TINJAUAN PUSTAKA
Ekologi Avian Influenza
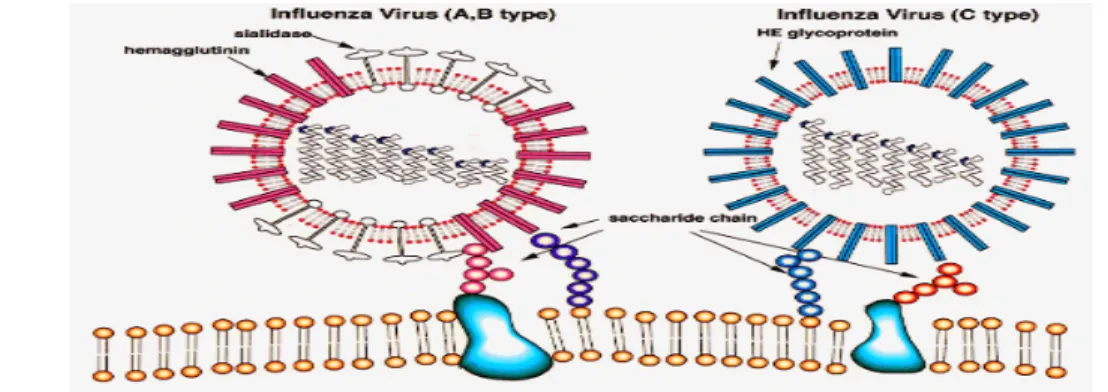
Virus influenza adalah partikel berselubung berbentuk bundar atau bulat panjang, merupakan genom RNA rantai tunggal dengan 8 segmen, serta berpolaritas negatif. Virus influenza termasuk famili Orthomyxoviridae dan diklasifikasikan dalam tipe A, B atau C berdasarkan perbedaan sifat antigenik dari nukleoprotein dan matrix proteinnya.
Gambar 1. Virus Influenza tipe A, B, C
Berdasarkan sifat antigenisitas glikoprotein, virus influenza dikelompokkan ke dalam 16 subtipe H (H1-H16) dan 9 subtipe N (N1-N9). Kelompok-kelompok tersebut ditetapkan berdasarkan analisis filogenetik terhadap nukleotida dan penetapan sekuen gen-gen HA dan NA melalui cara deduksi asam amino (Fouchier et al. 2005).
Hemaglutinin merupakan protein yang mengalami glikosilasi dan asilasi (glycosylated and acylated protein) terdiri dari 562-566 asam amino yang terikat dalam selubung virus. Bagian distal berbentuk bulat yang berkaitan dengan kemampuannya melekat pada reseptor sel. Hemaglutinin terdiri dari oligosakharida yang menyalurkan derivat asam neuroaminik (Watowich et al. 1994). Daerah eksternal (exodomain) dari glikoprotein transmembran merupakan neuroamidase
(NA), enzim bertugas melakukan aktivitas enzimatik sialolitik (sialolytic ensymatic activity) selain itu enzim neuraminidase bertugas melepaskan progeni virus yang terjebak di permukaan sel yang terinfeksi sewaktu dilepaskan. Fungsi ini mencegah bertumpuknya virus serta memudahkan gerakan virus dalam selaput lendir dari jaringan epitel sasaran (Matrosivich et al. 2004). Menempelnya virus ke target infeksi membuat neuroamidase merupakan sasaran yang menarik bagi obat antivirus (Garman & Laver 2004). Kegiatan yang terkoordinasi dari glikoprotein HA dan NA dari strain virus tertentu merupakan hal yang penting bagi proses pelekatan dan pelepasan virion (Wagner et al. 2002).
