i
DISERTASI
PENGARUH VIRGIN COCONUT OIL DALAM
FORMULA WHO DIBANDINGKAN DENGAN
MINYAK JAGUNG TERHADAP GLUTATHIONE,
MALONDIALDEHYDE, TNF-
α
DAN PERLEMAKAN
HATI SERTA BERAT BADAN TIKUS WISTAR
MALNUTRISI BERAT
I GUSTI AYU PUTU EKA PRATIWI
PROGRAM PASCASARJANA
UNIVERSITAS UDAYANA
ii
DISERTASI
PENGARUH VIRGIN COCONUT OIL DALAM
FORMULA WHO DIBANDINGKAN DENGAN
MINYAK JAGUNG TERHADAP GLUTATHIONE,
MALONDIALDEHYDE, TNF-
α
DAN PERLEMAKAN
HATI SERTA BERAT BADAN TIKUS WISTAR
MALNUTRISI BERAT
I GUSTI AYU PUTU EKA PRATIWI NIM 1190271003
PROGRAM DOKTOR
PROGRAM STUDI ILMU KEDOKTERAN
PROGRAM PASCASARJANA
UNIVERSITAS UDAYANA
DENPASAR
iii
PENGARUH VIRGIN COCONUT OIL DALAM
FORMULA WHO DIBANDINGKAN DENGAN
MINYAK JAGUNG TERHADAP GLUTATHIONE,
MALONDIALDEHYDE, TNF-
α
DAN PERLEMAKAN
HATI SERTA BERAT BADAN TIKUS WISTAR
MALNUTRISI BERAT
Disertasi Untuk Memperoleh Gelar Doktor
Pada Program Doktor, Program Studi Ilmu Kedokteran, Program Pascasarjana Universitas Udayana
I GUSTI AYU PUTU EKA PRATIWI NIM 1190271003
PROGRAM DOKTOR
PROGRAM STUDI ILMU KEDOKTERAN
PROGRAM PASCASARJANA
UNIVERSITAS UDAYANA
DENPASAR
iv
Lembar Persetujuan Promotor/Kopromotor
UJIAN TERTUTUP DISERTASI INI TELAH DISETUJUI PADA TANGGAL: 26 APRIL 2016
Promotor,
Prof. dr. Soetjiningsih, Sp.A(K) NIP. 194501241971062001
Kopromotor I, Kopromotor II,
Prof. drh. I Nyoman Mantik Astawa, Ph.D Dr.dr.IGA Trisna Windiani,Sp.A(K) NIP. 196001231988031003 NIP. 196912091999032001
Mengetahui:
Ketua Program Studi Ilmu Kedokteran Direktur Program Pascasarjana Program Pascasarjana Universitas Udayana, Universitas Udayana,
v
Ujian Tertutup Disertasi ini Telah Diuji dan Dinilai oleh Panitia Penguji pada
Program Pascasarjana Universitas Udayana pada Tanggal 26 April 2016
Berdasarkan SK Direktur Program Pascasarjana Universitas Udayana No. : 1570/UN14.4/HK/2016
Tanggal : 13 April 2016
Panitia Penguji Usulan Penelitian Disertasi adalah: Ketua : Dr. dr. I Wayan Putu SutirtaYasa, M.Si Anggota :
1. Prof. dr. Soetjiningsih, Sp.A(K)
2. Prof.drh. I Nyoman Mantik Astawa, Ph.D 3. Dr. dr. I Gusti Ayu Trisna Windiani, Sp.A(K) 4. Prof. Dr. dr. I Made Bakta, Sp.PD(KHOM) 5. Prof. Dr. dr. N Adiputra, M.OH
vi
SURAT PERNYATAAN BEBAS PLAGIARISME
Yang bertanda tangan di bawah ini, saya :
Nama : dr. I Gusti Ayu Putu Eka Pratiwi, M.Kes., Sp.A Program Studi Doktor : Ilmu Kedokteran UNUD
NIM : 1190271003
No. Telpon/HP : 08123920750
Email : [email protected]
Judul Disertasi : Pengaruh Virgin Coconut Oil dalam Formula WHO Dibandingkan dengan Minyak Jagung terhadap Glutathione, Malondialdehyde, TNF-α, dan Perlemakan Hati serta Berat Badan Tikus Wistar Malnutrisi Berat
Merupakan hasil karya original yang bisa dipertanggungjawabkan keasliannya dan tidak mengandung unsur plagiarisme. Pernyataan ini saya buat dengan sebenar - benarnya, apabila dikemudian hari ditemukan adanya pelanggaran, maka saya bersedia untuk mempertanggungjawabkan sesuai peraturan yang berlaku.
Denpasar, 24 Maret 2016 Yang membuat pernyataan,
vii
UCAPAN TERIMA KASIH
Puji syukur penulis panjatkan ke hadapan Ida Sang Hyang Widhi Wasa
Tuhan Yang Maha Esa, atas asung kertha wara nugraha-Nya, sehingga disertasi ini dapat diselesaikan pada waktunya.
Pada kesempatan ini, penulis mengucapkan terima kasih yang sebesar-besarnya kepada Prof. dr. Soetjiningsih, Sp.A(K), sebagai Promotor yang dengan penuh kesabaran dan perhatian telah memberikan dorongan, semangat, bimbingan dan saran selama penulis mengikuti program doktor, khususnya dalam penyelesaian disertasi ini. Begitu pula penghargaan dan rasa terima kasih yang sebesar-besarnya penulis sampaikan kepada Prof. drh. I Nyoman Mantik Astawa, Ph. D, sebagai Kopromotor I dan Dr. dr. I Gusti Ayu Trisna Windiani, Sp.A(K) sebagai Kopromotor II yang telah memberikan bimbingan dan saran dengan penuh kesabaran dan tiada henti-hentinya memberikan semangat kepada penulis dalam menyelesaikan disertasi ini.
viii
kepada penulis selama menempuh pendidikan di Pascasarjana Universitas Udayana.
Pada kesempatan yang baik ini, penulis juga menyampaikan rasa terimakasih kepada Prof. Dr. dr. I Putu Astawa, M.Kes, Sp.OT selaku Dekan Fakultas Kedokteran UNUD dan Dr. dr. Dewa Putu Gde Purwa Samatra, SpS (K) sebagai ketua Program Studi Pendidikan Dokter atas izin yang diberikan kepada penulis dalam mengikuti Program Doktor ini.
Penulis juga mengucapkan terimakasih yang sebesar-besarnya kepada: Prof. dr. Soetjiningsih, Sp.A(K), Prof.drh. I Nyoman Mantik Astawa, Ph.D, Dr. dr. I Gusti Ayu Trisna Windiani, Sp.A(K), Prof. Dr. dr. I Made Bakta, Sp.PD(KHOM), Prof. Dr. dr. N Adiputra, M.OH, Prof. Dr. Ir. Ida Bagus Putra Manuaba, M.Phill, Dr. dr. I Wayan Putu SutirtaYasa, M.Si dan Prof. Dr. dr. Boerhan Hidajat, Sp.A(K) sebagai penguji disertasi ini mulai dari tahap awal, atas semua masukan dan bimbingannya yang dengan penuh kesabaran dan perhatian telah memberikan dorongan semangat, saran, sanggahan, dan koreksi sehingga disertasi ini akhirnya dapat terwujud.
ix
Ayahanda dr, I Gusti Gede Djelantik, Sp.A(K) yang telah mengasuh dan membesarkan penulis, mengajarkan disiplin dan nilai-nilai luhur, serta terus menerus memberikan motivasi dan semangat yang tidak habis-habisnya sehingga penulis akhirnya bisa menyelesaikan pendidikan ini. Terimakasih juga penulis ucapkan kepada Bapak mertua Prof. Dr. dr I Gede Putu Surya, SpOG (K) dan Ibu mertua Ni Nyoman Murtini atas dorongan dan dukungannya kepada penulis dalam menempuh pendidikan ini. Terima kasih pula kepada adik dan adik ipar, dr. I Gusti Ayu Made Riantarini, Sp.S, I Gusti Ayu Nyoman Rani Dewi Yani, S.T., I Gusti Ketut Ari Wijaya Saputra, B.Sc., serta I Gusti Parisuda Mangun Sidanta, S.T., dr. I Putu Oka Darmawan, MARS, Natalia, S.E., dr. Ira Yunitasari S, S. Ked, I Gusti Bagus Siddhajapa HS, S.I.P., M.M., I Gede Teddy Prananda S, S.T., M.T. dan dr. Ni Nym Yunita KB, S. Ked., atas dukungan dan pengertiannya selama ini.
Akhirnya penulis menyampaikan terimakasih kepada suami tercinta Dr. dr. I Gede Ngurah Harry Wijaya Surya, Sp.OG yang telah berusaha mengerti dan bersabar mendampingi penulis selama ini, anak-anak tersayang Putu Ayu Adindhya Saraswati Surya, Made Ayu Nadine Indira Surya dan I Gede Nyoman Arvin Adhyasta Surya, yang telah memberikan kerelaan, pengorbanan dan pengertiannya sehingga memungkinkan penulis bisa lebih berkonsentrasi menyelesaikan naskah disertasi ini. Penulis juga mengucapkan terimakasih yang sebesar-besarnya kepada teman-teman di Program Studi S3 Ilmu Kedokteran Universitas Udayana, khususnya teman-teman angkatan 2011/2012, atas motivasi, semangat dan kebersamaannya.
Untuk pihak-pihak yang belum dapat penulis sebutkan satu persatu, penulis juga ucapkan terima kasih yang sebesar-besarnya.
