TANGGAP KARAKTER AGRONOMI DAN HASIL
KEDELAI BERUMUR DALAM YANG DIPUPUK
:
PADA SISTEM BUDIDAYA JENUH AIF
OLEH :
MOCHTAR
PROGRAM PASCASARJANA
INSTITUT PERTANIAN BOGOR
ABSTRAK
M 0 C H T A R. Tanggap Karakter Agronomi dan Hasil Genotipe Kedelai Beru~nur Dalam yang Dipupuk Dolomit pada Sistem Budidaya Jenuh Air. Di bimbing oleh
FRED RUMAWAS, MUNIF GHULAMAHDI, dan WAHJU QA;YIARA
MUGNISJAH.
Percobaan ini bertujuan mempelajari (1) tanggap genotipe kedelai berumur dalam terhadap Ca dan Mg yang berasal dari pupuk dolomit pada sistem budidaya jenuh air (BJA) dan (2) tanggap hasil dan karakter agronomi kedelai beru~nur dalam terhadap pemupukan dolomit pada sistem budidaya jenuh air.
Percobaan dilaksanakan di Kebun Percobaan Cikarawang Darmaga, IPB Bogor, dari bulan Mei sampai Agustus 2000. Rancangan disusun dalam bentuk petak terpisah dengan tiga ulangan. Dosis dolomit (0, 2, 4, dan 6 ton ha-') sebagai petak utama dan genotipe (PTR6, PTR178, PTR190, PTR201, PTR288, dan PTR293) sebagai anak petak
SURAT PERNYATAAN
Dengan ini saya menyatakan bahwa tesis yang berjudul
TANGGAP KARAKTER AGRONOMI DAN HASIL GE KEDELAI BERUMUR DALAM YANG DIPUPUK DOLl SISTEM BUDIDAYA JENUH AIR.
Adalah benar merupakan hasii karya saya sendiri dan belum pernah Semua sumber data dan informasi yang digunakan telah dinyatakan dapat diperiksa kebenarannya.
Nrp. 98049 /Agr.
lOTIPE IMIT PADA
TANGGAP ICARAKTER AGRONOMI DAN HASlLGEN ICEDELAI BERUMUR DALAM YANG DIPUPUIC DOL
PADA SISTEM BUDIDAYA JENUH AIR
MOCHTAR
Tesis
sebagai salah sato syarat untuk memperoleh gelar Magister Sains pada
Program Studi Agronomi
ITIPE MIT
PROGRAM PASCASARJANA INSTITUT PERTANIAN BOGOR
Judul Tesis : Tanggap Karakter Agronomi dan Hasil Genotip Berumur Dalam yang Dipupuk Dolomit pada Si Budidaya Jenuh Air.
Nama Mahasiswa : Mochtar
Nomor Pokok : 98049
Program Studi : Agronomi
Menyetujui,
Dr. Ir. Fred Rumawas. MSc. Ketua
Anggota
Mengetahui,
2. Ketua Program Studi Agronomi
C _ _ _
Dr. Ir. Hairial Aswidinnoor. MSc.
: ICedelai
tem
-
-
. . ih. M.Agr. icasarjana@
~woto, MSc.
RIWAYAT HIDUP
Penulis dilahirkan di Gowa, Sulawesi Selatan, pada tanggal
sebagai anak keempat dari Bapak Anwar dan Ibu Halipah. Penulis r
Iis Nurliani pada tanggal 17 Juli 1992 dan dikaruniai tiga orang putri.
Tahun 1980 penulis lulus Sekolah Dasar Negeri Panaikang
1983 lulus Sekolah Menengah Pertama (SMP) Negeri Bontomanai, GI
lulus SPP-SPMA Negeri Gowa. Pada tahun 1986 penulis melanjutka
Fakultas Pertanian Universitas "45" Makassar dan lulus tahun 1991.
Semenjak tahun 1991 sampai dengan sekarang penulis bekerja
pada Fakultas Pertanian Universitas "45" Makassar. Pada tahun
mendapat kesempatan melanjutkan pendidikan S2 pada Program Pascc
Pertanian Bogor dengan program studi Agronomi yang dibiayai
Program Pascasarjana (BPPS).
0 April 1966
:nikah dengall
Gowa; tahun
va; tahu 1986
pendidikan di
;ebagai dosen
1998 penulis
lrjana Institut
PRAKATA
Puji dan syukur penulis panjatkan ke hadirat Allah SWT, atas berkat dan rahmat-
Nya penulis telah dapat melaksanakan penelitian hingga penyusunan
salah satu syarat untuk memperoleh gelar Magister Sains pada Prog
Institut Pertanian Bogor.
Dalam kesempatan ini penulis mengucapkan terima kas
besarnya kepada
1 Bapak Dr. Ir. Fred Rumawas, MSc sebagai Ketua Komi
Bapak Dr. Ir. Munif Ghulamahdi, MS., dan Bapak Dr. Ir
Mugnisjah, M. Agr., masing-masing sebagai anggota.
2 Bapak Pimpinan Beasaswa Program Pascasarjana (BPP
menyediakan beasiswa selama tugas belajar.
3 Bapak Kepala dan Staf Kebun Percobaan Cikarawang Darmaga,
Bogor yang telah membantu penulis selama penelitian
4 Isteri yang tercinta, Iis Nurliana, dan anak-anak tersayang.
Mochtar, Annisa Nuramaliah Mochtar, dan Alifah Nurhadizah Mochtar, serta
seluruh keluarga yang telah memberi dorongan dan semangat sela
pendidikan baik dalam suka maupun duka.
5 Rekan-rekan mahasiswa Program Studi Agronomi dan semua
ikut membantu penulis mulai dari pelaksanaan penelitian sampai selesainya
Penulis berharap semoga sesala kebaikan diberi imbalan yang
Allah SWT dan tesis ini dapat bermanfaat bagi pihak-pihak yar
DAPTAR IS1
DAFTAR TABEL
...
.
.
.
...DAFTAR GAMBAR
...
...DAFTAR LAMPIRAN ...
PEWAHULUAN
.
...
...
Latar Belakang
...
Tujuan Percobaan
...
.... ...Hipotesis Percobaan
...
.
.
.
...TlNJAIJAN PUSTAKA
...
Budidaya Jenuh Air dan Tanggap Tanaman Kedelai ...
...
Keadaan Tergenang
... ...
Pengapuran
.
.
.
.
.
...
Serapan Hara Tanaman
BAHAN DAN METODE ...
Tempat dan Waktu Percobaan ...
.
.
...Metode Percobaan
...
....
......
...Pelaksanaan Percobaan ...
Pengamatan dan Analisis
H A S I L ...
Serapan dan Kadar Ca Mg Tajuk ...
.
.
.
...Pertumbuhan dan Umur Tanaman ... ...
I<omponen Hasil
...
...
Hasil BIJI
PEMBAHASAN ...
... ...
KESIMPULAN DAN SARAN
.
.
.
.
Kesimpulan ...
.
.
...S a r a n
...
.
.
.
... .
.
...DAFTAR TABEL
1 Genotipe kedelai berumur dalam dan asal persilangannya ... 13
2 Pengaruh dolomit dan genotipe terhadap kadar Mg tajuk ... 2 1
3 Pengaruh dolomit dan genotipe terhadap bobot kering bintil akar,
bobot kering akar, dan bobot kering tajuk ... 22
4 Pengaruh dolomit dan genotipe terhadap tinggi tanaman ... 23
5 Pengaruh dolomit dan genotipe terhadap bobot
...
100 biji dan -1bobot ~ I J I tanaman
...
286 Pengaruh dolomit dan genotipe terhadap bobot biji petak-l dan
. . . .
DAFTAR GAMBAR
1 Hubungan dolomit dengan kadar Ca tajuk beberapa genotipe kedelai berumur dalam ...
2 Serapan Ca tajuk genotipe kedelai berumur dalam pada beberapa dosis dolomit
...
.
.
...
3 Serapan M g tajuk genotipe kedelai berumur dalam pada beberapa dosis dolomit
...
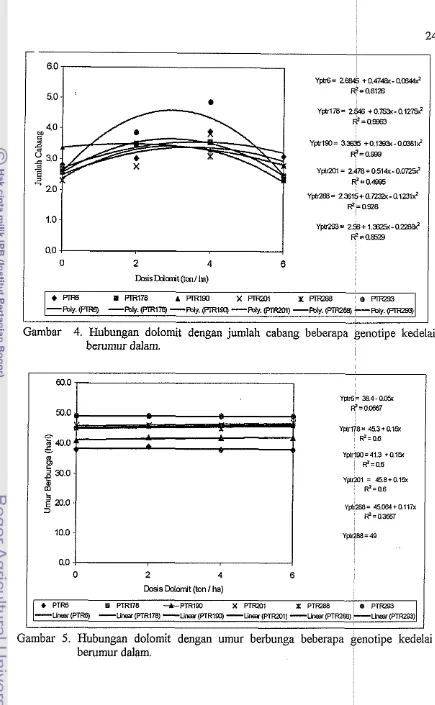
4 Hubungan dolomit dengan jumlah cabang beberapa genotipe
...
kedelai berumur dalam...
.
.
.
5 Hubungan dolomit dengan umur berbunga beberapa genotipe
...
kedelai berumur dalam...
.
...
...
...
