SELEKSI SEGREGAN TRANSGRESIF PADA GENERASI F3
DAN F4 KACANG TANAH (Arachis hypogaea L.)
SITI NURHIDAYAH
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2016
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa tesis berjudul Seleksi Segregan Transgresif pada Generasi F3 dan F4 Kacang Tanah (Arachis hypogaea L.) adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, April 2016
Siti Nurhidayah
RINGKASAN
SITI NURHIDAYAH. Seleksi Segregan Transgresif pada Generasi F3 dan F4 Kacang Tanah (Arachis hypogaea L.). Dibimbing oleh YUDIWANTI WAHYU ENDRO KUSUMO dan WILLY BAYUARDI SUWARNO.
Kacang tanah (Arachis hypogaea L.) merupakan tanaman yang banyak dibudidayakan di berbagai negara sebagai sumber minyak nabati dan protein tinggi yang baik untuk kesehatan. Permintaan kacang tanah mengalami peningkatan seiring dengan meningkatnya jumlah penduduk dan kebutuhan masyarakat juga ikut meningkat. Kacang tanah memiliki adaptasi cukup luas, tahan terhadap kekeringan dan cocok ditanam pada dataran rendah sampai sedang, relatif suhu rata-rata 27-32 ⁰C. Penelitian ini dilakukan dengan tujuan memanfaatkan fenomena segregasi transgresif agar diperoleh genotipe kacang tanah dengan potensi hasil tinggi yang dapat diprediksi pada generasi awal segregasi sehingga efektivitas seleksi dapat tercapai.
Penelitian dilaksanakan dua tahap. Tahap pertama pada bulan Agustus-Desember 2014 untuk mendeteksi segregan transgresif pada populasi F3. Galur segregan transgresif merupakan galur yang memiliki nilai tengah terbaik daripada tetua terbaiknya dengan ragam dalam famili yang rendah atau sama dengan varietas pembanding. Tahap kedua dilaksanakan pada bulan Februari-Mei 2015 untuk memverifikasi galur-galur segregan transgresif pada populasi F4 kacang tanah. Galur-galur hasil identifikasi di generasi F3 yang diduga sebagai segregan transgresif kemudian diverifikasi nilai tengah dan ragam dalam familinya dengan melihat kekonsistenan nilai tengah dan ragam yang seragam atau sama dengan varietas pembanding. Penelitian dilakukan di lapangan percobaan Leuwikopo Dramaga IPB pada ketinggian 250 m dpl. Bahan genetik yang digunakan terdiri atas 218 galur F3 dan F4 yang berasal dari 5 persilangan biparental (Jerapah/GWS79A1, GWS79A1/Zebra, GWS79A1/Jerapah, Zebra/ GWS79A1, dan Zebra/GWS18) dan 4 varietas komersial yang digunakan sebagai pembanding (Gajah, Jerapah, Sima, dan Zebra).
Hasil studi keragaman, heritabilitas dan aksi gen pada populasi F3 menunjukkan bahwa genotipe berpengaruh nyata terhadap karakter jumlah polong isi dan jumlah polong total sehingga galur-galur yang diuji diprediksi memiliki potensi keragaman yang cukup tinggi. Hasil analisis skewness dan kurtosis memperlihatkan bahwa karakter-karakter kuantitatif secara umum dipengaruhi oleh banyak gen dengan aksi gen aditif, dominan, epistasis komplementer atau epistasis aditif. Karakter jumlah polong isi, jumlah polong total dan bobot biji per tanaman memiliki nilai heritabilitas tinggi dengan koefisien keragaman genetik yang sedang sehingga efektif untuk dilakukan seleksi. Seleksi pada jumlah polong total didapatkan 22 galur yang memiliki keragaan fenotipe dengan jumlah polong terbaik dengan ragam yang sudah seragam atau masih beragam. Terhadap galur-galur yang masih beragam dapat dilakukan seleksi pada generasi berikutnya. Galur U2-39 terdeteksi sebagai galur segregan transgresif dengan jumlah polong total sebanyak 30 polong dengan ragam rendah daripada varietas Sima.
Hasil studi generasi F4 menunjukkan bahwa hampir semua galur-galur yang baik pada generasi F3 tidak menunjukkan kekonsistenan nilai tengah meskipun ragam dalam famili diduga telah seragam. Terdapat 2 galur memiliki jumlah polong
tinggi dan seragam yaitu galur U2-108 dan U3-118 sehingga dapat dijadikan sebagai galur harapan untuk diteruskan pada generasi selanjutnya. Tidak ditemukannya galur segregan transgresif pada percobaan ini karena galur U2-39 yang diduga sebagai segregan transgresif tidak menunjukkan kekonsistenan nilai tengah pada generasi F4 bahkan cenderung mengalami penurunan nilai tengah.
Hasil studi analisis korelasi dan lintasan pada karakter agronomi kacang tanah menunjukkan bahwa karakter bobot polong kering total memiliki pengaruh langsung dan korelasi yang tinggi, sementara karakter jumlah polong total memiliki pengaruh tidak langsung melalui karakter bobot polong kering total terhadap perbaikan bobot biji per tanaman. Kedua karakter tersebut dapat dijadikan sebagai karakter seleksi untuk merakit varietas unggul kacang tanah berdaya hasil tinggi. Kata kunci: jumlah polong total, kacang tanah, korelasi, segregan transgresif
SUMMARY
SITI NURHIDAYAH. Selection of Transgressive Segregants on F3 and F4 Populations of Peanut (Arachis hypogaea L.). Supervised by YUDIWANTI WAHYU ENDRO KUSUMO and WILLY BAYUARDI SUWARNO.
Peanut (Arachis hypogaea L.) is widely cultivated in many countries as a source of vegetable oil and protein which is good for the human body. Demand for peanuts has increased along with the increasing population and consumption. Peanut is widely adapted, tolerant to drought, and suitable to be planted in the lowlands and mid-altitudes with average temperatures of 27-32 ⁰C. The main objective of this research was to study the phenomenon of transgressive segregation in peanut, so that high yielding genotypes can be identified in early segregating generations.
The study consisted of two experiments that were conducted in different planting seasons. The first was performed in August-December 2014 to identify putative transgressive segregants in F3 population. The transgressive segregant lines should have high mean and small variance (i.e. smaller than or equal to that of check varieties). The second was conducted in February-May 2015 to verify the performance of the putative transgressive segregant lines on F4 population. This research was conducted at Leuwikopo Experimental Station of Bogor Agricultural University (IPB), Darmaga, Bogor (250 masl). Genetic material used consisted of 218 F3 and F4 lines derived from 5 biparental crosses (Jerapah/GWS79A1, GWS79A1/Zebra, GWS79A1/Jerapah, Zebra/GWS79A1, and Zebra/GWS18) and 4 commercial varieties as checks (Gajah, Jerapah, Sima, and Zebra).
The study of diversity, heritability and gene action in the F3 populations showed that the genotype factor significantly affected the number of filled pods and total number of pods, indicating that the tested lines have some extent of genetic variability for yield. Skewness and kurtosis analysis results showed that the quantitative characters in general are influenced by many genes with additive, dominant, epistasis complementary, or additive epistasis gene actions. Number of filled pods, total number of pods, and seed yield per plant have high heritability with moderate value of genetic diversity coefficient, indicating an opportunity for obtaining effective selection. Selection on the total number of pods resulted in 22 families having largest number of pods with different extent of variance within family. Lines with large variances within family may be further selected in an individual plant basis in in subsequent generations. Line U2-39 is considered as a putative transgressive segregant with total number of pods of 30 and have smaller variance within family than that of Sima variety.
Results from the F4 experiment indicated that almost all well-performing F3 genotypes did not have similar performance in F4 despite variance within family was reduced. At least two promising F4 lines have good and uniform number of pods, namely U2-108 and U3-118, to be evaluated further in subsequent generations. The putative transgressive segregant selected in F3, U2-39, exhibited a decrease on average total number of pods in F4.
Correlation and path analysis of agronomic traits of peanut showed that there were differences among generations for the correlations between traits. Total dry weight of pods have high direct and positive correlations with seed weight per plant
in the two generations. Total number of pods have high indirect and positive correlations with seed weight per plant through total dry weight of pods character. These characters may be used as selection criteria for developing high-yielding peanut varieties.
© Hak Cipta Milik IPB, Tahun 2016
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
Dilarang mengumumkan dan memperbanyak sebagian atau seluruh karya tulis ini dalam bentuk apa pun tanpa izin IPB
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Pemuliaan dan Bioteknologi Tanaman
SELEKSI SEGREGAN TRANSGRESIF PADA GENERASI F3
DAN F4 KACANG TANAH (Arachis hypogaea L.)
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2016
Penguji Luar Komisi pada Ujian Tesis: Dr Ir Hajrial Aswidinnoor, MSc
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah subhanahu wa ta’ala atas segala karunia-Nya sehingga karya ilmiah ini berhasil diselesaikan. Perakitan varietas unggul kacang tanah masih perlu dilakukan untuk memenuhi permintaan kacang tanah dalam negeri. Seleksi merupakan salah satu tahapan penting yang perlu dilalui khususnya dalam pemuliaan konvensional. Seleksi segregan transgresif merupakan modifikasi seleksi pada generasi awal segregasi.
Tesis yang berjudul Seleksi Segregan Transgresif pada Generasi F3 dan F4 Kacang Tanah (Arachis hypogaea L.) merupakan salah satu upaya dalam menghasilkan galur-galur berdaya hasil tinggi dengan ragam dalam galur yang rendah sehingga efektivitas seleksi dapat tercapai dengan mudah dan efisien.
Penulis menyampaikan rasa hormat dan terima kasih kepada:
1. Dr Ir Yudiwanti Wahyu E.K., MS dan Dr Willy Bayuardi Suwarno, SP MSi selaku komisi pembimbing atas bimbingan, arahan, saran, dan dorongan kepada penulis selama proses penelitian sampai akhir dari penyelesaian tesis.
2. Dr Ir Hajrial Aswidinnoor, MSc dan Dr Ir M. Rahmad Suhartanto, MSi selaku dosen penguji luar komisi dan perwakilan dari program studi atas
tambahan ilmu filosofi, masukan dan sarannya untuk menyempurnakan penulisan makalah tesis.
3. Kedua orang tua (Ibunda Karsiah dan Ayahanda Abdul Karim), kakanda (Asmawati, SP; Dede Kurniawan; Dian Rodiani; Lutfi) dan adinda (Nurmala, SkomI; Erna Muliana; Erni Muliani; Husna Ginanisa; Lita Irmawati; Hana Mujtahidah Fitriani) atas dukungan do’a, moril, motivasi, dan kasih sayangnya kepada penulis.
4. Bapak Rahmat, Bapak Supriatna, dan seluruh teknisi kebun percobaan Leuwikopo yang telah menyediakan fasilitas alat, ruang, dan tenaga selama proses penelitian.
