27 III. HASIL DAN PEMBAHASAN
3.1 Karakterisasi Morfologi dan Fisiologis Kandidat Probiotik
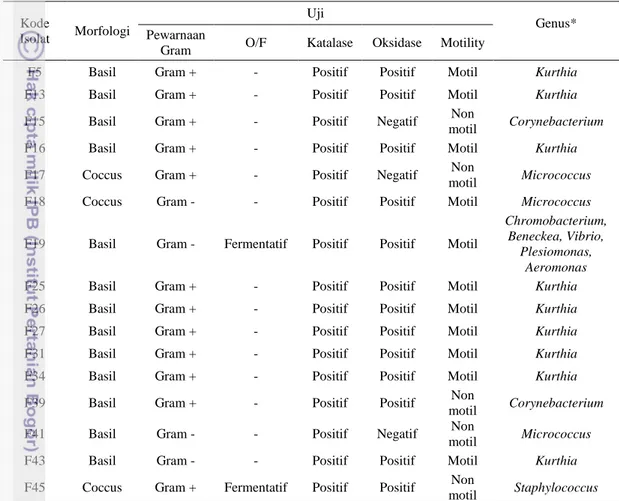
Hasil karakterisasi morfologi dan fisiologis yang dilakukan terhadap 16 jenis bakteri hasil isolasi Ardiani (2011) tertera pada tabel 1.
Tabel 1. Karakterisasi morfologi dan fisiologis kandidat probiotik
Kode
Isolat Morfologi
Uji
Genus* Pewarnaan
Gram O/F Katalase Oksidase Motility
F5 Basil Gram + - Positif Positif Motil Kurthia
F13 Basil Gram + - Positif Positif Motil Kurthia
F15 Basil Gram + - Positif Negatif Non
motil Corynebacterium
F16 Basil Gram + - Positif Positif Motil Kurthia
F17 Coccus Gram + - Positif Negatif Non
motil Micrococcus
F18 Coccus Gram - - Positif Positif Motil Micrococcus
F19 Basil Gram - Fermentatif Positif Positif Motil
Chromobacterium, Beneckea, Vibrio,
Plesiomonas, Aeromonas
F25 Basil Gram + - Positif Positif Motil Kurthia
F26 Basil Gram + - Positif Positif Motil Kurthia
F27 Basil Gram + - Positif Positif Motil Kurthia
F31 Basil Gram + - Positif Positif Motil Kurthia
F34 Basil Gram + - Positif Positif Motil Kurthia
F39 Basil Gram + - Positif Positif Non
motil Corynebacterium
F41 Basil Gram - - Positif Negatif Non
motil Micrococcus
F43 Basil Gram - - Positif Positif Motil Kurthia
F45 Coccus Gram + Fermentatif Positif Positif Non
motil Staphylococcus *Mengacu pada metode identifikasi Cowan (1974)
** Hasil (-) menunjukkan tidak ada reaksi yang terjadi
Berdasarkan hasil karakterisasi morfologi dan fisiologis, diketahui bahwa keenam belas jenis isolat hasil isolasi Ardiani (2011) dari saluran pencernaan udang vaname didominasi oleh bakteri berbentuk basil dan berjenis Gram positif. Sedangkan hasil uji oksidatif/fermentatif menunjukkan hasil fermentatif untuk isolat F19 dan F45 sedangkan isolat lainnya menunjukkan hasil negatif (tidak ada reaksi yang terjadi). Uji katalase menunjukkan hasil positif pada semua jenis isolat. Uji oksidase menunjukkan hasil negatif pada isolat F15, F17, dan F41, sedangkan isolat lainnya menunjukkan hasil yang positif. Isolat F15, F17, F39,
28 F41, dan F45 bersifat tidak motil sedangkan isolat lainnya bersifat motil (mampu bergerak). Berdasarkan identifikasi tingkat genus yang mengacu pada metode Cowan (1974), isolat berhasil diidentifikasi sebagai anggota dari genus Kurthia, Corynebacterium, Micrococcus, Chromobacterium, Beneckea, Vibrio, Plesiomonas, Aeromonas, dan Staphylococcus.
3.2 Uji Aktivitas Amilolitik dan Proteolitik
Aktivitas amilase ditandai dengan terbentuknya zona berwarna kuning terang disekitar isolat sedangkan isolat yang tidak mampu menghidrolisa karbohidrat, area disekitar isolat tetap berwarna gelap setelah penambahan reagen KI. Sedangkan aktivitas protease ditandai dengan terbentuknya zona bening disekitar isolat. Hasil uji amilolitik dan proteolitik disajikan pada gambar 1 di bawah ini.
a. b.