Semoga Ida Sang Hyang Widhi Wasa/Tuhan Yang Maha Esa selalu melimpahkan karunia-Nya kepada semua pihak yang telah membantu pelaksanaan dan penyelesaian disertasi ini.
Denpasar, 26 April 2016 Penulis
x
ABSTRAK
PENGARUH VIRGIN COCONUT OIL DALAM FORMULA WHO DIBANDINGKAN DENGAN MINYAK JAGUNG TERHADAP
GLUTATHIONE, MALONDIALDEHYDE, TNF-α DAN PERLEMAKAN HATI SERTA BERAT BADAN TIKUS WISTAR MALNUTRISI BERAT
Malnutrisi berat (MB) masih merupakan penyebab kematian utama anak-anak di bawah usia lima tahun. Kadar antioksidan kurang, radikal bebas, perlemakan hati, dan sitokin proinflamasi meningkat pada MB, terjadi kondisi hiperkatabolik berkepanjangan. Berat badan (BB) sulit meningkat. Virgin coconut oil
mengandung medium chain triglyseride (MCT) dan antioksidan tinggi diharapkan dapat mengatasi masalah tersebut. Penelitian pemberian VCO dalam penanganan MB belum ada.
Penelitian ini bertujuan untuk mengetahui pengaruh VCO dalam penanganan MB, terhadap kandungan glutathione (GSH), malondialdehyde (MDA), tumor necrotizing factor-α (TNF-α), perlemakan hati, dan peningkatan BB. Penelitian ini merupakan penelitian true experimental, randomized posttest only control group design, membandingkan efek terapi VCO dengan minyak jagung dalam F75 dan F100 untuk penanganan tikus MB. Tikus MB dibagi 2 kelompok secara random, kelompok yang mendapat F75 dan F100 dengan kandungan VCO (Kelompok A) dan kandungan minyak jagung (Kelompok B). Pengamatan dilakukan setelah pemberian selama 28 hari.
xi ABSTRACT
INFLUENCE OF VIRGIN COCONUT OIL COMPARED TO CORN OIL IN WHO FORMULA ON GLUTATHIONE, MALONDIALDEHYDE, TNF-α,
FATTY LIVER AND BODY WEIGHT IN SEVERE MALNOURISHED WISTAR RAT
Severe malnutrition remain a major killer of children under five years old. On such condition, there is lack of antioxidants, increase free radical agents, fatty liver, and increase of proinflammatory cytokine. Those cause prolong hypercatabolic state that make difficulty in weight increament. Virgin coconut oil
(VCO) contain medium chain triglyceride (MCT) and antioxidants. It is expected to solve that problem compared with corn oil which is precursor of proinflammation cytokine and low level of antioxidants. Study of VCO use for severe malnutrition is still not exist.
The aim of this study is to understand the influence of VCO for severe malnutrition treatment in GSH level, MDA, TNF-α, fatty liver, and weight increament. This study is a true experimental with using randomized posttest only control group design, to compare treatment effect of using VCO vs corn oil in Formula 75 and Formula 100 in the treatment of severe malnourished rat. Severe malnourished rats were divided by 2 groups randomly which were group given F75 and F100 with VCO content (Group A) and group given F75 and F100 with corn oil content (Group B). Those groups were given treatment for 28 days. There were 19 rats for each group. One rat from goup B died before the end of this study. Analysis was done for the last 37 rats. GSH levels were not significantly different from both of two group 1.35 pg/g (SD (0.74) and 1.27 pg/g (SD 0.48) for Group A and B respectively (p = 0.703). Low expression of MDA (< 2.9%) was more prominence in Group A than Group B (p=0.046). TNF-α level for Group A (1087.2 pg/g (320.00-2525.46)) was higher than Group B (711.32 pg/g (403.19-2400.91)) but not statistically significant (p = 0.08). Fatty liver in Group A (13.74 cell in five view field (SD 1.32)) was lesser than Group B (20.74 cell in five view field (SD 2.01) and statistically significant (p = 0,000). Means of weight increament were not significantly different from both of two group, 73.45 g (SD 20.08) and 68.97 g (SD 11.49) for Group A and B respectively (p = 0.414).
Low expression of MDA was more prominence and fatty liver was lesser in VCO compare to corn oil group. There were no differencies in liver GSH, liver TNF-α and body weight increament for VCO and corn oil groups. Lesser fatty liver in VCO group could be because of MCT content in VCO.
xii
PENETAPAN PANITIA PENGUJI ... iv
xiii
4.9.2 Penyiapan tikus percobaan menjadi malnutrisi berat... 96
4.9.3 Prosedur pemeriksaan………..……….. 98
4.10 Alur Penelitian... 99
4.11 Prosedur Penelitian... 100
4.12 Analisis Data... 101
BAB V HASIL PENELITIAN ... 103
5.1 Karakteristik Sampel Penelitian ... 103
5.2 Efek Pemberian VCO dan Minyak Jagung pada Tikus Malnutrisi Berat terhadap Kadar GSH, Ekspresi MDA, Kadar TNF-α, Perlemakan Hati dan Peningkatan Berat Badan ……… 104
BAB VI PEMBAHASAN ... .. 109
6.1 Kadar Glutathione Hati pada Pemberian VCO Dibanding Minyak Jagung dalam Penanganan Tikus Malnutrisi Berat (MB)……... 109
6.2 Ekspresi Malondialdehyde Hati pada Pemberian VCO Dibanding Minyak Jagung dalam Penanganan Tikus Malnutrisi Berat……... 114
6.3 Kadar TNF-α Hati pada Pemberian VCO Dibanding Minyak Jagung dalam Penanganan Tikus Malnutrisi Berat ………..………. 116
6.4 Perlemakan Hati pada Pemberian VCO Dibanding Minyak Jagung dalam Penanganan Tikus Malnutrisi Berat………. 124
xiv
DAFTAR GAMBAR
2.1 Klasifikasi Mekanisme Pertahanan Seluler Antioksidan……….. 26
2.2 Mekanisme Aktivitas Scavenger……….. 29
2.3 Interrelasi di antara LMWA………..… 31
2.4 Skema Sistem Kekebalan Tubuh……….… 39
2.5 Efek Lokal dan Sistemik Sitokin dalam Inflamasi………... 44
3.1 Kerangka Berpikir……… 80
3.2 Kerangka Konsep Penelitian... 83
4.1 Bagan Randomized Posttest Only Control Group Design…...……… 85
4.2 Alur Penelitian………... 99
5.1 Gambaran pemeriksaan histologi jaringan hati tikus Kelompok A ... 107
5.2 Gambaran pemeriksaan histologi jaringan hati tikus Kelompok B ... 108
xv
DAFTAR TABEL
2.1 Kadar Aktivitas Antioksidan Total (µmol/L) pada Kontrol Marasmus dan Kwashiorkor saat Presentasi, Follow-up Pertama dan Kedua………… 37 2.2 Kadar Aktivitas Antioksidan Total (µmol/L) pada Marasmus dan Kwashiorkor dengan dan tanpa Suplementasi Antioksidan selama Follow-up…... 37 2.3 Tipe dan Frekuensi Pemberian Diet pada Malnutrisi Berat ……… 55 2.4 Komposisi F75 dan F100 ………. 57 2.5 Komposisi Modisco ½, I, II, dan III ……….. 58 2.6 Komposisi Asam Lemak VCO, Minyak Kelapa Biasa, dan Minyak Jagung 76 2.7 Persentase Asam Lemak Saturated (SFA), Monounsaturated (MUFA), Polyunsaturated (PUFA) dan Total Unsaturated (MUFA+PUFA) Minyak Kelapa dan Minyak Jagung ………. 76 2.8 Komposisi Asam Lemak Minyak Kedelai, Minyak Canola, Minyak
Cottonseed, Minyak Bunga Matahari, Minyak Kacang Tanah, Minyak Zaitun, Minyak Wijen, Minyak Safflower, dan Minyak Flaxseed……… 77 5.1 Karakteristik Sampel Penelitian ... 104 5.2 Efek Pemberian VCO dan Minyak Jagung pada Tikus Gizi Buruk terhadap Kadar GSH, Kadar TNF-α, Perlemakan Hati, Peningkatan
xvi
DAFTAR SINGKATAN
AA : Arachidonic acid
ABTS : 2,2’-azinobis-(3-ethylbenzothiazoline-6sulfonic acid
ALA : α-linolenic acid
APC : Antigen presenting cells
BB : Berat badan
CRP : C-reactive protein
CSAD : Center Study of Animal Diseases DPPH : α,α-diphenyl-β-picrylhydrazyl
EGCG : Epigallocatechingallate
ELISA : Enzyme linked immunosorbent assay EPA : Eicosapentaenoic acid
F75 : Formula 75
F100 : Formula 100
FKH : Fakultas Kedokteran Hewan
G-CSF : Granulocyte colony-stimulating factor
GM-CSF : Granulocyte-macrophage colony-stimulating factor
JAKs : Janus-activated kinases
JNK : Jun N-terminal kinase
LA : Linoleic acid
LDL : Low density lipoprotein
LLA : Lingkar lengan atas
LMWA : Low Molecular Weight Antioxidants
LTB4 : Leukotriene B4
MAP : Mitogen-activated protein
MB : Malnutrisi berat
MCP-1 : Monocyte chemotactic protein-1
MCT : Medium chain fatty acid
MDA : Malondialdehyde
MFR : Mannose-fucose receptor
MIP-2 : Macrophage inflammatory protein 2 NAC : N-acetyl cysteine
NICU : Neonatal Intensive Care Unit
xvii
STATs : Signal transducers and activators of transciption
xviii
DAFTAR LAMPIRAN
1. Keaslian Penelitian
2. APCC Standards for Virgin Coconut 3. Daftar Randomisasi
4. Cara Pembuatan Virgin Coconut Oil Metode Pendiaman 5. Prosedur Pemeriksaan Glutathione Hati
6. Pemeriksaan Imunohistokimia MDA 7. Prosedur Pemeriksaan TNF-α Hati Tikus 8. Cara Pemeriksaan Perlemakan Hati 9. Time Table Penelitian
10.Cara Bekerja dengan Hewan Coba (Tikus Putih) 11.Komposisi Bahan Makanan Tikus
12.Keterangan Kelaikan Etik 13.Aktivitas Antioksidan VCO
14.Aktivitas Antioksidan Minyak Jagung
BAB I
PENDAHULUAN
1.1Latar Belakang
Malnutrisi berat (MB) masih merupakan penyebab kematian utama anak-anak di bawah usia lima tahun. Sekitar 20 juta anak menderita MB di seluruh dunia. Sebagian besar penderita MB terdapat di Asia Selatan dan Afrika SubSahara. Angka kematian anak MB sebesar 5-20 kali lebih tinggi dibanding anak dengan status nutrisi baik. Malnutrisi berat merupakan penyebab langsung dan tidak langsung kematian pada anak. Penyebab tidak langsung adalah meningkatkan
case fatality rate anak-anak yang menderita penyakit yang umum diderita pada masa anak-anak seperti diare dan pneumonia. Perkiraan saat ini sekitar satu juta anak meninggal setiap tahun karena MB (World Health Organization, dkk., 2007). Data Riset Kesehatan Dasar (RISKESDAS) 2010 menyatakan bahwa prevalens status gizi balita nasional untuk gizi buruk, gizi kurang, gizi baik, dan gizi lebih masing-masing adalah 4,9%, 13%, 76,2%, dan 5,8%. Daerah Istimewa Yogyakarta dan Bali menunjukkan prevalens terendah gizi buruk yaitu 1,4% dan 1,7%. Provinsi Gorontalo dan NTB menduduki posisi tertinggi gizi buruk yaitu 11,2% dan 10,6% (Kementerian Kesehatan RI, 2010).
