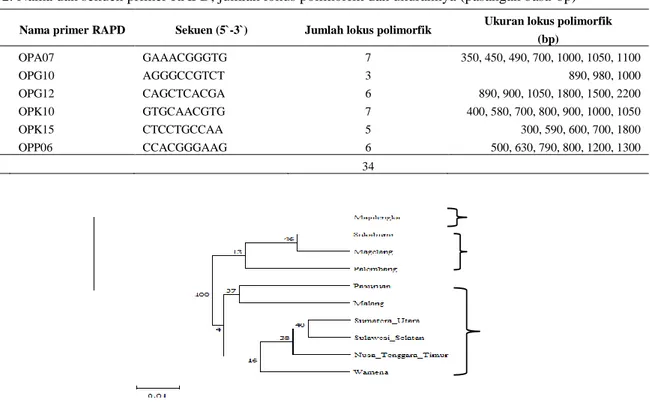
The differences of stem xylem structures of sengon (Falcataria moluccana) from Solomon
and Wamena Provenances
Lucy Ana Cahya Inkasari1, Liliana Baskorowati2, dan Anti Damayanti1 1Fakultas Biologi, Universitas Islam Negeri Sunan Kalijaga
Jl. Marsda Adisucipto Yogyakarta 55281
2
Balai Besar Penelitian dan Pengembangan Bioteknologi dan Pemuliaan Tanaman Hutan Jl. Palagan Tentara Pelajar, Km.15, Purwobinangun, Pakem, Sleman, Yogyakarta
e-mail: l_baskorowati@yahoo.co.id
Tanggal diterima: 12 Oktober 2015, Tanggal direvisi: 30 Oktober 2015, Disetujui terbit: 10 Juni 2016
ABSTRACT
Sengon (Falcataria moluccana) is fast growing species mostly planted by farmers due to its high productivity. Sengon originating from Solomon Island has been known with the high productivity eventhough susceptible to gall rust attack; on the other hand, sengon originating from Wamena is known to be more tolerant to gall rust attack. There is no previous study in terms of stem xylem structures comparing sengon from those seed origins. Therefore, this study was undertaken to identify the differences in anatomical structure of sengon stem; and to compare the xylem cell based on proportions and dimensions of the stems between the two provenances. Six stem samples of tolerant Wamena provenance and six samples of susceptible Solomon provenance were used in this study. Samples were collected from progeny trial of sengon in Lumajang, Jawa Timur. The observations include the anatomical structures and vessel element, parenchyma apotracheal cell, parenchyma paratracheal cell, xylem fiber, fiber length, and fiber diameter. The result showed that there was no difference in terms of anatomical structures between sengon Wamena (tolerant) and Solomon (susceptible) stem in cross section, tangential and radial section of periderm, phloem, secondary xylem (vessel cell, xylem fiber, and parenchyma xylem) and pith. However, in one of susceptible stem sample, a black reaction zone on the secondary xylem was found. Analysis of variance showed that parenchyma paratracheal cell, apotracheal cell, number of xylem fibers, xylem fiber diameter and length were not significantly different between stem of sengon Wamena (tolerant) and Solomon (susceptible).
Keywords: anatomy, sengon stem, xylem, Solomon, Wamena
ABSTRAK
Sengon (Falcataria moluccana) merupakan salah satu jenis tanaman cepat tumbuh yang banyak ditanam masyarakat, karena produktifitas yang tinggi. Sengon provenan Solomon ditengarai mempunyai produktivitas yang tinggi meskipun tidak tahan terhadap penyakit karat tumor, sedangkan sengon provenan Wamena diketahui mempunyai ketahanan yang lebih baik terhadap penyakit karat tumor. Studi tentang perbandingan struktur xylem batang sengon dari provenan tersebut belum pernah dilakukan sebelumnya. Oleh karena itu penelitian ini dilakukan untuk mengetahui perbedaan struktur anatomi dan perbandingan berdasarkan proporsi dan dimensi sel xilem antara batang sengon Solomon dan Wamena. Enam contoh batang rentan penyakit karat tumor provenan Solomon dan 6 contoh batang tahan karat tumor provenan Wamena digunakan dalam penelitian ini. Contoh kayu diambil dari petak uji keturunan sengon di Lumajang, Jawa Timur. Hasil penelitian menunjukkan bahwa secara penampang melintang, tangensial dan radial pada bagian periderm hingga bagian empulur tidak ada perbedaan struktur anatomi batang antara sengon Solomon dan Wamena. Namun demikian, pada salah satu contoh Solomon, terdapat zona reaksi berwarna hitam pada bagian xilem sekunder. Hasil analisis varian menunjukkan bahwa sel parenkim paratrakeal, apotrakeal, jumlah serat xilem, panjang serat xilem dan diameter serat xilem tidak menunjukkan perbedaan nyata antara batang sengon dari Wamena (toleran) dan Solomon (rentan).
I. PENDAHULUAN
Sengon (Falcataria moluccana)
merupakan spesies cepat tumbuh (fast growing
species) yang banyak dibudidayakan oleh
masyarakat karena kemudahannya beradaptasi dengan lingkungan dan nilai ekonomi yang tinggi. Sengon mempunyai produktivitas yang tinggi dengan riap rata-rata pertahun antara 10– 25 m3/ha/th (8 tahun) dan 30-40 m3/ha/th (12 tahun), dengan pola usaha tani, riap dapat mencapai 16,78 m3/ha/th; dan pada tanaman sengon umur 7 tahun dengan perlakuan seleksi riap dapat ditingkatkan menjadi 27,26 m3/ha/th (Soerianegara & Lemmens, 1993; Rimbawanto, 2008). Sampai saat ini, sengon yang berasal dari kepulauan Solomon diyakini merupakan sengon dengan produktivitas yang paling tinggi (Hardiyanto, 2010). Beberapa penelitian sebelumnya menyatakan bahwa sengon Solomon memiliki pertumbuhan yang lebih cepat dibandingkan dengan sengon lokal, dengan produktivitas 3 kali lipat dibandingkan dengan sengon lokal. Sengon Solomon mempunyai rerata pertumbuhan tinggi 5 m dan diameter 5,7 cm pada umur 1 tahun; serta memiliki rerata pertumbuhan diameter 16 cm saat 2 tahun, dan 19 cm pada umur 3 tahun (Hardiyanto, 2010; Setiadi et al., 2014b). Hasil penelitian pada petak uji keturunan sengon Solomon oleh Setiadi et al. (2014b) menyatakan bahwa rerata pertumbuhan tinggi dan diameter umur 6, 12 dan 18 bulan berturut-turut adalah 2,42 m dan 2,97 cm; 4,74 m dan 5,56 cm, serta 17,35 m dan 7,39 cm.
Namun demikian, produktivitas yang tinggi dari sengon Solomon tersebut sangat terkendala dengan adanya serangan penyakit karat tumor. Sengon yang berasal dari Solomon merupakan jenis sengon yang sangat mudah tertular penyakit karat tumor dibandingkan dengan sengon dari Indonesia. Penelitian pada petak uji keturunan sengon di Bondowoso oleh Setiadi et al. (2014a) memperlihatkan bahwa pada umur 6 bulan sengon Solomon mulai
terserang karat tumor dengan nilai luas serangan 0,85% dan intensitas serangan 0,51%; yang meningkat secara nyata pada saat umur tanaman 1 tahun yaitu 39,60% (luas serangan) dan 17,99% (intensitas serangan). Semua famili sengon Solomon yang tumbuh dalam petak uji sengon Solomon di Bondowoso (25 famili, 4 tree plot, 8 blok) terserang penyakit karat tumor sejak umur 6 bulan dengan luas serangan dan intensitas serangan yang bervariasi antar famili (Setiadi et al., 2014b).
Sengon yang berasal dari Wamena, sebaliknya, diketahui merupakan jenis sengon yang lebih tahan terhadap penyakit karat tumor, karena menunjukkan luas dan intensitas serangan yang lebih rendah (Baskorowati & Nurrohmah, 2011; Baskorowati et al., 2012). Lebih lanjut Rahayu et al., (2009) dengan hasil penelitiannya terkait inokulasi buatan jamur U.
falcatarium pada semai sengon umur 6 minggu
di persemaian menemukan bahwa semai yang berasal dari Wamena lebih tahan terhadap penyakit karat tumor jika dibandingkan dengan Kediri, Timor Timur, Morotai, 2S/75 (asal Sabah) dan Walang Gintang. Hasil penelitian awal Charomaini & Ismail (2008) juga menyebutkan bahwa individu-individu yang berasal dari Papua seperti Waga-waga, Wamena, Hubikosi, dan Muliama Bawah lebih tahan terhadap serangan penyakit karat tumor. Setiadi et al., (2014a) menambahkan bahwa individu-individu yang berasal dari Papua, seperti Holima, Meagama, dan Elagaima tahan terhadap penyakit karat tumor sampai umur 12 bulan pada uji keturunan di Bondowoso. Lebih lanjut, Diputra (2015) menyatakan bahwa tanaman sengon yang berasal dari Wamena A, Wamena B lebih tahan terhadap terhadap penyakit karat tumor di lapangan dibandingkan dengan sengon asal Nabire, Manokwari, Serui dan ras lahan Jawa.
Tanggapan tanaman sengon terhadap penyakit karat tumor sangat dipengaruhi oleh faktor genetik dan lingkungan. Beberapa penelitian tersebut di atas menunjukkan bahwa tanaman tersebut memiliki materi genetik yang
tahan terhadap penyakit. Salah satu faktor yang dapat mempengaruhi kemunculan mekanisme ketahanan tersebut adalah cekaman lingkungan (Hopkins & Huiiner, 2008). Tanggapan tanaman terhadap berbagai cekaman dapat menghasilkan perbedaan morfologi, anatomi dan fisiologi tanaman, misalnya perubahan pada pertumbuhan tanaman,volume sel menjadi lebih kecil, penurunan luas daun, daun menjadi lebih tebal, penurunan jumlah akar, penurunan laju fotosintesis, perubahan metabolism serta perubahan ekspresi gen (Salisbury & Ross, 1995). Lebih lajut Salisbury & Ross (1995) menyebutkan bahwa penyakit tanaman dan serangan hama penyakit merupakan kategori cekaman biotik yang dapat dialami tanaman selama daur hidupnya.
