LAPORAN GENETIKA (BI-2105)
ISOLASI DNA PLASMID pET-32b(+) DAN AMPLIFIKASI
GEN phoR
Tanggal Praktikum : 30 Oktober 2015 – 13 November 2015 Tanggal Pengumpulan : 20 November 2015
disusun oleh : Dhia Shofi S 10614032 Kelompok 9 Asisten : Annisa Rizkia 10613016 Muh. Agung Saputra 10613040
PROGRAM STUDI BIOLOGI
SEKOLAH ILMU DAN TEKNOLOGI HAYATI INSTITUT TEKNOLOGI BANDUNG
BANDUNG 2015
BAB I PENDAHULUAN
1.1 Latar Belakang
Isolasi DNA merupakan proses pemurnian DNA dari sampel menggunakan kombinasi metode fisik dan kimiawi. Tujuan dilakukannya isolasi DNA adalah untuk mendapatkan DNA yang benar-benar murni untuk kemudian dilakukan investigasi lebih lanjut. DNA sendiri merupakan materi genetik yang diwariskan dan bersifat spesifik pada setiap individu dan dapat diaplikasikan pada kepentingan identifikasi pelaku kejadian (kepentingan forensik), identifikasi penyakit, dan penyisipan gene of interest untuk diamplifikasi (Dahm, 2008).
Salah satu DNA yang banyak digunakan untuk kepentingan ini adalah DNA plasmid pada bakteri. DNA genom tidak digunakan untuk proses amplifikasi dengan mesin PCR karena ukuran DNA genom lebih besar dibanding DNA Plasmid. Selain itu, DNA plasmid juga bersifat independen yang artinya bisa menduplikasi dirinya sendiri terlepas dari kegiatan DNA genom. Selain itu, DNA plasmid lebih mudah ditransfer dengan proses transformasi yang memungkinkan penyisipan gen ke organisme lain seperti ke tanaman karena berukuran lebih kecil (Lipps, 2008).
Amplifikasi gen merupakan metode perbanyakan suatu gen dengan cara polymerase chain reaction (PCR). Amplifikasi gen dengan metode PCR bekerja spesifik yang bisa segera mengenali daerah sampel DNA yang terkena translokasi. Amplifikasi gen dilakukan untuk menganalisis suatu sampel gen dengan jumlah yang sangat sedikit. Amplifikasi gen dapat digunakan untuk mengetahui agen penginfeksi dan diskriminasi strain patogen dan non-patogen, diagnosis awal penyakit malignant seperti
leukemia dan limfoma, perbanyakan gen yang diinginkan, serta untuk mengidentifikasi sampel DNA berumur ribuan tahun.
Elektroforesis adalah perpindahan materi dari katoda menuju anoda didalam daerah elektrik. Faktor-faktor yang mempengaruhi perpindahan partikel saat elektroforesis adalah ukuran DNA, konformasi DNA, konsentrasi gel, dan jumlah muatan diberikan. Elektroforesis dilakukan untuk memisahkan DNA dan protein. Contoh aplikasi dari elektroforesis adalah untuk mengidentifikasi korban dan pelaku berdasarkan hasil band DNA, melihat kecocokan DNA orang sehat dengan yang mengalami mutasi. Selain itu, elektroforesis dapat digunakan untuk mengetahui apakah gene of interest yang diisolasi berhasil diamplifikasi atau tidak (Robyt & White, 1990).
1.2 Tujuan
Tujuan dari praktikum kali ini adalah :
1. Mengisolasi DNA plasmid pET 32 dengan metode alkali lisis
2. Mengamplifikasi Gen phoR dari DNA plasmid pET 32 yang diisolasi dengan metode PCR
BAB II
TINJAUAN PUSTAKA
2.1 Plasmid
Plasmid merupakan molekul DNA kecil di dalam sel yang secara fisik terpisah dari DNA kromosom dan bisa bereplikasi secara independen. Plasmid biasanya ditemukan pada bakteri sebagai bentuk yang kecil, sirkular, double stranded. Plasmid juga kadang ditemukan pada archaea dan organisme eukariot. Plasmid bisa memiliki panjang dari 1 hingga 1000 kbp (Lederberg, 1952).
Plasmid digunakan sebagai vektor perbanyakan/ekspresi suatu gen. Gen yang akan diamplifikasi disisipkan ke dalam plasmid. Setelah itu, plasmid dimasukkan ke dalam bakteri melalui proses transformasi dan bakteri dikultur. Selain untuk mengekspresikan gen tertentu, plasmid juga digunakan untuk membuat protein dalam jumlah besar (Lederberg, 1952).
Perbedaan DNA plasmid dengan DNA genom ada pada ukuran, bentuk, organisasi konformasi, dan kandungan yang dibawa. DNA plasmid memiliki ukuran sekitar 1-1000 kbp, sementara DNA genom memiliki panjang 160-12.200 kbp pada prokaryot. DNA plasmid berbentuk sirkular, sementara DNA genom berbentuk linear pada eukaryot, namun bisa sirkular pada bakteri. Organisasi dari DNA plasmid kebanyakan memiliki organisasi konformasi supercoiled, sedangkan DNA genom memiliki organisasi konformasi melingkar di sekitar histon. DNA genom mengandung informasi penting bagi kehidupan organisme, sementara DNA plasmid mengandung informasi tambahan bagi organisme tersebut (Tranbichler & Shapiro, 2006).