Gambar 1. Hasil aktivitas (a) amilolitik dan (b) proteolitik
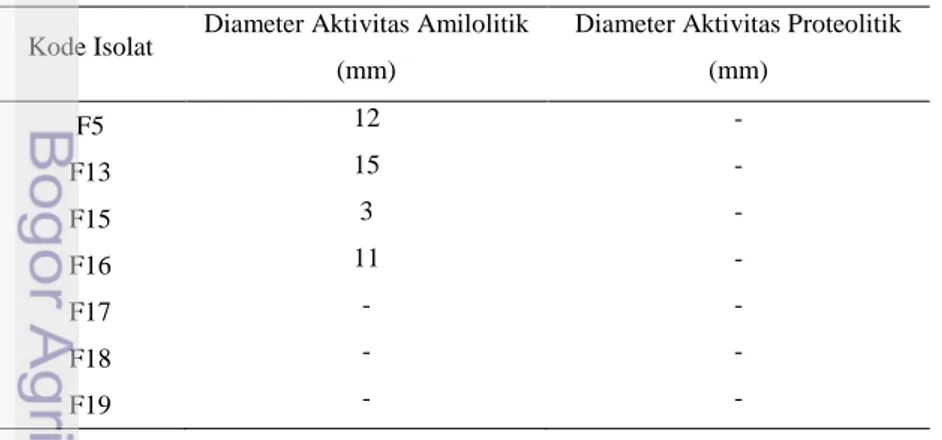
Hasil pengukuran diameter aktivitas amilolitik dan proteolitik dapat dilihat pada tabel 2.
Tabel 2. Diameter aktivitas amilolitik dan proteolitik kandidat probiotik
Kode Isolat Diameter Aktivitas Amilolitik (mm)
Diameter Aktivitas Proteolitik (mm) F5 12 - F13 15 - F15 3 - F16 11 - F17 - - F18 - - F19 - -
29 Tabel 2. Diameter aktivitas amilolitik dan proteolitik kandidat probiotik (lanjutan)
Kode Isolat Diameter Aktivitas Amilolitik (mm)
Diameter Aktivitas Proteolitik (mm) F25 6 - F26 11 - F27 10 - F31 12 - F34 - 15 F39 5 19 F41 - - F43 - - F45 - -
Berdasarkan hasil uji amilolitik isolat diketahui bahwa isolat yang memiliki aktivitas amilolitik terbesar berturut-turut terdapat pada isolat F13 dengan diameter zona amilolitik sebesar 15 mm, serta F5 dan F31 dengan diameter zona amilolitik sebesar 12 mm. Aktivitas proteolitik hanya terdapat pada isolat F34 dan F39. Aktivitas proteolitik terbesar terdapat pada isolat F39 dengan diameter zona sebesar 19 mm diikuti dengan isolat F34 dengan diameter zona sebesar 15 mm. Adanya kemampuan menghidrolisis karbohidrat dan protein menunjukkan isolat-isolat tersebut mampu memanfaatkan sumber energi berupa pati dan protein yang ditambahkan pada media menjadi sumber karbon.
Menurut Price dan Stevens (1996) dalam Yandri et al. (2008), protease merupakan enzim yang berfungsi memecah ikatan peptida untuk menghasilkan asam amino dan peptida sederhana lainnya. Enzim ini dapat diisolasi dari berbagai sumber seperti tanaman, hewan, dan mikroba (fungi dan bakteri). Protease atau disebut juga endopeptidase merupakan salah satu enzim pencernaan yang penting, termasuk di dalamnya tripsin dan kemotripsin yang bertanggung jawab terhadap hampir 60% dari proses pencernaan udang (Lemos et al. 2000).
3.3 Uji Sensitivitas Antibiotik
Uji sensitivitas antibiotik dilakukan pada empat jenis antibiotik yaitu streptomisin, kanamisin, rifampisin, dan penisilin G. Uji ini dilakukan untuk mengetahui tingkat sensitivitas bakteri hasil isolasi Ardiani (2011) terhadap
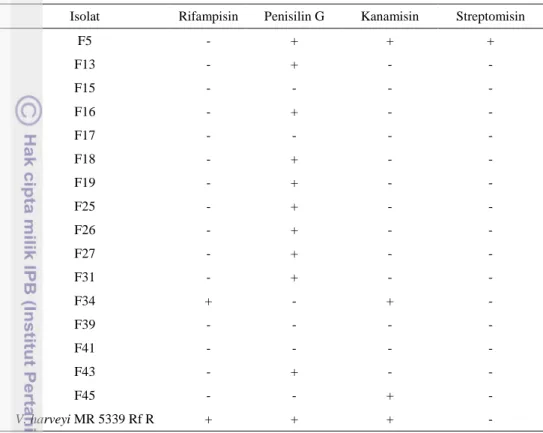
30 antibiotik. Konsentrasi antibiotik yang digunakan sebesar 50 µg/ml dan 100 µg/ml media. Hasil uji sensitivitas disajikan pada tabel di bawah ini.
Tabel 3. Hasil uji sensitivitas antibiotik pada konsentrasi 50 µg/ml
Isolat Rifampisin Penisilin G Kanamisin Streptomisin
F5 - + + + F13 - + - - F15 - - - - F16 - + - - F17 - - - - F18 - + - - F19 - + - - F25 - + - - F26 - + - - F27 - + - - F31 - + - - F34 + - + - F39 - - - - F41 - - - - F43 - + - - F45 - - + - V. harveyi MR 5339 Rf R + + + -
Keterangan: (-):sensitif; (+):resisten
Berdasarkan tabel 3, isolat F34 dan V. harveyi MR 5339 Rf R bersifat resisten terhadap antibiotik rifampisin. Isolat F15, F17, F34, F39, F41, F45 bersifat resisten terhadap penisilin G. Isolat F5, F34, F45 dan V. harveyi MR 5339 Rf R bersifat resisten terhadap kanamisin sedangkan semua isolat (kecuali isolat F5) bersifat sensitif terhadap streptomisin. Sifat resisten terhadap antibiotik pada bakteri dapat disebabkan oleh dua jenis mekanisme yaitu mutasi kromosom dan akuisisi plasmid. Mutasi pada kromosom tidak dapat ditransfer pada bakteri lain tetapi plasmid mampu mentransfer dengan cepat sifat resisten tersebut (Lewin 1992 dalam Schnick 2001). Sifat resistensi pada mikroorganisme juga diakibatkan oleh: (1) kemampuan menghasilkan enzim yang mampu menginaktivasi antibiotik; (2) adanya penambahan substansi kimia pada struktur kimia lain oleh
31 (3) perubahan struktur protein pada dinding sel; (4) berkurangnya permeabilitas dinding sel sehingga antibiotik tidak dapat masuk ke dalam sel (Guilfoile 2007).
