalam suatu kurun waktu tertentu. Informasi tersebut penting untuk mengetahui perkembangan populasi parasitoid, perbanyakan di laboratorium dan pelepasan di lapangan. Lama perkembangan jantan dan betina juga dapat mempengaruhi keturunan selanjutnya karena perbedaan waktu perkembangan antara jantan dan betina akan membuat peluang terjadinya kopulasi semakin besar. Jantan yang telah muncul terlebih dahulu dapat mengawini betina yang baru muncul. Informasi lama perkembangan parasiotid juga dapat dijadikan dasar dalam penentuan waktu pelepasan di lapangan. Pelepasan parasitoid Trissolcus sp. ke lapang dapat dilakukan pada hari ke-9 setelah telur C. javanus terparasit atau pada saat parasitoid memasuki stadium pupa.
Keberhasilan Hidup
merupakan inang yang sesuai bagi perkembangan Trissolcus sp. Oleh sebab itu, Trissolcus sp. mempunyai potensi yang besar untuk perbanyakan massal maupun pelepasan di lapangan.
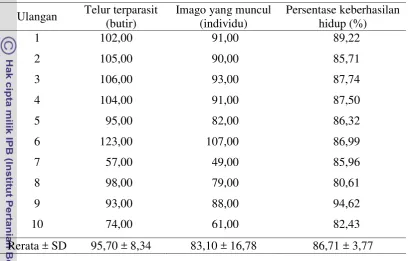
Tabel 4 Keberhasilan hidup imago Trissolcus sp. Ulangan Telur terparasit
(butir)
Imago yang muncul (individu)
Persentase keberhasilan hidup (%)
1 102,00 91,00 89,22
2 105,00 90,00 85,71
3 106,00 93,00 87,74
4 104,00 91,00 87,50
5 95,00 82,00 86,32
6 123,00 107,00 86,99
7 57,00 49,00 85,96
8 98,00 79,00 80,61
9 93,00 88,00 94,62
10 74,00 61,00 82,43
Rerata ± SD 95,70 ± 8,34 83,10 ± 16,78 86,71 ± 3,77 Kematian pradewasa Trissolcus sp. tidak diketahui penyebabnya secara pasti namun berdasarkan pengamatan terlihat bahwa sejumlah imago yang telah berkembang tidak bisa menggigit kulit telur sehingga imago tersebut tidak bisa keluar dari inang. Jenis inang (Kivan dan Kilic 2002) dan suhu (Chabi-Olaye et al. 2001) juga mempengaruhi keberhasilan hidup parasitoid.
Parameter Kehidupan Parasitoid
25
[image:36.595.108.517.147.423.2]suhu rendah (Foerster dan Doetzer 2006) maupun saat kondisi normal (Chabi-Olaye et al. 2001; Iranipour et al. 2010).
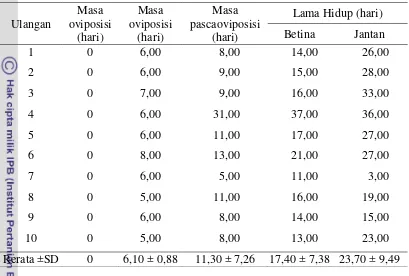
Tabel 5 Parameter kehidupan imago betina Trissolcus sp. Ulangan Masa oviposisi (hari) Masa oviposisi (hari) Masa pascaoviposisi (hari)
Lama Hidup (hari) Betina Jantan
1 0 6,00 8,00 14,00 26,00
2 0 6,00 9,00 15,00 28,00
3 0 7,00 9,00 16,00 33,00
4 0 6,00 31,00 37,00 36,00
5 0 6,00 11,00 17,00 27,00
6 0 8,00 13,00 21,00 27,00
7 0 6,00 5,00 11,00 3,00
8 0 5,00 11,00 16,00 19,00
9 0 6,00 8,00 14,00 15,00
10 0 5,00 8,00 13,00 23,00
Rerata ±SD 0 6,10 ± 0,88 11,30 ± 7,26 17,40 ± 7,38 23,70 ± 9,49 Rendahnya lama hidup betina dibandingkan jantan disebabkan oleh aktifitas oviposisi yang dilakukan oleh betina. Alasan yang dikemukakan oleh Godfray (1994) adalah dibutuhkannya energi dan usaha yang lebih besar dari induk untuk persiapan nutrisi pada jumlah keturunan yang banyak hingga lama hidupnya jadi lebih pendek.
Hasil penelitian menunjukkan bahwa imago betina Trissolcus sp. meletakkan telur pada hari pertama setelah kemunculannya dan rata-rata jumlah telur yang dihasilkan dapat mencapai 35,50 butir. Kondisi ini menguntungkan karena segera setelah parasitoid betina keluar dari telur inang dapat langsung memarasit inangnya, sehingga pengaruh faktor-faktor luar seperti suhu dan kelembaban terhadap potensi parasitisasi dapat diperkecil.
(Hemiptera: Scutelleridae) berkisar antara 4,7 – 22,4 hari tergantung suhu lingkungan dan asal populasi parasitoid.
