BAB VI
ISOLASI PROTOPLAS, FUSI PROTOPLAS DAN REGENERASI HASIL FUSI ANTARA TANAMAN JERUK SIAM SIMADU
DENGAN MANDARIN SATSUMA
Ringkasan
Penelitian isolasi protoplas, fusi protoplas, dan regenerasi hasil fusi antara jeruk siam Simadu dengan Mandarin Satsuma untuk mendapatkan hibrida baru antara jeruk siam Simadu dengan Mandarin Satsuma.Penelitian dilakukan di laboratorium Biologi sel dan Jaringan BB-Biogen Bogor dari bulan Februari – juni 2008. Dari hail penelitian diperoleh bahwa jenis enzim, konsentrasi, dan kombinasi enzim yang digunakan sangat berpengaruh dalam keberhasilan isolasi protoplas. Kombinasi enzim selulase Onozuka R10-Yakult 1% dengan maserozim R10-Yakult 1% dalam larutan CPW yang dimurnikan dengan campuran sukrosa 25% dengan manitol 13% dapat menghasilkan protoplas dengan densitas 15.7x105 protoplas/g kalus dan 13.0x105 protoplas/g daun. Konsentrasi PEG yang digunakan untuk menginduksi fusi protoplas dari kalus jeruk siam Simadu dengan protoplas mesopil daun in vitro berpengaruh terhadap jumlah rata-rata protoplas berfusi yang dihasilkan. Rata-rata jumlah protoplas yang dihasilkan dari induksi fusi dengan PEG 4% adalah 3.3 fusan yang hetero fusi, 5 fusan yang homo fusi dan multi fusi. Rata-rata jumlah protoplas yang dihasilkan dari induksi fusi dengan PEG 30% adalah 4.7 fusan hetero fusi, 6.7 fusan homo fusi, dan 7.7 fusan multi fusi. Frekuwensi fusi meningkat menjadi 4.3 fusan hetero fusi, 6.7 fusan homofusi, dan 7.7 multi fusi dari induksi PEG 4% serta menjadi 5.7 fusan hetero fusi, 7.7 fusan homo fusi, dan 6.3 fusan multi fusi dari induksi fusi dengan PEG 30%. Keberhasilan regenerasi protoplas hasil fusi pada media kultur yang digunakan dipengaruhi oleh konsentrasi PEG yang digunakan untuk induksi terjadinya fusi. Protoplas yang yang difusikan dengan PEG 4% dapat beregenerasi membentuk dinding sel, melakukan pembelahan sel, koloni sel, mikro kalus, dan embrio somatik pada media, sedangkan protoplas yang difusikan dengan PEG 30% hanya dapat beregenerasi membentuk didnding sel dan pembelahan sel. Penambahan ABA 0.5 mg/l pada media MW dapat mendewasakan embrio somatik dan GA3 0.5 mg/l dapat menginduksi perkecambahan embrio somatik menjadi plantlet dengan efisisensi perkecambahan sebesar 76%.
Kata kunci: Isolasi protoplas, fusi protoplas, regenerasi fusan, jeruk siam Simadu, dan Mandarin Satsuma.
PROTOPLAST ISOLATION AND FUSION AND FUSAN REGENERATION BETWEEN SIMADU TANGERINE WITH SATSUMA MANDARIN
Abstract
Research protoplast isolation, protoplast fusion and regeneration of fusion between Simadu tangerines with Satsuma Mandarin to get a new hybrid seedless. The study was conducted in the laboratory of Cell and tissue Biology, BB BIOGEN Bogor, from February - June 2008. The result showed that protoplasts from embryogenic callus of Simadu tangerine and in vitro leaf of mandarin Satsuma can be isolated in large numbers by using a combination of cellulase Onozuka R10, Yakult Yakult% 1% with R10 Maserozim in CPW solution. Protoplasts are purified with a mixture of 25% sucrose with 13% mannitol. Protoplast density produced from embryogenic callus is 15.7x105 protoplasts / g callus and 13x105 protoplasts/ g in vitro leaf. The concentration of PEG used to induce fusion between mesophyll protoplasts mandarin Satsuma and siam Simadu callus affect the average number of protoplasts fusion. Average number of protoplasts were fused by using PEG 4% are 3.3 hetero fusion, 5.0 homo fusion and multi-fusion. Average number of protoplasts were fused by using PEG 30% are 4.7 hetero fusion, 6.7 homo fusion 6.7 and 7.7multi-fusion. The success of fusan regeneration on regeneration medium is influenced by PEG concentration used for induction of fusion. Protoplasts were fused with PEG only 4%, which can regenerate to form cell walls, making cell division, colony cells, micro-callus and somatic embryo. Protoplasts were fused with PEG 30% can only be regenerated to form the cell wall and cell division. Giving light to the culture after 2 weeks may accelerate cell division that can form colonies of cells. Dilutions of cell suspension with the same medium (without 2, 4-D) can accelerate the growth and development of the protoplasts formed colonies of cells, micro-callus and somatic embryos. MW medium is best used in the fusion because it can encourage of somatic embryos directly. Addition of ABA 0.5 mg / l in the media can produce somatic embryos and GA3 0.5 mg / l can germinate mature somatic embryos to plantlets with 76% germination efficiency.
Keywords : Protoplast isolation, protoplast fusion, fusan regeneration, siam Simadu, and Mandarin Satsuma
Pendahuluan
Trend kebutuhan pasar dunia secara global akan buah jeruk yang dikonsumsi segar saat ini dan masa mendatang adalah perlu memenuhi kategori buah yang tidak berbiji (seedless), mudah dikupas (easy peeling) dan mempunyai tipe mandarin dengan warna yang menarik (pigmented). Jeruk siam Pontianak dan Simadu adalah dua dari jenis jeruk batang atas komersial (scion) yang banyak dikenal di Indonesia. Akan tetapi kedua jenis jeruk tersebut masih mempunyai biji yang relatif banyak (15-23 biji per buah) dan warna (pigmented) belum begitu menarik sehingga kalah bersaing dengan jeruk produk negara lain. Untuk menghindari tekanan buah jeruk impor tersebut maka diperlukan sentuhan inovasi teknologi terhadap jeruk lokal tersebut untuk meningkatkan kualitas buah sehingga dapat diterima dan bersaing di pasar global (Husni 2007).
Untuk mendapatkan tanaman jeruk yang mempunyai karakter buah seedless pada tanaman jeruk sudah dimulai dilakukan beberapa dekade yang lalu melalui pemuliaan konvensional. Satsuma mandarin (C. Unshiu Marc.) adalah merupakan jenis jeruk yang secara alami mempunyai sifat seedless (Kunittake et al. 1991; Spiegel-Roy and Goldschmidt 1996). Sifat seedless tersebut telah terbukti disebabkan oleh pollennya yang steril (male strility) yang termasuk dalam cytoplasmic male strility biasa disebut dengan istilah CMS (Yamamoto et al. 1997). Untuk memindahkan sifat CMS tersebut dari jeruk mandarin Satsuma kepada kultivar jeruk siam Simadu sangat sulit dilakukan melalui pemuliaan konvensional karena adanya faktor genetik pembatas (incompatible). Oleh karena itu perlu dicari cara lain untuk memindahkan sifat seedless dari jeruk mandarin Satsuma ke kultivar jeruk lainnya.
Salah satu teknologi yang dapat digunakan adalah teknik fusi protoplas (Grosser et al. 1996; Moriguchi et al. 1996; Grosser and Gemitter 2005). Pada tanaman jeruk, teknik fusi protoplas mulai berkembang setelah Ohgawara et al. (1985) melaporkan keberhasilannya mendapatkan hibrida somatik antara C. sinensis dengan Poncirus tripoliata yang secara genetik inkompatibel. Semenjak itu, teknik tersebut banyak digunakan dalam program pemuliaan tanaman jeruk di dunia seperti
di Jepang oleh Kobayashi et al. (1988), Israel oleh Vardi et al. (1987), Amerika Serikat oleh Grosser dan Gemitter (1990), dan di Prancis oleh Ollitrault dan Luro (1996). Pada saat ini telah diperoleh lebih dari 40 kombinasi dari 250 jenis tetua jeruk melalui fusi protoplas (Grosser et al. 2000; Cabasson et al. 2001; Guo et al. 2004).