Virion masuk dan menyatu ke dalam sebuah ruang endosom setelah berhasil melekat pada reseptor yang sesuai (Rust et al. 2004). Dalam ruang endosom, virus mengalami degradasi dengan cara menyatukan membran virus dengan membran endosom yang dimediasi oleh pemindahan proton melalui terowongan protein dari matrix-2 (M2) virus pada nilai pH di endosom sekitar 5,0. Selanjutnya akan terjadi serangkaian penataan ulang protein matrix-1 (M1) dan kompleks glikoprotein homotrimerik HA. Hasilnya, terbuka (exposed) sebuah bidang (domain) yang sangat lipofilik dan fusogenik dari setiap monomer HA yang masuk ke dalam membran endolisomal, sehingga mulai terjadi fusi antara membran virus dengan membran lisomal (Wagner et al. 2005). Kedelapan segmen RNA genomik dari virus, yang terbungkus dalam lapisan pelindung protein (ribonucleoprotein complex, RNP) nukleokapsid (N), dilepaskan ke dalam sitoplasma. Di sini mereka disalurkan ke nukleus untuk melakukan transkripsi mRNA virus dan replikasi RNA genomik melalui proses rumit yang diatur oleh faktor virus dan faktor sel (Whitaker et al. 1996). Polimerase yang dependen terhadap RNA (RdRp) dibentuk oleh sebuah kompleks (gabungan) dari PB1, PB2 dan protein PA virus, serta memerlukan RNA (RNP) yang terbungkus (encapsidated RNA (RNPs)) untuk hal ini. Setelah terjadi translasi protein virus dan perangkaian nukleokapsid yang membawa RNA genomik yang sudah ter-replikasi, virion-virion progeni tumbuh dari membran sel yang di dalamnya sudah dimasukkan glikoprotein virus sebelumnya. Penataan antara nukleokapsid berbentuk lonjong dan protein pembungkus virus dimediasi oleh
protein matrix-1 virus (M1) yang membentuk struktur serupa cangkang tepat di bawah pembungkus virus. Reproduksi virus di dalam sel peka bisa berlangsung sangat cepat (kurang dari sepuluh jam). Proses ini akan efisien, apabila gen optimal tersedia di sana (Rott et al. 1995).
Akibat aktivitas Polimerase yang dependen terhadap RNA (RdRp), virus mudah mengalami mutasi dan siklus replikasinya cepat. Jika ada tekanan selektif seperti antibodi penetral, ikatan reseptor yang tidak optimal, atau obat antiviral yang bekerja selama proses replikasi virus pada inang, dapat menyebabkan terjadi mutan dengan keunggulan selektif (mis. reseptor terlepas saat proses netralisasi serta membentuk unit pengikat reseptor baru) dan kemudian menjadi varian yang dominan dalam spesies virus di dalam tubuh inang. Determinan antigenik dari glikoprotein HA dan NA yang dipengaruhi oleh mekanisme yang dipicu oleh sistem kekebalan, prosesnya disebut sebagai Antigenic drift (Fergusson et al. 2003). Sebaliknya, Antigenic shift menunjukkan adanya perubahan mendadak dalam determinan antigenik, yaitu pertukaran subtipe H dan/atau N, dalam satu siklus tunggal replikasi. Hal ini terjadi dalam sebuah sel yang secara bersamaan terinfeksi oleh dua atau lebih virus influenza A dari subtipe yang berbeda. Karena distribusi segmen genomik virus yang sudah tereplikasi ke dalam progeni yang baru tumbuh berlangsung tanpa tergantung kepada subtipe asal dari tiap segmen, dan dapat muncul progeni yang berkemampuan untuk bereplikasi membawa informasi genetik dari virus induk yang berbeda-beda (disebut sebagai reassortant) (Webster & Hulse 2004, WHO 2005).
