POTENSI RIZOBAKTERI UNTUK PENGHAMBATAN
PENYEBAB BLAS PADI (Pyricularia oryzae Cav.)
RIAN ANDINI
DEPARTEMEN PROTEKSI TANAMAN FAKULTAS PERTANIAN
INSTITUT PERTANIAN BOGOR BOGOR
PERNYATAAN MENGENAI SKRIPSI DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa skripsi berjudul Potensi Rizobakteri untuk Penghambatan Penyebab Blas Padi (Pyricularia oryzae Cav.) adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir skripsi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Januari 2015
ABSTRAK
RIAN ANDINI. Potensi Rizobakteri untuk Penghambatan Penyebab Blas Padi (Pyricularia oryzae Cav.). Dibimbing oleh MEITY SURADJI SINAGA.
ABSTRACT
RIAN ANDINI. Rhizobacteria Potential for Inhibiting The Growth of Rice Blast Pathogen (Pyricularia oryzae Cav.). Supervised by MEITY SURADJI SINAGA.
Pyricularia oryzae Cav. is the causal agent of the blast disease on upland rice cultivation. Late 1980, P.oryzae also infected the irrigated rice and cause 10%-30% field loss. Disease biocontrol using bacteria has been developed. Rhizobacteria are the bacteria live around the roots of plants and have the symbiotic relationship with the plants. Non-pathogenic rhizobacteria can reduce inoculum of pathogen. Objective of this study is to evaluate potential uses of rhizobacteria to control the growth of P. oryzae. Rhizobacteria had been isolated from the irrigated rice cultivation soil at 4 villages around IPB Dramaga: Babakan Lebak (Blk), Balumbang Jaya (Blb), Cikarawang (Ckr), dan Situgede (Stgd). Rhizobacteria which had been isolated were 33 isolates. After hypersensitive test, has been known 17 from 33 rizhobacteria isolates are non plant pathogenic. Among non plant pathogenic rhizobacteria, 2 isolates are Gram negative and 15 isolates are Gram positive. The antagonism test has shown that Blk 3.6, Blk 5.3, Blb 3.11, dan Ckr 3.2 isolates have significantly suppressed growth of P.oryzae. Isolate Blk 3.6 was the most effective in suppressing growth of P.oryzae (91.98%).
© Hak Cipta Milik IPB, tahun 2015 Hak Cipta dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan yang wajar IPB.
2
POTENSI RIZOBAKTERI UNTUK PENGHAMBATAN
PENYEBAB BLAS PADI (Pyricularia oryzae Cav.)
RIAN ANDINI
Skripsi
sebagai salah satu syarat untuk memperoleh gelar Sarjana Pertanian
pada
Departemen Proteksi Tanaman
DEPARTEMEN PROTEKSI TANAMAN FAKULTAS PERTANIAN
INSTITUT PERTANIAN BOGOR BOGOR
Judul Skripsi : Potensi Rizobakteri dalam Penghambatan Penyebab Blas Padi (Pyricularia oryzae Cav.)
Nama : Rian Andini
NIM : A34100019
Disetujui oleh
Prof.Dr. Ir. Meity Suradji Sinaga, M.Sc. Dosen Pembimbing
Diketahui oleh
Dr.Ir.Abdjad Asih Nawangsih, M.Si. Ketua Departemen
iii
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah subhanahu wa ta’ala atas segala karunia-Nya sehingga karya ilmiah ini berhasil diselesaikan. Tugas akhir ini dilaksanakan sejak bulan Februari 2014 sampai Oktober 2014 dengan topik eksplorasi rizobakteri untuk mengevalusi potensinya sebagai pengendali blas padi. Terima kasih penulis ucapkan kepada Ibu Prof. Dr. Ir. Meity Suradji Sinaga, M.Sc selaku pembimbing dan Bapak Dr.Ir.Yayi Munara Kusumah, M.Si selaku dosen penguji yang banyak memberi saran untuk penulisan skripsi. Ucapan terima kasih juga disampaikan kepada Dr.Ir.Abdul Muin Adnan, M.S sebagai dosen pembimbing akademik untuk dukungan moral selama penelitian.
Terima kasih juga penulis ucapkan kepada para petani di sekitar lingkungan kampus IPB Dramaga yang sudah mengizinkan kegiatan sampling di lahan mereka; Bapak Dadang Surachman atas bantuan teknis di dalam kegiatan laboratorium; teman-teman lorong 7 asrama A1; teman-teman TPB B23-B24; teman-teman wisma Padasuka untuk dukungan dan kerjasamanya; teman-teman Laboratorium Mikologi Tumbuhan yang selalu memberikan bantuan dan keceriaan.
Terima kasih juga penulis ucapkan kepada orang tua, adik, seluruh keluarga yang tidak pernah lelah memberikan semangat dan doa, serta seluruh dosen, tenaga kependidikan Departemen Proteksi Tanaman, rekan-rekan Proteksi Tanaman angkatan 2010, dan seluruh rekan-rekan yang membantu kelancaran studi program pendidikan S1. Semoga penelitian ini bermanfaat.