Beberapa hasil penelitian yang telah menggunakan teknologi fusi protoplas untuk perbaikan tanaman jeruk antara lain adalah fusi protoplas antara C. unshiu dengan C. sinensis (Yamamoto dan Kobayashi 1995; Guo et al. 2004; Xu et al. 2006; Cai et al. 2007) dan C. sinensis dengan Clausena lansium (Fu et al. 2003). De Carvalho Costa et al. (2003) juga telah menggunakan teknologi fusi protoplas untuk mendapatkan tanaman jeruk batang bawah yang toleran terhadap citrus blight, tristeza virus dan phytopthora dan Tusa et al. (2000) untuk ketahanan terhadap infeksi Phoma tracheiphila. Calixto et al. (2004) mendapatkan hibrida somatik dari C. sinensis dengan C. grandis yang toleran terhadap virus Citrus tristeza, Phytophthora, dan berpotensi digunakan sebagai batang bawah. Cai et al. (2007) juga menggunakan teknologi fusi protoplas untuk mendapatkan tanaman jeruk yang seedless hasil fusi antara C. unshiu dengan C. grandis dan C. sinensis. Untuk itu maka perlu dilakukan penelitian untuk mendapatkan hibrida baru dari jeruk siam kultivar Simadu dengan jeruk mandarin kultivar Satsuma.
Bahan dan Metode
Penelitian dilaksanakan di Laboratorium Kultur Jaringan, Kelompok Peneliti Biologi Sel dan Jaringan, Balai Besar Penelitian dan Pengembangan Bioteknologi dan Sumberdaya Genetik Pertanian, Bogor dari bulan Februari-Juni tahun 2008. Penelitian terdiri dari tiga tahap penelitian yang saling berhubungan yaitu: 1)isolasi protoplas, 2)fusi protoplas, dan 3)regenerasi protoplas hasil fusi.
Gambar 19. Penampakan kalus embriogenik dan daun in vitro yang digunakan sebagai sumber protoplas (A= kalus embriogenik dari jeruk siam Simadu dan B= daun in vitro dari jeruk mandarin Satsuma).
Isolasi protoplas
Bahan tanaman yang digunakan sebagai sumber protoplas pada penelitian ini adalah kalus embriogenik dari jeruk siam kultivar Simadu dan daun in vitro dari tanaman jeruk mandarin Satsuma (Gambar 19). Metode isolasi protoplas yang digunakan adalah metode isolasi yang terbaik dari hasil penelitian sebelumnya (penelitian 2) menggunakan larutan kombinasi selulase Onozuka RS10-Yakult 1% dengan macerozim RS10-Yakult 1% dalam larutan CPW (0.7 M manitol, 24.5 mM CaCl2, 0.92 mM NaH2PO4, dan 6.15 mM MES. Inkubasi dalam larutan enzim dilakukan selama 16 jam dalam keadaan gelap (tanpa cahaya). Pemurnian protoplas dilakukan dengan campuran larutan sukrosa 25% dengan manitol 13%.
Fusi protoplas
Induksi fusi dengan PEG dilakukan dengan metode fusi protoplas dari hasil penelitian sebelumnya (penelitian 3) menggunakan PEG 4% dan 30% dengan cara menambahkan 25 µl larutan PEG di empat titik di sekeliling suspensi protoplas yang telah dicampur dengan waktu induksi fusi selama 15 menit. Pengamatan dilakukan secara mikroskopik (in verted) dengan cara menghitung jumlah, persentase protoplas yang mengalami fusi, dan tipe fusi yang dihasilkan (hetro fusi, homo fusi, dan multi fusi) setelah diberi perlakuan PEG.
B A
Peningkatan frekuensi fusi
Untuk meningkatkan terjadinya frekuensi fusi juga menggunakan metode yang terbaik dari hasil penelitian sebelumnya (penelitian 3) dengan cara menambahkan 200 µl larutan pencuci (0.5 M manitol + 0.5 mM CaCl2).
Kultur protoplas hasil fusi
Kultur protoplas dilakukan dengan metoda Sihachakr (1998) dan Husni et al. (2004). Setelah dilakukan penetapan jumlah protoplas maka protoplas ditaburkan dalam cawan petri (∅ 50 mm x 15 mm) yang masing-masing berisi 5 ml media cair dengan densitas 104 protoplas/ml. Kultur diinkubasi dalam keadaan gelap tanpa cahaya dalam inkubator yang suhunya diatur pada temperatur 25 0
Media awal pertumbuhan protoplas yang digunakan adalah empat jenis media dasar yaitu KM, VKM, MW, dan MT ( Tabel Lampiran 3 dan 4). Sedangkan zat pengatur tumbuh yang digunakan adalah 0.1 mg/l 2,4_D + 3 mg/l BA + 500 mg/l EM + 50 g/l sukrosa. Kemasaman media (pH) ditetapkan pada kisaran 5.7-5.8. Media diseterilkan dengan filter ukuran 0.22 µm. Kultur disimpan dalam inkubator dalam keadan gelap tanpa cahaya pada suhu 25
C sampai terjadi pembentukan dinding sel (1-2 minggu). Setelah terjadi pembelahan sel membentuk koloni sel, kultur dipindahkan dan diberi cahaya dengan intensitas 1000 lux selama 16 jam sehingga terbentuk koloni sel yang lebih banyak.
0
C sampai terjadi pembentukan dinding sel dan pembelahan sel selama dua minggu. Setelah terjadi pembentukan dinding dan pembelahan sel, maka kultur dipindahkan ke ruang kultur yang diberi cahaya dengan intensitas 1000 lux selama 16 jam sampai terbentuk koloni sel. Pengamatan dilakukan secara mikroskopis dengan mikroskop inverted (Olymphus) dengan cara menggoyang kultur terlebih dahulu secara perlahan sehingga protoplas/sel merata keseluruh cawan petri. Kemudian dilakukan penghitungan protoplas yang dapat membentuk dinding sel dan sel yang telah melakukan pembelahan setiap bidang pandang pengamatan. Pengamatan dilakukan terhadap persentasi kemampuan protoplas beregenerasi membentuk dinding sel dan koloni sel setiap bidang pandang.
Pengenceran kultur
Pengenceran media dilakukan dengan tujuan untuk mendorong pertumbuhan dan perkembangan sel membentuk koloni sel dan mikrokalus. Pengenceran dilakukan dengan cara membagi 3 suspensi protoplas/sel setiap cawan petri ke cawan petri baru yang telah berisi media dasar yang sama (KM, VKM, MW dan MT), tetapi komposisi zat pengatur tumbuh yang ditambahkan hanya 3 mg/l BA (hasil penelitian 1). Kultur diinkubasi kembali pada ruang inkubasi yang diberikan cahaya dengan intensitas 1000 lux selama 16 jam sampai terbentuk mikrokalus dan kalus. Pengamatan dilakukan terhadap banyaknya rata-rata jumlah mikrokalus setiap minggu setelah pengenceran, tipe kalus yang dihasilkan, dan rata-rata jumlah embrio somatik yang dihasilkan.
Regenerasi tunas
Regenerasi tunas dilakukan dengan cara memindahkan struktur embrio somatik yang terbentuk secara langsung dalam media pengenceran. Struktur embrio somatik yang diperoleh dipindahkan pada media baru yang dipadatkan dengan penambahan 0.5 mg/l ABA pada media MW untuk pendewasaan dan 0.5 mg/l GA3
Setiap botol di masukkan 8 embrio somatik fase globular ke dalam setiap botol kultur dan diulang sebanyak 5 kali sehingga diperoleh 40 botol kultur dari semua perlakuan. Semua kultur disimpan di ruang kultur dengan penyinaran dengan intensitas 1000 lux selama 16 jam dengan suhu 23 - 27
untuk perkecambahan. Pendewasaan embrio somatik dilakukan dengan cara memindahkan struktur globuler yang diperoleh ke media pendewasaan strukruktur globular menjadi fase hati dan torpedo.
