P
lanktonic microalgae (phytoplankton) live in diverse and highly variable environments. Their photosynthetic apparatus is subject to significant stresses because of rapid changes or imbalances in irradiance and nutrient supply imposed by the physics and chemistry of natural water bodies. The ways in which the pho-tosynthetic apparatus adjusts to these temporally variable and complex environments are of interest for both practical and fun-damental reasons, because phytoplankton photosynthesis is respon-sible for ~50% of global productivity. Modern techniques for assessing productivity on global scales rely on remotely sensing plankton’s optical properties, using the signature of chlorophyll as an index of abundance (Fig. 1). The emphasis on chlorophyll is inevitable given chlorophyll’s distinctive optical characteristics, which enables plant material to be distinguished from other sus-pended matter. The relationship between chlorophyll and organic carbon (the desired currency for productivity models) is highly plastic, varying with growth, irradiance, nutrient availability and temperature. However, the variability is ordered, not random, and knowledge about the effects of environmental variables on the efficiency of light absorption and its conversion to biomass during photosynthesis allows estimates of chlorophyll abundance to be translated into carbon equivalents.Understanding the effect of varying environmental conditions on photosynthetic rates can be considered in terms of the regulation of the amounts and the specific activities of components of the photo-synthetic apparatus. Regulation can be accomplished by variations in the relative abundance of the constituents [e.g. chlorophyll and ribulose-1,5-bisphosphate carboxylase/oxygenase (Rubisco)] or, on a shorter timescale, by varying the efficiency of their coupling and activity (e.g. the activation state of Rubisco). The distinction is blurred at some levels but serves to distinguish between those means of photosynthetic regulation that depend on synthesis or turnover (usually of chlorophyll) in photoacclimation, and those that depend on more rapid changes in activation states or efficiencies, independent of turnover. Work in this area is complicated by the fact that phytoplankton include representatives of four kingdoms1 and are highly variable in their molecular, structural and optical properties. Whereas physiological studies have focused largely on taxa that are easy to maintain in culture, particularly chlorophytes and diatoms. It has been challenging to bring qualitative information on photosynthetic mechanisms together with quantitative kinetic information on rates of response to understand cellular photosyn-thesis in the natural environment. Here we review recent developments in our understanding of how phytoplankton
photo-synthesis adjusts to naturally variable environments.
The principal environmental factors that affect phytoplank-ton photosynthesis are light, nutrient availability and tem-perature. In nature, these factors operate independently on time-scales that match photosynthetic physiology, presenting a com-plex and unpredictable envi-ronment to the cells. Phyto-plankton respond to such stochastic environments using a variety of physiological processes that affect light harvesting efficiency and photosynthetic capacity. Two key issues are at the forefront of ecophysiological research: Fig. 1.Composite false-color image of mean annual ocean chlorophyll concentrations, as detected by satellite
remote-sensing. Chlorophyll is used as an index of biomass in seawater because, unlike other organic compounds, it is unique to phytoplankton and common to all autotrophs. Note the higher concentrations near the land
The effect of water motion on
short-term rates of photosynthesis
by marine phytoplankton
Hugh L. MacIntyre, Todd M. Kana and Richard J. Geider
• What are the kinetic constraints on processes that modulate photosynthesis in rapidly fluctuating environments?
• What are the mechanisms by which cells integrate multiple en-vironmental factors in regulating their physiological responses? These questions are related: the photosynthetic mechanisms that modulate energy and material flow through the cell are common to phytoplankton in spite of their diversity. Significant conceptual advances have been made recently that tie whole-cell physiology with photosynthetic regulatory processes operating at the bio-chemical and biophysical levels.