v
Isolasi dan Identifikasi Penyebab Blas 4
Eksplorasi Rizobakteri 4
Uji Non Patogenik IsolatRizobakteri 5
Identifikasi Rizobakteri 5
Uji Antagonisme antara Rizobakteri dan P.oryzae 5
Analisis Data 5
HASIL DAN PEMBAHASAN 6
Isolasi dan Identifikasi Penyebab Blas 6
Eksplorasi Rizobakteri 7
Mekanisme Antagonisme Rizobakteriterhadap P.oryzae 13
vii
DAFTAR TABEL
1 Hasil eksplorasi rizobakteri dan uji hipersensitif dari Desa Balumbang Jaya (Blb), Situgede (Stgd), Cikarawang (Ckr), dan Babakan Lebak
(Blk). 8
2 Hasil pengamatan ciri koloni pada isolat rizobakteri yang tidak menunjukkan gejala nekrotik pada uji hipersensitif 9 3 Hasil uji Gram dan uji antagonisme rizobakteri terhadap P.oryzae 13
DAFTAR GAMBAR
1 Gejala blas pada permukaan daun (a,b) 6
2 Inokulasi isolat P.oryzae pada daun padi (a),gejala blas pada daun
setelah diinokulasi (b,c) 6
3 Mikroskopis konidia P.oryzae dengan perbesaran 40x10 7 4 Hasil uji hipersensitif pada daun tembakau; isolat rizobakteri yang
menimbulkan gejala nekrotik pada daun tembakau (a), isolat rizobakteri yang tidak menimbulkan gejala nekrotik pada daun
tembakau (b) 8
5 Hasil uji Gram pada isolat rizobakteri; isolat bakteri Gram negatif (a), hasil uji Gram pada isolat bakteri Gram positif (b) 14 6 Zona bening yang terbentuk saat uji antagonisme antara rizobakteri
dan P.oryzae 14
7 Hasil uji antagonisme antara rizobakteri dan P.oryzae; perlakuan rizobakteri yang tidak membentuk zona bening (a), perlakuan kontrol
(b) 15
DAFTAR LAMPIRAN
1 Pertumbuhan diameter P.oryzae pada uji antagonisme dengan
PENDAHULUAN
Latar Belakang
Padi merupakan sumber utama makanan pokok di Indonesia. Produksi padi di dunia menempati urutan ketiga dari semua serealia setelah jagung dan gandum (Kharisma et al. 2013). Penurunan hasil padi disebabkan banyak faktor seperti iklim, ketersediaan air, kesuburan tanah, varietas yang digunakan, sistem pengelolaan tanaman, perkembangan hama dan penyakit. Penyakit blas adalah salah satu penyakit penting pada padi yang terjadi hampir di seluruh belahan dunia pada daerah yang memiliki curah hujan tinggi dan menggunakan pupuk nitrogen secara berlebihan (Agrios 2005).
Pyricularia oryzae Cav. merupakan cendawan penyebab penyakit blas yang menyerang padi gogo. Pada akhir 1980, dilaporkan bahwa cendawan ini juga menyerang padi sawah (Puslitbang Tanaman Pangan 2007a). Luas serangan penyakit blas di Indonesia dapat mencapai 1.285 juta ha atau sekitar 12% dari total luas areal pertanaman padi di Indonesia sedangkan konsumsi beras pada tahun 2010, 2015, dan 2020 diproyeksikan berturut-turut sebesar 32.13, 34.12, dan 35.97 juta ton. Jumlah penduduk pada ketiga periode itu diperkirakan berturut-turut 235, 249, dan 263 juta jiwa (Puslitbang Tanaman Pangan 2007b). Epidemi penyakit ini dapat disebabkan oleh nitrogen yang berlebihan, tingginya adaptasi cendawan ini dalam menghadapi perubahan-perubahan lingkungan hidupnya, penggunaan fungisida, penggunaan varietas tahan secara monokultur dan terus menerus. Cendawan inidapat menjadi patogen pada beberapa tanaman penting lainnya seperti gandum, sorgum, serealia lainnya, dan gulma rumput-rumputan serta gulma lainnya (Santoso dan Nasution 2009).
Cendawan P.oryzae dapat menyerang daun (leaf blast), buku (node blast), leher malai (neck blast), bulir padi (spikelet blast) dan daun pelepah (collar rot) (Scardaci et al. 1997). Penyakit blas dapat menimbulkan dua gejala khas yakni bercak coklat kehitaman berbentuk belah ketupat, dengan pusat bercak berwarna putih pada blas daun dan pada blas leher berupa bercak coklat kehitaman pada pangkal leher yang dapat mengakibatkan leher malai tidak mampu menopang malai dan patah (Puslitbang Tanaman Pangan 2007a). Bentuk dan ukuran bercak dipengaruhi oleh perbedaan kultivar dan kondisi lingkungan. Keadaan yang lembap dan varietas inang yang rentan dapat mengkondisikan bercak terus membesar.
P.oryzae adalah cendawan yang termasuk dalam kelompok Deuteromycetes,
ordo Moniliales, famili Moniliaceae (Barnett dan Hunter 1998). Cendawan ini ditemukan di alam dalam bentuk aseksualnya saja sedangkan bentuk seksualnya yaitu Magnaporthe grisea hanya dihasilkan dengan pengkulturan di laboratorium (Agrios 2005). Secara morfologi, cendawan P.oryzae mempunyai konidia berbentuk seperti buah pir dan bersekat dua (Barnett dan Hunter 1998).
2
kondisi kelembapan dan suhu yang mendukung, P.oryzae mengalami banyak daur penyakit dalam satu musim dan menghasilkan spora yang melimpah pada akhir musim. Tingkat inokulum yang tinggi sangat berbahaya bagi tanaman padi yang rentan (Scardaci et al. 1997).
Daur penyakit blas meliputi fase infeksi, kolonisasi, dan sporulasi. Fase infeksi diawali dengan pembentukan konidia bersepta tiga yang dilepaskan dari konidiafora. Konidia berpindah ke permukaan daun melalui percikan air atau angin. Konidia kontak pada daun karena adanya perekat atau getah di ujungnya (Hamer 1988). Konidia akan berkecambah pada kondisi optimum dengan cara membentuk miselium yang selanjutnya akan membentuk apresoria dan kapak penetrasi. Kapak penetrasiakan menembus kutikula daun dengan bantuan melanin yang ada pada dinding apresoria. Hifa yang terus berkembang dan menginfeksi akan menimbulkan bercak sebagai tanda sel dan jaringan mengalami nekrosis. Pada kelembapan yang tinggi, bercak pada tanaman yang rentan menghasilkan konidia selama 3-4 hari. Konidia ini sangat mudah tersebar dan merupakan inokulum untuk infeksi selanjutnya (Agrios 2005).
Bercak pertama akan muncul 4-5 hari setelah inokulasi pada suhu 26-28 °C. Perkembangan dari bercak kecil menjadi bercak besar akan berlangsung cepat pada suhu 32 °C selama 8 hari. Sporulasi berlangsung optimum pada suhu 28 °C, kelembapan 95%, dan kondisi gelap selama 15 jam. Sporulasi tidak terjadi jika kelembapan kurang dari 89%. Suhu optimum untuk perkecambahan spora, pembentukan bercak dan sporulasi adalah 32-35 °C (Scardaci et al. 1997).