Sifat resistensi ini dapat dipindahkan baik secara horizontal maupun vertikal (Madigan et al. 2003 dalam Reboucas et al. 2011). Baticados et al. (1990) dalam Tendencia and de la Pena (2001) melaporkan bahwa patogen penyebab luminous disease seperti Vibrio harveyi dan Vibrio splendidus yang diisolasi dari larva udang resisten terhadap erythromycin, kanamycin, pencillin G, dan streptomycin.
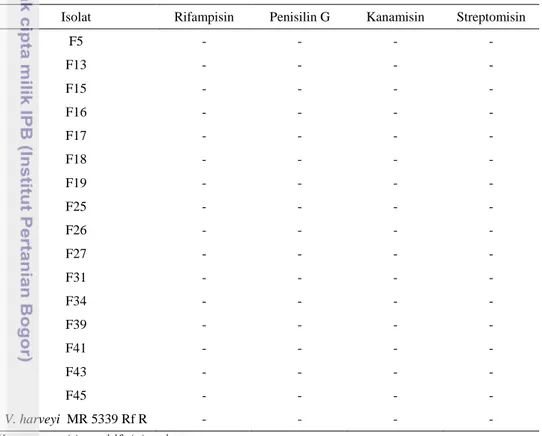
Tabel 4. Hasil uji sensitivitas antibiotik pada konsentrasi 100 µg/ml
Isolat Rifampisin Penisilin G Kanamisin Streptomisin
F5 - - - - F13 - - - - F15 - - - - F16 - - - - F17 - - - - F18 - - - - F19 - - - - F25 - - - - F26 - - - - F27 - - - - F31 - - - - F34 - - - - F39 - - - - F41 - - - - F43 - - - - F45 - - - - V. harveyi MR 5339 Rf R - - - -
Keterangan: (-):sensitif; (+):resisten
Berdasarkan tabel 4, diketahui bahwa pada konsentrasi 100 µg/ml semua jenis isolat sensitif terhadap jenis antibiotik rifampisin, penisilin G, kanamisin, dan streptomisin. Hal tersebut sesuai dengan mekanisme kerja antibiotik menurut Guilfoile (2007) sehingga mampu membunuh bakteri yaitu: (1) merusak kerja ribosom dalam mensintesis protein; (2) menghambat sintesis DNA sebagai sumber informasi genetik; (3) menghambat proses biokimia dalam sel; (4) menghambat sintensis komponen penyusun dinding sel; serta (5) mengganggu fungsi kerja membran sel.
32 3.4 Metode Kultur Bersama
Uji in vitro kultur bersama dilakukan untuk mengetahui kemampuan bakteri kandidat dalam menghambat pertumbuhan bakteri patogen (V. harveyi MR 5339 Rf R). Uji ini dilakukan dengan membandingkan jumlah kepadatan bakteri V. harveyi MR 5339 Rf R yang tumbuh di tabung kontrol dengan tabung kultur bersama.
Berdasarkan hasil analisa statistik (one way ANOVA, P=0,05) diketahui terdapat delapan jenis isolat yang menunjukkan kemampuan menekan pertumbuhan bakteri patogen secara berbeda nyata jika dibandingkan dengan tabung kontrol. Isolat-isolat tersebut yaitu F5, F17, F18, F19, F25, F31, F39, dan F43. Isolat F5, F17, F19 dan F43 mampu menekan populasi patogen sebesar 102 -103 CFU/ml sedangkan kepadatan bakteri patogen pada tabung kontrol mencapai 109 CFU/ml. Isolat F18, F25, F31, dan F39 juga mampu menekan populasi patogen namun dengan kemampuan yang berbeda (Gambar 2).
33 Gambar 2. Penghambatan V. harveyi MR 5339 Rf R oleh kandidat probiotik pada uji in vitro metode kultur bersama
7,08 ± 0,47 9,29 ± 2,17 9,23 ± 2,25 9,27 ± 2,37 7,14 ± 0,45 8,45 ± 1,30 6,85 ± 0,10 8,77 ± 1,89 9,46 ± 1,45 9,35 ± 2,30 8,36 ± 1,17 9,39 ± 1,93 8,08 ± 1,42 10,15 ± 3,30 7,84 ± 1,08 9,17 ± 2,23 9,30 ± 2,17 0,0 0,5 1,0 1,5 2,0 2,5 3,0 3,5 4,0 4,5 5,0 5,5 6,0 6,5 7,0 7,5 8,0 8,5 9,0 9,5 10,0 10,5 11,0 11,5 12,0 12,5 13,0 13,5 14,0 F 5 F 13 F 15 F 16 F 17 F 18 F 19 F 25 F 26 F 27 F 31 F 34 F 39 F 41 F 43 F 45 Vibrio harveyi K epa da ta n ba k ter i (lo g CF U/m l) Vibrio harveyi MR 5339 Rf R ab a bc cd bc cd a a a d cd a e cd a a cd
34 Tidak semua jenis bakteri mampu memproduksi inhibitory metabolites (senyawa penghambat) pada media agar karena terdapat kemungkinan produksi inhibitory metabolites akan lebih banyak jika kultur dilakukan pada media broth. Adanya perbedaan komposisi medium juga mempengaruhi produksi senyawa penghambat (Verschuere et al. 2000). Hal tersebut membuktikan bahwa penghambatan pertumbuhan populasi tidak selalu dapat dilihat melalui metode zona bening (Kirby Bauer Method).