Environmental control of photosynthetic processes
By definition, phytoplankton are incapable of sustained direc-tional movement and therefore are subject to the environmental conditions in their parent body of water. Photosynthesis is respon-sive to changes in nutrient availability on short timescales2
but in marine systems the changes are more likely to occur over rela-tively long timescales (days to seasons). Although nutrient avail-ability is critical for determining population dynamics3
and the ultimate determinant of the geographic abundance of phytoplankton (Fig. 1), nutrient availability is less likely to drive short-term changes in photosynthetic rates. Because of water’s capacity for high levels of latent heat, temperature is more likely to vary on daily to seasonal scales, and in a manner predictable from the balance of radiative transfer and evaporative cooling. By contrast, phyto-plankton are subject to relatively rapid changes in both the inten-sity and the spectral quality of light as they move vertically in a water column (Box 1). In addition to the changes imposed by mixing, there are variations in the light field with timescales of hours, minutes and milliseconds4
as a result of changes in solar elevation, cloud cover and subsurface focusing by waves. Photo-synthesis and growth rates appear to be insensitive to variability in the millisecond domain5
, therefore we will focus on variations that occur on a timescale of minutes and hours.
Photoacclimation of pigment content
For a given temperature and nutrient status, photoacclimation in phytoplankton can be described in terms of regulation of the cell concentrations of the catalysts that determine light-limited and light-saturated photosynthetic rates. The rate of light absorption, which co-varies with a cell’s chlorophyll concentration, is often the primary determinant of light-limited photosynthesis, whereas the maximum rate of carbon dioxide fixation, which co-varies with Rubisco concentration, is the primary determinant of light-saturated photosynthesis. The
cell chlorophyll content is usu-ally higher in cells that have grown under low light6
. Vari-ations in other constituents of the photosynthetic apparatus appear to be taxon-specific: of two studies of cells grown under nutrient-replete condi-tions, the concentration of Rubisco decreased at low growth irradiance in a diatom7 whereas it remained unchanged in a chlorophyte8
. Synthesis of pigment–protein complexes appears to be transcriptionally regulated by a mechanism that is under the control of the redox state of the plasto-quinone pool9
. Under low-light
conditions, plastoquinone is largely oxidized, and transcription of the mRNAs that code for the proteins that bind chlorophyll in the antenna occurs at maximal rates. As irradiance increases, photo-synthesis control passes from light harvesting to the maximum capacity for carbon dioxide fixation, the plastoquinone becomes increasingly reduced, and transcription of the mRNAs for pig-ment–protein complexes declines. The end results of this mode of regulation can be mimicked by ‘energy balance’ models de-veloped recently10,11
. These are based on the concept of a physio-logical light sensor that regulates the pigment quota to maintain a balance between the harvesting of excitation energy by means of light absorption and photochemistry on the one hand and the ener-getic demands of growth on the other. Conceptually, this is com-parable to the redox regulation of the antenna protein LHCII (Refs 9,12). These models can account for steady-state responses and for the differences in the rates of acclimation that are observed following shifts from low-to-high versus high-to-low irradiance, without the potential errors imposed by specifying a mathematical function to describe the kinetics of photoacclimation13
.
Photosynthetic Induction
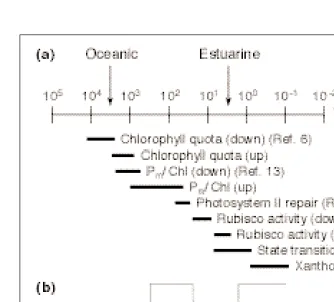
The time-course of light intensity changes in estuarine waters is much faster than in coastal or open-ocean waters because both the rate of light attenuation and the rate of mixing are much higher (Box 1). Two mechanisms, the activation and deactivation of Rubisco and state transitions, have time constants that are compa-rable to the timescale of mixing in estuarine waters and probably dominate short-term rates of photosynthesis (Fig. 2).