5. Arina Saniaty, SP; Trias Sitaresmi, SP; Sofia Hanum, SP; Hasrat Enggal Prayogi, SP; Silvi Nur Arifah SSi; Fithriya Y.R., SSi; Inggit Radesiyani, SSi; Siti Aminah, SPt; Nopi Elida, SPt; yang telah membantu dan memotivasi penulis.
6. Rekan-rekan PBT 2013, FORSCA-AGH, dan kost Wisma Tanjung atas kebersamaannya untuk saling memotivasi satu sama lain.
7. Direktorat Jenderal Pendidikan Tinggi, Kementerian Pendidikan dan Kebudayaan atas dukungan dana penelitian melalui Beasiswa Pendidikan Pascasarjana Dalam Negeri tahun 2013 dan skim Penelitian Strategis Nasional tahun 2014 serta Beasiswa Pemerintah Provinsi Jawa Barat atas dukungan materi sehingga penulis dapat menyelesaikan studi dan tugas akhir.
Bogor, April 2016
DAFTAR ISI
DAFTAR TABEL xvii
DAFTAR GAMBAR xviii
DAFTAR LAMPIRAN xviii
1 PENDAHULUAN 1
Latar Belakang 1
Tujuan Penelitian 2
Hipotesis Penelitian 2
2 TINJAUAN PUSTAKA 4
Sejarah Penyebaran Kacang Tanah 4
Biologi dan Morfologi Kacang Tanah 4
Syarat Tumbuh Kacang Tanah 5
Komponen Genetik dan Aksi Gen 5
Segregan Transgresif 7
3 KERAGAMAN GENETIK DAN SELEKSI SEGREGAN
TRANSGRESIF PADA POPULASI F3 KACANG TANAH 8
Abstrak 8
Pendahuluan 10
Metode Penelitian 11
Hasil dan Pembahasan 14
Simpulan 22
4 VERIFIKASI GALUR-GALUR SEGREGAN TRANSGRESIF
PADA POPULASI F4 KACANG TANAH 23
Abstrak 23
Pendahuluan 25
Metode Penelitian 25
Hasil dan Pembahasan 27
Simpulan 36
4 ANALISIS KORELASI DAN ANALISIS LINTAS PADA
DUA GENERASI KACANG TANAH 37
Abstrak 37
Pendahuluan 39
Metode Penelitian 39
Hasil dan Pembahasan 41
6 PEMBAHASAN UMUM 46
7 SIMPULAN DAN SARAN 49
Simpulan 49
Saran 49
DAFTAR PUSTAKA 51
LAMPIRAN 57
DAFTAR TABEL
1. Analisis ragam rancangan Augmented 12
2. Analisis ragam dengan kuadrat tengah karakter agronomi kacang tanah
generasi F3 14
3. Keragaan nilai tengah dan standar deviasi karakter pada tiap kombinasi
persilangan kacang tanah 15
4. Keragaan nilai tengah dan standar deviasi karakter pada varietas
pembanding kacang tanah 15
5. Ukuran dan aksi gen karakter kuantitatif berdasarkan sebaran data
galur-galur F3 pada 5 persilangan kacang tanah 17
6. Nilai P dari uji kenormalan dan kehomogenan ragam, serta uji skewness dan kurtosis karakter kuantitatif pada 4 varietas kacang tanah 18 7. Komponen nilai ragam genetik, heritabilitas dan koefisien keragaman
genetik galur-galur kacang tanah generasi F3 19
8. Galur terbaik hasil seleksi berdasarkan jumlah polong total kacang tanah 21 9. Kuadrat tengah karakter agronomi kacang tanah generasi F4 27 10. Komponen ragam genetik, heritabilitas arti luas, dan koefisien
keragaman genetik karakter agronomi kacang tanah generasi F4 28 11. Keragaan galur-galur hasil seleksi berdasarkan jumlah polong total
kacang tanah pada 2 generasi 29
12. Nilai F-hitung hasil analisis ragam pengaruh musim terhadap karakter agronomi pada 4 varietas pembanding kacang tanah 31 13. Koefisien korelasi rataan karakter semua galur antara F3 dan F4 pada tiap
kombinasi persilangan kacang tanah 32
14. Verifikasi nilai tengah dan ragam galur segregan transgresif kacang tanah
berdasarkan karakter jumlah polong total 34
15. Heritabilitas tiap kombinasi persilangan biparental dengan metode
Parent-Offspring pada kacang tanah 35 16. Matriks korelasi antar karakter agronomi galur-galur kacang tanah
generasi F3 42
17. Matriks korelasi antar karakter agronomi galur-galur kacang tanah
generasi F4 42
18. Matriks analisis lintas terhadap karakter bobot biji per tanaman generasi
F3 kacang tanah 43
19. Matriks analisis lintas terhadap karakter bobot biji per tanaman generasi
DAFTAR GAMBAR
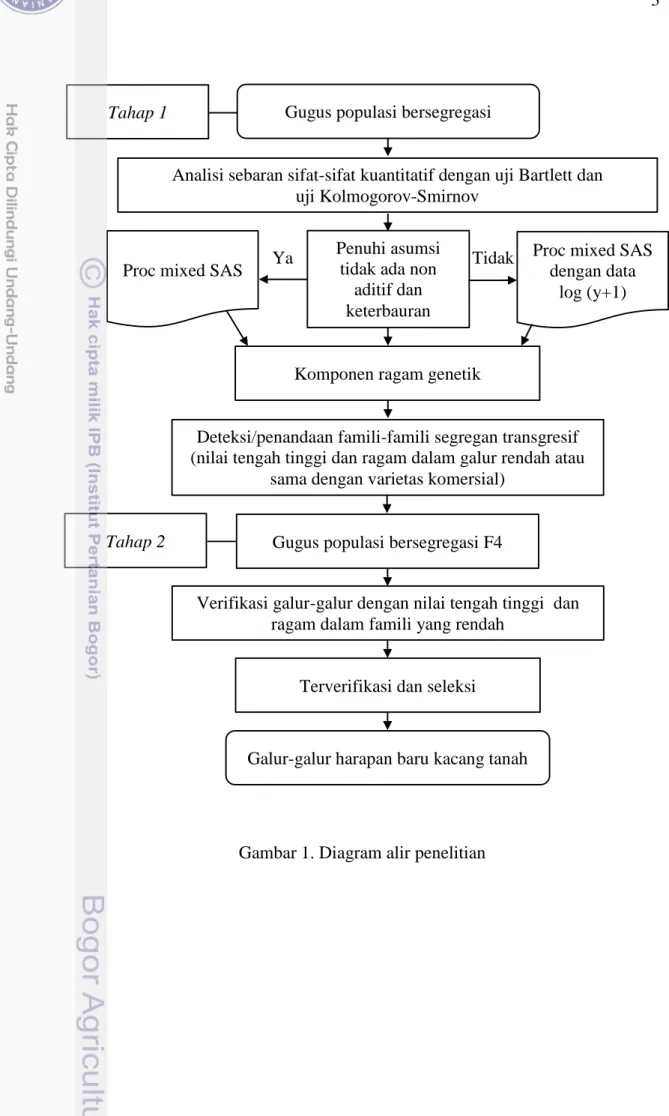
1. Diagram alir penelitian 3
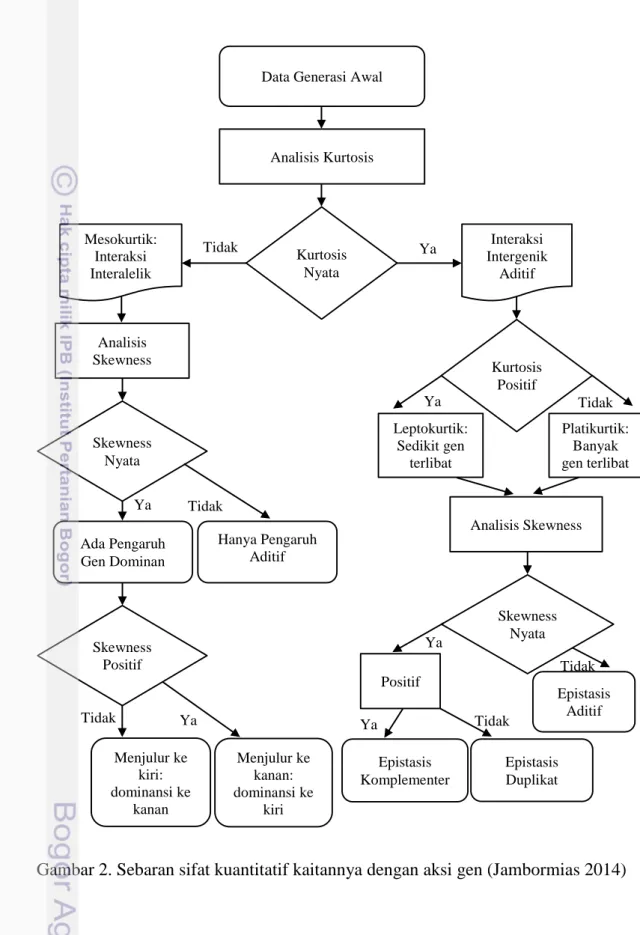
2. Sebaran sifat kuantitatif kaitannya dengan aksi gen 13 3. Sebaran nilai tengah jumlah polong total dan ragam dalam famili pada
218 galur kacang tanah 20
4. Sebaran 22 galur kacang tanah hasil seleksi 10% berdasarkan karakter
jumlah polong total 21
5. Keragaan rataan dan ragam jumlah polong total galur-galur kacang tanah
terseleksi pada generasi F3 30
6. Keragaan rataan dan ragam jumlah polong total galur-galur kacang tanah
terseleksi generasi F4 30
7. Verifikasi nilai tengah dan ragam karakter jumlah polong total
galur-galur generasi F3 dan F4 kacang tanah 33
8. Diagram lintasan karakter bobot biji per tanaman generasi F3 kacang
tanah 43
9. Diagram lintasan karakter bobot biji per tanaman generasi F4 kacang
tanah 44
DAFTAR LAMPIRAN
1. Deskripsi kacang tanah varietas Jerapah 59
2. Deskripsi kacang tanah varietas Zebra 60
3. Rata-rata curah hujan, temperatur, kelembaban dan intensitas cahaya
selama penelitian percobaan tahun 2014 61
4. Rata-rata curah hujan, temperatur, kelembaban dan intensitas cahaya
1 PENDAHULUAN
Latar BelakangKacang tanah adalah tanaman yang berasal dari Amerika Selatan dengan daya adaptasi luas di negara subtropis dan tropis. Menurut Wang et al. (2012) kacang tanah menempati urutan ke 4 sebagai sumber minyak nabati dan urutan ke 13 sebagai pangan penting dunia yang ditanam luas lebih dari 80 negara. Menurut Janila et al. (2013), kacang tanah ditanam lebih dari 24 juta hektar di dunia. Sumarno (1993) memaparkan bahwa tanaman ini ditanam luas di Indonesia khususnya di daerah Jawa Timur, Jawa Tengah, dan Jawa Barat. Produktivitas kacang tanah berkisar 0.5 ton ha-1 hingga 5 ton ha-1 polong kering. Amerika dan Australia merupakan negara dengan produktivitas tertinggi berkisar 3 ton ha-1. Sedangkan produktivitas negara tropis hanya berkisar 0.7 hingga 1.3 ton ha-1 polong kering. Sumarno (1993); Adisarwanto (2001); Yudiwanti et al. (2008) menjelaskan bahwa rendahnya produktivitas dipengaruhi oleh berbagai faktor seperti teknologi budi daya, sifat agroklimat, hama penyakit, jenis varietas, umur panen, dan usaha taninya.