Lactid Acid Bacteria (LAB) yang diisolasi dari saluran pencernaan mampu memproduksi komponen bakteriosin yang mampu menghambat pertumbuhan mikroorganisme lainnya dan secara tidak langsung akan mempengaruhi kompetisi dalam memperoleh energi di lingkungannya. LAB yang ditemukan dalam saluran pencernaan organisme akuatik seperti ikan mampu meningkatkan status kesehatan organisme tersebut (Verschuere et al. 2000). Sasanti (2005) melaporkan bahwa dari 110 isolat hasil isolasi dari terumbu karang, hanya 10 isolat saja yang potensial sebagai kandidat probiotik. Lima diantaranya terbukti mampu menghambat pertumbuhan Vibrio harveyi MR 5339 Rf R walaupun tidak menunjukkan adanya zona hambat pada uji in vitro.
Untuk mengetahui tingkat kepadatan bakteri yang dikultur sebelum masuk ketahapan kultur bersama, dilakukan pengukuran kepadatan bakteri dengan menggunakan spektrofotometer. Kepadatan bakteri diketahui dengan mengukur nilai optical density (OD) dari populasi sel yang dikultur selama 24 jam. Pengukuran OD dilakukan pada panjang gelombang 600 nm (lampiran 1).
3.5 Uji Patogenisitas Bakteri Kandidat Probiotik
Uji ini dilakukan untuk melihat tingkat kelangsungan hidup pascalarva (PL17) udang vaname setelah diberi kandidat probiotik. Uji patogenisitas dilakukan terhadap isolat-isolat yang menunjukkan daya hambat terhadap pertumbuhan Vibrio harveyi MR 5339 Rf R pada tahap kultur bersama. Uji patogenisitas dilakukan terhadap empat bakteri kandidat terbaik berdasarkan hasil kultur bersama (Gambar 2) yaitu isolat F5, F17, F19, dan F43. Kontrol positif yang digunakan adalah patogen V. harveyi MR 5339 Rf R. Uji patogenisitas dilakukan dengan enam perlakuan dimana masing-masing perlakuan dan dua
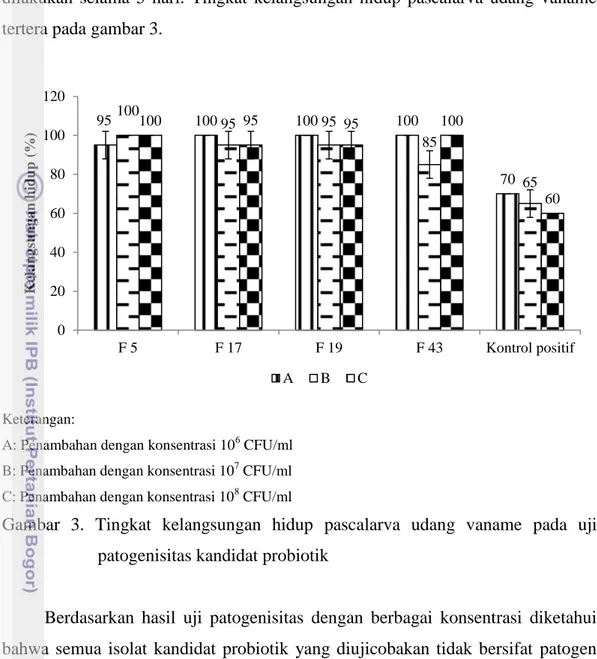
35 ulangan. Pengamatan tingkat kelangsungan hidup pascalarva udang vaname dilakukan selama 5 hari. Tingkat kelangsungan hidup pascalarva udang vaname tertera pada gambar 3.
Keterangan:
A: Penambahan dengan konsentrasi 106 CFU/ml
B: Penambahan dengan konsentrasi 107 CFU/ml
C: Penambahan dengan konsentrasi 108 CFU/ml
Gambar 3. Tingkat kelangsungan hidup pascalarva udang vaname pada uji patogenisitas kandidat probiotik
Berdasarkan hasil uji patogenisitas dengan berbagai konsentrasi diketahui bahwa semua isolat kandidat probiotik yang diujicobakan tidak bersifat patogen terhadap pascalarva udang vaname (Gambar 3). Hal ini dibuktikan dengan tingkat kelangsungan hidup pascalarva udang vaname yang diberi kandidat probiotik yang tinggi yaitu berkisar antara 85-100%, sedangkan kontrol positif (penambahan V. harveyi) sebesar 60-70%.