Photosynthetic regulation operating by means of the catalytic activity of Rubisco can be controlled by variations in the enzyme concentration or, in the short-term, by its activation state. Although the enzyme cellular concentration might not be regu-lated during photoacclimation under nutrient-replete conditions, depending on the taxon8
, it is regulated in response to chronic phosphorus or nitrogen limitation14
. Because there is a correlation between the maximum quantum efficiency (fm) and the ratio of Rubisco to the PSII reaction center protein D1, changes in the pool size of Rubisco might play a role in regulating acclimated photo-synthetic rates during nutrient-limited growth.
A potential role for activation and deactivation of Rubisco in the short-term limitation of photosynthetic rates hinges on the assumption that at light saturation, the rate of photosynthe-sis depends on the enzyme’s activity. Broad conclusions are complicated by the diversity in microalgal Rubisco structure and
Box 1. Variations in the submarine light field
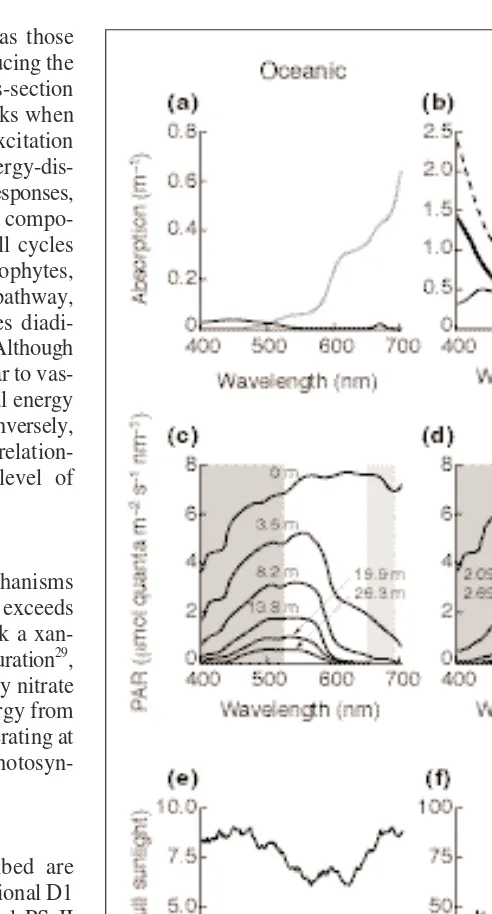
The light attenuation within any body of water follows (approximately) the Lambert–Beer law, decreasing expo-nentially with depth. The rate and spectral dependence of attenuation depends on the abundance of phytoplankton, detritus (which includes organic material of biogenic origin and suspended sediment) and chromophoric dissolved organic matter (CDOM, mainly humic and fulvic acids of terrestrial origin).
In estuarine waters, terrestrial runoff that is high in humics and plant nutrients, and close coupling between the benthos and water column results in high absorption by CDOM, phytoplankton and detritus in blue light. The dom-inant penetrating wavelengths are therefore red and the intermediate green (Fig. 3). Attenuation is rapid: in an extreme case, the euphotic zone (the depth to which 1% of sunlight penetrates) is ,0.5 m.
In oceanic waters, the abundance of CDOM, detritus and phytoplankton is low and the dominant attenuator is water itself, which is highly absorbent in red light. The dominant penetrating wavelengths are therefore blue light (Fig. 3). The euphotic zone might be up to 140 m deep. Coastal waters are intermediate with respect to the mag-nitude and spectral dependence of attenuation.