Pertambahan jumlah penduduk dunia dan semakin meningkatnya kebutuhan protein dan lemak mengakibatkan permintaan kacang tanah terus meningkat. Setiap negara mencoba untuk memenuhi kecukupan kacang tanah dalam negerinya (Sumarno 1993). Hasil penelitian Naeem-ud-Din et al. (2012) melaporkan bahwa negara Pakistan memiliki varietas BARI-2011 dengan potensi hasil 2.5 – 6.3 ton ha-1 polong kering, toleran kekeringan, tahan penyakit bercak daun dan busuk akar. Varietas kacang tanah dalam negeri yang telah dilepas tahun 1995 sampai 2014 berjumlah 34 varietas dengan produktivitas masih rendah yaitu 1.2-3.0 ton ha-1 polong kering (Kasno & Harnowo 2014). Produksi yang masih rendah belum mampu mencukupi kebutuhan dalam negeri sehingga perakitan varietas unggul dengan produktivitas tinggi masih perlu dilakukan agar kebutuhan masyarakat terpenuhi.
Hasil tinggi merupakan tujuan utama dalam perkembangan genetika tanaman (Wang et al. 2012). Salah satu langkah yang ditempuh dalam pemuliaan tanaman adalah seleksi (Sa’diyah et al. 2009). Kegiatan seleksi merupakan tahapan penting dalam produksi kacang tanah (Smith & Simpson 1995). Ada beberapa hal yang perlu diperhatikan pada seleksi berdasarkan fenotipe tanaman antara lain keragaman, heritabilitas, dan korelasinya pada karakter agronomi yang dapat meningkatkan hasil kacang tanah (Sa’diyah et al. 2009; Jogloy et al. 2011).
Keefektifan seleksi dipengaruhi oleh tersedianya keragaman dalam populasi yang akan diseleksi. Makin besar tingkat keragaman dalam populasi efektifitas seleksi untuk memilih suatu karakter yang sesuai dengan keinginan makin besar (Sa’diyah et al. 2009). Selain keragaman, faktor lain yang menentukan keefektifan suatu seleksi adalah nilai duga heritabilitas. Heritabilitas merupakan parameter genetik yang dapat mengukur sejauh mana suatu genotipe mewariskan sifat-sifat yang dimiliki oleh suatu tanaman. Informasi hubungan antar karakter dapat dihitung dengan analisis korelasi. Analisis korelasi dapat melihat sejauh mana keeratan hubungan antar karakter. Persaingan antar karater dapat menimbulkan korelasi
2
negatif antar karakter. Faktor genetik yang dapat menyebabkan terjadinya korelasi antar karakter adalah pautan (linkage) dan pleiotropi (Falconer 1989).
Korelasi genetik dapat dimanfaatkan untuk seleksi tidak langsung apabila karakter utama yang diseleksi mempunyai heritabilitas tinggi. Adanya korelasi antara dua karakter dapat dipengaruhi oleh peristiwa pleiotropi dan linkage.
Pleiotropi merupakan fenomena yang terjadi bila suatu gen pada lokus atau suatu
set gen pada beberapa lokus mengendalikan dua karakter yang berbeda atau lebih. Apabila sebagian dari lokus mengendalikan karakter A juga mengendalikan karakter B, maka kedua karakter tersebut dikatakan berkorelasi secara genetik.
Linkage adalah fenomena terdapatnya dua gen atau lebih yang mengendalikan dua
atau lebih karakter berbeda yang terletak pada unit kromosom yang sama. Kedua karakter tersebut cenderung akan diwariskan secara bersama-sama sebab kendali gen terletak pada kromosom yang sama seperti kehadiran karakter A selalu diikuti karakter B (Sa’diyah et al. 2009). Menurut Kotzamanidis (2006) bahwa karakter hasil dan kualitas hasil dapat ditingkatkan dengan memanfaatan fenomena segregasi transgresif pada persilangan Virginia dan Spanish. Jambormias (2014) melaporkan bahwa fenomena segregasi transgresif pada kacang hijau didapatkan 18-24% famili segregan transgresif.
Berbagai macam cara telah dilakukan oleh para peneliti dari lembaga penelitian maupun peneliti bagian Genetika dan Pemuliaan Tanaman IPB dalam menghasilkan varietas unggul kacang tanah. Salah satunya adalah pemanfaatan fenomena segregasi transgresif dengan cara seleksi segregan transgresif yang mampu mempercepat fiksasi gen sampai generasi F4 sehingga efektivitas waktu dapat tercapai. Penelitian ini terdiri atas dua tahapan percobaan (Gambar 1). Percobaan pertama mempelajari keragaan galur-galur F3 dan mendeteksi segregan transgresif dengan mencari informasi nilai ragam genetik, heritabilitas, koefisien keragaman genetik, dan aksi gen yang mengendalikan karakter kuantitatif. Percobaan kedua memverifikasi galur-galur yang diduga sebagai segregan transgresif pada generasi F4.
Tujuan Penelitian Penelitian ini bertujuan untuk:
1. Memperoleh informasi keragaan dan keragaman galur-galur hasil 5 persilangan biparental kacang tanah
2. Mendapatkan galur-galur yang teridentifikasi sebagai segregan transgresif pada generasi F3 kacang tanah
3. Memverifikasi galur-galur segregan transgresif pada populasi F4 kacang tanah 4. Memperoleh informasi korelasi dan pengaruh langsung dan tidak langsung
antar karakter agronomi kacang tanah
Hipotesis Penelitian
1. Terdapat keragaman antar galur-galur pada populasi F3 dan F4 kacang tanah 2. Terdapat minimal satu genotipe kacang tanah yang teridentifikasi sebagai
segregan transgresif
3. Terdapat minimal satu galur yang terverifikasi sebagai segregan transgresif 4. Terdapat korelasi dan pengaruh yang besar antara karakter agronomi terhadap
3
Gambar 1. Diagram alir penelitian
Tahap 2
Tahap 1 Gugus populasi bersegregasi
Proc mixed SAS
Proc mixed SAS dengan data
log (y+1) Analisi sebaran sifat-sifat kuantitatif dengan uji Bartlett dan
uji Kolmogorov-Smirnov Penuhi asumsi
tidak ada non aditif dan keterbauran
Komponen ragam genetik
Deteksi/penandaan famili-famili segregan transgresif (nilai tengah tinggi dan ragam dalam galur rendah atau
sama dengan varietas komersial)
Verifikasi galur-galur dengan nilai tengah tinggi dan ragam dalam famili yang rendah
Terverifikasi dan seleksi Gugus populasi bersegregasi F4
Galur-galur harapan baru kacang tanah
4
2 TINJAUAN PUSTAKA
Sejarah Penyebaran Kacang TanahKacang tanah merupakan komoditas kedua terpenting setelah kedelai di Indonesia. Kacang tanah berasal dari Amerika Latin tepatnya Brazil (Pusdatin 2013), kemudian tanaman ini mulai masuk ke Indonesia pada abad ke 17 yang dibawa oleh pedagang Cina dan Portugis (Ristek 2000). Saat ini kacang tanah telah menyebar ke seluruh dunia baik yang beriklim tropis maupun subtropis termasuk Indonesia (Pusdatin 2013). Produktivitas rata-rata di tingkat petani sebesar 1 ton ha-1 polong kering (Faisal & Polakitan 2014). Sementara produktivitas rata-rata nasional mencapai 1.25 ton ha-1 polong kering (Kasno & Harnowo 2014). Varietas unggul kacang tanah ditandai dengan karakteristik daya hasil tinggi, ukuran biji seragam, umur genjah antara 85-90 hari, hasilnya stabil, tahan terhadap penyakit utama (karat dan bercak daun), dan toleran cekaman kekeringan (Ristek 2000; Syamsiar 2010).
Biologi dan Morfologi Kacang Tanah
Kacang tanah memiliki kromosom rangkap empat (tetraploid) yaitu pada tiap selnya berjumlah 40 buah kromosom atau 10 pasang kromosom rangkap empat. Kacang tanah yang telah dibudidayakan saat ini diperkirakan berasal dari persilangan dua kacang tanah jenis liar berkromosom rangkap dua. Pewarisan sifat kacang tanah ini mirip dengan tanaman yang berkromosom rangkap dua sehingga disebut dengan amfiploid atau allotetraploid (Kasno 1993; Syukur 2013; Syukur & Wahyu 2013). Kacang tanah tergolong ke dalam tanaman menyerbuk sendiri (Kasno 1993). Persarian terjadi sebelum bunga mekar (kleistogami), sehingga jarang sekali terjadi penyerbukan silang (Kotzamanidis 2006; Kasno & Harnowo 2014).
Kacang tanah dibagi atas tiga tipe yaitu tipe Valensia, Virginia dan Spanish. Tipe Virginia tumbuh menjalar, cabang banyak, batang utama tidak berbunga, berbiji besar dan berumur dalam. Tipe Valensia dan Spanish tumbuh tegak, cabang sedikit, batang utama berbunga, berbiji kecil, berumur sedang, polong berbiji 2 atau lebih. Perbedaan tipe Valensia dan Spanish terletak pada jumlah biji per polong. Spanish memiliki 2 biji per polong, sebaliknya Valensia lebih dari 2 biji per polong. Penyebaran di Indonesia lebih banyak pada tipe Spanish (Trustinah 1993). Taksonomi kacang tanah menurut Acquaah (2007) adalah sebagai berikut:
Kingdom : Plantae Subkingdom : Tracheobionta Divisi : Magnoliophyta Sub Divisi : Angiospermae Kelas : Magnoliopsida Subclass : Rosidae Ordo : Fabales Famili : Fabaceae Genus : Arachis
5
Berdasarkan letak cabang lateral, pertumbuhan kacang tanah dibagi atas dua tipe yaitu tipe menjalar dan tipe tegak. Tipe menjalar meliputi runner, trailing,
procumbent, dan postate sedangkan tipe tegak meliputi upright, erect bunch, dan bunch. Perakaran kacang tanah termasuk akar tunggang dan akar-akar lateral. Daun
pada kacang tanah memiliki empat helai sehingga disebut tetra foliate. Bunga kacang tanah termasuk bunga sempurna yaitu kelamin jantan dan betina terdapat pada satu bunga. Pada saat pembuahan terjadi kemudian muncul bakal buah yang tumbuh memanjang yang biasa disebut dengan ginofor. Ginofor bersifat geotropik sehingga dengan sendirinya ginofor masuk ke dalam tanah sedalam 2-7 cm. Warna ginofor umumnya hijau atau merah dan ungu bila terdapat pigmen antosianin. Polong kacang tanah pun bervariasi baik bentuk, ukuran, dan kontriksinya (Trustinah 1993).