0
C. Pengecambahan embrio somatik dewasa dilakukan dengan cara memindahkan embrio somatik dewasa ke media perkecambahan. Pengamatan dilakukan terhadap persentase keberhasilan perkembangan embrio somatik fase globuler menjadi fase hati, torpedo dan kotiledon serta perkecambahan embrio somatik menjadi individu baru yang lengkap mempunyai tunas dan akar (plantlet). Untuk mempercepat pertumbuhan dan perkembangan regeneran (klon), setiap embrio somatik diisolasi dan dipindahkan
dalam botol kultur yang berisi media MW tanpa penambahan zat pengatur tumbuh untuk mendorong pertumuhan dan perkembangannya sehingga lebih sempurna.
Hasil dan Pembahasan Isolasi protoplas
Jenis sumber protoplas, komposisi larutan enzim, lama inkubasi dalam larutan enzim dan larutan purifikasi yang digunakan dalam isolasi protoplas merupakan faktor yang berpengaruh terhadap jumlah dan viabilitas protoplas yang dihasilkan. Metode isolasi protoplas yang digunakan pada penelitian ini adalah metode yang terbaik dari hasil penelitian sebelumnya. Berdasarkan hasil penelitin sebelumnya, kombinasi enzim selulase Onozuka R10-Yakult 1% dan maserozim R10-Yakult 1% dalam larutan CPW terbukti baik digunakan untuk mengisolasi protoplas dari kalus dan daun tanaman jeruk (hasil penelitian 2). Inkubasi dalam larutan enzim juga digunakan selama 16 jam dalam keadaan gelap dan dimurnikan dengan campuran manitol 13% dan sukrosa 25% untuk mengapungkan protoplas (hasil penelitian 3). Dari hasil isolasi protoplas yang dilakukan menggunakan metodologi dari hasil penelitian sebelumnya diperoleh bahwa rata-rata jumlah protoplas yang dihasilkan juga cukup tingi (Tabel 17). Berdasarkan tabel tersebut dapat dilihat bahwa densitas protoplas yang dihasilkan dari kalus embriogenik lebih banyak dari pada protoplas yang dihasilkan dari daun in vitro. Hal ini disebabkan oleh adanya lignin di jaringan daun sehingga aktifitas enzim kelompok pektinase seperti meserozim lebih sulit mendegradasi zat pektin yang berada diantara sel yang satu dengan sel lainnya sehingga sel yang satu dengan sel lainnya menjadi terpisah (sel tungga l). Rata-rata Tabel 17. Produksi protoplas dari kalus dan mesofil daun menggunakan kombinasi
enzim Selualse 1%+ Maserozim 1% yang diinkubasi selama 16 jam dan dimurnikan dengan campuran 25% sukrosa+13% manitol dalam larutan CPW.
Sumber protoplas Rata-rata jumlah protoplas
(protoplas/g sumber protoplas) Kalus embriogenik Mesofil daun 15.7x105 13.0x10 ± 8.0 5 ± 9.8
.
Gambar 20. Isolasi protoplas mesofil daun mandarin satsuma dan kalus jeruk siam simadu dengan kombinasi enzim selulase 1%+maserozim 1% yang dimurnikan dengan larutan sukrosa 25% + manitol 13% (A dan C= mesofil daun dan protoplas yang dihasilkan, B dan D=kalus embriogenik dan protoplas yang dihasilkan) perbesaran 20x.
jumlah protoplas yang dihasilkan dari kalus embriogenik adalah sebanyak 15.7x105 protoplas/g kalus dan 13.0x105
Selain itu, dari gambar protoplas tersebut juga terlihat adanya perbedaan kepadatan isi protoplas yang mencirikan sel tua dan muda. Protoplas yang berasal dari sel muda mempunyai vakuola yang lebih kecil daripada vakuola sel dewasa. Besar kecilnya vakuola sel yang dihasilkan juga akan berpengaruh terhadap terjadinya fusi. Protoplas yang megandung vakuola yang besar akan lebih sulit berfusi sehingga jumlah protoplas hasil fusi yang dihasilkan juga berbeda.
protoplas/g daun yang dihasilkan dari mesofil daun. Protoplas yang dihasilkan dari mesofil daun berwarna kehijauan karena adanya klorofil sedangkan protoplas yang berasal dari kalus tidak berwarna hijau karena kalus belum mengandung klorofil (Gambar 20). Viabilitasnya juga sangat baik yang ditandai dengan bentuk protoplasnya berbentuk bulat sempurna.
C D
Fusi protoplas
Keberhasilan dalam menginduksi terjadinya fusi protoplas sangat tergantung dari kadar konsentrasi dan periode inkubasi yang digunakan dalam larutan PEG. Konsentrasi dan lama inkubasi yang digunakan dalam penelitian ini untuk memacu terjadinya fusi adalah PEG 4% dan 30% yang diberikan pada 4 titik yang berlawanan disekitar suspensi protoplas dengan waktu inkubasi dalam larutan PEG selama 15 menit (hasil penelitian 3).
Dari hasil penelitian tersebut diperoleh bahwa rata-rata jumlah protoplas berfusi yang dihasilkan lebih banyak dari induksi fusi dengan PEG 30% dari pada induksi fusi dengan PEG 4% (Gambar 21). Hal ini sesuai dengan hasil penelitian sebelumnya, dimana rata-rata jumlah protoplas berfusi yang dihasilkan dari induksi fusi dengn PEG 30% lebih banyak dari rata-rata jumlah protoplas berfusi dari induk si PEG 4%. Rata-rata jumlah protoplas berfusi yang dihasilkan dari induksi fusi dengan PEG 4% adalah 3.3 fusan yang hetero fusi, 5 fusan yang homo fusi dan multi fusi. Sedangkan rata-rata jumlah protoplas berfusi yang dihasilkan dari induksi fusi dengan PEG 30% adalah 4.7 fusan yang hetero fusi, 6.7 fusan yang homo fusi, dan 7.7 fusan. Penambahan larutan pencuci (0.5 M manitol+0.5 mM CaCl2
3,3 5 5 4,7 6,7 7,7 0 1 2 3 4 5 6 7 8 Ju m la h p ro to pl as berf us i 4% 30% Konsentrasi PEG Hetero fusi Homo fusi Multi fusi ) setelah 15 menit induksi fusi juga dapat meningkatkan frekuwensi fusi baik dari induksi fusi dengan PEG 4% maupun induksi fusi dengan PEG 30% kecuali multi fusi dari PEG 30%
Gambar 21. Pengaruh konsentrasi PEG (4% dan 30%) terhadap keberhasilan fusi (hetero, homo, dan multi fusi) protoplas jeruk siam Simadu dengan mandarin Satsuma, inkubasi 15 menit
4,3 6,7 7,7 5,7 7,7 6,3 0 1 2 3 4 5 6 7 8 Jum la h pr ot opl as be rf us i 4% 30% Konsentrasi PEG Hetero fusi Homo fusi Multi fusi
Gambar 22. Peningkatan jumlah protoplas berfusi setelah penambahan larutan pencuci ke dalam suspensi protoplas yang telah difusi selama 15 menit dengan PEG 4 dan 30%.