catalytic characteristics15,16
. However, activity changes in those few microalgae that have been studied are consistent with models of regulation by means of carbamylation–decarbamylation and, in addition, possibly, by a tight-binding inhibitor, such as carboxy-arabinitol-1-phosphate17
. The kinetics of activation and deacti-vation could affect photosynthetic rates in response to rapid increases in light intensity during mixing. Because activation is much faster than deactivation, the effect is less pronounced during rapid mixing because the interval between decreases in illumi-nation as a cell is mixed away from the well-lit surface and increases
in illumination as it is mixed back up are insufficient to allow much deactivation18
. There appears to be little role for Rubisco oxygenase activity in phytoplankton. Although the abundance of free CO2in sea-water (~10–15 mM) is well below the half-saturation concentration of Rubisco (30–170 mM)15,16
, carbon concentrating mechanisms that involve either active transport of HCO3
2
or coupled dehydration of HCO3
2
by a cell-surface carbonic anhy-drase and CO2 transport, have been docu-mented for cyanobacteria, chlorophytes19 , diatoms20
and dinoflagellates21,22
. In addi-tion, microalgal Rubiscos have high CO2 specificities compared with those from ter-restrial plants. This might be because of differences in the structure of the loop-6 region15
or in the length of the large subunit C-terminus16
, both of which appear to play a role in the conformation of the active site during catalysis. The dinoflagellates are distinct from all other eukaryotes in having a prokaryotic-type form II Rubisco23
(i.e. one lacking the small subunit characteristic of the eukaryotic form I). Although the speci-ficity of dinoflagellate form II Rubisco is higher than bacterial form II Rubisco24
, it is unlikely that it could support measured rates of photosynthesis without the presence of a carbon-concentrating mechanism.
A second phenomenon that might drive the short-term photosynthetic response in estuarine waters is the occurrence of state transitions25
, which are rapid adjustments in the relative magnitudes of the photosys-tem I (PS I) and photosysphotosys-tem II (PS II) antennae. State 2 (preferential excitation of PS I) is favored under conditions of high absolute irradiance and under darkness, whereas State 1 (preferential excitation of PS II) is favored under conditions in which the spectrum is dominated by red light. Consequently cells in estuarine waters are driven to State 2 at aphotic depths (depths to which light does not penetrate because of complete attenuation by the overlying water), to State 1 at intermediate depths where the spectrum is weighted towards long wavelengths (Fig. 3), and back to State 2 at the surface4,25
. It is not clear what effect, if any, state transitions have on pho-tosynthetic rates, but the interchange time constants are close enough to the rate of mixing that the transitions can be observed in cyanobacteria and chlorophytes from turbid waters25
. State transitions are unlikely to limit photosynthetic rates in the ocean because the timescale of mixing is slower, and because red light is more rapidly attenuated than blue light (Fig. 3). State transitions have yet to be docu-mented from chromophytic microalgae.
Photoprotection
The most complex group of photosynthetic responses includes the transients associated with photoprotection and photoinhibition. Fig. 2.(a) The first-order reaction time constants (t, in minutes) of photosynthetic responses
to increases and decreases in light intensity vary from 104min for changes in the chlorophyll
quota to 1021min for inter-conversion of the xanthophylls diadinoxanthin and diatoxanthin.
Note that time constants for increases in irradiance (denoted ‘up’) are shorter than for decreases (denoted ‘down’), except in the case of the chlorophyll-specific light-saturated rate of photosynthesis, Pm/Chl, where the kinetics are driven by changes in the chlorophyll quota.
The time constant defines the rate of change of species X over a time-step of duration Dt as: Xt 1 Dt5Xtexp(Dt/2tdown)
for a decrease in irradiance and as:
Xt 1 Dt5Xt[1 2exp(Dt/2tup)]
for an increase.
The time taken for the light level to either double or halve for cells in simulated oceanic and estuarine water (Fig. 3) is indicated by arrows. Sources for the time constants are given in parentheses. (b) The essence of mixing-based productivity models is defining a step-change in irradiance because of a cell’s motion through a light gradient (compare with Fig. 3) and imple-menting the time-dependence of photosynthetic response via time constants (tupand tdown) that
pho-Photoprotective mechanisms can broadly be described as those that either decrease the absorption of light energy (by reducing the absorption cross section or effective photosynthetic cross-section of the reaction centers), or provide alternative energy sinks when photosynthetic capacity is exceeded. Change in the excitation delivery includes rapid responses, such as induction of energy-dis-sipating pigments in xanthophyll cycles26,27
, and slower responses, such as photoacclimative changes in the size and pigment compo-sition of the antennae (Fig. 2). Two different xanthophyll cycles are found in chlorophytes and chromophytes: in chlorophytes, violaxanthin is de-epoxidated to zeaxanthin in a two-step pathway, with antheraxanthin as an intermediate; in chromophytes diadi-noxanthin is de-epoxidated to diatoxanthin in a single step. Although the chlorophytes have a pathway that is structurally similar to vas-cular plants’, the relationship between non-photochemical energy quenching and the level of de-epoxidation is different28
. Conversely, although chromophytes have a different pathway, the relation-ship between non-photochemical quenching and the level of de-epoxidation is comparable to that in vascular plants.