Syarat Tumbuh Kacang Tanah
Kacang tanah merupakan sejenis tanaman tropika yang tumbuh secara perdu setinggi 30 hingga 50 cm dan mengeluarkan daun-daun kecil (Pusdatin 2013). Curah hujan yang sesuai untuk kacang tanah sekitar 800–1 300 mm th-1. Hujan yang terlalu keras dapat mengakibatkan rontok dan bunga tidak terserbuki oleh lebah. Selain itu, hujan yang terus-menerus akan meningkatkan kelembaban di sekitar per tanaman kacang tanah. Suhu udara sekitar 28º–32 ºC, sedangkan menurut Janila et
al. (2013) suhu optimum antara 25°C dan 30°C. Suhu di bawah 10 ºC menyebabkan
pertumbuhan tanaman sedikit terhambat, bahkan menjadi kerdil dikarenakan pertumbuhan bunga yang kurang sempurna. Kelembaban udara untuk tanaman kacang tanah berkisar antara 65–75%. Adanya curah hujan yang tinggi akan meningkatkan kelembaban terlalu tinggi di sekitar per tanaman. Penyinaran sinar matahari secara penuh amat dibutuhkan bagi tanaman kacang tanah, terutama kesuburan daun dan perkembangan besarnya kacang (Ristek 2000). Pembentukan polong merupakan periode yang sangat peka terhadap kekurangan air karena dapat mengurangi pembungaan, pembentukan polong dan penurunan hasil akhir. Pada fase ini merupakan akumulasi bahan kering yang maksimum (Trustinah 1993).
Komponen Genetik dan Aksi Gen
Karaker sederhana seperti warna bunga dan ketahanan terhadap penyakit tidak dipengaruhi atau sedikit dipengaruhi lingkungan serta hanya dikendalikan oleh satu atau dua gen dominan atau resesif atau disebut dengan karakter kualitatif. Namun karakter yang mempunyai nilai ekonomi dan agronomi sangat penting seperti ukuran tanaman, daya hasil, tahan kekeringan, tahan rebah, kualitas hasil umumnya dipengaruhi oleh banyak gen (Makmur 1992) yang masing-masing mempunyai pengaruh kecil pada karakter itu. Karakter demikian disebut karakter kuantitatif yang banyak dipengaruhi oleh faktor lingkungan. Peranan pengaruh genetik dan pengaruh lingkungan sulit diketahui secara langsung (Poespodarsono 1988). Permasalahannya adalah seberapa jauh suatu karakter disebabkan faktor genetik sebagai akibat aksi gen dan seberapa jauh disebabkan oleh faktor lingkungan (Syukur et al. 2012).
Karakter yang teramati dari suatu tanaman merupakan hasil dari genetik dan lingkungan, yaitu P = G + E. Ragam fenotipe terdiri dari ragam genetik (𝜎2G) dan ragam lingkungan (𝜎2E) serta interaksi antara keduanya. Rumus matematisnya:
6 𝜎2p = 𝜎2
G + 𝜎2E + 𝜎2GxE (Syukur et al. 2012). Ragam genetik (𝜎2G) dibagi menjadi ragam aditif (𝜎2A), dominan (𝜎2D), dan interaksinya (𝜎2I). Ragam aditif dan dominan berasosiasi dengan alel-alel pada satu lokus sedangkan ragam epistasis sebagai interaksi non-alelik pada dua atau lebih lokus (Kosev 2014).
Kontribusi ragam genetik dapat diduga dengan pendugaan heritabilitas. Heritabilitas merupakan perbandingan antara ragam genotipe dan total ragam fenotipe dari suatu karakter. Hubungan ini menggambarkan seberapa jauh fenotipe yang tampak merupakan refleksi dari genotipe (Syukur et al. 2012). Selain itu, heritabilitas merupakan komponen genetika yang menunjukkan seberapa besar suatu sifat diwariskan kepada turunannya (Rosidah et al. 2014).
Heritabilitas dibedakan menjadi heritabilitas arti luas dan heritabilitas arti sempit. Nilai heritabilitas berkisar antara 0–1 atau dapat dinyatakan ke dalam persentase (Poespodarsono 1988). Heritabilitas dikatakan tinggi apabila nilainya > 50%, sedang apabila nilainya 20–50% dan rendah apabila nilainya < 20% (Mangoendidjojo 2004).
Sifat kualitatif umumnya mempunyai heritabilitas tinggi, sebaliknya sifat kuantitatif mempunyai heritabilitas rendah. Sifat kualitatif dikendalikan oleh gen sederhana sehingga penampakan sifat tidak dikaburkan oleh lingkungan (Pospodarsono 1988). Nilai duga heritabilitas perlu diketahui untuk menduga kemajuan seleksi, apakah karakter tersebut banyak dipengaruhi oleh faktor genetik atau lingkungan. Jika keturunannya mempunyai ragam genetik lebih tinggi daripada ragam lingkungan maka heritabilitas akan tinggi (Syukur et al. 2012). Jika ragam genetik rendah daripada lingkungan maka heritabilitas rendah. Seleksi akan efektif ketika ragam genetik lebih tinggi daripada ragam lingkungan (Poehlman 1983).
Keragaman genetik dan heritabilitas sangat bermanfaat dalam proses seleksi. Pengaruh genetik mempunyai arti penting untuk menentukan nilai pemuliaan tanaman. Semakin tinggi perbedaan nilai genetik berarti seleksi makin efektif yaitu jika populasi tersebut mempunyai keragaman genetik yang luas dan heritabilitas yang tinggi (Poespodarsono 1988; Syukur et al. 2010). Efektifitas seleksi tanaman dengan hasil tinggi dalam populasi yang beragam akan tergantung pada (a) sejauh mana variabilitas hasil tanaman individu dalam populasi merupakan hasil dari faktor genetik pada tanaman terpilih, (b) sejauh mana variabilitas antara tanaman dengan lingkungan tumbuh. Seleksi tanaman untuk hasil tinggi tidak efektif jika variasi lingkungan lebih tinggi dari pada variasi genetik (Poehlman 1983).
Selain keragaman genetik dan heritabilitas (Syukur et al. 2012; Makmur 1992), agar seleksi efektif perlu pengetahuan tentang sistem genetik yang mengontrol ekspresi gen pada karakter kuantitatif (Shoba et al. 2010), pola segregasi, jumlah gen yang mengedalikan dan tipe aksi gen (Barmawi 2007) dan hubungan antar sifat (Poespodarsono 1988). Penelitian yang membahas tentang hubungan aksi gen terhadap karakter kuantitatif tanaman pada komoditas kedelai antara lain dilaporkan oleh Wang dan Gai (2001). Menurut Poespodarsono (1988), hubungan antar sifat digunakan untuk mengetahui seberapa besar pengaruh sifat lain terhadap sifat yang akan dituju. Jika ada hubugan yang erat dan positif maka akan memudahkan dalam proses seleksi.
7 Segregan Transgresif
Dari hasil perkawinan akan diperoleh keturunan yang mengalami segregasi atau pemisahan gen. Bila sepasang tetua keduanya homozigot menyerbuk sendiri maka akan terjadi segregasi pada F2. Segregasi terjadi pada proses meiosis yang menyebabkan gen-gen pada suatu lokus terpisah dan masing-masing dapat membentuk gamet yang berbeda. Semakin banyak pasangan gen yang mengalami segregasi maka makin banyak kombinasi baru pada keturunannya (Poespadarsono 1988). Segregasi transgresif adalah segregasi gen pada sifat-sifat kuantitatif dari zuriat hasil persilangan dua tetua yang memiliki jangkauan sebaran yang melampaui jangkauan sebaran kedua tetuanya atau yang memiliki nilai ekstrim dari tetuanya (Poehlman & Sleper 1996; Rieseberg et al. 1999; Rieseberg 2003). Segregasi transgresif membentuk dua gugus segregan transgresif dalam spektrum sebaran, yaitu lebih kecil dari sebaran tetua dengan keragaan rendah, dan lebih besar dari sebaran tetua dengan keragaan tinggi. Bila menggunakan seleksi positif, misalnya seleksi untuk memperoleh varietas yang produksi bijinya tinggi, kandungan protein biji tinggi, dan berbagai sifat yang ingin ditingkatkan nilai fenotipenya, maka gugus segregan transgresif dengan keragaan yang lebih besar dari keragaan tetua tertinggi yang akan ditingkatkan frekuensi genotipenya, sedangkan gugus segregan trasgresif dengan sebaran yang lebih kecil dari keragaan tetua rendah dibuang. Keadaan sebaliknya berlaku untuk seleksi negatif, misalnya seleksi untuk memperoleh varietas berumur genjah (Jambormias & Riry 2009).
8
3 KERAGAMAN GENETIK DAN SELEKSI SEGREGAN
TRANSGRESIF PADA POPULASI F3 KACANG TANAH
(Genetic Variability and Selection of Transgressive Segregant in F3 Population of Peanut)
Abstrak
Seleksi segregan transgresif adalah salah satu cara efektif dalam memperoleh genotipe kacang tanah dengan hasil tinggi. Penelitian ini bertujuan untuk menduga parameter genetik dan mengidentifikasi segregan transgresif kacang tanah generasi F3. Penelitian ini dilaksanakan pada bulan Agustus-Desember 2014 di kebun percobaan Leuwikopo, IPB, Dramaga, Bogor. Bahan genetik yang diuji sebanyak 218 galur generasi F3 dari 5 populasi biparental (Jerapah/GWS79A1, GWS79A1/Zebra, GWS79A1/ Jerapah, Zebra/ GWS79A1, dan Zebra/GWS18) dan 4 varietas komersial sebagai kontrol (Gajah, Jerapah, Sima, dan Zebra). Rancangan yang digunakan adalah augmented-Rancangan Kelompok Lengkap Teracak (RKLT) dengan 4 cek sebagai ulangan. Hasil penelitian menunjukkan bahwa karakter-karakter kuantitatif bersifat kontinu dikendalikan oleh banyak gen dengan aksi gen aditif, dominan, epistasis aditif, atau epistasis komplementer. Karakter jumlah polong total, jumlah polong isi memiliki nilai heritabilitas arti luas yang tinggi dan KKG yang sedang. Seleksi 10% menghasilkan 22 galur yang memiliki jumlah polong total terbaik, dimana 6 galur memiliki jumlah polong lebih tinggi daripada kedua tetuanya. Galur U2-39 terdeteksi sebagai galur segregan transgresif dengan jumlah polong total 30 dengan ragam dalam famili yang rendah daripada varietas Sima.