(Gambar 22). Adanya peningkatan frekuwensi fusi tersebut disebabkan oleh adanya CaCl2 dalam larutan pencuci yang digunakan. Frekuensi fusi protoplas dapat meningkat apabila pada suspensi protoplas yang difusikan dengan PEG di cuci dengan larutan pencuci atau hipotonik. Adanya ion Ca2+
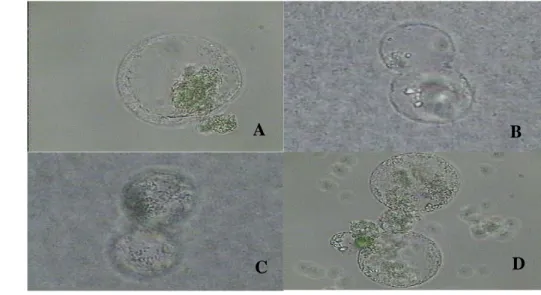
Penampakan protoplas fusi yang hetero fusi, homo fusi dan multi fusi jelas terlihat karena adanya perbedan warna protoplas yang digunakan (Gambar 23). Berdasarkan gambar 23 jelas terlihat bahwa protoplas yang berfusi dari dua jenis protoplas yang berbeda (hetero fusi) terlihat adanya penyatuan dari dua jenis protoplas yang berwarna hijau dengan yang tidak berwarna yang berasal dari
tinggi dalam larutan hipotonik juga dapat meningkatkan frekuensi fusi antar protoplas (Kao dan Michayluk 1975; Veilleux et al. 2005). Dengan bertambahnya frekuwensi fusi yang diperoleh akan memberikan peluang lebih tinggi untuk mendapatkan hibrida somatik yang dihasilkan setelah dikulturkan. Rata-rata jumlah protoplas berfusi yang dihasilkan setelah penambahan larutan pencuci dari induksi fusi dengan PEG 4% adalah menjadi 4.3 fusan yang hetero fusi, 6.7 yang homo fusi, dan 7.7 yang multi fusi. Sedangkan rata-rata jumlah protoplas berfusi yang dihasilkan dari induksi fusi dengan PEG 30% adalah menjadi 5.7 fusan yang hetero fusi, 7.7 fusan yang homo fusi, dan 6.3 fusan yang multi fusi.
Gambar 23. Penampakan keadaan suspensi protoplas pada saat penambahan PEG (A) dan jenis fusi yang dihasilkan dari protoplas yang diinduksi dengan PEG selama 15 menit (B =hetero fusi, C=homo fusi, D=multi fusi, dan E= total fusi).
protoplas mesofil daun dan kalus. Demikian juga halnya yang homo fusi memperlihatkan penggabungan dari dua protoplas yang berwarna hijau yang berasal dari dua protoplas mesofil daun. Multi fusi ditunjukkan oleh penggabungan tiga protoplas berwarna hijau yang berasal dari mesofil daun. Dari tipe hasil fusi yang dihasilkan juga akan menghasilkan regeneran yang mempunyai variasi genetik yang berbeda antara satu dengan yang lainnya.
Regenerasi protoplas hasil fusi
Komposisi zat pengatur tumbuh yang digunakan dalam medium merupakan faktor sangat penting dalam pertumbuhan dan perkembangan protoplas. Protoplas yang ditanaman di dalam media kultur akan membentuk diding sel sehingga membentuk sel yang sempurna yang dapat membelah dan memperbanyak diri
A B
C D
PEG
sehingga terbentuk koloni sel. Tahap pertama kesuksesan di dalam kultur protoplas adalah keberhasilan protoplas membentuk dinding sel yang baru dan keberhasilan protoplas atau sel yang baru melakukan pembelahan mitosis. Keberhasilan regenerasi dinding sel tanaman dari protoplas merupakan tahap yang sangat sulit dan sering mendapatkan hasil yang berbeda jika diulang pada waktu dan tempat yang berbeda (Evan dan Bravo 1983). Biosintesis dari dinding sel bervariasi tergantung material tanaman yang digunakan sebagai sumber protoplas. Sintesis dinding sel protoplas memerlukan adanya penyediaan sumber karbon yang mudah dimetabolisasikan seperti sukrosa dan penambahan osmolytikum dalam media kultur (Carlson et al. 1995).
Gautheret (1977) mengatakan bahwa dinding sel protoplas dapat terbentuk apabila enzim yang melarutkan dinding sel pada saat isolasi protoplas harus dihilangkan dengan cara mencucinya dengan larutan pencuci sampai hilang. Dalam media kultur perlu ditambahkan suatu osmotic stabilizer atau zat anti pecah karena protoplas belum mempunyai dinding sel. Untuk pembentukan dinding sel biasanya ditambahkan manitol atau sukrosa secukupnya. Pembentukan dinding sel protoplas juga ditentukan oleh adanya auksin dan sitokinin yang mempengaruhi pembentukan permukaan dinding.
Protoplas biasanya dikulturkan dalam medium cair pada erlenmeyer atau cawan petri dalam jumlah sedikit dalam bentuk tetesan (Bawa dan Torrey 1971; Constabel 1982). Umumnya untuk merangsang pembelahan, protoplas harus ditanam dalam kerapatan tidak lebih dari 104
Hasil kultur protoplas yang dilakukan setelah difusi denga PEG 4 % dan 30% selama 15 menit memperlihatkan bahwa protoplas dapat melakukan regenerasi dinding sel pada minggu pertama pengamatan setelah kultur pada semua media kultur yang digunakan (Tabel 18). Hal ini diduga disebabkan karena jumlah sukroksa yang
protoplas/ml medium (Albersheim 1974). Selain itu, faktor ada tidaknya cahaya selama kultur juga sangat mempengaruhi keberhasilan regenerasi dinding sel sehingga kultur protoplas yang telah difusikan disimpan dalam keadaan gelap dalam kaleng steril yang tertutup.
Tabel 18. Pengaruh media kultur terhadap persentase kemampuan protoplas membentuk dinding dan pembelahan sel yang diinduksi fusi denga PEG 4 dan 30%, 2 minggu setelah kultur.
Media kultur Protoplas berdinding setiap bidang pandang (%)
Protoplas membelah setiap bidang pandang (%) PEG 4% PEG 30% PEG 4% PEG 30% KM VKM MT MW 12.2 11.7 21.8 21.1 7.8 5.3 11.6 16.7 6.1 5.8 15.0 15.0 0.0 0.0 3.0 5.9 Keterangan: KM= Kao dan Michayluk (1975) +0.1 mg/l 2,4_D+3 mg/l BA+500 mg/l EM+50 g/l Sukrosa,
VKM= Binding dan Nels (1977)+0.1 mg/l 2,4_D+3 mg/l BA+500 mg/l EM+50 g/l sukrosa, MT= Murashige and Tucker (1969)+0.1 mg/l 2,4_D+3 mg/l BA+500 mg/l EM+50 g/l sukrosa, MW= Morel and Wetmore (1951) )+0.1 mg/l 2,4_D+3 mg/l BA+500 mg/l EM+50 g/l sukrosa
diberikan pada semua media kultur yang digunakan sebagai sumber karbon yang mudah dimetabolisasikan adalah sama, yaitu sebanyak 5%. Gautheret (1977) mengatakan bahwa untuk sintesis dinding sel perlu adanya penyediaan sumber karbon yang mudah dimetabolisasikan seperti sukrosa. Adanya sukrosa yang tinggi dalam media juga dapat mencegah penurunan pH pada media kultur sehingga tidak mengganggu pertumbuhan sel pada awal kultur (Kowalczyk et al. 1983). Proses regenerasi dinding sel protoplas dimulai dalam beberapa jam sampai beberapa hari untuk melengkapinya. Proses regenerasi dimulai dari adanya akumulasi bahan dinding sel secara perlahan-lahan pada permukaan plasmalema dalam beberapa jam setelah kultur. Selulosa dikumpulkan diantara plasmalema dan antara bahan dinding multilamelar atau langsung pada plasmalema (media kultur mulai meregenerasikan dinding (Bawa dan Torrey 1971). Selain itu, kondisi fisik kultur dan zat pengatur tumbuh yang digunakan juga sama, yaitu media cair yang disimpan dalam keadaan gelap dengan penambahan 0.1 mg/l 2,-D + 3 mg/l BA dan 500 mg/l EM. Constabel (1982) menyarankan untuk budidaya protoplas hasil fusi lebih baik menggunakan media cair dengan kondisi gelap (tanpa cahaya) pada temperatur 25 – 28 0
Media MT dan MW merupakan media kultur yang baik digunakan untuk meregenerasi dinding sel. Persentase keberhasilan regenerasi protoplas membentuk
dinding sel per bidang pandang paling baik berasal dari media MT dengan keberhasilan regenerasi sebanyak 21.8% dari induksi fusi dengan PEG 4% dan 11.6% dari induksi fusi dengan PEG 30%. Kemudian diikuti oleh media MW sebanyak 21.1% dari induksi fusi dengan PEG 4% dan 16.7% dari induksi fusi dengan PEG 30%, KM sebanyak 12.2% dari induksi fusi dengan PEG 4% dan 7.8% dari induksi fusi dengan PEG 30%, dan VKM sebanyak 11.7% dari induksi fusi dengan PEG 4% dan 5.3% dengan PEG 30%.