Masa pascaoviposisi Trissolcus sp. mencapai 11,30 ± 7,26 hari (Tabel 5). Menurut Awan et al. (1990) masa pascaoviposisi T. basalis dapat mencapai 21,1 hari pada inang N. viridula. Hasil penelitian Iranipour et al. (2010) menunjukkan bahwa masa pascaoviposisi T. grandis pada inang E. integriceps dapat mencapai 30 hari. Sedangkan T. semistriatus pada inang E. integriceps hanya 5,7 hari (Kivan dan Kilic 2006). Hasil ini mengindikasikan bahwa Trissolcus sp. merupakan serangga pro-ovigenic. Serangga seperti ini memiliki cadangan telur untuk seumur hidupnya dan dapat meletakkannya pada berbagai inang (Driesche et al. 2008). Informasi siklus hidup parastoid diperlukan untuk mengetahui potensi parasiotoid dalam mengendalikan hama sebagai inangnya. Siklus hidup Trissolcus sp. 11,91 - 12,66 hari lebih pendek dari C. javanus yaitu 60 – 80 hari.
Kemunculan Imago Trissolcus sp.
27
Gambar 10 Kemunculan imago Trissolcus sp.
Nisbah Kelamin
Sebagian besar Hymenoptera parasitoid memilki tipe reproduksi arenotoki atau haplodiploid (Doutt et al. 1989; Driesche 2008). Telur yang tidak dibuahi akan berkembang secara partenogenesis menjadi serangga jantan sedangkan yang dibuahi menjadi serangga betina. Imago betina dan jantan berwarna hitam dan mudah dibedakan, terutama dari ukuran tubuh dan bentuk antenanya. Imago betina lebih besar dibandingkan dengan jantan. Pada antena jantan, ruas flagelomernya membulat sedangkan pada antena betina tidak.
Hasil penelitian pada tabel 6 menunjukkan adanya keragaman dan fluktuasi nisbah kelamin yaitu 1 : 3,83 dimana jumlah imago betina cenderung lebih banyak. Hasil penelitian Awan et al. (1990) menunjukkan nisbah kelamin beragam menurut asal daerah geografis T. basalis pada inang N. viridula berkisar dari 1: 0,94 sampai 1: 1,79. Kivan dan Kilic (2002) menemukan bahwa nisbah kelamin Trissolcus semistriatus bervariasi menurut jenis inang. Doutt (1973) menyatakan bahwa keragaman dan fluktuasi nisbah kelamin adalah karakteristik utama reproduksi halplodiploid. Persentase imago Trissolcus sp. betina yang lebih banyak akan menguntungkan karena imago betina menentukan perkembangan suatu populasi dibandingkan jantan, semakin banyak imago Trissolcus sp. betina akan semakin banyak keturunan yang dihasilkan.
0,00 40,00 80,00 120,00 160,00 200,00 240,00 280,00
10 11 12 13 14 15 16 17 18
Tabel 6 Nisbah kelamin keturunan F1 dari imago betina Trissolcus sp. Ulangan
Jumlah keturunan F1
(individu)
Jantan (individu)
Betina (individu)
Nisbah Kelamin
1 91,00 17,00 74,00 1 : 4,35
2 90,00 11,00 79,00 1 : 7,18
3 93,00 27,00 66,00 1 : 2,44
4 91,00 4,00 87,00 1 : 21,75
5 82,00 8,00 74,00 1 : 9,25
6 107,00 17,00 90,00 1 : 5,29
7 49,00 6,00 43,00 1 : 7,17
8 79,00 15,00 64,00 1 : 4,27
9 88,00 28,00 60,00 1 : 2,14
10 61,00 39,00 22,00 1 : 0,56
Rerata ± SD 83,10 ± 16,78 17,20 ± 11,13 65,90 ± 20,60 1 : 3,83
Kemampuan Reproduksi
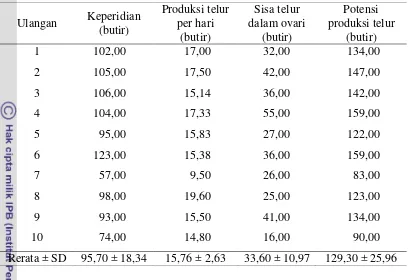
Hasil penelitian pada tabel 7 menunjukkan bahwa parasitoid Trissolcus sp. betina mampu menghasilkan telur sebanyak 95,7 ± 18,34 butir, produksi telur harian sebanyak 15,76 ± 2,63 butir. Hasil pembedahan ovari betina yang telah mati ditemukan 33,60 ± 10,97 butir telur sehingga potensi produksi telur Trissolcus sp. mencapai 129,30 ± 25,96 butir per betina. Kemampuan reproduksi dapat mempengaruhi jumlah keturunan dan menentukan seberapa cepat perkembangan populasi parasitoid tersebut (Doutt 1973).