balita yang mengidap berbagai penyakit tersebut juga menderita malnutrisi (World Health Organization, 1999a). Infeksi dan malnutrisi merupakan suatu mata rantai yang saling berkaitan. Anak kurang gizi amat rentan terhadap infeksi dan infeksi menyebabkan anak kehilangan napsu makan, yang berakhir dengan kematian. Balita yang bertahan hidup dalam proses perkembangan selanjutnya banyak mengalami hambatan seperti keterbelakangan mental (World Health Organization, 2003).
Konsentrasi antioksidan (konsentrasi glutathione (GSH) dan vitamin E plasma) didapatkan berkurang pada kwashiorkor (Sauerwein dkk.,1997). Konsentrasi antioksidan yang rendah ini merupakan suatu akibat bukan sebagai penyebab terjadinya kwashiorkor (Ciliberto, A dkk., 2005). Pada tikus yang mengalami malnutrisi protein jika dibandingkan dengan tikus tanpa malnutrisi protein didapatkan adanya penurunan glutathione total hati sebesar 65% (Ling dkk., 2004). Kondisi ini serupa dengan respon selama fase inflamasi. Kadar
glutathione hati berhubungan terbalik dengan tingkat aktivasi NF-KB hati, menyebabkan peningkatan transkripsi IL-1β dan TNF-α (Sies, 1999). Deplesi
aktivasi sitokin, sehingga sulit membedakan antara perubahan primer dan sekunder dari glutathione (Ling dkk., 2004).
Produksi yang berlebihan dari reactive oxygen intermediates (superoxide anion (O2–), hydroxyl radical (OH•), singlet oxygen dan hydrogen peroxide (H2O2) dalam eritrosit terjadi pada kondisi malnutrisi yang menimbulkan terjadinya stres oksidatif. Malondialdehyde (MDA) merupakan suatu produk antara teroksidasi yang sering digunakan sebagai petanda yang dapat dipercaya terhadap lipid peroxidation pada malnutrisi. Malonndialdehyde (MDA) serum meningkat jumlahnya dan terjadi penurunan kadar zinc (Ghone, dkk., 2013, Jain, dkk., 2008), vitamin E serum, dan erythrocyte superoxide dismutase pada pasien MB. Setelah diberikan suplementasi antioksidan selama sebulan didapatkan kadar MDA menurun secara bermakna dan kadar zinc serta kapasitas erythrocyte superoxide dismutase meningkat secara bermakna. Kadar vitamin E mengalami peningkatan yang tidak bermakna jika dibandingkan dengan kadar sebelum disuplementasi. Defisiensi yang berat dari berbagai macam nutrisi pada MB menimbulkan generasi stres oksidatif berat. Efek ini dapat diminimalisasi dengan pemberian suplementasi antioksidan (Ghone, dkk., 2013).
pada MB adalah kemungkinan karena aktivasi kronik nonspesifik dari sistem kekebalan tubuh karena inflamasi kronik (Bosnak, dkk., 2010).
Pada kondisi malnutrisi terjadi disfungsi berbagai elemen imun yang lebih disebabkan disregulasi dibandingkan dengan imunodefisiensi, tetapi gangguan pada proses tersebut masih belum jelas (Hughes dan Kelly, 2006). Malnutrisi protein menginduksi suatu kondisi inflamasi derajat rendah pada tikus, yang dibuktikan dengan adanya aktivasi mitogen-activated protein (MAP) kinase pada hati, peningkatan kadar serum Tumor Necrosis Factor (TNF), interleukin-1 (IL-1), interleukin-6 (IL-6), dan serum α1-acid glycoprotein, disertai pengurangan kadar serum albumin. Jadi deplesi diet protein itu sendiri (tanpa adanya stimulus eksogen) merupakan salah satu faktor stimulus untuk terjadinya inflamasi sistemik (Ling, dkk., 2004).
berlangsung berkepanjangan maka akan menimbulkan hal-hal yang merugikan (Madeddu dan Mantovani, 2006).
Tumor necrosis factor (TNF)-α merupakan suatu sitokin yang bersifat pleiotropik. Sitokin ini menginduksi respon seluler yaitu proliferasi, produksi mediator inflamasi, dan kematian sel. Tumor necrosis factor (TNF)-α memegang peranan penting dalam patofisiologi dari syok septik dan wasting syndrome.
Tumor necrosis factor (TNF)-α pada jaringan hati terlibat dalam patofisiologi hepatitis virus, penyakit hati karena alkohol, nonalcoholic fatty liver disease, dan
ischemia-reperfusion injury. Tumor necrosis factor (TNF)-α memegang peranan dikotomi pada jaringan hati, sebagai mediator kematian sel dan juga menginduksi proliferasi hepatosit dan regenerasi hati (Schwabe dan Brenner, 2006).
Ltk mouse fibroblast, 70Z/3 mouse pre B cell). Efek antioksidan dalam translokasi NF-KB tampaknya berupa cell-type-dependent dan stimulus-dependent.
Infiltrasi lemak yang ekstensif merupakan salah satu gambaran kardinal dari MB. Akumulasi lemak terjadi melalui salah satu atau kombinasi dari tiga mekanisme yaitu peningkatan sintesis lemak hati, gangguan pada transportasi pengeluaran lemak hati, atau penurunan pemecahan lemak dalam hati. Mekanisme peningkatan sintesis lemak hati tidak dapat dikonfirmasi dengan didapatkannya penurunan dari aktivitas glucose-6-phosphatase. Mekanisme tidak adekuatnya sintesis lipoprotein dan sekresinya secara teori tampaknya merupakan penjelasan yang tepat. Konsentrasi lipoprotein sirkulasi didapatkan rendah pada anak dengan malnutrisi berat dan sintesis protein mengalami gangguan. Lemak hati tidak berkorelasi baik dengan konsentrasi sirkulasi lipoprotein.
Mekanisme yang ke tiga yaitu penurunan pemecahan lemak hati. Pada penelitian Leung dan Peter (1986) melihat biopsi hati yang diambil dari pasien perlemakan hati karena alkohol dan mendapatkan adanya gangguan -oksidation, menyimpulkan bahwa penurunan -oksidation ini penting dalam patogenesis perlemakan hati. Beta-oxidation lemak terutama terjadi dalam mitokondria tetapi fungsi mitokondria pada anak malnutrisi berat masih baik (Waterlow, 1961),
sehingga hipotesis gangguan β-oxidation sebagai penyebab perlemakan hati
Aktivitas sistem peroxisomal -oxidation dipengaruhi oleh banyak faktor eksternal. Peroxisome memiliki waktu paruh yang pendek dan ini dapat berkurang lagi dengan adanya stres tertentu. Glutathione (GSH) serum dan hati didapatkan menurun pada kondisi MB. Glutathione berhubungan terbalik dengan perlemakan hati pada MB. Pada kondisi malnutrisi, status karnitin dan riboflavin marginal, sehingga peranan peroxisomal -oxidation secara relatif akan meningkat.