9 Abstract
Selection of transgressive segregants is considered as one effective way for obtaining peanut genotypes with high yield. The objective of the study was to estimate genetic parameters and to identify transgressive segregants of peanut lines in F3 generation. The experiment was conducted at Leuwikopo experimental station of IPB, Dramaga, Bogor (250 masl) from August to Desember 2014. The genetic materials evaluated were 218 lines of F3 generation from 5 biparental populations (Jerapah/GWS79A1, GWS79A1/ Zebra, GWS79A1/Jerapah, Zebra/GWS79A1, and Zebra/GWS18) and 4 commercial varieties as checks (Gajah, Jerapah, Sima, and Zebra). The experiment was arranged in an augmented randomized complete block design with 4 replications for the checks. The results showed that quantitative characters had continuous distribution and controlled by many genes with additive, dominance, complementary epistasis, or additive epistasis gene actions. Total number of pod and number of filled pod have high heritability and broad genetic coefficient of variation. Selection with 10% intensity showed that 22 genotypes had large total number of pods, where 6 lines among them had larger total number of pods than both parents. Line U2-39 was identified as a putative transgressive segregant with total number of pod of 30 and have smaller within family variance than Sima variety.
10
Pendahuluan
Kacang tanah (Arachis hypogaea L.) digunakan sebagai sumber minyak nabati dan sumber protein yang baik, selain itu digunakan sebagai bahan penganekaragaman pangan, juga sebagai bahan pakan ternak (Rukmana 1998; Santosa 2010; Shoba et al. 2010; Naeem-ud-Din et al. 2012; Wijayanti et al. 2014). Meningkatnya jumlah penduduk dan semakin berkembangnya industri pangan dan pakan berhubungan dengan peningkatan permintaan kacang tanah (Sudjadi & Supriyati 2001; Saleh 2010).
Produktivitas rata-rata kacang tanah dalam negeri mencapai kisaran 1.07-1.35 ton ha-1 polong kering (BPS 2014), sementara produktivitas di tingkat petani sebesar 1.02-1.11 ton ha-1 polong kering (Sudjadi dan Supriati 2001). Padahal potensi hasil kacang tanah yang telah dilepas selama kurun waktu tahun 1950-2004 dapat mencapai 2.8 ton ha-1 polong kering (Suhartina 2005). Peningkatan produksi dan produktivitas kacang tanah masih belum mengimbangi kebutuhan konsumsi dalam negeri sehingga Indonesia mengimpor sebanyak 132 ribu ton pada tahun 2009 dan menjadi 136 ribu ton pada tahun 2012 (Pusdatin 2013), bahkan dapat mencapai 200 ribu ton membengkak seiring dengan tingginya permintaan untuk kebutuhan pangan (Kasno & Harnowo 2014)
Pemuliaan tanaman memainkan peranan yang penting dalam meningkatkan produktivitas atau hasil tanaman (Jambormias & Riry 2009; Syukur et al. 2012). Salah satu hal penting dari program pemuliaan tersedianya keragaman genetik. Semakin besar keragaman genetik semakin besar pula peluang keberhasilan untuk memperoleh sifat-sifat genetik yang diinginkan (Supeno 2004). Adanya keragaman genetik dapat mempengaruhi efektivitas seleksi (Frey 1981; Chahal & Gosal 2003) Seleksi merupakan kegiatan penting dalam produksi kacang tanah (Smith & Simpson 1995). Pelaksanaan seleksi bertujuan untuk meningkatkan frekuensi genotipe-genotipe segregan yang dikehendaki dari dalam populasi homozigositas dan heterozigositas pada setiap generasi, hingga diperoleh genotipe-genotipe segregan transgresif homozigot untuk semua gen yang telah mengalami fiksasi (Jambormias & Riry 2009). Segregasi transgresif adalah segregasi gen pada sifat-sifat kuantitatif dari zuriat hasil persilangan dua tetua yang memiliki jangkauan sebaran melampaui jangkauan sebaran kedua tetuanya atau yang memiliki nilai ekstrim dari tetuanya (Poehlman & Sleper 1996; Rieseberg et al. 2003). Jambormias dan Riry (2009) menambahkan bahwa genotipe-genotipe dengan perilaku demikian dapat disebut sebagai segregan transgresif. Secara teoritis bila tidak ada pengaruh lingkungan yang besar maka suatu segregan transgresif telah ada pada generasi segregasi F2 atau pada generasi seleksi S1.
Menurut Makmur (1992), bila tanaman yang heterozigot pada satu lokus diserbuki sendiri, untuk mencapai fiksasi menjadi homozigot setidaknya memerlukan waktu sampai 5 generasi seleksi (S1-S5) atau sedikitnya sampai generasi F6. Karakter kuantitatif yang mempunyai nilai ekonomi dan agronomi sangat penting seperti ukuran tanaman, daya hasil, ketahanan, kualitas hasil umumnya dipengaruhi oleh banyak gen. Jambormias (2014) menyatakan bahwa periode seleksi akan semakin panjang jika melibatkan lebih dari satu gen untuk satu sifat kuantitatif. Periode seleksi dapat diperpendek dengan menggunakan seleksi nilai tengah tinggi dan ragam genotipe terpilih yang rendah sampai generasi F4. Penelitian Jambormias dan Riry (2009) telah berhasil menemukan terobosan untuk
11
mendeteksi segregan transgresif dan Jambormias (2014) meneliti segregan transgresif pada generasi awal segregasi atau pada generasi F3 komoditas kacang hijau. Hal inilah yang menjadi perhatian tujuan dilaksanakannya penelitian ini. Pada penelitian ini diharapkan seleksi dapat diperpendek dengan deteksi segregan transgresif pada generasi F3. Produk akhir dari deteksi segregan transgresif adalah didapatkannya galur-galur yang memiliki nilai tengah tinggi daripada sebaran kedua tetuanya dan ragam galur yang rendah atau sama dengan varietas komersial sehingga efektivitas waktu untuk mendapatkan varietas unggul baru dapat tercapai.
Metode Penelitian Waktu dan Tempat
Penelitian dilaksanakan pada bulan Agustus – Desember 2014 di kebun percobaan Leuwikopo, Darmaga, Bogor. Penghitungan komponen hasil dilaksanakan di Laboratorium Benih, Departemen Agronomi dan Hortikultura, Institut Pertanian Bogor.
Bahan dan Alat
Bahan pengujian terdiri atas 218 famili generasi F3 yang berasal dari 5 populasi biparental (Jerapah/GWS79A1, GWS79A1/Zebra, GWS79A1/ Jerapah, Zebra/ GWS79A1, dan Zebra/GWS18) dan 4 varietas komersial (Gajah, Jerapah, Sima, dan Zebra) sebagai varietas pembanding. Bahan lain yang digunakan adalah pupuk phonska dosis 200 kg ha-1, kaptan 0.5 ton ha-1 dan pupuk kandang dosis 1 ton ha-1. Alat yang digunakan berupa peralatan budi daya, ajir bambu, gunting, tali plastik, meteran, label, timbangan digital, dan kamera digital.
Prosedur Penelitian
Percobaan disusun dengan menggunakan Rancangan Augmented-Rancangan Kelompok Lengkap Teracak (RKLT) satu faktor yaitu galur kacang tanah. Menurut Petersen (1994), rancangan augmented digunakan karena bahan genetik yang dimiliki sangat banyak namun jumlah benih yang sedikit sehingga tidak memungkinkan untuk melakukan pengulangan pada tiap galurnya. Bahan yang diulang hanya pada genotipe pembanding. Pada percobaan ini, seluruh bahan genetik masing-masing ditanam sebagai barisan famili berukuran 20 tanaman yang tersebar pada 4 blok percobaan. Tiap galur ditanam tanpa ulangan sedangkan varietas komersial diacak pada 4 blok dengan blok sebagai ulangan.
Sebelum dilakukan penanaman, tanah diolah terlebih dahulu kemudian dibuat petakan sebanyak 4 petak yang sama. Pada saat penanaman, jarak tanam yang digunakan 40 cm x 22 cm satu benih per lubang. Pupuk phonska, pupuk kandang, dan kapur tanah diaplikasikan pada saat tanam. Pemeliharaan terdiri atas penyulaman, pengairan, penyiangan sekaligus pembumbunan dan pemberantasan hama dan penyakit yang sesuai dengan tingkat serangan yang timbul. Tanaman dipanen pada umur 105 HST (hari setelah tanam) dengan mencabut tanaman kemudian dipisahkan tiap galurnya. Panen ditandai oleh guratan nyata pada kulit polong, polong terisi penuh, dan kulit polong bagian dalam berwarna kecoklatan. Pengeringan polong dilakukan dengan cara dijemur ± 8 jam per hari pada cuaca cerah selama 3 hari. Pengamatan karakter agronomi pada 10 tanaman contoh tiap satuan percobaan meliputi: tinggi tanaman (cm), bobot brangkasan basah (g), bobot
12
polong basah (g), indeks panen, jumlah polong total, jumlah polong isi, bobot polong kering total (g), polong kering bernas (g), dan bobot biji per tanaman (g). Analisis Data
Data yang diperoleh dianalisis dengan menghitung:
a. Komponen ragam taraf α=5%. Penggunaan proc mixed di SAS pada rancangan
augmented yaitu perlakuan dan blok bersifat acak (Santos et al. 2002). Jika
perlakuan yang diuji berpengaruh nyata terhadap karakter yang diamati maka dilanjutkan dengan uji BNT taraf α=5% dengan bantuan aplikasi SAS 9.1.3. Komponen ragam dapat dilihat pada Tabel 1.