29
Tabel 7 Kemampuan reproduksi imago betina Trissolcus sp. Ulangan Keperidian
(butir)
Produksi telur per hari
(butir)
Sisa telur dalam ovari
(butir)
Potensi produksi telur
(butir)
1 102,00 17,00 32,00 134,00
2 105,00 17,50 42,00 147,00
3 106,00 15,14 36,00 142,00
4 104,00 17,33 55,00 159,00
5 95,00 15,83 27,00 122,00
6 123,00 15,38 36,00 159,00
7 57,00 9,50 26,00 83,00
8 98,00 19,60 25,00 123,00
9 93,00 15,50 41,00 134,00
10 74,00 14,80 16,00 90,00
Gambar 11 Reproduksi harian imago betina Trissolcus sp. Persentase Parasitisasi
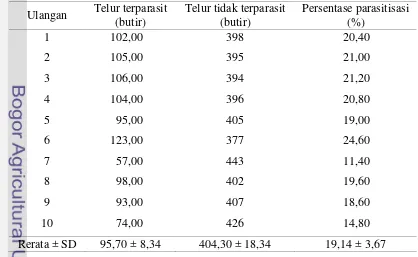
Hasil penelitian pada tabel 8 menunjukkan rata-rata persentase parasitisasi Trissolcus sp. adalah 19,14 ± 3,67 % lebih rendah jika dibandingkan dengan rata-rata tingkat parasitisasi T. basalis inang N. viridula mencapai 41,8 - 72 % (Awan et al. 1990). Hasil penelitian Kivan dan Kilic (2002) menunjukkan persentase parasitisasi Trissolcus semistriatus mencapai 24,0 % pada inang Eurydema ornatum (Hemiptera : Pentatomidae) dan 94,8 % pada inang Graphosoma lineatum (Hemiptera : Pentatomidae).
Tabel 8 Persentase parasitisasi Trissolcus sp. Ulangan Telur terparasit
(butir)
Telur tidak terparasit (butir)
Persentase parasitisasi (%)
1 102,00 398 20,40
2 105,00 395 21,00
3 106,00 394 21,20
4 104,00 396 20,80
5 95,00 405 19,00
6 123,00 377 24,60
7 57,00 443 11,40
8 98,00 402 19,60
9 93,00 407 18,60
10 74,00 426 14,80
Rerata ± SD 95,70 ± 8,34 404,30 ± 18,34 19,14 ± 3,67
0,00 5,00 10,00 15,00 20,00 25,00 30,00 35,00 40,00
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17
P rodu k si te lur har ian (bu tir )
31
[image:42.595.117.505.86.376.2]Persentase parasitisasi harian Trissolcus sp. tertinggi terjadi pada hari pertama kemunculan imago betina yang mencapai 70,60 % (Gambar 12). Hasil ini dapat digunakan untuk menentukan kebutuhan imago betina yang harus dilepaskan di lapang.
Gambar 12 Persentase parasitisasi berdasarkan umur imago Trissolcus sp.
0,00 10,00 20,00 30,00 40,00
50,00
60,00 70,00 80,00 90,00 100,00
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17
P
re
se
n
tas
e
P
ar
as
it
is
as
i
(%
)
KESIMPULAN DAN SARAN
Kesimpulan
Siklus hidup Trissolcus sp. meliputi fase telur (satu hari) , larva (empat hari) yang terdiri tiga instar, prapupa (satu hari), pupa (lima hari) dan dewasa. Lama hidup imago jantan 23,70 ± 9,49 hari dan betina 17,40 ± 7,38 hari. Telur parasitoid Trissolcus sp. bertipe stalked, larva instar pertama bertipe teleaform, dan larva instar ketiga bertipe hymenopteriform.
Parasitoid telur Trissolcus sp. berpotensi dikembangkan sebagai parasitoid telur untuk mengendalikan kepik jarak pagar C. javanus ditinjau dari:
i) Lama perkembangan yang relatif singkat (10 - 12 hari) ii) Nisbah kelamin (1 : 3,83)
iii) Kemampuan reproduksi (95,7 butir/betina selama masa hidupnya) iv) Masa praoviposisi yang singkat (<1 hari)
Saran
DAFTAR PUSTAKA
Arakawa R, Miura M, Fujita M. 2004. Effects of host species on the body size, fecundity, and longevity of Trissolcus mitsukurii (Hymenoptera: Scelionidae), a solitary egg parasitoid of stink bugs. Appl. Entomol. Zool. 39 (1): 177–181.
Austin AD, Johnson NF, Dowton M. 2005. Systematics, evolution, and biology of scelionid and platygastrid wasps. Annu. Rev. Entomol 50: 553–582. Awan MS, Wilson LT, Hoomann MP. 1990. Comparative biology of three
geographic populations of Trissolcus basalis (Hymenoptera: Scelionidae). Environ. Entomol. 19(2): 387-392.
Bebawi FF, Lockett CJ, Davis KM, Lukitsch BV. 2007. Damage potential of an introduced biological control agent Agonosoma trilineatum (F.) on bellyache bush (Jatropha gossypiifolia L.). Biological Control 41: 415–422. Borror DJ, Triplehorn CA, Johnson NF. 1996. Pengenalan Pelajaran Serangga
Edisi ke-6. Partosoedjono S, penerjemah. Brotowidjoyo M D, penyunting. Yogyakarta: UGM-Press. Terjemahan dari: An Introduction to the Study of Insects, Sixth edition.
Chabi-Olaye A, Schulthess F, Poehling HM, Borgemeister C. 2001. Factors affecting the biology of Telenomus isis (Polaszek) (Hymenoptera: Scelionidae), an egg Parasitoid of cereal stem borers in West Africa. Biological Control 21: 44–54.
Chandra D. 2008. Inventarisasi hama dan penyakit pada pertanaman jarak pagar (Jatropha curcas Linn.) di Lampung dan Jawa Barat. [skripsi]. Bogor: Fakultas Pertanian, Institut Pertanian Bogor.
Clausen CP. 1940. Entomophagus Insect. New York: McGraw Hill Book Company Inc.