Hubungan antara infeksi dan malnutrisi telah diketahui dengan baik, infeksi bakteri juga diketahui mengurangi jumlah peroxisome. Reaksi awal sistem peroxisomal akan menghasilkan hidrogen peroksida yang bersifat radikal sementara pada kondisi MB kadar antioksidan berkurang. Radikal bebas tersebut akan merusak peroxisome itu sendiri, sehingga akan terjadi bunuh diri metabolik dari peroxisome. Hal ini dapat dilihat dari hasil biopsi anak-anak MB yang meninggal jika dibandingkan dengan anak-anak MB yang dalam pemulihan nutrisi ditemukan adanya pengurangan jumlah peroxisome pada anak-anak yang meninggal tersebut. Radikal bebas diperkirakan memegang peranan dalam patogenesis gambaran klinis malnutrisi anak, dan defek dari -oxidation
kemungkinan berkontribusi terhadap perlemakan hati, jadi kemungkinan disfungsi
peroxisome menghubungkan ke dua hal tesebut (Doherty, dkk., 1991).
kandungan protein, lemak dan natrium yang cukup sesuai dengan usianya, tetapi harus lebih rendah disertai dengan kandungan karbohidrat yang tinggi. World Health Organization mengajukan formula 75 (F75) dan formula 100 (F100) untuk tahap awal dan rehabilitasi. Formula 75 diberikan untuk tahap awal, dan setelah napsu makan mulai pulih diberikan F100. Salah satu komposisi F75 dan F100 adalah minyak sayur (Kementerian Kesehatan Republik Indonesia, 2013). Minyak sayur yang sering digunakan adalah minyak jagung. Penyediaannya pada beberapa daerah di Indonesia terkadang susah. Alternatif minyak lain diperlukan untuk menggantikan minyak jagung. Kelapa banyak terdapat hampir di seluruh daerah Indonesia. Virgin coconut oil (VCO) mudah untuk dibuat sendiri dari bahan kelapa. Penggunaan VCO dalam F75 dan F100 perlu dipertimbangkan untuk menggantikan minyak jagung.
Minyak jagung terdiri dari 99% triacyglycerols dengan polyunsaturated fatty acid (PUFA) 59%, monounsaturated fatty acid 24%, dan saturated fatty acid
(SFA) 13% (Dupont, dkk., 1990). Polyunsaturated fatty acid (PUFA) minyak jagung terdiri dari 98% omega-6 linoleic acid (C18:2 n-6 c,c) dan 2% omega-3 alpha-linolenic acid (C18:3 n-3 c,c,c). Monounsaturated fatty acid minyak jagung mengandung 99% oleic acid (C18:1c). Saturated fatty acid minyak jagung terdiri dari 80% palmitic acid (C16:0), 14% stearic acid (C18:0), dan 3% arachidic acid
(C20:0) (U.S. Department of Agriculture, Agricultural Research Service, 2007). Berdasarkan strukturnya, lemak jenuh dibagi menjadi lemak rantai pendek, menengah dan panjang, sementara monounsaturated fatty acid dan
2008). Minyak jagung tidak mengandung asam lemak rantai pendek dan menengah. Minyak jagung mengandung antioksidan sebanyak 0,33-0,34 mmol/100 g minyak jagung (Halvorsen, dkk., 2006).
Mediator inflamasi antara lain adalah n-6 eicosanoid, prostaglandin E2 (PGE2), dan leukotriene B4 (LTB4) yang terbentuk dari n-6 PUFA arachidonic acid (AA; 20:4n-6). Lemak n-6 PUFA ini banyak terdapat pada diet dengan
linoleic acid (LA; 18:2n-6) yaitu pada minyak kedele, jagung, safflower dan bunga matahari. n-3 homolog LA adalah -linolenic acid (ALA; 18:3n-3) yang terdapat dalam sayuran hijau berdaun, minyak flaxseed dan canola. Saat dicerna,
18-carbon fatty acid tersebut akan mengalami desaturasi dan perpanjangan menjadi 20-carbon n-6 fatty acids. Linoleic acid menjadi AA, dan ALA menjadi
eicosapentaenoic acid (EPA; 20:5n-3). n-6 PUFA arachidonic acid merupakan progenitor dari PGE2 dan LTB4 lewat jalur enzim cyclooxygenase dan 5-lipoxygenase. Eicosapentaenoic acid dapat menginhibisi metabolisme AA secara kompetitif melalui jalur enzimatik, sehingga dapat menekan produksi dari mediator inflamasi n-6 eicosanoid. Eicosapentaenoic acid juga dapat menekan produksi IL-1 dan TNF- dengan mekanisme yang belum jelas. Semakin tinggi kandungan EPA dalam bahan makanan maka kandungan AA akan semakin berkurang (James, dkk., 2000). Minyak jagung dengan kadar AA tinggi maka akan menurunkan kadar EPA menyebabkan efek proinflamasi yang akan meningkat.
selama 8 minggu minyak jagung menimbulkan peningkatan produksi 1 dan IL-6. Pada pemberian minyak kelapa selama 8 minggu menekan produksi IL-1 (Tappia dan Grimble, 1994).
Intake linoleic acid juga dapat meningkatkan katalase, glutathione peroxidase
dan superoxide dismutase pada hepar. Aktivitas peroxisomal catalase juga meningkat. Hal ini menunjukkan bahwa mekanisme eliminasi peroxide diaktivasi oleh intake linoleic acid. Saat tikus diberi minyak jagung dalam makanannya, maka linoleic acid menimbulkan peroksidasi lipid dan mengaktivasi mekanisme pertahanan terhadap peroksidasi lipid (stres oksidatif) dengan meningkatkan aktivitas glutathione peroxidase (Iritani dan Ikeda, 1982).
Virgin coconut oil adalah minyak yang didapatkan dari kelapa tua segar dengan menggunakan alat atau secara alami, tanpa pemanasan, tanpa menggunakan pemurnian, tanpa pemutihan, dan tanpa pemberian aroma secara kimia, sehingga didapatkan suatu minyak yang tidak mengalami perubahan seperti minyak alami apa adanya. Minyak ini banyak mengandung asam lemak rantai sedang (medium-chain fatty acid /MCFAs) sekitar 64%, dengan asam lemak laurat (C12) terbanyak kandungannya (47-53%) (Bawalan dan Chapman, 2006). Kandungan linoleic acid VCO rendah yaitu berkisar antara 0,90-1,72% (Marina, dkk., 2009a). Sementara kandungan linolenic acid juga rendah yaitu berkisar dari tidak terdeteksi sampai 0,2% (Bawalan dan Chapman, 2006).
tidak saponifiable dibuktikan dengan peningkatan kadar enzim antioksidan dan pencegahan peroksidasi lipid secara in vivo dan in vitro (Nevin dan Rajamohan, 2006). Virgin coconut oil mengandung aktivitas antioksidan antara 52-80% jika dibandingkan dengan kontrol tokoferol dan BHA. Aktivitas antioksidan ini berkorelasi dengan total kandungan fenolik (Marina, dkk., 2009a). Pada penelitian mengenai pengaruh senyawa fenolik dan sumber makanan terhadap produksi sitokin dan antioksidan oleh sel A549, didapatkan bukti bahwa senyawa fenolik secara bermakna mengubah produksi sitokin dan antioksidan, terjadi inhibisi produksi IL-6 dan interleukin-8 (IL-8) (Gaulliard, dkk., 2008).
detoksifikasi fase II. Keterlibatan jalur Nrf2 terdapat dalam phenolic acid-mediated antioxidant enzyme gene induction (Yeh dan Yen, 2006).
Beberapa penelitian dilakukan untuk mengetahui manfaat pemberian MCT. Perubahan mukosa dan pertumbuhan bakteri yang berlebihan pada usus halus bagian atas terjadi pada MB (Peny, 2003). Medium chain triglyceride (MCT) menginhibisi formasi radikal bebas hati dan produksi TNF-α sel Kuffer yang diaktivasi endotoksin. Medium chain triglyceride (MCT) mengubah struktur usus halus dan memengaruhi permeabilitas atau mikroflora saluran cerna. Aktivasi sel Kupffer mengeluarkan mediator (sitokin, eicosanoids, dan radikal bebas). Medium chain trigyceride (MCT) menghilangkan peningkatan Ca2+ intraseluler yang disebabkan lipopolisakarida. Aktivasi sel Kupffer memerlukan Ca2+, yang mengandung voltage-dependent Ca2+ channel (Kono, dkk., 2000).
digantikan dengan lemak saturated (18:82, lemak sapi: minyak MCT) pada konsentrasi 20% sampai 65% selama 21 hari. Peningkatan konsentrasi minyak jagung meningkatkan steatosis hati dan serum alanine amino transferases (p<0,05). Penggantian lemak unsaturated seperti minyak jagung dengan MCT pada diet dapat digunakan sebagai terapi yang potensial dalam penanganan NAFLD (Ronis, dkk., 2013).
Pada kondisi MB didapatkan kondisi kadar antioksidan berkurang, peningkatan radikal bebas, perlemakan hati, dan peningkatan sitokin proinflamasi sehingga menimbulkan kondisi hiperkatabolik yang berkepanjangan. Kondisi ini menimbulkan sulitnya peningkatan berat badan. Virgin coconut oil dengan kandungan MCT dan antioksidan yang tinggi diharapkan dapat mengatasi hal tersebut jika dibandingkan dengan minyak jagung dengan kandungan prekursor proinflamasi dan kadar antioksidan yang rendah. Penelitian mengenai VCO yang digunakan dalam penanganan MB belum ada (Lampiran 1). Peneliti ingin mengetahui bagaimana pengaruh pemberian VCO dalam penanganan MB. Peneliti membandingkan pemberian VCO dengan minyak jagung dalam komposisi F75 dan F100 untuk memperbaiki sistem imun tikus MB sehingga mempercepat kenaikan berat badan.
1.2Rumusan Masalah
Rumusan masalah penelitian ini adalah sebagai berikut:
2. Apakah ekspresi MDA hati yang rendah lebih banyak pada pemberian VCO dibanding minyak jagung dalam penanganan tikus MB?