Cendawan P.oryzae memiliki keragaman genetik yang tinggi. Populasi
P.oryzae terdiri dari individu-individu ras yang mempunyai sifat virulensi yang berbeda. Ras P.oryzae dapat berubah dan membentuk ras baru dengan cepat apabila populasi tanaman atau sifat ketahanan tanaman berubah. Tingginya tingkat keragaman pada patogen P.oryzae disebabkan kemampuannya melakukan perkawinan antar-haploid hifa yang berlainan materi genetiknya (parasexual
exchanged DNA) sehingga memungkinkan terjadinya segregasi dan rekombinasi
antar ras (Zeigler et al. 1997). Reflinur et al. (2005) melaporkan bahwa amplifikasi DNA 230 isolat cendawan dari 5 lokasi yaitu Lampung, Sukabumi, Sumatera Utara, Sumatera Barat, dan Bogor dengan menggunakan primer spesifik gen virulensi, menghasilkan 8 haplotip.
3 agens hayati yang dikembangkan adalah mikroba alami, baik yang hidup sebagai saprofit di dalam tanah, air, bahan organik, maupun jaringan tanaman. Salah satu agens hayati yang melimpah paling banyak di tanah adalah bakteri. Rizobakteri adalah bakteri yang berkolonisasi di daerah rizosfer dan memberikan hubungan simbiotik terhadap tanaman dan lingkunganya (Maheshwari dan Saraf 2013).
Interaksi mikroba tanaman di daerah perakaran tanaman menjadi penentu atas kesehatan tanaman dan kesuburan tanah. Interaksi ini memainkan peranan penting dalam transformasi, mobilisasi sumber hara di tanah dan penyerapan nutrisi tanaman esensial untuk memaksimalkan pertumbuhan tanaman (Fatima et al. 2009). Menurut Glick dan Pasternak (2003), rizobakteri yang menguntungkan seperti Plant Growth Promoting Rhizobacteria (PGPR) dapat mempengaruhi pertumbuhan tumbuhan dengan 2 cara yaitu secara langsung dan tidak langsung. Pengaruh secara langsung pada tumbuhan yaitu menyediakan mineral-mineral yang sudah disintesis dalam bentuk yang dapat digunakan oleh tanaman seperti mengikat nitrogen, mensistesis hormon tanaman, dan membantu tanaman dalam pengambilan nutrisi di alam. Pengaruh PGPR secara tidak langsung yaitu mencegah perkembangan organisme yang bersifat patogen pada tanaman dengan mekanisme antagonisme, predasi, dan parasitisme. Antagonisme adalah salah satu mekanisme biokontrol PGPR, di mana bakteri dapat menghasilkan antibiotik, misalnya Bacillus cereus. Antibiotik yang dihasilkan dan telah digunakan secara luas dalam pengendalian penyakit tanaman yaitu streptomisin, tetrasiklin, sikloheksamid, dan blastisidin. Mekanisme biokontrol predasi dan parasitisme biasanya ditemukan pada cendawan, seperti Trichoderma yang dapat merusak dinding sel cendawan lainya (Lugtenberg dan Kamilova 2009).
Bakteri yang sudah sering dipakai sebagai pengendalian adalah
Pseudomonas fluorescens. Pseudomonas fluorescens sering dipakai sebagai
perlakuan benih, tanah, dan foliar untuk pengendalian blas. Pseudomonas
fluorescens mengendalikan patogen tanaman dengan kompetisi nutrisi, antibiosis,
produksi sideropor, dan enzim litik (Shymala dan Sivakumaar 2012). Pengendalian hayati penyakit blas pada padi sawah masih belum banyak dilakukan. Pengendalian hayati yang paling memungkinkan untuk padi sawah adalah mengembangkan agens hayati bakteri atau aktinomiset.
Tujuan Penelitian
Penelitian ini bertujuan untuk mengeksplorasi bakteri dari rizosfer padi dan mengevaluasi potensinya sebagai agens biokontrol terhadap P.oryzae.
Manfaat Penelitian
4
BAHAN DAN METODE
Waktu dan Tempat Penelitian
Penelitian ini dilaksanakan dari bulan Februari 2014 sampai Oktober 2014 di Laboratorium Mikologi Tumbuhan, Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian Bogor.
Bahan dan Alat
Bahan yang digunakan ialah daun padi bergejala blas, tanah perakaran dari padi sawah di Desa Babakan Lebak, Balumbang Jaya, Cikarawang, dan Situgede, media Nutrient Agar (NA), media Potato Dextrose Agar (PDA), media Luria
Isolasi penyebab blas dilakukan dari daun padi sawah di Desa Cikarawang, Bogor. Daun padi sawah yang bergejala blas dipotong dengan ukuran 0.5 x 0.5 cm. Setiap potongan terdiri atas bagian yang sakit dan sehat. Daun disterilisasi dengan pencelupan dalam larutan NaOCl 2% selama 1 menit, dibilas dengan air steril sebanyak 3 kali dan ditiriskan dengan tisu steril. Potongan daun bergejala blas tersebut diinokulasi pada cawan petri berisi media PDA (Potato Dextrose Agar) yang telah ditambahkan NaCl 0.1% dan diinkubasi pada suhu ruang. Pemurnian segera dilakukan setelah hifa tumbuh saat 3 hari setelah inokulasi (HSI).
Isolat patogen penyebab blas berumur 6 HSI diinokulasikan pada daun padi berumur 21 hari yang sudah dilukai dengan jarum steril. Inokulasi dilakukan dengan penempelan potongan inokulum pada daun yang telah dilukai. Selanjutnya, daun tersebut ditutup dengan kapas steril yang sudah dilembapkan dengan akuades steril dan dilapisi dengan plastic wrapping. Setelah 7 HSI, kapas dilepas dan diamati perkembangan gejala yang terjadi.
Bila daun padi tersebut menunjukkan gejala penyakit blas, dilakukan reisolasi pada media PDA yang telah ditambahkan NaCl 0.1% dan diidentifikasi secara mikroskopis. Pengamatan mikroskopis dilakukan dengan mengamati bentuk konidia dan hifa.