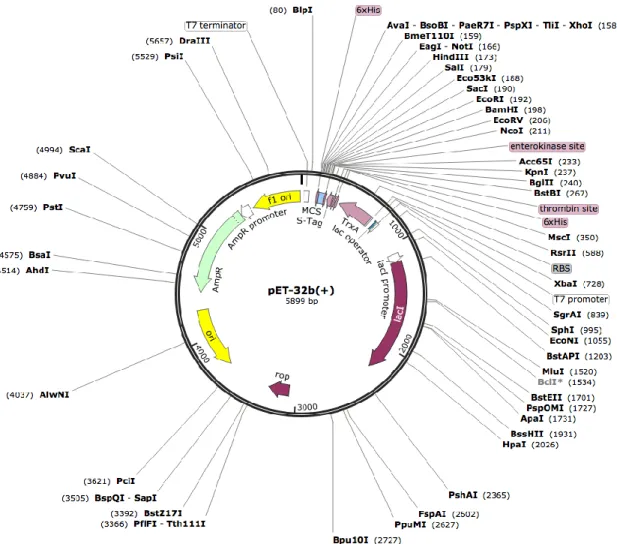
Pada percobaan isolasi DNA digunakan DNA plasmid pET-32b(+). DNA plasmid pET-32b(+) memiliki panjang 5899 bp. Vektor ini memiliki peta seperti pada gambar 2.1.
Gambar 2.1 Peta DNA Plasmid pET-32b(+) (LaVallie, et al., 1993)
Vektor ini memiliki beberapa sequence landmarks, yaitu T7 promoter 764-780 bp, T7 transcription start 763 bp, Trx•Tag coding sequence 366-692 bp, His•Tag coding sequence 327-344 bp, S•Tag coding sequence 249-293 bp, Multiple cloning sites, (Nco I - Xho I) 158-217 bp, His•Tag coding sequence 140-157 bp, T7 terminator 26-72 bp, lacI coding sequence 1171-2250 bp, pBR322 origin 3684 bp, bla coding sequence 4445-5302 bp, f1 origin 5434-5889 bp (LaVallie, et al., 1993).
DNA Plasmid bisa muncul dalam bentuk - bentuk tertentu yang disebut konformasi. Ada 5 tipe konformasi pada DNA plasmid yakni:
1. Nicked open-circular DNA yang memiliki satu strand yang dipotong (Kroll, et al., 2010)
2. Relaxed circular DNA yang memiliki bentuk sirkular utuh, tidak terpotong sama sekali kedua strandnya, namun telah direlaksasi secara enzimatik (tidak supercoiled) (Kroll, et al., 2010)
3. Linear DNA yang memiliki ujung bebas, bisa jadi karena kedua strand telah dipotong atau karena DNA linear in vivo (Kroll, et al., 2010)
4. Supercoiled (covalently closed-circular) dimana DNA memiliki kedua strand yang tidak terpotong sama sekali dengan pilinan yang terintegrasi, yang menghasilkan bentuk yang kompak (Kroll, et al., 2010)
5. Supercoiled denatured DNA, memiliki bentuk seperti supercoiled namun memiliki area yang tidak berpasangan yang membuatnya jadi kurang kompak yang bisa jadi terjadi karena suasana yang terlalu basa saat preparasi plasmid (Kroll, et al., 2010)
2.2 PCR (Polymerase Chain Reaction)
Polymerase Chain Reaction atau PCR merupakan suatu teknik untuk mengamplifikasi satu atau beberapa copy dari sebuah DNA dengan prinsip thermal cycling (Bartlett & Stirling, 2003). Thermal cycling merupakan siklus pengulangan pemanasan dan pendinginan untuk reaksi pelelehan DNA dan replikasi enzimatik DNA. Prinsip kerjanya adalah DNA dipanaskan atau didinginkan pada temperature tertentu yang merupakan temperature efektif untuk setiap proses enzimatiknya. Proses enzimatik itu terdiri dari beberapa tahapan yakni: inisiasi, denaturasi, annealing, ekstensi, dan penyimpanan. Tahap denaturasi, annealing, dan ekstensi dilakukan secara berulang kali dalam suatu siklus dimana 1 siklus = elongasi + denaturasi + annealing + elongasi sehingga dengan demikian jumlah gen yang akan diamplifikasi meningkat berdasar deret eksponensial 2n dengan n merupakan jumlah siklus (Logan, et al., 2009)
2.3 Elektroforesis
Elektroforesis merupakan metode pemisahan partikel-partikel bermuatan di dalam medan listrik yang homogen. Pada elektroforesis terdapat beberapa komponen yakni chamber elektroforesis, sumber tegangan, gel, dan materi yang akan dielektroforesis (protein, RNA, DNA). Materi yang akan dielektroforesis seperti DNA ditaruh pada well di gel elektroforesis (agarosa / polyacrilamide gel) yang berada dalam chamber berisi buffer. Kemudian chamber dihubungkan dengan sumber tegangan. Prinsip kerja dari elektroforesis ini adalah sumber tegangan akan menghasilkan suatu medan listrik homogen, di dalam medan listrik ini, DNA akan mengalami gaya gerak listrik (ggl) karena muatan yang dimilikinya dan tempatnya yang berada di dalam medan listrik. DNA akan bergerak ke arah kutub medan yang memiliki muatan berlawanan dengannya (Robyt & White, 1990).