3. Apakah kadar TNF-α hati lebih rendah pada pemberian VCO dibanding minyak jagung dalam penanganan tikus MB?
4. Apakah perlemakan hati lebih sedikit pada pemberian VCO dibanding minyak jagung dalam penanganan tikus MB?
5. Apakah peningkatan berat badan pada tikus yang diberikan VCO lebih tinggi dibanding minyak jagung dalam penanganan tikus MB?
1.3Tujuan Penelitian
Tujuan umum: untuk mengetahui pengaruh antioksidan VCO dalam memodulasi sitokin proinflamasi pada tikus malnutrisi berat guna meningkatkan kecepatan kenaikan berat badan.
Tujuan khusus:
1. Membuktikan bahwa kadar glutathione hati lebih tinggi pada pemberian VCO dibanding minyak jagung dalam penanganan tikus MB.
2. Membuktikan bahwa ekspresi MDA hati yang rendah lebih banyak pada pemberian VCO dibanding minyak jagung dalam penanganan tikus MB. 3. Membuktikan bahwa kadar TNF-α hati lebih rendah pada pemberian VCO
dibanding minyak jagung dalam penanganan tikus MB.
5. Membuktikan bahwa peningkatan berat badan pada tikus yang diberikan VCO lebih tinggi dibanding minyak jagung dalam penanganan tikus MB.
1.4 Manfaat Penelitian
1.4.1 Manfaat akademik
Manfaat akademik dari penelitian ini adalah:
1. Pemahaman terhadap patofisiologi dan pengaruh nutrisi yang diberikan dalam penanganan MB dapat ditingkatkan dengan mengetahui mekanisme imunologis yang berpengaruh terhadap peningkatan kecepatan peningkatan berat badan pada MB.
2. Landasan bagi penelitian selanjutnya dalam bidang nutrisi dengan mengetahui mekanisme VCO dapat memperbaiki metabolisme tubuh yang terganggu pada MB dan peranannya dalam memperbaiki sistem kekebalan tubuh.
1.4.2 Manfaat praktis
2.1 Malnutrisi Berat
Istilah malnutrisi meliputi undernutrition dan obesitas. Pada penelitian ini malnutrisi yang dimaksudkan adalah malnutrisi undernutrition. Malnutrisi (undernutrition) didefinisikan sebagai suatu kondisi ketidakseimbangan antara kebutuhan dan masukan nutrien, yang menghasilkan secara kumulatif adanya defisit energi, protein, atau mikronutrien yang secara negatif memengaruhi pertumbuhan, perkembangan, dan akibat lainnya yang berhubungan. Malnutrisi berdasarkan etiologinya disebabkan oleh penyakit (1 atau lebih penyakit atau cedera yang secara langsung mengakibatkan ketidakseimbangan nutrien) atau akibat dari lingkungan/faktor behavioral yang berhubungan dengan penurunan masukan nutrien atau akibat dari ke dua hal tersebut (Mehta, dkk., 2013).
Malnutrisi berat (MB) atau disebut juga dengan gizi buruk merupakan suatu keadaan dimana seorang anak tampak sangat kurus yang ditandai dengan berat badan/panjang badan (BB/PB) < -3 SD dari median WHO child growth standard
2006, atau didapatkan edema nutrisional, dan pada anak usia 5-59 bulan Lingkar Lengan Atas (LLA) < 110 mm. World Health Organization dan United Nations
Children’s Fund (Unicef) menggunakan cut-off BB/PB < -3 SD median baku
Anak ini (WHO, 2009). Marasmus dan kwashiorkor adalah hasil akhir dari tingkat keparahan penderita gizi buruk. Marasmus ditandai dengan tubuh yang sangat kurus dengan berbagai tanda ikutannya. Kwashiorkor ditandai dengan edema, diawali edema pada punggung kaki yang dapat menyebar ke seluruh tubuh (Susanto, dkk., 2011). Manifestasi klinis malnutrisi berhubungan dengan tipe, keparahan, dan lamanya gangguan nutrisi tersebut terjadi, sehingga dapat terjadi manifestasi subklinis, reversibel, atau menetap tergantung pada ketersediaan terapi, penyakit lain atau penyakit komplikasi, dan derajat kerusakannya (Cunningham-Rundles dan McNeeley, 2003).
adalah terpaparnya bayi dan balita terhadap penyakit infeksi yang secara langsung merupakan penyebab kematian balita seperti diare, infeksi saluran pernapasan akut, campak, malaria, HIV/AIDS, dan lain-lain. Secara global lebih dari 50% balita yang mengidap berbagai penyakit tersebut juga menderita malnutrisi (WHO, 1999a). Infeksi dan malnutrisi merupakan suatu mata rantai yang saling berkaitan, dimana anak kurang gizi amat rentan terhadap infeksi dan infeksi menyebabkan anak kehilangan napsu makan, yang berakhir dengan kematian. Anak-anak yang bertahan hidup dalam proses perkembangan selanjutnya banyak mengalami hambatan seperti keterbelakangan mental (WHO, 2003).
Pada kondisi malnutrisi terjadi suatu proses adaptasi tubuh terhadap kondisi
transferin dan apolipoprotein B (Peny, 2003).
Infeksi menyebabkan terjadinya perubahan metabolisme pula sehingga produksi protein lebih diutamakan ke arah pembentukan protein fase akut. Produksi dari protein fase akut ini dan konsekuensi metabolik dari infeksi dimediasi oleh protein sitokin dan lipid derived factor. Produksi sitokin inflamasi yang menimbulkan suatu peradangan, juga akan menyebabkan mekanisme balik sehingga tidak terjadi proses inflamasi yang berlebihan. Pada malnutrisi, kondisi ini mengalami gangguan. Pada malnutrisi terjadi suatu kondisi pengurangan respon imun dan respon demam (Peny, 2003).
2.2Stres Oksidatif
Reactive oxygen species (ROS) dan radikal lainnya terlibat dalam berbagai peristiwa biologi (mutasi, karsinogenesis, proses degenerasi, inflamasi, penuaan dan perkembangan). Reactive oxygen species dapat berperan menguntungkan dan merugikan (Kohen dan Nyska, 2002).
Senyawa yang dapat menerima elektron disebut oksidan atau bahan yang mengoksidasi. Bahan yang memberikan elektron disebut reduktan atau bahan yang mereduksi (Prior dan Cao, 1999). Reaksi kimia dimana suatu bahan mendapatkan elektron disebut reduksi. Oksidasi adalah suatu proses dimana suatu bahan mengalami kehilangan elektron. Jika reduktan mendonasikan elektronnya, maka menyebabkan bahan lain mengalami reduksi, dan jika oksidan menerima elektron, maka menyebabkan bahan lain mengalami oksidasi. Suatu bahan yang mereduksi bertindak sebagai donasi elektron, biasanya dengan mendonorkan hidrogen atau melepas oksigen. Suatu proses oksidasi selalu ditemani oleh proses reduksi. Pada proses reduksi biasanya terjadi kehilangan oksigen, sementara pada proses oksidasi akan mendapatkan oksigen. Reaksi ini disebut reaksi redox. Reduktan dan oksidan merupakan istilah kimia, pada lingkungan biologi disebut dengan istilah antioksidan dan prooksidan (Kohen dan Nyska, 2002).
sekurang-karena afinitasnya mendonasikan atau mendapatkan elektron lain untuk mendapatkan stabilitas.
Molekul oksigen sendiri jika berdasarkan definisi tersebut juga dapat dikatakan radikal, karena mengandung dua elektron yang tidak berpasangan dalam dua orbit yang berbeda, sehingga dikatakan biradikal. Radikal oksigen tidak reaktif, meskipun demikian karena restriksi putaran yang tidak memungkinkan terjadinya donasi atau menerima elektron sebelum dilakukan pengaturan kembali arah putaran sekitar atom.
Kelompok senyawa nonradikal terdiri dari berbagai macam jenis bahan, dimana beberapa sangat reaktif meskipun tidak radikal perdefinisi. Senyawa-senyawa yang diproduksi dalam konsentrasi tinggi pada sel hidup adalah
hypochlorous acid (HClO), hydrogen peroxide (H2O2), organic peroxides,
Keseimbangan antara prooksidan dan antioksidan sangat ketat dan penting untuk mempertahankan fungsi sel dan biokimia yang vital (Hrbac dan Kohen, 2000). Keseimbangan ini sering disebut sebagai potensial redox. Potensial redox ini spesifik untuk setiap organela dan tempat biologis, dan setiap gangguan pada keseimbangan ini akan menimbulkan kerusakan sel dan organisme. Perubahan keseimbangan ke arah peningkatan prooksidan di atas kapasitas antioksidan disebut stres oksidatif dan dapat menimbulkan kerusakan oksidatif. Perubahan keseimbangan ke arah peningkatan kekuatan yang mereduksi, atau antioksidan, juga akan menimbulkan kerusakan dan disebut sebagai stres reduktif (Kohen dan Nyska, 2002).
besar terhadap waktu paruh ROS.