Other electron sinks
In addition to xanthophyll cycling, there are other mechanisms that might act as sinks for electrons when PS II activity exceeds photosynthetic capacity. For example, cyanobacteria lack a xan-thophyll cycle but exhibit strong Mehler activity at light saturation29
, and diatoms appear to be unique in using non-assimilatory nitrate reduction as a sink30
. Both pathways bleed off excess energy from the electron transport chain when NADPH turnover is operating at the maximum rate, preventing over-excitation of the photosyn-thetic antennae.
Photoinhibition
When the photoprotective mechanisms already described are exceeded, damaged PS II reaction centers, lacking a functional D1 protein, can accumulate. The reconstitution of functional PS II reaction centers can be described by first-order reaction kinetics when cells are moved to non-inhibitory irradiance31
. Modeling the accumulation of damaged D1 might be more difficult because of the diversity of photoprotective responses, the protective capacity of which depends on both the light history and nutrient status of the cell26
. However, photo-inactivation of D1 can be modeled by target theory32
, opening the possibility that photosynthesis could then be described using the relationship between the proportion of inactive PS II reaction centers and the quantum efficiency.
Kinetic models and the time-dependence of photosynthetic physiology
Short-term variability in photosynthetic rates can be accounted for in productivity models using time-dependent relationships. Generally, two assumptions are made, namely that the ocean can be considered as a one-dimensional system in the vertical and (in many models) that the time-dependence of photosynthetic processes follow first-order reaction kinetics. With an adequate understanding of the ver-tical variation in mixing rate, the trajectories of groups of cells can be described by a random walk simulation (the vertical steps of which are defined in time by the local turbulent diffusivity) using a Lagrangian ensemble model. Given the rate of light attenuation, changes in light intensity can be described as a function of depth and time (Fig. 3). The sensitivity of photosynthesis to mixing can then be described by specifying the time-dependence of the response to changing light intensity with one or more time constants (Fig. 2). Separate time constants for upward and downward movements are usually employed to account for different physiological rate constants associated with responses to increases and decreases in irradiance.
The approach has been used to model the sensitivity of photosyn-thesis to changes that are characteristic of photoacclimation33
, photoinhibition34
and photosynthetic induction18
. The relative importance of different photosynthetic mechanisms depends on the timescale and the magnitude of the changes in the light field (i.e. if the time constant for the metabolic response is longer than the characteristic timescale of change, the mechanism has the potential to limit the reaction rate13
; Fig. 2). Constructing a gen-eral model of the effect of mixing on photoinhibition that is analo-gous to the acclimative models depends on a somewhat arbitrary level of calibration because both the degree and rate of accumu-lation of photoinhibitory damage are less constrained than accli-mative changes (Fig. 2). One approach, which is based on first principles, involves defining an action spectrum of the dose-response of photoinhibition and using a biological weighting function to drive the onset of photoinhibition and the subsequent recovery as cells are mixed through a water column35
.
