PERAN IMMUNOMODULATOR DALAM
MENGAKTIFKANKAN RESPON IMUN TERHADAP
INFEKSI VIRUS
OLEH :
A.A. Gde Arjana
KATA PENGANTAR
Atas asung kerta wara nugraha Ida Sang Hyang Widhi Wasa
penulis dapat menyelesaiakan penulisan karya ilmiah dengan judul :
Peran Immunomodulator Dalam Mengaktifkan Respon immun
Terhadap Infeksi Virus
Penulisan karya ilmiah ini bertujuan untuk menambah pengetahuan
tentang bagaimana peran immunomodulator dalam mengaktifkan respon
immun terhadap infeksi virus.
Terima kasih penulis sampaikan kepada semua pihak yang telah
membantu sehingga karya ilmiah ini dapat diselesaikan dengan baik.
Demikianlah penulis dapat sampaikan semoga Ida Sang Hyang
Widhi Wasa melimpahkan segala rahmatNya kepada kita semua.
Denpasar, Januari 2016
DAFTAR ISI
2.4. Respon imun non Spesifik Terhadap Virus ………. 7
2.4.1. Makrofag ……… 7
2.4.2. Sel Natural Killer (NK) ……….. 10
2.4.3. Interferon (INF ) ………. 12
2.4.4. Interleukin 6 (IL-6) ………. 12
2.5. Respon Imum Spesifik Terhadap Virus ………. 14
2.5.1. Limfosit T ……….... 14
2.5.2. Limfosit B ……… 16
2.5.3. Interleukin 2 (IL-2) ………. 17
BAB III. KESIMPULAN ………... 18
DAFTAR GAMBAR
GAMBAR 1. Mekanisme Stimulasi Imun Non Spesifik ………. 5
RINGKASAN
Sistem Immun tubuh terdiri dari banyak komponen. Semua komponen tersebut akan bekerja serentak manakala tubuh mendapat serangan dari penyakit yang berasal dari luar maupun dari dalam tubuh.
Tubuh dalam melindungi diri dari serangan mikroorganisme pathogen terutama virus dengan cara mengembangkan sistem pertahanan tubuh. Sistem pertahanan tubuh dapat diaktifkan dengan memberikan suatu senyawa yang dapat digunakan untuk meningkatkan respon immun yang disebut immunomodulator.
BAB I
PENDAHULUAN
1.1. Latar Belakang
Sistem immun tubuh terdiri dari banyak komponen. Semua
komponen tersebut akan bekerja serentak manakala tubuh mendapat
serangan dari penyakit yang berasal dari luar maupun dari dalam tubuh .
Sistem imun yang bertugas mengatur keseimbangan, dengan
menggunakan komponennya yang beredar diseluruh tubuh, sehingga
dapat mencapai sasaran yang jauh dari pusat. Untuk melaksanakan
fungsi imunitas, didalam tubuh terdapat suatu sistem yang disebut dengan
sistem limforetikuler. Sistem limforetikuler merupakan jaringan atau
kumpulan sel yang letaknya tersebar diseluruh tubuh, misalnya didalam
sumsum tulang, kelenjar limfe, limfa, timus, sistem saluran napas, saluran
cerna dan beberapa organ lainnya. (Tizard, 2004; Rabson dkk., 2005).
Rangsangan terhadap sel-sel tersebut terjadi apabila kedalam
tubuh terpapar suatu zat yang oleh sel atau jaringan tadi dianggap asing.
Konfigurasi asing ini dinamakan antigen atau imunogen dan proses serta
fenomena yang menyertainya disebut dengan respons imun yang
menghasilkan suatu zat yang disebut dengan antibodi. Jadi antigen atau
imunogen merupakan potensi dari zat-zat yang dapat menginduksi
respons imun tubuh yang dapat diamati baik secara seluler ataupun
membedakan zat asing (non-self) dari zat yang berasal dari tubuhnya
sendiri (self), sehingga sel-sel dalam sistem imun membentuk zat anti
terhadap jaringan tubuhnya sendiri. Kejadian ini disebut dengan
Autoimmun(Abbas dkk., 2007; Baratawijaya, 2010).