6 Hubungan dolomit dengan umur panen beberapa genotipe kedelai
...
berumur dalam7
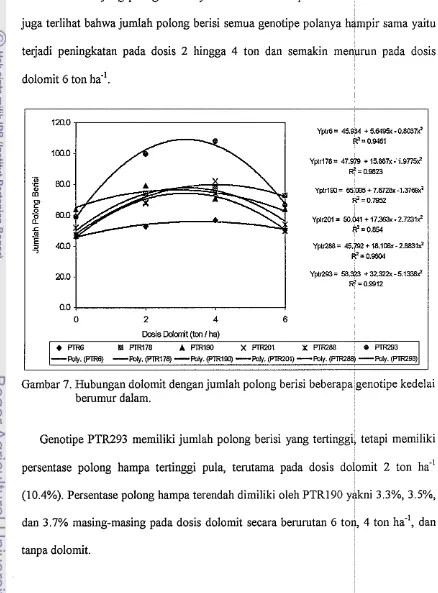
Hubungan dolomit dengan jumlah polong berisi beberapa genotipe...
kedelai berumur dalamDAPTAR LAMPIRAN
1 Analis ragam bobot kering bintil a k a , bobot kering akar bob kering tajuk, dan serapan Ca dan Mg tajuk ...
.
.
.
.
.
2 Analisis ragam kadar Ca dan Mg tajuk, umur berbunga, umur pan! dan tinggi tanaman
... .
.
.
...
3 Analisis ragam jumlah cabang, polong berisi, polong hampa, bobc
100 biji, dan bobot biji tanaman-'
...
.
.
...4 Analisis ragam bobot biji petak"
...
5 Pengaruh dolomit dan genotipe terhadap serapan Ca dan Mg taju kadar Ca tajuk, umur berbunga, dan umur panen
... ...
6 Pengaruh dolomit dan genotipe terhadap jumlah polong berisi, pe sentase polong hampa, dan jumlah cabang ...
7 Hasil analisis sifat kimia dan fisika tanah lokasi percobaan sebelum perlakuan dan analisis dolomit
...
8 Rata-rata curah hujan, frekuensi curah hujan, suhu udara, kelen baban relatif, dan penyinaran pada stadia vegetatifa dan generatif.
PENDAHULUAN
Latar Belakang
Kedelai (Glycine n~a. (L.) Memll) merupakan komoditi pang:
yang terjangkau oleh semua lapisan masyarakat. Selain untuk pe
kedelai banyak digunakan untuk pakan dan bahan baku industri.
nasional masih belum cukup untuk memenuhi kebutuhan sendiri a
mengimpornya dengan jumlah yang cukup besar. Dalam periode
rata-rata volume impor kedelai sebesar 1.1 juta ton tahun-' dalam be
bungkil, dan minyak kedelai. Impor tertinggi terjadi pada tahun 1991
ton karena berkurangnya luas panen (Manurung 1999).
Penurunan luas panen mengakibatkan terjadinya penurunan prod
Pada periode tahun 1983-1993 luas panen rata-rata 1.4 juta ha deng
rata 1.5 juta ton, periode tahun 1993-1998 rata-rata luas panen 1.
produksi rata-rata 1.4 juta ton sementara kebutuhan diperkirakan sa
+
2.5 juta ton (Manurung 1999)Strategi peningkatan kedelai untuk mencapai kebutuhan nasi
melalui perluasan areal. Menurut Sumarno (1999), perluasan areal
pada berbagai tipe lahan dan agroekologi, antara lain, lahan sav
kemarau, lahan bukaan yang berasal dari lahan pasang surut dan lak
lahan sawah tadah hujan pada awal musim hujan sebelum padi. Dul
diperlukan untuk perluasan areal tanam, antara lain, berupa pengal
dengan harga
an, dewasa ini
oduksi kedelai
ngga terpaksa
un 1991-1998
~k biji, tepung,
:besar 1.7 juta
;i dalam negri.
produksi rata-
uta ha dengan
~ a i tahun 2001
11 diutamakan
tpat dilakukan
pada musim
gambut, serta
lgan teknologi
masam, perbaikan sistem hidrologi, penyediaan benih bermutu
struktur serta kesuburan tanah.
Penelitian selama dekade terakhir menunjukkan bahwa tanam
beradaptasi dengan kondisi lahan basah yang jenuh air serta
menambat nitrogen lebih tinggi daripada dengan budidaya sec,
(Troedson et 01. 1983; Lawn 1985;Garside et
crl.
1992; Raka 1993;1991).
Menurut Sumarno (1986), budidaya kedelai pada lahan basah
tanah di bawah daerah perakaran jenuh air secara terus-men
pertumbuhan tanaman hingga masak fisiologi. Hunter et rrl. (1980) r
budidaya jenuh air merupakan penanaman dengan memberikan irig:
dan membuat tinggi muka air tetap (f 5 cm di bawah permukaar
lapisan di bawah perakaran jenuh air.
Hasil penelitian Ghulamahdi (1999) pada budidaya jenuh air (BJ
tadah hujan (BTH) dengan berbagai genotipe kedelai, baik berumi
genjah yang diberi kapur 2 ton ha.', menunjukkan bahwa BJA men
leher akar, bobot kering bintil, aktivitas nitrogenase, serapan h
nitrogen, kalium, besi, mangan, bobot kering tanaman, dan bobot kc
tetapi nyata menurunkan kandungan kalsium dan magnesium da
dengan BTH. Produksi kedelai tertinggi dicapai pada genotipe ben
dibandingkan yang berumur genjah, juga pada BJA.
Berdasarkan uraian di atas perlu diteliti dosis pemupuka
dibutuhkan dan genotipe kedelai berumur dalam yang dapat beradap
2
dan perbaikan
kedelai dapat
rproduksi dan
I konvensional
iulamahdi et 01.
:mbuat kondisi
.IS sejak awal ~yatakan bahwa
terus-menenis
anah) sehingga
) dan budidaya
dalam maupun
:katkan lingkar
3 daun, kadar
ng biji petak-',
dibandingkan
ur lebih dalam
dolomit yang
T i ~ j u a n P e r c o b a a ~ ~
1. Mempelajari tanggap genotipe kedelai berumur dalam terhadap
berasal dari pupuk dolomit pada sistem
BJA.
2. Mempelajari tanggap hasil dan karakter agronomi kedelai
terhadap pemupukan dolomit pada sistem
BJA.
Hipotesis Percobaan
1. Tanggap masing-masing genotipe berbeda terhadap Ca I
pemupukan dolomit pada sitem
BJA.
2. Karakter agronomi dan hasil masing-masing genotipe memberi:
berbeda terhadap pemupukan dolomit pada sistem
BJA.
3
:a dan Mg yang
berumur dalam
.n Mg dengan
TINJAUAN PUSTAKA
Budidaya J e n u l ~ Air dan Tanggap Tanaman Kedelai
Masalah kelebihan air sesaat merupakan keadaan umuln yang terjadi pada pola
penanaman di daerah tropis dan subtropis. Kelebihan air dapat terjadi karena periode
yang panjang dari cuaca basah dan curah hujan tinggi setelah irigasi (Troedson et al.
1983). Sumamo (1986) mengemukakan bahwa di Indonesia ma
juga terjadi pada lahan sawah yang akan dimanfaatkan untuk
setelah padi dipanen. Keadaan ini disebabkan oleh adanya lap
kedalaman 15-20 cm di bawah permukaan tanah. Sebaliknya j
lapisan kedap air membatasi penetrasi perakaran dan tanaman
et al. 1985).
Kedelai relatif toleran terhadap kelebihan air sesaat dibandi
kacangan lain, dan cepat memperbaiki pertumbuhannya setelah air berkurang
(Stanley et al. 1980). Percobaan yang dilakukan Talnpubolon (1988) menunjukkan
bahwa penggenangan terputus-putus menghambat pertumbuha
produksi tanaman kedelai. Menurut Garside et al. (1992), pembe
inenerus pada budidaya jenuh air dapat memperbaiki status ai
sehingga pertumbuhan vegetatif dari tanaman terpacu. Menurut
budidaya jenuh air merupakan penanainan dengan memberikan irig i taus-menerus
dan menlbuat tinggi muka air tetap (f 5 cm di bawah permuka
lapisan di bawah perakaran jenuh air. Air diberikan sejak beru
5 Menurut Lawn (1985), budidaya jenull air pada tanaman kedelai hampir sama
dengan budidaya tanarnan padi sawah. Perbedaamya terletak pada ketinggian
permukaan air. Pada budidaya jenuh air tinggi muka air berada beberapa centimeter
di bawah permukaan tanah, sedangkan pada padi sawah beberapa centimeter di atas
permukaan tanah. Irigasi pada budidaya jenuh air dilakukan deng ara alur (fiurrow
irrigation).
Fase aklimatisasi tanaman kedelai terhadap kondisi lahan je air berlangsung
selama 2 minggu (Troedson et al. 1983) atau 2-4 minggu setel ngairan dimulai
(Lawn 1985). Pada fase tersebut akar dan bintil akar di bawah permukaan air mati.
Matinya akar dan biitil akar menyebabkan berkurangnya penyerapan nitrogen
sehingga tanaman menunjukkan gejala klorosis (Troedson et al. 1983). Lebih ianjut
klorosis menyebabkan proses fotosintesis berjalan tidak normal, dan terjadi
translokasi hasil fotosintesis ke bagian bawah tanalnan untuk pertumbuhan akar dan
bintil akar yang baru, menyebabkan bobot kering tanaman pada budidaya jenuh air
lebih rendah dibandingkan dengan cara konvensional. Walaupun terjadi gangguan
pertumbuhan pada tahap aklimatisasi, setelall tahap tersebut tanaman kedelai
menunjukkan pertumbuhan akar dan bintil akar baru yang cepat dan banyak pada
lapisan tanah di atas permukaan air. Selanjutnya tanaman menjadi hijau dan tumbuh
cepat dengan laju pertumbuhan lebih tinggi pada budidaya jen r dibandingkan
pada budidaya biasa atau konvensional (Ralph 1985).