Eksplorasi Rizobakteri
5 Setelah itu, cawan petri diinkubasi pada suhu ruang hingga berbagai macam bentuk koloni rizobakteri tumbuh. Pemurnian dilakukan dengan metode penggoresan kuadran hingga didapatkan koloni rizobakteri yang murni (Hadioetomo 1993).
Uji Non Patogenik IsolatRizobakteri
Uji non patogenik isolat rizobakteri dilakukan dengan uji hipersensitif pada daun tembakau berumur 3 bulan yang diperoleh dari Balai Besar Litbang Bioteknologi dan Sumber Daya Genetik Pertanian Bogor. Rizobakteri dibiakkan pada media LB (Luria Broth) lalu dikocok menggunakan shaker dengan kecepatan 80 rpm selama 24 jam. Biakan rizobakteri disuntikkan pada bagian bawah daun tembakau sebanyak 1 ml kemudian diamati setelah 48 jam. Rizobakteri yang bersifat patogen akan menyebabkan gejala nekrotik pada daun tembakau.
Identifikasi Rizobakteri
Identifikasi rizobakteri yang tidak bersifat patogenik dilakukan dengan uji Gram. Uji Gram menggunakan KOH 3% yang diteteskan ke 1 lup bakteri pada gelas preparat. Uji Gram ditentukan dengan melihat ada atau tidaknya pembentukan lendir dari campuran rizobakteri dan KOH 3%. Jika tidak terbentuk lendir, maka isolat rizobakteri tergolong ke dalam Gram positif dan jika terbentuk lendir, maka termasuk bakteri Gram negatif.
Uji Antagonisme antara Rizobakteri dan P.oryzae
Pengujian dilakukan menggunakan metode umpan beracun. Isolat rizobakteri yang akan diuji terlebih dahulu dilakukan pengenceran bertingkat hingga 10-1 menggunakan 9 ml akuades steril. Suspensi rizobakteri 10-1 diteteskan ke cawan petri sebanyak 1 ml, kemudian media PDA+NaCl 0.1% bersuhu 50 ºC dituang. Cawan petri digoyang mengikuti angka 8 sebanyak 10 kali agar isolat rizobakteri dan media tercampur. Isolat P. oryzae diletakkan dengan menggunakan pelubang gabus 0.5 cm di tengah media yang sudah mengeras. Pengamatan dan pengukuran dilakukan setiap hari terhadap diameter zona bening yang terbentuk dari interaksi keduanya dan diameter pertumbuhan P.oryzae. Rumus persentase penghambatan:
Keterangan :
d = presentase penghambatan pertumbuhan P.oryzae (%) d1 = diameter pertumbuhan P.oryzae perlakuan kontrol (cm) d2 = diameter pertumbuhan P.oryzae perlakuan rizobakteri (cm)
Analisis Data
6
HASIL DAN PEMBAHASAN
Isolasi dan Identifikasi Penyebab Blas
Pengamatan dan isolasi penyakit blas pada tanaman padi dilakukan di Desa Cikarawang, Bogor. Sebelum dilakukan isolasi, survei penyakit blas juga dilakukan di Desa Situgede, tetapi kejadian dan keparahan penyakit blas di sana lebih rendah dibandingkan dengan Desa Cikarawang. Penyakit blas daun paling mendominasi di Desa Cikarawang. Semua stadia umur padi dapat terserang penyakit blas, namun pada fase generatif gejala terlihat lebih jelas dan bila sudah parah terdapat blas leher malai.
Penyakit blas yang ditemukan di bagian daun (leaf blast) berupa bercak coklat kehitaman berbentuk belah ketupat dengan pusat bercak berwarna putih atau kelabu, di sekitar bercak terlihat dikelilingi oleh warna kuning pucat (Gambar 1a, 1b). Bercak belah ketupat dapat membesar, menyatu, dan mematikan keseluruhan daun. Blas juga menyebabkan tanaman menjadi kerdil serta mengurangi jumlah pembentukan malai (Ou 1985). Gejala blas daun biasanya diawali dengan bercak kecil berwarna keputihan, abu-abu, atau kebiruan.
Hasil reinokulasi penyebab blas pada daun padi (Gambar 2a) menunjukkan gejala yang sama yaitu berupa bercak coklat kehitaman berbentuk belah ketupat
dengan pusat bercak berwarna putih atau kelabu dan di sekitar bercak terlihat dikelilingi oleh warna kuning pucat (Gambar 2b, 2c).
Gambar 1 Gejala blas pada permukaan daun (a,b)
b
Gambar 2 Inokulasi isolat P.oryzae pada daun padi (a),gejala blas pada daun setelah diinokulasi (b,c)
a b
7
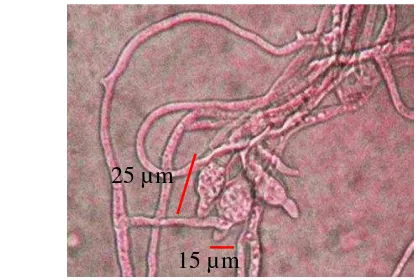
Setelah daun tersebut direisolasi, dilakukan pengamatan mikroskopis yang menunjukkan bahwa konidia berbentuk seperti buah pir berukuran 15µm x 25µm dengan miselium bersekat (Gambar 3). Hal ini sesuai dengan Barnett dan Hunter (1998) serta Agrios (2005) yang menjelaskan bahwa konidia P.oryzae berbentuk seperti buah pir, berbentuk hampir elips dengan ujung konidia yang membulat.
Menurut Ou (1985) ukuran konidia P.oryzae berkisar antara 19-23 × 7-9µm. Ukuran dan bentuk konidia bisa berbeda-beda tergantung dari ras patogen dan kondisi lingkungan. Kondisi lingkungan yang optimum untuk infeksi dan sporulasi P.oryzae saat kelembapan 95% dengan suhu 26-27 °C (Munoz 2008). Perkembangan bercak dipengaruhi kerentanan varietas dan umur bercak itu sendiri. Bercak dapat berkembang hingga mencapai panjang 1-1.5 cm dan lebar mencapai 0.3-0.5 cm. Bercak pada varietas rentan yang berkembang pada kelembapan tinggi dan di bawah naungan menunjukkan bercak kecoklatan yang tidak lebar tetapi memiliki daerah halo berwarna kekuningan di daerah sekitar bercak. Pada varietas tahan, bercak tidak akan berkembang dan tetap seperti titik . Hal ini terjadi karena adanya penghambatan perkembangan P.oryzae dalam jaringan inang.