Elektroforesis bisa memisahkan partikel-partikel karena perbedaan kecepatan partikel tersebut dalam menempuh matriks elektroforesis (gel). Ada beberapa hal yang dapat mempengaruhi kecepatan suatu partikel menempuh gel elektroforesis yakni:
a. Ukuran DNA
Ukuran DNA menentukan kecepatan pergerakan pada gel. Molekul DNA yang lebih besar akan lebih tertahan pada gel, sementara molekul yang lebih kecil lebih tidak tertahan sehingga akan menempuh gel lebih cepat (Lucotte & Baneyx, 1993).
b. Konformasi DNA
DNA yang memiliki konformasi supercoiled akan bergerak lebih cepat daripada DNA relaxed. Hal ini dapat terjadi karena DNA supercoiled tergulung secara ketat sehingga lebih kompak dan memudahkan untuk bergerak lebih cepat (Lucotte & Baneyx, 1993). c. Konsentrasi gel
Konsentrasi gel menentukan ukuran pori gel yang mempengaruhi migrasi DNA. Jika konsentrasi gel semakin besar, pori pada gel akan
semakin kecil sehingga lebih menahan DNA dan mengurangi kecepatan migrasinya. Agarose dengan konsentrasi tinggi baik untuk memisahkan partikel DNA kecil, sementara agarose dengan konsentrasi tinggi untuk memisahkan DNA berukuran besar (Lucotte & Baneyx, 1993).
d. Medan listrik yang diberikan
Pada voltase rendah, migrasi DNA proporsional dengan besarnya tegangan yang diberikan, yakni semakin besar tegangan, semakin cepat DNA bergerak. Namun, pada medan listrik yang meningkat, mobilitas fragmen DNA berat akan meningkat secara diferensial, dan keefektifan pemisahan menuruh dan resolusinya kemudian lebih rendah pada voltase tinggi (Lucotte & Baneyx, 1993).
BAB III METODOLOGI
3.1 Alat dan Bahan
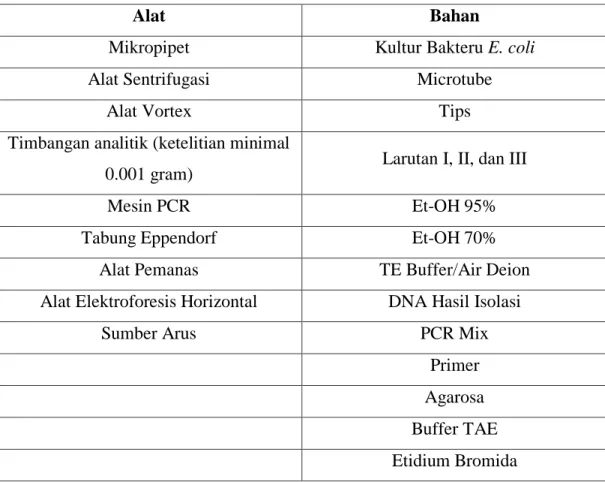
Alat dan bahan yang digunakan dalam praktikum isolasi DNA, PCR, dan elektroforesis terdapat dalam Tabel 3.1.
Tabel 3.1 Alat dan bahan praktikum isolasi DNA, PCR, dan elektroforesis
Alat Bahan
Mikropipet Kultur Bakteru E. coli Alat Sentrifugasi Microtube
Alat Vortex Tips
Timbangan analitik (ketelitian minimal
0.001 gram) Larutan I, II, dan III
Mesin PCR Et-OH 95%
Tabung Eppendorf Et-OH 70%
Alat Pemanas TE Buffer/Air Deion Alat Elektroforesis Horizontal DNA Hasil Isolasi
Sumber Arus PCR Mix
Primer Agarosa Buffer TAE Etidium Bromida
3.2 Cara Kerja
3.2.1 Isolasi DNA Plasmid
Kultur bakteri E.coli diambil sebanyak 1000 µL dengan mikropipet. Kemudian kultur dimasukkan ke dalam microtube 1.5 mL. Setelah itu microtube disentrifugasi dengan kecepatan maksimum selama 1 menit. Dari hasil sentrifugasi akan didapatkan supernatan dan pellet. Bagian supernatant
dibuang kemudian pellet dikeringkan. Setelah itu, pelet ditambahkan 200 µl solution I. Setelah itu microtube divortex, lalu ditambahkan 200 µL solution II. Micortube kemudian dibolak-balik hingga larutan homogen. Setelah itu ditambahkan 150 µL solution III dan dibolak-balik lagi hingga larutan homogen. Berikutnya microtube kembali disentrifuga dengan kecepatan maksimum selama 5 menit. Didapatkan kembali pellet dan supernatan dari hasil sentrifuga. Supernatan hasil sentrifuga dipisahkan ke dalam microtube baru. Setelah itu supernatan ditambahkan ethanol absolut bervolume dua kali lebih banyak dengan volume larutan plasmid, kemudian divortex dan didiamkan dua menit. Kemudian microtube disentrifugasi dengan kecepatan maksimum selama 15 menit pada suhu ruang. Kembali didapatkan supernatan dan pellet. Supernatan dibuang dan pellet ditambahkan 0.5 mL alkohol 70% kemudian dibolak-balik hingga homogen dan disentrifugasi lagi dengan kecepatan maksimum selama 5 menit pada suhu ruang. Setelah itu kembali didapatkan supernatan dan pellet. Supernatan dibuang sementara pellet disimpan sampai alkohol menguap. Setelah itu pellet ditambahkan 30 µL nuclease free water kemudian divortex dan diinkubasi pada temperatur -20oC. 3.2.2 PCR (Polymerase Chain Reaction)
Mula-mula DNA target diambil sebanyak 1 µL. Kemudian dimasukkan ke dalam microtube dan diencerkan dengan air deion sampai total larutan 25 µL. Setelah itu larutan ditambahkan PCR mix sebanyak 10 µL. Lalu larutan dihomogenisasi dan kemudian disentrifuga. Setelah itu, microtube dimasukkan ke dalam mesin PCR kemudian didenaturasi awal pada suhu 95oC selama 5 menit. Setelah itu DNA memasuki siklus denaturasi pada suhu 95oC selama 30 detik, annealing pada suhu 65oC selama 30 detik, dan pemanjangan primer pada suhu 72oC selama 7 menit 45 detik. Siklus dilakukan sebanyak 25x.