Nathanson et al. (1984) menyatakan bahwa kandungan N pada budidaya
jenuh air meningkat, bahkan menjadi sama dengan yang diperoleh pada budidaya
6 varietas kedelai terhadap keadaan jenuh air berbeda-beda. Kedelai yang berunlur
lebih panjang, pertumbuhannya lebih baik dan produksinya lebih tinggi dibanding
kedelai berumur pendek (Hunter et al. 1980; CSIRO 1983; Ghulamahdi et al. 1991).
Hasil penelitian Nurlianti (1997) menunjukkan ballwa b
menyebabkan kemunduran waktu panen bila dibandingkan u
berdasarkan diskripsinya di lahan kering. Mundurnya waktu pa rsebut berkisar
6-32 hari. Mugnisjah (1996) mendapatkan kisaran keterlambat
hari. Pada penelitian Raka (1993) menyatakan kisaran waktu p
sedangkan hasil penelitian Ghulamahdi (1999) waktu panen mundur 2-3 hari.
Nurlianti (1997) menyatakan bahwa kelompok varietas yang berumur genjah di lahan
kering bila dibudidayakan di lahan jenuh air umur panennya
kelompok berumur sedang atau berumur dalam; varietas berumur
menjadi berumur dalam; varietas berumur dalam tetap berada
berumur dalam.
Menurut Ghulamahdi (1999), perubahan yang terjadi pada b
sejak awal sampai panen yaitu dari perubahan fisiologi sampai ke peningkatan bobot
biji petak", dimulai dari meningkatnya ACC (I-aminocyclopropane-1-carboxylic
acid) akar dan etilen akar. Etilen akar menyebabkan meningkatnya kandungan
glukosa dan sukrosa akar yang dipergunakan untuk pertumbuhan perakaran; etilen
akar juga menyebabkan terbentuknya jaringan aerenkima sehin
lingkar leher akar. Keadaan ini meri~pakan tahap penyesuaian tan
budidaya jenuh air. Pertumbuhan akar-akar baru yang berlangsu
7 dan serapan hara ole11 akar. Ole11 karena itu, pertu~nbuhan akar-akar baru
meningkatkan serapan hara daun sehingga meningkatkan jumlah polong isi
tanaman" dan akhirnya meningkatkan bobot kering biji petak-'.
Keadaan Tergenang
Segera setelah tanah digenangi kadar 0 2 tanah berkurang drastis dan akhirnya
habis karena didesak air dan difksinya terhanlbat oleh lapisan air (Leiwakabessy
1993). Selanjutnya kadar beberapa gas seperti C02, N2, C&, dan Hz yang terbentuk
selanla penggenangan meningkat karena terhalang keluarnya oleh lapisan air.
Konsentarsi NO-3 menjadi lebih rendah dalam jaringan tanaman pada keadaan
tergenang. Pengamatan menunjukkan bahwa nitrat tanah akan direduksi menjadi
komponen gas seperti N2 dan N2O yang tidak tersedia bagi tanaman. Nitrogen
diangkut dari daun tua ke daun muda pada keadaan tergenang (Drew d m Sisworo
1979). Kahat nitrogen disebabkan oleh denitrifikasi dan pencucian, serta menurunnya
absorbsi dan translokasi N dalam jaringan tanaman (Tsai dan Chu 1992).
Menurut Drew dan Sisworo (1979), pengan~h penggenangan terhadap
pengambilan P tidak konsisten, sedangkan terhadap ion K konsisten, menurun dalam
jaringan tanaman pada keadaan tergenang. Pengaruh mangan dan besi dapat ditekan
dengan CzH4 (Drew 1992). Pembentukan jaringan aerenkiina meningkatkan kapasitas
0 2 yang mempercepat perubahan ACC menjadi etilen (Ghulamahdi 1999).
Sebahagian besar P tersedia dan kalsium yang dapat dipertukarkan berada di
dalam tanah permukaan. Kalium d m magnesium yang dapat dipertukarkan
8
dan magnesium di dalam propil tanah menunjukkan penurunan. Kandungan
N&+N
bergerak dengan cepat ke bawah pada kedalaman 50 cm (Yoo et nl. 1987).
Pengapuran
Bahan kapur menurut definisi Barber (1967) terdiri dari bahan-bahan kalsium dan
magnesium yang mampu menetralkan tanah masam. Bahan kapur berupa dolomit
yang diberikan ke dalam tanah akan mengalami reaksi sampai terbentuk
keseimbangan baru. Reaksi yang terjadi pertama kali adalah penguraian bahan kapur
mernbentuk ion CO j serta ion ~ a + + dan M ~ + + . Selanjutnya ion C03 yang terbentuk
menarik ion
HC
dari kompleks jerapan.Soepardi (1979) menyatakan bahwa kation yang paling cocok untuk memperbaiki
kemasaman tanah adalah Ca dan Mg. Keduanya mempunyai perbedaaan dalam ha1
kecepatan reaksinya. Kalsit bereaksi lebih cepat daripada dolomit. Bila kecepatan
reaksi bukan merupakan pertimbangan penggunaan bahan kapur, dolomit dianggap
lebih menguntungkan dibanding kalsit karena dalam bahan dolomit terdapat sejumlah
unsur Mg.
Secara uinum pemberian kapur dapat mempengaruhi sifat fisik, kimia, dan biologi
tanah (Nurhayati 1982). Pengapuran menjadikan tanah lebih gembur sehingga
menguntungkan pertumbuhan tanaman dan akar dapat menjangkau unsur hara pada
lapisan yang lebih dalam. Pemberian kapur dapat menurunkan kadar dan kejenuhan
Al, meningkatkan pH tanah yang diikuti dengan peningkatan ketersediaan P, K, Ca,
dan Mg serta C organik, dan menurunkan ketersediaan Mn dan Fe. Penambahan
9 diperkiran Ca untuk pembentukan bintil akar lebih besar daripada keperluan Ca untuk
pertunlbuhan tanaman sendiri (Kamprath 1970). Serapan Ca sangat penting untuk
pertunbuhan bintil akar pada tanaman kacang-kacangan (Suhartatik 1986).
Gemawi (1986) menyatakan bahwa pernberian kapur sangat nyata mempengaruhi
tinggi tanaman, jumlah cabang, jumlah polong, dan produksi ha". Lebih lanjut
dikatakan bahwakanaman yang tidak dikapur memiliki pertambahan tinggi tanaman
yang kurang dan lebih cepat mengering, bobot bintil akamya kurang dan jumlah
polong serta prodnksinya rendah emberian
P
6 ton kalsit dan dolomit ha-' nyata dapatmeningkatkan pertumbuhan tanaman, terutama tinggi tanaman, bobot kering
brangkasan, jumlah bintil akar, dan jumlah cabang produktif serta nyata pada
peningkatkan produksi biji kering (Djuhartono 1989).
Serapan Hara Tanaman
Kalsium dan magnesium merupakan kation-kation utama pada kompleks
pertukaran. Keduanya inempunyai sifat dan perilaku yang mirip di dalam tmah
(Sopardie 1979). Menurut Jones (1998), kalsium berperan dalam pemeliharaan sel,
permeabilitas membran, peningkatan pertumbuhan dan perkecambahan, berperan
aktif pada enzim dalam penyusunan, pembelahan, dan pemanjangan sel. Daerah
ineristematik di akar, batang, dan daun yang selnya aktif membelah merupakan
bagian paling peka karena Ca dibutuhkan untuk membentuk lamela tengah baru pada
lempeng sel, yang tumbuh di atas dua sel anak (Salisbury dan Ross 1992).
Kadar Ca dalam tanaman berkisar antara 0.2 sampai beberapa persen
10 pada kebutuhan minimum. Banyak jenis tanaman dapat tumbuh dengan nornlal dalam
larutan pada konsentrasi kalsium 2 ppm atau 0.05 ppm, yang menghasilkan 0.01 1
persen Ca dalam daun jagung dan 0.08 persen dalam daun tembakau dengan
ketentuan bahwa kadar ion-ion divalen lainnya rendah. Jones (1998) menduga
kandungan Ca dalam tanaman 0.20-5.00 persen berdasarkan bobot kering, dan
dengan nilai kecukupan 0.30-3.00 persen.
Di rizosfer Ca mencapai permukaan akar melalui aliran massa. Dari analisis
diketahui bahwa kebutuhan Ca temyata cukup rendah, juga terjadi penulnpukan pada
permukaan akar, pada dinding sel (apoplas) dan pada permukaan eksterior membran
plasma (Marschner 1995). Berbagai enzim di aktifkan oleh Ca tetapi banyak pula
yang dihambat. Penghainbatan ini mendorong sel me~npertahankan konsentrasi Ca
lebih rendah lagi dalam sitosol tempat enzim berada (Salisbu~y dan Ross 1992).
Akibat dari kahat Ca lebih parah pada pertumbuhan akar jika dibandingkan dengan
pertumbuhan pucuk. Pertumbuhan akar sangat terhambat, akar rusak, berubah warna
dan mati. Kekurangan Ca pada bunga dan buah juga sangat parah dan dikenal sebagai
penyakit "blossom ent-rot" (Sopandie 1997). Kandungan Ca yang berlebihan
lnengakibatkan kekurangan pada Mg atau K, bergantung pada konsentrasi kedua
unsur tersebut dalam tanaman (Jones 1998).