Serangan patogen juga merupakan salah satu jenis cekaman. Beberapa penelitian terkait perubahan anatomi akibat serangan patogen diantaranya menunjukkan perubahan anatomi tumor kayu sengon trubusan yang terserang jamur U. tepperanium (Rukhama & Nugroho, 2014). Selain itu, Batang pinus yang terkena tumor (gall rust) memiliki jari-jari xilem (xylem
ray) dan jari-jari floem (floem ray) lebih rapat,
peningkatan jumlah sel parenkim floem, hiperplasia di korteks serta batas kambium yang tidak terlihat jelas dibandingkan batang yang sehat atau normal (Jewell, 1988). Lebih lanjut, cabang yang terinfeksi pada Pinus densiflora memiliki jumlah trakeid, jumlah saluran resin dan jari-jari yang lebih banyak daripada cabang yang tidak terinfeksi (Yamamoto et al., 1998). Disebutkan juga bahwa keberadaan patogen juga menyebabkan penebalan pada sel sklerenkima, sel parenkim di daerah sekitar berkas pembuluh (Yamamoto et al., 1998; Zalasky, 1976).
Penelitian – penelitian tersebut menunjukkan bahwa struktur anatomi dapat digunakan sebagai petunjuk ciri adanya suatu cekaman. Jaringan yang sering mengalami perubahan karena cekaman adalah jaringan xilem. Oleh karena itu, kajian anatomi memungkinkan untuk memahami dasar adaptasi
tanaman dalam berbagai kondisi cekaman lingkungan. Penelitian ini diharapkan dapat mengetahui perbandingan struktur anatomi batang sengon Wamena yang tahan dan sengon Solomon yang rentan terhadap penyakit karat tumor, ditinjau dari jaringan xilemnya.
II. METODE PENELITIAN Waktu dan Lokasi
A.
Pengambilan sampel dilapangan dilakukan pada pada bulan November 2014. Sedangkan pengamatan di labolatorium dilakuan pada bulan Januari sampai Maret 2015. Sampel kayu diambil dari plot uji keturunan Sengon B2P2BPTH Yogyakarta di Lumajang (dusun Kayu Enak, desa Kandang Tepus, Kecamatan Senduro, Kabupaten Lumajang). Sedangkan pengamatan di labolatorium dilakukan di labolatorium kayu B2P2BPTH Yogyakarta.
Bahan dan Alat B.
Bahan yang digunakan dalam penelitian ini adalah batang pohon sengon umur 2 tahun yang ditanam pada Plot Uji Keturunan Sengon di Lumajang, Jawa Timur. Tanaman uji keturunan ini menggunakan rancangan penelitian Incomplete Block Design (IBD) dengan 97 seedlot (famili) 4 treeplot dan 7 blok (replikasi) di lokasi Lumajang. Tanaman sengon dalam plot tersebut berasal dari Wamena, Serui, Manokwari, Nabire dan Kepulauan Solomon. Sedangkan bahan di laboratorium meliputi alkohol 96%, xilol, safranin 0,25%, glyserin, albumin, aquades, asam asetat glasial (CH3COOH) dan hydrogen peroksida (H2O2).
Alat yang digunakan dalam penelitian ini adalah mikroskop, mikrotom, gelas benda, gelas penutup, cawan petri, pipet tetes, pinset, gelas beker, erlemeyer, aluminium foil, gelas ukur, oven, spatula, cutter atau pisau, kuas, kertas saring, dan kawat.
Prosedur Kerja C.
1. Koleksi contoh
Pengambilan contoh kayu dilakukan pada batang pohon sengon hasil seleksi Plot Uji Keturunan Sengon umur 2 tahun di Dusun Kayu Enak, Desa Kandang Tepus, Kecamatan Senduro, Kabupaten Lumajang yang ditanam oleh Balai Besar Penelitian Bioteknologi dan Pemuliaan Tanaman Hutan bekerjasama dengan Dinas Kehutanan Lumajang. Batang diambil dengan melakukan penebangan dan untuk menyeragamkan contoh, batang diambil dengan jarak 1 meter dari permukaan tanah. Jumlah batang pohon yang diambil sebanyak 12 pohon dengan 6 pohon berasal Solomon dan 6 pohon dari Wamena. Contoh batang yang berasal dari Solomon merupakan pohon yang terserang penyakit karat tumor yang ditunjukkan dengan adanya gall (tumor) yang terbentuk pada pohon tersebut, umumnya terdapat di bagian batang. Sedangkan contoh batang yang berasal dari Wamena merupakan pohon yang sehat tidak terindikasi adanya serangan penyakit di semua bagian tanaman.
2. Penentuan proporsi sel (Kasmudjo, 1985) Contoh uji potongan kayu pada bagian batang pohon (setinggi 1,3 m diatas permukaan tanah disiapkan untuk analisis labolatorium. Preparat dibuat dengan terlebih dahulu mempersiapkan contoh uji berupa potongan kayu arah vertikal dengan ukuran sekitar 1,5 x 1,5 x 3 cm.Potongan kayu tersebut dimasukkan ke dalam tabung yang berisi campuran akuades dan gliserin dengan perbandingan 1:3 selama 4 hari untuk pelunakan kayu (Schweingruber, 2007). Contoh dibuat menjadi tiga macam irisan yaitu penampang melintang (x), tangensial (t) dan radial (r) dengan menggunakan mikrotom dengan ketebalan 15-20 mikron. Irisan dipilih yang terbaik yaitu irisan yang tipis dan tidak sobek. Irisan ditampung dalam cawan petri yang berisi akuades. Pewarnaaan dilakukan dengan menggunakan safranin 0,25%.
Contoh dicuci dengan menggunakan alkohol 96% sebanyak tiga kali selama 5 menit, dicelupkan ke dalam xylol selama 3 menit. Contoh kemudian dikeringkan dengan menggunakan kertas saring, lalu diletakkan di atas kaca preparat dan ditutup dengan menggunakan kaca penutup. Preparat kemudian diamati di bawah mikroskop.
Pengamatan pada penampang x meliputi sel parenkim apotrakeal dan sel trakea. Pada penampang t yaitu sel parenkim paratrakeal dan pada penampang r diamati serat xilem (xylem
fibers). Dilakukan perhitungan pada data yang
telah dikumpulkan untuk mengetahui persentase selnya.
3. Penentuan dimensi sel dengan maserasi kayu (Kasmudjo, 1985)
Pembuatan contoh uji dilakukan dengan memotong kayu berukuran 0,2 cm x 0,2 cm x 1,5 cm. Contoh diambil pada bagian tengah batang antara kulit dengan empulur. Cairan maserator yaitu campuran antara asam asetat glasial (CH3COOH) dan hydrogen peroksida
(H2O2) dengan perbandingan 1:1, kemudian
disiapkan dalam tabung reaksi. Contoh dimasukkan dalam botol, kemudian cairan maserator dituang sampai contoh terendam. Contoh dioven selama 3 hari pada suhu 60°C. Contoh kemudian dicuci dengan menggunakan air sampai 5 kali sehingga benar-benar terbebas dari zat kimia.
Untuk memisahkan serat-seratnya, tabung reaksi yang berisi contoh diisi dengan akuades hingga ¾ volume tabung kemudian digoyangkan secara perlahan hingga preparat menjadi serabut yang saling terlepas. Serat kemudian diambil dengan menggunakan pipet dan diletakkan diatas kaca preparat, ditetesi dengan safranin 0,25% sebanyak 2-3 tetes dan didiamkan selama 5 menit. Sisa zat warna kemudian dihilangkan dengan menggunakan kertas saring, dan preparat ditutup dengan kaca penutup. Preparat kemudian diamati di bawah mikroskop. Pengamatan meliputi panjang serat serta diameter serat. Jumlah panjang serat yang
diukur sebanyak 100 serat. Hal ini dilakukan untuk mengetahui n kali serat yang diukur. Penentuan n serat dilakukan dengan menggunakan pendekatan menurut Kasmudjo (1985): dengan: ∑ ∑ ∑ Keterangan:
N = jumlah serat yang diukur S = standar deviasi
L = nilai rata-rata panjang serat kali 0,05 (error 5% dianggap memadai)
Xi = panjang serat Fi = frekuensi serat
N = jumlah serat yang diukur dalam pengukuran pendahuluan (n=100)
4. Analisis Data
Data kuantitatif seperti sel parenkim apotrakeal, sel trakea, sel parenkim paratrakeal, serat xilem, panjang serat, dan diameter serat dianalisis dengan menggunakan analisis one
way ANOVA. Jika terdapat perbedaan, maka
dilanjutkan dengan uji Duncan pada tingkat signifikasi 5% untuk menunjukkan famili dan provenan yang berbeda nyata.
III. HASIL DAN PEMBAHASAN Struktur Anatomi
A.
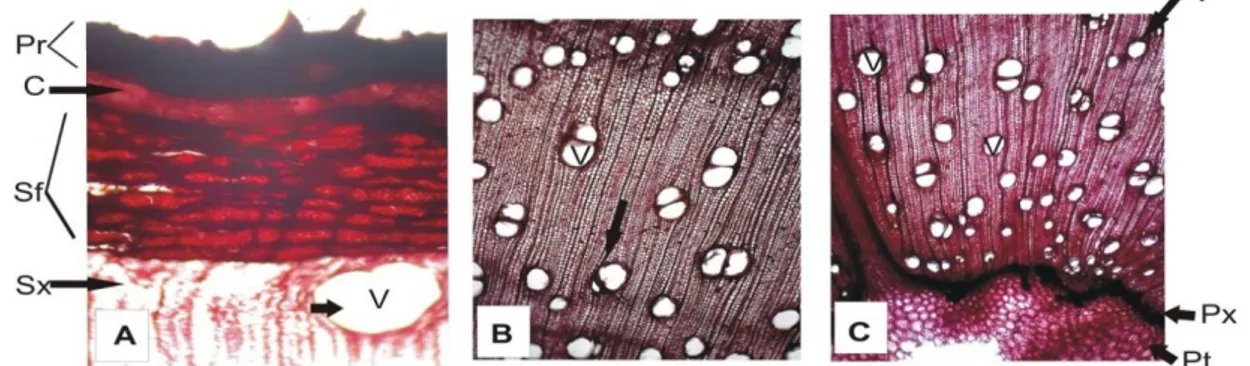
Hasil penelitian menunjukkan bahwa penampang melintang batang sengon Wamena (Gambar 1a) bagian periderm, floem, xilem sekunder (sel pembuluh, sel serat xilem, dan parenkim xilem), serta bagian empelur tidak berbeda dengan struktur anatomi pada batang sengon Solomon (Gambar 1b). Gambar tersebut juga menunjukkan bahwa jarak antar sel parenkim apotrakeal relatif sama, begitu pula kepadatan sel trakea pada kedua batang sengon.