Dadang. 2005. Pengendalian hama dan penyakit tanaman jarak pagar (Jatropha curcas Linn.) Dalam : Seminar Nasional Pengembangan Jarak Pagar (Jatropha curcas Linn.) untuk Biodiesel dan Minyak Bakar. Prosiding. Bogor, 22 Desember 2006. Bogor: Pusat Penelitian Surfaktan dan Bioenergi, hlm 90-103.
Dadang. 2006. Pengendalian terpadu hamatanaman jarak pagar (Jatropha curcas Linn.). Dalam : Workshop Hama dan Penyakit Tanaman Jarak (Jatropha curcas Linn.) : Potensi Kerusakan dan Teknik Pengendaliannya. Bogor,5-6 Desember 2006. Bogor: Pusat Penelitian Surfaktan dan Bioenergi, hlm 17-22.
Doutt RL, DeBach P. 1973. Some biological control concepts and question. Di dalam: DeBach P,editor. Biological Control of Insect Pest and Weeds. London: Chapman and Hall Ltd. hlm 118-142.
Doutt RL, Annecke DP, Tremblay E. 1989. Biologi dan hubungan hospes parasitoida. Di dalam: Huffaker CB, Messenger PS,editor. Teori dan Praktek Pengendalian Biologis. Mangoendiharjo S,penerjemah. Untung K,pendamping. Jakarta: UI-press. Terjemahan dari: Theory and Practice of Biological Control.
Driesche RV, Hoddle M, Center T. 2008. Control of Pests and Weeds by Natural Enemies: an Introduction to Biological Control. Singapore: Blackwell Publishing.
Foerster LA, Doetzer AK. 2006. Cold storage of the egg parasitoids Trissolcus basalis (Wollaston) and Telenomus podisi Ashmead (Hymenoptera: Scelionidae). Biological Control 36: 232–237.
Godfray HCJ. 1994. Parasitoids, Behavioral and Evolutionary Ecology. New Jersey: Princeton University Press.
Gordh G, Headrick D. 2001. A Dictionary of Entomology. UK: CABI Publishing. Goulet H, Huber JT,editor. 1993. Hymenoptera of the World: An Identification
Guide to Families. Kanada: Agriculture Canada Publication.
Hagen KS. 1973. Developmental stages of parasites. Di dalam: DeBach P,editor. Biological Control of Insect Pest and Weeds. London: Chapman and Hall Ltd. hlm 168-246.
Hambali E. 2005. Kontribusi perguruan tinggi dan lembaga litbang untuk pengembangan jarak pagar (Jatropha curcas Linn.) menjadi biodiesel dan minyak bakar). Dalam : Seminar Nasional Pengembangan Jarak Pagar (Jatropha curcas Linn.) untuk Biodiesel dan Minyak Bakar. Prosiding. Bogor, 22 Desember 2006. Bogor: Pusat Penelitian Surfaktan dan Bioenergi, hlm 6-24.
Hambali E, Suryani A, Dadang, Hariyadi, Hanafie H, Reksowardojo IK, Rivai M, Ihasanur M, Suryadarma P, Tjitrosemito S, Soerawidjaja TH, Prawitasari T, Prakoso T, Purnama W. 2007. Jarak Pagar Tanaman Penghasil Biodiesel Cet ke-4. Jakarta: Penebar Swadaya.
Heard TA, Chan RR, Senaratne KADW, Palmer WA, Lockett C, et al. 2009. Agonosoma trilineatum (Heteroptera: Scutelleridae) a biological control agent of the weed bellyache bush, Jatropha gossypiifolia (Euphorbiaceae). Biological Control 48: 196–203.
Iranipour S, Nozadbonab Z, Michaud JP. 2010. Thermal requirements of Trissolcus grandis (Hymenoptera: Scelionidae), an egg parasitoid of sunn pest. Eur. J. Entomol. 107: 47–53.
35
Kalshoven LGE. 1981. Pests of Crops in Indonesia. Van Der Laan PA, penerjemah. Jakarta: PT Ichtiar Baru-Van Hoeve. Terjemahan dari: De Plagen van de Cultuurgewassen in Indonesie.
Karmawati E, Rumini W. 2009. Dinamika populasi dan pengendalian hama utama jarak pagar. Warta Penelitian dan pengembangan Pertanian 31(5): 12-14.
Kementerian Perhubungan. 2009. Perhubungan Darat dalam Angka 2009. Jakarta: Direktorat Jenderal Perhubungan Darat.
Kivan M, Kilic N. 2002. Host preference: parasitism, emergence and development of Trissolcus semistriatus (Hymenoptera: Scelonidae) in various host eggs. J. Appl. Ent 126: 395-399.
Kivan M, Kilic N. 2006. Age-specific fecundity and life table of Trissolcus semistriatus, an egg parasitoid of the sunn pest Eurygaster integriceps. Entomological Science 9: 39-46.
Koçak E, Kilinçer N. 2003. Taxonomic studies on Trissolcus sp. (Hymenoptera: Scelionidae), egg parasitoids of the sunn pest (Hemiptera: Scutelleridae: Eurygaster sp.), in Turkey. Turk J Zool 27: 301-317.
Krisnamurthi B. 2005. Pengembangan biofuel berbahan baku “jarak pagar” sebagai bagian dari kebijakan diversifikasi energi nasional. Dalam : Seminar Nasional Pengembangan Jarak Pagar (Jatropha curcas Linn.) untuk Biodiesel dan Minyak Bakar. Prosiding. Bogor, 22 Desember 2006. Bogor: Pusat Penelitian Surfaktan dan Bioenergi, hlm 1-5.