Magnesium merupakan bagian dari klorofil dan mempengaruhi aktivitas enzim
yang berhubungan dengan transfer P. Karenanya, kahat Mg melnpengaruhi aspek
n~etabolisrne tanaman (Sopandie 1997). Menurut Salisbury dan Ross (1992), Mg
halnpir tidak pemah menjadi pembatas pada tumbuhan yang hidup di tanah. Di
~nenjadikan ATP berfungsi dalam berbagai reaksi, mengakibatk
yang diperlukan dalam fotosintesis, respirasi dan pembentukan
nucleic acid) serta RNA (ribonucleic acid).
Kanduulgan Mg dalam tanaman berkisar antara 0.15 dau 1 .OO
bobot kering dam, dengan nilai kecukupan sekitar 0.25 persen p
beberapa tanaman. Nilai kritis Mg sangat bervariasi pada seti:
terendah pada tanaman biji-bijian dan tinggi pada tanaman leg
tanaman sayuran serta buah-buahan (Jones 1998).
Kahat Mg didahului dengan terjadinya klorosis yang diikuti d
laju fotosintesis dan asimilasi, kemudian senyawa N larut tertimb~
akhirnya kahat Mg menyebabkan pertumbuhan kloroplas, mit~
abnormal (Sopandie 1997). Magnesium berakibat antagonisme d
artinya peningkatan penyerapan Mg mengurangi penyerapan Ca dan
Menurut Small dan Ohlrogge (1973), kandungan hara d
mencukupi untu~k perturnbuhan kedelai pada saat berbunga adalaf
0.26-0.50%,
K
1.71-2.50%, Ca 0.36-2.00%, dan Mg 0.26-1Ghulamahdi (1999) dengan budidaya jenuh air pada tiga genot
(Wilis, PTR 8, dan PTR 322) menghasilkan kandungan hara (
masing-inasing sebagai berikut: Ca 0.25-0.35%, dan Mg 0.27-0.29
0.23-0.24%, dan Mg 0.27-0.28% pada PTR8; Ca 0.31-0.34%, dai
pada PTR322.
Menurut Marschner (1995), ada beberapa faktor penyebab l a j ~
dan air yang menurun dari ujumg akar ke pangkal akar, antara lain:
11 banyak enzim
NA (deoxyribo
;en berdasarkan
2 jaringan daun
jenis tanaman,
I dan berbagai
:an merosotnya
ialam tanaman,
~ndria menjadi
;an Ca dan K,
I dalaln batas
.1 4.26-5.5%, P
)%. Penelitian
: kedelai yaitu
n Ca dan Mg
pada Wilis; Ca
dg 0.27-0.29%
myerapan hara
12 suberin (gabus) pada rizodermis; (2) pembentukan endodermis sekunder dan tersier
secara radial ke stele dan; (3) degradasi sel-sel korteks sehingga membentuk celah-
celah dalam daerah kotteks yang disebut aerenkima. Sepanjang perjalanan di dalam
xylem, zat hara diserap kembali oleh sel-sel hidup sejak dati akar sampai ke daun.
Karenanya tanaman yang ditanam dalam larutan hara menghasilkan konsentrasi ion-
ion tertentu yang menurun drastis dari akar sampai ke pucuk. Gejala ini jelas sekali
pada tanaman jenis kacang-kacangan (Sopandie 1997).
Sorooshzadeh dan Batthakur (1995) dari hasil penelitiannya melaporkan bahwa
pada biji dan polong tanaman kedelai yang diairi (irigasi) mengakibatkan absorbsi
kalsium lebih banyak daripada yang tanpa diairi. Lebih lanjut dinyatakan bahwa di
bawah cekaman air, Ca tidak ditranslokasikan dari polong ke daun selama periode
pengisian polong. Menurut Marschner (1995), tingkat remobilisasi hara mikro dan
Ca selama tahap reproduktif seringkali sangat tinggi bila dibandingkan dengan saat
pertumbuhan vegetatif
Kandungan N, K, Fe, dan Mn dari kedelai pada budidaya jenuh air lebih tinggi
dibanding dengan pada budidaya tadah hujan, sedangkan kandungan P daun sama
pada kedua perlakuan tersebut. Akan tetapi, kandungan Ca dan M g daun pada
budidaya jenuh air lebih rendah dibandingkan dengan pada budidaya tadah hujan.
Rendahnya kandungan Ca dan Mg tersebut disebabkan Ca dan Mg banyak diperlukan
di bagian bawah untuk membentuk akar-akar barn sehingga transportasi ke bagian
atas menurun dan dengan bettambahnya bobot daun, Ca dan Mg daun menjadi lebih
BAHAN DAN METODE
Tempat dan Waktu
Percobaan dilaksanakan di Kebun Percobaan Cikarawang I
Pertanian Bogor, pada ketinggian tempat 240 m dpl., dengan jer
-.
(Oxic Dystropept). Percobaan ini berlangsung dari bulan Mei samp
Proses pengeringan dan analisis kadar Ca dan Mg tajuk tanarr
laboratorium Balai Penelitian Bioteknologi Tanaman Pangan, Bogo
sampai Agustus 2000.
Metode
Rancangan yang digunakan dalam percobaan ini adalah rancang
dalam acak kelompok dengan tiga ulangan. Petak utama adalah dosi:
dan 6 ton ha-') dan anak petak adalah enam genotipe kedelai berun:
1). Denah percobaan dan penempatan perlakuan secara acak di lapar
[image:102.595.84.519.46.717.2]pada Lampiran 1.
Tabel 1. Genotipe kedelai berumur dalam dan asal persilangannya
91 - 070 :
Dempo x
-
Amerikan Kerinci xNomor 1 2 3 4 5 6
I I I
Keterangan : #) Berasal dari koleksi Bapak Dr.
II.
Fred Rutnawas, M! Budidaya Pertanian Fakultas Pertanian, Institut PertaniarGenotipe# PTR6 PTR178 PTR190 PTR 201 PTR288 PTR293
maga, Institut
tanah Latosol
4gustUs 2000.
dilakukan di
lari bulan Juni
petak terpisah
~lomit (0, 2, 4,
dalam (Tabel
n dapat dilihat
Benih kedelai diinokulasi dengan inokulum Bradyrhizobiunz
berasal dari inokulum yang diperoleh dari Dr. Ir. Rasti Saraswati,
BioteknoIogi Tanaman Pangan Bogor. Rhizoplus merupakan
multiguna (PMMg), pengernbangan dari inokulum Rhizobiunl
beberapa jenis mikroba multiguna (Bradyrhizobitrm dan mikroba
Indonesia (Saraswati et al. 1997; Sunarlim 1997).
Model linear yang gunakan adalah sebagai berikut :
i = 1,2,3 ( ulangan)
j = 1,2,3,4 (dosis dolomit)
k = 1,2, 3,4, 5, 6 (genotipe beru~nur dalam) dengan
Yijk = hasil pengamatan dosis dolomit ke-j dan genotipe 1
ulangan ke-r
P = nilai rata-rata umum
pi = pengamatan aditif ulangan ke-r
'4 = pengaruh aditif perlakuan petak utama ke-j
Yij = pengaruh aditif galat perlakuan petak utama ke-j I
Pk
= pengaruh aditif perlakuan anak petak ke-k(ap)jk = pengamh interaksi antara dosis dolomit ke-j dan ge Eijk = pengaruh galat yang timbul dari dosis dolomit ke-j
ke-k pada ulangan ke-r.
Analisis data yang menunjukkan pengaruh perlakuan yang berl
taraf 5% uji
F
dalam analisis ragam dilanjutkan dengan uji beda nilaMultiple Range Test (DMRT). Untuk melihat hubungan antara pen
dan karakter agronomi yang interaksinya nyata dilakukan uji re(
?panicurn yang
lalai Penelitian
upuk mikroba
romersial dari
3elarut P) asal
-k pada
mgan ke-r
otipe ke-k an genotipe
:da nyata pada
tengah Duncan
ipuran dolomit
15 Pelaksanaan
Tanah untuk percobaan diolah dengan menggunakan bajak kemudian diratakan
dengan cangkul, akhirnya dibersihkan dari sisi-sisa gulina. Anak petak percobaan
berukuran 2 m x 5 m.Yang berperan sebagai saluran air adalah Jarak antarpetak
utaina dan antarulangan 0.30 m. Petak utama dikapur sesuai dengan dosis masing-
masing perlakuan, dan diinkubasi selama 4 minggu. Pupuk SP 36 sebanyak 200 kg
ha-' dan KC1 100 kg ha" diberikail bersamaan pada saat pengapuran dengan
mencampur pupuk dan kapur menjadi satu.
Benih kedelai ditanam langsung di lapangan setelah terlebih dahulu diinokulasi
dengan rhizobiu~n jenis Biolestari (5 g kg-' benih). Jarak tanam yang digunakan
adalah 50 cm x 7.5 cm dengan 1 butir setiap lubang. Untuk ~nengatasi lalat kacang,
sebelum tanam benih diberi karbosulfun (Marshal 25 ST) sebanyak 15 glkg benih.