Tabel 1. Analisis ragam rancangan Augmented (Sharma 2006)
Sumber keragaman DB JK KT E(KT)
Blok r-1 JKu KTu
Perlakuan (g+k)-1 JKp KTp
Genotipe g-1 JKg KTg σ2 +σg2
Kontrol k-1 JKk KTk σ2 +rσk2
G vs K 1 JKg vs k KTg vs k
Galat k(r-1) JKe KTe σ2
Total terkoreksi (rk+g)-1 JKt
b. Model informasi famili dan informasi pembanding diuji asumsi kenormalan dengan uji Kolmogorov-Smirnov dan kehomogenan ragam uji Levene dan uji
Bartlett. Jika informasi famili dan pembanding menyebar normal, maka tidak
ada pengaruh aksi gen non aditif.
c. Bila terdapat pengaruh non aditif, maka digunakan uji skewness (kemenjuluran kurva) dan kurtosis (keruncingan kurva) menggunakan aplikasi Minitab 14, dan Ms. Excel (Gambar 2).
d. Pendugaan komponen ragam dan heritabilitas arti luas mengacu pada Allard (1960) dan Kasno (1993) serta ragam dalam galur mengacu pada Petersen (1994). Formula perhitungan sebagai berikut:
σe2=KTer ; σg2=(KTg-KTe)r ; σp2=σg2+σe2 hbs2 = σg2 σp2 x 100% ; KKG= √σg2 x̅ x 100% ; s 2= ∑xi2- ( ∑ xi) 2 n n-1 Keterangan: σ2g = ragam genetik σ2e = ragam lingkungan σ2p = ragam fenotipe
KTg = kuadrat tengah genotipe KTe = kuadrat tengah galat h2bs = heritabilitas arti luas
KKG = koefisien keragaman genetik
s2 = nilai ragam antar famili
r = jumlah ulangan
xi = nilai individu tanaman contoh tiap galur
x̅ = rataan umum dalam galur n = jumlah tanaman contoh
13
Gambar 2. Sebaran sifat kuantitatif kaitannya dengan aksi gen (Jambormias 2014)
Ya
Data Generasi Awal
Analisis Kurtosis
Kurtosis Nyata
Tidak Ya Intergenik Interaksi
Aditif Mesokurtik: Interaksi Interalelik Analisis Skewness Skewness Nyata Kurtosis Positif Leptokurtik: Sedikit gen terlibat Platikurtik: Banyak gen terlibat Ya Analisis Skewness Skewness Nyata Epistasis Aditif Positif Epistasis Komplementer Epistasis Duplikat Ya Tidak Ada Pengaruh Gen Dominan Skewness Positif Ya Tidak Menjulur ke kiri: dominansi ke kanan Menjulur ke kanan: dominansi ke kiri Tidak Hanya Pengaruh Aditif Ya Tidak Tidak
14
Hasil dan Pembahasan Analisis Komponen Ragam
Hasil analisis ragam pada Tabel 2 menunjukkan bahwa populasi yang diuji tidak berpengaruh nyata pada hampir semua sifat kuantitatif diantaranya karakter tinggi tanaman, bobot brangkasan basah, bobot polong basah, indeks panen, bobot polong kering isi, bobot polong kering total dan bobot biji per tanaman. Hal ini menunjukkan bahwa mayoritas pada karakter tersebut memiliki tingkat keseragaman yang tinggi antar galur. Hanya karakter jumlah polong isi dan jumlah polong total yang menunjukkan perbedaan yang nyata pada galur-galur yang diuji. Terdapatnya perbedaan yang nyata pada karakter tersebut mengindikasikan bahwa galur-galur yang diuji memiliki keragaman genetik yang tinggi untuk kedua karakter tersebut.
Tabel 2. Analisis ragam dengan kuadrat tengah karakter agronomi kacang tanah generasi F3
Karakter KT genotipe KT galur KT kontrol
Tinggi tanaman 86.795tn 80.794tn 569.704** Bobot brangkasan basah 0.015tn 0.015tn 0.047tn Bobot polong basah 0.015tn 0.015tn 0.040* Indeks panen 0.004tn 0.004tn 0.008tn Bobot polong kering total 33.316tn 32.908tn 73.176tn Bobot polong kering isi 32.400tn 31.962tn 75.034tn Jumlah polong total 29.626* 28.871* 92.095** Jumlah polong isi 24.371* 23.709* 80.134** Bobot biji per tanaman 0.144tn 0.134tn 0.013*
db= derajat bebas, tn,*,**= berturut-turut hasil uji F tidak nyata pada taraf 5%, nyata pada taraf 5%, dan nyata pada taraf 1%
Sebaran Sifat-sifat Kuantitatif pada Kacang Tanah
Kelima kombinasi persilangan merupakan populasi bersegregasi yang keragaan secara fenotipe pada generasi awal terdapat famili-famili seragam dan beragam (Tabel 3 dan 4). Karakter tinggi tanaman dengan nilai tengah maksimum pada populasi GWS79A1/Zebra (73.41 ± 8.14) sedangkan nilai tengah minimum pada populasi Zebra/GWS18 (58.62 ± 4.10). Karakter bobot brangkasan basah dan jumlah polong isi dengan nilai maksimum pada populasi GWS79A1/Zebra (143.24 ± 39.31 dan 18.35 ± 5.23) dan minimum pada Zebra/GWS79A1 (97.26 ± 34.15 dan 12.56 ± 3.57). Karakter indeks panen dengan nilai tengah maksimum pada Zebra/GWS18 (0.29 ± 0.11) dan minimum pada Jerapah/GWS79A1 (0.23 ± 0.06). Karakter bobot polong basah, bobot polong kering total, bobot polong kering isi, jumlah polong total dan bobot biji per tanaman dengan nilai maksimum pada populasi persilangan GWS79A1/Jerapah terdapat sedangkan nilai tengah minimum pada populasi Zebra/GWS7A1. Secara umum populasi GWS79A1/Jerapah memiliki penampilan fenotipe lebih baik dari persilangan lainnya. Pada varietas pembanding, keragaan nilai tengah pada hampir seluruh karakter lebih unggul pada varietas Jerapah (Tabel 4).
15
Tabel 3. Keragaan nilai tengah dan standar deviasi karakter pada tiap kombinasi persilangan kacang tanah
Karakter Nilai tengah ± SD Jerapah/ GWS79A1 GWS79A1/ Zebra GWS79A1/ Jerapah Zebra/ GWS79A1 Zebra/ GWS18A TT (cm) 66.65 ± 9.46 73.41 ± 8.14 69.62 ± 7.47 58.97 ± 7.85 58.62 ± 4.10 BB (g) 126.80 ± 39.69 143.24 ± 39.31 136.89 ± 37.29 97.26 ± 34.15 103.82 ± 25.66 BPB (g) 28.97 ± 8.33 34.01 ± 8.69 35.07 ± 7.91 28.64 ± 9.60 31.70 ± 11.41 IDX 0.23 ± 0.06 0.24 ± 0.06 0.26 ± 0.06 0.27 ± 0.05 0.29 ± 0.11 BPKT (g) 18.60 ± 5.05 21.63 ± 6.13 22.33 ± 5.88 17.36 ± 5.14 18.29 ± 6.73 BPKI (g) 18.20 ± 5.02 21.16 ± 6.12 21.79 ± 5.77 16.94 ± 5.09 17.92 ± 6.75 JPT 16.77 ± 4.18 20.58 ± 5.48 20.64 ± 5.49 14.18 ± 3.93 14.71 ± 4.81 JPI 15.00 ± 3.75 18.35 ± 5.23 18.14 ± 4.96 12.56 ± 3.57 12.83 ± 4.43 BBPT (g) 14.42 ± 3.61 16.98 ± 4.52 17.51 ± 4.49 13.33 ± 4.05 13.89 ± 5.20
Tabel 4. Keragaan nilai tengah dan standar deviasi karakter pada varietas pembanding kacang tanah
Karakter Nilai tengah ± SD
Gajah Jerapah Sima Zebra
TT (cm) 70.95 ± 11.44 63.43 ± 7.94 82.10 ± 11.79 53.88 ± 2.66 BB (g) 175.83 ± 59.32 136.95 ± 40.31 126.15 ± 33.15 92.26 ± 17.47 BPB (g) 35.16 ± 5.67 36.33 ± 6.64 34.60 ± 9.50 22.11 ± 5.09 IDX 0.21 ± 0.03 0.30 ± 0.06 0.29 ± 0.03 0.24 ± 0.09 BPKT (g) 20.08 ± 1.84 23.91 ± 3.29 20.91 ± 4.21 13.73 ± 3.64 BPKI (g) 19.64 ± 1.91 23.53 ± 3.34 20.44 ± 4.31 13.22 ± 3.32 JPT 19.85 ± 0.74 22.40 ± 2.13 15.96 ± 2.42 11.40 ± 3.22 JPI 17.85 ± 0.73 20.13 ± 2.00 14.49 ± 2.42 9.83 ± 2.74 BBPT (g) 15.38 ± 1.31 18.06 ± 2.31 14.79 ± 2.87 9.85 ± 2.07
TT= tinggi tanaman, BB= bobot brangkasan basah, BPB= bobot polong basah, IDX= indeks panen, BPKI= bobot polong kering isi, BPKT= bobot polong kering total, JPI= jumlah polong isi, JPT= jumlah polong total, BBPT= bobot biji per tanaman
Pendugaan Aksi Gen pada Generasi Awal
Hasil uji kenormalan dengan Kolmogorov-Smirnov dan uji kehomogenan ragam dapat dilihat pada Tabel 5 dan 6. Hampir seluruh karakter pada hampir semua kombinasi persilangan memperlihatkan sebaran yang tidak normal (p<0.05). Sebaran normal (p≥0.05) diperlihatkan oleh beberapa karakter diantaranya karakter tinggi tanaman pada 4 kombinasi persilangan serta jumlah polong total pada 3 kombinasi persilangan. Ragam homogen (p≥0.05) hanya terjadi pada kombinasi Zebra/GWS18 sebaliknya pada kombinasi lainnya memperlihatkan ragam yang tidak homogen (p<0.05). Menurut Jambormias et al. (2013), populasi generasi awal (F3) antar famili cenderung heterogen dan menyebar tidak normal. Selanjutnya Jambormias (2014) menyatakan bahwa ketidaknormalan dan ketidakhomogenan ragam selain karena pengaruh faktor aditif, pengaruh faktor non-aditif dan keterbauran berkontribusi terhadap keragaan tanaman. Distribusi sebaran yang tidak normal dan tidak homogen dapat dianalisis lebih lanjut berdasarkan nilai
skewness dan kurtosis.
Perbaikan karakter tanaman memerlukan pemahaman parameter genetik, yang pendugaannya dapat dilakukan dengan pendekatan analisis genetik yaitu aksi gen (Wahyu et al. 2015). Roy (2000) telah menyatakan bahwa analisis skewness dan kurtosis dapat digunakan untuk menentukan terjadi tidaknya epistasis pada zuriat persilangan. skewness dipengaruhi oleh gen letal, linkage, epistasis, gen
non-16
aditif (dominan atau over dominan), interaksi genotipe x lingkungan, pleiotropi, kompetisi, serta stres lingkungan suboptimum. Selanjutnya Roy (2000) menyatakan bahwa kurtosis negatif (platikurtik) atau positif (leptokurtik) berturut-turut mengindikasikan banyak atau sedikitnya gen yang terlibat dalam mengendalikan suatu karakter. Kurtosis nyata dengan skewness nyata negatif atau positif berturut-turut mengindikasikan aksi gen epistasis duplikat atau epistasis komplementer.
Kurtosis nyata tetapi skewness tidak nyata mengindikasikan aksi gen epistasis
aditif. Bila kurtosis tidak nyata maka sebarannya disebut mesokurtik. Kurtosis tidak nyata dengan skewness tidak nyata atau nyata berturut-turut mengindikasikan keterlibatan aksi gen aditif atau dominan. Kurtosis tidak nyata dengan skewness nyata negatif atau positif berturut-turut mengindikasikan adanya aksi gen dominansi ke kanan atau ke kiri.