Sifat dan Patogenitas Virus Avian Influenza
Sifat virus Avian influenza antara lain mengaglutinasi sel darah merah ayam, mudah mengalami mutasi, virus mudah mati di luar sel tubuh ayam (tidak stabil di lingkungan). Virus ini juga mudah mati oleh panas, kekeringan, sinar ultraviolet, serta berbagai desinfektan yang umum di lapangan (deterjen, formalin, yodium, ammonium, kuarterner, hipoklorit, klorin, serta senyawa fenol). Tingkat keganasan virus Avian influenza sangat bervariasi, dan secara garis besar dibedakan atas dua
bentuk yaitu Low Pathogenic AI (LPAI) dan Highly Pathogenic AI (HPAI). Kejadian LPAI umumnya tidak menunjukkan gejala yang khas atau asimptomatik, misalnya berupa gangguan pernapasan ringan atau gangguan reproduksi. Kasus HPAI memiliki ciri-ciri antara lain bersifat sangat kontagius, dapat menimbulkan penyakit multisistemik dengan mortalitas tinggi. Bentuk HPAI dapat disebakan oleh virus AI dari beberapa suptipe H5 atau H7, namun penentuan bentuk HPAI tidak didasarkan pada subtipe H5 atau H7 tetapi sifat keganasan penyakit. Pada setiap wabah, subtipe H5 atau H7 mempunyai karekteristik internal gene yang mungkin bervariasi. Virus HPAI dapat berasal dari virus LPAI yang bermutasi pada protein permukaan hemaglutinin (OIE 2005)
Replikasi Virus
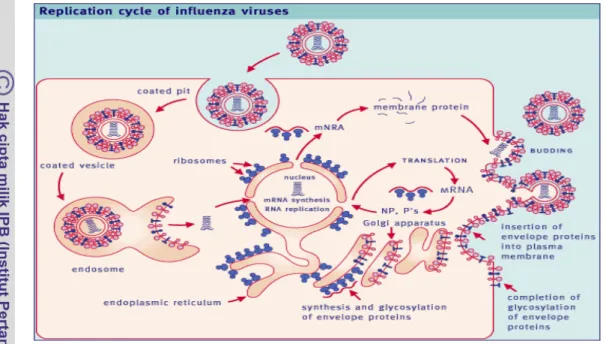
Replikasi virus dimulai dengan adsorbsi virus ke reseptor glikoprotein yang mengandung asam sialic pada permukaan sel. Virus kemudian memasuki sel dengan jalan endositosis melalui reseptor. Kondisi pH rendah dalam endosom, menghasilkan perubahan konformasi dalam hemaglutinin, yang memperantarai fusi membran. Nukleokapsid kemudian memasuki sitoplasma dan bermigrasi ke nukleus. Virus influenza menggunakan mekanisme yang unik untuk menginisiasi transkripsi, yaitu menggunakan viral transkriptase. Enam mRNA monosistronik dihasilkan dan ditranslasi menjadi HA, NA, NP dan tiga polimerase (PB1, PB2, dan PA). Melalui pembelahan mRNA untuk gen NS dan M masing-masing menjadi dua mRNA, yang ditranslasi dalam reading frame berbeda dan menghasilkan protein NS1, NS2, M1 dan M2. Hemaglutinin dan neuraminiase diglikosilasi dalam retikulum endoplasma kasar, dilengkapi di dalam badan golgi, ditransportasikan ke permukaan dan melekat pada membran sel (Easterday et al. 1997).
Syarat penting protein HA dapat bekerja adalah adanya pembelahan oleh protease sel inang menjadi HA1 dan HA2 yang dihubungkan dengan ikatan disulfida. Pembelahan dibutuhkan untuk dihasilkannya virus yang infeksius. Setelah produksi
dan pemasangan protein viral dan RNA, virus keluar sel dengan jalan menguncup dari membran plasma (Easterday et al. 1997).