Tubuh dalam melindungi diri dari serangan mikroorganisme
pathogen dengan mengembangkan sistem pertahanan tubuh. Sistem
pertahanan tubuh dapat diaktifkan dengan memberikan suatu senyawa
yang dapat digunakan untuk meningkatkan respon immun yang disebut
immunomodulator. Immunomudulator ini dapat meningkatkan mekanisme
pertahanan tubuh baik secara spesifik (adaptive immune system) maupun
non spesifik ( innate immune system). . Kedua respon imun tersebut
dalam bekerjanya melibatkan berbagai komponen seluler maupun zat
terlarut seperti sitokin, kemokin dan komplemen (Tizard, 2004;
Baratawidjaya, 2010) .
1.2. Rumusan Masalah
Bagaimana peran immunomodulator dalam mengaktifkan respon
BAB II
PEMBAHASAN
2.1 Penyakit Infeksi
Penyakit infeksi merupakan salah satu penyakit yang sering
menggangu kesehatan, baik kesehatan manusia maupun kesehatan
hewan. Penyakit ini, dapat disebabkan oleh berbagai jenis bakteri, virus,
jamur dan parasit. Secara umum gangguan yang ditimbulkan dapat
bersifat, akut dan kronis, yang disertai dengan berbagai gejala dari ringan
sampai berat dan bahkan tidak sedikit mengakibatkan kematian. Salah
satu penyakit infeksi yang sering menyebabkan kerugian yang cukup
besar baik pada manusia maupun pada hewan adalah penyakit virus.
Penyakit virus pada hewan merupakan salah satu faktor
penghambat perkembangan peternakan. Penyakit ini banyak
menimbulkan kerugian yang besar pada dunia peternakan terutama
kerugian materiil terutama karena dapat menimbulkan kematian.
Disamping menimbulkan kerugian materiil penyakit virus dapat bersifat
zoonosis yaitu dapat menular dari hewan ke manusia. Beberapa contoh
penyakit virus yang bersifat zoonosis adalah Avian Influenza (AI/Flu
Burung) dan rabies. (Deptan, 2005, Davidson, dkk, 2008).
Untuk menanggulangi penyakit virus usaha pengendalian dilakukan
dengan jalan vaksinasi , akan tetapi wabah penyakit virus pada hewan
Kegagalan vaksinasi disebabkan oleh berbagai faktor antara lain : faktor
vaksinnya, pelaksananya (vaksinator), dan faktor hewannya.
Vaksinator dapat melakukan kesalahan dalam pemilihan vaksin,
pengangkutan, penyimpanan, pengenceran dan pengaplikasiannya.
Gangguan pada hewannya bisa disebabkan oleh adanya infeksi dari
mikroorganisme yang merugikan seperti infeksi parasit atau infeksi oleh
mikroorganisme yang dapat mengakibatkan imunodefisiensi. Aplikasi
vaksin pada hewan dapat dilakukan melalui tetes mata (intra oculer), tetes
hidung (intra nasal), air minum (drinking water), dan melalui suntikan
(injection) (Tizad, 2004; Webster dkk, 2004).
Untuk mengatasi hal tersebut diatas dipandang perlu utnuk
memberikan bahan-bahan yang bersifat sebagai immunomodulator,
sehingga dapat meningkatkan antibodi. Imumunomodulator atau
imunopotensiasi adalah cara memperbaiki fungsi sistem imun dengan
menggunakan imunostimulan yaitu bahan yang merangsang sistem imun.
2.2 Immunomodulator
sebagai mitogen yaitu menaikkan prolifirasi sel yang berperan pada
immunitas. Sel tujuan adalah makrofag, granulosit, limfosit Tdan B, karena
induktor paraimunitas ini terutama menstimulasi mekanisme pertahanan
selluler. Mitogen ini dapat bekerja langsung maupun tidak langsung
(misalnya melalui sistem komplemen atau limfosit, melalui produksi
interferon atau enzim lisosomal) untuk meningkatkan fagositosis mikro
dan makro.
Beberapa bahan yang sering digunakan sebagai immunomodulator
adalah levamisol, isoprinosin, muramil dipeptida (MDP), hidroksiklorokin,
arginin, antioksidan, mikroorganisme, polinukleotida, limfokin dan
dengan herbal dapat digunakan sebagai immunomodulator adalah
temuputih, ketepeng cina, lidah buaya, buah merah,mahkota dewa .