3.6 Pembuatan Mutan Bakteri Kandidat Probiotik
Empat isolat terpilih hasil seleksi in vitro yang terbukti mampu menekan pertumbuhan bakteri V. harveyi MR 5339 Rf R dan tidak bersifat patogen pada pascalarva udang vaname dibuat mutan terhadap antibiotik sebelum digunakan
95 100 100 100 70 100 95 95 85 65 100 95 95 100 60 0 20 40 60 80 100 120 F 5 F 17 F 19 F 43 Kontrol positif K ela ng sun g a n hid up ( %) A B C
36 pada uji in vivo. Empat isolat yaitu F5, F17, F19, dan F43 dibuat mutan secara spontan dengan cara menyebar biakan cair isolat pada media SWC agar yang mengandung streptomisin 50 µg/ml. Isolat yang tumbuh (resisten) pada media SWC yang mengandung streptomisin diambil sebagai inokulan untuk uji in vivo.
Bentuk dan ciri fisik isolat mutan tidak berbeda dengan isolat awalnya. Isolat F5 memiliki ciri berbentuk bulat, tepian rata, elevasi cembung, dan koloni berwarna putih susu (Gambar 4a). Hasil identifikasi genus merujuk pada metode identifikasi Cowan (1974) menunjukkan isolat F5 tergolong anggota genus Kurthia. Isolat F17 memiliki ciri koloni berbentuk bulat, tepian rata, elevasi cembung, berwarna putih, dan teridentifikasi sebagai anggota genus Micrococcus (Gambar 4b). Isolat F19 memiliki ciri koloni berbentuk bulat, berwarna putih susu, tepian rata, berelevasi cembung, dan teridentifikasi sebagai anggota genus Chromobacterium, Beneckea, Vibrio, Plesiomonas, dan Aeromonas. Isolat F43 memiliki ciri koloni berwarna putih, berbentuk bulat kecil, tepian rata, berelevasi cembung dan teridentifikasi sebagai anggota genus Kurthia.
a. b.
c. d.
Gambar 4. Penampilan mutasi spontan isolat (a) F5; (b) F17; (c) F19; dan (d) F43 pada media SWC agar
37 3.7 Uji in vivo Kandidat Probiotik
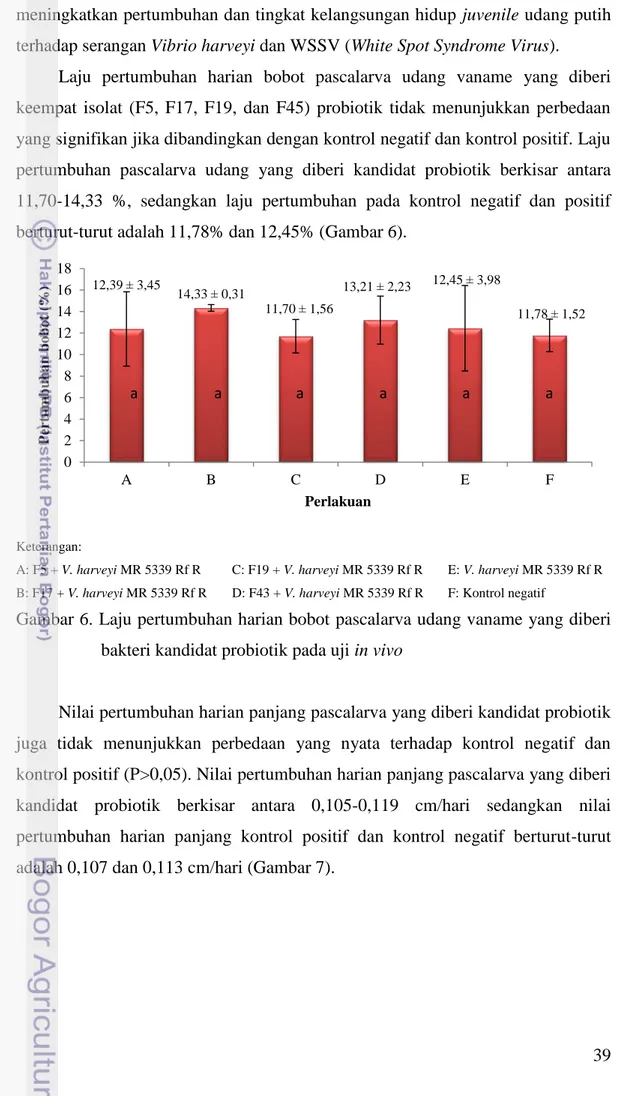
Empat isolat hasil uji in vitro yaitu isolat F5, F17, F19, dan F43 diuji kemampuannya dalam menghambat pertumbuhan V. harveyi MR 5339 Rf R pada pascalarva (PL 26) udang vaname. Pengamatan yang dilakukan pada uji in vivo meliputi survival rate (tingkat kelangsungan hidup), laju pertumbuhan harian, total populasi V. harveyi MR 5339 Rf R, dan total populasi probiotik.
Berdasarkan hasil pengamatan diketahui bahwa tingkat kelangsungan hidup pascalarva udang vaname yang diberi keempat kandidat probiotik memiliki perbedaan yang signifikan (P<0,05) jika dibandingkan dengan kontrol positif (67,5%). Namun jika dibandingkan dengan kontrol negatif tidak menunjukkan perbedaan yang signifikan. Tingkat kelangsungan hidup pascalarva udang vaname yang diberi kandidat probiotik berkisar antara 78,75-92,50% sedangkan kontrol positif sebesar 67,5% (Gambar 5).