Gambar 2. Replikasi Virus Influenza (Paul 2003)
Gambaran Klinis
Gambaran klinis penyakit Avian influenza (AI) pada unggas bervariasi serta gejalanya sering tidak spesifik, oleh karena itu tidak mungkin untuk menegakkan diagnosis hanya berdasarkan gambaran klinis. Virus berkembang selama beberapa hari, tergantung pada karakteristik isolat, dosis inokulum, spesies, dan usia unggas. Unggas yang menderita AI dapat mengeluarkan virus dalam jumlah yang besar di kotorannya. Virus tersebut dapat bertahan hidup di air sampai 4 hari pada suhu 22 0C dan lebih dari 30 hari dalam suhu 30 0C. Di dalam feses unggas dan dalam tubuh unggas yang sakit, virus dapat bertahan lebih lama, tetapi akan mati pada pemanasan 60 0C selama 30 menit (Soejoedono & Handharyani 2005). Gejala klinis infeksi virus AI berpatogenesis rendah tidak terlalu jelas, seperti bulu-bulu kusut, produksi telur secara perlahan menurun, penurunan berat badan disertai gangguan pernapasan ringan (Capua & Mutinelli 2001). Beberapa strain berpatogenesis rendah misalnya
strain H9N2 dari garis Asia virus ini telah beradaptasi sehingga menghasilkan replikasi yang efisien dalam unggas, dan menimbulkan gejala klinis yang nyata serta mengakibatkan kematian secara signifikan. Infeksi virus AI yang patogenitasnya tinggi, menimbulkan penyakit yang ditandai dengan serangan mendadak dan gejala hebat serta kematian mendekati 100% dalam jangka waktu 48 jam (Swayne & Suarez 2000). Penyebaran virus dalam kelompok unggas tergantung bentuk pemeliharaannya. Pada kelompok unggas yang dilepas di tempat kotor dan terjadi hubungan langsung serta percampuran dengan hewan lain, penyebaran infeksi berlangsung lebih cepat daripada yang dipelihara dalam kandang. Namun masih diperlukan beberapa hari untuk terjadinya penularan yang sempurna (Capua et al. 2000). Infeksi virus pada peternakan unggas skala besar, terjadinya penurunan konsumsi air dan makanan yang drastis dalam waktu singkat. Hal ini dapat menjadi tanda akan adanya penyakit sistemik. Pada ayam petelur, terhentinya produksi telur sangat nyata. Secara individual, ayam yang terkena HPAI sering hanya menunjukkan apatis dan imobilitas. Pembengkakan nampak pada daerah kepala yang tidak ditumbuhi bulu, terjadi sianosis pada jengger, gelambir dan kaki, diare dengan kotoran berwarna kehijauan, dan susah bernafas, dapat dijumpai meskipun tidak selalu (inkonsisten). Pada ayam petelur, awalnya telur yang dihasilkan berkulit lembek, tetapi kemudian produksi telur berhenti secara cepat sejalan dengan perkembangan penyakit (Elbers et al. 2005). Gejala sistem saraf termasuk tremaor, tortikolis, dan ataxia mendominasi gambaran klinis pada spesies yang tidak begitu rentan seperti bebek, angsa, serta burung onta.
Vaksin dan Vaksinasi Terhadap Penyakit Avian Influenza
Vaksinasi bertujuan untuk memberikan perlindungan terhadap timbulnya penyakit secara klinis. Perlindungan terhadap serangan virus yang virulen, merupakan perlindungan terhadap ekskresi virus. Vaksinasi yang digunakan saat ini belum ada yang secara eksperimental memenuhi persyaratan di atas (Lee & Suarez 2005).