Kemampuan keberhasilan menginduksi pembelahan sel yang terbaik juga terjadi pada media MT dan MW. Hal ini dapat terlihat dari besarnya persentase kemampuan sel baru melakukan pembelahan mitosis sebesar 15% pada kedua media tersebut baik pada kultur hasil induksi fusi dengan PEG 4%. Kemampuan pembelahan sel pada budidaya protoplas ditentukan oleh komposisi media kultur dan kondisi eksplan yang digunakan sebagai sumber protoplas. Reiner dalam Gautheret (1982) melaporkan bahwa sel yang berasal dari protoplas baru dapat membelah setelah 32 hari. Kegagalan mitosisi tersebut mungkin disebabkan olaeh kurangnya nutrisi dalam medium. Besaran kemampuan pembelahan sel pada media MT dan MW pada kultur yang diinduksi dengan PEG 4% adalah sebesar 15% kemudian diikuti media KM sebesar 6.1% dan VKM sebesar 5.8% pada kultur yang diinduksi dengan PEG 4%. Pembelahan sel pada kultur yang diinduksi fusi dengan PEG 30% hanya terjadi pada media MT (3.0%) dan MW (5.9%) saja. Terbentuknya dinding sel protoplas secara sempurna ditandai dengan bentuk protoplas yang tidak bulat lagi dan disekeliling sel protoplas terlihat lebih tebal (Gambar 24A). Proses awal pembelahan mitosis sel ditandai dengan adanya dua bakal sel hasil pembelahan yang belum sempurna dan terbentuknya dua sel pada akhir pembelahan sehingga terbentuk sel-sel baru yang sempurna (24B, 24C dan 24D).
Untuk mempertebal dinding dan mempercepat pembelahan sel, tutup kaleng tempat inkubasi kultur setelah dua minggu di buka sehingga kultur dapat menyerap cahaya. Sihachakr (1998) mengatakan bahwa pemberian cahaya pada sel protoplas yang telah membentuk dinding sel setelah dikulturkan dapat mempercepat penebalan dinding sel dan mampu mempercepat terjadinya pembelahan sel. Hasil pengamatan
Gambar 24. Pembentukan dinding sel dan proses pembelahan sel (A=protoplas yang telah meregenerasi dinding selnya secara sempurna, B= awal pembelahan sel, C= akhir pembelahan sel dan D= sel baru hasil pembelahan).
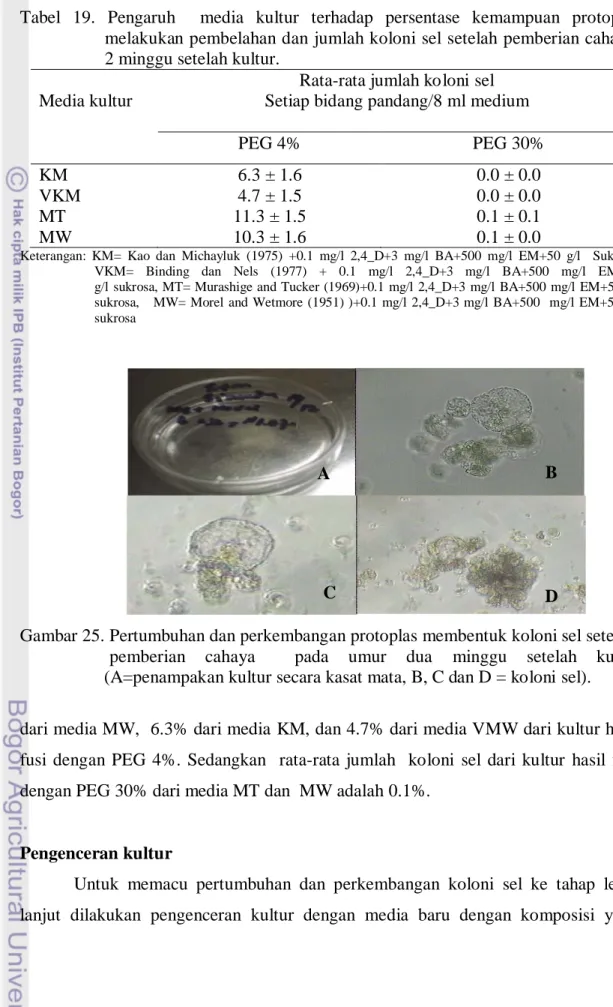
dua minggu setelah pemberian cahaya diperoleh adanya pertumbuhan dan perkembangan sel (4 minggu setelah kultur) pada semua media yang digunakan pada kultur hasil fusi dengan PEG 4%, sedangakan kultur hasil fusi dengan PEG 30% hanya pada media MT dan MW saja (Tabel 19). Hal ini jelas terlihat dari penampakan media kultur yang semakin keruh yang dipenuhi dengan noktah kecil tersebar dalam media dengan warna putih (Gambar 25A). Berdasarkan pengamatan yang dilakukan secara mikroskopik ternyata noktah-noktah tersebut adalah koloni-koloni sel dari hasil pembelahan sel yang terus-menerus yang ditandai dengan adanya sel-sel muda baru disekitarnya dengan ukuran lebih kecil (Gambar 25 B, C, dan D). Koloni sel tersebut ditandai dengan adanya sel-sel baru di sekitar sel protoplas yang mempunyai plasma yang penuh, inti besar, vacuola kecil dan ukurannya lebih kecil. Banyaknya rata-rata jumlah koloni sel yang dihasilkan per setiap bidang pandang pengamatan dari media MT dan MW berbeda dibandingkan media KM dan VMW. Rata-rata jumlah koloni sel yang dihasilkan dari media MT adalah 11.3% dan 10.3%
A B
Tabel 19. Pengaruh media kultur terhadap persentase kemampuan protoplas melakukan pembelahan dan jumlah koloni sel setelah pemberian cahaya, 2 minggu setelah kultur.
Media kultur
Rata-rata jumlah koloni sel Setiap bidang pandang/8 ml medium
PEG 4% PEG 30% KM VKM MT MW 6.3 ± 1.6 4.7 ± 1.5 11.3 ± 1.5 10.3 ± 1.6 0.0 ± 0.0 0.0 ± 0.0 0.1 ± 0.1 0.1 ± 0.0
Keterangan: KM= Kao dan Michayluk (1975) +0.1 mg/l 2,4_D+3 mg/l BA+500 mg/l EM+50 g/l Sukrosa,
VKM= Binding dan Nels (1977) + 0.1 mg/l 2,4_D+3 mg/l BA+500 mg/l EM+50 g/l sukrosa, MT= Murashige and Tucker (1969)+0.1 mg/l 2,4_D+3 mg/l BA+500 mg/l EM+50 g/l
sukrosa, MW= Morel and Wetmore (1951) )+0.1 mg/l 2,4_D+3 mg/l BA+500 mg/l EM+50 g/l sukrosa
Gambar 25. Pertumbuhan dan perkembangan protoplas membentuk koloni sel setelah pemberian cahaya pada umur dua minggu setelah kultur
(A=penampakan kultur secara kasat mata, B, C dan D = koloni sel). dari media MW, 6.3% dari media KM, dan 4.7% dari media VMW dari kultur hasil fusi dengan PEG 4%. Sedangkan rata-rata jumlah koloni sel dari kultur hasil fusi dengan PEG 30% dari media MT dan MW adalah 0.1%.