Untuk inengatasi tumbuhnya gulma dilakukan penyemprotan herbisida pratumbuh
0
dengan oxyflourfen (Goal 2E) bertakaran 1,25 1 ha.' (2 ml 1.' air) satu hari setelah
tanam. Gangguan hama diatasi dengan penyemprotan insektisida Deltametrin (Decis
2.5 EC) pada takaran 2.5 ml I-' dengan volume semprot 500
1
ha" pada umur 3, 5,6,dan 8 minggu setelah tanam (MST). Pengairan diberikan dari umur 14 hari setelah
tanain sampai polong benvama coklat dengan tinggi muka air tetap dipertahankan i 5
cm di bawah permukaan tanah sehingga lapisan di bawah perakaran jenuh air (Hunter
et 01. 1980). Pengaturan tinggi nluka air diatur sedemikian rupa sehingga diharapkan
16
Panen dilakukan jika daun telah mulai rontok dan 90 % polong benvarna coklat.
Pemanenan dilakukan dengan memotong pangkal batang dengan sabit bergerigi,
kemudian brangkasan dimasukkan ke dalam karung panen sesuai perlakuan. Hasil
panen kemudian dijemur sampai kering atau polong mulai pecah, lalu biji
dikeringkan sampai mencapai kadar air 14 %.
Pengamatan dan Analisis
Peubah yang diamati meliputi komponen peitumbuhan, kadar hara, komponen
hasil, hasil, data iklim, dan sifat fisik dan kimia tanah selama perlakuan. Komponen
pertumbuhan dan komponen hasil diukur berdasarkan rata-rata dari 10 tanaman
contoh, kecuali bobot biji dan hasil biji.
1. Serapan dan kadar C a dan M g tajuk.
-4nalisis hara tajuk dilakukan dengan mengambil sampel tajuk dari lapangan,
kemudian dikeringkan dalam oven dengan suhu 105' C selama 3 hari. Tajuk
dihaluskan dengan alat penggiling. Ca dan Mg dianalisis dengan metode HC104
+
HN03 dengan alat atomic absorbtion spectrometer (AAS). Pengamatandilakukan pada umur 8 minggu setelah tanam.
2. Bobot kering bintil a k a r (g).
Pengamatan bobot kering bintil akar dilakukan dengan memisahkan bintil akar
yang aktif, dikeringkan dengan oven pada suhu 105' C selama 72 jam lalu
3. Bobot kering tajuk dan a k a r t a ~ ~ a m a n - ' ( ~ ) .
Tanaman contoh untuk pengukuran bobot bintil akar di atas dipisahkar, antara
tajuk dan akarnya. Tajuk dan akar dikeringkan dalam oven pada suhu 105' C
selama 72 jam, atau sampai bobotnya konstan, dan ditimbang.
4. W a l t u berbunga (hari).
Waktu berbunga ditetapkan jika 50% dari total tanaman telah berbunga pada
petakan.
Waktu panen ditentukan jika 90% polong telah benvarna coklat di setiap petak.
6 . Tinggi tanaman (em).
Tinggi tanaman diukur dari permukaan tanah sampai titik tumbuh pada setiap
tanaman sampel. Pengamatan dilakukan pada saat panen.
7. Jumlah cabang tmaman-' (buah).
Jumlah cabang per tanaman dilakukan dengan menghitung semua cabang yang
berada pada batang utama pada setiap tanaman sampel saat panen.
8. Jumlah polong isi (buah) dan polong hampa tanaman-' (%).
Pengamatan polong isi dan hampa per tanaman dilakukan pada saat panen
dengan menghitung persentase polong yang berisi dan hampa dari seluruh polong
yang terbentuk pada tanaman sampel.
9. Bobot 100 biji (g).
Pengamatan dilakukan dengan menimbang 100 butir biji yang berkadar air 14%
10 Bobot biji tanaman" (g).
Pengamatan dilakukan dengan menimbang biji kering yan,
tanaman sampel dari masing-masing petak percobaan.
11. Hasil biji (g).
Pengamatan dilakukan dengan menimbang biji kering yang dih
percobaan.
12. Hasil (ton ha-').
Hasil tanaman dihitung dari hasil biji per petak, selanjutnya
satuan hektar.
13. Analisis t m a h awal dan data agroklimatologi.
Pada analisis tanah ini diamati kandungan pasir, debu, liat; pH,
P total, K total, PzOs, dan KzO; kation K, Na, Ca, Mg, dan KTE
Mn, Zn, dan Cu, serta A d d . Kandungan pasir, liat, dan debu (
metode pipet; pH dengan metode pH meter; N dengan me
dengan metode Kurmis;
P
total dan K total dengan metodedengan metode Bray I; kation K, Na, Ca, dan Mg dengan ml
dengan metode titrasi; dan unsur mikro Fe, Mn, Zn dan Cu den,
A d d dengan metode titrasi. Pengamatan data agroklimatologi
hujan, dan kelembaban relatif dilakukan sejak tanam sampai
percobaan selesai dipanen.
ihasilkan tiap
;an tiap petak
~nversi dalam
dungan N, C,
isur mikro Fe,
tukan dengan
: Kjeldahl; C
:I 25%; PzOs
e AAS; KTK
metode AAS;
.I
suhu, curahH A S I L
Serapan dan I<adar Ca Mg Tajuk
Dolomit dan genotipe serta interaksinya tidak nyata mempen
dan M g tajuk, tetapi dolomit nyata mempengaruhi kadar Ca dan
tidak nyata mempengaruhi kadar Ca tajuk, tetapi interaksi dolo
nyata mempengaruhi kadar Ca tajuk (Lampiran 1).
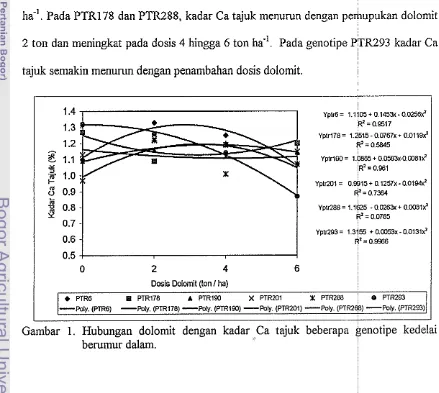
Gambar 1 menunjukkan bahwa kadar Ca tajuk setiap genoti
dosis dolomit. PTR6, PTR190, dan PTR201 polanya hampir sama yaitu kadar Ca
tajuk meningkat pada dosis 2 hingga 4 ton dan menurun pada dosis dolomit 6 ton
ha-'. Pada PTR178 dan PTR288, kadar Ca tajuk menurun dengan p
2 ton dan meningkat pada dosis 4 hingga 6 ton ha.'. Pada genotipe
tajuk semakin menurun dengan penambahan dosis dolomit
0.5
5
0 2 4 6
Dmts D o l m ~ t (ton / ha)
+ PTR6 U PTR178 A PTRIEO X PTR201 X PTR288 e PTR253
-Pow (PTRG) --Pow (PTR178) -Pow (PTRIEO) -Poly (PTR201) -Poly (PTR288) -Poly (PTR253)
[image:108.595.77.516.334.727.2]140
-
120s"
100-
$3 z 80C 60
8
40 "I 20 00 2 4 6
Dosis Dolonlit (ton/ Ira)
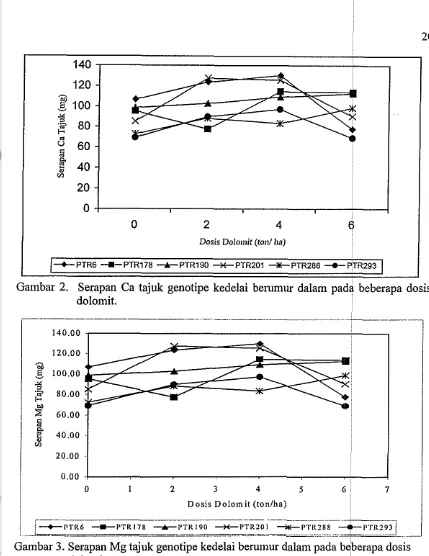
Garnbar 2. Serapan Ca tajuk genotipe kedelai berumur dalam pada beberapa dosis dolomit.
0.00
[image:109.602.86.515.40.596.2]0 1 2 3 4 5 6 7 Dosis Dolomit (tonlha)
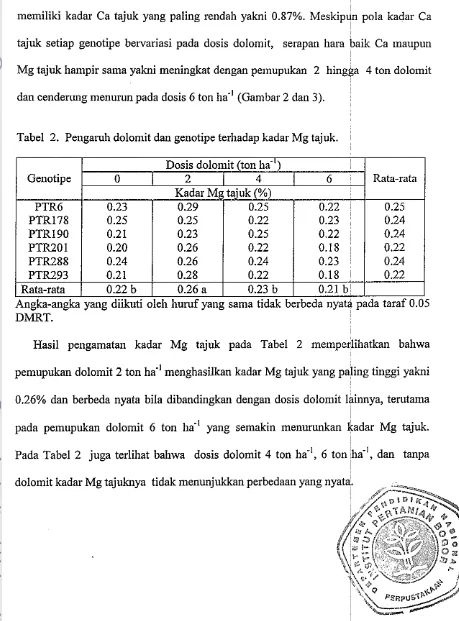
Gambar 3. Serapan Mg tajuk genotipe kedelai berumur dalam pada beberapa dosis dolomit.
Pada Lampiran 5 dan Gambar 1 juga memperlihatkan bahwa kadar Ca tajuk
genotipe PTR6 nyata lebih tinggi yakni 1.33% pada dosis dolomit
2
ton ha" dan2 1 PTR201 tanpa dolomit, PTR178 pada dosis dolomit 4 dan 6 ton ha-', PTR288 pada
dosis dolomit 4 ton ha-', dan PTR293 pada dosis dolomit 6 ton ha.' yang juga
memiliki kadar Ca tajuk yang paling rendah yakni 0.87%. Meskipun pola kadar Ca
tajuk setiap genotipe bel-variasi pada dosis dolomit, serapan hara baik Ca maupun
Mg tajuk hampir sama yakni meningkat dengan pelnupukan 2 hingga 4 ton dolomit
dan cendert~ng menurun pada dosis 6 ton ha" (Gambar 2 dan 3).