Eksplorasi Rizobakteri
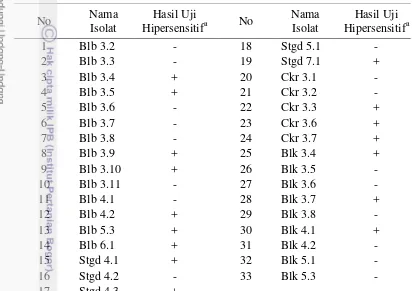
Eksplorasi rizobakteri dilakukan di 4 desa sekitar kampus IPB yaitu Cikarawang (Ckr), Situgede (Stgd), Balumbang Jaya (Blb), dan Babakan lebak (Blk). Isolasi rizobakteri yang dilakukan di Desa Balumbang Jaya menghasilkan 14 isolat, Desa Situgede 5 isolat, Desa Cikarawang 5 isolat, dan Desa Babakan Lebak 9 isolat (Tabel 1). Sebanyak 16 isolat merupakan bakteri patogenik karena menyebabkan daun tembakau mengalami gejala nekrosis sedangkan 17 isolat lainnya tidak menyebabkan gejala nekrotik di daun tembakau (Gambar 4b). Gejala nekrotik yang ditunjukkan dalam penelitian ini hampir seluruhnya sama yaitu bercak kecoklatan yang menyebar dan membesar disertai halo berwarna kuning di pinggiran bercak (Gambar 4a). Pada uji hipersensitif ketahanan tanaman mengenali adanya molekul sinyal khusus yang dihasilkan oleh patogen yang disebut elisitor. Pengenalan elisitor oleh ketahanan tanaman mengaktifkan reaksi biokimia di sekitar sel yang terserang dan menyebabkan hancurnya membran seluler dari sel-sel yang berkontak dengan bakteri. Sel-sel tersebut akan mengering dan daun tembakau pada percobaan ini mengalami nekrosis. Jaringan
Gambar 3 Mikroskopis konidia P.oryzae dengan perbesaran 40x10 15 µm
8
daun yang mengalami nekrosis bertujuan mengisolasi patogen dari substansi hidup di sekitarnya sehingga menyebabkan patogen tersebut mati (Agrios 2005).
Gambar 4 Hasil uji hipersensitif pada daun tembakau; isolat rizobakteri yang menimbulkan gejala nekrotik pada daun tembakau (a), isolat rizobakteri yang tidak menimbulkan gejala nekrotik pada daun tembakau (b)
Perbedaan jumlah isolat yang didapat dipengaruhi oleh keragaman mikroorganisme yang ada di rizosfer. Mikroorganisme yang menyokong Tabel 1 Hasil eksplorasi rizobakteri dan uji hipersensitif dari Desa Balumbang
Jaya (Blb), Situgede (Stgd), Cikarawang (Ckr), dan Babakan Lebak
menujukkan tidak ada gejala apapun saat uji hipersensitif
9 pertumbuhan tanaman berkembang baik di tanah yang mengandung banyak unsur hara. Faktor fisik dan kimia tanah seperti kelembapan, suhu, kandungan oksigen, karbon dioksida, hara, dan pH akan mempengaruhi aktivitas mikroorganisme di dalamnya (Johnson dan Curl 1972). Beberapa jenis tanah yang mampu membuat mikroorganisme antagonis berkembang baik sehingga dapat menyebabkan patogen tanaman tertekan perkembanganya disebut sebagai suppressive soil. Mekanismenya melibatkan faktor abiotik dan biotik seperti menghasilkan antibiotik, kompetisi terhadap makanan, dan parasitisme langsung (Agrios 2005).
Menurut Hadiwiyono (2010), tanah supresif dibentuk dan dipengaruhi oleh ekosistem yang kompleks. Faktor yang paling berperan pada supresivitas tanah adalah faktor hayati sedangkan faktor nonhayati berperan sebagai pendukung dengan memberikan kondisi yang sesuai untuk aktivitas hayati dan sebagai sumber nutrisi pada aktivitas mikrob yang ada di dalamnya. Usaha yang umum dilakukan adalah memanipulasi kondisi fisik tanah seperti tekstur, struktur, temperatur, dan kelembaban tanah adalah melakukan solarisasi tanah. Praktik budidaya juga dapat diarahkan agar tidak menghambat perkembangan aktivitas mikrob yang berperan pada supresivitas tanah seperti melakukan rotasi tanaman.
Pengamatan karakterisitk koloni dilakukan pada 17 isolat rizobakteri yang tidak menunjukkan gejala nekrotik pada uji hipersensitif. Pemurnian dilakukan dengan metode penggoresan kuadran (Hadioetomo 1993). Ciri koloni yang diamati yaitu warna, elevasi, tepian dan bentuk (Tabel 2).