A description based on energy-balance models is an alternative to the descriptions of photoacclimation based on explicit time-constants of chlorophyll-specific photosynthetic responses. In energy-balance models, a physiological sensor regulates the pig-ment quota as a means of balancing excitation-energy harvesting with the energetic demands of growth. By specifying both as rates and using units of inverse time (which necessitates converting the currency of electrons to carbon equivalents), the imbalance can be used to drive changes in energy harvesting, either by inducing chlorophyll synthesis or D1 turnover10,11
until equilibrium is re-established. Embedding such an energy-balance model into a physical model of mixing produces results that are consistent with field measurements over wide temporal and spatial scales (annual cycles from 0 to 608N)36
. Because the regulated term is the ratio of phytoplankton chlorophyll to carbon (an index of the pigment quota), energy-balance models can be reconciled with large-scale estimates of phytoplankton biomass (Fig. 1), and the response has an explicit dependence on temperature and nutrient (nitrogen) availability. These dependencies have not been quantified for first-order kinetic models, in which both the time constants and the end-points of acclimation might depend on temperature and nutrient status.
The future
It is now possible to evaluate how the photosynthetic apparatus is regulated under complex environmental forcing. The com-plexity of the environment–photosynthesis relationship is tractable using the notion of energy-balance regulation: the redox state of the light reactions is a universal signal in micro-algae for regulating cellular light harvesting efficiency. The success of recent pigment-photoacclimation models that incor-porate an energy-balance regulatory ‘signal’ based on the redox state, holds promise for more sophisticated models that incorpo-rate short-term energy modulation mechanisms. These models can be made species-specific by setting parameters according to the specific energy modulation processes that are present. Future work on environmental effects will distinguish short-term kinetically constrained responses from longer-term energy-balance-constrained responses. This will clarify the relative importance of kinetic versus acclimative responses to particular environments.
Acknowledgements
We thank two anonymous reviewers for their comments on the manuscript. This work was supported by Grants OCE-9730098 and OCE-9633633 from the National Science Foundation (USA) and is contribution No. 3239 from Horn Point Laboratory.
References
1Falkowski, P.G. and Raven, J.A. (1997) Aquatic Photosynthesis, Blackwell Science 2Holmes, J.J. et al.(1989) Chlorophyll afluorescence predicts total
photosynthetic electron flow to CO2or NO32/NO 2
2under transient conditions.
Plant Physiol.91, 331–337
3Broekhuisen, N. et al.(1998) Seasonal photoadaptation and diatom dynamics in temperate waters. Mar. Ecol. Prog. Ser.175, 227–239
4Schubert, H. and Forester, R.M. (1997) Sources of variability in the factors used for modelling primary productivity in eutrophic waters. Hydrobiolgia349, 75–85 5Mouget, J-L. et al.(1995) Long-term acclimatization of Scenedesmus
bicellularisto high-frequency intermittent lighting (100 Hz). I. Growth, photosynthesis and photosystem II activity. J. Plankt. Res.17, 859–874 6Geider, R.J. et al.(1997) Dynamic model of phytoplankton growth and acclimation: responses of the balanced growth rate and the chlorophyll
a:carbon ratio to light, nutrient-limitation and temperature. Mar. Ecol. Prog. Ser.148, 187–200
7Orellana, M.V. and Perry, M.J. (1992) An immunoprobe to measure Rubisco concentrations and maximal photosynthetic rates of individual phytoplankton cells. Limnol. Oceanogr.37, 478–490
8Sukenik, A. et al.(1987) Light-saturated photosynthesis – limitation by electron transport or carbon fixation? Biochim. Biophys. Acta891, 205–215 9Escoubas, J-M. et al.(1995) Light-intensity regulation of CAB gene
transcription is signaled by the redox state of the plastiquinone pool. Proc. Natl. Acad. Sci. U. S. A.92, 10237–10241
10Geider, R.J. et al.(1998) A dynamic regulatory model of phytoplanktonic acclimation to light, nutrients and temperature. Limnol. Oceanogr.43, 679–694 11Kana, T.M. et al.(1997) Photosynthetic pigment regulation in microalgae by
multiple environmental sensors: a dynamic balance hypothesis. New Phytol.