Tabel 2. Pengaruh dolomit dan genotipe terhadap kadar Mg tajuk.
Hasil pengamatan kadar Mg tajuk pada Tabel 2 mernperlihatkan bahwa Rata-rata 0.25 Genotipe PTR6 PTR178 PTR190 PTR201 PTR288 PTR293 Rata-rata
pemupukan dolomit 2 ton ha-' menghasilkan kadar Mg tajuk yang paling tinggi yakni
0.26% dan berbeda nyata bila dibandingkan dengan dosis dolomit lainnya, terutama Dosis dolomit (ton ha-')
Angka-angka yang diikuti oleh huruf yang sama tidak berbeda nyata pada taraf 0.05 DMRT. 0.25 0.21 0.20 0.24 0.21 0.22 b
pada pemupukan dolomit 6 ton ha-' yang semakin menurunkan kadar Mg tajuk. 0
Pada Tabel 2 juga terlihat bahwa dosis dolomit 4 ton hd', 6 ton ha-', dan tanpa 0.25 0.23 0.26 0.26 0.28 0.26 a 2
Kadar Mg tajuk (%)
0.22 0.25 0.22 0.24 0.22 0.23 b 4 0.23 6 0.23 0.22 0.18 0.23 0.18 0.21 b 0.29 0.24 0.24 0.22 0.24 0.22
[image:110.595.85.544.131.752.2]Pertumbuhan dan Umur Tanaman
Dolomit tidak nyata mempengaruhi bobot kering bintil akar, bobot kering akar,
bobot kering tajuk, dan tinggi tanaman, tetapi genotipe nyata mempengaruhi bobot
kering bintil akar, bobot kering tajuk, tinggi tanaman, dan jumlah cabang. Interaksi
dolomit dan genotipe nyata mempengaruhi jumlah cabang (Lampiran 1,2, dan 3).
Tabel 3. Pengaruh dolomit dan genotipe terhadap bobot kering bintil akar, bobot kering akar, dan bobot kering tajuk
[image:111.595.76.514.188.724.2]23 Genotipe PTR293 menghasilkan bobot kering bintil akar terendah dan berbeda
nyata dengan PTR6, PTR178, PTR190, PTR201, dan PTR288, tetapi kelima genotipe
yang disebut terakllir tidak berbeda nyata (Tabel 3). Pada Tabel 3 juga terlihat bahwa
bobot kering bintil akar paling tinggi pada PTR190. Bobot kering akar juga tinggi
pada PTR190 dan rendah pada PTR288 meskipun secara statistik tidak nyata
perbedaannya. Tinggi tanaman tertinggi juga pada PTR190 dan berbeda nyata dengan
[image:112.595.85.513.200.734.2]genotipe lainnya, terutama PTR6 yang karakter tinggi tanamannya terendah
(Tabel 4).
Tabel 4. Pengaruh dolomit dan genotipe terhadap tinggi tanaman.
Genotipe
I
Tinggi tanaman (cm)I
I
PTR 293 42.10 40.20 41.73 39.731
40.94 bcI
PTR 6 PTR 178 PTR 190 PTR 20 1 PTR 288
I
Rata-rata 42.50 44.11 44.1 1 40.68Angka-angka yang diikuti oleh huruf yang sama tidak berbeda nyata pada taraf 0.05 Rata-rata Dosis dolomit (ton ha-')
Interaksi antara dolomit dan genotipe nyata mempengaruhi jumlah cabang 0 35.63 39.13 58.56 41.26 38.30
(Lampiran 3). Jumlah cabang terbanyak terdapat pada PTR293 dengan dosis 4 ton
ha-' dan paling sedikit pada PTR288 tanpa dolomit (Lampiran 6 Gambar 4). Pada 2 41.20 41.70 58.83 44.76 39.10
Gambar 4 terlihat bahwa jumlah cabang semua genotipe pola interaksinya hampir
sama terhadap dosis dolomit, cenderung meningkat dengan peningkatan dosis 4 39.43 44.73 53.83 43.23 41.70
dolomit 2 hingga 4 ton, dan menurun dengan penambahan 6 ton ha-'. 6
Gambar 4. Hubungan dolomit dengan jumlah cabang beberapa genotipe kede berumur dalam.
0.0
-1
0 2 4 6
DrmsDolomt (ton/ ha)
+
PTR6 B PTR178 -k-PTRIEO X P T m l X PTFa88 b PTFa93 -Lin%~(PTffi) -Uw(PTR178) -Lmesr(PTRlSO) -Lmer(PTRZOl) -Uner(PTRZffl) -Lln%U(PTR293)Dolomit, genotipe, dan interaksinya nyata mempengaruhi umur berbunga dan
umur panen (Lampitan 5). Pada Gambar 5 dan 6 terlihat bahwa baik umur berbunga
maupun umur panen pola interaksinya linear dengan nilai konstan setiap peningkatan
dosis dolomit. Pada Gambar 5 dan 6 juga terlihat bahwa hampir semua genotipe
1
0 2 4 6 [image:114.595.83.515.37.771.2]1
Cosls Ddmit (ton / ha)Gambar 6. Hubungan dolomit dengan umur panen beberapa genotipe kedelai berumur dalam.
mempunyai umur berbunga dan umur panen berbeda kecuali PTR178, PTR201, dan
PTR288 pada semua tingkat dolomit, umur berbunganya sama tetapi umur panennya
sedikit berbeda. Umur panen PTR6 sedikit meningkat dengan meningkatnya dosis
dolomit.
Komponen Hasil
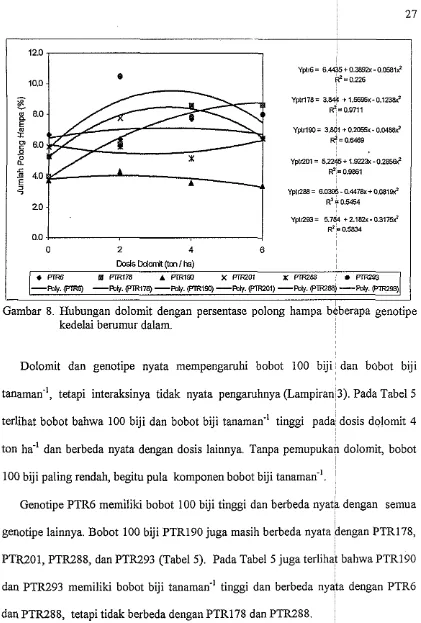
Dololllit tidak nyata mempengaruhi persentase polong hampa, tetapi genotipe dan
interaksinya nyata mempengaruhi ju~nlah polong berisi dan persentase polong han~pa
26 polong berisi yang lebih banyak (108.1 buah) pada dosis dolomit 4 ton ha*' dan
99.8 buah pada dosis 2 ton ha.' dan berbeda nyata dengan semua perlakuan lainnya
terutama PTR6 yang paling sedikit yakni 46.3 buah tanpa dolomit. Pada Gambar 7
juga terlihat bahwa jumlah polong berisi semua genotipe polanya hampir sama yaitu
terjadi peningkatan pada dosis 2 hingga 4 ton dan semakin menurun pada dosis
dolomit 6 ton ha.'.
[image:115.595.81.519.156.749.2]YplQOl= 50.041 + 17.363~-2.7231*
Gambar 7. Hubungan dolomit dengan jumlah polong berisi beberapagenotipe kedelai berumur dalam.
Genotipe PTR293 memiliki jumlah polong berisi yang tertinggi, tetapi memiliki
persentase polong hampa tertinggi pula, terutama pada dosis dolomit 2 ton ha.'
(10.4%). Persentase polong hampa terendah dimiliki oleh PTR190 yakni 3.3%, 3.5%,
dan 3.7% masing-masing pada dosis dolomit secara berurutan 6 ton, 4 ton ha-', dan
R2 = 0 5454
Yp10?3= 5.784 t 2.18h- 031759 $ = o m
0.0
20 0
0
2 4 6Dmis Ddwnt (ton / k)
PIR6 f# FIR178 A FIR153 X m 0 1 X m e a
*
m-POW. WW -W. eTR178) -Pot$ eTR153) -pohl. (F'lRZo1) -mty. (~1~289) -pohl. ~ 9 3 )
Gambar 8. Hubungan dolomit dengan persentase polong hampa beberapa genotipe kedelai berumur dalam.
Dolomit dan genotipe nyata mempengaruhi bobot 100 biji dan bobot biji
tanaman-', tetapi interaksinya tidak nyata pengaruhnya (Lampiran 3). Pada Tabel 5
terlihat bobot bahwa 100 biji dan bobot biji tanaman-' tinggi pada dosis dolomit 4
ton ha-' dan berbeda nyata dengan dosis lainnya. Tanpa pemupukan dolomit, bobot
100 biji paling rendah, begitu pula kornponen bobot biji tanaman-'.