Krombein KV, Hurd,Jr. PD, Smith DR, Burks BD. 1979. Catalog of Hymenoptera in America North of Mexico. Washington DC: Smithsonian Institution Press. Laumann RA, Moraes MCB, Pareja M, Alarcão GC, Botelho AC, Maia AHN, Leonardecz E, Borges M. 2008. Comparative biology and functional response of Trissolcus spp. (Hymenoptera: Scelionidae) and implications for stink bugs (Hemiptera: Pentatomidae) biological control. Biological Control 44: 32–41.
Mahmud Z. 2006. Budi daya jarak pagar untuk sumber energi masa depan. Dalam: Warta Penelitian dan pengembangan Pertanian 28(4): 1-3.
Masner L. 1993. Superfamily Platygastroidea. Di dalam: Goulet H, Huber JT, editor. Hymenoptera of The World: an Identification Guide to Families. Ottawa: Canada Communication Group. hlm 558 – 620.
Nurcholis M, Sumarsih S. 2007. Jarak Pagar dan Pembuatan Biodisel, Seri Budi Daya. Yogyakarta: Kanisius.
Prihandana R, Hendroko R. 2006. Petunjuk Budi Daya Jarak Pagar. Tangerang: Agromedia Pustaka.
Priyanto U. 2007. Menghasilkan Biodisel Jarak Pagar Berkualitas. Jakarta: Agromedia.
(Jatropha curcas Linn.) di Kabupaten Bogor. [skripsi]. Bogor: Fakultas Pertanian, Institut Pertanian Bogor.
Rahat S, Gurr GM, Wratten SD, Mo J, Neeson R. 2005. Effect of plant nectars on adult longevity of the stinkbug parasitoid, Trissolcus basalis. International Journal of Pest Management 51(4): 321-324.
Rajmohana K. 2006. A checklist of the Scelionidae (Hymenoptera: Platygastroidea) of India. Zoos Print Journal 21(12): 2506-2613.
[RI] Presiden Republik Indonesia. 2006. Peraturan Presiden Republik Indonesia Nomor 5 Tahun 2006 Tentang Kebijakan Energi Nasional. Jakarta: RI. [RI] Presiden Republik Indonesia. 2006. Instruksi Presiden Republik Indonesia
Nomor 1 Tahun 2006 Tentang Penyediaan Dan Pemanfaatan Bahan Bakar Nabati (Biofuel) Sebagai Bahan Bakar Lain. Jakarta: RI.
Rider DA. 2009. Hymenoptera parasitoid records list by Pentatomoid species. Department of Entomology. North Dakota State University. http://www.ndsu.nodak.edu [21 Mei 2011].
Rosi MC, Isidoro N, Colazza S, Bin F. 2001. Source of the host marking pheromone in the egg parasitoid Trissolcus basalis (Hymenoptera: Scelionidae). Journal of Insect Physiology 47: 989–995.
Rumini W, Karmawati E. 2007. Hama pada tanaman jarak pagar (Jatropha curcas). Dalam: Status Teknologi Tanaman Jarak Pagar (Jatropha curcas L.). Prosiding. Lokakarya II Jarak Pagar. Bogor, 29 Nopember 2006. Bogor: Pusat Penelitian dan Pengembangan Perkebunan, hlm 302 – 306.
Shanker C, Dhyani SK. 2006. Insect pests of Jatropha curcas L. and the potential for their management. Current Science 91(2): 162-163.
Sodiq A. 2006. Inventarisasi hama dan penyakit tanaman jarak pagar (Jatropha curcas Linn.) di Bogor. [skripsi]. Bogor: Fakultas Pertanian, Institut Pertanian Bogor.
Sosromarsono S, Wardojo S, Adisoemarto S, Suhardjono YR. 2007. Nama Umum Serangga. Ed ke-2. Bogor: Perhimpunan Entomologi Indonesia.
Tohir AM. 2004. Teknik pembiakan massal parasitoid telur pengisap polong kedelai Trissolcus basalis Wollaston (Hymenoptera: Scelionidae). Buletin Teknik Pertanian 9(2): 66-69.
Torres JB, Musolin DL, Zanuncio JC. 2002. Thermal requirements and parasitism capacity of Trissolcus brochymenae (Ashmead) (Hymenoptera: Scelionidae) under constant and fluctuating temperatures, and assessment of development in field conditions. Biocontrol Science and Technology 12: 583-593.
Tabel Lampiran 1 Ukuran pradewasa dan imago Trissolcus sp.
Stadia Perkembangan Rerata ± SD
Panjang (mm) Lebar (mm)
Telur 0,38 ± 0,03 0,17 ± 0,02
Larva instar pertama 0,66 ± 0,19 1,03 ± 0,18 1,34 ± 0,07
0,35 ± 0,17 0,82 ± 0,24 1,13 ± 0,10
Larva instar kedua Larva instar ketiga
Prapupa 1,32 ± 0,09 0,97 ± 0,10
Pupa 1,38 ± 0,05 0,84 ± 0,08
Imago jantan 1,33 ± 0,06 0,69 ± 0,03
Imago betina 1,51 ± 0,06 0,73 ± 0,03
39
Gambar Lampiran 2 Imago parasitoid Trissolcus sp.; (a) jantan menunggu kemunculan betina, (b) kopulasi sesaat betina muncul dari inang, (c) dan (d) betina melakukan oviposisi.