(Mathilda., 1987; Janeway dkk., 1999; Tizard, 2004; Abbas dkk., 2007;
Bratawijaya, 2010)
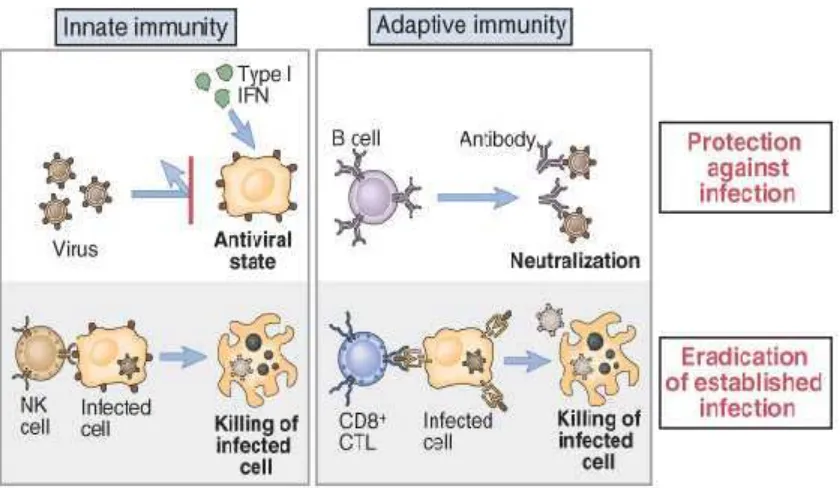
2.3 Imunitas terhadap infeksi virus
Virus, merupakan organisme obligat, umumnya terdiri atas
potongan DNA atau RNA yang diselubungi envelop dari protein atau
lipoprotein. Respon imun terhadap protein virus melibatkan respon imun
secara nonspesifik yaitu makrofag, sel NK, interferon, interleukin (IL)- 1,
dan IL-6, sedangkan peningkatan respon imun secara spesifik melibatkan
limfosit T, limfosit B dan IL-2 (Abbas dkk., 2007; Baratawidjaya, 2010).
Gambar 2.
Imunitas terhadap Infeksi Virus
2.4 Respon Imun non Spesifik terhadap Infeksi Virus
2.4.1 Makrofag
Adanya agen infeksi yang masuk ke dalam tubuh akan
menyebabkan makrofag mengalami aktivasi. Aktivasi makrofag ini diikuti
dengan meningkatnya kemampuan morfologis, metabolisme, dan
fungsional. Secara morfologis, makrofag tampak lebih besar dengan
asam fosfatase, lisozim, beta glukoronidase, esteroprotease, hidrolise,
myeloperoksidase, dan arilsulfatase akan meningkat. Meningkatnya
kemampuan fungsional makrofag ditandai dengan meningkatnya aktivitas
makrofag, kapasitas fagosit makrofag, dan produksi interleukin (Janeway
dkk., 1999; Fernandes, dkk, 2000; Kusmardi dkk., 2007).
Fungsi utama makrofag dalam imunitas nonspesifik adalah
memfagosit partikel asing yang masuk tubuh seperti kuman, virus, parasit,
dan sel tumor. Fagositosis juga dilakukan terhadap sel atau jaringan
sendiri yang mengalami kerusakan atau mati. Antigen yang berada di
dalam fagolisosom tersebut akan didenaturasi atau didegradasi menjadi
partikel peptida. Selanjutnya peptida ini diikat oleh MHC dan dibawa ke
permukaan sel untuk disajikan ke sel T. Selama proses fagositosis dan
penyajian antigen, makrofag mengeluarkan bahan biologik yang dikenal
dengan interleukin. Interleukin ini merupakan alat komunikasi antar sel.
Ada beberapa interleukin (IL) yang dikeluarkan oleh makrofag yaitu IL-1,
IL2, IL-4, IL-6, dan TNF. Pada dasarnya interleukin ini berperan penting
enzim di dalam makrofag berhubungan dengan digesti intraseluler
material yang difagosit, perkembangan dan mempertahankan reaksi
radang dan pembunuhan mikrobia (Rabson, 2005; Tizard, 2004). Lebih
lanjut dikatakan bahwa aktivasi makrofag akan memacu aktivitas
mikrobisidal, meningkatkan kapasitas tumorisidal, mempercepat
pergerakannya, meningkatkan kemampuan produksi sejumlah
bahan-bahan yang penting, dan memacu sel imunokompeten untuk
menghasilkan antibodi.