Genotipe PTR6 memiliki bobot 100 biji tinggi dan berbeda nyata dengan semua
genotipe lainnya. Bobot 100 biji PTR190 juga masih berbeda nyata dengan PTR178,
PTR201, PTR288, dan PTR293 (Tabel 5). Pada Tabel 5 juga terlihat bahwa PTR190
dan PTR293 memiliki bobot biji tanaman-' tinggi dan berbeda nyata dengan PTR6
[image:116.599.101.522.55.678.2]Tabel 5. Pengaruh dolomit dan genotipe terhadap bobot 100 biji dan bobot biji tanaman-'.
DMKT.
Hasil Biji
Genotipe nyata mempengaruhi bobot biji petak-' dan hasil biji ha*', tetapi dolomit
dan interaksi~lya tidak nyata mempengaruhi kedua peubah ini (Lampiran 4).
Pada Tabel 6 terlihat bahwa bobot biji petak-' dan hasil biji tertinggi pada PTR190
yakni masing-masing 1139.92 g atau 2.849 ton ha-', bobot dan hasilnya sama dengan
PTR6 (yakni 1016.30 g atau 2.540 ton) serta PTR293 (yakni 1017,42 g atau 2.543 ton
ha-'). Bobot biji dan hasil biji terendah terdapat pada PTR178 yakni 860.50 g p e t a ~ '
atau 2.151 ton ha-'. Tabel 6 juga terlihat bahwa bobot biji petak-' PTR178 tidak
berbeda nyata dengan PTR201 dan PTR288 dengan nilai masing-masing 922.75 g
[image:117.595.87.526.69.592.2]PEMBAHASAN
Hasil percobaan ini inenunjukkan bahwa kandungan kalsiu
tajuk telah cukup untuk pertuinbuhan normal tanainan tanpa
(Lampiran 5 dan Gambar 1, 2, dail 3). Hal ini tidak terlepas
percobaan masih cukup subur untuk peituinbuhan tanaman kede
tidak terlalu inasam kandungan Ca dan Mg tersedia inasih dal
serta unsur-unsur beracun seperti A1 dalain jumlah kecil b
berdasarkan hasil analisis tanah awal ( Lanlpiran 7).
Dalam kondisi BJA kesuburan tanah semakin meningkat
ineningkatnya pH tanah dan lebih tingginya P dail I<
dibandingkan dengan pada BTH (Ghulamahdi, 1999). Ono (1991) inenyatalcall bahwa
tanah yang digenangi hampir tidak dipengaruhi oleh tingkat p
halnya tanah tanpa penggenangan. Tanah yang digenangi da
perlakuail pengapuran ternyata pH-nya juga ineili~lgkat dari p
6.44, sebaliknya tanpa penggenangan pH bahkan inenurun dari
5.3. Penggenangan dengan pengapuran dapat inendorong berkurangnya juinlah ion-
ion hidrogen dan aluminium yang dapat dipertukarkan sehingga pH
Dolomit dan genotipe serta interaksi~lya tidak nyata mempen
dan Mg tajuk, tetapi dolomit nyata mempengarul~i kadar
(Tabel 2). Genotipe tidak nyata meinpengaruhi kadar Ca taj
dolomit dengan genotipe nyata ineinpengaruhi kadar Ca taj
31 ha-' 0.26%, paling tinggi dan berbeda nyata dengaii lainnya. Disustll dengan
menurtmya kadar Mg tajuk dengan penlambahan dosis dolomit.
Gambar 1 menunjulkan ballwa Ca tajuk setiap genotipe bervariasi menurtit dosis
dolomit. Pada genotipe PTR6, PTR190, dan PTR201 berpola hampir sazna yaitu
kadar Ca tajuk meningkat pada dosis 2 hingga 4 ton dan lnenurun pada dosis dolomit
hingga 6 ton ha-'. Pada PTR178 dan PTR288, kadar Ca tajuk lnenurun dengan
penlupukan 2 ton dolomit dan meningkat pada dosis 4 hingga 6 ton ha-'. Pada
genotipe PTR293 kadar Ca semakin menurun dengan penamballan dosis doloinit.
Meskipun pola kadar Ca tajulc genotipe beirvariasi pada setiap dosis dolomit, pola
serapan hara baik Ca maupun Mg tajuk hampir sama yaitu meningkat dengan
pe~nupukan 2 hingga 4 ton dan cenderung inenuirun pada dosis 6 ton doloinit ton
ha-' (Gambar 2 dan 3). Pada Gambar 1 juga meinperlihatkan ballwa PTR6 kadar Ca
tajuknya tinggi yakni 1.33% pada dosis dolomit 2 ton, dan rendah pada PTR293
(0.85%) pada dosis dolomit 6 ton ha.'. Meskipun berbeda dengan hasil penelitian
Ghulamahdi (1999), yang rendah kadar Ca dan Mg-nya pada B
dan Mg 0.23%. Berdasarkan pengamatan pada BTH, Ca menc
0.32% dengan pemupukan kalsit 2 ton ha-' dan pengamatan pad
Mg hasil penelitian ini diduga sama dengan penga~natan pada
Mg banyak diperlukan di bagian bawah untuk pertumbuhan a
1999), juga diduga diperlukan untuk pe~nbentukan batang, dan
polong dan pengisian biji. Hal ini dapat dibuktikan pada Ga
yang lebih cepat berbunga kadar Ca tajuknya lebih tingg
32 miho dan Ca selama tahap reproduksi sering tinggi jika dibandingkan dengan selalna
tahap vegetatif. Di bawah cekaman air kalsium tidak ditranslokasikan dari polong ke
daun selama pengisian polong (Sorooshcades et al. 1995).
Rendalmya kadar Ca dan Mg daun pada BJA seperti yang dilaporkan Ghula~nahdi
(1999), juga dilaporkan ole11 Seong et al. (1999). Para peneliti ittipun mendapatlcan
kandungan Ca dan Mg daun yang rendah pada pet~ggenangan dibandinglcan dengall
pada BTH tetapi kandumgan hara Fe dall Mg daun tinggi. Meskipun demikian
pertunbuhan tanaman terutama daun kelihatan normal (tidak terlihat gejala
kekurangan Ca dan Mg). Ini menunjukkan baliwa Ca dan Mg dam yang rendah pada
BJA cukup ultuk pertumbuhan normal tanaman terutama pada daun. Sopandie
(1997) lnenyatakan bahwa kadar Ca dala~n tanaman berkisar antara 0.2 sa~npai
beberapa persen (berdasarkan bobot kering), ballkan mungkin jumlahnya lebih tinggi
dari pada kebutuhan minimum. Lebih lanjut dinyatakan bahwa banyak jenis tanaman
yang dapat tumbuh normal dalam larutan dengall konsentrasi Ca 2 ppm atau 0.05
ppm seperti pada tanaman tell dan tenlbakau, dengan ketentuan ballwa kadar ion-ion
divalen lai~uiya rendah. Rumawas menyebutkan kadar Ca 0.16% cukup untuk
pertumbuhan normal tanaman kedelai (komunikasi pribadi, 2002). Hal lain yang
mungkin juga mendukung pertumbuhan normal adalah adanya unsur Fe dan Mn daun
yang tinggi pada BJA, yang diduga menggantikan fungsi Ca dan Mg pada daul.
Sopandie (1997) dan Salisbury dan Ross (1992) menyataka~l bahwa kahat Fe
dicirikan dengan meluasnya klorosis yang jelas di daerah antara urat-urat daun yang
mirip gejala kahat Mg. Jones (1998) menyatakan baliwa Ca bersaing dan
33 0.075 nm, berada di antara Mg (0.065 nm) dan Ca (0.099 nm) sehingga dapat saling
bertukar atau bersaing pada bermacam-macam reaksi, dalarn beberapa ha1 Mg dapat
digantikan oleh Mn atau sebaliknya dan kaliat Mn dapat znenyebabkan defisiensi Fe,
Mg, terutama Ca.
Serapan l~ara yang tinggi menyebabkan kandungan hara yang tinggi pula dan
berpengaruh pada bobot kering akar. Pada percobaan ini genotipe PTR190 bobot
kering akamya tinggi (0.93 g tanaman-') da~i rendah pada PTR6 (0.77 g) meskipun
secara statistik nilainya sama. Pada penelitia~i Jafri (2002) bobot kering akar tinggi
pada PTR293 (0.57 g tanaman-') dan rendah pada P'TR288 (0.36 g).
Bobot kering akar yang tinggi pada BJA tidak terlepas dari pertumbuhan
perakaran yang terus-.lnene~us (karena terjadi akumulasi hasil fotosintesis kebagian
bawah tanaman pada tahap aklimatisasi) (Troedson et 01. 1983). Tidak terjadinya
penuaan akar me~lyebabkan pe~nbentukan bintil akar terus-menerus yang selanjutnya
berpengaruh pada peni~igkatan bobot bintil aka]. seliingga ~neningkatkan nitrogenase,
peningkatan serapan hara akar la111 lneningkatkan serapan 11ara daun (Gl~ulnmahdi
1999). Pada percobaan ini bobot kering bintil akar tanaman" tinggi pada genotipe
PTR190 (0.35 g) hrunpir sania yang dilaporkan Ghulamahdi (1999) pada PTR322
yakni 0.34 g pada BJA, dilakukan penga~natan pada u~nur 8 MST, dan bobot kering
bintil akar yang rendah terdapat pada genotipe PTR293 yakni 0.1 5 g. Sementara Jafri
(2002) melaporkan bahwa pada BTH, bobot keri~lg bintil akar te~tinggi juga pada
PTR190 (59.59 mg) dan terendal~ pada PTR178 (40.00 n~g). Namun bobot kering
bintil akar percobaan Jafri (2002) masill Iebih rendah dengan sisteni BTI-I
34 karena pada BJA akar tumbuh terus-menerus dan tidak terjadi penuaan altar dan bintil
akar dibandingkan dengan BTH, disanlping itu kondisi lahan percobaan ~nasih cukup
mengandung unsur hara Ca dan Mg se~ta adanya tambahall dolomit yang sangat
dibutuhkan untuk pertulnbuhan bintil akar dan tanaman. Menu~ut Yutono (1983) dan
Sul~artatik et al. (1987), selain menaikkan pH pemberian kapur juga berpengaruh
terhadap peitumbul~an tananlan leguninosa dan bintil akar. Kalsiuln diperlukan oleh
tanaman dan bakteri rhizobium dalaln julnlah yang besar untuk proses infeksi
rhizobium pada akar tanalnau dan pembentukan bintil akar selanjutnya.