Hasil penelitian menunjukkan bahwa karakter bobot polong kering total, bobot kering isi, jumlah polong total, jumlah polong isi, dan bobot biji per tanaman dikendalikan oleh banyak gen dengan aksi gen dominansi pada kombinasi persilangan Jerapah/ GWS79A1. Hal ini tidak sejalan dengan penelitian Halward dan Wayne (1991) pada persilangan kacang tanah NC-V11/GP-NC343 menunjukkan bahwa karakter bobot polong dan bobot biji dikendalikan oleh aksi gen aditif, dan pada persilangan NC-V11/FESR-11-P11-32 hampir semua karakter dikendalikan oleh aksi gen aditif. Karakter bobot brangkasan basah basah polong, bobot polong basah dan indeks panen dikendalikan oleh sedikit gen dengan aksi gen epistasis komplementer. Karakter tinggi tanaman dikendalikan oleh banyak gen dengan aksi gen aditif. Pada kombinasi persilangan GWS79A1/Zebra hampir seluruh karakter dikendalikan oleh sedikit gen dengan aksi gen epistasis komplementer kecuali karakter tinggi tanaman dan bobot biji per tanaman berturut-turut dikendalikan oleh banyak gen dengan tipe aksi gen aditif dan banyak gen dengan aksi gen epistasis komplementer. Pada kombinasi persilangan GWS79A1/Jerapah masing-masing karakter dikendalikan oleh sedikit atau banyak gen dengan aksi gen epistasis komplementer kecuali tinggi tanaman dikendalikan oleh aksi gen epistasis aditif.
Pada kombinasi persilangan Zebra/GWS79A1 hampir seluruh karakter dipengaruhi oleh banyak gen dengan aksi gen dominansi kecuali indeks panen dengan sifat aksi gen aditif. Hal ini sejalan dengan hasil penelitian Nigam et al. (2001) yaitu karakter indeks panen bersifat aditif. Pada persilangan yang sama, karakter bobot brangkasan basah dan bobot biji per tanaman dikendalikan oleh sedikit gen dengan aksi gen epistasis komplementer dan epistasis aditif. Pada populasi persilangan Zebra/GWS18, seluruh karakter dikendalikan oleh banyak gen dengan sifat aksi gen berbeda-beda. Karakter bobot brangkasan basah, bobot polong basah dan bobot biji dikendalikan oleh aksi gen dominansi. Karakter bobot polong kering total, bobot polong kering isi dan jumlah polong isi dikendalikan oleh aksi gen epistasis aditif. Karakter tinggi tanaman, indeks panen dan jumlah polong total dikendalikan oleh aksi gen aditif. Hal ini sejalan dengan hasil penelitian Hartati et al. (2013) dan Sihaloho et al. (2015) bahwa hampir semua karakter kuantitatif dikendalikan oleh banyak gen pada komoditas kedelai.
17
Tabel 5. Ukuran dan aksi gen karakter kuantitatif berdasarkan sebaran data galur-galur F3 pada 5 persilangan kacang tanah
Karakter Kenormalan Kehomogenan ragam Skewness Kurtosis Ukuran dan
aksi gen Bartlett Levene Jerapah / GWS79A1 TT >0.15 0.00 0.00 -0.07tn -0.13tn B, Ad BB <0.01 0.00 0.00 1.04** 2.12** S, EK BPB 0.04 0.00 0.00 0.66** 0.90** S, EK IDX <0.01 0.00 0.00 0.92** 2.82** S, EK BPKT 0.08 0.00 0.00 0.40** -0.23tn B, Dm BPKI 0.07 0.00 0.00 0.39** -0.25tn B, Dm JPT 0.12 0.00 0.01 0.46** 0.37tn B, Dm JPI >0.15 0.00 0.00 0.37** -0.07tn B, Dm BBPT 0.08 0.00 0.03 0.38** -0.25tn B, Dm GWS79A1/Zebra TT >0.15 0.01 0.34 -0.20tn -0.10tn B, Ad BB <0.01 0.00 0.00 0.84** 0.52* S, EK BPB <0.01 0.00 0.01 0.69** 0.67** S, EK IDX 0.03 0.00 0.00 0.46** 1.06** S, EK BPKT 0.03 0.00 0.01 0.71** 1.34** S, EK BPKI <0.01 0.00 0.01 0.71** 1.26** S, EK JPT 0.06 0.00 0.01 0.78** 1.58** S, EK JPI 0.03 0.00 0.00 0.81** 1.51** S, EK BBPT <0.01 0.00 0.01 0.47** -0.22tn B, EK GWS79A1/Jerapah TT 0.09 0.00 0.02 0.18tn 1.65** S, EA BB <0.01 0.00 0.00 0.76** 0.77** S, EK BPB 0.02 0.00 0.02 0.45** 0.05tn B, EK IDX <0.01 0.00 0.00 0.44** 1.09** S, EK BPKT <0.01 0.00 0.01 0.62** 1.03** S, EK BPKI 0.02 0.00 0.01 0.64** 1.10** S, EK JPT 0.03 0.00 0.01 0.52** 0.35tn B, EK JPI 0.07 0.00 0.01 0.56** 0.68** S, EK BBPT <0.01 0.00 0.01 0.67** 1.14** S, EK Zebra/GWS79A1 TT <0.01 0.00 0.29 0.32* 0.60tn B, Dm BB 0.14 0.07 0.74 0.73** 1.70** S, EK BPB 0.02 0.02 0.17 0.32* -0.61tn B, Dm IDX <0.01 0.00 0.01 -0.20tn -0.40tn B, Ad BPKT <0.01 0.25 0.49 0.38* -0.57tn B, Dm BPKI <0.01 0.30 0.53 0.41** -0.51tn B, Dm JPT 0.03 0.03 0.16 0.43** -0.21tn B, Dm JPI 0.02 0.06 0.24 0.44** -0.29tn B, Dm BBPT <0.01 0.17 0.36 0.29tn -0.83** S, EA Zebra/GWS18 TT >0.15 0.05 0.63 0.06tn -0.03tn B, Ad BB <0.01 0.01 0.20 0.75** 0.44tn B, Dm BPB <0.01 0.04 0.35 0.54** 0.06tn B, Dm IDX >0.15 0.19 0.26 0.08tn -0.62tn B, Ad BPKT <0.01 0.21 0.65 0.34tn -0.75* B, EA BPKI <0.01 0.19 0.62 0.36tn -0.74* B, EA JPT 0.09 0.15 0.59 0.31tn -0.67tn B, Ad JPI 0.02 0.08 0.57 0.33tn -0.75* B, EA BBPT <0.01 0.11 0.56 0.52** -0.29tn B, Dm
18
Tabel 6. Nilai P dari uji kenormalan dan kehomogenan ragam, serta uji skewness dan kurtosis karakter kuantitatif pada 4 varietas kacang tanah
Karakter Kenormalan Kehomogenan ragam Skewness Kurtosis Pengaruh
Bartlett Levene TT 0.15 0.07 0.09 0.20tn 0.10tn - BB <0.01 0.00 0.00 1.24** 2.31** Lingkungan BPB 0.03 0.04 0.01 0.36tn -0.21tn - IDX >0.15 0.00 0.00 1.05** 3.43** Lingkungan BPKT >0.15 0.10 0.05 0.16tn -0.42tn - BPKI >0.15 0.12 0.06 0.18tn -0.41tn - JPT >0.15 0.05 0.11 0.27tn -0.06tn - JPI >0.15 0.14 0.29 0.16tn -0.47tn - BBPT >0.15 0.03 0.06 0.25tn -0.31tn -
TT= tinggi tanaman, BB= bobot brangkasan basah, BPB= bobot polong basah, IDX= indeks panen, BPKI= bobot polong kering isi, BPKT= bobot polong kering total, JPI= jumlah polong isi, JPT= jumlah polong total, BBPT= bobot biji per tanaman, nilai P <0.05 menunjukkan terjadinya pelanggaran asumsi kenormalan atau kehomogenan ragam, tn,*,**= berturut-turut hasil uji F tidak nyata pada taraf 5%, nyata pada taraf 5%, dan nyata pada taraf 1%, S = sedikit gen terlibat, B = banyak gen terlibat, Ad = aditif, EA = epistasis aditif, Dm = dominan, EK = epistasis komplementer Seleksi dilakukan pada generasi berikutnya jika karakter yang mengendalikan suatu sifat dikendalikan oleh aksi gen dominansi atau epistasis (Barmawi 2007). Sedangkan bila karakter dikendalikan oleh aksi gen aditif maka karakter dapat diwariskan pada generasi berikutnya (Sulistyowati et al. 2015). Hasil penelitian Shoba et al. (2010) pada komoditas kacang tanah persilangan TMV2/ ICCGV97150 menunjukkan bahwa karakter tinggi tanaman, jumlah polong per tanaman, bobot polong per tanaman, bobot biji per tanaman, bobot 100 biji dan persentase pengisian dikendalikan oleh aksi gen aditif. Zhang et al. (2011) karakter hasil dikendalikan oleh banyak gen (poligenik) dengan dua gen mayor bersifat aditif yang dapat meningkatkan hasil kacang tanah.
Pada seluruh kombinasi persilangan yang diuji (Tabel 5), setiap persilangan menunjukkan aksi gen yang berbeda-beda. Kejadian ini mirip dengan penelitian Wahyu et al. (2014) bahwa setiap pasangan persilangan kedelai menunjukkan aksi gen yang berbeda pula. Kisman et al. (2008) melaporkan bahwa pola adaptasi kedelai cekaman naungan pada karakter-karakter yang diamati dikendalikan oleh aksi gen dominan parsial, aditif, atau epistasis. Pada penelitian Machfud dan Sulistyowati (2009), karakter yang mengendalikan kelebatan bulu daun pada kapas bersifat dominan sebagian negatif dan dominan sebagian positif. Menurut Roy (2000) dan Jayaramachandran et al. (2010), penyebaran karakter kuantitatif pada tanaman yang menjulur ke kiri (skewed left) atau ke kanan (skewed right) menunjukkan adanya pengaruh non-aditif seperti pengaruh lingkungan, interaksi genotipe x lingkungan, epistasis dan pautan gen.
Varietas pembanding yang disajikan pada Tabel 6 menunjukkan bahwa ketidakaknormalan dan ketidakhomogenan ragam untuk karakter bobot brangkasan basah dan indeks panen mengindikasikan bahwa percobaan belum benar-benar homogen di lapangan percobaan. Varietas pembanding yang merupakan varietas generasi lanjut yang homogen homozigot mengalami keterbauran, berkompetisi dengan famili-famili yang secara genetik dan fenotipe masih beragam. Ketahomogenan dan ketaknormalan tersebut dipengaruhi oleh faktor non-aditif yaitu interaksi antara genotipe x lingkungan.