3.2.3 Elektroforesis
Agarosa dicampurkan dengan buffer TAE dengan volume sama dengan agarosa hingga konsentrasi akhir 0.3%. Setelah itu agarosa dididihkan di atas pemanas kemudian didinginkan. Selanjutnya campuran agarosa ini dituangkan
pada cetakan gel yang telah diletakkan comb di atasnya. Kemudian agarosa didiamkan hingga mengeras. Sebelum mendingin, ditambahkan etidium bromide kedalam agarosa. Setelah agarosa mengeras, comb dilepaskan dari cetakan gel. Setelah itu cetakan gel diletakkan pada tangki elektroforesis. Selanjutnya tangki elektroforesis ditambahkan buffer TAE sampai 1 mm di atas gel. Kemudian, DNA target diambil sebanyak 10 µL kemudian diletakkan pada parafilm dengan mikropipet. Setelah itu, loading buffer diambil dengan mikropipet sebanyak 2 µL kemudian diletakkan di parafilm tepat di atas DNA target yang diambil. Selanjutnya kedua larutan diresuspensi dengan cara menarik dan mengeluarkan kembali kedua cairan dengan mikropipet hingga homogen. Setelah larutan homogen, larutan dimasukkan ke dalam well agarosa pada tangki elektroforesis dengan mikropipet. Setelah itu, tangki elektroforesis dihubungkan dengan generator 100V dan DNA dielektroforesis selama + 25 menit. Selanjutnya, generator dimatikan dan pencetak gel dikeluarkan dari tangki elektroforesis. Terakhir, hasil elektroforesis gen dapat dilihat di bawah sinar UV.
BAB IV
HASIL PENGAMATAN DAN PEMBAHASAN
4.1 Hasil
Hasil pengamatan untuk isolasi DNA plasmid pET-32b(+) ditunjukkan pada gambar 4.1. Untuk hasil elektroforesis gen phoR yang telah di-PCR ditunjukkan pada gambar 4.2.
Gambar 4.1 Hasil Foto Elektroforesis DNA Plasmid pET-32b(+) (dokumentasi pribadi,
2015)
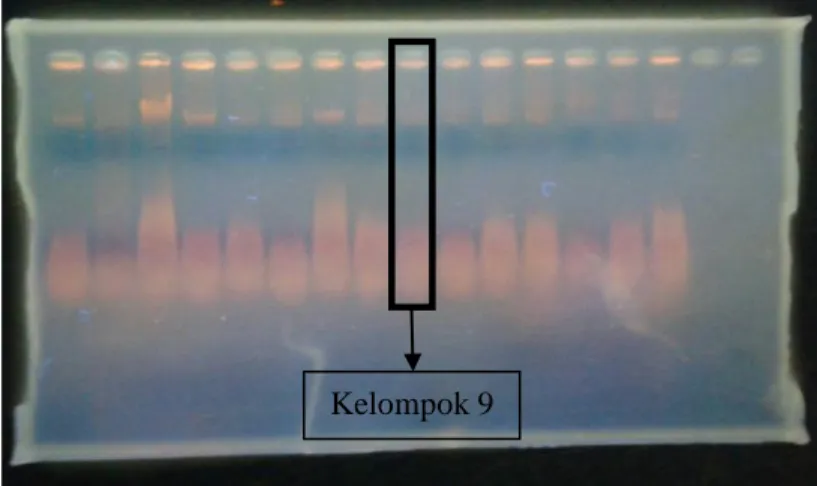
Gambar 4.2 Hasil Foto Elektroforesis Amplifikasi Gen phoR (dokumentasi pribadi,
2015) Kel 9
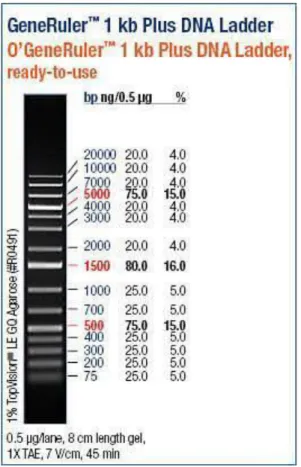
Gambar 4.3 Ukuran DNA Ladder 1 kb
4.2 Pembahasan
4.2.1 Metode Isolasi dengan Alkali Lisis
Dalam percobaan kali ini, digunakan metode alkali lisis untuk mengisolasi DNA plasmid. Metode alkali lisis adalah metode pemisahan menggunakan tiga larutan, yaitu larutan I, II, dan III yang masing-masing memiliki fungsi di tiap tahap penambahan. Metode alkali lisis meliputi beberapa tahapan sebagai berikut:
1. Sentrifugasi
Kultur bakteri yang telah dipanen dimasukkan ke dalam microtube kemudian disentrifugasi. Dari hasil sentrifugasi akan didapatkan bagian pellet (bagian padat) dan supernatan (bagian cair). Bagian pellet merupakan sel bakteri yang akan diisolasi plasmidnya, sementara bagian supernatan adalah media tumbuh
bakteri tersebut. Supernatan dibuang sementara bagian pelletnya diambil agar tersisa bagian sel bakterinya saja (Oswald, 2014).