Antigen yang disajikan oleh makropag sebagai antigen precenting
cell (APC) ke limfosit T merupakan tahap awal terjadinya respon imun. Di
dalam makrofag, antigen diproses dengan cara denaturasi atau
proteolisis. Sementara itu molekul Major Histocompatibility Complex
(MHC) yang terdapat dalam lisosom mengenali segmen antigen lalu
dibawa ke permukaan sel dan disajikan kepada sel T. Makrofag yang
memiliki CD38 merupakan penghubung antara makrofag dengan sel.
Molekul MHC pada APC bertindak sebagai reseptor primer antigen
(Tizard, 2004; Bryniarski dkk., 2005).
Pada infeksi virus maka MHC I menangkap peptida yang dihasilkan
oleh virus di dalam retikulum endoplasma, lalu dibawa ke permukaan sel
dan disajikan ke sel T. MHC yang ada di dalam retikulum endoplasma
tidak bisa leluasa bergerak ke organel lainnya, jika rantai pendeknya tidak
ke permukaan sel dan disajikan ke sel T sitotoksik melalui reseptor (TCR).
Proses selanjutnya sel T sitotoksik mengeluarkan bahan toksik sehingga
sel penyajinya akan terbunuh (Tobian dkk., 2003).
Pada fagolisosom terjadi pemrosesan antigen yang meliputi proses
oksidasi, non oksidasi, dan degranulasi. Fragmen-fragmen antigen yang
terbentuk, akan diikat oleh molekul MHC II selanjutnya dibawa ke
permukaan sel untuk disajikan ke sel T. Sel T helpermelalui reseptornya
(TCR) akan mengenal antigen yang disajikan oleh makrofag. Ligan antara
kompleks antigen-MHC pada sel penyaji dengan kompleks CD3-TCR
pada sel T helper membangkitkan aktivitas inositol pada membran sel T
menjadi inositol trifosfat dan senyawa gliserol dalam sitoplasma. Inositol
trifosfat akan meningkatkan ion Ca++ dalam sitoplasma, sedangkan
diasilgliserol akan mengaktifkan enzim proteinkinase C. Keduanya
merupakan sinyal untuk mengaktifkan sel T. Namun kedua sinyal itu
belum cukup untuk mengaktifkan sel T, karena itu masih memerlukan
sinyal ketiga yang diawali oleh IL-1 yang dilepaskan oleh makrofag.
memiliki petanda sel B atau sel T atau imunoglobulin permukaan. Sel NK
memiliki banyak sitoplasma, granul sitoplasma azurofilik, pseudopodia dan
nukleus eksentris. Sel NK merupakan sumber interferon γ (INF-γ) yang
mengaktifkan makrofag dan berfungsi dalam imunitas nonspesifik
terhadap virus dan sel tumor. Sel NK mengenal dan membunuh sel
terinfeksi atau sel yang menunjukkaan transformasi ganas, tetapi tidak
membunuh sel sendiri yang normal oleh karena dapat membedakan sel
sendiri dari sel yang potensial berbahaya, akibat adanya reseptor inhibitori
dan reseptor aktivasi (Bottino dkk., 2005; Baratawidjaya, 2010)
Sel NK mengenal MHC-I yang diekspresikan semua sel sehat dan
tidak oleh sel terinfeksi virus dan kanker. Pengaruh reseptor inhibitori akan
dominan dan mengikat MHC-I yang normal diekspresikan pada sel sehat.
Sel NK membunuh sel yang terinfeksi oleh berbagai jenis virus dan
merupakan efektor imunitas penting terhadap infeksi virus secara dini,
sebelum respon imun spesifik bekerja. Sel NK mengenal sel terinfeksi
walaupun tidak mengekspresikan MHC-I. Untuk membunuh virus sel NK
tidak memerlukan bantuan molekul MHC- I. Sel NK memiliki reseptor
aktivasi dapat merupakan pembunuh poten sel terinfeksi virus, jamur dan
tumor dengan langsung, tanpa bantuan komplemen. Fenomena ini
disebut Antibody Dependent Cell Cytotoxicity (ADCC) (Bottino dkk., 2005;
2.4.3 Interferon γ (INF-γ)
Interferon γ diproduksi oleh berbagai sel imun, merupakan sitokin
utama Makrofag Activating Cytokain (MAC) dan berperan terutama dalam
imunitas non spesifik. Interferon γ adalah sitokin yang mengaktifkan
makrofag untuk membunuh virus. INF-γ merangsang ekspresi Major
Histocompatibility Complex(MHC-I) , MHC-II dan konstimulator APC.