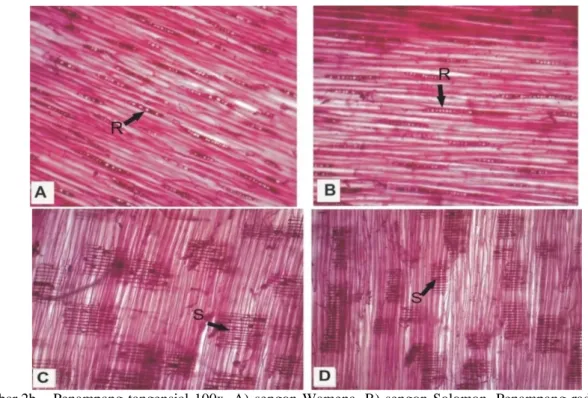
Dengan membandingkan penampang tangensial (Gambar 2a) dan penampang radial (Gambar 2b) terlihat bahwa tidak terdapat perbedaan antara struktur anatomi batang tahan (Wamena) dengan batang yang rentan (Solomon); yang terlihat dari kepadatan sel parenkim paratrakeal dan serat xilem yang sama. Namun demikian, pada salah satu contoh batang sengon Solomon, terdapat bercak karat tumor yang meskipun belum sampai terbentuk pembengkakan tumor, telah menunjukkan perubahan anatomi (Gambar 3). Perubahan anatomi yang ditimbulkan adalah adanya lapisan sel atau zona infeksi yang berwarna hitam di xilem sekunder. Selain itu, infeksi juga terdapat di bagian lain yaitu sekitar pembuluh karena parenkim berwarna hitam (Gambar 3a). Zona infeksi ini terletak dekat dengan korteks. Pada bagian xilem sekunder yang dekat dengan empelur tidak terdapat zona infeksi (Gambar 3b).
Gambar 1a. A.) Penampang melintang batang sengon Wamena bagian periderm (kulit batang) 100x, B.) bagian xilem sekunder (tengah), C.) bagian empelur (bagian tengah batang) 40x. documentasi oleh:
Gambar 1b. D.) Penampang melintang batang sengon Solomon, bagian periderm (kulit batang) 100x, E.) bagian xilem sekunder (tengah), F.) bagian empelur (bagian tengah batang) 40x.
Keterangan: periderm (pr), korteks (c), floem sekunder (sf), pembuluh/sel trakea (v), xilem sekunder (sx), sel parenkim apotrakeal (pa), sel parenkim paratrakeal (pp), xilem primer (px), empelur (pt).
Gambar 2b. Penampang tangensial 100x, A) sengon Wamena, B) sengon Solomon, Penampang radial 100x, C) sengon Wamena, D) sengon Solomon.
Keterangan: sel parenkim paratrakeal (r), serat xilem (s)
Seperti diketahui, kolonisasi fungi karat hanya terjadi pada daerah yang terinfeksi (Widyastuti et al., 2005). Hal ini berhubungan dengan ketahanan tanaman terhadap patogen yang melibatkan berbagai macam tanggapan, salah satunya adalah gum yang berfungsi sebagai penghalang atau barrier sehingga patogen tidak dapat melanjutkan serangannya dan gum yang dibentuk pada pembuluh dapat mencegah pergerakan patogen (Agrios, 1996). Salah satu yang menyebabkan reaksi tersebut
terjadi dikarenakan perkembangan miselia terhenti dan diisolasi di jaringan xilem sekunder (Gramacho et al., 2013; Allen et al., 1990a). Hal ini juga berhubungan dengan pembentukan periderm (Gramacho et al., 2013; Allen et al., 1990b), sedangkan dalam penelitian Allen et al. (1990a), perkembangan periderm di sekitar sel yang terkena infeksi menghasilkan tanin, yang dilaporkan sebagai mekanisme ketahanan.
Gambar 3. A. Penampang melintang batang sengon Solomon yang mengalami infeksi jamur U. tepperianum terdapat zona infeksi berwarna hitam disekitar xilem sekunder (anak panah kuning) dan disekitar pembuluh (anak panah biru) 100x. B. Bagian xilem sekunder dari batang sengon Solomon yang dekat dengan empelur tidak mengalami infeksi 40x.
Keterangan: pembuluh (v), sel parenkim apotrakeal (pa), xilem primer (px), empelur (pt)
Proporsi sel xilem B.
Hasil analisis varians proporsi sel, dari 4 variabel yang dianalisis yaitu serat xilem (db=1, ms= 4,08, Fpr=0,691), sel parenkim paratrakeal (db=1, ms= 43,13, Fpr=0,093), sel parenkim apotrakeal (db=1, ms= 0,422, Fpr=0,535) dan sel trakea (db=1, ms= 0,880, Fpr=0,565),
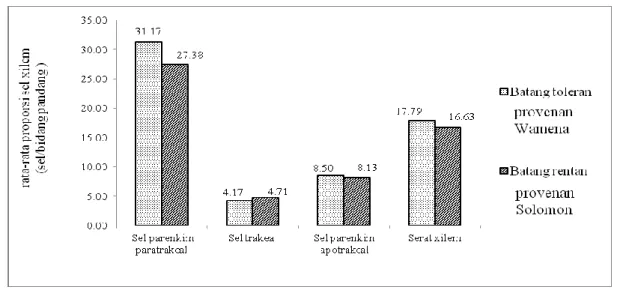
menunjukkan tidak ada perbedaan nyata antara batang yang rentan (Solomon) dengan batang tahan (Wamena). Namun demikian, dari rerata proporsi sel xilem terlihat bahwa batang toleran mempunyai proporsi sel yang lebih banyak dibandingkan batang rentan (Gambar 4).
Gambar 4. Rata-rata proporsi sel penyusun jaringan xilem pada batang sengon Wamena (tahan) dan Solomon (rentan).
Tidak adanya perbedaan struktur anatomi batang rentan (Solomon) dengan batang tahan
(Wamena) dapat disebabkan karena beberapa faktor. Salah satu faktor tersebut adalah
A
kekebalan bawaan yang dimiliki oleh tanaman (Freeman & Beattie, 2008). Pada dasarnya tanaman tahan terhadap infeksi patogen karena tanaman tersebut memang tahan terhadap infeksi patogen. Beberapa pertahanan yang dapat dilakukan merupakan hasil dari perubahan stuktur jaringan atau senyawa-senyawa yang dikeluarkan di daerah sekitar serangan patogen. Penyebab lain adalah karena tanaman tersebut telah terinfeksi oleh patogen namun patogen tersebut mampu membatasi aktivitas patogen sehingga kerusakan yang ditimbulkan tidak berkembang atau tidak meluas ke daerah lain dan tidak mempengaruhi aktivitas inang (Agrios, 1996; Anonim, 2014).
Menurut Rahayu (2008), benih sengon yang diketahui asal usulnya dan berasal dari indukan yang memiliki kualitas yang baik cenderung lebih kuat dan tahan terhadap penyakit karat tumor, sedangkan benih yang tidak diketahui asal usulnya atau benih yang memiliki kualitas rendah lebih rentan terserang penyakit. Namun demikian, berdasarkan hasil penelitian, ketahanan terhadap patogen tidak tertunjukkan dalam struktur anatomi batang khususnya xilem. Hasil analisis menunjukkan bahwa sifat yang dikaji tidak berbeda nyata karena pada batang rentan dan tahan mempunyai proporsi sel penyusun jaringan xilem yang sama, hal ini mengindikasikan tidak terkait dengan cekaman patogen. Kenampakan secara morfologi mungkin terlihat pada batang yang berasal dari Solomon yaitu pada bagian atas pohonnya terdapat pembengkakan karat tumor. Hal ini mungkin disebabkan karena perbedaan genetik sengon dari Solomon lebih rendah. Kepulauan Solomon hanya merupakan pulau-pulau kecil sehingga tegakannya dapat dikategorikan dalam satu provenan, serta memiliki geografis yang sama sehingga adaptasinya sama (Rahayu, 2009; Susanto et al., 2014; Setiadi et al., 2014a).
Penggunaan jaringan xilem ini saja belum mewakili secara keseluruhan. Hal ini
disebabkan perubahan pada jaringan xilem hanya terjadi pada bagian yang mengalami infeksi karena serangan patogen, sehingga pengamatan secara anatomi membutuhkan sifat lain misalnya ketebalan dinding sel. Dinding sel yang mengalami lignifikasi sangat kedap terhadap patogen (Freeman & Beattie, 2008). Lignifikasi dinding sel berhubungan dalam pembentukan jaringan yang tahan terhadap infeksi jamur dan pelukaan tanaman dan dianggap penting dalam proses regenerasi felogen (Biggs et al., 1984 dalam Allen et al., 1990b). Pertahanan juga dapat terjadi pada berkas pembuluh, sel parenkim dan sel sklerenkima. Sel-sel ini menebal dengan proses esterifikasi, lignifikasi dan deposisi suberin. Lignifikasi pada sel sklerenkima pada sekitar berkas pembuluh membantu penebalan dan berfungsi sebagai mekanisme pertahanan.
Jika dilihat dari panjang serat (db=1, ms= 15557, Fpr=0,090) dan diameter serat xilem (db=1, ms= 24,04, Fpr=0,323) antara batang sengon rentan (Solomon) dengan batang sengon tahan (Wamena) hasil analisis varians juga tidak menunjukkan perbedaan nyata. Namun demikian Gambar 5 memperlihatkan bahwa rata-rata panjang dan diameter serat xilem kayu provenan Solomon memiliki panjang dan diameter serat yang lebih tinggi dari pada kayu provenan Wamena. Hal tersebut mengindikasikan bahwa penyakit karat tumor menyebabkan perubahan susunan ukuran serta jumlah sel penyusunnya pada kayu yang terinfeksi dan yang terserang. Seperti yang terjadi pada batang Fraxinus sp. yang terserang kanker, terdapat modifikasi serat yang berombak; sedangkan pada tanaman Quercus
robur yang terserang tumor terjadi perubahan
susunan sel pada batang, serat memiliki panjang lebih pendek dari sel batang normal (Gűlsoy et al., 2005). Menurut Rukhama & Nugroho (2014), serabut pada kayu yang terserang karat tumor lebih pipih dan panjang dari kayu sehat, dan ini diduga karena pengaruh hormon auksin
Gambar 5. Rata-rata panjang dan diameter serat xilem (µm) pada batang sengon Wamena (tahan) dan
Solomon (rentan)
Hasil analis varians juga tidak menunjukkan terdapatnya perbedaan yang nyata pada diameter serat antar sampel pohon Wamena dan Solomon (db=1, ms= 24,04, Fpr=0,323), meskipun rata-rata diameter serat Solomon (36,44 µm) lebih tinggi dari batang Wamena (33,61 µm). Diameter serat xilem sengon memang bervariasi antara penelitian yang satu dengan yang lainnya. Untuk sengon Jawa bervariasi antara 33,74 - 48,55 µm (Martawijaya et al., 1986; Praptoyo, 2001; Manggala, 2013). Lebih lanjut Praptoyo (2001) menyatakan bahwa diameter serat sengon Solomon memiliki nilai rata-rata 35,68 μm, yang umumnya lebih besar dibandingkan dengan sengon lokal.