Teknologi produksi vaksin untuk penyakit AI saat ini diarahkan ke tiga tipe produksi yaitu vaksin konvensional homolog, vaksin konvensional heterolog, dan
vaksin rekombinan homolog. Vaksin konvensional homolog H5N1, memiliki homologi yang mirip virus lapang sehingga tidak terbentuk mutant akibat tekanan imunologis (Antigenic drift). Namun vaksinasi ini memiliki kelemahan antara lain hingga saat ini bentuk virus H5N1 bersifat ganas sehingga pada proses produksinya secara konvensional membahayakan. Vaksin konvensional heterolog H5N2, H5N9, H5N1, dipakai pada vaksin konvensional karena sifatnya Low Pathogenic sehingga tidak berbahaya pada proses produksi vaksin dan dapat menghasilkan titer virus tinggi. Namun vaksin konvensional heterolog memiliki kelemahan antara lain antigenisitas tidak kompatibel (homologi yang mirip) sepenuhnya terhadap virus lapang sehingga besar kemungkinan terbentuk mutant akibat tekanan imunologis (Antigenic drift). Vaksin rekombinan homolog memiliki kelebihan dapat menginduksi imunitas seluler dan humoral, tidak menimbulkan reaksi sakit akibat vaksinasi, tidak memerlukan adjuvan, lebih effektif dan efisien, dapat diberikan pada anak ayam umur muda. Pembedaan antara hewan terinfeksi atau divaksinasi lebih nyata secara klinis. Namun vaksin rekombinan homolog juga memiliki kelemahan antara lain kemampuan bereplikasi yang rendah dan menginduksi kekebalan parsial pada unggas, pemberian vaksin rekombinan juga berarti memaparkan vaksin atau virus vektornya. Efek antibodi asal juga dapat mempengaruhi efikasi vaksin, sampai saat ini efikasi vaksin perlu diteliti terlebih dahulu, serta penggunaan vaksin tersebut harus dibatasi untuk spesies tertentu.
Antibodi
Hewan yang terpapar oleh suatu antigen, akan membentuk respon kekebalan pada tubuh hewan tersebut. Respon humoral tubuh yang terinfeksi menghasilkan antibodi. Antibodi merupakan molekul protein yang diproduksi oleh sel B dan saling berhubungan secara spesifik dengan molekul asing (antigen).
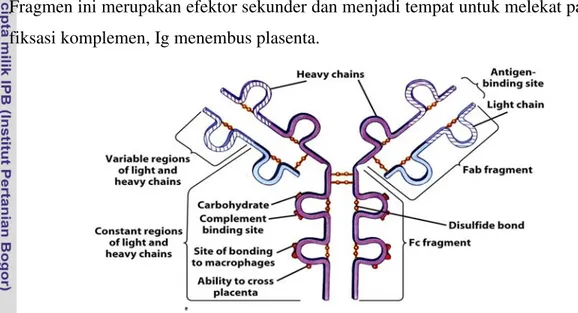
Molekul antibodi merupakan globulin, sehingga umumnya dikenal sebagai immunoglobulin (Ig). Dikenal 5 kelas utama imunoglobulin dalam serum, IgG, IgA, IgM, IgD dan IgE. Struktur dasar imunoglobulin terdiri atas 2 rantai berat (H-chain)
yang identik dan 2 rantai ringan (L-chain) yang juga identik. Setiap rantai ringan terikat pada rantai berat melalui ikatan disulfida (S – S). Fragmen Fab dengan antigen binding site, berfungsi mengikat antigen, karena itu susunan asam amino dibagian ini berbeda antara molekul imunoglobulin satu dengan yang lain dan bervariasi sesuai dengan variabilitas antigen yang merangsang pembentukannya. Sebaliknya fragmen Fc merupakan fragmen yang konstan. Fragmen ini tidak mempunyai kemampuan mengikat antigen tetapi dapat bersifat sebagai antigen (Determinan antigen). Fragmen ini merupakan efektor sekunder dan menjadi tempat untuk melekat pada sel, fiksasi komplemen, Ig menembus plasenta.