Keterangan:
A: F5 + V. harveyi MR 5339 Rf R C: F19 + V. harveyi MR 5339 Rf R E: V. harveyi MR 5339 Rf R B: F17 + V. harveyi MR 5339 Rf R D: F43 + V. harveyi MR 5339 Rf R F: Kontrol negatif
Gambar 5. Tingkat kelangsungan hidup pascalarva udang vaname yang diberi bakteri kandidat probiotik pada uji in vivo
Pemberian bakteri probiotik mampu meningkatkan kelangsungan hidup larva udang vaname secara signifikan dapat disebabkan oleh adanya kompetisi dalam mendapatkan sumber energi. Menurut Smith and Davey (1993) dalam Verschuere et al. (2000) melaporkan bahwa Pseudomonas fluorescent F19 mampu menghambat pertumbuhan Aeromonas salmonicida pada media
92,50 ± 6,45 86,25 ± 4,79 78,75 ± 8,54 82,50 ± 6,45 67,50 ± 6,45 83,75 ± 8,54 0 20 40 60 80 100 120 A B C D E F Sur v iv a l ra te ( %) Perlakuan a a a a b a
38 pemeliharaan Atlantic salmon. Mekanisme penghambatannya diketahui melalui kompetisi dalam memperoleh zat besi. Adanya produksi senyawa penghambat juga turut memberikan pengaruh dalam proses penghambatan pertumbuhan patogen oleh bakteri probiotik. Senyawa penghambat yang dimaksud dapat berupa antibiotik, senyawa yang bersifat bakteriostatik maupun bakteriosida, lysozyme, ataupun protease. Senyawa-senyawa tersebut diketahui mampu menghambat pertumbuhan mikroorganisme lain. Adanya kompetisi dalam memperoleh energi dan produksi senyawa penghambat inilah yang diduga mampu menekan pertumbuhan bakteri patogen.
Rengpipat et al. (2000) juga melaporkan bahwa penambahan Bacillus S11 mampu meningkatkan kelangsungan hidup pada larva udang windu (Penaeus monodon). Hasil penelitian Vaseeharan and Ramasamy (2003) menunjukkan bahwa tingkat kematian udang windu yang tidak diberi probotik Bacillus subtilis BT23 mencapai 50% pada hari ke-9 dan mencapai 100% pada hari ke-17.
Probiotik umumnya diaplikasikan pada media atau kolam pemeliharaan sebagai bentuk tindakan preventif terhadap serangan penyakit serta aktivator regenerasi nutrien (Yasuda dan Taga 1980 dalam Verschuere et al. 2000). Beberapa kelompok bakteri diketahui sebagai agen pengontrol biologis dalam akuakultur diantaranya kelompok Lactid Acid Bacteria (genus Lactobacillus dan Carnobacterium), anggota genus Vibrio (Vibrio alginolyticus), anggota genus Bacillus, serta anggota genus Pseudomonas. Menurut Verschuere et al. (2000), mekanisme kerja probiotik diantaranya adalah adanya kemampuan memproduksi senyawa inhibitor, kompetisi dalam memperoleh energi, kompetisi tempat pelekatan, meningkatkan respon imun, dan memperbaiki kualitas air.
Salah satu mekanisme kerja probiotik adalah mampu meningkatkan respon imun non spesifik inang. Rengpipat et al. (2000) melaporkan bahwa Bacillus sp. strain S11 mampu meningkatkan aktivitas fagositik leukosit pada udang windu (Penaeus monodon). Tingkat kelangsungan hidup udang windu yang diberi probiotik Bacillus sp. strain S11 memiliki perbedaan yang signifikan (54,3%) jika dibandingkan dengan yang tidak diberi probiotik (35,5%). Hal tersebut juga didukung oleh penelitian Balaczar (2003) dalam Ghosh et al. (2008) yang menyebutkan bahwa penggunaan campuran Bacillus sp. dan Vibrio sp. mampu
39 meningkatkan pertumbuhan dan tingkat kelangsungan hidup juvenile udang putih terhadap serangan Vibrio harveyi dan WSSV (White Spot Syndrome Virus).
Laju pertumbuhan harian bobot pascalarva udang vaname yang diberi keempat isolat (F5, F17, F19, dan F45) probiotik tidak menunjukkan perbedaan yang signifikan jika dibandingkan dengan kontrol negatif dan kontrol positif. Laju pertumbuhan pascalarva udang yang diberi kandidat probiotik berkisar antara 11,70-14,33 %, sedangkan laju pertumbuhan pada kontrol negatif dan positif berturut-turut adalah 11,78% dan 12,45% (Gambar 6).
Keterangan:
A: F5 + V. harveyi MR 5339 Rf R C: F19 + V. harveyi MR 5339 Rf R E: V. harveyi MR 5339 Rf R B: F17 + V. harveyi MR 5339 Rf R D: F43 + V. harveyi MR 5339 Rf R F: Kontrol negatif
Gambar 6. Laju pertumbuhan harian bobot pascalarva udang vaname yang diberi bakteri kandidat probiotik pada uji in vivo
Nilai pertumbuhan harian panjang pascalarva yang diberi kandidat probiotik juga tidak menunjukkan perbedaan yang nyata terhadap kontrol negatif dan kontrol positif (P>0,05). Nilai pertumbuhan harian panjang pascalarva yang diberi kandidat probiotik berkisar antara 0,105-0,119 cm/hari sedangkan nilai pertumbuhan harian panjang kontrol positif dan kontrol negatif berturut-turut adalah 0,107 dan 0,113 cm/hari (Gambar 7).