41
ABSTRAK
YULIUS DIKA CIPTADI. Biologi, Siklus Hidup, dan Potensi Parasitoid Telur Trissolcus sp. Pada Chrysocoris javanus Westw., Hama Tanaman Jarak Pagar (Jatropha curcas L.). Dibimbing oleh DHAMAYANTI ADIDHARMA.
Beberapa tahun terakhir, jarak pagar (Jatropha curcas L.) mulai diusahakan dalam skala luas dan ditanam secara monokultur karena potensinya sebagai sumber bahan bakar nabati. Cara budi daya seperti ini dapat menyebabkan perubahan keadaan agroekosistem yaitu pengurangan keanekaragaman hayati sehingga menyediakan lingkungan yang sesuai untuk perkembangan hama. Salah satunya adalah hama Chrysocoris javanus. Hama ini menyerang buah sehingga menyebabkan buah rusak dan tidak dapat dipanen. Musuh alami seperti parasitoid telah ditemukan menyerang telur C. javanus. Tujuan dari penelitian ini adalah untuk mengetahui biologi, siklus hidup dan potensi Trissolcus sp. sebagai parasitoid telur pada inang C. javanus. Telur C. javanus direkatkan pada pias kertas dengan menggunakan gom arab kemudian dimasukkan ke dalam tabung gelas yang telah berisi parasitoid betina. Penelitian tentang bentuk, ukuran, warna, dan ciri-ciri lain setiap stadium pradewasa dilakukan dengan membedah telur setiap 24 jam kecuali stadia larva setiap 12 jam dan diamati menggunakan mikroskop stereo. Penelitian tentang biologi, siklus hidup dan potensi Trissolcus sp. sebagai parasitoid telur pada inang C. javanus dilakukan tanpa pembedahan telur dan pengamatan dilakukan terhadap keperidian, produksi telur harian, potensi produksi telur, lama hidup parasitoid, lama perkembangan, kemunculan imago, nisbah kelamin dan lama masa reproduksi. Hasil penelitian menunjukkan bahwa telur parasitoid Trissolcus sp. bertipe stalked, larva instar pertama bertipe teleaform, larva instar ketiga bertipe hymenopteriform. Lama stadium telur Trissolcus sp. satu hari, larva empat hari, prapupa satu hari, pupa lima hari, lama hidup imago jantan 23,70 ± 9,49 hari dan betina 17,40 ± 7,38 hari. Keperidian 95,70 ± 18,34 butir, produksi telur harian 15,76 ± 2,63 butir, potensi produksi telur 129,30 ± 25,96 butir, lama masa reproduksi 6,10 ± 0,88 hari, nisbah kelamin 1 : 3,83. Tingkat parasitisasi 19,14 ± 3,67 % dan keberhasilan hidup mencapai 86,71 ± 3,77 %. Parasitoid telur Trissolcus sp. berpotensi dikembangkan sebagai parasitoid telur untuk mengendalikan kepik jarak pagar C. javanus.
PENDAHULUAN
Latar Belakang
Kebutuhan bahan bakar minyak Indonesia saat ini cukup tinggi. Konsumsi energi terbesar adalah minyak bumi yang mencapai 54,4 % (Krisnamurthi 2005). Menurut data Direktorat Lalu Lintas Angkutan Jalan – Ditjen Perhubungan Darat tahun 2009, kebutuhan solar mencapai 12.382.242 liter per hari dan jumlah kendaraan akan mencapai 110 juta kendaraan pada tahun 2020. Sedangkan cadangan energi fosil hanya akan bertahan kurang lebih 18 tahun (Krisnamurthi 2005).
Melihat kenyataan tersebut, perlu adanya tindakan untuk mengurangi ketergantungan terhadap bahan bakar minyak dan menanggulangi masalah harga minyak yang makin meningkat dan cadangan yang makin menipis. Pemerintah telah menerbitkan Peraturan Presiden Republik Indonesia Nomor 5 Tahun 2006 tentang Kebijakan Energi Nasional untuk mengembangkan sumber energi alternatif sebagai pengganti bahan bakar minyak. Presiden Indonesia juga menerbitkan Instruksi Presiden Nomor 1 Tahun 2006 tentang penyediaan dan pemanfaatan bahan bakar nabati (biofuel) sebagai energi alternatif.
Hambali (2005) menyatakan salah satu sumber minyak nabati yang sangat prospektif sebagai bahan baku biodiesel adalah biji jarak pagar yang bernama latin Jatropha curcas Linn. Program pemerintah untuk mensubtitusi 5% minyak bumi sampai tahun 2025 dapat memacu penanaman jarak pagar dalam skala luas, monokultur dan intensif. Cara budi daya seperti ini dapat menyebabkan perubahan keadaan ekosistem yaitu berkurangnya keanekaragaman hayati. Kondisi ini akan menyediakan lingkungan yang sesuai untuk perkembangan hama (Dadang 2005).