IV. KESIMPULAN
Hasil penelitian menunjukkan bahwa antara batang sengon yang berasal dari Wamena yang kemungkinan tahan karat tumor dengan batang sengon yang berasal dari Solomon yang kemungkinan rentan karat tumor tidak menunjukkan perbedaan dalam struktur xilemnya. Hal ini terlihat dari tidak terdapatnya perbedaan pada bagian periderm hingga empulur dari sampel uji tersebut. Namun demikian, salah satu contoh dari Solomon (rentan) memperlihatkan zona infeksi yang berwarna hitam pada jaringan xilem sekundernya. Tidak adanya perbedaan tersebut
dapat dikaitkan dengan mekanisme ketahanan tanaman terhadap patogen, maupun sifat genetik yang dimiliki tanaman untuk menghindar atau mengurangi kerusakan yang disebabkan patogen. Hasil penelitian menunjukkan bahwa sel parenkim paratrakeal, apotrakeal, sel trakea, serat xilem, panjang dan diameter serat xilem tidak berbeda nyata antara batang yang rentan dengan batang yang tahan.
UCAPAN TERIMAKASIH
Penulis mengucapkan terimakasih banyak kepada tim penelitian jenis sengon B2P2BPTH Yogyakarta, yang telah menyediakan contoh kayu untuk penelitian ini. Kepada bapak/ibu teknisi B2P2BPTH disampaikan terimakasih atas bantuannya selama pengamatan di laboratorium.
DAFTAR PUSTAKA
Adinugroho, C. W. (2008). Konsep Timbulnya Penyakit Tanaman. Mayor Silvikultur Tropika Sekolah Pascasarjana Institut Pertanian Bogor.
Agrios, G. N. (1996). Ilmu Penyakit Tumbuhan. Yogyakarta: Gajah Mada University Press. Allen, E. A., Blenis. P. V., & Hiratsuka, Y. (1990a).
Early Symptom Development in Lodgepole Pine Seedling Infected with Endocronartium harknessii. Canadian Journal of Botany, 68, 270-277.
Allen, E. A., Blenis. P. V., & Hiratsuka, Y. (1990b). Histological evidence of Resistant to Endocronartium harknessii in Pinus contoria
var. latifolia. Canadian Journal of Botany, 68, 1728-1737.
Anonim. (2014). Bab V Patologi dan Patogenitas. Universitas Gajah Mada. Diakses tanggal 2 November 2014, dari
http://elisa.ugm.ac.id/user/archive/download/ 29095/59708175da718f7d67039d7314983 Baskorowati, L., & Nurrohmah, S.H. (2011). Variasi
Ketahanan Terhadap Penyakit Karat Tumor Pada Sengon Tingkat Semai. Jurnal Pemuliaan Tanaman Hutan, 5(3), 129-138. Baskorowati, L., Susanto, M., & Charomaini, M.
(2012). Genetic Variability in Resistance of Falcataria moluccana (Miq.) Barbeby & J. W. Grimes to Gall Rust Disease. Journal of Forestry Research, 9(1), 1-9.
Charomaini, M. Z., & Burhan, I. (2008). Indikasi Awal Ketahanan Sengon (Falcataria moluccana) Provenan Papua Terhadap Jamur Uromycladium tepperianum Penyebab Penyakit Karat Tumor (Gall Rust). Jurnal Pemuliaan Tanaman Hutan. 2(2), 1-9. Diputra, I. M. M. M. (2015). Respons Sengon
(Falcataria moluccana (Miq.) Barbeby & J. W. Grimes) Provenans Papua dan Ras Lahan Jawa Terhadap Penyakit Karat Tumor (Tesis tidak dipublikasikan). Program Pascasarjana Program Studi Ilmu Kehutanan, UGM. Freeman, B. C., & Beattie, G. A. (2008). An
Overview of Plant Defenses against Pathogens and Herbivores: The Plant Health Instructor. Iowa State University. doi: 10.1094/PHI-I-2008-0226-01
Gramacho, K. P., Thomas, M., & Robert, A. S. (2013). Comparative Histopathology of Host Reaction in Slash Pine Resistant to
Cronartium quercuum f. sp. fusiform. Diakses tanggal 3 November 2014, dari
www.mdpi.com/journal/forest
Gülsoy, S., Eroĝlu., K.H., & Merev, N. (2005). Chemical and Wood Anatomical Properties of Tumorous Wood in A Turkish White Oak (Quercus robursubsp. robur). IAWA Journal, 26(4), 469-476.
Hardiyanto, E. B. (2010). Pemuliaan Pohon Lanjut (Tidak Dipublikasikan). Fakultas Kehutanan, Universitas Gadjah Mada, Yogyakarta. Hopkins, W. G., & Huiiner N. P. A. (2008).
Introduction to Plant Physiology (4th ed.). United States of America: John Wiley & Sons, Inc.
Jewell, F. F., (1988). Histopathology of Fusiform Rust-Inoculated Progeny from (Shortleaf X
Slash) X Shortleaf Pine Crosses. Phytopathology, 78(4), 397-402.
Kasmudjo. (1985). Teknologi Hasil Hutan. Fakultas Kehutanan, Universitas Gajah Mada, Yogyakarta.
Martawijaya, A., Kartasujana, I., Mandang, Y.I., Prawira, S.A., & Kadir, K. (1989). Atlas Kayu Indonesia Jilid II. Bogor: Pusat Penelitian dan Pengembangan Hasil Hutan. Manggala Z.S. (2013). Sifat Fisika dan Dimensi
Serat Kayu Sengon dengan Gejala Tumor di Daerah Cangkringan. (Skripsi tidak dipublikasikan). Fakultas Kehutanan Universitas Gadjah Mada, Yogyakarta. Praptoyo, H. (2001). Studi Proporsi Sel dan Dimensi
Serat pada Arah Aksial dan Radial Kayu Sengon Laut (Paraserianthes falcataria) Salomon. Jurnal Ilmu Dan Teknologi Kayu Tropis, 3(2).
Praptoyo, H., & Puspitasari, R.(2012). Variasi sifat anatomi kayu sengon (Paraserianthes falcataria (L) Nielsen) dari dua jenis pemudaan yang berbeda. Seminar Nasional Mapeki XV 6-7 November 2012 (pp. 33-41), Makassar.
Rahayu, S. (2008). Penyakit karat tumor pada sengon (Falcataria moluccana (Miq.) Barneby & J.W. Grimes). Workshop penanggulangan serangan karat puru pada tanaman sengon 19 Nop 2008. Balai Besar Penelitian Bioteknologi dan Pemuliaan Tanaman Hutan, Yogyakarta.
Rimbawanto, A. (2008). Pemuliaan tanaman dan ketahanan penyakit pada sengon. Workshop penanggulangan serangan karat puru pada tanaman sengon 19 Nop 2008. Balai Besar Penelitian Bioteknologi dan Pemuliaan Tanaman Hutan, Yogyakarta.
Rukhama, S. & Nugroho, W. D. (2014). Anatomi Tumor pada Kayu Sengon Trubusan yang Terserang Jamur Uromycladium tepperianum (Skripsi tidak dipublikasikan). Fakultas Kehutanan, Universitas Gadjah Mada, Yogyakarta.
Salisbury, F. B., & Ross, C.V. (1995). Fisiologi Tumbuhan (D. R. Lukman, Trans.). Bandung: Penerbit ITB.
Setiadi, D., Susanto, M., & Baskorowati, L. (2014a). Ketahanan serangan penyakit karat tumor pada uji keturunan sengon di Bondowoso, Jawa Timur. Jurnal Pemuliaan Tanaman Hutan, 8(1), 121-136.
Setiadi, D., Susanto, M., & Baskorowati, L. (2014b). Pertumbuhan sengon Solomon dan responnya
terhadap penyakit karat tumor di Bondowoso, Jawa Timur. Jurnal Pemuliaan Tanaman Hutan, 8(2), 1-13.
Soerianegara, I., & Lemmens, R.H.M.J. (1993) Plant resources of South-East Asia 5(1): Timber trees: major commercial timbers. Belanda, Wageningen: Pudoc Scientific Publishers. Susanto, M., Baskorowati, L. & Setiadi, D. (2014).
Estimasi Peningkatan Genetik Falcataria moluccana di Cikampek Jawa Barat. Jurnal Hutan Tanaman, 11(2), 85-76.
Widyastuti, S. M., Sumardi & Harjono. (2005). Patologi Hutan. Yogyakarta: Gadjah Mada University Press.
Yamamoto, F., Nakamura, K., & Hiratsuka, Y. (1998). Is Ethylene a Trigger af Stem Hyperplasia Caused by Eastern Gall Rust In Pinus densiflora. Research Papers 712: 243-251. Pfoc. First IUFRO Rusts Of Forest Trees WP Conf., 2-7 Aug. Saariselklj: Finland Finnish Forest Research Institute. Zalasky, H. (1976). Xylem in galls of lodgepole pine
caused by western gall rust, Endocronartium harknessii. Canadian Journal of Botany, 54, 1586-1590.