Toksisitas tidak berkorelasi dengan reaktifitas. Semakin panjang waktu paruh suatu spesies menunjukkan semakin toksik senyawa tersebut karena memiliki waktu yang cukup untuk berdifusi dan mencapai lokasi yang sensitif sehingga dapat berinteraksi dan menimbulkan kerusakan dalam perjalanannya dari tempat produksinya. Radikal superoksid yang memiliki waktu paruh yang relatif panjang memiliki waktu untuk berpindah lokasi dimana radikal tersebut dapat berinteraksi dengan molekul lainnya. Radikal ini dapat diproduksi dari membran mitokondria, berdifusi ke arah genome mitokondria, dan mengurangi transisi ikatan logam ke
metabolit oksigen. Salah satu enzim tersebut adalah xanthine oxidase. Sistem perbaikan yang efisien merupakan salah satu metode yang paling penting pada organisme untuk menanggulangi kerusakan oksidatif, terdiri dari enzim dan molekul-molekul kecil yang secara efisien memperbaiki tempat kerusakan oksidatif pada makromolekul. Sistem perbaikan DNA dapat mengidentifikasi
DNA-oxidized adduct (8-hydroxy-2-deoxyguanosine), thiamine glycol, dan tempat
Gambar 2.1 Klasifikasi mekanisme pertahanan seluler antioksidan (Kohen dan Nyska, 2002)
Sistem ini terdiri dari dua kelompok utama yaitu enzim antioksidan dan antioksidan berat molekul ringan (LMWA) (Gambar 2.1). Kelompok enzim terdiri dari direct-acting proteins, misalnya SOD. Protein dari keluarga ini berbeda dalam struktur dan kofaktornya. Cu-Zn SOD merupakan enzim dengan berat molekul sekitar 32.000, terdiri dari dua subunit, dimana salah satunya memiliki
Nyska, 2002).
Tipe SOD lainnya yang ada antara lain extracellular SOD (EC-SOD) dan Fe-SOD pada tanaman. Enzim ini memiliki struktur, berat molekul dan konstanta laju reaksi yang berbeda. Aktivitas enzim itu sendiri mampu untuk meningkatkan
dismutase spontan radikal superoksida terhadap H2O2. Perubahan yang bermakna terhadap konstanta laju reaksi dari berbagai macam SOD tergantung pada pH dan tempat aktivitasnya. Produk akhir reaksi dismutase yaitu H2O2, dapat dipindahkan dengan aktivitas enzim katalase dan anggota keluarga peroxidase termasuk
glutathione peroxidase (Chance, dkk., 1979).
membedakannya antara reaksi oleh peroxidase dan katalase.
Enzim-enzim lainnya dalam lingkungan sel mendukung aktivitas antioksidan.
Glucose-6-phosphate dehydrogenase menyediakan bahan yang mereduksi (NADPH) yang diperlukan untuk fungsi sel dan penting untuk regenerasi oxidized antioxidants. Regenerasi oxidized glutathione (GSSG) dalam bentuk tereduksi (GSH) dilakukan oleh reduced nicotinamide dinucleotide (NADH) (Chance, dkk., 1979). Beberapa enzim pendukung dapat mengeliminasi oksidan seperti misalnya xanthine dehydrogenase yang menghasilkan uric acid (antioksidan endogen yang efektif) (Gul, dkk., 2000).
Kelompok low-molecular-weight antioxidant (LMWA) terdiri dari sejumlah senyawa yang dapat mencegah kerusakan oksidatif secara langsung dan tidak langsung terhadap ROS (Kohen dan Gati, 2000). Mekanisme tidak langsung melibatkan chelation of transition metals yang mencegahnya untuk berpartisipasi dalam metal-mediated Haber-Weiss reaction (Beckman dan Koppenol, 1996). Molekul dengan aktivitas langsung memberikan turunan kimia serupa sehingga dapat memberikan elektronnya kepada radikal oksigen, sehingga dapat mengatasi radikal tersebut dan mencegahnya menyerang target biologinya. Scavengers
memiliki banyak manfaat di atas Kelompok Antioksidan enzimatik, karena
berkompetisi dengan target biologi pada spesies yang merusak tersebut (Kohen dan Gati, 2000). Aksi LMWA adalah sinergis dan interrelasi di antara LMWA adalah penting untuk perkembangan pedoman terapi antioksidan. Scavengers
yang berasal dari sumber endogen seperti proses biosintesis dan generasi produk sampah dari sel dan sumber eksogen dari diet. Sejumlah LMWA yang disintesis oleh sel hidup atau yang berasal dari produk sampah adalah sedikit saja (misalnya
histidine dipeptides, glutathione, uric acid, lipoic acid, dan bilirubin). Kebanyakan LMWA berasal dari sumber diet (Kohen dan Nyska, 2002).
Karakteristik scavenger adalah aktivitasnya yang bereaksi secara langsung dengan radikal dan menghilangkannya dengan memberikan elektron kepada spesies reaktif tersebut. Reaksi ini menghasilkan konversi dari scavenger itu sendiri menjadi radikal, meskipun tidak reaktif (Gambar 2.2).
Gambar 2.2 Mekanisme aktivitas scavenger (Kohen dan Nyska, 2002)
Oksidasi lebih lanjut dari scavenger
kembali menjadi bentuk tereduksinya (ascorbic acid) dengan bantuan glutathione.
Ascorbic acid sendiri akan menjadi radikal, yang dapat menerima elektron dari donor lainnya, misalnya NADH. Proses regenerasi dapat merupakan murni kimiawi atau suatu enzim dapat terlibat dalam transfer elektron (Gambar 2.3). Aktivitas kooperatif ini menjelaskan sinergisitas yang didapatkan saat beberapa
scavenger terlibat dan dapat dipergunakan untuk mendapatkan manfaat dari kombinasi LMWA dalam terapi antioksidan.
Glutathione merupakan suatu massa dengan berat molekul rendah.
Glutathione merupakan suatu tripeptide yang mengandung thiol, dengan bentuk tereduksinya glutamic acid-cysteine-glycine (GSH) dan bentuk teroksidasinya berupa GSSG (dimana 2 molekul GSH bergabung melalui oksidasi grup SH dari residu cysteine untuk membentuk jembatan disulphide (Kohen dan Nyska, 2002). Glutathione terdapat pada manusia, hewan, tumbuh-tumbuhan, dan bakteri aerob dengan konsentrasi tinggi mencapai milimolar. Glutathione berlaku sebagai suatu kofaktor dari enzim peroksidase, jadi sebagai antioksidan tidak langsung mendonasikan elektron yang diperlukan untuk mendekomposisikan H2O2. Senyawa ini juga terlibat dalam berbagai jalur biokimia dan fungsi sel lainnya (Barhoumi, dkk., 1993).
Glutathione mencegah oksidasi kelompok protein -SH dan untuk transportasi copper (Gul, dkk., 2000). Glutathione dapat berlaku chelating agent untuk ion
secara langsung. Glutathione berinteraksi dengan radikal OH., ROO., dan RO.
seperti juga HCLO dan „O2 saat bereaksi dengan ROS, yang menghasilkan radikal
glutathione, sehingga dapat meregerasikannya menjadi bentuk tereduksi dari
glutathione (Gul, dkk., 2000).
2.3Stres Oksidatif pada Malnutrisi Berat (MB)
Produksi berlebihan dari reactive oxygen intermediates (superoxide anion
(O2–), hydroxyl radical (OH•), singlet oxygen dan hydrogen peroxide (H2O2) dalam eritrosit terjadi pada kondisi malnutrisi yang menimbulkan terjadinya stres oksidatif (Ghone, dkk., 2013). Malondialdehyde (MDA) merupakan suatu produk antara teroksidasi yang sering digunakan sebagai petanda yang dapat dipercaya terhadap lipid peroxidation pada malnutrisi. Malonndialdehyde (MDA) serum meningkat jumlahnya dan terjadi penurunan kadar vitamin E serum, zinc dan
erythrocyte superoxide dismutase pada pasien MB. Setelah diberikan suplementasi antioksidan selama sebulan didapatkan kadar MDA menurun secara bermakna dan kadar zinc serta kapasitas erythrocyte superoxide dismutase
menyebabkan terjadinya peningkatan stres oksidatif pada MB. Faktor terpenting adalah asupan yang kurang dari nutrien misalnya karbohidrat, protein, vitamin, sehingga menimbulkan akumulasi ROS. Pada malnutrisi didapatkan kekurangan konsentrasi antioksidan enzimatik dan nonenzimatik bersamaan dengan trace elemen. Mekanisme yang ke dua terhadap terjadinya peningkatan stres oksidatif pada MB adalah kemungkinan karena aktivasi kronik nonspesifik dari sistem kekebalan tubuh karena inflamasi kronik. Peningkatan aktivasi MDA pada anak MB kemungkinan karena deplesi sebagian besar enzim antioksidan sebagai mekanisme kompensasi untuk melindungi membran sel dari efek merugikan radikal bebas (Bosnak, dkk., 2010).
Pada anak-anak yang menderita MB (dengan edema tetapi tidak yang tanpa edema) terjadi pengurangan konsentrasi GSH dalam plasma dan darah jika dibandingkan dengan anak-anak dengan status nutrisi yang baik (Becker, dkk., 1995). Pada penelitian lainnya menunjukkan hasil yang berbeda yaitu didapatkan bahwa pada ke dua tipe malnutrisi baik marasmus maupun kwashiorkor terjadi pengurangan aktivitas antioksidan. Pada anak dengan marasmus didapatkan adanya pengurangan red cell glutathione dan peningkatan lipid peroxidation
peningkatan beberapa petanda oxidant-induced lipid peroxidation seperti misalnya malondialdehyde, hexanal (Lenhartz, dkk., 1998), dan lipid hydroperoxide (Reid, dkk., 2000). Kerusakan pada pertahanan antioksidan pada kwashiorkor dan marasmus kwashiorkor menimbulkan terjadinya kerusakan radikal bebas pada membran sel dan kerusakan ini akan memegang peranan penting dalam patogenesis penyakit (Jackson, 1986). Glutathione selular merupakan senyawa antioksidan utama yang bereaksi secara langsung dalam menghilangkan ROS dan sebagai substrat dari beberapa peroksidase. Penelitian lain menunjukkan bahwa pada malnutrisi protein menginduksi terjadinya pengurangan pertahanan antioksidan tetapi tidak menyebabkan peningkatan radikal bebas. Pada kondisi malnutrisi protein terjadi penurunan metabolisme mitokondria di otak dan hati, yang kemungkinan menghasilkan penurunan produksi ROS (Wu, dkk., 2001).