Pengenceran kultur
Untuk memacu pertumbuhan dan perkembangan koloni sel ke tahap lebih lanjut dilakukan pengenceran kultur dengan media baru dengan komposisi yang
A B
sama. Pengenceran kultur suspensi koloni sel dilakukan dengan cara setiap kultur dari cawan petri dipipet dan dimasukkan ke dalam 3 cawan petri baru (± 2 ml) yang telah berisi 6 ml media. Cara tersebut telah barhasil dilakukan untuk mendorong pertumbuhan dan perkembangan koloni sel menjadi mikro kalus dan kalus pada tanaman kentang (Sihachakr 1998; Purwito 1999) dan tanaman terung (Sihachakr et al. 1999; Husni et al. 2003).
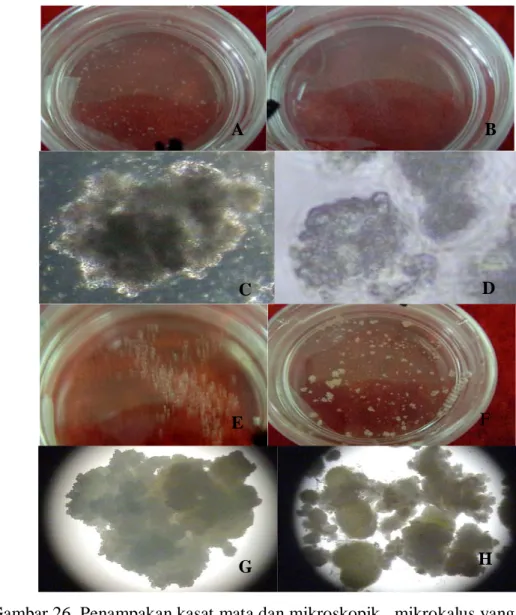
Satu minggu setelah pengenceran koloni sel belum terlihat adanya mikrokalus yang terbentuk, akan tetapi jumlah koloni sel yang terlihat semakin banyak. Mikro kalus mulai muncul pada pengamatan minggu ke-2 hanya diperoleh dari kultur yang difusi dengan PEG 4% saja, sedangkan kultur hasil fusi dengan PEG 30% tidak menghasilkan mikrokalus. Hal ini memeperlihatkan bahwa konsentrasi PEG yang digunakan untuk induksi fuasi berpengaruh terhadap kemampuan regenerasi. Mikro kalus yang terbentuk dapat terlihat secara jelas dengan kasat mata berwarna putih susu (Gambar 26). Kecepatan pertumbuhan dan perkembangan koloni sel membentuk mikrokalus dipengaruhi oleh media dasar dan konsentrasi PEG yang digunakan untuk induksi fusi. Hal ini terbukti dari mikro kalus yang terbentuk hanya diperoleh dari protoplas hasil fusi dengan PEG 4% pada media MT dan MW saja pada minggu ke-2, sedangkan pada media KM dan VKW belum ada.
Sampai akhir pengamatan (4 minggu setelah pengenceran) belum diperoleh adanya mikro kalus pada kultur hasil fusi degan PEG 30%, sedangkan mikrokalus yang terbentuk dari kultur hasil fusi dengan PEG 4% semakin banyak (Tabel 20). Dari tabel 20 dapat dilihat bahwa semakin lama umur kultur semakin padat jumlah mikrokalus yang terbentuk pada kultur hasil fusi dengan PEG 4%, sedangkan kultur hasil fusi dengan PEG 30% tidak menghasilkan mikrokalus sampai akhir pengamatan.
Gambar 26. Penampakan kasat mata dan mikroskopik mikrokalus yang terbentuk pada media kultur KM, VKM, MT dan MW 4 minggu setelah pengenceran (A dan C= mikro kalus pada media KM, B dan D= mikro kalus pada media VKM, E dan G= mikro kalus pada media MT serta F dan H= mikro kalus pada media MW).
Rata-rata jumlah mikrokalus yang dihasilkan tidak berbeda nyata antara media MT dan MW, akan tetapi berbeda nyata dengan media KM dan VKM. Mikro kalus yang terbentuk pada minggu ke-2 hanya terdapat pada media MT dan MW saja dengan rata-rata 4.3 dan 4.7. Mikrokalus kalus mulai terbentuk pada media KM dan VKM setelah 3 minggu pengenceran. Rata-rata jumlah mikrokalus yang dihasilkan pada
A B
C D
E F
H G
Tabel 20. Pengaruh media kultur terhadap kemampuan koloni sel membentuk mikrokalus (1, 2, dan 3 minggu setelah pengenceran).
Media kultur
Rata-rata jumlah mikrokalus/8 ml medium
Minggu ke-2 Minggu ke-3 Minggu ke-4
KM VKM MT MW 0 0 4.3 ± 1.2 4.7± 1.5 2.3 ± 1.5 1.0 ± 1.0 15.3 ± 3.1 18.7 ± 1.5 5.7± 2.3 4.3 ± 0.6 23.7± 4.0 28.3± 2.5
Keterangan: KM= Kao dan Michayluk (1975) +0.1 mg/l 2,4_D+3 mg/l BA+500 mg/l EM+50 g/l Sukrosa, VKM= Binding dan Nels (1977)+0.1 mg/l 2,4_D+3 mg/l BA+500 mg/l EM+50 g/l sukrosa, MT= Murashige and Tucker (1969)+0.1 mg/l 2,4_D+3 mg/l BA+500 mg/l EM+50 g/l sukrosa, MW= Morel and Wetmore (1951) )+0.1 mg/l 2,4_D+3 mg/l BA+500 mg/l EM+50 g/l sukrosa.
media KM dan VKM adalah 2.3 dan 1 mikrokalus serta 15.3 pada media MT dan 18.7 pada media MW.
Bila dilihat dari tipe mikrokalus yang dihasilkan pada pengamatan minggu ke 4, mikrokalus yang berasal dari media KM dan VKM berwarna coklat kehitaman. Hal ini mengindikasikan bahwa viabilitas dari mikrokalus tersebut menurun sehingga kalus yang dihasilkan bersifat tidak embriogenik (4 minggu setelah kultur) jumlah mikrokalus terus bertambah banyak pada seluruh media yang digunakan (Tabel 21). Rata-rata embrio somatik yang dihasilkan adalah 5.7 pada media KM, 4.3 pada media VKM, 23.7 pada media MT dan 28.3 pada media MW. Pada media kultur MW, Tabe 21. Pengaruh media kultur terhadap kemampuan koloni sel membentuk mikro
kalus dan embrio somatik, 4 minggu setelah pengenceran. Media
kultur
Rata-rata mikro kalus/8 ml
medium
Rata-rata embrio /8 ml medium
Fase globular Fase torpedo Fase hati KM VKM MT MW 5.7 ± 2.3 4.3 ± 0.6 23.7 ± 4.0 28.3 ± 2.5 0 0 0 7.0 (42/6) 0 0 0 3.0 (18/6) 0 0 0 3.5 (21/6) Keterangan: KM= Kao dan Michayluk (1975) +0.1 mg/l 2,4_D+3 mg/l BA+500 mg/l EM+50 g/l Sukrosa,
VKM= Binding dan Nels (1977)+0.1 mg/l 2,4_D+3 mg/l BA+500 mg/l EM+50 g/l sukrosa, MT= Murashige and Tucker (1969)+0.1 mg/l 2,4_D+3 mg/l BA+500 mg/l EM+50 g/l sukrosa, MW= Morel and Wetmore (1951) )+0.1 mg/l 2,4_D+3 mg/l BA+500 mg/l EM+50 g/l sukrosa.