2. Resuspensi
Pada tahap ini, pellet hasil sentrifugasi diresuspensi dengan larutan I yang berisi Tris, EDTA, glukosa, dan RNase A. Fungsi dari EDTA adalah untuk menangkap kation divalent seperti Mg2+ dan Ca2+ yang berfungsi dalam
aktivitas DNase dan mengintegrasikan sel bakteri. Dengan adanya EDTA, aktivitas DNase untuk menghancurkan plasmid dapat dihambat serta dinding sel diganggu kestabilannya. Glukosa berfungsi untuk menjaga tekanan osmotik sehingga sel tidak meledak. Tris atau lengkapnya tris(hydroxymethyl)aminomethane merupakan buffer pH dengan rumus molekul (HOCH2)3CNH2. Tris memiliki pKa 8.07 pada suhu 25oC dan
memiliki range pH efektif antara 7.07 dan 9.07. Sementara RNase A berfungsi untuk mendegradasi RNA tepat setelah sel lisis. Setelah itu, pellet divortex. Tujuan dari proses ini adalah untuk menghomogenisasi pellet dengan larutan I (Oswald, 2014).
3. Lisis
Pada tahap ini, ditambahkan larutan II ke dalam pellet. Larutan II berisi basa natrium hidroksida (NaOH) dan detergent natrium dodecyl sulfat (SDS). SDS berfungsi untuk melarutkan membran sel serta mendenaturasi protein-protein dalam sel yang berfungsi kemudian untuk pemisahan protein-protein dengan DNA. Sementara itu, NaOH berfungsi untuk menghancurkan dinding sel, namun fungsi utamanya adalah untuk mengganggu ikatan hidrogen diantara basa-basa nitrogen DNA sehingga mengubah DNA double stranded menjadi DNA single stranded. Setelah itu, microtube dibolak-balik untuk dibuat homogen. Pada tahap ini, untuk menghomogenisasi larutan tidak digunakan metode vortex karena jika microtube diguncang terlalu kuat, maka DNA kromosomal yang sudah berada di luar sel bisa terpecah menjadi fragmen-fragmen kecil sehingga bisa mengkontaminasi hasil plasmid (Oswald, 2014).
4. Netralisasi
Pada tahap ini larutan ditambahkan larutan III yang berisi potassium (kalium) asetat (CH3COOK). Tujuan dari penambahan larutan II ini adalah
untuk mengurangi alkalinitas dari larutan karena potassium asetat bersifat asam dalam air. Dalam kondisi ini, hubungan antara basa-basa DNA single stranded bisa terbentuk kembali sehingga ssDNA bisa berenaturasi menjadi dsDNA. Bagian ini merupakan bagian yang selektif. DNA plasmid akan mudah untuk merenaturasi karena ukurannya yang kecil, sementara DNA kromosomal akan sulit terenaturasi karena ukurannya yang sangat besar. Pada bagian ini untuk menghomogenisasi tidak boleh digunakan metode vortex, sehingga microtube hanya dibolak-balik saja secara manual. Tujuannya adalah untuk mencegah putusnya DNA kromosomal menjadi fragmen-fragmen kecil yang bisa terenaturasi. Selain berfungsi untuk mengasamkan larutan, K+ dalam potassium asetat berfungsi untuk menangkap ion dodecyl sulphate pada larutan (Oswald, 2014).
dsPlasmid mudah larut dalam larutan, sementara DNA kromosomal, SDS, protein terdenaturasi akan berinteraksi secara hidrofobik membentuk endapan. Untuk mempermudah terbentuknya endapan, dilakukan sentrifugasi pada larutan. Hasilnya adalah supernatan yang mengandung DNA genom dan pellet yang mengandung DNA kromosom, SDS, dan protein terdenaturasi. Setelah itu, supernatant dipisah dengan pellet dan diambil supernatannya (Oswald, 2014).
5. Pencucian
Bagian supernatant yang telah didapat dari proses netralisasi masih belum mengandung DNA plasmid saja, tetapi masih mengandung garam-garam, EDTA, RNase dan protein residual selular sehingga masih perlu dibersihkan. Cara pembersihannya yaitu pertama-tama supernatan hasil sentrifugasi ditambahkan isopropanol. Tujuan dari tahapan ini adalah untuk melarutkan protein-protein sisa yang bersifat nonpolar, sementara DNA yang bersifat polar akan tertinggal di bawah. Setelah itu, diinkubasi pada suhu -80oC, tujuannya adalah untuk mempercepat reaksi isopropanol dengan campuran.