Gambar 3 . Struktur Antibodi (Jacquelyn 2005)
Fungsi antibodi sebagian besar ditentukan oleh spesifitas antigen binding site dan isotype H-chain. Subkelas yang berbeda menunjukkan perbedaan pula dalam hal kemampuan berikatan dengan reseptor Fc, komplemen dan reseptor lain . IgG, IgA dan IgE mempunyai afinitas terhadap antigen yang lebih tinggi dibanding IgM, walaupun hal ini juga bergantung pada pematangan (maturasi) afinitas yang bersaingan dengan Class switching pada sel B dan tidak ada hubungannya dengan domain terminal – C. Dalam tubuh terdapat 2 bentuk imunoglobulin yang berbeda, yaitu sebagai reseptor permukaan untuk antigen dan sebagai antibodi yang disekresikan kedalam cairan ekstraselular. Antibodi yang disekresikan dapat berfungsi sebagai adaptor yang berfungsi untuk mengikat antigen pada struktur
binding-sitenya yang spesifik. Agar terjadi proses netralisasi, antigen harus dikenal oleh antibodi. Bagian antibodi yang dikenal atau bereaksi dengan antibodi disebut epitop, sedangkan bagian antibodi yang dapat mengenal antigen disebut paratop.
Bentuk Alami Dari Reaksi Antibodi Antigen
Studi X-ray crystallography dari interaksi antibodi antigen menunjukkan adanya sebuah bentuk celah lewat tempat gabungan antibodi pada jaring determinan antigen. Selanjutnya konsep reaksi antibodi antigen merupakan sebuah kunci (yaitu antigen) yang cocok masuk dalam sebuah gembok (yaitu antibodi) (Li et al. 2000).
Ikatan antibodi antigen mempengaruhi antigen pada lokasi kombinasi semua antibodi non-kovalen dalam bentuk alaminya, diantaraya termasuk ikatan hidrogen, ikatan elektrostatik, kekuatan Van der waals serta ikatan hidrofobik. Banyaknya ikatan diantara antigen dan antibodi yang terjadi menyebabkan antigen menjadi berbentuk seperti papan bagi antibodi. Sejak terjadi reaksi antibodi antigen melalui ikatan non kovalen, maka dikatakan sebagai bentuk reversibel alami.
Gambar 4. Bentuk alami reaksi antibodi antigen (Li et al. 2000)
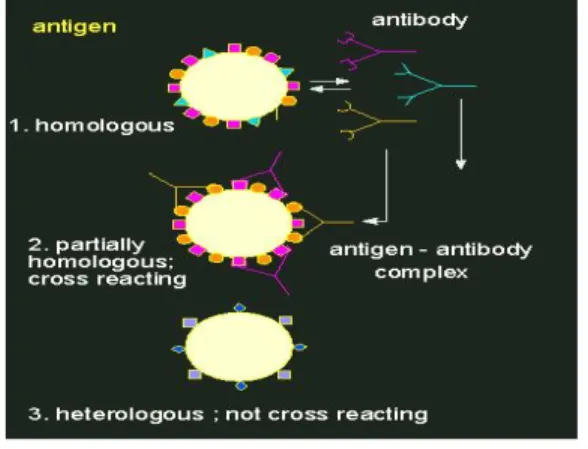
Bentuk antibodi (digambarkan dengan struktur Y) berbeda jenisnya untuk molekul antigen, dimana perbedaannya sangat spesifik. Reaksi silang pada sebuah
populasi antibodi dengan antigen asing hanya terjadi, jika terdapat homologi dengan antigen asing. Setiap antibodi mempunyai dua tempat penanda ikatan untuk determinan antigen.