Penelitian mengenai suplementasi diet awal dengan menggunakan cysteine
pada anak dengan malnutrisi berat dengan edema didapatkan konsentrasi dan laju sintesis absolut GSH meningkat pada kelompok yang mendapatkan suplementasi
penyediaan bahan atau defek pada jalur sintesis GSH atau karena ke duanya (Badaloo, dkk., 2002). Penelitian sebelumnya pada tikus menunjukkan bahwa baik aktivitas maupun jumlah -glutamylcysteine synthetase hati, rate limiting enzyme for GSH synthesis adalah tidak menurun dengan konsumsi diet rendah protein. Fakta ini menunjukkan bahwa jalur sintesis GSH tidak mengalami kerusakan pada malnutrisi protein (Hunter dan Grible, 1997). Jadi berdasarkan hal tersebut keterlambatan restrorasi kadar GSH sel pada anak dengan malnutrisi berat dengan edema adalah akibat dari kekurangan cysteine (Badaloo, dkk., 2002). TAOS terdiri dari kapasitas antioksidan yang berasal dari protein total (85%, terutama albumin, juga transferrin dan ceruloplasmin), uric acid (12%), bilirubin (4%), carotinoids (3%), tocopherols (1%), dan ascorbic acid (1%). TAOS pada anak-anak yang mengalami kwashiorkor menurun sampai kurang dari 50% dibandingkan dengan kontrol anak-anak dengan status nutrisi normal. Penurunan TAOS terjadi pada hari ke 1, 4, 8, 14, dan 20 (keseluruhan periode observasi). Taos terendah terjadi pada hari ke-4, kemudian mengalami peningkatan sampai hari ke-14, dan kemudian menurun kembali pada hari ke-20. Konsentrasi
yang hati-hati terhadap berkurangnya status antioksidan pasien (Fechner, dkk., 2001).
Kadar zinc serum pada anak MB lebih rendah secara bermakna jika dibandingkan dengan anak nutrisi baik. Kadar zink ini juga lebih rendah secara bermakna pada anak-anak MB dengan lesi kulit jika dibandingkan dengan tanpa lesi kulit. Kapasitas antioksidan total juga didapatkan lebih rendah pada anak-anak MB. Konsentrasi MDA pada anak-anak MB didapatkan lebih tinggi dibandingkan dengan kontrol. Kapasitas antioksidan total dan hipoalbuminemia juga berkorelasi positif dengan kadar zinc serum yang rendah. Defisiensi trace elemen serum menimbulkan penurunan proteksi antioksidan kemungkinan merupakan suatu faktor yang berkontribusi dalam patofisologi malnutrisi energi protein dan penggantian elemen ini dalam penatalaksanaan kondisi MB adalah penting (Jain, dkk., 2008).
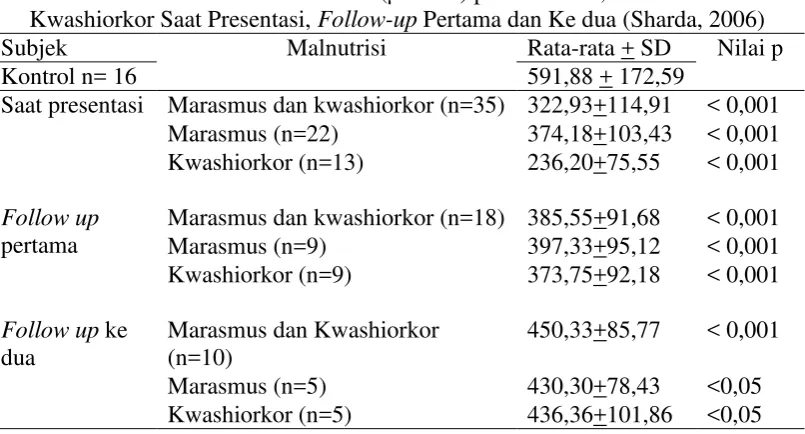
Subjek Malnutrisi Rata-rata + SD Nilai p
Kontrol n= 16 591,88 + 172,59
Saat presentasi Marasmus dan kwashiorkor (n=35) 322,93+114,91 < 0,001 Marasmus (n=22) 374,18+103,43 < 0,001 Kwashiorkor (n=13) 236,20+75,55 < 0,001
Follow up
pertama
Marasmus dan kwashiorkor (n=18) 385,55+91,68 < 0,001 Marasmus (n=9) 397,33+95,12 < 0,001 Kwashiorkor (n=9) 373,75+92,18 < 0,001
Follow up ke
Setelah diberikan suplementasi antioksidan, terjadilah meningkatan kadar aktivitas antioksidan total, tetapi sampai akhir bulan ke dua belum juga mencapai kadar yang sama dengan kontrol. Peningkatan kadar aktivitas antioksidan total lebih tinggi pada kelompok yang mendapatkan antioksidan pada follow up
pertama dan ke dua (Tabel 2.2).
Tabel 2.2
Kadar Aktivitas Antioksidan Total (µmol/L) pada Marasmus dan Kwashiorkor dengan dan Tanpa Suplementasi Antioksidan selama Follow up (Sharda, 2006)
Subjek Rata-rata + SD Nilai P
Follow up pertama dengan suplementasi n=13
405,61 + 95,47 <0,05
Follow up pertama tanpa suplementasi n=5 373,39 + 69,57 Tidak bermakna
Follow up ke dua dengan suplementasi n=8 428,03 + 83,28 <0,05
Follow up ke dua tanpa suplementasi n=2 454,55 + 128,6 Tidak bermakna
Saat presentasi n = 35 322,93 +
tikus dengan diet rendah protein selama 4 minggu didapatkan aktivitas katalase pada mukosa intestinal kelompok tikus dengan diet rendah protein meningkat lebih tinggi jika dibandingkan dengan kontrol. Konsentrasi glutathione dan aktivitas superoxide dismutase dan Se-dependent glutathione peroxidase pada mukosa intestinal adalah sama dengan kelompok kontrol. Basal short-circuit current (Isc) dan agonis Isc yang diinduksi oleh glukosa dan forskolin, seperti juga rf-lactoglobulin fluxes, lebih tinggi pada kelompok protein rendah. Stres H2O2 eksogen meningkatkan Isc secara bermakna pada kelompok protein rendah lebih tinggi daripada kelompok protein normal, tetapi tidak mengubah permeabilitas protein. Hasil ini menunjukkan bahwa malnutrisi menginduksi kerusakan radikal bebas intestinal dan mengubah transpor epitel, yang menunjukkan bahwa stres oksidatif berkontribusi terhadap disfungsi intestinal yang berhubungan dengan malnutrisi (Darmon, dkk., 1993)
2.4 Respon Imun pada Malnutrisi Berat (MB)
meregulasi dan mengkoordinasi berbagai aktivitas sel kekebalan alamiah) (Abbas, dkk., 2012b) (Gambar 2.4).
Gambar 2.4 Skema sistem kekebalan tubuh
Sel-sel utama pada kekebalan didapat adalah limfosit (limfosit B dan limfosit T (sel T helper, sel T sitotoksik, sel T regulator)), antigen presenting cells (APC), dan effector cells. Sel T helper akan mensekresikan sitokin apabila terdapat stimulasi antigenik. Sitokin ini bertanggung jawab terhadap berbagai respon seluler pada kekebalan alamiah dan didapat, jadi berfungsi sebagai molekul
messenger sistem imun. Sitokin yang disekresikan tersebut akan menstimulasi proliferasi dan diferensiasi sel T itu sendiri dan mengaktivasi sel lainnya (sel B, makrofag, dan leukosit lainnya). Sitokin bekerja secara autokrin, parakrin, atau endokrin (Abbas, dkk., 2012b).
Hubungan yang fundamental antara malnutrisi dan imunitas pertama kali digambarkan oleh Smythe dkk., sebagai defisiensi timolimfatik yang disebabkan
berhubungan dengan malnutrisi klinis dan gangguan fungsi timus (Paren, dkk., 1994).
Berbagai abnormalitas pada respon imun didapatkan berhubungan dengan kondisi malnutrisi energi protein, antara lain dalam hal jumlah sel T, rasio subset set T, aktivitas NK cells, dan produksi sitokin. Hasil dari penelitian-penelitian tersebut masih kontroversi. Beberapa penelitian menunjukkan bahwa malnutrisi menurunkan fungsi sel T, produksi sitokin, dan kemampuan limfosit berespon terhadap sitokin (Rodriguez, dkk., 2005).
Respon sel T terhadap stimulus tergantung pada signal yang diterima dari APC. Sel dendritik (salah satu APC) pada penderita malnutrisi jumlahnya berkurang. Jumlah APC meningkat setelah diterapi dengan terapi standar. Pada malnutrisi terdapat kegagalan maturasi sel dendritik yang berhubungan dengan endotoksemia. Anergi pada sel dendritik ini menyebabkan kegagalan proliferasi dari sel T (Hughes dkk, 2009).