PENGARUH MEDIA ORGANIK SEBAGAI MEDIA SAPIH TERHADAP KUALITAS
BIBIT BIDARA LAUT (Strychnos lucida R. Brown)
The effect of Organic Media as growing media on seedling quality of Bidara Laut (Strychnos
lucida R. Brown)
Anita Apriliani Dwi Rahayu, dan Resti Wahyuni
Balai Penelitian dan Pengembangan Teknologi HHBK
Jl. Dharmabhakti No. 7, Ds. Langko, Kec. Lingsar, Lombok Barat NTB e-mail: anita_forester03@yahoo.co.id
Tanggal diterima: 20 Maret 2015, Tanggal direvisi: 24 Maret 2015, Disetujui terbit: 10 Juni 2016
ABSTRACT
Timber utilization of Strychnos lucida R. Brown as a medicine caused uncontrolled exploitation. Currently unknown nursery technique appropriate to Strychnos lucida. The aim of this research wastesting the organic media such as eceng gondok compost, rice skin ash and cocopeat as growing media so was produced the best quality of Strychnos lucida seedling. The research used Randomized Complete Block Design with four treatments: B0 (top soil + organic compost (1:1)), B1 (eceng gondok compost + rice husk ash + cocopeat (2:2:1)), B2 (eceng gondok compost + rice husk ash + cocopeat (2:1:2)), and B3 (eceng gondok compost + rice husk ash + cocopeat (1:2:2)). The results showed that growing media only significantly influence on the total of leave, while others parameter such as life percentage, height, diameter, root/shoot ratios (R/S), seedling quality index (QI), and seedling sturdiness were not significantly influence. Duncan test showed average of total of leave was significantly different. The best of total of leave obtained in media B0 (7.4 strands).
Keywords: growth, organic media, seedling, Strychnos lucida R. Brown
ABSTRAK
Pemanfaatan kayu bidara laut (Strychnos lucida R. Brown) sebagai obat menyebabkan eksploitasi yang kurang terkendali. Saat ini belum diketahui teknik pembibitan yang tepat untuk jenis bidara laut. Tujuan penelitian ini adalah mengujimedia organik seperti kompos eceng gondok, arang sekam dan cocopeat sebagai media sapih, sehingga dihasilkan kualitas bibit bidara laut yang terbaik. Rancangan yang digunakan adalah Rancangan Acak Lengkap Berblok dengan empat perlakuan media sapih yaitu B0 (top soil + kompos organik (1:1)), B1 (kompos eceng gondok + arang sekam + cocopeat (2:2:1)), B2 (kompos eceng gondok + arang sekam + cocopeat (2:1:2)), dan B3 (kompos eceng gondok + arang sekam + cocopeat (1:2:2)). Hasil penelitian menunjukkan bahwa perlakuan media sapih hanya berpengaruh nyata terhadap jumlah daun, sedangkan parameter lain seperti persentase hidup, tinggi, diameter, nisbah pucuk akar, indeks mutu bibit dan kekokohan semai tidak berpengaruh nyata. Uji Duncan menunjukkan nilai rata-rata jumlah daun berbeda nyata. Jumlah daun terbaik didapatkan pada media B0 (7,4 helai).
Kata kunci: media organik, pertumbuhan, semai, Strychnos lucida R. Brown
I. PENDAHULUAN
Bidara laut (Strychnos lucida R. Brown) sudah terkenal sejak dahulu sebagai bahan pembuatan obat tradisional. Tumbuhan ini mengandung zat yang berguna untuk obat malaria, diabetes, darah tinggi, kurang darah, gangguan pencernaan, cacar air, kurang nafsu makan, penguat lambung, ejakulasi dini, dan lainnya (Tim PNPM-MP Dompu, tanpa tahun). Hasil penelitian Balai Penelitian Kehutanan Mataram tahun 2009 menunjukkan bagian yang
paling banyak mengandung khasiat obat adalah kulit kayu bidara laut dibandingkan bagian tanaman lainnya (Hasan et al., 2011).
Semula masyarakat hanya memanfaatkan biji bidara laut untuk dijadikan sebagai obat. Saat ini, pemanfaatan bidara laut sebagai obat tidak hanya digunakan secara tradisional akan tetapi sudah banyak dijadikan sebagai peluang bisnis. Para pengusaha memanfaatkan kayu bidara laut untuk dibuat cangkir/gelas. Hal ini yang menyebabkan eksploitasi kayu songga di kawasan hutan di Kabupaten Dompu kurang
terkendali (Hasan et al., 2010). Oleh karena itu, upaya pelestarian bidara laut perlu segera dilakukan sebelum jenis ini menjadi jenis yang langka. Salah satu upaya yang dapat dilakukan adalah dengan kegiatan budidaya.
Salah satu faktor yang menentukan keberhasilan budidaya adalah bibit yang digunakan berkualitas baik. Bibit yang berkualitas baik dihasilkan dari teknik pembibitan yang tepat. Sampai saat ini belum diketahui teknik pembibitan yang tepat untuk jenis bidara laut. Penelitian yang pernah dilakukan oleh Nandini & Agustarini (2011) menunjukkan pembibitan bidara laut dari biji menggunakan media sapih tanah dan pasir (3:2) menghasilkan persen hidup 62,67% pada umur semai 7 bulan. Hal ini menunjukkan penggunaan media tanah masih belum dapat mengoptimalkan pertumbuhan bibit.
Media top soil mempunyai banyak kekurangan yaitu berat per satuan bibit lebih tinggi, sifat fisik medium lebih jelek, kadang-kadang mengandung hama atau penyakit, dan pengambilan dalam skala luas dapat merusak ekosistem di aeral pengambilan (Hendromono, 2003b). Alternatif media yang dapat digunakan untuk pembuatan bibit adalah media organik. Bahan-bahan organik yang bersifat limbah lebih murah dan mudah didapatkan, serta bobot persatuan lebih rendah dibandingkan top soil. (Hendromono, 1998 dalam Putri, 2008).
Terkait permasalahan kelestarian jenis bidara laut di alam dan beberapa keunggulan media organik dibandingkan media top soil sebagai media pembibitan, maka penelitian ini dilaksanakan dengan tujuan memperoleh media sapih yang sesuai untuk pertumbuhan bibit bidara laut dari berbagai media organik yang diuji sehingga diperoleh bibit yang berkualitas tinggi.
II. BAHAN DAN METODE Waktu dan Lokasi
A.
Penelitian ini dilaksanakan di persemaian Balai Penelitian Teknologi Hasil Hutan Bukan
Kayu di Kabupaten Lombok Barat, NTB.Benih diambil dari Kabupaten Dompu dan Bima, Pulau Sumbawa, NTB. Kegiatan penelitian dilakukan selama 6 bulan, yaitu mulai bulan Juli sampai dengan bulan Desember 2013.
Bahan dan Alat B.
1. Bahan
Bahan yang diperlukan adalah benih bidara laut, media perkecambahan (pasir 100%) dan media penyapihan (kompos eceng gondok, arang sekam, cocopeat, top soil dan kompos organik).
2. Alat
Peralatan yang digunakan meliputi bak kecambah, polybag, kantong plastik, penggaris, kaliper, label, shading net, kamera, dan lain-lain.
Metode Pengamatan C.
Benih didapatkan dari ekstraksi buah dengan cara memisahkan biji dari kulit dan daging buahnya. Perkecambahan dilakukan dengan menabur benih pada bak kecambah yang berisi media pasir 100%. Penyapihan dilakukan pada kecambah yang telah memiliki dua pasang daun. Empat macam perlakuan media sapih yaitu:
- B0 = kontrol (top soil + kompos organik (1:1))
- B1 = kompos eceng gondok + arang sekam + cocopeat (2:2:1)
- B2 = kompos eceng gondok + arang sekam + cocopeat (2:1:2)
- B3 = kompos eceng gondok + arang sekam + cocopeat (1:2:2)
Pengukuran parameter pertumbuhan dan mutu bibit dilakukan dua minggu sekali. Parameter yang diukur/diamati terdiri atas:
1. Persentase hidup
Persentase hidup bibit dihitung dari keseluruhan jumlah bibit yang masih hidup sampai akhir penelitian dibandingkan dengan jumlah seluruh bibit yang ditanam pada awal penelitian.
2. Tinggi dan diameter batang bibit
Tinggi bibit diukur dari permukaan tanah sampai pucuk tanaman. Diameter batang diukur pada bagian bawah (± 2 cm di atas permukaan tanah).
3. Jumlah daun
Jumlah daun dihitung sudah ada berapa helai daun yang muncul sampai akhir pengamatan.
4. Nisbah pucuk akar (NPA)
Nisbah pucuk akar (NPA) adalah perbandingan berat kering pucuk (batang dan daun) dengan berat kering akar yang diukur pada akhir pengamatan dengan rumus (Darwo & Sugiarti, 2008):
Keterangan:
BKP = Berat Kering Pucuk BKA = Berat Kering Akar
5. Kekokohan bibit
Rumus yang digunakan untuk mendapatkan nilai kekokohan bibit (Hendromono, 2003 dalam Junaedi et al., 2010) adalah sebagai berikut:
6. Indeks mutu bibit
Penilaian angka indeks mutu morfologis bibit dilakukan dengan rumus Indeks Mutu Bibit (Dickson, et al., 1960 Dalam Putri, 2008) yaitu indeks yang menyatakan tinggi rendahnya mutu bibit tanaman. Cara perhitungan Indeks Mutu Bibit adalah sebagai berikut: Keterangan:
BKP = Berat Kering Pucuk BKA = Berat Kering Akar T = Tinggi
D = Diameter
Rancangan D.
Penelitian ini dilakukan di Green House menggunakan Rancangan Acak Lengkap Berblok, dengan 3 macam perlakuan media dan 1 kontrol yang dibuat dalam 3 blok (ulangan). Tiap ulangan terdiri dari 24 semai, sehingga total semai yang diamati sebanyak 4 perlakuan media x 3 blok x 24 semai = 288 semai. Blok (ulangan) berfungsi untuk meminimalisir galat percobaan (Gomez & Gomez, 2007) akibat perbedaan kemiringan lahan pada tempat persemaian.
Analisis Data E.
Untuk mengetahui pengaruh perlakuan dan blok terhadap parameter yang diamati, maka dilakukan analisis varians. Khusus untuk parameter persentase hidup dilakukan tranformasi terlebih dahulu sebelum dianalisis dengan menggunakan transformasi Arcsin. Model yang digunakan dalam analisis varians adalah sebagai berikut (Ott & Longnecker, 2015):
Yij = µ + ri + βj + εij
Keterangan:
Yij = rata-rata pengamatan pada blok ke-i dan media ke-j; µ = rata-rata umum;
ri = pengaruh blok ke-i; βj = pengaruh media ke-j;
εij = galat.
Apabila hasil analisis varians menunjukkan perbedaan nyata, maka dilakukan uji lanjut dengan metode Duncan Multiple
Range Test (DMRT) pada taraf uji 5%.
III. HASIL DAN PEMBAHASAN Pertumbuhan bibit
A.
Pertumbuhan bibit adalah komponen penting dalam menentukan kualitas suatu bibit. Pertumbuhan bibit biasanya terbagi menjadi dua yaitu pertumbuhan atas permukaan tanah (pucuk/tajuk) dan pertumbuhan bawah permukaan tanah (akar) (Junaedi et al., 2010).