137, 628–638
12Maxwell, D.P. et al.(1995) Redox regulation of light-harvesting complex II and cab mRNA abundance in Dunaliella salina. Plant Phys.109, 787–795 13Cullen, J.J. and Lewis, M.R. (1988) The kinetics of algal photoadaptation in
the context of vertical mixing. J. Plankt. Res.10, 1039–1063 14Geider, R.J. et al.(1998) Responses of the photosynthetic apparatus of
Dunaliella teriolecta(Chlorophyceae) to nitrogen and phosphorus limitation.
Eur. J. Phycol.33, 315–332
15Read, B.A. and Tabita, F.R. (1994) High substrate specificity factor ribulose bisphosphate carboxylase/oxygenase from eukaryotic marine algae and properties of recombinant cyanobacterial Rubisco containing ‘algal’ residue modifications. Arch. Biochem. Biophys.312, 210–218
16Zhu, G. et al.(1998) Dependence of catalysis and CO2/O2specificity of Rubisco on the carboxy-terminus of the large subunit at different temperatures. Photosynth. Res.57, 71–79
17MacIntyre, H.L. et al.(1997) Activation and deactivation of ribulose-1,5-bisphosphate carboxylase/oxygenase (Rubisco) in three marine microalgae.
Photosynth. Res.51, 93–106
18MacIntyre, H.L. and Geider, R.J. (1996) Regulation of Rubisco activity and its potential effect on photosynthesis during mixing in a turbid estuary. Mar. Ecol. Prog. Ser.144, 247–264
19Badger, M.R. and Price, G.D. (1992) The CO2concentrating mechanism in cyanobacteria and microalgae. Physiol. Plant.84, 606–615
20Tortell, P.D. et al.(1997) Active uptake of bicarbonate by diatoms. Nature
390, 243–244
21Berman-Frank, I. et al.(1998) Changes in inorganic carbon uptake during the progression of a dinoflagellate bloom in a lake ecosystem. Can. J. Bot.76, 1043–1051
22Nimer, N. et al.(1999) Extracellular carbonic anhydrase facilitates carbon dioxide availability for photosynthesis in the marine dinoflagellate
Prorocentrum micans. Plant. Physiol.120, 105–111
23Morse, D. et al.(1995) A nuclear-encoded form II Rubisco in dinoflagellates.
Science268, 1622–1624
24Whitney, S.M. and Andrews, T.J. (1998) The CO2/O2specificity of single-subunit ribulose-bisphosphate carboxylase from the dinoflagellate,
F
loral induction is the process by which stimuli originating outside the shoot apex induce the formation of flower pri-mordia (Fig. 1). The photoperiodic induction of flowering was discovered 86 years ago by Julien Tornois in hops1. Shortly afterwards, additional experiments suggested that the photoperi-odic control of flowering was a general phenomenon, which con-trolled flowering in most plants2
. Later, focused-light experiments showed that leaves perceive photoperiodic signals3
. These studies, and numerous grafting experiments, indicate that the production of the photoperiod-induced floral stimulus4
occurs in the leaves of a wide variety of flowering plants5–7
.
In contrast with floral induction, floral determination can be defined as the assignment of flower(ing) fate, which is persistent even when the flower-inducing conditions no longer exist8,9
. Assays for floral determination include:
• Changing environmental conditions (from inductive to non-inductive).
• Microsurgical removal of shoot apices, and the placement of those apices into neutral environments8,10
.
However, both types of determination assay have limitations, and it is important to note that different determination assays might yield alternative conclusions for the same primordia (the caveats associated with determination experiments are discussed in Ref. 11). A third type of assay has been used to test leaf commitment to the continued production of floral stimulus: in this assay, photo-induced leaves are removed from the plant following an inductive treatment12
.