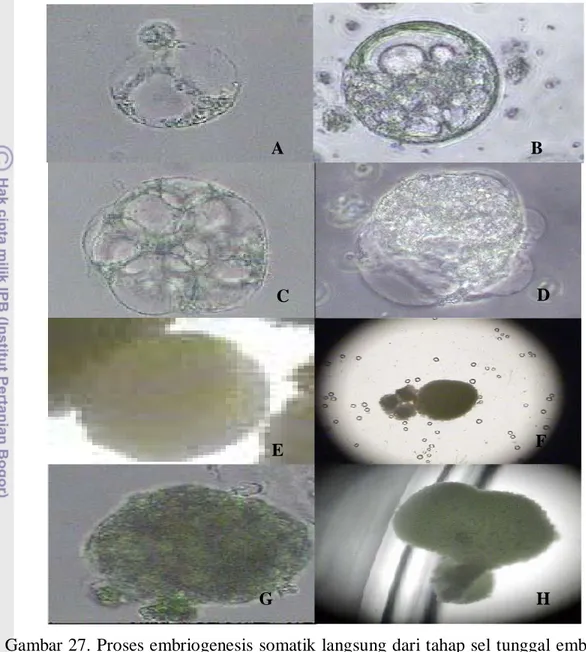
Gambar 27. Proses embriogenesis somatik langsung dari tahap sel tunggal embryoid hasil fusi antara jeruk siam Simadu dengan mandarin Satsuma sampai menjadi plantlet pada media MW (. A dan B= Sel embrioid yang aktif membelah, C dan D= Pre-embrio (pem), E= embrio fase globuler, F=embrio fase torpedo, G dan H = embrio fase hati.
selain mikro kalus juga diperoleh adanya embrio somatik fase globular, torpedo, dan hati. Adanya embrio somatik pada media MW mengindikasikan bahwa telah terjadi proses embriogenesis langsung dari sel-sel embrioid (Gambar 27). Rata-rata jumlah embrio somatik fase globular adalah sebanyak 7 embrio, fase torpedo sebanyak 3 embrio, dan fase hati hati sebanyak 3.5 embrio.
A B
C D
E F
Embriogenesis somatik mempunyai potensi morfogenik yang tinggi membentuk individu baru dari hasil proses embriogenesis dari sel somatik yang bersifat embrioid (von Arnold et al. 2002). Terjadinya proses embriogenesis somatik secara langsung pada penelitian ini antara lain disebabkan oleh salah satu sumber protoplas yang digunakan adalah kalus embriogenik dan komposisi media yang digunakan juga sangat mendukung. Embriogenesis somatik secara langsung terjadi dari sel-sel tunggal yang meristemoid yang bersifat embrioid (Gambar 27A) yang aktif membelah secara terus-menerus sehingga selnya bertambah banyak (Gambar 27B ), tumbuh dan berkembang membentuk bakal embrio/preembrio (Gambar 27C dan D), embrio somatik fase globular (Gambar 27 E), embrio somatik fase torpedo (27F), dan fase hati (Gambar 27G dan H).
Pendewasaan dan Perkecambahan embrio somatik
Berdasarkan pertumbuhan dan perkembangan 42 embrio somatik tahap globular yang dipindahkan pada media MW baru (padat) dengan penambahan 0.5 mg/l ABA diperoleh bahwa media tersebut dapat mendorong pertumbuhan dan perkembangan struktur globular menjadi tahap lebih lanjut (fase torpedo dan hati). Semakin lama umur kultur semakin banyak pula embrio somatik yang tumbuh dan berkembang ke fase torpedo dan hati (Tabel 22). Setelah dua minggu pemindahan, jumlah globular yang tumbuh dan berkembang ke fase torpedo dan hati tidak terlalu menyolok. Jumlah embrio somatik fase torpedo sebanyak 12 dan fase hati dan kotiledon sebanyak 6 embrio. Akan tetapi, perubahan tersebut sangat menyolok pada Tabel 22. Pendewasaan embrio somatik fase globular menjadi fase torpedo dan hati
dalam media MW + 1.5 mg/l ABA + 500 mg/l EM umur 2 dan 4 minggu. Fase embrio somatik Jumlah embrio somatik minggu ke - Keterangan 2 4 Globular Torpedo
Hati dan kotiledon
24 12 6 32 19 25
Kultur membentuk kalus embriogenik dan embrio somatik sekunder disekitar kultur dengan persentase sebesar
Gambar 28. Proses pendewasaan dalam media MW+0.5 mg/l ABA dan perkecambahan embrio somatik dalam media MW + 0.5 mg/l GA3 menjadi plantlet (A= embrio somatik langsung dari hasil fusi, B dan C= pembentukan embrio somatik sekunder pada media pendewasaan, D dan E= perkecambahan embrio somatik, dan F= plantlet dalam media MW).
pengamatan minggu ke empat dalam media pendewasaan. Bahkan, banyak kultur yang membentuk kalus embriogenik (Gambar 28B) dan menghasilkan embrio somatik sekunder (Gambar 28C) sehingga jumlahnya bertambah banyak (76 embrio). Jumlah embrio somatik fase globular menjadi 32, fase torpedo menjadi 19 dan fase hati dan kotiledon menjadi 25. Dari 25 embrio somatik dewasa yang dikecambahkan dalam media yang sama dengan mengganti ABA dengan 0.5 mg GA3
A
diperoleh bahwa media tersebut dapat menginduksi pertumbuhan dan perkembangannya lebih lanjut. Kemampuan menginduksi perkecambahan yang diperoleh menghasilkan plantlet sebanyak 19 kecambah menjadi menjadi benih somatik dengan efisisensi
B
C D
perkecambahan sebesar 76%. Setiap individu fusan yang berkecambah dipindakan ke media MW tanpa zat pengatur tumbuh sehingga terbentuk plantlet (Gambar 28F). Proses pendewasaan dan perkecambahan embrio somatik dapat dilihat pada Gambar 28.
Simpulan
1. Kombinasi enzim selulase Onozuka Yakult 1% dengan maserozim R10-Yakult 1% dalam larutan CPW yang dimurnikan dengan campuran sukrosa 25% dengan manitol 13% dapat menghasilkan protoplas dengan densitas 15.7x105 protoplas/g kalus dan 13x105
2. Konsentrasi PEG yang digunakan untuk menginduksi fusi protoplas dari kalus jeruk siam Simadu dengan protoplas mesofil daun in vitro berpengaruh terhadap jumlah rata-rata protoplas berfusi yang dihasilkan. Rata-rata jumlah protoplas berfusi yang dihasilkan dari induksi fusi dengan PEG 4% adalah 3.3 fusan yang hetero fusi, 5 fusan yang homo fusi dan multi fusi. Sedangkan rata-rata jumlah protoplas berfusi yang dihasilkan dari induksi fusi dengan PEG 30% adalah 4.7 fusan yang hetero fusi, 6.7 fusan yang homo fusi, dan 7.7 fusan.
protoplas/g daun.
3. Frekuensi fusi meningkat menjadi 4.3 fusan yang hetero fusi, 6.7 yang homo fusi, dan 7.7 multi fusi yang diinduksi dengan PEG 4% serta menjadi 5.7 fusan yang hetero fusi, 7.7 fusan yang homo fusi, dan 6.3 fusan yang multi fusi yang diinduksi dengan PEG 30%.
4. Keberhasilan regenerasi protoplas hasil fusi pada media kultur yang digunakan dipengaruhi oleh konsentrasi PEG yang dipakai untuk menginduksi terjadinya fusi. Protoplas yang difusikan dengan PEG 4% dapat beregenerasi membentuk dinding sel, melakukan pembelahan sel, koloni sel, mikro kalus dan embrio somatik pada media, sedangkan protoplas yang difusikan dengan PEG 30% hanya dapat beregenerasi membentuk dinding sel dan pembelahan sel.