Pertumbuhan bibit di permukaan dapat dilihat dari persentase hidup bibit, tinggi dan diameternya. Berdasarkan hasil analisis varians,
semua parameter pertumbuhan bibit tidak berbeda nyata pada blok yang berbeda. Hal ini menunjukkan pengelompokkan perlakuan media
sapih ke dalam blok-blok tidak dapat meningkatkan keberagaman pertumbuhan bibit antar blok.
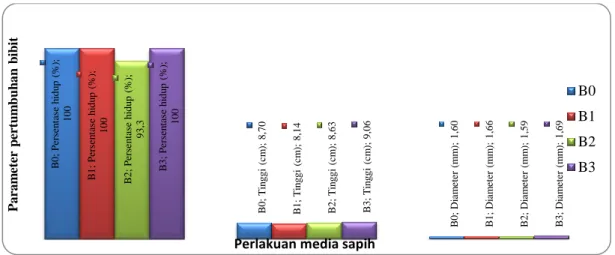
Gambar 1. Rata-Rata persentase hidup, tinggi dan diameter bibit Bidara Laut pada perlakuan media sapih
Persentase hidup pada semua perlakuan media sapih menunjukkan tidak ada perbedaan yang nyata. Perlakuan media organik sebagai media sapih juga tidak berpengaruh nyata
meningkatkan pertumbuhan tinggi dan diameter bibit bidara laut, secara statistik dapat dilihat pada hasil analisis varians yang tidak berbeda nyata (Tabel 1).
Tabel 1. Hasil analisis varians perlakuan media sapih terhadap persentase hidup, tinggi, dan diameter bibit Bidara Laut umur 2 bulan setelah disapih
Parameter Sumber variasi
Blok Media sapih
Persentase Hidup 1,038ns 0,321ns
Tinggi 0,487ns 0,675ns
Diameter 3,815ns 0,889ns
Keterangan: ns = tidak berpengaruh nyata
Kandungan unsur hara yang hampir sama pada semua media perlakuan menyebabkan pertumbuhan tinggi dan diameter tidak berbeda
nyata (Tabel 2). Begitu juga untuk parameter persentase hidup, pada semua menunjukkan hasil yang tidak berbeda nyata.
Tabel 2. Rerata persentase hidup, tinggi, dan diameter bibit Bidara Laut umur 2 bulan setelah sapih
Perlakuan Persentase Hidup Tinggi Diameter
(%) (cm) (mm)
B0 100,00±0,00 8,70±0,70 1,60±0,16
B1 100,00±0,00 8,14±1,13 1,66±0,17
B2 93,33±2,89 8,63±0,28 1,59±0,01
B3 100,00±0,00 9,06±0,59 1,69±0,07
Salah satu faktor lingkungan yang menentukan pertumbuhan tanaman adalah unsur hara yang tersedia pada media tanam (Winarso, 2005). Hasil analisis media organik perlakuan
didapatkan kandungan N antara 0,85-0,98% atau tergolong sangat tinggi, sedangkan media kontrol (top soil + kompos) kandungan N nya rendah (0,20%). Kandungan P media organik
B 0; P er se n tas e h idu p (% ); 100 B 0; T in g gi ( cm ); 8, 70 B 0; Di am eter ( m m ); 1, 60 B 1; P er se n tas e h idu p (% ); 100 B 1; T in g gi ( cm ); 8, 14 B 1; Di am eter ( m m ); 1, 66 B 2; P er se n tas e h idu p (% ); 93, 3 B 2; T in g gi ( cm ); 8, 63 B 2; Di am eter ( m m ); 1, 59 B 3; P er se n tas e h idu p (% ); 100 B 3; T in g gi ( cm ); 9, 06 B 3; Di am eter ( m m ); 1, 69 Pa ra m ete r p er tu m bu h a n b ib it
Perlakuan media sapih
B0 B1 B2 B3
tergolong sangat rendah (0,41-0,47 ppm), sedangkan media kontrol sangat tinggi (100,15 ppm). Kandungan K pada semua media tergolong sangat tinggi (1,07-2,77 Cmol/kg). Dari hasil tersebut dapat dilihat bahwa media organik mengandung hara yang cukup, sama dengan media top soil untuk menunjang pertumbuhan bibit. Tingkat pertumbuhan bibit yang terbaik pada media organik menunjukkan bahwa media organik efektif untuk pertumbuhan fisiologis tanaman (Ahmadloo et al., 2012). Hanya saja kekurangan pada media organik perlakuan adalah kandungan P nya yang rendah. Unsur P sangat berperan untuk pertumbuhan dan metabolisme tanaman (Munawar, 2011), sehingga jika akan
mengaplikasikan media organik perlu meningkatkan kandungan P. Jumlah P dapat ditingkatkan dengan penambahan pupuk atau mikroorganisme seperti bakteri pelarut fosfat (Noor, 2003).
Kualitas bibit B.
Kualitas bibit merupakan gambaran bibit yang diharapkan mampu beradaptasi dan tumbuh pada suatu kondisi tertentu (Nurhasybi & Sudrajat, 2006). Kualitas bibit dapat dilihat dari sifat morfologis dan fisiologisnya. Pada penelitian ini kualitas bibit dilihat dari parameter jumlah daun, Nisbah Pucuk Akar, kekokohan bibit dan Indeks Mutu Bibitnya.
Gambar 2. Rata-rata jumlah daun, nisbah pucuk akar, indeks mutu bibit, dan kekokohan bibit Bidara Laut pada perlakuan media sapih
Hasil analisis varians pengaruh pengelompokkan (blok) terhadap parameter kualitas bibit menunjukkan tidak berpengaruh nyata. Hal yang sama terjadi pada paramater pertumbuhan bibit, pembuatan blok-blok perlakuan juga tidak memberikan keragaman terhadap parameter kualitas bibit. Oleh karena itu, blok dianggap tidak diperlukan untuk meminimalisir perbedaan yang akan ditimbulkan akibat perbedaan kemiringan tempat persemaian.
Hasil analisis varians pada perlakuan media terhadap beberapa parameter kualitas bibit menunjukkan hanya jumlah daun yang berbeda nyata. Hal ini menunjukkan perlakuan media sapih berpengaruh terhadap perbedaan
jumlah daun yang muncul pada bibit bidara laut, sedangkan parameter kualitas bibit yang lain tidak terpengaruh dengan adanya perbedaan media sapih.
Unsur hara yang mempengaruhi pertumbuhan dan perkembangan daun pada tanaman adalah nitrogen (Lakitan, 1996 dalam Fatimah &Handarto, 2008). Bentuk nitrogen yang diserap tanaman dari dalam tanah (top
soil) sudah siap digunakan untuk tanaman,
sedangkan pada bahan organik tanaman memerlukan waktu untuk memanfaatkan unsur hara yang terkandung di dalamnya. Hal ini yang dapat menyebabkan jumlah daun semai pada media kontrol lebih banyak dibandingkan campuran media organik.
B 0 ; Ju ml ah Dau n ; 7 ,4 0 B 0 ; NP A ; 0 ,8 9 B 0 ; IM B ; 0 ,0 8 B 0 ; K ek o k o h an ; 5 ,4 4 B 1 ; Ju ml ah D au n ; 6 ,5 5 B 1 ; N P A ; 0 ,9 7 B 1 ; IM B ; 0 ,1 0 B 1 ; K ek o k o h an ; 4 ,9 1 B2 ; Ju m lah D au n ; 6 ,1 4 B 2 ; NP A ; 0 ,9 2 B 2 ; IMB ; 0 ,0 8 B 2 ; K ek o k o h an ; 5 ,4 9 B 3 ; Ju ml ah D au n ; 6 ,2 8 B 3 ; N P A ; 0 ,8 8 B 3 ; IM B ; 0 ,1 5 B 3 ; K ek o k o h an ; 5 ,3 5 P a ra m et er k ua lita s bib it
Perlakuan media sapih
B0 B1 B2 B3
Kadar C/N juga sangat mempengaruhi pertambahan jumlah daun bibit bidara laut pada media organik. Kandungan C/N pada media organik tergolong sangat tinggi, hal ini mengindikasikan media yang belum matang atau belum terdekomposisi dengan sempurna (Danu & Kurniaty, 2013). Bahan organik yang
belum terdekomposisi akan mengalami dekomposisi yang dilakukan oleh bakteri di dalam medium. Akibatnya kandungan nitrogen medium berkurang karena digunakan bakteri untuk kelangsungan hidupnya (Hendromono, 2003b).
Tabel 3. Hasil analisis varians perlakuan media sapih terhadap jumlah daun, NPA, IMB, dan kekokohan bibit Bidara Laut umur 2 bulan setelah disapih
Parameter Sumber variasi
Blok Media sapih
Jumlah daun 0,190ns 5,672*
Nisbah Pucuk Akar 2,833ns 0,206ns
Kekokohan bibit 3,192ns 1,289ns
Indeks Mutu Bibit 1,277ns 1,251ns
Keterangan: * = berpengaruh nyata pada taraf 5%; ns= tidak berpengaruh nyata
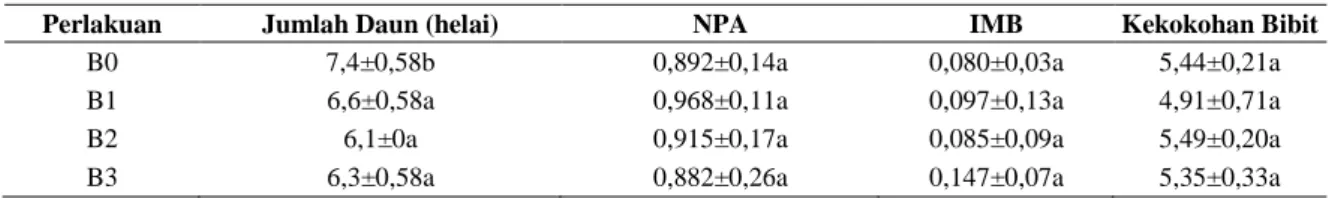
Tabel 4. Rerata jumlah daun, NPA, IMB dan kekokohan bibit Bidara Laut umur 2 bulan setelah sapih
Perlakuan Jumlah Daun (helai) NPA IMB Kekokohan Bibit
B0 7,4±0,58b 0,892±0,14a 0,080±0,03a 5,44±0,21a B1 6,6±0,58a 0,968±0,11a 0,097±0,13a 4,91±0,71a
B2 6,1±0a 0,915±0,17a 0,085±0,09a 5,49±0,20a
B3 6,3±0,58a 0,882±0,26a 0,147±0,07a 5,35±0,33a
Keterangan: angka yang diikuti huruf yang sama pada kolom yang sama menunjukkan tidak berbeda nyata
Tabel 5. Kandungan hara media perlakuan Media semai
Kandungan hara dalam media N-total (%) C-organik (%) C/N rasio P-tersedia (ppm) K-total (Cmol/kg) B0 0,20 1,78 8,90 100,15 1,07 B1 0,93 37,67 40,51 0,41 1,78 B2 0,85 38,14 44,87 0,43 1,78 B3 0,98 32,18 32,84 0,47 2,77
Parameter kualitas bibit lainnya seperti Nisbah Pucuk Akar, kekokohan bibit dan Indeks Mutu Bibit menunjukkan nilai yang tidak berbeda nyata. NPA menunjukkan kondisi fisiologis tanaman karena NPA mencerminkan nilai total produksi pertumbuhan yaitu berat kering pucuk dan perakarannya (Sukendro & Sugiarto, 2012). Besarnya berat kering pucuk akan menyebabkan nilai NPA yang besar pula. Seperti hasil penelitian Sukendro & Sugiarto (2012), pertumbuhan daun yang lebih banyak akan meningkatkan nilai NPA. Hal ini tidak terlihat jelas pada hasil penelitian ini. Pada media B0, meskipun jumlah daun tertinggi tidak
menunjukkan nilai NPA yang tertinggi, begitu juga pada media B2 yang menunjukkan jumlah daun paling sedikit, nilai NPAnya masih lebih tinggi dibandingkan dengan media B0.