Setelah itu, campuran disentrifugasi untuk mempercepat pemisahan DNA dengan protein-protein sisa tersebut. Selanjutnya DNA dibersihkan dengan etanol yang teruji efektif meningkatkan konsentrasi DNA dari 0.1 menjadi 0.5 M. Etanol mengubah struktur DNA sehingga molekul DNA beragregasi dan mengendap dari larutan, sementara itu molekul-molekul organik dan garam-garam terlarut pada etanol sehingga tetap berada pada fasa cair. Campuran ini bisa dipisahkan dengan cara sentrifugasi dimana kemudian terdapat supernatan dan pellet. Pellet inilah yang mengandung DNA sementara supernatan hanya mengandung molekul-molekul organik lain dan garam-garam. Pellet dipisahkan dengan supernatan (Oswald, 2014).
6. Penyimpanan
Pellet DNA yang telah didapatkan disentrifugasi quick spin pada tahap ini. Tujuan dilakukan quick spin adalah untuk mengeringkan DNA. DNA yang telah benar-benar kering kemudian dicampurkan dengan larutan TE pH 8.0. TE atau tris-EDTA merupakan buffer pH yang komposisinya terdiri dari 10mM tris pH 8.0 dengan HCl, dan 1mM EDTA. Menurut penelitian, pada pH 8.0 ini DNA menjadi kurang aktif, sehingga pH ini efektif untuk penyimpanan DNA. SEmentara itu EDTA berfungsi untuk mengikat kation metal seperti magnesium dan kalsium yang mungkin masih tertinggal di larutan untuk mencegah aktivitas enzimatis. Setelah itu, larutan disimpan pada temperatur -20oC dengan tujuan untuk mengurangi aktivitas DNA sehingga DNA tidak terdegradasi dan lebih tahan lama (Oswald, 2014).
Pada hasil percobaan, hasil elektroforesis isolasi DNA Plasmid pET-32b(+) menyatakan bahwa DNA Plasmid berhasil diisolasi. Tetapi, panjang dari DNA Plasmid pET-32b(+) tidak dapat dibaca karena tidak adanya gene ladder yang dapat dijadikan acuan pembacaan panjang DNA. Dari literatur, panjang dari DNA Plasmid pET-32b(+) adalah 5899 bp (LaVallie, et al., 1993). Terlihat pula dari gambar, panjang dari DNA yang dielektroforesis tidak sepanjang kelompok lain. Hal ini bisa disebabkan saat DNA dipipet, DNA tidak seluruhnya terambil karena jumlahnya yang sangat sedikit. Saat DNA ditaruh pada gel bisa saja masih terdapat DNA yang tertinggal pada tips
mikropipet ataupun gel tidak seluruhnya masuk ke dalam well namun ada yang terlepas ke larutan buffer karena kurang berhati-hati dalam prosedur.
4.2.2 Amplifikasi PCR
Pada percobaan PCR digunakan beberapa reagen sebagai berikut beserta fungsinya:
1. DNA target
DNA target merupakan DNA yang mengandung area yang akan diamplifikasi atau dengan kata lain rantai DNA yang mengandung gene of interest (Stock, et al., 2009).
2. Primer
Primer merupakan rantai nukleotida yang berfungsi sebagai starting point untuk sintesis DNA yang komplemen dengan ujung 3’ dari sense dan antisense DNA target. Primer digunakan untuk replikasi DNA sebab enzyme yang mengkatalisis proses ini, DNA polymerase hanya bisa menambahkan nukleotida baru pada strand DNA yang telah ada. Polymerase memulai replikasi pada ujung 3’ dari primer. Primer yang digunakan untuk amplifikasi gen biasanya memiliki panjang tidak lebih dari 30 nukleotida dan memiliki ujung yang sama dari awal dan akhir fragmen DNA yang akan diamplifikasi (Stock, et al., 2009).
3. Dream Taq polymerase
Dream Taq polymerase adalah DNA polymerase yang termostabil yang diisolasi dari bakteri Thermus aquaticus. Thermus aquaticus merupakan bakteri yang hidup di air panas dan pintu-pintu hidrotermal sehingga DNA polymerasenya tahan terhadap denaturasi pada suhu tinggi. DNA polymerase ini digunakan untuk menambahkan nukleotida pada rantai DNA yang akan direplikasi. Temperatur optimum untuk Taq ini adalah 75-80oC dan bisa mereplikasi 1000 bp DNA dalam waktu kurang dari 10 detik pada temperatur 72oC (Chien, et al., 1976).
4. dNTPs (deoxy-Nucleotide Tri-Phospates)
dNTPs merupakan suatu molekul yang mengandung suatu nukleosida yang terikat pada 3 fosfat. Nukleosida merupakan suatu kompleks gula pentose yang terikat pada satu basa nitrogen. Gula pentosa pada dNTPS merupakan gula deoxyribosa yang merupakan gula ribosa yang kehilangan 1 atom oksigennya. dNTPs terdiri atas dATP (deoxyadenosine triphospate), dTTP (deoxytimine triphopate), dGTP (deoxyguanine triphospate), dan dCTP (deoxycytocine triphospate). dNTPs ini berfungsi sebagai building blocks dalam replikasi DNA. dNTPs akan direkatkan ke ujung 3’ primer oleh DNA polymerase dan saat proses tersebut berlangsung, dNTPs akan kehilangan 2 gugus fosfatnya (Alberts, et al., 2002).