INF-γ meningkatkan diferensiasi sel CD4+ naif kesubset sel Th1 dan
mencegah proliferasi sel Th2. INF-γ bekerja terhadap sel B dalam
pengalihan subkelas IgG yang mengikat Fcγ-R pada fagosit dan
mengaktifkan komplemen.
Efek protektif INF-γ terjadi melalui reseptor di membran sel dan
mengaktifkan gen yang menginduksi sel untuk memproduksi protein anti
virus yang mencegah translasi mRNA virus. INF-γ dapat mengaktifkan
fagosit, APC dan induksi pengalihan sel B, menginduksi tidak langsung
efek Th1 atas peran peningkatan produksi Il-12 dan ekspresi reseptor
keratinosit, fibroblas, adiposit, sel otot polos vaskuler, osteoblas, sel
stroma sumsum tulang, sinoviosit, kondrosit, sel Leydig testis, sel stroma
endometrium, dan trofoblas. Pada sistem saraf pusat, IL–6 diekspresikan
oleh astrosit, sel mikroglia, dan sel folikulostelata hipotalamus (Dostatni
dkk., 1996; Klipinen, 2003; Baratawidjaja, 2006).
Pada percobaan in vivo dan in vitro keikutsertaan IL–6 dalam
aktivasi sel T dan differensiasi sel B dapat diperlihatkan. IL–6 bersama IL–
2 dapat mengendalikan differensiasi sel T menjadi sel T sitotoksik.
Aktivasi sel T sitotoksik ini menghasilkan IL-4. IL-4 bersama dengan IL-6
merangsang diferensiasi sel B menjadi sel plasma untuk menghasilkan
imunoglobulin.. Disamping IL-6 berperan dalam diferensiasi sel B, IL-6
juga berperan sebagai faktor hemopoetik yaitu merangsang proliferasi dan
diferensiasi sel megakariosit dan meningkatkan jumlah platelet (Dostatni
dkk., 1996; Klipinen, 2003 ; Baratawidjaja, 2010). IL–6 bekerja sebagai
faktor diferensiasi sel B yang bertanggung jawab dalam pematangan akhir
sel B menjadi sel plasma dan meningkatkan produksi IgM, Ig G dan Ig A.
IL–6 merangsang proliferasi Thymosit dan sel T perifer, serta mendukung
aktivasi, proliferasi, dan diferensiasi sel NK. (Dostatni dkk., 1996;
2.5 Respon Imun Spesifik terhadap Infeksi Virus
Respon imun spesifik terhadap infeksi virus diperankan oleh :
2.5.1 Limfosit T
Progenitor limfosit T berasal dari sumsum tulang yang bermigrasi
ke timus, berdiferensiasi menjadi sel T. Sel T yang non aktif disirkulasikan
melalui kelenjar getah bening (KGB) dan limfa yang dikonsentrasikan
dalam folikel dan zona marginal sekitar folikel. Sel T imatur dipersiapkan
dalam timus untuk memperoleh reseptor. Timosit imature hanya dapat
menjadi matang bila reseptornya tidak berintegrasi dengan peptida sel
tubuh sendiri (self antigen) yang diikat MHC dan dipresentasikan oleh
APC. Sawar darah timus melindungi timosit dari kontak dengan antigen
sendiri. Sel T yang self reaktip akan mengalami apoptosis. Proses ini
disebut seleksi positip timosit yang menghasilkan sel Tcytotoxic (Tc) atau
sel Thelper(Th) (Abbas dkk., 2007; Baratawidjaya, 2010). Kemampuan
limfosit T matang untuk mengenal benda asing, karena adanya T Cell
Sel T umumnya berperan pada inflamasi, aktivasi fagositosis
makrofag, aktivasi dan proliferasi sel B dalam produksi antibodi. Sel T juga
berperan dalam pengenalan dan penghancuran sel yang terinfeksi virus.