2
Kepik C. javanus menjadi salah satu hama penting pada pertanaman jarak pagar karena menyerang tanaman jarak fase generatif yaitu pada saat pembungaan, menjelang pembentukan buah dan saat pembentukan buah. C. javanus menyerang tanaman dengan cara menghisap buah jarak sehingga menimbulkan kerusakan pada kapsul buah yang sedang berkembang. Gejala yang ditimbulkan pada buah jarak pagar yaitu adanya bekas tusukan pada buah dan buah berwarna coklat kehitaman kemudian membusuk dan mengering (Rumini dan Karmawati 2007; Qodir 2010). Berbeda dengan di Indonesia, serangga anggota famili Scutelleridae yang menyerang jarak pagar di India adalah Scutellera nobilis Fabr. Sedangkan Pachycoris klugii Burmeister dan Agonosoma trilineatum Fabr. menyerang jarak pagar di Nicaragua. Serangga ini dilaporkan dapat menyebabkan gugurnya bunga, aborsi buah dan kerusakan benih. (Shanker dan Dhyani 2006). Bahkan A. trilineatum menjadi agens hayati untuk mengendalikan gulma famili Euphorbiaceae, Jatropha gossypiifolia (Bebawi et al. 2007; Heard et al. 2009).
Tujuan Penelitian
Penelitian ini bertujuan mengetahui biologi, siklus hidup, dan potensi parasitoid telur Trissolcus sp. pada inang C. javanus.
Manfaat Penelitian
TINJAUAN PUSTAKA
Tanaman Jarak Pagar (Jatropha curcas Linn.)
Tanaman jarak pagar (Jatropha curcas Linn.) termasuk dalam famili Euphorbiaceae merupakan salah satu tanaman yang baik sebagai sumber bahan bakar nabati (Prihandana dan Hendroko 2006; Hambali et al. 2007). Jarak pagar berasal dari Amerika Tengah dan didistribusikan oleh pelaut Portugis melalui pulau Cape Verde ke berbagai negara di Afrika dan Asia (Hambali 2005). Jarak pagar mempunyai 4 varietas, yaitu varietas Cape Verde, Nicaragua, Ife-Nigeria, dan varietas tidak beracun Mexico. Varietas Cape Verde merupakan varietas yang umum terdapat di seluruh dunia dan bersifat toksik karena mengandung senyawa lektine dan ester forbol (Henning 2005 dalam Nurcholis dan Sumarsih 2007).
C. javanus memiliki nama umum kepik buah jarak (Sosromarsono et al. 2007). Tubuhnya berbentuk perisai yang khas dengan skutelum yang berkembang dengan baik menutupi abdomen, warna tubuh imago cerah dan mencolok dengan elitra berwarna merah dan corak hitam. Serangga ini memiliki antena tiga ruas, lebih panjang dari kepala (Dadang 2006; Kalshoven 1981). Telur berbentuk silinder seperti drum, bagian bawah datar sedangkan bagian atas cembung. Telur berdiameter 1,24 mm dan tinggi 1,34 mm. Telur yang baru diletakkan berwarna krem terang dan ada yang berwarna agak kehijauan. Telur diletakkan secara berkelompok di bawah permukaan daun, pada batang atau ranting dan pada permukaan buah jarak pagar saat pagi hingga menjelang siang. Nimfa tubuhnya berwarna hitam dengan bintik merah, kuning, dan hijau mengkilat, sementara bagian dorsal toraks berwarna hijau metalik. Ukuran tubuh imago betina relatif lebih besar dari pada imago jantan, yaitu panjang 17,65 mm dan lebar 9,55 mm sedangkan ukuran tubuh jantan panjang 15,95 mm dan lebar 8,1 mm. Kemampuan reproduksi cukup tinggi, satu kelompok telur terdiri dari 28 sampai 126 butir dengan rerata 84,80 butir (Qodir 2010). Siklus hidupnya berkisar 60-80 hari (Rumini dan Karmawati 2007).
6 insektisida berbahan aktif imidaklorpid dan karbamat (Dadang 2006; Mahmud 2006; Rumini dan Karmawati 2007).
Parasitoid Trissolcus sp.
Trissolcus sp. merupakan parasitoid yang termasuk ke dalam Superfamili Platygastroidea Famili Scelionidae Subfamili Telenominae (Rajmohana 2006). Semua spesies yang telah diketahui dalam famili ini merupakan parasitoid telur dan hidup pada berbagai habitat (Hagen 1973; Austin et al. 2005; Driesche et al.. 2008). Trissolcus sp. telah menjadi spesies yang penting dalam penelitian maupun praktek pengendalian hayati serangga hama khususnya serangga ordo Hemiptera (Awan et al. 1990; Weber et al. 1996; Justo et al. 1997; Koçak dan Kilinçer 2003; Arakawa et al. 2004; Tohir 2004; Laumann et al. 2008)
Tahap Perkembangan Parasitoid Famili Scelionidae
Parasitoid famili Scelionidae terbagi pada tiga kelompok subfamili yaitu Scelioninae, Telesinae, dan Telenominae (Krombein 1979; Masner 1993). Parasitoid ini dapat menyerang telur serangga ordo Lepidoptera, Hemiptera, Orthoptera, Diptera (Tabanidae) dan Arachnida. Adakalanya beberapa spesies menyerang telur serangga ordo Coleoptera dan Neuroptera (Clausen 1940).
Imago parasitoid famili Scelionidae hidup secara soliter (Clausen 1940), umumnya berada pada lingkungan terbuka dan terpapar sinar matahari seperti padang rumput, gurun pasir, hutan, tanah, dan air (Masner 1993).
Telur
Hagen (1973) menyatakan hampir semua parasitoid yang meletakkan telur dan berkembang di dalam inang mengalami perkembangan hipermetamorfosis. Artinya, larva instar awal memiliki perbedaan bentuk dengan larva instar akhir. Jumlah instar larva sangat beragam antar genus dan spesies. Pada larva ektoparasit terdapat lima instar dan pada larva endoparasit kurang dari lima instar.