5. Media kultur yang baik digunakan untuk regenerasi adalah MW (Morel dan Wetmor) dan MT (Murashige dan skoog). Penambahan ABA 0.5 mg/l dalam media MW dapat mendewasakan embrio somatik dan GA3 0.5 mg/l dapat menginduksi perkecambahan embrio somatik menjadi plantlet dengan efisiensi sebesar 76%.
Daftar Pustaka
Albersheim P. 1974. Strucktur and growth of cell wall in culture. In: Street HE (eds). Tissue Culture and Plant Science. Pp379-404. Academic Press. London. Bawa SB, Torrey JG. 1971. Budding and nuclear division in cultured protoplast of
corn, Convolvulus and union. Botan Gaz 132: 240-245.
Cabasson CM, Luro F, Ollitrault O, Grosser JW. 2001. Non-random inheritance of mithocondrial genomes in Citrus hybrids froduced by protoplast fusion. Plant Cell rep 20:604-609.
Cai XD, Fu J, Deng XX, Guo WW. 2007. Production and molecular characterization of potential seedless cybrid plants between pollen steril Satsuma mandarin and two seedy Citrus cultivars. Plant Cell Tiss Organ Cult. 90:275-283.
Calixto MC, Filho FAAM, Mendes BMJ, Vieira MLC. 2004. Somatic hybridization between Citrus sinensis (L.) Osbeck and C. grandis (L.) Osbeck. Pesq. Agropec. Bras. 39(7):1-6.
Carlson C, Ollitrault P, Cote FX, Michaux-Ferriere N, Dambier D, Teisson C. 1995. Characteristics of citrus cell cultures during undifferentiated growth on sucrose and somatic embryogenesis on galactose. Physiologa Plantarum. Vol 93: 464-470.
Constabel, F., H. Koblitz, J.W. Kirkpatrick and S. Rambold. 1980. Fusion of cell sap vacuoles subsequent to protoplast fusion. Can. J. Bot. 58:1032-1034.
De Carvalho Costa MAP, Mendes BMJ, Filho FAAM. 2003. Somatic hybridization for improvement of citrus rootstock: production of five new combinations with potensial for improved disease resistance. Aust. J. Exp. Agr. 43: 1151-1156.
Evans DA, Bravo JE. 1983. Protoplasts isolation and culture. In Ammirato PV, Evans DA, Sharp WR, Yamada Y (eds.). Handbook of Plant Cell Culture. MacMillan Pub Co. New York, London.
Fu CH, Guo WW, Liu JH, Deng XX. 2003. Regeneration of Citrus sinensis + Clausena lansium intergeneric triploid ang tetraploid somatic hybrids and their molecular identification. In Vitro Cell Dev. Sci.20:251-255.
Gautheret RJ. 1977. La Culture des Tissus et des celluler des vegetaur p 158-165. Masson : Paris, New York, Barcelona, Milan.
Grosser JW and Gmitter FG Jr. 1990. Protoplast fusion and citrus improvement. Plant Breeding Reviews. Portland, V.8, p.339-374.
Grosser JW, Gmitter FG, Tusa N, Reforgiato G, Cucinotta. 1996. Further evidence of a cybridization requirement for plant regeneration from citrus leaf protoplast following somatic fusion. Plant Cell Rep. 15:672-676.
Grosser JW, Ollitrault P, Olivares-Fuster O. 2000. Somatic hybridization in Citrus: an effective tool to facilitate variety improvement. In Vitro Cell Dev Biol Plant 36:434-449.
Grosser JW, Gmitter FG. 2005. Application of somatic hybridization and cybridization in crop improvement, with citrus as a model. In vitro Cell Dev. Biol Plant 39:360-364.
Guo WW., Prassad D., Cheng YJ., Serrano P., Deng XX, and grosser.2004. Targeted cybridization in citrus: transfer of Satsuma cytoplasm to seedy cultivars for potential seedlessness. Plant Cell rep 22:752-758.
Husni A, Wattimena GA, Mariska I, Purwito A. 2003. Keragaman genetic tanaman terung hasil regenerasi protoplas. Jurnal bioteknologi Pertanian. 8(2):52-59. Husni A. 2007. Penerapan teknik fusi protoplas dalam dalam perakitan jeruk lokal
tipe baru. Laporan Akhir Penelitian Riset Insentif Terapan TA.2007.Kejasama BB-Biogen dengan Menristek. 33h.
Kao KN and Michayluh MR. 1975. Nutrition requirements for growth of Vicia hajastana cell and protoplast at a very low population density inliquid media. Planta. 125:105-110.
Kobayashi S, Ohgawara T, Ohgawara E, Oiyima I, Ishii IS.1988. A somatic hybirid plant obtained by protoplast fusion between navel orange (Citrus sinensis) and Satsuma mandarin. Plant Cell Tissue and Organ Culture 14:63-69.
Kowalezyk TP, Mackenzie IA, andCocking C. 1983. Plant regeneration from organ explants and protoplasts of medicinal plant Solanum khasianum CB clarke
var. Chatterjeeanum Sengupta (syn Solanum viarum Dunal). Z Pflanzenphysiol 111: 55-68.
Kunitake H, Kagami H, Mii M. 1991. Somatic embrtogenesis and plant regeneration from protoplasts of Stsuma?mandarin (Citrus unshiu Marc.) Scientia Horticilturae, 47:27-33.
Kuntarsih S. 2007. Pengelolaan rantai pasok dengan bisnis jeruk (kasus jeruk siam Pontianak Kabupaten Sambas). Prosiding Seminar Nasional jeruk. Yogyakarta, 13-14 Juni 2007.
Moriguchi T, Hidaka T, Omura M, Motomura T, and Akihama T. 1996. Genotypes and parental combination influence efficiency of cybrid induction in citrus by electrofusion. Hort Science 31:275-278.
Ohgawara T, Kobayashi S, Ohgawara E, Uchi miya H, Ishii S. 1985. Somatic hybrids plants obtained by protoplast fusion between (Citrus sinensis and Poncirus tripoliata). Theor Appl Genet. 71: 1-4.
Ollitrault P, Dambier D, and Luro F. 1996. Somatic hybridization in Citrus; some new hybrids and alloplasmic plants. Proc. Int. Soc. Citricult.2:907-912. Purwito A. 1999. Fusi protoplas intra dan interspesies pada tanaman kentang.
Disertasi Program Pasca Sarjana. IPB. Bogor.
Sihachakr D. 1998. Culture Media and Protocols for Isolation and Fusion of Prtoplasts of Eggplant. Universite Paris sud, France (Tidak dipublikasi). Spiegel-Roy P, Goldschmidt EE. 1996. Biology Of Citrus. Cambridge University
Press. 221 p.
Tusa N, Patta del Bosco S, Nigro F, and Ippolito A. 2000. Response of cybrids and a somatic hybrid of lemon to Phoma tracheiphila infections. HortScience 35:125-127.
Vardi A, Breiman A, Galun E. 1987. Citrus cybrids: production by donor-recipient protoplast fusion and verification by mitochondrial-DNA restriction profiles. Theor. Appl. Genet., 75:51-58.
Veilleux RE, Compton ME, and Saunders JA. 2005. Use of Protoplasts for Plant Improvement In R.N. Trigiano and D.J. Gray (Eds) Plant Development and Biotechnology.187-200pp. CRC Press LLC.
Von Arnold S, Sabala I, Bozhkov P, Kyachok J, Filonova L. (2002).Developmental pathway of somatic embryogenesis. Plant Cell Tissue Organ Cult 69:233-249.
Xu XY, Liu JH, Deng XX. 2006. Isolations of citoplats from Satsuma mandarin (Citrus unshiu Mrc.) and production of alloplasmic hybrid calluses via cytoplast-protoplsat fussion. Plant Cell rep. 25:533-539.
Yamamoto M, Kobayashi S. 1995. A cybrid plant produced by electrofusion between Citrus unshiu and C. Sinensis. Plant Tiss Cult Lett 12:131-137. Yamamoto M, Matsumoto R, Okudai N, and Yamada Y. 1997. Aborted anthers of