12,39 ± 3,45 14,33 ± 0,31 11,70 ± 1,56 13,21 ± 2,23 12,45 ± 3,98 11,78 ± 1,52 0 2 4 6 8 10 12 14 16 18 A B C D E F P er tum bu ha n bo bo t (%) Perlakuan a a a a a a
40
Keterangan:
A: F5 + V. harveyi MR 5339 Rf R C: F19 + V. harveyi MR 5339 Rf R E: V. harveyi MR 5339 Rf R B: F17 + V. harveyi MR 5339 Rf R D: F43 + V. harveyi MR 5339 Rf R F: Kontrol negatif
Gambar 7. Nilai pertumbuhan harian panjang pascalarva udang vaname yang diberi bakteri kandidat probiotik pada uji in vivo
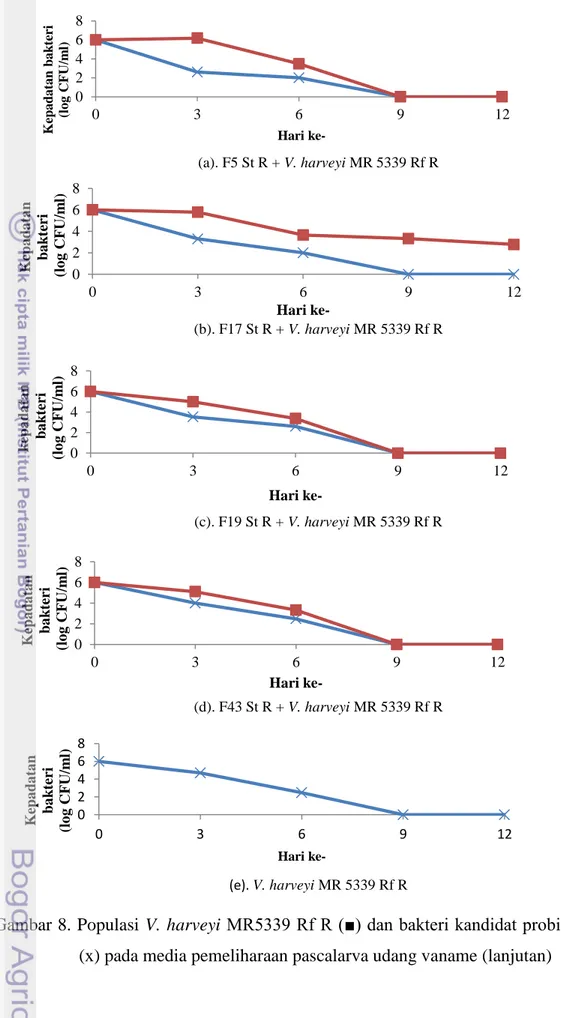
Populasi patogen V. harveyi MR 5339 Rf R dan bakteri kandidat probiotik (F5, F17, F19, dan F43) dimonitor keberadaannya setelah ditandai terlebih dahulu melalui pembuatan mutan spontan antibiotik. Populasi V. harveyi MR 5339 Rf R pada keempat isolat yang diberikan mampu menekan jumlah bakteri V. harveyi MR 5339 Rf R. Hal ini dapat dilihat pada hari ke-3 dimana keempat isolat tersebut mampu menekan patogen sebesar 102-104 CFU/ml (Gambar 8) sedangkan
kepadatan bakteri V. harveyi MR 5339 Rf R pada kontrol positif sebesar 105 CFU/ml. Gambar 8 menunjukkan kepadatan V. harveyi MR 5339 Rf R pada
semua perlakuan mengalami penurunan hingga mencapai kepadatan 102 CFU/ml pada hari ke-6 dan sudah tidak terdeteksi lagi pada hari ke-9. Hal ini menunjukkan adanya kemampuan menekan pertumbuhan bakteri patogen pada media pemeliharaan pascalarva udang vaname.
0,105±0,014 0,119±0,013 0,110±0,016 0,105±0,014 0,107±0,021 0,113±0,015 0,050 0,070 0,090 0,110 0,130 A B C D E F L a ju per tum bu ha n pa nja ng (cm /ha ri) a a a a a a
41 (a). F5 St R + V. harveyi MR 5339 Rf R (b). F17 St R + V. harveyi MR 5339 Rf R (c). F19 St R + V. harveyi MR 5339 Rf R (d). F43 St R + V. harveyi MR 5339 Rf R (e). V. harveyi MR 5339 Rf R
Gambar 8. Populasi V. harveyi MR5339 Rf R (■) dan bakteri kandidat probiotik (x) pada media pemeliharaan pascalarva udang vaname (lanjutan)
0 2 4 6 8 0 3 6 9 12 K ep a d a tan b a k teri (l o g CF U/ m l) Hari ke- 0 2 4 6 8 0 3 6 9 12 K epa da ta n ba k ter i ( lo g CF U/m l) Hari ke- 0 2 4 6 8 0 3 6 9 12 k epa da ta n ba k ter i (lo g CF U/m l) Hari ke- 0 2 4 6 8 0 3 6 9 12 K epa da ta n ba k ter i ( lo g CFU/m l) Hari ke- 0 2 4 6 8 0 3 6 9 12 K epa da ta n ba k ter i (lo g CF U/m l) Hari ke-
42 Adanya peningkatan kelangsungan hidup pada pascalarva udang disebabkan oleh adanya penekanan terhadap pertumbuhan V. harveyi MR 5339 Rf R. Penekanan pertumbuhan populasi patogen ini ditunjukkan dengan berkurangnya jumlah bakteri patogen pada media pemeliharaan yang diberi kandidat probiotik jika dibandingkan dengan kontrol positif (Gambar 9). Populasi V. harveyi MR 5339 Rf R pada kontrol positif mencapai 105 CFU/ml pada hari ke-3, ini lebih tinggi daripada populasi V. harveyi MR 5339 Rf R pada perlakuan penambahan probiotik yang mencapai kepadatan 102-104 CFU/ml. Penekanan populasi patogen terbaik terdapat pada perlakuan penambahan isolat F5 yakni mampu mengeliminasi patogen sebesar 103 CFU/ml pada hari ke-3. Kemampuan penekanan V. harveyi MR 5339 Rf R oleh isolat F5 diikuti pula oleh isolat F19, F17, dan F43. Kemampuan menghambat ini diduga menyebabkan tingginya tingkat kelangsungan hidup pascalarva udang vaname pada perlakuan penambahan isolat F5 (Gambar 9).