Sel T terdiri atas sel T helper (Th) yang mengaktifkan makrofag untuk
membunuh mikroba dan sel Tcytotoxic(Tc) yang membunuh sel terinfeksi
mikroba atau virus dan menyingkirkan sumber infeksi. Sel T terdiri atas sel
CD4+, CD8+, sel T naif dan sel Natural Killer T (NKT) (Germain, 2002;
Baratawidjaya, 2010).
Sel limfosit naif adalah sel limfosit matang yang meninggalkan
timus dan belum berdiferensiasi, belum pernah terpapar antigen dan
menunjukkan molekul permukaan CD45RA. Sel T helper disebut juga sel
T inducer merupakan subset sel T yang diperlukan dalam induksi respon
imun terhadap antigen asing. Antigen yang ditangkap, diproses dan
dipresentasikan makrofag dalam konteks MHC-II ke sel CD4+. Selanjutnya
sel CD4+ diaktifkan dan memproduksi IL-2 autokrin yang merangsang sel
CD4+ untuk berproliferasi menjadi subset sel Th1 dan Th2, mensintesis
sitokin yang mengaktifkan sel imun lain seperti CD8+, sel B makrofag dan
sel NK (Germain, 2002; Abbas dkk., 2007; Baratawidjaya, 2010).
Sel T CD8+ naif yang keluar dari timus disebut juga Cytolitic T
(CTL) atau Citotoxic T (Tc). CD8+ mengenal kompleks antigen MHC-I
yang dipresentasikan APC. Molekul MHC I ditemukan pada semua sel
terinfeksi virus, menghancurkan sel ganas dan sel histoin kompatibel yang
menimbulkan penolakan pada transplantasi. Sel Tc menimbulkan sitolisis
melalui perforin/granzim (apoptosis), TNF-α dan memacu produksi sitokin
Th1 dan Th2 (Hewitt, 2003; Baratawidjaya, 2010).
2.5.2 Limfosit B
Sel B diproduksi pertama selama fase embrionik dan berlangsung
terus selama hidup. Sebelum lahir yolk sac, hati dan sumsum tulang janin
merupakan tempat pematangan utama sel B dan setelah lahir
pematangan sel B terjadi di sumsum tulang. Pematangan sel B terjadi
dalam berbagai tahap. Pada unggas, sel B berkembang dalam bursa
fabricius yang terbentuk dari epitel kloaka. Pada manusia belum
didapatkan hal yang analog dengan bursa tersebut dan pematangan sel B
terjadi di sumsum tulang atau ditempat yang belum diketahui. Setelah
matang sel B bergerak ke organ limpa, kelenjar getah bening dan tonsil
(Busslinger, 2004; Baratawidjaya, 2010).
perkembangan selanjutnya memerlukan rangsangan antigen. Sel B yang
diaktifkan berkembang menjadi limfoblas, selanjutnya menjadi sel plasma
yang memproduksi antibodi dan sel memori (Busslinger, 2004; Abbas
dkk., 2007).
2.5.3 Interleukin 2 (IL-2)
Interleukin 2 adalah faktor pertumbuhan sel T yang dirangsang
antigen dan berperan pada ekspansi klon sel T setelah antigen dikenal.
Ekspresi reseptor IL-2 ditingkatkan oleh rangsangan antigen, oleh karena
itu sel T yang mengenal antigen merupakan sel utama yang berproliferasi
pada respons imun spesifik. IL-2 meningkatkan proliferasi dan
diferensiasi sel T, sel B dan NK. IL-2 juga mencegah respons imun
terhadap antigen sendiri melalui peningkatan apoptosis sel T
(Baratawijaya, 2010).
Peningkatan IL-2 dalam tubuh akan meningkatkan produksi CD4+,
dengan demikian IL-2 juga berfungsi sebagai imunomodulator yaitu
pengaturan menyeluruh sistem imun di dalam tubuh, baik dalam keadaan
normal maupun abnormal. Pemberian IL-2 telah terbukti dapat menekan
pertumbuhan beberapa tipe kanker. Treatmen penyakit HIV dengan
menggunakan IL-2 jugasudah pernah dilakukan walaupun hasilnya belum
BAB III
KESIMPULAN
Sistem immun tubuh terdiri dari banyak komponen. Semua
komponen tersebut akan bekerja serentak manakala tubuh mendapat
serangan dari penyakit yang berasal dari luar maupun dari dalam tubuh.