19 Komponen Ragam dan Heritabilitas
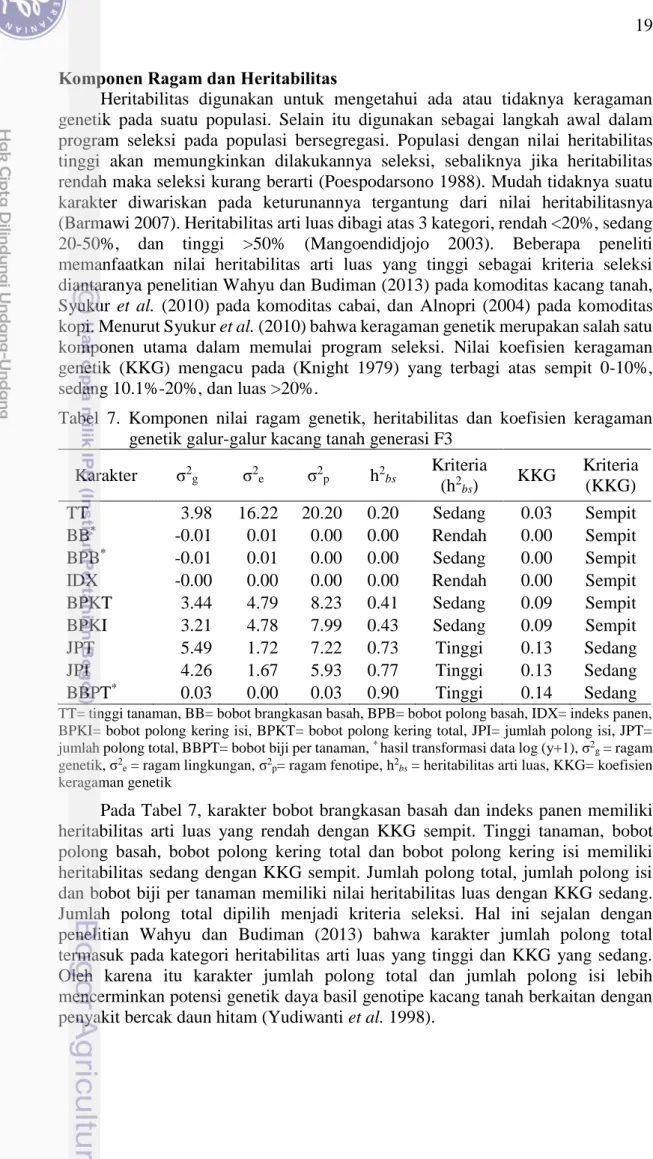
Heritabilitas digunakan untuk mengetahui ada atau tidaknya keragaman genetik pada suatu populasi. Selain itu digunakan sebagai langkah awal dalam program seleksi pada populasi bersegregasi. Populasi dengan nilai heritabilitas tinggi akan memungkinkan dilakukannya seleksi, sebaliknya jika heritabilitas rendah maka seleksi kurang berarti (Poespodarsono 1988). Mudah tidaknya suatu karakter diwariskan pada keturunannya tergantung dari nilai heritabilitasnya (Barmawi 2007). Heritabilitas arti luas dibagi atas 3 kategori, rendah <20%, sedang 20-50%, dan tinggi >50% (Mangoendidjojo 2003). Beberapa peneliti memanfaatkan nilai heritabilitas arti luas yang tinggi sebagai kriteria seleksi diantaranya penelitian Wahyu dan Budiman (2013) pada komoditas kacang tanah, Syukur et al. (2010) pada komoditas cabai, dan Alnopri (2004) pada komoditas kopi. Menurut Syukur et al. (2010) bahwa keragaman genetik merupakan salah satu komponen utama dalam memulai program seleksi. Nilai koefisien keragaman genetik (KKG) mengacu pada (Knight 1979) yang terbagi atas sempit 0-10%, sedang 10.1%-20%, dan luas >20%.
Tabel 7. Komponen nilai ragam genetik, heritabilitas dan koefisien keragaman genetik galur-galur kacang tanah generasi F3
Karakter σ2g σ2e σ2p h2bs Kriteria (h2 bs) KKG Kriteria (KKG) TT 3.98 16.22 20.20 0.20 Sedang 0.03 Sempit BB* -0.01 0.01 0.00 0.00 Rendah 0.00 Sempit BPB* -0.01 0.01 0.00 0.00 Sedang 0.00 Sempit IDX -0.00 0.00 0.00 0.00 Rendah 0.00 Sempit BPKT 3.44 4.79 8.23 0.41 Sedang 0.09 Sempit BPKI 3.21 4.78 7.99 0.43 Sedang 0.09 Sempit
JPT 5.49 1.72 7.22 0.73 Tinggi 0.13 Sedang
JPI 4.26 1.67 5.93 0.77 Tinggi 0.13 Sedang
BBPT* 0.03 0.00 0.03 0.90 Tinggi 0.14 Sedang
TT= tinggi tanaman, BB= bobot brangkasan basah, BPB= bobot polong basah, IDX= indeks panen, BPKI= bobot polong kering isi, BPKT= bobot polong kering total, JPI= jumlah polong isi, JPT= jumlah polong total, BBPT= bobot biji per tanaman, * hasil transformasi data log (y+1), σ2
g = ragam
genetik, σ2
e = ragam lingkungan, σ2p= ragam fenotipe, h2bs = heritabilitas arti luas, KKG= koefisien keragaman genetik
Pada Tabel 7, karakter bobot brangkasan basah dan indeks panen memiliki heritabilitas arti luas yang rendah dengan KKG sempit. Tinggi tanaman, bobot polong basah, bobot polong kering total dan bobot polong kering isi memiliki heritabilitas sedang dengan KKG sempit. Jumlah polong total, jumlah polong isi dan bobot biji per tanaman memiliki nilai heritabilitas luas dengan KKG sedang. Jumlah polong total dipilih menjadi kriteria seleksi. Hal ini sejalan dengan penelitian Wahyu dan Budiman (2013) bahwa karakter jumlah polong total termasuk pada kategori heritabilitas arti luas yang tinggi dan KKG yang sedang. Oleh karena itu karakter jumlah polong total dan jumlah polong isi lebih mencerminkan potensi genetik daya basil genotipe kacang tanah berkaitan dengan penyakit bercak daun hitam (Yudiwanti et al. 1998).
20
Identifikasi Galur-galur Segregan Transgresif
Sebelum melakukan seleksi terlebih dahulu melihat berapa proporsi nilai heritabilitas yang mencerminkan potensi genetik yang sudah dibahas sebelumnya. Menurut Chahal dan Gosal (2003), seleksi akan efektif jika tersedia keragaman dalam populasi. Sa’adiyah et al. (2009) menambahkan bahwa efektivitas seleksi semakin tinggi seiring dengan besarnya tingkat keragaman pada populasi yang diuji sehingga tujuan yang diinginkan dapat tercapai.
Nilai heritabilitas yang tinggi dan KKG sedang terdapat pada karakter jumlah polong isi, jumlah polong total dan bobot biji per tanaman. Namun seleksi lebih ditekankan pada karakter jumlah polong di mana pada laporan Kusumo (1996) disebutkan bahwa karakter jumlah polong mencerminkan potensi genetik daya hasil. Seleksi pada karakter jumlah polong total lebih efektif dilakukan daripada bobot biji per tanaman karena karakter bobot lebih dipengaruhi oleh faktor lingkungan selama proses pengisian polong. Selanjutnya Puangbut et al. (2013) menambahkan bahwa jumlah polong isi merupakan faktor utama dalam meningkatkan daya hasil kacang tanah. Purnamawati (2012) menyebutkan bahwa karakter jumlah polong berpengaruh langsung positif terhadap peningkatan hasil polong. Wahyu dan Budiman (2013) memilih karakter jumlah polong karena karakter tersebut merupakan faktor utama dalam meningkatkan daya hasil kacang tanah.
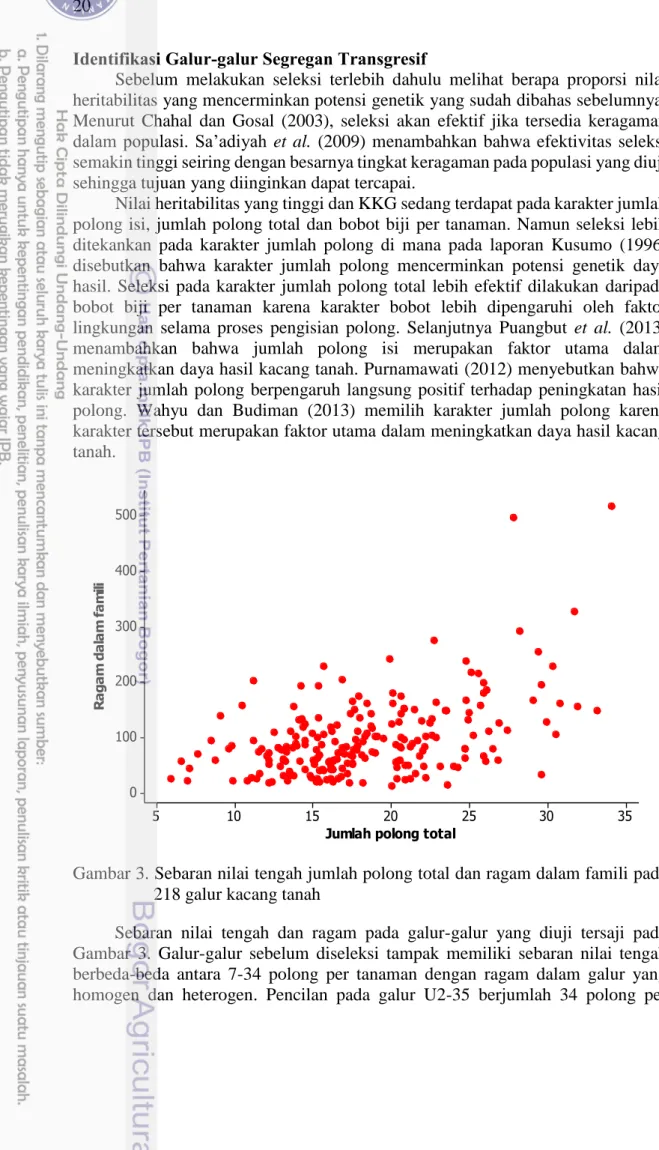
Gambar 3. Sebaran nilai tengah jumlah polong total dan ragam dalam famili pada 218 galur kacang tanah
Sebaran nilai tengah dan ragam pada galur-galur yang diuji tersaji pada Gambar 3. Galur-galur sebelum diseleksi tampak memiliki sebaran nilai tengah berbeda-beda antara 7-34 polong per tanaman dengan ragam dalam galur yang homogen dan heterogen. Pencilan pada galur U2-35 berjumlah 34 polong per
35 30 25 20 15 10 5 500 400 300 200 100 0
Jumlah polong total
R a g a m d a la m f a m ili