Limfosit CD4+ anak gizi baik yang mengalami infeksi memiliki kemampuan cukup untuk berdiferensiasi. Fraksi sel CD4+CD45RO+ (memori) meningkat dan sel CD4+CD45RA+ (naive) menurun dalam darah tepi. Limfosit CD4+ pada anak malnutrisi tidak mampu mencapai jumlah fraksi sel memori yang cukup untuk memberikan perlindungan terhadap antigen luar dan untuk menimbulkan aktivitas
peningkatan proporsi limfosit B (CD20+) bila dibandingkan anak gizi baik tanpa infeksi bakteri. Pada anak malnutrisi dengan infeksi juga terjadi penurunan proporsi limfosit T CD4+ tetapi proporsi limfosit B (CD20+) juga menurun jika dibanding anak gizi baik yang mengalami infeksi. Hal ini menunjukkan bahwa penurunan proporsi T limfosit pada anak gizi baik yang mengalami infeksi tersebut berhubungan dengan penyakit infeksinya, dan ketidakmampuan untuk meningkatkan proporsi limfosit B pada anak malnutrisi berhubungan dengan menurunnya sintesis beberapa molekul yang terlibat dalam respon imunologis, sebagai akibat kurangnya nutrisi pada anak malnutrisi (Najera dkk., 2004).
Produksi IL-4 dan IL-10 dari sel CD4+ dan CD8+ anak malnutrisi meningkat, sementara produksi IL-2 dan IFN- menurun jika dibandingkan antara anak
malnutrisi dengan anak gizi baik tanpa dan dengan infeksi. Anak malnutrisi menunjukkan gangguan pada kemampuan aktivasi, akibat dari intensitas fluoresensi sel CD69+ dan CD 25+ lebih rendah dari sel-sel pada anak dengan gizi baik baik yang tidak terinfeksi maupun yang terinfeksi. Hasil ini menunjukkan bahwa malnutrisi mengubah kapasitas sel CD4+ dan CD 8+ untuk memproduksi IL-2, IFN- , IL-4 dan IL-10 terhadap respon dari infeksi. Kerusakan fungsional
macrophage inflammatory protein 2 (MIP-2), penurunan kemokin spesifik PMN (TNF-α, IL-6, IL-10), dan peningkatan reactive oxygen intermediate (ROI). Setelah perbaikan pemberian makan, maka dalam waktu singkat terjadi perbaikan pertahanan tubuh dengan perbaikan fungsi PMN dan produksi kemokin pada tempat inflamasi lokal (Ikeda, dkk., 2001).
Tikus model malnutrisi (yang diberikan restriksi diet selama 4 minggu) mengalami peningkatan konsentrasi sitokin proinflamasi dalam darah (TNF, IL-1, dan IFN-gamma sitokin Th-1) dibandingkan dengan kontrol, akan tetapi tidak terdapat perbedaan dalam sitokin antiinflamasi TGF-beta. Ekspresi mRNA yang mengkoding TNF dan IL-1 pada jantung juga meningkat. Ekspresi sitokin Th-1 (IFN-gamma) dan sitokin Th-17 (IL-17 dan IL-23p19) juga meningkat. Ekspresi mRNA sitokin pada hati tidak mengalami perubahan pada malnutrisi (Stevanovic, dkk., 2010).
Malnutrisi kalori protein menginduksi terjadinya supresi kekebalan tubuh, terjadi kerusakan macrophage respiratory burst activity (superoxide anion [O2-]
Penelitian tentang IL-6 pada malnutrisi ini memberikan hasil yang masih kontroversial, ada yang mendapatkan konsentrasinya meningkat dan ada yang mendapatkan ekspresi gennya menurun. Hal ini mungkin karena sampel dari penelitian tersebut berbeda yaitu menggunakan anak dengan malnutrisi ringan dan sedang saja (Gonzalez-Martinez dkk., 2008). Interleukin-6 selain berfungsi sebagai sitokin proinflamasi juga sebagai sitokin antiinflamasi. Interleukin-6 setelah berikatan dengan reseptor permukaan merangsang sel hati memproduksi protein fase akut dengan beberapa jalur. Jalur pertama melalui jalur aktivasi
receptor-associated Janus-activated kinases (JAKs) yang mengaktifkan faktor transkripsi signal tranducers and activators of transcription (STATs). Pengaktivan dari STATs ini memegang peranan penting dalam menginduksi dan memodulasi transkripsi gen multipel temasuk yang mengkode protein fase akut. Salah satu protein fase akut pada tikus yaitu α1-acid glycoprotein meningkat kadarnya saat terjadi inflamasi dan dipengaruhi oleh TNF, IL-1, dan IL-6. Jalur ke dua adalah yang melibatkan berbagai macam mitogen-activated protein (MAP)
Umumnya AFLD dan NAFLD diawali dengan steatosis hati. Jika steatosis ini terus berlangsung maka akan berkembang menjadi steatohepatitis, cirrhosis, dan kanker hati. Steatosis hati menunjukkan adanya akumulasi berlebih lemak (trigliserida) pada sel parenkim hati (hepatosit), dan terjadi karena berbagai penyebab.
Steatosis hati secara morfologi bermanifestasi sebagai akumulasi droplet lemak intrasitoplasma besar (makro vesikular) atau kecil (mikro vesikular) dalam sel parenkim hati. Diagnosis steatosis dibuat jika kandungan lemak hati melebihi 5-10% dari beratnya.
Pada kondisi malnutrisi misalnya kwashiorkor, hepatic steatosis yang terjadi utamanya adalah makro vesikular. Pada bentuk makro vesicular didapatkan suatu vakuola tunggal lemak besar dalam sel hepatosit yang mengisi sitoplasma dan menggeser inti ke perifer, sehingga didapatkan gejala khas yang disebut signet ring appearance. Steatosis hati itu sendiri tidak berbahaya, reversibel, dan tidak progresif jika penyebabnya dihilangkan. Perkembangan menjadi steatohepatitis baik pada ASH maupun NASH dipengaruhi oleh masih tetapnya dan keparahan dari penyebab steatosis hati tersebut (Reddy dan Rao, 2006).
penggunaan energi dan meningkatkan lipogenesis (dimediasi PPAR- ), menghasilkan steatosis dan steatohepatitis. Steatohepatitis alkohol dan nonalkohol disebabkan karena gangguan pada sistem oksidasi asam lemak di hati.
Abnormalitas yang berhubungan dengan sistem oksidasi asam lemak yang berbeda disebabkan karena genetik, racun (termasuk obat-obatan), dan gangguan metabolisme juga akan menghasilkan penurunan pembakaran energi di hati, sehingga menimbulkan penumpukan lemak di sel hati. Efisiensi dari PPAR-α sensing di hati penting untuk mengenal dan merespon influx asam lemak dalam kasus kelaparan atau puasa, dimana influx asam lemak secara kuat menginduksi aktivitas ke tiga sistem oksidasi asam lemak untuk mencegah steatosis hati. Puasa semalaman atau berkepanjangan menimbulkan steatosis hati berat jika PPAR-α sensing tidak efisien (Reddy dan Rao, 2006).
mekanisme yaitu peningkatan sintesis lemak hati, gangguan pada transportasi pengeluaran lemak hati, atau penurunan pemecahan lemak dalam hati. Mekanisme peningkatan sintesis lemak hati tidak dapat dikonfirmasi dengan didapatkannya penurunan dari aktivitas glucose-6-phosphatase. Mekanisme tidak adekuatnya sintesis lipoprotein dan sekresinya secara teori tampaknya merupakan penjelasan yang tepat. Konsentrasi lipoprotein sirkulasi didapatkan rendah pada anak dengan malnutrisi berat dan sintesis protein pun mengalami gangguan. Lemak hati tidak berkorelasi baik dengan konsentrasi sirkulasi lipoprotein.
Mekanisme yang ke tiga yaitu penurunan pemecahan lemak hati (Doherty, dkk., 1991). Penelitian Leung dan Peter (1986) melihat biopsi hati yang diambil dari pasien perlemakan hati karena alkohol dan mendapatkan adanya gangguan -oksidation, menyimpulkan bahwa penurunan -oksidation ini penting dalam patogenesis perlemakan hati. Beta-oxidation lemak terutama terjadi dalam mitokondria tetapi fungsi mitokondria pada anak malnutrisi berat masih baik (Waterlow, 1961), sehingga hipotesis gangguan -oxidation sebagai penyebab perlemakan hati menjadi jauh. Lazarow dan de Duve (1976) mendapatkan peranan peroxisomal -oxidation serupa tetapi merupakan suatu sistem yang terpisah dari
memiliki kapasitas untuk menyelesaikan -oxidation substratnya, tetapi hal ini belum pernah diobservasi secara in vivo. Peroxisome memendekan asam lemak rantai panjang yang kemudian ditransportasi ke mitokondria untuk proses oksidasi lebih lanjut. Secara teori, walaupun dalam kondisi fungsi mitokondria masih baik, maka suatu defek yang terjadi pada peroxisome ( -oxidation) masih memungkinkan menimbulkan akumulasi lemak hati. Akumulasi lemak kemudian cenderung pada rantai yang lebih panjang. Transportasi asam lemak menyeberangi membran peroxisome terjadi tidak tergantung dari karnitin, sementara pada mitokondria sebaliknya. Tidak seperti mitokondria, peroxisomal -oxidation tidak terganggu pada kondisi tanpa adanya riboflavin. Aktivitas sistem peroxisomal -oxidation dipengaruhi oleh banyak faktor eksternal.
Peroxisome memiliki waktu paruh yang pendek dan ini dapat berkurang lagi dengan adanya stres tertentu (Doherty, dkk., 1991).