5. Larutan buffer
Larutan buffer merupakan larutan yang mengandung campuran dari asam lemah dan basa konjugasinya atau basa lemah dan asam konjugasinya. Larutan buffer akan bergeser sangat sedikit pHnya ketika sejumlah asam atau basa kuat ditambahkan ke dalamnya sehingga digunakan untuk mencegah perubahan pH pada larutan. Larutan buffer digunakan untuk menjaga pH pada suatu nilai yang hampir konstan. Pada percobaan digunakan tris-HCl yang memiliki pKa 8.06 dan memiliki buffer range 7.5-9.0, pH larutan = 8.0 (Oswald, 2014).
6. MgCl2
Magnesium klorida (MgCl2) adalah suatu garam yang akan terurai menjadi
ion magnesium (Mg2+) dan ion klorida (Cl-) dalam air. Ion magnesium berfungsi sebagai kofaktor enzim DNA polymerase (Oswald, 2014).
Pada amplifikasi gen, dilakukan tahapan-tahapan tertentu dalam suatu siklus yang dinamakan siklus termal. Siklus termal untuk PCR terdiri dari beberapa tahapan yakni:
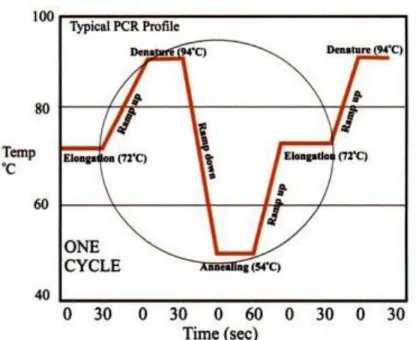
1. Tahap inisiasi
Tahap ini diatur pada temperatur 94-96oC atau 98oC jika polymerase yang sangat termostabil digunakan selama 1-9 menit (Logan, et al., 2009).
2. Tahap denaturasi
Tahap ini merupakan tahap siklus pertama yang terjadi pada temperatur 94-98oC selama 20-30 detik. Pada tahap ini terjadi DNA melting dimana
template DNA diganggu ikatan hidrogennya sehingga menghasilkan DNA single stranded (Logan, et al., 2009).
3. Tahap annealing
Tahap ini terjadi pada temperatur 50-65oC selama 20-40 detik. Pada tahap ini terjadi penempelan primer pada ssDNA template. Tahap annealing harus berada pada temperatur yang cukup rendah untuk memungkinan hibridisasi primer pada strand DNA, namun cukup tinggi agar hibridisasi bisa berjalan secara spesifik, yakni primer hanya menempel pada area template yang komplemen dengan primer. Jika temperatur terlalu rendah, primer tidak dapat menempel dengan sempurna, namun jika terlalu tinggi, primer tidak dapat menempel sama sekali. Biasanya temperatur untuk proses annealing adalah sekitar 3-5oC dibawah titik leleh primer yang digunakan (Logan, et al., 2009). 4. Tahap ekstensi / elongasi
Temperatur tahap ini bergantung pada DNA polymerase yang digunakan, Taq polymerase memiliki aktivitas optimum pada temperatur 75-80oC dan
biasanya temperatur yang digunakan adalah 72oC. Pada tahap ini, DNA polymerase mensintesis strand DNA baru yang komplemen dengan template DNA dengan menambahkan dNTPs dari arah 5’ ke 3’. Waktu ekstensi berganung pada DNA polymerase yang digunakan dan panjang fragmen DNA yang diamplifikasi (Logan, et al., 2009).
5. Elongasi final
Tahap ini biasanya dilakukan pada temperature 70-74oC selama 5-15 menit setelah siklus PCR terakhir untuk memastikan setiap single stranded DNA yang tersisa diekstensi secara utuh (Logan, et al., 2009).
6. Tahap penyimpanan
Temperatur tahap ini adalah 4-15oC untuk waktu yang tak terhingga. Tujuan dari tahap ini adalah penyimpanan jangka pendek dari DNA yang telah direaksikan (Logan, et al., 2009).
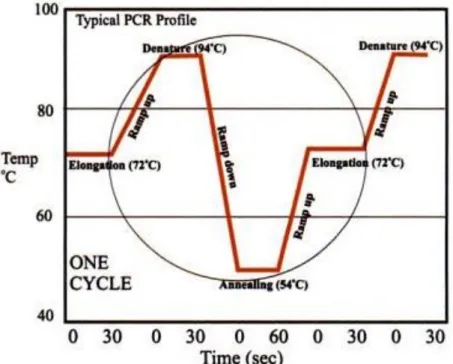
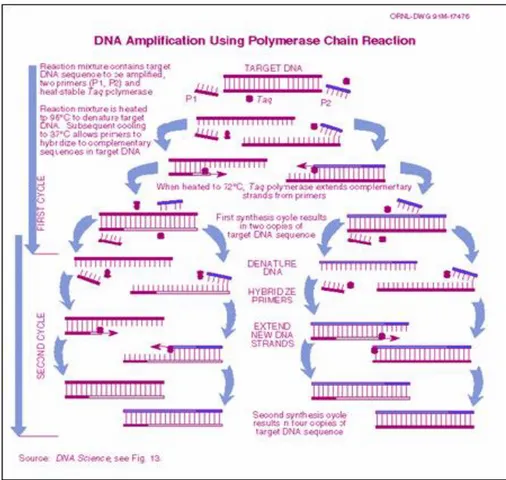
Gambar 4.4 menunjukkan proses molekuler yang terjadi pada siklus termal PCR, sementara gambar 4.5 menunjukkan diagram siklus termal PCR.