No Isolat Bentuk Tepian Elevasi Warna
10
No Isolat Bentuk Tepian Elevasi Warna
3
Blb 3.6
bundar licin cembung putih
4
Blb 3.7
bundar berombak datar putih
5
Blb 3.8
bundar berombak datar putih
6
Blb 3.11
bundar berombak datar putih
7
Blb 4.1
tak beraturan
dan menyebar
11
No Isolat Bentuk Tepian Elevasi Warna
8
Stgd 4.2
bundar berombak datar kuning pekat
9
Stgd 5.1
bundar licin datar putih kekuningan
10
Ckr 3.1
bundar licin cembung putih
11
Ckr 3.2
bundar licin berombak putih
12
Blk 3.5
bundar tak
12
No Isolat Bentuk Tepian Elevasi Warna
13
Blk 3.6
tak beraturan
dan menyebar
berombak datar putih
14
Blk 3.8
bundar licin datar putih kekuningan
15
Blk 4.2
bundar licin timbul putih
16
Blk 5.1
bundar licin cembung putih
17
Blk 5.3
13
Mekanisme Antagonisme Rizobakteriterhadap P.oryzae
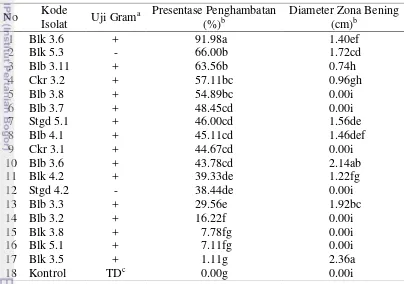
Isolat rizobakteri yang tidak menyebabkan gejala nekrotik di daun tembakau diuji Gram dan antagonisme dengan isolat P.oryzae. Hasil uji Gram menunjukkan bahwa terdapat 2 isolat bakteri Gram negatif dan 15 isolat bakteri Gram positif (Tabel 3). Isolat rizobateri Gram negatif akan menunjukkan pembentukan lendir setelah tercampur dengan KOH 3% (Gambar 5a) sedangkan isolat Gram positif tidak terdapat pembentukan lendir (Gambar 5b). Menurut Shivas dan Beasley (2005) lendir yang terbentuk pada bakeri Gram negatif disebabkan dinding sel bakteri Gram negatif lebih sensitif dan tidak memiliki ketahanan terhadap penghambat basa seperti KOH sehingga dinding sel akan pecah dan melisis. Setelah itu, DNA akan keluar dari sel dan terbentuklah benang-benang lendir. Pada bakteri Gram positif tidak terdapat pembentukan lendir karena dinding selnya lebih resisten terhadap KOH sehingga dinding sel tidak pecah.
Tabel 4 Hasil uji Gram dan uji antagonisme rizobakteri terhadap P.oryzae No Kode
Tanda + merupakan bakteri Gram positif, tanda -merupakan bakteri Gram negatif. b Nilai tengah
yang memiliki huruf yang sama pada kolom yang sama tidak berbeda nyata berdasarkan DMRT
α=5%.c
14
Uji antagonisme menunjukkan 14 isolat rizobakteri mampu menghambat pertumbuhan P.oryzae dan 3 isolat lainya menunjukkan hasil yang tidak berbeda nyata dengan kontrol ((Tabel 3). Isolat Blk 3.6 mampu menghambat pertumbuhan cendawan P.oryzae dengan persentase paling besar yaitu 91.98% dengan pertumbuhan diameter P.oryzae 0.72 cm saat uji antagonisme (Lampiran 1). Isolat yang tidak mampu menghambat pertumbuhan P.oryzae yaitu Blk 3.5, Blk 5.1, dan Blk 3.8. Terdapat 10 isolat rizobakteri menunjukkan pembentukan zona bening (Gambar 6) dan 7 isolat tidak membentuk zona bening. Isolat Blk 3.5 memiliki diameter zona bening paling besar yaitu 2.36 cm dan isolat Blb 3.11 memiliki diameter paling kecil yaitu 0.74 cm.
Gambar 6 Zona bening yang terbentuk saat uji antagonisme antara rizobakteri dan P.oryzae
Keragaman kemampuan antagonisme isolat rizobakteri tergantung pada jenis zat antifungal yang dihasilkan dan keragaman isolat itu sendiri (William and Asher 1996). Zona bening yang dihasilkan dalam uji antagonisme in vitro pada percobaan ini diduga karena adanya produksi zat antifungal seperti antibiotik. Gamard dan De Boer (1995) melaporkan bahwa salah satu mekanisme rizobakteri dalam menekan perkembangan patogen adalah dengan menghasilkan antibiotik yang ditunjukkan dengan adanya zona bening di sekitar inokulum patogen saat uji antagonisme. Antibiosis yang ditunjukkan dalam perlakuan in vitro akan Gambar 5 Hasil uji Gram pada isolat rizobakteri; isolat bakteri Gram negatif (a),
hasil uji Gram pada isolat bakteri Gram positif (b)
a b
15 memudahkan dalam menyeleksi mikroorganisme yang berpotensi sebagai agens biokontrol. Beberapa mekanisme antibiotik dalam menekan mikroorganisme lain ialah penghambatan sinstesis dinding sel, penghambatan sintesis protein, penghambatan sintesis DNA/RNA (Toha 2004). Chaiharn et al. (2008) juga menggunakan zona bening sebagai indikator adanya enzim litik yang dapat mendegradasi dinding sel patogen. Produksi enzim litik adalah salah satu mekanisme antagonis yang penting karena mampu mendegradasi dinding sel cendawan. Walaupun demikian, tidak ada bukti yang pasti untuk menunjukkan mekanisme rizobakteri yang tepat karena banyaknya aktivitas biokontrol yang terlibat dalam penghambatan pertumbuhan patogen (Wahyudi et al. 2011).
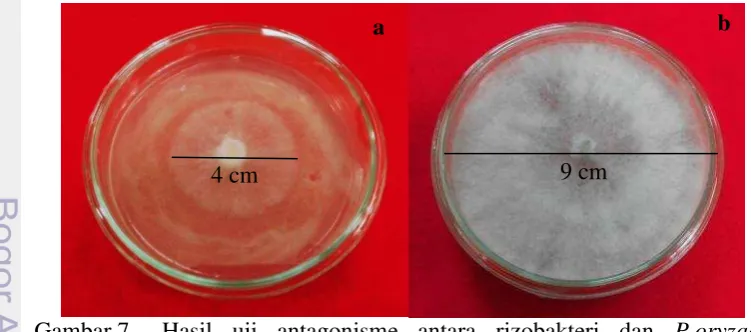
Wahyudi et al. (2011) melakukan penelitian pada Pseudomonas sp. sebagai agens antagonis terhadap Fusarium oxysporum, Rhizoctonia solani, dan Sclerotium rolfsii. Beberapa isolat yang menunjukkan adanya produksi enzim kitinase ataupun aktivitas siderofor tidak dapat menghambat pertumbuhan patogen yang diuji tetapi beberapa isolat yang tidak menunjukkan adanya produksi enzim kitinase ataupun aktivitas siderofor dapat menghambat pertumbuhan patogen yang diuji. Hal ini diduga karena adanya mekanisme biokontrol yang lain seperti produksi HCN atau antibiotik yang lain. Lima isolat rizobakteri dalam percobaan ini tidak menunjukkan adanya pembentukan zona bening seperti Blb 3.8, Blb 3.7, Ckr 3.1, Stgd 4.2, dan Blb 3.2, namun tetap menunjukkan adanya penghambatan pertumbuhan P.oryzae yang lebih tinggi (Gambar 7a) dibandingkan dengan perlakuan kontrol (Gambar 7b). Zona bening yang tidak terbentuk pada beberapa isolat dalam percobaan ini diduga karena mekanisme biokontrol lain yang lebih mendominasi dibandingkan dengan produksi antibiotik. Isolat Blk 3.5 yang memiliki zona bening terbesar namun tidak dapat menghambat pertumbuhan P.oryzae karena pembentukan zona bening yang besar tidak menentukan adanya antagonisme yang tinggi dari isolat tersebut.