Clausen (1940) menyebutkan ada 14 tipe larva instar awal pada parasitoid dari ordo Hymenoptera. Larva instar awal memiliki variasi bentuk yang terbesar dalam perkembangan pradewasa parasitoid. Tipe larva instar awal tersebut adalah sacciform, hymenopteriform, caudate, vesiculate, encyrtiform, mandibulate, teleaform, microtype, mymariform, planidium, agriotypiform, polypodeiform, eucoiliform dan cyclopiform.
Larva instar pertengahan tidak memilki perbedaan karakteristik yang mendasar dari bentuk larva instar awalnya (Hagen 1973). Bentuk sebenarnya dari larva instar kedua serangga famili Scelionidae masih dalam perdebatan. Pada beberapa spesies hanya ditemukan dua instar larva, ini ditentukan berdasarkan persamaan bentuk antara larva instar kedua dan ketiga (Clausen 1940). Larva instar akhir pada parasitoid dari ordo Hymenoptera adalah hymenopteriform (Hagen 1973).
Prapupa dan Pupa
8 Pupa serangga ordo Hymenoptera bertipe eksarata dengan embelan bebas (Borror et al. 1996). Serangga ordo Hymenoptera khususnya parasitoid berpupa di dalam inang. Seringkali larva parasitoid berpupa di dalam kokon atau puparium inang atau berpupa di dalam terowongan yang dibuat inang dimana inang tersebut terlindungi. Parasitoid yang hidup di pada inang yang tersembunyi biasanya tidak berkokon (Hagen 1973).
Imago
BAHAN DAN METODE
Tempat dan Waktu Penelitian
Penelitian dilaksanakan di lapang dan di Laboratorium Bioekologi Parasitoid dan Predator Departemen Proteksi Tanaman Institut Pertanian Bogor, pada bulan Mei sampai Agustus 2011.
Metode Penelitian
Survei Awal Bioekologi C. javanus dan Trissolcus sp.
[image:63.595.99.493.114.783.2]Survei awal tentang bioekologi serangga uji parasitoid Trissolcus sp dan C. javanus dilakukan di kebun jarak pagar di Kebun Percobaan IPB desa Leuwikopo, kecamatan Darmaga, kabupaten Bogor. Informasi tentang cara hidup, perilaku dan tempat peletakkan telur serangga uji ini diperlukan sebagai dasar dalam teknik perbanyakannya.
Gambar 1 Lahan jarak pagar di Leuwikopo Perbanyakan C. javanus
10
tanaman jarak pagar dalam polibag berukuran 18 cm x 25 cm dengan media tanah dan selanjutnya dimasukkan ke dalam sangkar berkerangka kayu berdinding kain kasa dengan ukuran 45 cm x 45 cm x 60 cm. Setiap hari tanaman disiram untuk menjaga kesegaran tanaman. Sebagai pakan C. javanus, buah jarak pagar segar digantungkan pada tanaman jarak. Pada setiap kaki-kaki sangkar diberi alas wadah plastik yang diisi oli untuk mencegah semut dan serangga lain masuk kedalam sangkar. Telur-telur C. javanus yang dihasilkan diambil setiap hari sebagai bahan penelitian.
Gambar 2 Perbanyakan C. javanus, (a) sangkar pemeliharaan nimfa dan imago, (b) kelompok telur C. javanus yang diletakkan di bawah permukaan daun tanaman jarak pagar dalam sangkar.
Perbanyakan Parasitoid Telur Trissolcus sp.
Parasitoid Trissolcus sp. diperoleh dengan cara mengumpulkan kelompok telur kepik C. javanus terparasit dari pertanaman jarak pagar di desa Leuwikopo, Darmaga dan desa Lulut, Citeureup. Telur-telur C. javanus diambil beserta tempat melekatnya telur-telur tersebut (daun, ranting atau buah), dimasukkan ke dalam kantung plastik dan diberi label keterangan asal telur dan waktu pengambilan. Di laboratorium, telur tersebut dimasukkan ke dalam tabung reaksi berukuran 0,5 cm x 10 cm yang ditutup kapas. Imago parasitoid yang keluar dipelihara dengan diberi pakan larutan madu 10% yang dioleskan pada permukaan dinding dalam tabung. Imago-imago yang baru muncul tersebut akan digunakan untuk perbanyakan.
parasitoid Trissolcus sp. berumur satu hari dan telah mengalami kopulasi. Telur inang yang telah terparasit dipindahkan ke dalam tabung reaksi lainnya. Imago parasitoid yang keluar dipelihara dengan diberi pakan larutan madu 10% yang dioleskan pada permukaan dinding dalam tabung. Tabung-tabung tersebut disusun rapi diatas papan kayu yang ditopang wadah plastik yang diisi oli untuk mencegah semut dan serangga lain mencapai tabung. Seluruh tabung disimpan dalam ruangan dengan suhu 28,18 ± 0,59 0C dan kelembaban relatif 56,33 ± 4,27 %. Imago-imago yang baru muncul tersebut akan digunakan untuk penelitian.
Penelitian Biologi dan Siklus Hidup Parasitoid Trissolcus sp. pada Inang C. javanus
Telur C. javanus umur satu hari sebanyak 5 butir yang diambil dari