Tubuh dalam melindungi diri dari serangan mikroorganisme
patogen terutama virus dengan cara mengembangkan sistem pertahanan
tubuh. Sistem pertahanan tubuh dapat diaktifkan dengan memberikan
suatu senyawa yang dapat digunakan untuk meningkatkan respon immun
yang disebut immunomodulator.
Immunomodulator adalah senyawa yang dapat meningkatkan
mekanisme pertahanan tubuh baik secara spesifik maupun non spesifik.
Immunomudulator dapat mengaktifkan respon immun yang non spesifik
yaitu makrofag, sel NK, interferon, interleukin (IL)1 dan IL6, dan respon
DAFTAR PUSTAKA
Abbas, A.K., Lichtman, A.H., Pillai S. 2007. Cellular and Molecular Immunology. 6th ed. WB Saunders Company Saunders, Philadelphia.Pp : 19-351.
Busslinger M. 2004. Transcriptional control of early B cell Development. Annual Review of immunology 22:55-79.
Baratawidjaja, K.G., Rengganis I. Imunologi Dasar ed. 9. Jakarta. BP.FKUI. 2010. hal: 27-217.
Beckerman, K.P. and Dudley, D.J. 2001. Reproduction & the Immun System. In Medical Immunology a Lange medical Book 10 th ed. Philadhelpia, Mc Graw Hill. p:563-67.
Bryniarski ,K., Szczepanik, M., Ptak, M., Ptak, W. 2005. Modulation of testicular macrophage activity by collagenase. Folia histochemica et cytobiologica Vol. 43, No. 1. p : 37-41.
Bottino C,R Castriconi, L Moretta, and A Moretta. 2005. Cellular Ligands of Activating NK Receptors. Trends in Immunology 26:221-226.
Davison, F., Kaspers, B.,Karel A. Schat. 2008. Avian Immunology. An Introduction First edition. Elsevier. USA Pp : 51-101.
Departemen Pertanian (2005). Buku pedoman dan Pencegahan Flu Burung (Avian Influenza) pada Peternakan Unggas skala kecil. Buku Petunjuk Mengenai Avian Influenza. Direktorat Jendral Peternakan Departenen Pertanian. Jakarta.
Dostatni, R., Berthold, S., Biermann. 1996. Interleukin-6 in Intensive care Medicine. Diagnostic Products Corporation. p: 1-15.
Fernandez, M.E.H. and Lopez, D.M. 2000. Isolation of macrophages from tissues, fluids, and immune response sites. Macrophages. Apractical Approach. Edited By Paulnock DM. Oxford University Press. Pp 1-4.
Germain RN. 2002. T cell development and the CD4-CD8 liniage decision. Nature Reviews Immunology 2:309-322.
Janeway, C.A. Jr, Travers, P., Walport, M., Capra, J.D. 1999. Immunobiology. The Immune System in Health and Disease. 4 th ed. USA. Garland Publishing. Pp. 79-263.
Klipinen, S. Inflammatory cytokines and their promoter polymorphisms. Academic dessertation University of tampere, Tampere, 2003. p:19-27
Kusmardi, Kumala, S., Triana, E.E. 2007. Efek immunomodulator ekstrak daun ketepeng cina (Cassia alataL.) terhadap aktivitas fagositosis makrofag. Makara, Kesehatan,Vol. 11, NO. 2. Pp 50-53.
Mathilda, B.W. (1987). Immunomodulator. Cermin Dunia Kedokteran. No.44. 43-45
Muthmainah. 2004. Studi tentang aktivitas sekresi reactive oxygen intermediates (ROIs) makrofag mencit yang distimuli dengan stimulant spesifik dan non spesifik selama infeksi Toxoplasma gondii. Laboratorium Histologi. Fakultas Kedokteran. Universitas Sebelas Maret. Surakarta. Jurnal BioSMART Vol 6, No. 2. 2004 : 1-2.
Noss, E.H., Pai, R.K., Sellati, T.J., Radolf, J.D., Belisle, J., Golenbock, D.T., Boom, W.H., Harding, C.V. 2001. Toll-like Receptor 2-dependent inhibition of macrophage class II MHC expression and antigen processing by 19-kDa lipoprotein of M. tuberculosis. J Immunol. Jul 15;167(2):910-8.
Rabson A, Roitt IM, Delves PJ. Really. 2005. Essential Immunology Second Edition. Oxford. Blackwell Publishing Pp : 27-98.