Gambar 5. Reaksi antibodi - antigen (Peter 2007)
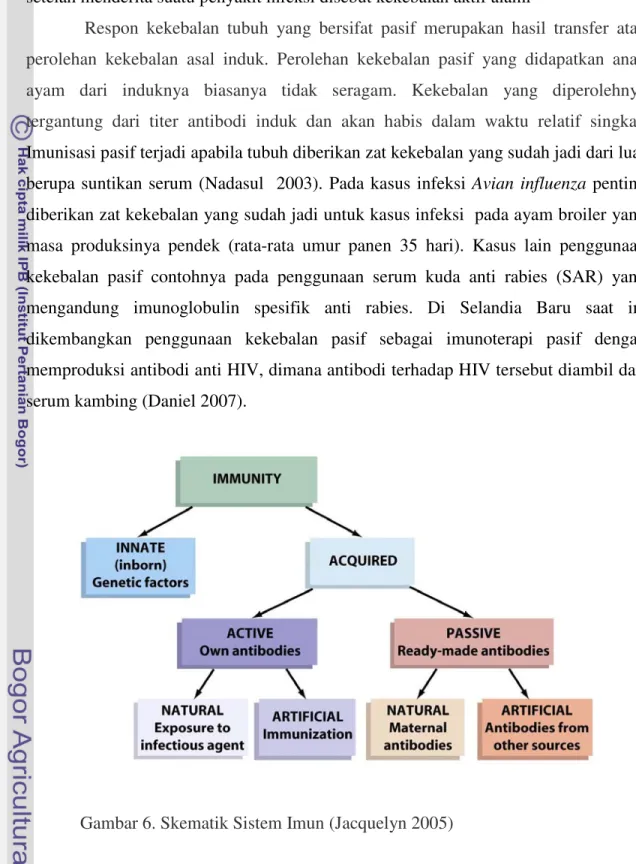
Imunisasi Pasif
Pertahanan tubuh dibagi dua, yaitu pertahanan tubuh non spesifik dan pertahanan tubuh spesifik. Sistem pertahanan tubuh non spesifik merupakan sistem pertahanan tubuh yang melindungi dari berbagai ancaman penyakit secara umum. Sistem pertahanan non spesifik berupa kulit, mukosa, mukus dan silia pada saluran pernapasan. Selain itu pertahanan non spesifik juga berupa fagositosis, sistem komplemen dan sel pembunuh. Sistem pertahanan tubuh spesifik, berkaitan dengan adanya respon kekebalan tubuh berperantara seluler maupun humoral.
Respon kekebalan tubuh humoral dapat bersifat aktif maupun pasif. Sistem ini mampu mengenali antigen sebagai benda asing, mempunyai spesifitas tertentu serta memori terhadap antigen. Respon kekebalan tubuh yang bersifat aktif dapat diperoleh sebagai hasil vaksinasi, dan paparan materi yang berkaitan dengan respon kekebalan
humoral aktif seperti antigen. Kekebalan aktif timbul karena adanya vaksinasi oleh
karena tubuh secara aktif membentuk setelah diberikan rangsangan oleh vaksin (berisi bibit penyakit) yang disuntikan. Oleh karena sengaja dibuat, kekebalan
vaksinasi disebut sebagai kekebalan aktif buatan. Sementara antibodi yang terbentuk setelah menderita suatu penyakit infeksi disebut kekebalan aktif alami
Respon kekebalan tubuh yang bersifat pasif merupakan hasil transfer atau perolehan kekebalan asal induk. Perolehan kekebalan pasif yang didapatkan anak ayam dari induknya biasanya tidak seragam. Kekebalan yang diperolehnya tergantung dari titer antibodi induk dan akan habis dalam waktu relatif singkat. Imunisasi pasif terjadi apabila tubuh diberikan zat kekebalan yang sudah jadi dari luar berupa suntikan serum (Nadasul 2003). Pada kasus infeksi Avian influenza penting diberikan zat kekebalan yang sudah jadi untuk kasus infeksi pada ayam broiler yang masa produksinya pendek (rata-rata umur panen 35 hari). Kasus lain penggunaan kekebalan pasif contohnya pada penggunaan serum kuda anti rabies (SAR) yang mengandung imunoglobulin spesifik anti rabies. Di Selandia Baru saat ini dikembangkan penggunaan kekebalan pasif sebagai imunoterapi pasif dengan memproduksi antibodi anti HIV, dimana antibodi terhadap HIV tersebut diambil dari serum kambing (Daniel 2007).