Gambar 4.5 Siklus PCR dengan Suhu Optimum (Viljoen, et al., 2005)
Pada hasil percobaan, didapatkan bahwa hasil elektroforesis PCR dari gen phoR berada di sekitar 500bp. Selain itu, terlihat bahwa tidak ada zat lain yang berada di bawah DNA yang dielektroforesis. Ini tidak sesuai dengan literatur yang menunjukkan bahwa panjang protein dari gen phoR adalah 1740 bp (Seki, et al., 1988). Hal ini bisa disebabkan karena DNA tidak seluruhnya terambil ataupun masih tertinggal dalam microtube. Lalu, saat DNA ditaruh pada gel bisa saja masih terdapat DNA yang tertinggal pada tips mikropipet ataupun gel tidak seluruhnya masuk ke dalam well namun ada yang terlepas ke larutan buffer karena kurang berhati-hati dalam prosedur. Selain itu, dapat pula terjadi karena konsentrasi primer yang diberikan tidak terlalu tepat sehingga DNA tidak tertranslasi secara benar.
BAB V KESIMPULAN
5.1 Kesimpulan
Kesimpulan dari percobaan ini adalah:
1. Vektor DNA Plasmid pET-32b(+) berhasil diisolasi. Hasil elektroforesis tidak dapat dibaca karena tidak adanya gene ladder yang dapat dijadikan acuan pembacaan panjang DNA. Seharusnya didapatkan panjang 5899 bp. 2. Pada percobaan dapat dilihat bahwa gen phoR berhasil diamplifikasi. Hasil
elektroforesis menunjukkan band yang didapat adalah 500 bp, sementara jika berhasil seharusnya didapatkan panjang basa 1740 bp.
DAFTAR PUSTAKA
Alberts, B. et al., 2002. Molecular Biology of the Cell. 4th ed. New York: Garland Science.
Bartlett, J. & Stirling, D., 2003. A Short History of the Polymerase Chain Reaction. Methods in Molecular Biology, Volume 226, pp. 3-6.
Chien, A., Edgar, D. & Trella, J., 1976. "Deoxyribonucleic Acid Polymerase from the Extreme Thermophile Thermus aquaticus". Journal of Bacterium, 127(3), pp. 1550-1557.
Dahm, R., 2008. "Discovering DNA: Friedrich Miescher and the Early Years of Nucleic Acid Research". Human Genetics, 122(6), pp. 565-581.
Focosi, D., 2014. NUCLEIC ACID ANALYSIS IN VITRO. http://www.ufrgs.br/imunovet/molecular_immunology/invitrocellfree_nucleic acid.html. Diakses tanggal 16 November 2015.
Kroll, J., Klinter, S., Schneider, C. & Steinbuchel, A., 2010. "Plasmid Addiction Systems: Perspectives and Applications in Biotechnology". Microbial Biotechnology, 3(6), pp. 634-657.
LaVallie, E. et al., 1993. "pET-32a-c(+) Vectors". Bio/Technology, Volume 11, p. 187–193.
Lederberg, J., 1952. "Cell Genetics and Hereditary Symbiosis". Physiol, 32(4), p. 403 – 430.
Lipps, G., 2008. Plasmids: Current Research and Future Trends.. Norfolk: Caister Academic Press.
Logan, J., Edwards, K. & Saunders, N., 2009. Real-Time PCR: Current Technology and Applications.. Norfolk: Caister Academic Press.
Lucotte, G. & Baneyx, F., 1993. Introduction to Molecular Cloning Techniques. New York: Willey-Blackwell.
NIH History, 2010. Polymerase Chain Reaction Test. https://history.nih.gov/nihinownwords/assets/images/archive/polymerasechai n_lg.jpg. Diakses tanggal 16 November 2015].
Oswald, N., 2014. The Basics: how Alkaline Lysis Works. http://bitesizebio.com/180/the-basics-how-alkaline-lysis-works/. Diakses tanggal 16 November 2015.
Robyt, J. F. & White, B. J., 1990. Biochemical Techniques Theory and Practice. Waveland: Waveland Press.
Seki, T., Yoshikawa, H., Takahashi, H. & Saito, H., 1988. "Nucleotide Sequence of the Bacillus subtilis phoR Gene". Journal of Bacteriology, 170(12), pp. 5935-5938.
Stock, S. P., Vandenberg, J., Glazer, I. & Boemare, N., 2009. Insect Pathogens: Molecular Approaches and Techniques. Oxfordshire: MPG Books Group. Tranbichler, M. & Shapiro, L., 2006. Chromosome Organization and Segregation
in Bacterial. Journal of Structural, 156(2), p. 292 – 303.
Viljoen, G. J., Nel, L. H. & Crowther, J. R., 2005. Molecular Diagnostic PCR Handbook. USA: Springer.