In this review we discuss firstly a variety of experiments that indicate the site(s) that control flowering. Secondly, we review recent studies that indicate how a few key molecular players regu-late the specification of flower primordia in Arabidopsis. Floral determination assays
Photoperiodic assays for floral determination
The simplest type of determination assay is one in which plants are moved to non-inductive conditions after various lengths of time under inductive conditions. Using this method, the duration of photoinduction treatment required to produce flowers can be
Floral induction and determination:
where is flowering controlled?
Frederick D. Hempel, David R. Welch and Lewis J. Feldman
Flowering is controlled by a variety of interrelated mechanisms. In many plants, the environment controls the production of a floral stimulus, which moves from the leaves to the shoot apex. Apices can become committed to the continuous production of flowers after the receipt of sufficient amounts of floral stimulus. However, in some plants, the commitment to continued flower production is evidently caused by a plant’s commitment to perpetually produce floral stimulus in the leaves. Ultimately, the induction of flowering leads to the specification of flowers at the shoot apex. In Arabidopsis, floral specification and inflorescence patterning are regulated largely by the interactions between the genes TERMINAL FLOWER, LEAFY and APETALA1/CAULIFLOWER.
25Schubert, H. et al.(1995) In situmeasurement of state transition in cyanobacterial blooms: kinetics and extent of the state change in relation to underwater light and vertical mixing. Mar. Ecol. Prog. Ser.128, 99–108 26Arsalane, W. et al.(1994) Influence of the pool size of the xanthophyll cycle
on the effects of light stress in a diatom: competition between photoprotection and photoinhibition. Biochem. Photobiol.60, 237–243
27Olaizola, M. et al.(1994) Non-photochemical fluorescence quenching and the diadinoxanthin cycle in a marine diatom. Photosynth. Res.41, 357–370 28Casper-Lindley, C. and Björkman, O. (1998) Fluorescence quenching in four
unicellular algae with different light-harvesting and xanthophyll-cycle pigments. Photosynth. Res.56, 277–289
29Kana, T.M. (1992) Relationship between photosynthetic oxygen cycling and carbon assimilation in SynechococcusWH7803 (Cyanophyta). J. Phycol.28, 304–308
30Lomas, M.W. and Glibert, P.M. (1999) Temperature regulation of nitrate uptake: a novel hypothesis about nitrate uptake and reduction in cool-water diatoms. Limnol. Oceanogr.44, 556–572
31Neidhardt, J. et al.(1998) Photosystem-II repair and chloroplast recovery from irradiance stress: relationship between chronic photoinhibition, light-harvesting chlorophyll antenna size and photosynthetic productivity in
Dunaliella salina(green algae). Photosynth. Res.56, 175–184 32Anderson, J.A. et al.(1998) Unifying model for the photoinactivation of
photosystem II in vivounder steady-state conditions. Photosynth. Res.56, 1–13
33Barkmann, W. and Woods, J.D. (1996) On using a Lagrangian model to calibrate primary production determined from in vitroincubation measurements. J. Plankt. Res.18, 767–788
34Franks, P.J.S. and Marra, J. (1994) A simple new formulation for
phytoplankton photoresponse and an application in a wind-driven mixed-layer model. Mar. Ecol. Prog. Ser.111, 143–153
35Neale, J.P. et al.(1998) Inhibition of marine photosynthesis by ultraviolet radiation: variable sensitivity of phytoplankton in the Weddell–Scotia confluence during the austral spring. Limnol. Oceanogr.43, 433–448 36Taylor, A.H. et al.(1997) Seasonal and latitudinal dependencies of
phytoplankton carbon-to-chlorophyll aratios: results of a modelling study.
Mar. Ecol. Prog. Ser.152, 51–66
Hugh L. MacIntyre*and Todd M. Kana are at the University of Maryland Center for Environmental Research, Horn Point Laboratory, PO Box 775, Cambridge, MD 21613, USA; Richard J. Geider is at the Dept of Biological Sciences, University of Essex, Colchester, UK CO3 4SQ.