Selain itu, besar kecilnya Nisbah Pucuk Akar juga menunjukkan kesiapan semai untuk dipindahkan ke lapangan (Danu et al., 2006). Menurut klasifikasi Duryea & Brown (1984) dalam Darwo & Sugiarti (2008) bahwa pertumbuhan dan kemampuan hidup semai terbaik terjadi pada Nisbah Pucuk Akar antara 1-3. NPA semai Bidara Laut pada semua perlakuan media sapih di bawah nilai minimum yaitu < 1. Nilai NPA yang paling mendekati
nilai 1 terlihat pada semai yang disapih pada media kompos eceng gondok + arang sekam +
cocopeat (2:2:1) yaitu 0,968. Nilai NPA yang
hanya di bawah 1 menunjukkan semai belum siap untuk ditanam di lapangan. Hal ini dikarenakan semai baru berumur 2 bulan setelah di sapih. Pada umur 2 bulan semai masih membutuhkan air dan unsur hara dalam jumlah yang besar untuk mendukung pertumbuhan, sehingga biasanya akar akan lebih panjang dan lebih banyak.
Nilai kekokohan bibit pada semua perlakuan media tidak menunjukkan perbedaan yang signifikan. Tidak adanya perbedaan yang nyata pada nilai kekokohan semai dikarenakan tinggi dan diameter semai juga tidak berbeda secara nyata pada semua perlakuan media sapih. Seperti dijelaskan Junaedi et al., (2010) bahwa kekokohan bibit ditentukan oleh besaran dan variasi dari tinggi dandiameter bibit. Nilai kekokohan bibit pada semua media berkisar antara 4,91-5,49. Bibit dengan nilai kekokohan semai lebih dari 6 sangat tidak diharapkan untuk ditanam. Semakin kecil nilai kekokohan semai maka semakin kokoh semai/bibit (Jaenicke, 1999 dalam Yudohartono & Fambayun, 2013). Nilai kekokohan bibit yang optimum adalah mendekati nilai 4-5 (Adinugraha, 2012). Hal ini dapat dikatakan bibit Bidara Laut pada semua media telah memenuhi nilai kekokohan bibit yang optimum.
Indeks Mutu Bibit menunjukkan bibit sudah siap untuk ditanam di lapangan. Nilai Indeks Mutu Bibit pada keempat perlakuan media tidak menunjukkan perbedaan yang signifikan. Hal ini dikarenakan IMB dipengaruhi oleh besarnya berat kering total, kekokohan bibit dan NPA (Junaedi et al., 2010). Nilai kekokohan bibit dan NPA yang tidak berbeda nyata pada semua perlakuan media menyebabkan nilai IMB pada semua media sapih juga tidak berbeda nyata.
Menurut Lackey & Alm (1982) dalam Hendromono (2003a), IMB minimal bibit agar dapat bertahan hidup yaitu sebesar 0,09. Nilai IMB yang memenuhi nilai minimal untuk
ditanam di lapangan adalah semai pada perlakuan media B1 (kompos eceng gondok + arang sekam + cocopeat (2:2:1)) dan B3(kompos eceng gondok + arang sekam +
cocopeat (1:2:2)) yaitu 0,097 dan 0,147.
Meskipun nilai kekokohan bibit pada bibit Bidara Laut umur 2 bulan dan Indeks Mutu Bibit pada perlakuan media B1 dan B3 telah memenuhi standar minimal bibit bisa ditanam di lapangan, akan tetapi nilai NPA nya belum menunjukkan kesiapan bibit di lapangan.
IV. KESIMPULAN DAN SARAN Kesimpulan
A.
Perlakuan media sapih berpengaruh nyata terhadap jumlah daun tetapi tidak memberikan pengaruh yang nyata terhadap persentase hidup, tinggi, diameter, nisbah pucuk akar, kekokohan semai dan indeks mutu bibit. Media perlakuan yang menunjukkan nilai rata-rata jumlah daun terbanyak didapatkan pada perlakuan kontrol (B0) yaitu 7,4 helai. Nilai rata-rata jumlah daun menunjukkan perbedaan, sedangkan persentase hidup, tinggi, diameter nisbah pucuk akar, kekokohan semai dan indeks mutu bibit tidak menunjukkan perbedaan.
Saran B.
Limbah bahan organik seperti eceng gondok, sekam padi, dan serbuk sabut kelapa
(cocopeat) dapat digunakan sebagai alternatif
media sapih jenis Bidara Laut , dengan mempertimbangkan komposisi media. Penambahan pupuk atau mikroorganisme diperlukan untuk meningkatkan unsur hara pada media organik guna mendapatkan pertumbuhan tanaman yang optimal.
UCAPAN TERIMA KASIH
Ucapan terima kasih disampaikan kepada Gipi Samawandana yang telah membantu selama pelaksanaan kegiatan penelitian.
DAFTAR PUSTAKA
Adinugraha, H. A. (2012). Pengaruh Cara Penyemaian dan Pemupukan NPK terhadap Pertumbuhan Bibit Mahoni Daun Lebar di Persemaian. Jurnal Pemuliaan Tanaman Hutan, 6(1), 1-10.
Ahmadloo, F., Tabari, M., Yousefzadeh, H., Kooch, Y., & Rahmani, A. (2012). Effect of Soil Nutritional Status on Seedling Nursery Performance of Arizona cypress (Cupressus arizonica var arizonica Greene) and Medite cypress (Cupressus sempervirens var. Horizantalis (Mill.) Gord). Afr. J. Plant Sci., 6(4), 140-149.
Awang, Y., Shaharom, A. S., Mohamad, R. B., & Selamat, A. (2009). Chemical and Physical Characteristics of Cocopeat-Based Media Mixtures and Their Effects on the Growth and Development of Celosia cristata Am. J. Agric. Biol. Sci., 4, 63-71.
Bayu. (n.d.). Ragam Informasi Tanaman Hias: Sekam Bakar Bagi Adenium. Diakses tanggal 4 Juli 2008, dari
http://tabloidgallery.wordpress.com/2008/07/ 04/sekam-bakar-bagi-adenium/.
Danu, Sudrajat, D. J., Verawati & Suhardi, E. (2006). Pengaruh Komposisi Media terhadap Pertumbuhan Bibit Sentang (Azadirachta exelsa (Jack) Jakob) Asal Cabutan di Persemaian. Prosiding Seminar Hasil-Hasil Penelitian (pp.109-115). Pusat Penelitian dan Pengembangan Hutan Tanaman.
Danu & Kurniaty, R. (2013). Pengaruh Media dan Naungan terhadap Pertumbuhan Pembibitan Gerunggang (Cratoxylom arborescens (Vahl) Blume). Jurnal Perbenihan Tanaman Hutan, 1(1), 43-50.
Darwo & Sugiarti. (2008). Pengaruh Dosis Serbuk Spora Cendawan Scleroderma citrinum Persoon dan Komposisi Media terhadap Pertumbuhan Tusam di Persemaian. Jurnal Penelitian Hutan dan Konservasi Alam, 5(5),461-472.
Deng, X. K., Yin, W., Li, W. D., Yin, F. Z., Lu, X. Y., Zhang, X. C., ... Cai, B. C. (2006). The Anti-Tumor Effect of Alkaloids from The Seeds of Strychnos nux-vomica on HepG2 cells and Its Possible Mechanism. Journal of Ethnopharmacology, 106(2), 179-186. Fatimah, S. & Handarto, B. M. (2008). Pengaruh
Komposisi Media Tanam terhadap Pertumbuhan dan Hasil Tanaman Sambiloto (Andrographis paniculata, Nees). Jurnal Embryo, 5(2), 133-148.
Gharagozloo, P., Lazareno, S., Popham, A. & Birdsall, N. J. M. (1999). Allosteric Interactions of Quaternary Strychnine and Brucine Derivatives with Muscarinic Acetycholine Receptors. J. Med. Chem, 42(3), 438-445.
Gomez, K. A. & Gomez, A. A.(2007). Prosedur Statistik untuk Penelitian Pertanian (edisi kedua). Jakarta: Penerbit Universitas Indonesia.
Hasan, R. A., Nandini, R. & Wahyuni, N. (2011). Kajian Pemanfaatan Tanaman Bidara Laut (Strychnos lucida) oleh Masyarakat di Kabupaten Dompu dan Buleleng. Prosiding Workshop: Sintesa Hasil Penelitian Hutan Tanaman (pp. 353-358). Bogor: Pusat Litbang Peningkatan Produktivitas Hutan. Hasriani, Kalsim, D. K. & Sukendro, A. (n.d.)
Kajian Serbuk Sabut Kelapa (Cocopeat) sebagai Media Tanam. Diakses tanggal 18 Desember 2013, dari
http://dedikalsim.files.wordpress.com/2013/1 2/jurnal-hasriani-ed-dkk-nov-2013.pdf. Hendromono (2003a). Kriteria Penilaian Mutu Bibit
Dalam Wadah yang Siap Tanam untuk Rehabilitasi Hutan dan Lahan. Buletin Penelitian dan Pengembangan Kehutanan , 4(1),11-20.