Beberapa bakteri rizosfer yang sudah sering dijadikan agens hayati dan sudah dikomersialkan adalah Psedomonas fluorescens dan Bacillus subtilis. Psedomonas fluorescens adalah contoh rizobakteri Gram negatif yang dapat menghasilkan enzim kitinolitik. Enzim ini berperan dalam mendegradasi senyawa
4 cm
a
Gambar 7 Hasil uji antagonisme antara rizobakteri dan P.oryzae; perlakuan rizobakteri yang tidak membentuk zona bening (a), perlakuan kontrol (b)
b
16
kitin yang merupakan salah satu komponen penyusun dinding hifa fungi (Folders et al. 2001). Mekanisme lainnya yaitu dengan menghasilkan beberapa antibiotik yaitu agrocin 84, agrocin 434, 2,4-diacetylphloroglucinol (DAPG), herbicolin, oomycin, phenazin, pyoluteorin, dan pyrrolnitrin (Glick dan Pasternak 2003). Bacillus subtilis adalah salah satu rizobakteri gram positif. B.subtilis dapat menghasilkan berbagai jenis antibiotik yaitu oligomycin A, kanosamine, zwittermicin A, dan xanthobaccin (Glick dan Pasternak 2003). Killani et al (2011) melaporkan bahwa B.subtilis menunjukkan kemampuan antibiosis yang tinggi dalam menekan Fusarium verticilloides, F.equiseti, F. Solani, F. Oxysporum, R. solani secara in vitro. Selain dengan antiboisis, bakteri Gram positif yang lain seperti Azospirillum yang memiliki keunggulan dalam kompetisi memperebutkan nutrisi seperti glukosa, asam amino, asam organik. Dalam aplikasinya di dalam tanah, Azospirillum memiliki kemotaksis untuk bergerak menuju ke arah senyawa tersebut (Compant et al. 2005).
Iaolat rizobakteri Blk 3.6, Blk 5.3, Blb 3.11, dan Ckr 3.2 menunjukkan pembentukan zona bening dan presentasi penghambatan pertumbuhan P.oryzae in vitro yang relatif tinggi dibandingkan isolat lainnya. Oleh karena itu, isolat-isolat tersebut berpotensi untuk digunakan sebagai agens pengendali penyakit blas.
17
SIMPULAN DAN SARAN
Simpulan
Hasil eksplorasi dari sawah di Desa Babakan Lebak, Balumbang Jaya, Cikarawang, dan Situgede diperoleh 33 isolat rizobakteri. Sebanyak 17 isolat merupakan bakteri non patogenik, 15 isolat diantaranya merupakan bakteri Gram positif dan 2 isolat bakteri Gram negatif. Rizobakteri yang relatif tinggi dalam menghambat pertumbuhan P.oryzae in vitro ialah isolat Blk 3.6 (Gram positif), Blk 5.3 (Gram negatif), Blb 3.11 (Gram positif), dan Ckr 3.2 (Gram positif) berturut-turut sebesar 91.98, 66, 63.56, dan 57.11% .
Saran
Perlu dilakukan uji lanjut yaitu identifikasi rizobakteri yang telah ditemukan, uji pada media agar darah untuk mengetahui apakah rizobakteri yang ditemukan bersifat parasit terhadap manusia atau tidak, dan aplikasi rizobakteri pada padi sawah.
18 rizobakteri for their plant growth promoting activities. Sci Tech J. 8(1): 18-23.
Compant S, Duffy B, Nowak J, Clement C, Barka EA. 2005. Use of plant growth promoting bacteria for biocontrol of plant disease: principles, mechanism of action, and future prospects. Appl Environ Microbiol. 71(9): 4951-4959. Dewi IM, Cholil A, Muhibuddin A. 2013. Hubungan karakteristik jaringan daun
dengan tingkat serangan penyakit blas daun (Pyricularia oryzae Cav.) pada beberapa genotype padi (Oryza sativa L.). J HPT. 2(1): 10-18.
Fatima Z, Saleemi M, Zia M, Sultan T, Aslam M, Rehman R, Chaudhary MF. 2009. Antifungal activity of plant growth-promoting rhizobacteria isolates against Rhizoctonia solani in wheat. Afr J Biotechnol. 8(2):219-225.
Folders J, Algra J, Roelofs MS, Leendert CL, Tommasen J, Bitter W. 2001. Characterization of Pseudomonas aeruginosa chitinase, a gradually secreted protein. J Bacteriol. 183(24): 7044-7052.
Gamard P, De Boer SH. 1995. Evaluation of antagonistic bacteria for suppression of bacterial ring rot of potato. Eur J Plant Path. 101(1): 519-525.
Glick BR, Pasternak JJ. 2003. Molecular Biotechnology Principles and Application of Recombinant DNA. 3rd ed. Washington (US): ASM press. Hadioetomo RS. 1993. Mikrobiologi Dasar dalam Praktek: Teknik dan Prosedur
Dasar Laboratorium. Jakarta: Gramedia Pustaka Utama.
Hadiwiyono. 2010. Tanah supresif dalam praktik pengelolaan penyakit tumbuhan. J Ilmu Tanah dan Agroklimatologi. 7(1): 31-40.
Hamer JE. 1988. Host species-specific conservation of repeated DNA elements in the genome of a fungal plant pathogen. Proc Natl Acad Sci. 86(1):9981-Borne Plant Pathogens. Minossota (US): Burgess Publishing Company.