Uji Netralisasi Virus
Uji netralisasi virus dapat digunakan untuk mengukur titer antibodi secara kuantitatif. Selain itu uji netralisasi dilakukan juga dalam idetifikasi virus yang tidak diketahui dengan menggunakan antisera yang sudah diketahui. Uji netralisasi terdiri dari dua tahap. Tahap pertama adalah virus dengan titer tertentu direaksikan dengan serum pada beberapa titer tertentu didalam sebuah tabung uji. Campuran virus dan serum diinkubasi bersama pada temperatur standar untuk jangka waktu tertentu. Tahap kedua, dilakukan pembiakan virus-virus yang tidak ternetralisasi ke sistem indikator (media biakan). Setelah diinkubasikan dilakukan pengamatan terhadap hasil pembiakan.
Serum yang akan diuji netralisasi harus disterilkan dahulu, bebas bahan kimia dalam penyimpanannya (phenol, formalin dan lainnya), serta telah diinaktivasi. Inaktivasi dilakukan dengan pemanasannya 56 0C selama 30 menit, pemanasan tersebut akan merusak substansi nonspesifik penghambatnya yang menghambat reaksi Ab dengan virus.
Strain virus yang digunakan untuk uji netralisasi harus mempunyai titer yang tinggi, tidak serumpun dengan virus uji, serta adaptasinya sangat baik dengan metode yang digunakan. Virus yang digunakan juga harus murni dan bebas dari bakteri, fungi, atau mikoplasma. Sebagai pelarut dapat digunakan media kultur sel.
Prosedur uji netralisasi yang digunakan atau dikenal saat ini yaitu prosedur uji netralisasi-ß dan prosedur uji netralisasi-α (Swayne et al. 1998). Pada uji netralisasi - ß, serum yang diuji diencerkan secara seri atau desimal dan virus standarnya bertiter tetap. Uji ini memiliki keuntungan yaitu volume serum uji yang digunakan sedikit. Pada uji netralisasi-α virus diencerkan secara serial serta diencerkan dengan serum tetap pada titer tertentu (tampa pengencer). Campuran virus dan serum diinkubasi dan dihitung untuk residual virus yang terkandung didalamnya yang dinyatakan dengan Lethal Dose 50 (LD50) atau Infectious Dose 50 (ID50).
Titrasi Virus
Titrasi virus diperlukan untuk kepentingan diagnosis, diantaranya untuk menghitung jumlah infektif virus dalam suatu material sampel. Ada beberapa metode yang digunakan untuk menghitung perkiraan jumlah (kuantitatif) virus antara lain nilai akhir pengenceran (dilution end-point), menghitung jumlah plak (Plaque counts), atau jumlah pock (Pock plaque). Nilai akhir pengenceran (dilution end-point) merupakan metode yang banyak digunakan. Perhitungan ini melibatkan dosis virus yang dibutuhkan untuk menginfeksi 50% populasi inang (hewan, telur, kultur jaringan) (Swayne et al. 1998). Titer dihitung ditandai dengan ID50 atau LD50 untuk tingkat infeksi atau kematian melalui pengenceran serial dari virus. Pengenceran serial ini kemudian diinokulasi pada sebuah Telur Ayam Berembrio (TAB) serta dilakukan pengamatan jumlah respon positif dan negatif. Dari hasil uji, dimungkinkan untuk menghitung 50% end-point dengan metode Reed and Muench. Respon atau nilai akhir (end-point) yang digunakan akan bervariasi dengan virus yang diuji. Namun diperlukan juga melihat kehadiran infeksi virus oleh faktor lain misalnya adanya lesi, munculnya virus akibat pewarnaan, hemagglutinasi atau lainnya.
Titrasi virus pada biakan jaringan biasanya dibuat dengan pengenceran virus atau efek cytophatogenic (kematian sel-sel tampa formasi plak). Metode plak sangat penting digunakan karena satu infeksi virus memberikan satu plak, jumlah plak pada pengenceran tertentu akan menghasilkan jumlah total partikel. Penghitungan jumlah Pock virus dalam suspensi yang dapat menginfeksi hampir serupa dengan metode Plaque counts hanya pada metode ini, perubahan yang terjadi, dilihat pada khorio alantois membran pada telur ayam berembrio.