Hendromono (2003b). Peningkatan Mutu Bibit Pohon Hutan dengan Menggunakan Medium Organik dan Wadah Yang Sesuai. Buletin Penelitian dan Pengembangan Kehutanan, 4(2), 135-143.
Junaedi, A., Hidayat, A., & Frianto, D. (2010). Kualitas Fisik Bibit Meranti Tembaga (Shorea leprosula Miq.) Asal Stek Pucuk Pada Tiga Tingkat Umur. Jurnal Penelitian Hutan dan Konservasi Alam, 7(3), 281-288. Kurniaty, R., Budiman, B., & Suartana, M.
(2010).Pengaruh Media dan Naungan terhadap Mutu BibitSuren (Toona sureni Merr.). Jurnal Penelitian Hutan Tanaman, 7(2), 77-83.
Munawar, A. (2011). Kesuburan Tanah dan Nutrisi Tanaman. Bogor: IPB Press.
Nandini, R., & Agustarini, R. (2011). Teknik Budidaya Tanaman Bidara Laut (Strychnos lucida R.Br) Secara Generatif. Prosiding Workshop: Sintesa Hasil Penelitian Hutan Tanaman (pp. 359-368). Bogor: Pusat Litbang Peningkatan Produktivitas Hutan. Noor, A. (2003). Pengaruh Fosfat Alam dan
Kombinasi Bakteri Pelarut Fosfat dengan Pupuk Kandang terhadap P Tersedia dan
Pertumbuhan Kedelai pada Ultisol. Bul. Agron, 31(3), 100-106.
Nugroho, D. S. (2011). Kajian Pupuk Organik Enceng Gondok terhadap Pertumbuhan dan Hasil Bayam Putih dan Bayam Merah (Skripsi tidak dipublikasikan). Universitas Sebelas Maret.
Ott, R. L. & Longnecker, M. (2015). An Introducation to Statistical Methods and Data Analysis (7th ed.). USA: Cengage Learning.
Putri, A. I. (2008). Pengaruh Media Organik terhadap Indeks Mutu Bibit Cendana. Jurnal Pemuliaan Tanaman Hutan, 21(1), 1-8. Sittadewi, E. H. (2007). Pengolahan Bahan Organik
Eceng Gondok Menjadi Media Tumbuh untuk Mendukung Pertanian Organik. Jurnal Teknologi Lingkungan, 8(3), 229-234. Sudomo, A. & Santosa, H. B. (2011). Pengaruh
Media Organik dan Tanah Mineral terhadap Pertumbuhan dan Indeks Mutu Bibit Mindi (Melia azedarach L.). Jurnal Penelitian Hutan dan Konservasi Hutan, 8(3), 263-271. Sukendro, A. & Sugiarto, E. (2012). Respon
Pertumbuhan Anakan Shorea leprosula Miq, Shorea mecistopteryx Ridley, Shorea ovalis (Korth) Blume dan Shorea selanica (DC) Blume terhadap Tingkat Intensitas Cahaya Matahari. Jurnal Silvikultur Tropika, 3(1), 22-27.
Tim PNPM-MP Dompu. (n.d.). KSPP Songga Menembus Pasar Luar Negeri: dengan SPP, Menembus Pasar Luar Negeri. Diakses 21 Maret 2014, dari http://pnpmmpd- ntb.blogspot.com/2013/10/kspp-songga-menembus-pasar-luar-negeri.html.
Winarni, E. (2008). Respon Pertumbuhan Semai Eucalyptus pellita terhadap Perbedaan Komposisi Bokhasi Eceng Gondok (Eichhornia crassipes Mort Solm) dan Top Soil. Jurnal Hutan Tropis Borneo, 23, 116-120.
Winarso, S. (2005). Kesuburan Tanah: Dasar Kesehatan dan Kualitas Tanah. Yogyakarta: Gava Media.
Wikipedia. (2014). Brucine. Diakses 23 Oktober 2014, dari
http://en.wikipedia.org/wiki/Brucine. Wikipedia. (2015). Strychnine. Diakses 27 Februari
2015, dari
http://en.wikipedia.org/wiki/Strychnine.
Yudohartono, T. P. & Fambayun, R. A. (2013). Karakteristik Pertumbuhan Semai Binuang Asal Provenan Pasaman Sumatera Barat. Jurnal Pemuliaan Tanaman Hutan, 6(3), 143-156.
Zuraida, Sukito, A. & Darmawan, S. (2012). Conservation and Protection of Songga Tree (Strychnos lucida R Brown) as Rare and Valuable Tree Species – A Case Study in Sumbawa Island, Indonesia. Proceeding IUFRO World Series Vol.30 (pp. 46-48). Vienna, Austria: IUFRO Headquarters.
KERAGAMAN GENETIK CENDANA
PADA TEGAKAN PENGHASIL BENIH DAN TEGAKAN REHABILITASI
DI NUSA TENGGARA TIMUR BERDASARKAN PENANDA ISOZIM
Genetic Diversity of Sandalwood on Seed Production Stand and Rehabilitation Stand in East
Nusa Tenggara Based on Isozyme Marker
Rini Purwiastuti, Sapto Indrioko, dan Eny Faridah
Fakultas Kehutanan, Universitas Gadjah Mada Jl. Agro No. 1 Bulaksumur, Yogyakarta, 55281
e-mail: rinipurwiastuti@gmail.com
Tanggal diterima: 27 Agustus 2015, Tanggal direvisi: 29 September 2015, Disetujui terbit: 30 Juni 2016
ABSTRACT
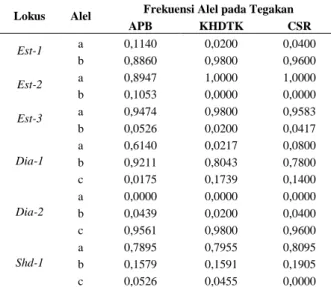
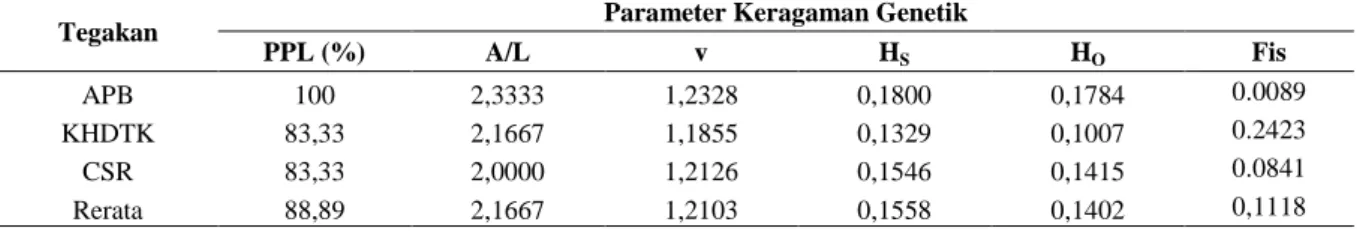
This study aimed to determine the genetic diversity of sandalwood on seed production stand and rehabilitation stand in East Nusa Tenggara using isozyme genetic marker. The study was conducted using samples collected from three stands, i.e. Seed Production Area (APB) representing seed production stand, KHDTK rehabilitation stand and CSR rehabilitation stand representing rehabilitation stands. Samples of sandalwood juvenile leaves are taken randomly from each location. Sample materials taken were juvenile leaves collected randomly from each stand. There were 57 samples taken from APB, while each 25 samples were collected from rehabilitation stands of KHDTK and CSR. Isozymes analyses were carried out in the laboratory using three kinds of enzyme systems i.e Esterase (EST), Diaphorase (DIA) and Shikimate Dehydrogenase (SHD). The results showed that for genetic diversity within stands, the mean of polymorphic loci was 88.89%, with a mean number of alleles per locus 2.1667 and a mean of effective alleles 1.2103. The expected heterozygosity within stands (HS) was 0.1558, with the observed heterozygosity (HO) of 0.1402, while the mean index of fixation (FIS) was 0.1118. On genetic diversity among stands, DST, and GST values were 0.0090 and 0.0545 respectively, while total expected heterozygosity of the three stands (HT) was 0.1648. To anticipate sandalwood genetic diversity decline, it is essential to identify and record the remaining sandalwood populations, then conserve rare alleles either through in-situ or ex-situ conservation programs.
Keywords: sandalwood, Seed Production Area, rehabilitation stands, genetic diversity, isozyme
ABSTRAK
Penelitian ini bertujuan untuk mengetahui keragaman genetik cendana pada tegakan penghasil benih dan tegakan rehabilitasi di Nusa Tenggara Timur dengan menggunakan isozim sebagai penanda genetik. Penelitian dilakukan menggunakan sampel yang diambil dari 3 tegakan yaitu Areal Produksi Benih (APB) yang mewakili tegakan penghasil benih, tegakan rehabilitasi Kawasan Hutan Dengan Tujuan Khusus (KHDTK) dan tegakan rehabilitasi Corporate Social Responsibility (CSR) yang mewakili tegakan rehabilitasi. Sampel berupa daun juvenil cendana diambil secara random dari masing-masing lokasi. Dari tegakan APB diambil sebanyak 57 sampel sedangkan dari tegakan rehabilitasi KHDTK dan CSR masing-masing 25 sampel. Analisis isozim dilakukan di laboratorium menggunakan 3 macam sistem enzim yaitu Esterase (EST), Diaphorase (DIA) dan Shikimate Dehydrogenase (SHD). Parameter keragaman genetik dalam tegakan menunjukkan rerata lokus polimorfik sebesar 88,89%, dengan rerata jumlah alel per lokus 2,1667 dan rerata alel efektif 1,2103. Heterozigositas harapan (HS) dalam tegakan diperoleh nilai 0,1558 dengan rerata heterozigositas teramati (HO)
sebesar 0,1402 dan rerata indeks fiksasi (FIS) sebesar 0,1118. Pada pengamatan parameter keragaman genetik
antar tegakan diperoleh nilai DST = 0,0090 dan GST = 0,0545, sedangkan heterozigositas harapan total ketiga
tegakan cendana (HT) sebesar 0,1648). Untuk mengantisipasi penurunan keragaman genetik cendana perlu
dilakukan identifikasi dan inventarisasi keberadaan populasi cendana yang masih tersisa, untuk kemudian dilakukan upaya konservasi bagi alel-alel langka baik secara in-situ maupun ex-situ.