Keterangan:
A: F5 + V. harveyi MR 5339 Rf R C: F19 + V. harveyi MR 5339 Rf R E: V. harveyi MR 5339 Rf R B: F17 + V. harveyi MR 5339 Rf R D: F43 + V. harveyi MR 5339 Rf R F: Kontrol negatif
Gambar 9. Tren populasi Vibrio harveyi MR 5339 Rf R pada berbagai perlakuan
Penambahan bakteri probiotik diawal pemeliharaan saja menyebabkan populasi kandidat probiotik menurun hingga hari ke-12 pengamatan (Gambar 10). Isolat F5, F19, dan F43 mengalami penurunan populasi hingga tidak terdeteksi sama sekali pada hari ke-9. Sedangkan isolat F17 mampu bertahan pada media pemeliharaan hingga hari ke-12 pengamatan. Isolat F5 dan F19 memiliki populasi
0 1 2 3 4 5 6 7 0 3 6 9 12 K epada ta n b a k teri (lo g CF U/m l) Hari ke- A B C D E F
43 yang lebih tinggi dari pada isolat F17 dan F43 pada hari ke-3 pengamatan. Lebih tingginya kepadatan isolat F5 pada hari ke-3 berkolerasi dengan kemampuannya menghambat pertumbuhan Vibrio harveyi MR 5339 Rf R (Gambar 9). Hubungan ini juga terdapat pada perlakuan penambahan isolat F17 dan F43.
Keterangan:
A: F5 + V. harveyi MR 5339 Rf R C: F19 + V. harveyi MR 5339 Rf R E: V. harveyi MR 5339 Rf R B: F17 + V. harveyi MR 5339 Rf R D: F43 + V. harveyi MR 5339 Rf R F: Kontrol negatif
Gambar 10. Tren populasi kandidat probiotik St R pada berbagai perlakuan
Vibriosis pada udang merupakan penyakit bakterial yang disebabkan oleh V. harveyi. Penyakit ini bersifat oportunis dimana akan menyerang pada saat sistem imun inang lemah (Defoirdt et al. 2008). Serangan vibriosis akan menyebabkan larva menjadi abnormal diantaranya saluran pencernaan menjadi kosong, hepatopankreas menyusut dan munculnya bintik-bintik melanisasi pada permukaan tubuh, serta kolonisasi bakteri pada permukaan kutikula larva (Brock and Main 1994). Patogen ini juga dilaporkan sebagai agen penyebab penyakit pada kuda laut (Hipocampus sp.), skin ucleration (bisul) pada teripang (Holothuria scabra) di Madagaskar, dan luminous disease pada larva phyllosoma (Jasus verreauxi). Gejala pada teripang yang terserang patogen ini berupa bintik putih dan kematian akan terjadi selama tiga hari (Austin and Zhang 2006).
Tingkat patogenisitas Vibrio harveyi berhubungan dengan mekanisme quorum sensing. Quorum sensing merupakan mekanisme komunikasi antar sel melalui produksi molekul-molekul tertentu (Defoirdt et al. 2007) atau mekanisme regulasi gen dimana bakteri mengkoordinasikan ekspresi gen tertentu dalam
0 1 2 3 4 5 6 7 0 3 6 9 12 K epa da ta n ba k ter i ( lo g CF U/m l) Hari ke- A B C D E F
44 menanggapi ada atau tidak adanya molekul sinyal tertentu (Defoirdt et al. 2008). Vibrio harveyi diketahui memiliki multi mekanisme quorum sensing. Mekanisme quorum sensing Vibrio harveyi diatur oleh harveyi autoinducer 1 (HAI-1), autoinducer 2 (AI-2), dan cholerae autoinducer 1 (CAI-1). Quorum sensing dilaporkan bertanggung jawab dalam mengatur sifat virulensi dari patogen V. harveyi. Menurut Natrah et al. (2011) sifat virulensi V. harveyi disebabkan oleh kemampuannya memproduksi ekstraseluler produk. Tingkat virulensi V. harveyi terkait dengan produksi ekstraselular produk berupa protease, sitotoksin, dan haemolisin yang memungkinkan bakteri mampu bertahan hidup dan bereplikasi pada inangnya (Montero and Austin 1999).