Meningkatnya bobot kering bintil akar berpengaruh pada peningkatan bobot
kering tajuk, tinggi tanaman, dan jumlah cabang. Bobot kering tajuk teltinggi pada
pada PTR201 (9.66 g) yang nilainya salna dengan PTR6, PTR178, dan PTR190.
Bobot kering tajuk terendah pada PTR 288 (7.63 g) dan PTR 293 (7.31 g) (Tabel 3).
Tinggi tanaman tertinggi pada PTR19O (54.76 cm) dall berbeda nyata dengan
genotipe lailmya, terendah pada PTR6 (38.38 cln) (Tabel 4). Jafii (2002) pada
penelitian BTH melaporkan genotipe te~tinggi juga pada PTR190 dan terendah pada
PTR6. Hal ini dapat dijadikan indikasi bahwa tinggi tanaman dipengaruhi oleh faltor
genetik tanaman (Dewi et
al.
1993). Zahara et 01. (1994) menyatakan bahwa tinggitanaman merupakan kriteria yang cukup penting karena secara tidak langsung dapat
meningkatkan hasil kedelai melalui jumlah polong berisi.
Tinggi tanaman tidak sejalan dengan julnlah cabang. Jumlah cabang terbanyak
pada PTR293 pada dosis dolomit 4 ton ha7' (4.8 buah) dan paling rendah pada
PTR288 tanpa dolomit (2.3 buah) dan dolomit 6 ton ha*' (2.3 buah) (Lampiran 5 dan
35 pola interalcsinya ha~npir sama terhadap dosis dolomit, cenderung meningkat dengan
peningkatan dosis dolomit 2 hingga 4 ton dan menurun pada penambahan 6 ton ha-'.
Hasil penelitian Jafri (2002) pada BTH juga dilaporkan, baliwa jumlah cabang
tertinggi pada PTR293 (3.6 buah) dan rendah pada PTR288 (2.5 buah). Menurut
Adam dan Weber (1998), jumlah cabang lebih banyak dipengaruhi oleh faktor
genetik. Pada kondisi lingkungan yang menguntungkan kedelai dapat menghasilkan
cabang dan buku lebih banyak.
Dolomit, genotipe dan interaksinya nyata mempengaruhi umur berbunga dan
umur panen (Lampiran 2 Gambar 5 dan 6). Pada Gambar 5 dan 6 memperlihatkan
bahwa baik umur berbunga maupun umur panen genotipe yang dicobakan cenderung
linier dengan nilai konstan setiap peningkatan dosis dolomit. Menurut Sumarno dan
Hartono (1983), lunur berbunga dan umur panen kedelai beragam tergantung varietas
yang ditanam. Di Indonesia umur panen kedelai dibedakan atas 3 golongan yaitu
berumur genjah (75-85 hari), berumur sedang (86-90 hari), dan berumur dalam (> 95
hari). Umur berbunga terkait dengan umur panen, semakii cepat berbunga semakin
cepat umur panen, begitu pula sebaliknya. Genotipe PTR6 umur berbunganya lebih
cepat (38.2 hari), umur panen 93.0 hari. Umur berbunga lebih lama pada PTR293
(49.0 hari), umur panen 106.0 hari. Jafri (2002) juga melaporkan bahwa pada BTH
umur berbunga tercepat dijumpai pada PTR6 (37.0 hari), umur panennya (87.3 hari),
dan terlama pada PTR293 (47.0 hari), umur panen (101.3 hari). Umur berbunga dan
umur panen pada BJA tidak jauh berbeda dengan BTH. Menurut Arsyad (1988)
36 diiendalikan oleh gen secara poligenik culup mantap dan stabil pada lingk~mgan
yang berbeda.
Komponen hasil nyata dipengaruhi ole11 genotipe dan dolomit, kecuali persentase
p ~ l o n g hamnpa, dolomit tidak nyata menlpengaruhi (Lampiran 3). Interaksi kedua
perlakuan juga nyata inempengaruhi jumlah polong berisi maupun persentase polong
hampa (Lampiran 3 Gambar 7 dan 8). Genotipe PTR293 menghasilkan polong berisi
yang lebih banyak (108.1 buah) pada dosis dolomit 4 ton ha" dan 99.8 buah pada
dosis dololllit 2 ton ha*', lebih sedikit pada PTR6 (46.3 buah) tanpa dolomit. Jafri
(2002) melaporkan, bahwa jumlah polong berisi tinggi pada PTR293 (71.2 buah) dan
rendah pada PTR6 (39.8 buah). Ganlbar 6 juga memperlihatkan ballwa juinlali polong
berisi semua genotipe polanya hampir sama yaitu terjadi peningkatan pada dosis
dolomit 2 hingga 4 ton ha-' dan semakin menurun dengan penambahan dosis dolomit.
Tigginya jumlah polong berisi pada PTR293 disebabkan oleh pertumnbd~an
(perkembangan akar, tajuk, dan cabang) yang lebih baik, disamping itu faktor genetik
tanaman, terutama umur yang panjang sehingga proses pembentukan dan pengisian
polong lebih lama dan didukung kondisi lahan jenuh air menyebabkan daun lambat
mengalami keguguran (senecence) sehingga proses fotosintesis berlangstmg lebih
lama dan menghasilkan asimilat yang lebih banyak. Nugraha (1994) menyatakan
bahwa tanaman kedelai berkorelasi positif dengan urnur. Dengan umur yang lebih
dalam hasil fotosintesis dapat dirnanfaatkan untuk pembentukan biji dan pengisian
polong. Board et al. (1999) menyatakan bahwa masa reproduktif yang lama dapat
37 Jumlah polong berisi yang tinggi pada PTR293 ternyata juga diilcuti ole11
persentase polong hampa yang tinggi pula, terutama pada dosis dolo~nit 2 ton 11a-l
(10.4%). Persentase polong hampa yang rendah dimiliki oleh PTR190 (3.3%) pada
dosis 6 ton ha-' (Lampiran 6, Gambar 5 dan 6). Jafri (2002) juga inelaporlcan bahwa
genotipe PTR393 memiliki persentase polong hainpa tinggi pula pada BTH. Ini
nlengindikasikan bahwa tingginya persentase polong hampa PTR293 disebabkan oleh
faktor genetik tanaman.
Dolomit dan genotipe nyata meinpengaruhi bobot 100 biji dan bobot biji
tanaman-', tetapi tidak nyata inempengaruhi bobot biji dan hasil biji ha-'
(Lanlpiran 3). Tabel 4 inemperlihatkan ballwa dosis dolomit 4 ton ha-' meninglcatkan
bobot 100 biji dan bobot biji tanaman' dan berbeda nyata deilgan yang lainnya. Pada
Tabel 5 juga terlihat bahwa Genotipe PTR190 dan PTR293 meiniliki bobot biji
tansunan-' yang tinggi yakni masing-masing 15.42 g dan 15.08 g. Bobot biji tanaman-'
yang rendah terdapat pada PTR6 dan PTR288, masing-masing 13.37 g dan 12.5 g.
Bobot 100 biji yang tinggi pada PTR6 (11.9 g) berbeda nyata dengan genotipe
lainnya, terutama PTR178 dan PTR293, masing-masing 9.80 g dan 9.74 g (Tabel 5).
Jafri (2002) pada BTH inelaporkan bobot biji tanaman-' juga tinggi pada PTR293
dan bobot 100 biji pada PTR6, masing-masing adalah 11.82 g tanaman-' dan
10.21 g. Bobot biji tanaman" PTR190 dan PTR293, dan bobot 100 biji PTR6 yang
tinggi tidalc terlepas dari tanggap yang tinggi genotipe-genotipe tersebut terhadap
lulgkungan tumbuhnya disamping faktor genetik. Arsyad dan Tanj~mg (2000)
ineyatakan ballwa hasil kedelai dipengarithi sangat nyata ole11 faktor genetik, kondisi
38 IComponen hasil yang tinggi berpengaluh pada peningkatan bobot biji petal<-' dan
akhirnya menghasilkan biji ha-' yang tinggi pula. Bobot biji petal<-' tinggi pada
PTiIl90 (1139.92 g) nilainya tidak berbeda dengan PTR293 dan PTR6, masing-
masing 1017.42 g dan 1016.33 g. Bobot biji petak' juga agak ren
PTR201, dan PTR288 masing-masing 860.50 g, 992.75 g, dan 9
Tabel 6 juga memperlihatkan hasil biji ha-' tinggi pada PTR190
tidak berbeda dengan PTR293 dan PTR6 masing-masing 2.543
Hasil biji ha-' juga agak rendah pada PTR178, PTR201, dan PTR288 masing-masing
2.306 ton, 2.420 ton, dan 2.151 ton. Penelitian Jafri (2002) pada BTH mendapatkan
hasil bij