Kharisma SD, Cholil A, Aini LQ. 2013. Ketahanan beberapa genotipe padi hibrida (Oryza sativa L.) terhadap Pyricularia oryzae Cav. penyebab penyakit blas daun padi [Internet] [diunduh 2014 Apr 20];1(2):19-27. Tersedia pada: http://jurnalhpt.studentjournal.ub.ac.id/index.php/jhpt/article/ download/14/23.
19
[Internet] [diunduh pada 2014 Jan 20]. Tersedia pada: http://www. sciencepub.net/researcher/research0303/02_4069research0303_11_18.pdf. Lugtenberg B, Kamilova F. 2009. Plant growth promoting rhizobacteria. Annu
Rev Microbiol. 63(1):541-556. doi:62.081307.162918.
Maheshwari DK, Saraf M. 2013. Bacteria in Agrobiology: Crop Productivity. Berlin(DE): Springer.
Menteri Pertanian RI. 1995. Peraturan Menteri Pertanian Nomor 411/KPTS/TP.120/6/1995 tentang Pemasukan Agens Hayati ke dalam Wilayah Negara Kesatuan Republik Indonesia. Deptan: Jakarta.
Munoz MC. 2008. The effect of temperature and realtiive humidity on the airborne concentration of Pyricularia oryzae spores and the development of rice blast in southern Spain. Spain J Agric Res. 6(1): 61-69.
Ou SH. 1985. Rice Disease 2nd ed. London (GB): The Cambrian News (Aberystwyth) Ltd.
Puslitbang Tanaman Pangan. 2007a. Masalah Lapang: Hama, Penyakit, dan Hara pada Padi ed ke-3. Jakarta (ID): Puslitbang Tanaman Pangan Press.
Puslitbang Tanaman Pangan. 2007b. Peningkatan produksi padi menuju 2020 [Internet] [diunduh 2014 Apr 20]. Tersedia pada: http://www. puslittan.bogor.net/index.php?bawaan=download/download_detail&&id=35. Reflinur, Bustama M, Widyastuti U, Aswidinoor H. 2005. Keragaman genetik
cendawan Pyricularia oryzae berdasarkan primer spesifik gen virulensi. J Bioteknol Pertan. 10(2):55-60.
Santoso, Nasution A. 2009. Pengendalian penyakit blas dan penyakit cendawan lainnya. Penelitian Pertanian Tanaman Pangan. [Internet] [diunduh pada 2014 Januari 20]. Tersedia pada: http://www.litbang.deptan.go.idspecial /padi/bbpadi_2009_itp_20.pdf.
Scardaci SC. 1997. Rice Blast . A New Disease in California. Agronomy Fact Sheet Series 1997-2. Davis(US): University of California.
Shivas R, Beasley D. 2005. Management of Plant Pathogens Collections. Australia (AUS): Queensland Department of Primary Industries and Fisheries.
Shymala, Sivakumaar. 2012. Integrated control of blast disease of rice using the antagonistic rhizobacteria Psedomonas fluorescens and the resistance inducing chemical salicylic acid [Internet] [diunduh 2014 April 20]. Tersedia pada: http://urpjournals.com/tocjnls/44_12v2i4_6.pdf.
Supriadi. 2006. Analisis risiko agens hayati untuk pengendalian patogen pada tanaman. J Litb Pertan. 25(3): 75-80.
Toha AHA. 2004. Ensiklopedia Biokimia dan Biologi Molekul. Manokwari (ID): Universitas Negeri Papua.
Wahyudi AT, Astuti RI, Giyanto. 2011. Screening of Pseudomonas sp. isolated from rhizosphere of soybean plant as plant growth promoter and biocontrol agent. Amr J Agri Biol Sci. 6(1): 134-141.
Williams GE, Asher MJC. 1996. Selection of rhizobacteria for the control of Phythium ultimum and Aphanomyces cochiliodes on sugar beet seedlings. Crop Prot. 15: 479-486.
20
21 Lampiran 1 Pertumbuhan diameter P.oryzae pada uji antagonisme dengan
rizobakteri
No Kode Isolat Pertumbuhan Diamater P.oryzae (cm)a
1 Kontrol 9.00 g
2 Blk 3.5 8.90 g
3 Blk 5.1 8.36 fg
4 Blk 3.8 8.30 fg
5 Blb 3.2 7.54 f
6 Blb 3.3 6.34 e
7 Blb 3.7 6.30 e
8 Stgd 4.2 5.54 e
9 Blk 4.2 5.46 cd
10 Blb 3.6 5.06 cd
11 Ckr 3.1 4.98 cd
12 Blb 4.1 4.94 cd
13 Stgd 5.1 4.86 cd
14 Blb 3.8 4.06 bc
15 Ckr 3.2 3.86 bc
16 Blb 3.11 3.28 b
17 Blk 5.3 3.06 b
18 Blk 3.6 0.72 a
a
Nilai tengah yang memiliki huruf yang sama pada kolom yang sama tidak berbeda
22
RIWAYAT HIDUP
Penulis dilahirkan di Samarinda pada tanggal 18 September 1992 dari pasangan Khairul Anam dan Farida. Penulis adalah putri pertama dari tiga bersaudara. Penulis menyelesaikan pendidikan TK Tunas Mulia Samarinda pada tahun 1998, SDN 027 Samarinda pada tahun 2004, SMPN 1 Samarinda pada tahun 2007, dan SMAN 10 Samarinda pada tahun 2010. Penulis diterima di Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian Bogor (IPB) pada tahun 2010 melalui jalur Undangan Seleksi Masuk IPB (USMI).
Kegiatan intra dan ekstrakurikuler yang pernah diikuti di IPB adalah menjadi asisten praktikum Entomologi Umum (2012/2013), Dasar-Dasar Proteksi Tanaman (2012/2013 dan 2014/2015), Hama Gudang dan Permukiman (2013/2014). Penulis juga aktif sebagai pengurus Badan Eksekutif Mahasiswa Fakultas Pertanian divisi Mitra Desa (2013) dan Badan Eksekutif Mahasiswa Keluarga Mahasiswa IPB divisi Komunikasi dan Informasi (2014). Penulis juga aktif mengikuti kepanitiaan dalam kegiatan departemen, fakultas, dan IPB.